
Acqua
Acqua
di Ven Te Chow, John W. Boylan e Kurt Kramer
ACQUA
Acqua di Ven Te Chow
sommario: 1. L'acqua in natura. a) La scoperta dell'acqua. b) Il ciclo idrologico. c) Gli studi idrologici. 2. L'acqua nella società. a) L'acqua e le civiltà. b) Lo sviluppo delle riserve d'acqua. 3. L'acqua nel futuro. a) Modifiche del ciclo idrologico. b) Trasporto dell'acqua su vasta scala. c) Trattamento delle acque. □ Bibliografia.
1. L'acqua in natura
L'acqua è la sostanza più diffusa sulla Terra. I soli oceani ricoprono più del 70% della superficie terrestre e contengono l'incredibile quantità di 1.350.000.000 di chilometri cubici d'acqua. Altri 770.000.000 di chilometri cubici si trovano nella litosfera sotto forma di acqua di idratazione e 28.000.000 di chilometri cubici sono contenuti nelle calotte polari e nei ghiacciai. Attorno alla Terra, l'acqua è presente in una regione dello spazio, detta ‛idrosfera', che si estende nell'atmosfera sino ad una quindicina di chilometri di altezza e nella litosfera sino alla profondità media di un chilometro.
L'acqua è anche il costituente principale di tutti gli organismi viventi. Senz'acqua non ci sarebbe vita, poiché la vita dipende dall'acqua. Ogni organismo vivente è costituito, infatti, in massima parte d'acqua. Il corpo umano ne contiene circa per il 65% del suo peso e un uomo morrebbe in breve tempo non appena perdesse il 12% dell'acqua presente nel proprio corpo. Un pollo contiene acqua per il 74% del proprio peso, un pomodoro per il 95%. Quasi ogni organismo è condizionato completamente dall'acqua per più del 50% del proprio peso corporeo. La vita stessa potrebbe avere avuto origine nell'acqua e precisamente nell'acqua salata del mare. Il sapore salso del sangue, del sudore e delle lacrime suggerisce chiaramente una tale possibilità.
L'acqua ha cominciato a plasmare la forma della Terra dal momento stesso della sua comparsa. La pioggia martella il terreno ed erode il suolo, le onde del mare si riversano sulle coste, cesellando gli scogli e trasportando via la terra. I fiumi formano le valli ed i delta alluvionali. I ghiacciai fendono le montagne e solcano le valli.
L'importanza dell'acqua non è limitata alle funzioni vitali di sostentamento degli organismi e alla determinazione della morfologia terrestre. L'acqua è un fattore chiave nel condizionamento climatico della Terra, per l'esistenza dell'uomo e per lo sviluppo della civiltà. Oggi si considera spesso l'acqua alla stregua di un bene di consumo che possiede un suo valore economico ed è oggetto di dispute legali, sociali e politiche.
Con l'espansione della popolazione del globo e col miglioramento delle condizioni di vita, la domanda d'acqua va rapidamente aumentando è la sua disponibilità sta diventando sempre più problematica. L'uomo, in definitiva, dovrà fare sempre miglior uso dell'acqua a sua disposizione. Requisito fondamentale per affrontare un tale impegno è ovviamente quello di possedere una conoscenza dell'acqua fondata su valide basi scientifiche.
a) La scoperta dell'acqua
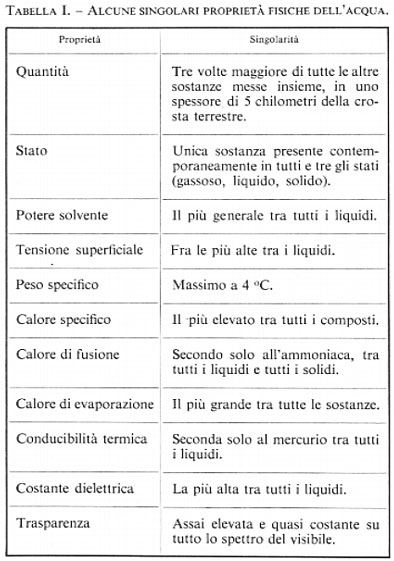
L'acqua è l'unica sostanza che sia presente in natura, simultaneamente e in quantità notevoli, nei tre distinti stati: gassoso, liquido e solido. Questa particolarità dell'acqua fu compresa e messa bene in evidenza da Talete di Mileto (624/23-548/45 a.C.) circa 2.500 anni or sono. Per questa ed altre singolari proprietà, che sono elencate nella tab. I, lo studio dell'acqua ha condotto a scoperte scientifiche importanti e ci si è avvalsi dell'acqua per introdurre dei campioni o dei punti di riferimento nei lavori scientifici: per es., ci si riferisce al livello medio del mare, al peso specifico di un corpo relativo a quello unitario dell'acqua, alla calibrazione della scala Celsius della temperatura effettuata sulla base dei punti di solidificazione e di ebollizione dell'acqua, ecc.
La sintesi dell'acqua fu ottenuta per la prima volta da H. Cavendish (1731-1810). Nel 1781 egli dette fuoco ad una miscela di ‛aria infiammabile' (idrogeno) ed aria comune (contenente ossigeno); ne ottenne un'esplosione e la formazione di goccioline di un liquido che immediatamente identificò come acqua. Successivamente a questa scoperta, J. Priestley (1733-1804), A.-L. Lavoisier (1743-1794) e J. Watt (1736-1819) dimostrarono sperimentalmente che l'acqua è composta, in peso, di otto parti di ossigeno e di una parte di idrogeno. Sir H. Davy (1778-1829) fece poi uso dell'elettrolisi per stabilire che la composizione dell'acqua è di due volumi di idrogeno e di un volume di ossigeno.
Sulla base delle leggi di J. Dalton (1766-1844) per gli atomi, J.-L. Gay-Lussac (1778-1850) determinò per primo sperimentalmente in modo assai ben approssimato la formula chimica dell'acqua, che A. Avogadro di Quaregna (1776-1856) correttamente propose nel 1811 come H2O. Fu soltanto nel 1934, tuttavia, che H. C. Urey (n. 1893) dimostrò che H2O non rappresenta compiutamente la composizione chimica dell'acqua. Egli scoprì un isotopo dell'idrogeno, detto idrogeno pesante (deuterio) D e l'acqua pesante D2O che fu separata per la prima volta dall'acqua normale nel 1932 da G. N. Lewis (1875-1946). Seguì quindi la scoperta di idrogeno ancor più pesante (trizio) T e dell'acqua T2O e fu anche stabilito che l'ossigeno possiede tre isotopi. Dal momento che ciascun tipo isotopico di acqua si ionizza in modo eguale agli altri, si possono avere, teoricamente, ben trentatré diverse entità fra molecole neutre e ioni da queste originatisi: diciotto composti molecolarmente distinti di acqua, più quindici tipi differenti di ioni.
Per interpretare coerentemente con i principi della scienza moderna le proprietà dell'acqua nei suoi tre diversi stati, una molecola d'acqua isolata può essere descritta in due modi complementari. Secondo l'uno, alcune proprietà, come le posizioni relative dei nuclei e la polarità della molecola, si determinano mediante esperienze effettuate su vapor d'acqua, a pressioni sufficientemente basse o temperature sufficientemente elevate da assicurare che non ci siano praticamente interazioni tra le molecole. Secondo l'altro, ci si avvale della teoria elettronica della valenza chimica per spiegare dettagliatamente quelle proprietà, come per esempio la disposizione della carica elettronica in seno alla molecola, che non possono essere rivelate dal primo metodo di indagine.
I tre stati in cui si presenta l'acqua (vapore, ghiaccio e liquido) si possono quindi descrivere mediante ulteriori considerazioni relative all'interazione tra le molecole. Per il vapore si sono già svolte indagini sulle forze interagenti tra molecole. Per interpretare le proprietà del ghiaccio, inoltre, vanno considerati la struttura cristallina ed i livelli energetici delle molecole stesse. Per ciò che riguarda l'acqua allo stato liquido, ci si è avvalsi delle sue caratteristiche macroscopiche per comprenderne la struttura. Inoltre, si sono sviluppati molti modelli teorici dell'acqua allo stato liquido, dei quali ricordiamo i più recenti: il modello a reticolo casualmente distribuito di J. D. Bernal (1964) ed il modello interstiziale di A. H. Narten, M. D. Danford e H. A. Levy (1967).
Tuttavia, nonostante i numerosi tentativi ed i continui sforzi compiuti dagli scienziati, la comprensione della struttura e delle proprietà dell'acqua non è ancor oggi completa.
b) Il ciclo idrologico
L'acqua presente in natura circola e si trasforma nell'idrosfera seguendo dei percorsi che costituiscono il cosiddetto ‛ciclo idrologico'. Il ciclo idrologico non ha principio né termine. A mano a mano che l'acqua evapora dagli oceani e dalle terre, essa diviene parte dell'atmosfera. Il vapore s'innalza ed è trasportato per l'atmosfera sino a che si condensa e precipita sulla terra o sul mare. L'acqua precipitata può essere intercettata dalle piante o traspirata, può scorrere in superficie o venir giù pei declivi fra strati diversi del terreno, oppure può infiltrarsi sottoterra. L'acqua che rimane alla superficie riempie le zone più basse, si raccoglie e poi evapora oppure, dopo essersi raccolta, scorre via in un secondo tempo. Gran parte dell'acqua intercettata e traspirata e di quella che scorre sul terreno ritorna nell'atmosfera in seguito ad evaporazione. L'acqua che si infiltra nel terreno può raggiungere le regioni più profonde ed ivi raccogliersi sotto forma di acqua sotterranea, per uscir quindi fuori come sorgente od incanalarsi in un corso d'acqua entrando a far parte delle acque di scorrimento, ed evaporare infine nell'atmosfera per completare così il ciclo idrologico. In tal modo, nel ciclo idrologico hanno luogo complicati processi di condensazione, precipitazione, evaporazione, intercettamento, traspirazione, infiltrazione, permeazione, ritenzione, detenzione, dilavamento di superficie, incanalamento, scorrimento. Si è soliti formulare vari diagrammi illustrativi del ciclo idrologico; taluni sono qualitativi, altri descrittivi ed altri ancora quantitativi.
L'acqua dell'atmosfera è, in gran parte, presente allo stato aeriforme, cioè di vapore, mentre allo stato liquido, come nel caso della pioggia o delle piccole gocce d'acqua presenti nelle nuvole, o allo stato solido, come nel caso della neve, della grandine o dei cristalli di ghiaccio nelle nuvole, essa appare solo saltuariamente e in zone circoscritte dello spazio. La quantità totale di vapore acqueo contenuto nell'atmosfera è relativamente piccola. Se si potesse condensare tutta sulla superficie della Terra, essa formerebbe uno strato d'acqua alto in media appena due centimetri e mezzo. Ad ogni istante, la quantità di vapore acqueo presente nell'atmosfera costituisce solo la cento- millesima parte di tutte le acque che partecipano al ciclo idrologico.
Sebbene il vapore acqueo sia presente in piccola quantità nell'atmosfera, pure esso è di fondamentale importanza perché costituisce la fase ascendente del ciclo idrologico. Il vapore acqueo si forma essenzialmente per evaporazione dell'acqua presente allo stato liquido sulla superficie terrestre. Sale quindi, per effetto delle correnti ascendenti, a migliaia di metri di altezza e viene trasportato fin sulle vette dei monti più alti e nel cuore dei continenti più estesi. Quando si condensa, esso dà luogo alle precipitazioni, le quali cadono sulla superficie della Terra e rappresentano la maggior parte della fase discendente del ciclo idrologico. Senza il vapore acqueo, il ciclo idrologico non potrebbe completarsi.
Il vapore acqueo adempie un'altra distinta funzione, assai meno evidente: mentre consente alla radiazione solare proveniente dall'alto di raggiungere il suolo, trattiene e assorbe la radiazione riflessa dal suolo, sì da non lasciarla sfuggire nello spazio. Questo fenomeno mantiene la temperatura della superficie terrestre ad un livello assai più elevato di quello che non sarebbe altrimenti. Se il vapore acqueo non regolarizzasse la temperatura terrestre, non vi sarebbe né acqua allo stato liquido, né vita sulla Terra.
Per condensazione, il vapore acqueo dell'atmosfera dà luogo alle precipitazioni. Il processo di condensazione può aver luogo non appena nell'aria vi sia più vapore di quanto questa non ne possa ricevere per evaporazione da una superficie di acqua liquida alla temperatura a cui avviene il processo. Una tale condizione può determinarsi in seguito a raffreddamento od a rimescolamento. Se una massa d'aria calda e satura si mescola ad una massa d'aria fredda, anch'essa satura, la pressione del vapore della mescolanza può essere maggiore della tensione del vapore saturo proprio della temperatura di equilibrio. Ciò, tuttavia, provoca raramente qualcosa di più di una tenue nebulosità o di una foschia sulla superficie della Terra.
Tutte le nuvole si formano per l'abbassamento della temperatura al di sotto del ‛punto di rugiada'. Se però l'aria che si raffredda non contiene gocce d'acqua, può anche darsi che non si abbia condensazione, pur essendo la temperatura scesa sino al ‛punto di rugiada' o anche un poco al di sotto, determinandosi in tal modo una condizione di ‛soprasaturazione' senza un'effettiva condensazione del vapore in gocce d'acqua. Tuttavia, la condizione di soprasaturazione è assai poco stabile e una piccola perturbazione può far riunire assieme un piccolo numero di molecole d'acqua sì da formare un nucleo ed iniziare il processo di condensazione.
Diversamente dal caso della nebbia, le nuvole si formano in seguito al raffreddamento, provocato dall'innalzamento di aria umida e non per cessione di calore all'ambiente circostante. A mano a mano che l'aria sale, si trova sottoposta ad una pressione sempre più bassa, si espande e quindi si raffredda. Questo tipo di raffreddamento provoca il fenomeno della precipitazione. Se l'innalzamento è dovuto al confluire dell'aria in una zona di bassa pressione, o ciclonica, ne consegue, specialmente nelle regioni di pianura, la cosiddetta ‛precipitazione ciclonica'. Se l'innalzamento è dovuto alla naturale salita d'aria più calda e leggera in zone più fredde e a densità maggiore, si ha, come nel caso dei temporali, una ‛precipitazione convettiva'. Se l'innalzamento avviene poi per via meccanica, per il moto dell'aria al di sopra di una barriera topografica, ad esempio una montagna, ne consegue, dal lato della barriera esposto al vento, una ‛precipitazione orografica'. In natura, gli effetti di queste varie forme di raffreddamento sono spesso collegati insieme, così che la precipitazione risultante non può chiaramente dirsi dovuta ad una sola di esse.
La precipitazione, nel corso del ciclo idrologico, può subire essenzialmente quattro diversi destini. Può evaporare durante la caduta o dopo aver toccato la superficie del suolo. Può essere intercettata dalla vegetazione ed evaporare quindi dalla superficie delle foglie. Può infiltrarsi nel terreno e costituire parte di quell'acqua propria dell'umidità della terra e di qui poi infine ricollegarsi alle acque sotterranee o fuoriuscire dal terreno, quindi evaporare o diventare parte delle acque di scorrimento. Può anche cadere direttamente su torrenti, fiumi, laghi o altre distese d'acqua e quindi evaporare o diventare parte delle acque di scorrimento superficiale.
Se la precipitazione cade sulla vegetazione, può seguire tre vie diverse. Può gocciolare giù a terra attraverso le foglie; può colare giù lungo i ramoscelli, i rami ed il tronco fino a terra o, infine, può essere trattenuta dalle foglie ed evaporare in seguito direttamente dalle superfici esposte di queste. A quest'ultima sequenza di eventi si dà il nome di ‛intercettamento'. In questo caso, l'acqua non raggiunge il terreno e così completa il ciclo idrologico senza passare attraverso la sua fase originaria in terra.
Nel trasferimento su vasta scala dell'acqua e del vapore acqueo tra oceani e continenti, l'evaporazione costituisce uno dei fattori più importanti del ciclo idrologico. Per effetto dell'evaporazione, l'acqua passa dallo stato liquido a quello aeriforme, cioè di vapore. Questo processo di trasformazione ha luogo nel caso in cui alcune molecole nella massa dell'acqua abbiano raggiunto un'energia cinetica sufficiente per uscir fuori dalla superficie libera di essa. Le molecole che sfuggono risentono dell'attrazione da parte delle altre molecole che tendono a trattenerle dentro l'acqua; possono quindi sfuggire via attraverso la superficie solamente quelle molecole che hanno un'energia cinetica maggiore di quella media delle molecole contenute dentro il liquido. L'evaporazione è perciò misurata dal numero delle molecole d'acqua che sono fuoriuscite. Durante il processo di evaporazione, la temperatura del liquido si abbassa in seguito alla fuoriuscita delle molecole d'acqua per effetto della trasformazione di energia da termica in cinetica. Perché ciò non avvenga, dev'essere disponibile una sorgente esterna di calore, quale può essere la radiazione solare o il calore ceduto dall'atmosfera o dal terreno.
Il moto delle molecole che vanno a formare il vapore attraverso la superficie dell'acqua produce una pressione. Questa pressione del vapore acqueo prende il nome di ‛tensione di vapore'. In realtà, essa è soltanto la pressione parziale del vapore acqueo nell'atmosfera, poiché in una miscela gassosa ciascun gas esercita una pressione parziale che non dipende dalla pressione degli altri gas nella miscela. La velocità di evaporazione dipende dalla differenza tra la tensione di vapore acqueo e la pressione dell'aria al di sopra della superficie. In certe condizioni, la quantità d'acqua che evapora è proporzionale alla differenza tra la pressione del vapore saturo alla temperatura dell'acqua e l'effettiva tensione di vapore nell'aria. Questo fatto è noto col nome di ‛legge di Dalton', poiché fu J. Dalton a porlo per primo in evidenza nel 1802.
Il processo di traspirazione dai vegetali consiste essenzialmente nell'evaporazione dell'acqua dalle cellule delle foglie. Per effetto di questo processo, l'acqua presente nel terreno viene inizialmente assorbita dal sistema delle radici delle piante e restituita quindi all'atmosfera attraverso le foglie sotto forma di vapore. In tale processo, soltanto una piccolissima parte dell'acqua, meno dell'1%, viene trattenuta per costruire i tessuti per la crescita e lo sviluppo della pianta.
Dopo aver raggiunto il terreno, parte dell'acqua precipitata sarà assorbita dagli strati superficiali. A questa fase del ciclo idrologico si dà il nome di ‛infiltrazione'. L'acqua infiltrata può scendere giù attraverso il suolo, il sottosuolo, gli strati di roccia e diventare parte, infine, di una o più delle zone umide del sottosuolo. L'acqua di infiltrazione si aggiunge all'umidità del suolo penetrando in un primo tempo nella zona chiamata propriamente ‛suolo', la quale costituisce uno strato relativamente sottile di terra vicino alla superficie. L'umidità, in genere, può andare via dal suolo attraverso l'evaporazione, la traspirazione od anche per effetto dell'infiltrazione verso la falda freatica. Poco al di sopra della falda freatica si trova la ‛frangia capillare', nella quale l'acqua è trattenuta dalle forze di capillarità che si contrappongono alla gravità. Tra la zona del suolo e la frangia capillare c'è la ‛zona intermedia', in cui l'acqua si muove generalmente verso il basso od anche in direzione orizzontale, purché si instauri in quella direzione un gradiente idraulico. Gli strati di terra al di sopra della falda freatica, che contengono quantità d'acqua inferiori al valore di saturazione, costituiscono la ‛zona di aerazione'. Tale zona è di transizione e quivi l'acqua è assorbita, trattenuta o spinta in basso verso la falda freatica, in alto verso la superficie del terreno o lateralmente in una direzione di flusso.
L'acqua che si trova sulla superficie del terreno deriva principalmente, per via diretta o indiretta, dalle precipitazioni. La maggior parte di quella che cade durante la prima fase di un temporale viene immagazzinata nelle depressioni del terreno, formando un ‛deposito superficiale', mentre la parte restante s'infiltra nel suolo o scorre pei pendii come acqua di dilavamento finché, trovato un solco, diviene parte di un corso d'acqua. Al continuare della precipitazione, il deposito superficiale aumenta se la quantità d'acqua che cade nell'unità di tempo è superiore alla capacità d'infiltrazione del terreno. Una buona parte dell'acqua contenuta nei depositi di superficie evapora infine e non subisce né il processo d'infiltrazione né quello di scorrimento superficiale sotto forma di acqua dilavante. Questa quantità d'acqua prende il nome di ‛deposito di ritenzione' o ‛ritenzione superficiale'. La parte restante del deposito superficiale, che è temporaneamente trattenuta e che diverrà in seguito acqua di infiltrazione o di dilavamento, prende il nome di ‛deposito di detenzione'. Estesi specchi d'acqua sulla superficie terrestre altro non sono che una combinazione di depositi di ritenzione e di detenzione. In generale, le acque di superficie sono costituite dalle acque di deposito superficiale, dalle acque dilavanti (o ‛selvagge') e da quelle incanalate.
Nel ciclo idrologico lo scorrimento delle acque superficiali ha una parte importante. ‛Scorrimento' viene detta quella frazione della precipitazione che compare in ultimo sotto forma di corsi d'acqua superficiali, aventi carattere perenne o intermittente. Esso è il flusso che si raccoglie per la maggior parte entro un bacino idrografico, del quale appare come lo scarico o sbocco. A seconda della sorgente da cui deriva il flusso, lo scorrimento può essere superficiale e sotterraneo più o meno profondo.
Lo ‛scorrimento superficiale' è costituito da quella parte delle acque di scorrimento che fluisce sulla superficie del terreno e attraverso solchi, per raggiungere lo sbocco del bacino. A quella porzione dello scorrimento superficiale che si sposta sul terreno verso i letti dei corsi d'acqua si dà il nome di ‛acque di dilavamento'. Quando queste sono divenute parte di un corso d'acqua, si uniscono ad altri flussi e costituiscono la totalità delle acque di scorrimento presenti nel letto di un tale corso. La precipitazione che cade direttamente nel letto del corso d'acqua è relativamente scarsa e viene abitualmente inclusa nello scorrimento superficiale.
Dello ‛scorrimento sotterraneo poco profondo' entra a far parte quella precipitazione che si infiltra nel sottosuolo e scorre lateralmente verso i fiumi e i torrenti lungo i primi strati del terreno. Una parte delle acque di tale scorrimento sotterraneo può subito riversarsi in un corso d'acqua, mentre la parte restante può confluirvi solo dopo molto tempo, oppure può ricomparire sulla superficie ed evaporare senza raggiungere alcun corso d'acqua.
Lo ‛scorrimento sotterraneo di profondità' è costituito da quella porzione delle acque di infiltrazione che ha raggiunto la falda acquifera attraverso una lunga migrazione e si scarica nei corsi d'acqua.
Dopo che l'acqua di precipitazione ha imbevuto il suolo, una parte di essa può infiltrarsi in profondità nella zona di aerazione fino alla falda freatica, diventando così acqua profonda. Poiché quest'acqua di infiltrazione deriva originariamente dalle precipitazioni, viene detta ‛acqua me- teorica'. In molte zone aride, ove vi sia un drenaggio sotterraneo, la sorgente principale di acqua meteorica è costituita dall'interramento dello scorrimento fluviale e dei laghi. L'acqua profonda, oltre che dall'acqua meteorica, deriva anche, per piccole quantità, da alcune sorgenti secondarie d'acqua localizzate nella crosta terrestre. Tra queste sorgenti vi è l'‛acqua fossile', che è rimasta racchiusa nelle rocce di sedimentazione sin dal tempo della loro formazione, e l'‛acqua giovanile', che si è formata per via chimica entro la terra e che è stata portata alla superficie per la prima volta dall'attività vulcanica. A causa della loro origine, le acque fossili e giovanili hanno spesso un contenuto minerale.
L'acqua profonda è in continuo movimento: in confronto, però, con l'acqua di superficie, essa si muove più lentamente e in maniera assai più varia. Può scorrere ad una velocità che va da qualche millesimo di centimetro al giorno nei suoli e in alcune rocce permeabili a grana sottile fino a qualche chilometro al giorno nelle formazioni geologiche fessurate. Per basse velocità di scorrimento, il flusso è laminare e la velocità dipende dalla permeabilità del materiale attraverso cui l'acqua fluisce e dal gradiente della pressione idraulica, cioè dall'inclinazione della falda freatica, nel caso in cui si tratti di acque sotterranee libere, ovvero dalla variazione della quota piezometrica, nel caso in cui si tratti di acque sotterranee artesiane confinate. Questo fenomeno del moto delle acque attraverso materiali porosi è descritto mediante la legge di Darcy, dal nome di H. Darcy, il quale lo studiò per primo nel 1856 nel caso di flusso attraverso la sabbia.
Il movimento delle acque profonde è necessario per il completamento del ciclo idrologico. L'acqua profonda, inoltre, non si presta soltanto a divenire altra parte del ciclo idrologico, ma rientra essa stessa in ciclo. Vicino alla falda freatica, il periodo medio del ciclo delle acque può essere di un anno o anche meno, ma in zone acquifere di profondità può diventare dell'ordine di migliaia d'anni. A grandi profondità, inoltre, si trovano delle acque fossili, di elevata concentrazione salina, con periodi di ciclazione dell'ordine delle ere geologiche, tali da escluderle dal ciclo idrologico attivo.
La quantità d'acqua totale, il suo bilancio, la sua ripartizione e il suo movimento nell'ambito dell'intero ciclo idrologico sono stati oggetto di ipotesi e di studi per più di un secolo. Ciò nondimeno, i dati quantitativi sono insufficienti per una conoscenza completa del ciclo idrologico, e si possono soltanto stabilire dei valori approssimati di stima per le sue varie componenti.
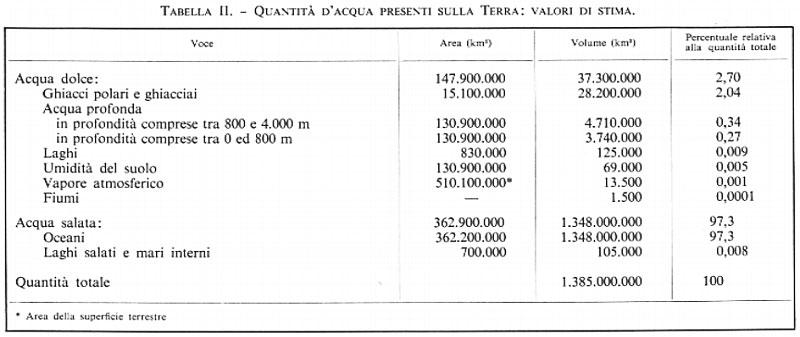
Nella tab. II sono riportati i valori di stima delle quantità d'acqua presenti sul globo (non sono state tenute in considerazione le acque di idratazione). Si può vedere da essa come gli oceani contengano circa il 97% di tutta l'acqua presente sul globo, cioè circa 1.350.000.000 di km3. Se la Terra fosse una sfera perfetta, tale quantità sarebbe sufficiente per ricoprirla con uno strato spesso 2.650 metri. La quantità d'acqua dolce si stima che sia circa 37.300.000 km3. Essa è distribuita pressappoco nel modo che segue: il 76% fa parte dei ghiacci polari e dei ghiacciai, il 13% è contenuto nel sottosuolo a profondità comprese tra gli 800 ed i 4.000 metri, il 10% nel sottosuolo a profondità inferiori agli 800 metri, lo 0,33% è contenuto nei laghi, lo 0,18% sotto forma di umidità del terreno, lo 0,036% si trova nell'atmosfera e una quantità trascurabile, pari allo 0,004%, si trova nei fiumi.
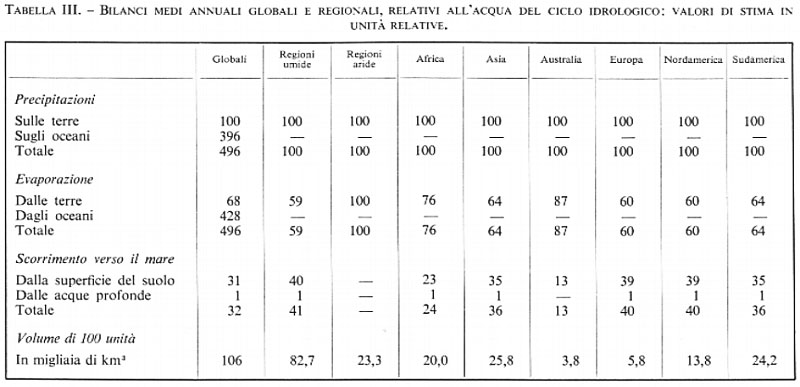
I valori dati delle quantità d'acqua sono valori stimati sulla base di un'ipotesi di distribuzione costante nel tempo. Mentre attraverso l'atmosfera e i fiumi passano ogni anno quantità enormi di acqua, essi ne contengono, ad ogni istante, delle quantità relativamente piccole. Nella tab. III sono riportati, in unità relative, i valori stimati del bilancio medio annuale complessivo dell'acqua. Nella tab. III sono riportati i valori relativi agli equilibri regionali dell'acqua, propri del ciclo idrologico, per zone umide e aride e pei diversi continenti. Si può notare che la precipitazione annuale media sulla Terra è circa 7,9 (=106.000/13.000) volte maggiore dell'acqua contenuta ad ogni istante sotto forma di vapore in tutta l'atmosfera; mentre il valore medio della precipitazione annuale sulla terraferma è circa 27 volte più grande del vapore acqueo nell'aria sovrastante i continenti. Così, il periodo del ciclo dell'umidità atmosferica è di circa 46 giorni per il ciclo idrologico computato nel suo complesso e soltanto di 13,5 giorni relativamente al ciclo che si svolge sui continenti. Inoltre, si può calcolare che la precipitazione annuale media sulla Terra sia di circa 104 centimetri all'anno: tale valore è bilanciato, in condizioni stazionarie, dall'evaporazione di un'eguale quantità d'acqua. Ciò equivale a dire che l'evaporazione media sulla Terra corrisponde a 2,84 millimetri d'acqua al giorno.
In linea di principio, il ciclo idrologico è semplice; in pratica, esso è enormemente complesso e intricato. Non è costituito da un unico grande ciclo, ma piuttosto da un gran numero di cicli, ristretti a continenti, regioni e zone locali, i quali, tutti, sono componenti collegate e interdipendenti del sistema globale. Anche se la quantità totale d'acqua è essenzialmente una costante del ciclo idrologico terrestre, la sua distribuzione varia continuamente sui continenti, sulle regioni e nell'ambito degli stessi bacini idrografici. Il comportamento idrologico dell'acqua in una certa zona è influenzato in maniera determinante dalle condizioni climatiche, le quali variano nel tempo e da regione a regione. Inoltre, la distribuzione dell'acqua in una certa area è influenzata da fattori naturali fisiografici, quali possono essere le caratteristiche topografiche, le formazioni geologiche e i vari tipi di vegetazione. Reciprocamente, i fattori fisiografici possono modificare quelli climatici, come l'intensità delle piogge, la loro distribuzione e frequenza, la formazione di neve e ghiaccio e gli effetti del vento, della temperatura, dell'umidità e della radiazione solare sulla traspirazione della vegetazione. A mano a mano che la società umana si evolve, ulteriori effetti sul regime delle acque dovuti alle opere ed alle attività dell'uomo si aggiungono prepotentemente alle condizioni ambientali dell'acqua e disturbano conseguentemente l'equilibrio dinamico del ciclo idrologico naturale, dando l'avvio a nuovi processi idrologici e a nuovi fenomeni.
c) Gli studi idrologici
Sin dalla prima antichità, e fin verso il 1400 d.C., il concetto di ciclo idrologico è stato argomento di speculazione da parte di molti pensatori fra cui Omero (1000 a.C. circa), Talete, Platone ed Aristotele in Grecia; Lucrezio, Seneca e Plinio a Roma.
Mentre la maggior parte dei concetti filosofici formulati in proposito è da considerare erronea dal punto di vista scientifico, Anassagora di Clazomene (500-428 a.C.) ideò una prima ipotesi sul ciclo idrologico complessivo; secondo tale ipotesi il Sole innalzerebbe l'acqua dal mare nell'atmosfera, donde ricadrebbe come pioggia: l'acqua piovana si raccoglierebbe poi in depositi sotterranei, i quali alimenterebbero il flusso dei fiumi. Un concetto più preciso lo formulò Teofrasto (371/370-288/287 a.C.), il quale descrisse con esattezza almeno la fase del ciclo idrologico relativo all'atmosfera, fornendo una corretta spiegazione della formazione delle precipitazioni, che si devono alla condensazione e al congelamento. Dopo un attento studio del lavoro di Teofrasto, Marco Vitruvio, trattatista romano vissuto all'incirca negli anni della vita del Cristo, completò l'interpretazione di Teofrasto ed affermò che le acque del sottosuolo derivano, per la maggior parte, dalle piogge e dalle nevi che si sono infiltrate nel terreno. Già da allora, quindi, si erano formate le basi essenziali della moderna interpretazione del ciclo idrologico.
Nel Rinascimento, si manifestò una leggera tendenza a spostare i concetti dell'idrologia dalla pura speculazione alla scienza sperimentale propria dei tempi moderni. Basandosi sui dati dell'osservazione, ad esempio, B. Palissy (1510-1589), P. Perrault (1608-1680) ed E. Mariotte (1620-1684) giunsero alla comprensione esatta dell'attuale concetto del ciclo idrologico. Palissy, notando come i fiumi e le sorgenti si originassero dalle piogge, confutò l'antica credenza che i corsi d'acqua fossero alimentati direttamente dal mare. Perrault misurò la quantità delle acque di scorrimento e trovò che esse non erano che una frazione delle acque piovane; dal che egli dedusse correttamente che, non essendo le piogge che una tra le sorgenti delle acque di scorrimento, le restanti acque di precipitazione dovevano andare perse per traspirazione, per evaporazione o per diversione. Mariotte precisò ulteriormente che esse si infiltrano nel terreno, e arricchiscono così le acque sotterranee.
Accanto all'aria, l'acqua è la sostanza più importante per la vita dell'uomo. Per superare i suoi problemi quotidiani e soddisfare la sua curiosità, l'uomo ha sempre cercato di approfondire la conoscenza dell'acqua presente nell'ambiente in cui vive. Sin da quando il concetto scientifico di ciclo idrologico è stato elaborato frammentariamente da uomini come Vitruvio, Palissy e Perrault, gli studiosi hanno cercato senza tregua di raggiungere una conoscenza complessiva del ciclo idrologico; il progresso, tuttavia, non è stato rapido, per le difficoltà e la complessità del fenomeno e la mancanza di un numero sufficiente ed adeguato di dati sperimentali. Attualmente, l'avvento dell'era atomica e spaziale ha portato nuove teorie e nuove conoscenze; oggi sono disponibili nuovi metodi e nuovi strumenti di indagine: i calcolatori elettronici, le tecniche nucleari e le tecnologie spaziali. Il progresso teorico più importante realizzato negli ultimi anni nel campo delle conoscenze idrologiche consiste in un'analisi del ciclo idrologico impostata su una base scientifica più rigorosa: si considera, cioè, il ciclo alla stregua di un sistema dinamico in evoluzione, costituito da un'‛entrata', da un'‛uscita' e da un mezzo di lavorazione, in questo caso l'acqua che attraversa il sistema. Lo studio di un intero ciclo idrologico o di una sua parte si può considerare come un problema del sistema o di un sottosistema. Un bacino idrografico può considerarsi, ad esempio, come un sistema, nel quale l'entrata è costituita dalle precipitazioni e dalle sorgenti d'acqua sotterranea, l'uscita dall'evaporazione, dalla traspirazione, dall'infiltrazione e dallo scorrimento e il mezzo di lavorazione dall'acqua che si muove attraverso il bacino. Secondo tale concetto, il ciclo idrologico può essere interpretato mediante le tecniche della moderna analisi dei sistemi e quindi simulato per mezzo di modelli matematici da risolversi con i calcolatori elettronici.
Poiché il ciclo idrologico interessa un'ampia regione dell'idrosfera e l'intera storia dell'acqua sulla Terra è tracciata nella continuità del ciclo, lo studio di questo coinvolge la fisica, la chimica, la biologia, la matematica ed altre scienze che si ricollegano ai vari processi del ciclo, quali la climatologia, la meteorologia, l'oceanologia, la geologia, la glaciologia, la geomorfologia, la limnologia e l'idraulica. Esso richiede, inoltre, la conoscenza di tutte le possibili correlazioni esistenti tra le acque dell'atmosfera, della superficie terrestre e del sottosuolo ed anche degli effetti provocati dai mutamenti che l'uomo ha apportato all'ambiente relativamente al regime delle acque. La conoscenza completa del ciclo idrologico diviene una necessità per lo sfruttamento e l'utilizzazione a fondo di tutte le risorse idriche disponibili sulla Terra.
Molti enti locali e nazionali ed organizzazioni internazionali, quali la Food and Agricultural Organization (FAO), la World Meteorological Organization (WMO) e la World Health Organization (WHO), sono attivamente impegnati in ricerche di idrologia, che hanno come tema centrale il ciclo idrologico. Il 1° gennaio 1965 l'UNESCO lanciò il programma del Decennio Internazionale di Idrologia, o IHD (dall'inglese International Hydrological Decade) cui aderirono più di cento paesi. L'IHD (1965-1974) è un vasto programma in cui le nazioni del mondo cooperano al fine di studiare le acque della Terra, essendo ormai dimostrato che, entro la fine di questo secolo, il progresso delle condizioni di vita dell'uomo potrà dipendere dalla sua capacità di risolvere i problemi connessi con l'acqua. È il primo tentativo concertato che l'uomo compie sia per fare l'inventario delle risorse idriche a sua disposizione, che vanno diminuendo a causa dell'aumento della popolazione del globo e del miglioramento delle condizioni di vita, sia per coordinare su scala mondiale gli studi sui metodi da adottare per fare il migliore uso possibile di queste risorse.
2. L'acqua nella società
a) L'acqua e le civiltà
È stato detto che si potrebbe scrivere la storia dello sviluppo dell'uomo attraverso il suo rapporto epico con l'acqua. Grandi civiltà del passato sono fiorite lungo i corsi dei fiumi, lungo il Nilo in Egitto (3000 a.C.), il Fiume Giallo in Cina (3000 a.C.), il Tigri-Eufrate in Mesopotamia (2400 a.C.) e l'Indo nell'India settentrionale (2500 a.C.). Le civiltà sono cadute quando sono venute a mancare le risorse d'acqua o queste sono state malamente amministrate.
Nelle antiche civiltà la maggior parte delle opere idrauliche era destinata a soddisfare alcune delle esigenze più immediate dell'uomo: quelle di dissetarsi e lavarsi. I qanat (o khanat) dell'Impero persiano e gli acquedotti dell'Impero romano sono famosi esempi di questo tipo di opere idrauliche. I qanat sono gallerie sotterranee che portano le acque del sottosuolo dai monti alle città. Ci sono oggi circa 22.000 qanat nella Persia, per un totale di oltre 270.000 km di canali sotterranei. Molti di questi furono costruiti 3.000 anni or sono. Essi forniscono ogni secondo una quantità di 550 m3 d'acqua, con i quali si provvede non solamente ai consumi domestici, ma anche all'irrigazione. Gli acquedotti della Roma imperiale fornivano un totale di circa un milione di metri cubi d'acqua giornalieri, 350.000 dei quali venivano utilizzati nella città stessa.
Nelle civiltà moderne, il tenore di vita può essere approssimativamente determinato sulla base del consumo individuale d'acqua. Il fabbisogno minimo per svolgere in modo opportuno tutte le funzioni vitali è di circa 80 litri d'acqua giornalieri a testa. Nei centri abitati, comunque, il consumo può raggiungere, oggi, i 400 litri d'acqua giornaliera pro capite e nelle zone metropolitane anche i 1.500 e più litri al giorno. Nel rifornimento cittadino assume dunque particolare importanza la qualità dell'acqua. Vari trattamenti sono in uso, quali la sedimentazione, la chiarificazione e l'infiltrazione, per liberare l'acqua da materiali estranei. Si suole poi aggiungere il cloro per impedire la crescita delle alghe ed uccidere i germi. Una parte di fluoro su un milione ha, inoltre, la proprietà di prevenire la carie dentaria. Per ridurre il grado di durezza, che è causa, fra l'altro, di sprechi di sapone e d'altri detersivi, si usano spesso agenti di addolcimento del tipo della zeolite.
Il bisogno d'acqua per l'irrigazione fu avvertito sin dai primi stadi della civilizzazione, in concomitanza con le necessità agricole della produzione alimentare. Un sistema organizzato d'irrigazione dei campi si era già sviluppato nelle pianure dell'Anatolia nel 4000 a.C. e nella valle del Nilo nel 3400 a.C. Le canalizzazioni di Tukiangyien, costruite in Cina 2.200 anni fa, irrigavano più di 200.000 ettari di terreno fertile. Come sistemi d'irrigazione moderni su vasta scala, basti qui ricordare il sistema del bacino dell'Indo nel Pakistan occidentale e in India, che porta acqua a più di nove milioni di ettari di terra e il sistema dell'Imperial Valley negli Stati Uniti, che irriga più di 200.000 ettari di terra. L'irrigazione consiste nel rifornire d'acqua un terreno allo scopo precipuo di far crescere le messi in climi secchi o in climi umidi che abbiano prolungati periodi di siccità. Si fa uso di irrigazione suppletiva anche in regioni umide che presentino precipitazioni moderate, allo scopo di ottenere colture più abbondanti o particolari. Per un'efficace irrigazione, non basta disporre di una quantità d'acqua sufficiente per il consumo, dovuto essenzialmente all'evaporazione e alla traspirazione delle piante, ma occorre che vi sia anche un movimento delle acque sotterranee e di drenaggio sufficiente a rimuovere i sali dalle aree irrigate. Ove il drenaggio non sia adeguato, i raccolti possono andare perduti a causa di un eccesso di salinità o di acqua nel suolo.
Col crescere della popolazione, la gente emigra e costruisce le città in zone alluvionali, per necessità economiche, per la vicinanza alle zone dei trasporti idrici o per altre esigenze. Una pianura alluvionale è un territorio formato da un fiume durante i periodi delle piene alluvionali. Successivamente, il fiume sorpassa di nuovo i propri argini e inonda le pianure alluvionali. Piccoli allagamenti sono un evento normalmente ricorrente nel caso di moltissimi fiumi e non provocano inondazioni preoccupanti. Allagamenti più estesi sono meno frequenti ma, quando capitano, possono produrre danni assai più ingenti. Le antiche civiltà di Mohenjo-Daro ed Harappa (2500-1500 a.C.), sviluppatesi sulle pianure dell'Indo, sembra che siano state distrutte da tali tipi di inondazioni ricorrenti. Il fatto che le pianure alluvionali siano densamente abitate implica ovviamente, in caso di alluvioni, danni molto gravi alle proprietà e, talvolta, perdita di molte vite umane. Questo problema non può essere risolto neppure con i mezzi oggi a disposizione, perché le inondazioni di portata maggiore non si possono evitare, ma soltanto contenere. Nonostante abbiano provocato rilevanti perdite, purtuttavia le inondazioni non hanno impedito all'uomo di stabilirsi sulle pianure alluvionali. Al contrario, è piuttosto evidente che la popolosità delle zone alluvionali è in continuo aumento in molte parti della Terra, con un ritmo di crescita più rapido rispetto a quello medio globale.
Gli sforzi che oggi si compiono per ridurre le perdite dovute agli allagamenti si possono classificare secondo i tipi di soluzioni seguenti: a) ricorrere a piani d'emergenza, come l'evacuazione di persone e cose dalla zona disastrata, e adottare misure di protezione temporanee, ammucchiando sacchi di sabbia e trasferendo in luoghi più alti le cose e le attrezzature che potrebbero essere danneggiate (lotta contro l'alluvione), oppure riprogrammare le proprie attività; b) modificare le strutture delle abitazioni in maniera da respingere le acque d'allagamento; c) adottare certe misure, come l'installazione di pompe di drenaggio e la costruzione di fondazioni impermeabili all'acqua (antialluvionali); d) regolare l'uso del territorio (suddivisione in zone delle pianure alluvionali); e) controllare le alluvioni mediante modifiche delle attività agricole (diminuzione degli effetti alluvionali) e costruzione di opere antiallagamento, come le dighe, gli argini e i bacini (protezione dalle alluvioni). Ciascuno di questi rimedi presenta vantaggi e svantaggi. La soluzione migliore è solitamente quella di combinare fra loro alcuni di essi.
I fiumi, i laghi e i mari costituiscono vie naturali di comunicazione; le costruzioni di canali per la navigazione ebbero inizio ai primordi della storia. Il Grande canale della Cina, completato nel sec. XIII, determinò il diffondersi della civiltà dalle regioni settentrionali a quelle del meridione. Nei secoli successivi, grande impulso alla navigazione fu dato dall'invenzione delle chiuse e dei motori a vapore e diesel delle imbarcazioni. Si aprirono i grandi canali di Suez, di Panama e del lago Erie.
Negli ultimi anni, le comunicazioni per via d'acqua hanno fatto grandi progressi grazie all'introduzione degli aliscafi e delle imbarcazioni a cuscino d'aria.
L'energia idrica contribuisce in maniera tanto più rilevante al progresso della civiltà moderna, quanto più intenso si fa il consumo di energia elettrica nelle città, nelle campagne e nelle industrie. I primi impianti idroelettrici furono costruiti in quelle regioni nelle quali più favorevoli erano le condizioni per la loro costruzione ed utilizzazione a basso costo. Oggi, però, il rendimento degli impianti a vapore, o termoelettrici, per la produzione di elettricità è divenuto ben maggiore di quello degli impianti idroelettrici. Attualmente è più economico produrre energia elettrica mediante un impianto a vapore piuttosto che costruirne uno idroelettrico, fatta eccezione ovviamente per quei casi in cui vi sia abbondanza di energia idrica e in pari tempo scarseggi o sia troppo costoso il combustibile per la produzione del vapore. Nel futuro, si potrà abbassare il costo degli impianti nucleari fino ad eguagliarlo a quello degli impianti a vapore. In questi casi, un impianto idroelettrico può in generale non essere economicamente competitivo se deve essere impiegato come sorgente costante di energia; ma, poiché la produzione idroelettrica di energia può essere regolata assai più facilmente sia di quella termoelettrica sia di quella nucleare, l'impianto idroelettrico dovrebbe conservare ancora la sua importanza economica per far fronte al fabbisogno di energia durante le ore di punta.
In molti tra i paesi più progrediti, si possono distinguere sei problemi connessi con l'acqua: il rifornimento, la variabilità, la distribuzione, gli allagamenti, la qualità (composizione chimica e sedimentazione) e l'inquinamento. Il rifornimento diviene un problema ogniqualvolta la richiesta superi la disponibilità delle risorse. Dallo studio del ciclo idrologico si è visto che la sorgente originale dell'acqua sulla Terra è costituita essenzialmente dalle precipitazioni. Dal momento che queste sono diversamente distribuite a seconda della località geografica, è ovvio che anche la disponibilità ne viene conseguentemente a dipendere. La distribuzione delle acque di scorrimento è simile a quella delle precipitazioni, ma è ulteriormente condizionata dalle formazioni geologiche, per il fatto che parte della precipitazione può infiltrarsi nel sottosuolo. La disponibilità dell'acqua derivante dalle acque di scorrimento superficiale e del sottosuolo varia con il consumo, che può essere maggiore in un luogo piuttosto che in un altro. Di conseguenza, la domanda d'acqua in una regione può essere non soddisfatta dalla disponibilità e la distribuzione diventare perciò un problema, anche se in altre parti della regione o in regioni vicine vi può essere sovrabbondanza di acqua. Una tale situazione si determina quando disponibilità e domanda non coincidono geograficamente. In certi luoghi, inoltre, anche se la disponibilità media dell'acqua è sufficiente per far fronte alla domanda media, pur tuttavia sorge il problema della variabilità se la richiesta, in certi momenti, supera la disponibilità. E sebbene la disponibilità sia soggetta alle variazioni temporali più della domanda, la variabilità di entrambe contribuisce a complicare il problema. Come si è detto precedentemente, le alluvioni sono conseguenza di un'eccessiva quantità d'acqua nel luogo non opportuno in un momento inopportuno e costituiscono pertanto un problema connesso con quello della distribuzione e della variabilità in molte regioni di molti paesi.
Fra tutti i problemi idrici che la società moderna si trova a dover affrontare, quelli connessi con la qualità delle acque si sono dimostrati i più complessi. Innanzitutto è difficile distinguere un'acqua naturalmente non buona, per sostanze chimiche naturali e per depositi, da un'acqua resa cattiva dall'inquinamento prodotto dall'uomo. Le sorgenti naturali, per ciò che si riferisce alla loro qualità, sono oltremodo variabili nel tempo e diverse a seconda della disposizione geologica. Per stabilire dei criteri di qualità delle acque naturali, occorre determinare la concentrazione e il carico dei materiali disciolti e del sedimento. I requisiti dell'acqua dipendono dall'uso che ne vien fatto e variano a seconda che essa venga impiegata per la casa, per le industrie o per irrigare i campi. A rendere ancora più complicato il problema della qualità, in talune zone l'inquinamento dovuto alle attività umane ha deteriorato le caratteristiche delle riserve idriche. Fattori di vario genere hanno contribuito a creare tale situazione. In seguito alla rapidissima crescita della popolazione avvenuta in molte parti della Terra negli ultimi decenni, l'eliminazione dei rifiuti è diventata un problema. Inoltre, gli sviluppi delle nuove tecnologie hanno introdotto nell'ambiente dell'uomo e nell'ecologia nuovi tipi di rifiuti industriali e d'altra origine, per esempio i prodotti chimici organici di sintesi, i detersivi e i materiali radioattivi. Nei tempi passati, l'acqua ha avuto un'importanza primaria nella formazione delle civiltà, mentre la civiltà d'oggi crea nuovi problemi per l'acqua.
b) Lo sviluppo delle riserve d'acqua
Nei tempi antichi, come ai nostri giorni, l'uomo ha sempre dovuto affrontare il problema di pianificare e controllare l'uso delle riserve idriche. L'impegno d'oggi è però ancor più grande di quello di una volta per via della rapida crescita della popolazione, della proliferazione industriale, dell'inurbamento e della produzione di rifiuti. Per di più le riserve idriche sono state male amministrate. Le riserve sotterranee sono state spesso sfruttate indiscriminatamente: alcune sono state depauperate temporaneamente, altre hanno subito danni permanenti. Un accumulo incontrollato dei rifiuti in zone acquitrinose inquina molte sorgenti acquifere e le rende inutilizzabili per molti scopi. In molte altre maniere l'uomo sconvolge i delicati equilibri naturali delle acque del ciclo idrologico. Per nostra buona sorte, le riserve idriche si ricostituiscono in maniera continua. Se le si protegge bene e le si amministra saggiamente, vi sarà più acqua a disposizione di quanta non sembri esservene in molti luoghi d'apparente carenza idrica.
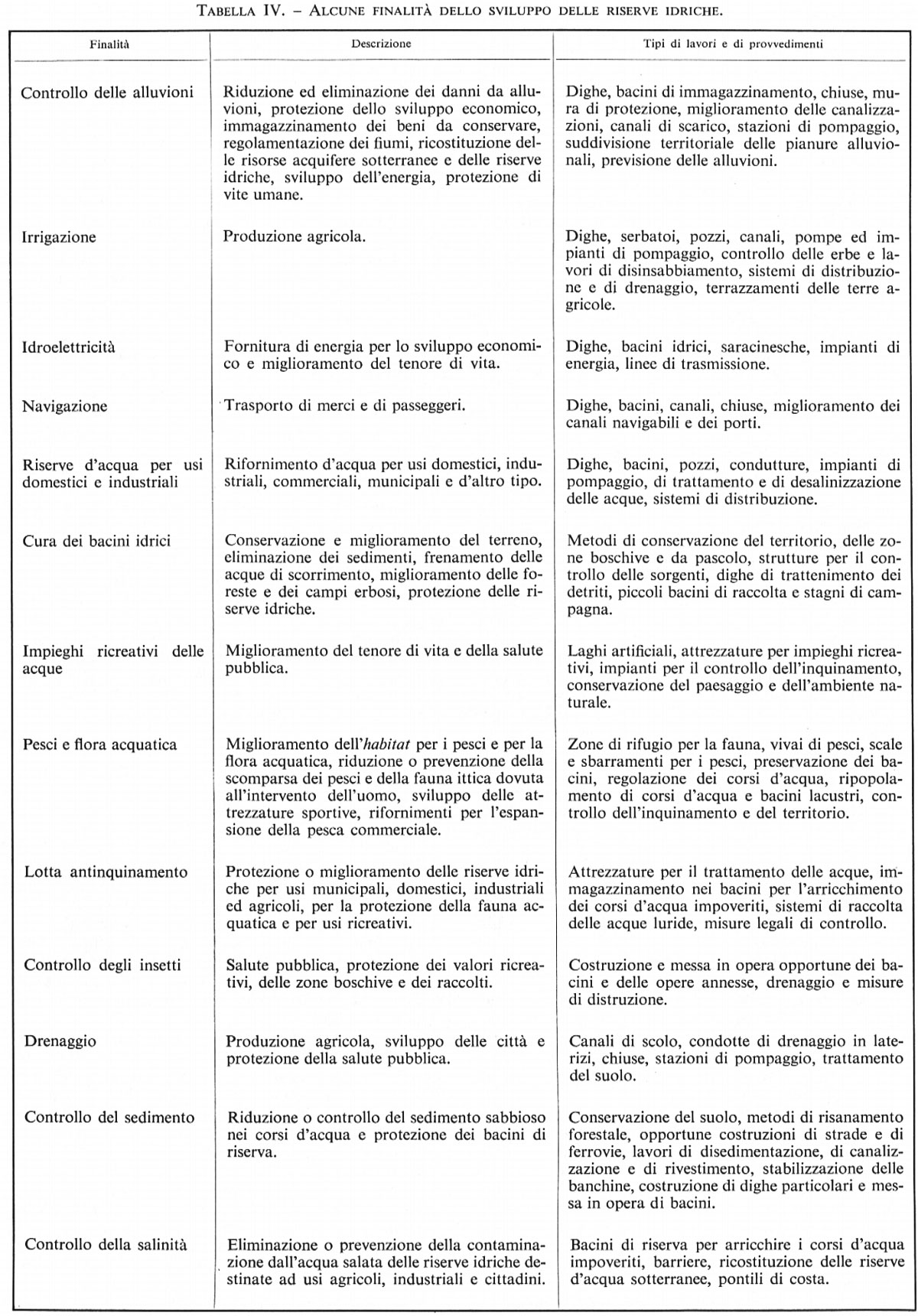
Lo sviluppo attuale delle risorse idriche è spesso programmato per conseguire più d'uno degli scopi tra quelli indicati nella tab. IV, poiché solitamente lo sviluppo a finalità multiple ha una giustificazione economica assai maggiore. Lo sviluppo delle riserve idriche può riguardare un certo numero di unità strutturali, del tipo delle dighe o dei serbatoi, e può al tempo stesso essere studiato per diversi tipi d'impiego. Uno sviluppo moderno delle riserve idriche può pertanto divenire molto complesso, se deve avere finalità multiple, essere a più unità e prestarsi a diversi tipi d'impiego.
Lo sviluppo delle risorse idriche, assai complesso tecnologicamente, deve tener conto delle necessità e dei problemi della regione in cui si attua. Queste necessità e questi problemi si pongono a livello internazionale e richiedono, per la loro soluzione, la cooperazione e il coordinamento di interessi pubblici e privati a livelli diversi. Oltre al conseguimento dell'efficienza economica, vanno tenuti di mira altri obiettivi, quali il miglioramento dell'ambiente e del benessere pubblico e il rafforzamento dello sviluppo regionale. Gran parte di questi obiettivi non è facilmente valutabile, dal momento che sfugge ad ogni criterio di quantificazione né si può esprimere in unità monetarie, come viceversa si richiede quando si vogliano fare, in economia, delle valutazioni matematiche. La parte non valutabile di questi obiettivi dev'essere giudicata da chi deve prendere le decisioni, in seguito a una stima o ad altro giudizio non matematico, sulla base di un'ampia informazione.
I moderni progetti riguardanti le riserve idriche possono presentarsi così complessi e complicati che anche i loro aspetti più concreti non sono facili da valutare con l'analisi economica convenzionale. Per risolvere questi problemi, negli ultimi decenni, ci si è serviti delle tecniche della ricerca operazionale e dell'analisi dei sistemi. Secondo tali tecniche, un progetto relativo alle riserve idriche viene trattato alla stregua di un sistema. Matematicamente, il sistema viene rappresentato da una ‛funzione obiettivo', che esprime gli obiettivi concreti del sistema. La funzione obiettivo viene quindi ottimizzata, sottoposta a tutte quelle condizioni che si possono esprimere in forma analitica. In generale, questi tipi di analisi si possono affrontare secondo due metodi distinti. Il primo è noto col nome di ‛simulazione matematica': si simula il sistema in un calcolatore elettronico e in conseguenza si sceglie la combinazione migliore delle variabili del sistema sulla base della risposta che il sistema simulato offre a varie combinazioni alternative. L'altro, cosiddetto a ‛modello matematico', utilizza un modello matematico semplificato che può essere direttamente risolto per la pianificazione migliore del progetto. Naturalmente, la pianificazione alla quale in tale maniera si perviene non è sempre quella definitiva, dal momento che questa viene scelta dopo aver considerato quegli altri obiettivi e quelle altre condizioni che, per loro natura, non sono valutabili analiticamente.
I progressi compiuti coi calcolatori hanno permesso di svolgere delle operazioni di computo con le quali si risolvono formulazioni matematiche, non altrimenti risolubili, relative a sistemi di riserve idriche di grande complessità. Per questi scopi si fa spesso uso dei calcolatori numerici. Attualmente, si sta studiando la possibilità di impiegare congiuntamente i calcolatori analogici e quelli numerici (calcolatori ibridi), per l'esecuzione di calcoli più veloci e più sicuri. Tuttavia, l'impiego dell'analisi dei sistemi per lo sviluppo e l'amministrazione delle riserve idriche è ancora in fase di studio, poiché le relative tecniche debbono ancor essere migliorate per aver pratica applicazione.
3. L'acqua nel futuro
a) Modifiche del ciclo idrologico
Per il futuro l'uomo ha molte possibilità di sfruttare le risorse idriche per la risoluzione dei suoi problemi relativi all'acqua. S'è fatto un certo numero di tentativi per modificare il naturale svolgersi del ciclo idrologico, senza però sbilanciarne gli equilibri e proyocare gravi conseguenze. Questi tentativi potranno essere un giorno utilmente impiegati su vasta scala per il bene dell'umanità. Vale qui la pena di ricordarne alcuni: il controllo delle precipitazioni, l'eliminazione dell'evaporazione e il rifornimento delle acque sotterranee.
È possibile aumentare artificialmente la quantità dell'acqua di precipitazione disperdendo tra le nuvole del materiale che provochi la cosiddetta ‛enucleazione', o formazione di nuclei (condensazione, deposizione di ghiaccio o congelamento) del vapore acqueo nell'atmosfera. La caduta di neve e di pioggia può essere provocata dalla presenza di piccoli cristalli di ghiaccio in nuvole sottoraffreddate (‛teoria di Wegener-Findeisen-Bergeron') o dalla presenza di goccioline d'acqua relativamente grosse (‛processo di coalescenza'). Certe nuvole danno luogo a scarse precipitazioni, o non ne danno affatto, perché mancano di questi componenti. Si può ovviare a questa deficienza seminando artificialmente le nuvole con ghiaccio secco o con ioduro d'argento per provocare la formazione di cristalli di ghiaccio, oppure introducendo gocce d'acqua o grossi nuclei igroscopici. Questa semina ha efficacia solamente in condizioni particolari di rilievo orografico e quando sia fatta in zone temporalesche, dove i nuclei non sono sufficientemente numerosi per provocare la pioggia secondo il processo naturale.
Oltre alla formazione delle piogge, si è dimostrato utile, al fine di dissipare le nebbie raggelate sugli aeroporti, disperdere nell'aria goccioline di propano gassoso, brina o ghiaccio secco, che provocano la caduta della neve e liberano l'atmosfera. Sono attualmente allo studio progetti più ambiziosi coi quali si vogliono eliminare i cicloni e disperdere l'enorme energia in essi concentrata, seminando le nuvole o persino provocando delle esplosioni nucleari. La violenza dei cicloni potrebbe anche essere notevolmente ridotta se si cospargesse di materiali oleosi la superficie del mare davanti ad essi, per eliminare l'evaporazione superficiale e il conseguente rifornimento di energia. Forse l'uomo è sul punto di apportare sensibili modifiche al ciclo idrologico in vaste zone, estese per centinaia di chilometri quadrati ed anche più. Questi tentativi di modificare su vasta scala i delicati equilibri del ciclo idrologico globale possono avere pericolose conseguenze; vanno perciò rinviati sino a quando non si sarà in grado di predirne con una certa ragionevolezza i possibili effetti.
Su scala locale, più ristretta, l'eliminazione dell'evaporazione e la ricostituzione delle riserve d'acqua sotterranea si sono dimostrati metodi di grande efficacia in molti casi; è perciò probabile che in futuro saranno largamente impiegati.
Si sono studiati vari metodi per eliminare l'evaporazione e conservare le quantità d'acqua disponibili. La pratica corrente consiste nel disperdere sulla superficie dell'acqua uno strato monomolecolare che ostacoli la fuoriuscita delle molecole dall'acqua, ritardando così l'evaporazione. Sebbene le ricerche chimiche per un tale metodo abbiano avuto inizio più di cinquant'anni or sono nei laboratori degli Stati Uniti, fu solo nel 1952 che questa idea trovò per la prima volta pratica applicazione in alcuni bacini idrici dell'Australia, ad opera essenzialmente di W.W. Mansfield. Diversi tipi di composti chimici sono capaci di disporsi in forma di strato monomolecolare sulla superficie dell'acqua. I composti organici migliori per questo scopo sono alcoli alifatici a lunga catena molecolare come l'‛ottadecanolo' e l'‛esadecanolo'. La molecola di questi composti possiede un gruppo OH ad un'estremità, il quale è attratto dall'acqua, mentre la catena degli idrocarburi all'estremità opposta è respinta, così che le molecole si orientano normalmente alla superficie dell'acqua e formano una pellicola aderente a questa ma penetrabile dalle molecole di ossigeno e di diossido di carbonio. Poiché la pellicola monomolecolare può essere facilmente rotta dai movimenti della superficie dell'acqua, il metodo perde di efficacia se viene applicato a vaste distese d'acqua. Teoricamente, un chilogrammo di esadecanolo forma una pellicola compatta di 4.500 metri quadrati; in pratica, ne occorrerà molto di più, perché viene eliminato parzialmente dal vento, dagli uccelli, dagli insetti e dalla fauna acquatica. Inoltre, poiché il composto chimico è biodegradabile, esso andrà di tanto in tanto rinnovato. La riduzione dell'evaporazione ottenibile con questo metodo può variare all'incirca dal 10 al 40% e oltre, a seconda delle condizioni.
Vari sono i metodi per la ricostituzione delle acque sotterranee. Il primo metodo, in uso nella California sin dal 1890, è quello della ‛diffusione dei canali', mediante il quale si fa deviare l'acqua superficiale in eccesso in fossi aperti, sì da aumentarne la rapidità d'infiltrazione. Questo metodo, tuttavia, è stato per lo più rimpiazzato da quello detto della ‛diffusione dei bacini'. È questo il sistema più efficace per la rigenerazione delle riserve, dal momento che si presta ad una diffusione prolungata di grandi quantità d'acqua. Nella contea di Los Angeles, in California, la velocità di infiltrazione combinata di una ventina di zone di diffusione è di circa 60.000 litri al secondo. Anche la ricarica per mezzo di pozzi ghiaiosi in disuso o appositamente costruiti si è dimostrata efficace per progetti su scala limitata, specialmente dove siano disponibili delle acque che non abbiano sedimenti naturali. Nei casi in cui una formazione di acque sotterranee sia isolata o non abbia comunque uscita agevole in superficie per la presenza di strati superficiali impermeabili, si può adottare il sistema di ricarica mediante pozzi o fosse d'infiltrazione.
La ricarica di bacini idrici per mezzo dei pozzi può anche costituire una barriera d'acqua fresca contro l'intrusione di acqua salata del mare e proteggere così le riserve d'acqua sotterranea vicine alle zone costiere.
b) Trasporto dell'acqua su vasta scala
Nella seconda metà del sec. XX si sono estese le tecniche riguardanti il trasporto dell'acqua su vasta scala, da un bacino idrografico o da una regione all'altra con la conseguenza di ottenere effetti determinanti, in particolare sullo sviluppo sociale ed economico della regione ricevente. L'acquedotto del California Water Plan, lungo 700 chilometri, porta verso le zone centrali e meridionali le acque della ricca regione del delta del Sacramento-San Joaquin, al nord. Ancora più ambizioso è il programma di trasportare l'acqua da uno Stato all'altro, oppure di farle superare la cresta spartiacque, come è per il caso delle acque del fiume Colorado verso Los Angeles.
Col modernissimo Texas Water Plan, circa 15.000 milioni di metri cubi d'acqua all'anno saranno prelevati dal Mississippi, nella bassa Luisiana. Circa 9.000 milioni di metri cubi di quest'acqua scorreranno per una lunghezza di 800 chilometri tra acquedotti e bacini e saranno trasportati per l'irrigazione sino a 1.000 m d'altezza sugli altipiani del Texas occidentale. L'acqua che risultasse eventualmente in eccesso sarà inviata nel Nuovo Messico.
Alcuni progetti per il trasporto su vasta scala dell'acqua possono forse sembrare sogni da fantascienza. Il programma NAWAPA (North American Water and Power Alliance) prevede il controllo di tutti i più grandi fiumi della zona occidentale del continente nordamericano, dallo Yukon, in Alasca, al Messico settentrionale. Arginando con dighe le prime anse dei fiumi Columbia, Kootenay e Fraser, si creerebbe sulle Montagne Rocciose un serbatoio lungo 800 chilometri che si estenderebbe dal Montana alla Columbia Britannica attraverso i confini del Canada. Il fondo di tale bacino di riserva si troverebbe a 900 m d'altezza sul livello del mare; di qui l'acqua verrebbe distribuita al sud, per mezzo di acquedotti, sino alle aride zone occidentali degli Stati Uniti ed al Messico. Accanto a questo bacino gigantesco vi dovrebbe essere una catena di serbatoi sviluppantesi dall'Alasca alla Columbia Britannica, dai quali si potrebbe pompare dell'acqua per alimentare un sistema di navigazione e per arricchire d'acqua le praterie canadesi, l'alto Mississippi ed i Grandi Laghi. Il sistema dovrebbe, infine, ricollegarsi con progetti analoghi nel Labrador, nel Quebec, nell'Ontario e nella zona settentrionale dello Stato di New York.
Paragonabile al programma NAWAPA per vastità di estensione è il progetto di deviazione dell'Ob-Jenisej-Irtyš. Con tale progetto si porterebbe l'acqua dei fiumi che nella Siberia occidentale scorrono verso il nord ai deserti della zona del Caspio, mediante la creazione di un mare artificiale in Asia, con una superficie avente un'area più grande dell'Inghilterra. Secondo un altro progetto, noto col nome di ‛progetto Sergei', si dovrebbe allagare il 10% circa del continente africano e si potrebbe un giorno forse irrigare l'arido deserto del Sahara.
c) Trattamento delle acque
Se la quantità di sostanze solide disciolte nell'acqua eccede 1.000 ppm, l'acqua viene classificata come ‛acqua salata'. Le acque salmastre contengono da 1.000 a 10.000 ppm. La quantità media nelle acque del mare è di circa 35.000 ppm. Per l'addolcimento delle acque salmastre si impiegano solitamente metodi di ‛elettrodialisi' e di ‛scambio di ioni'. Per dissalare l'acqua del mare vi sono vari processi. Quelli più largamente in uso si basano sull'evaporazione o la distillazione dell'acqua, secondo un adattamento tecnico dello stesso processo naturale con cui si ottiene l'acqua dolce dal mare. Attualmente, la quasi totalità delle acque dolci ricavate dal mare si ottiene mediante l'impiego di tre distinte versioni del metodo.
Il sistema maggiormente in uso, quanto a numero di impianti, è quello ad ‛evaporazione rapida a più stadi'. L'acqua marina, riscaldata a circa 120 °C, passa in una camera pressurizzata ad una pressione di poco inferiore a quella del punto di ebollizione dell'acqua. A questa pressione ridotta, parte dell'acqua evapora rapidamente e si condensa quindi per dare acqua dolce. Una versione raffinata dell'evaporazione rapida è quella cosiddetta ‛a tubi lunghi', nella quale l'evaporazione ha luogo in lunghi tubi anziché entro camere a vuoto. Un'altra versione, detta ‛distillazione a compressione di vapore', è stata largamente adottata per applicazioni su piccola scala. In questo processo, il vapore ottenuto dall'acqua di mare viene compresso e riportato nell'evaporatore, dove condensa per formare acqua dolce a una nuova temperatura più elevata; nel fare questo, produce calore sufficiente a far bollire una ulteriore quantità d'acqua di mare.
Altri procedimenti di dissalazione di grande interesse potenziale sono il congelamento sotto vuoto, l'osmosi inversa e la conversione per via chimica. Col procedimento di ‛congelamento sotto vuoto', l'acqua di mare raffreddata viene immessa in una camera di congelamento mantenuta a pressione ridotta. Parte dell'acqua evapora, assorbendo calore dalla parte restante. Poiché la temperatura è già vicina al punto di congelamento, si formano dei cristalli di ghiaccio, che costituiscono uno strato di brina, da cui si può ottenere acqua dolce. Nel procedimento per ‛osmosi inversa', l'acqua marina è posta ad una pressione superiore a quella osmotica sua propria; l'acqua dolce filtra allora attraverso una membrana osmotica. Anche la ‛conversione per via chimica' può assumere grande importanza nei processi atti ad ottenere acqua dolce dal mare. Il procedimento più noto è quello che va sotto il nome di ‛scambio ionico', il quale sembra ben promettere per il trattamento delle acque salmastre. Un altro nuovo procedimento assai promettente è quello di ‛estrazione per assorbimento', col quale si estrae l'acqua pura dal mare mediante un solvente organico.
Il costo di questi procedimenti sarà nel futuro il punto chiave per la dissalazione delle acque. Al momento attuale, i costi sono così elevati che la dissalazione viene limitata a quei casi in cui il costo dell'acqua ottenuta da altre fonti sia enormemente alto o in cui l'acqua dolce non sia praticamente ottenibile. La dissalazione troverà comunque impiego frequente per il rifornimento d'acqua quando i suoi costi si abbasseranno, in seguito al miglioramento della tecnologia.
Un altro modo importante e assai pratico per il trattamento delle acque consiste nel rinnovamento e nella riutilizzazione delle acque di scarico cittadine. Nei prossimi decenni, esso potrà diventare di uso corrente in molte zone.
Se si guarda al futuro, si può dire che gran parte della umanità sta muovendosi da un'economia di abbondanza d'acqua ad una di carenza. Per buona sorte, l'acqua è una sostanza indistruttibile e una risorsa che non può diminuire. La carenza d'acqua può manifestarsi in una certa regione geografica o si potrà avvertire un deterioramento qualitativo dell'acqua, ma la sua disponibilità globale non muterà nel tempo. Questa carenza è determinata largamente dal cattivo uso e dalla cattiva amministrazione delle riserve. Nel futuro, certamente, la scienza e la tecnologia contribuiranno in modo sempre più determinante alle soluzioni tecniche da dare a questi problemi, nei confronti dei quali sarà inoltre necessario mutare l'atteggiamento psicologico, sociale, economico e anche politico. Per conservare l'acqua, l'uomo dovrà vivere in futuro in modo diverso: dovrà conservare e riutilizzare la quantità d'acqua disponibile e non dovrà sprecare né inquinare le riserve. È anche importante convincere la gente che dare all'acqua un prezzo realistico aiuta a conservarne le riserve. Nella società contemporanea, l'acqua, in sostanza, è un prodotto: dovrebbe perciò essere considerata alla stregua di un manufatto che richiede manutenzione ed investimento di capitali per l'espansione e il miglioramento. Essa è anche un capitale preso in prestito che dev'essere restituito in condizioni tali da poter essere riutilizzato. Se non ci si convince di questa verità, forse veramente un giorno l'acqua verrà a scarseggiare.
bibliografia
Biswas, A. K., History of hydrology, Amsterdam-London 1970.
Chorley, R. J., Water, earth and man, London 1969.
Chow, V. T. (a cura di), Handbook of applied hydrology, New York 1964.
Davis, K. S., Day, J. A., Water. The mirror of science, New York 1961.
Eisenberg, D., Kauzmann, W., The structure and properties of water, New York-Oxford 1969.
King, T., Water. Miracle of nature, New York-London 1966.
Kuenen, P. H., Realms of water, New York 1963.
Rouse, H., Ince, S., History of hydraulics, New York 1963.
Vallentine, H. R., Water in the service of man, Baltimore 1967.
Water, in The yearbook of agriculture (a cura dell'U.S. Department of Agriculture), Washington 1955.
Acqua e vita di John W. Boylan, Kurt Kramer
sommario: 1. Introduzione. 2. Le proprietà dell'acqua e il loro significato biologico. 3. Assunzione, conservazione e perdita di acqua. a) Bilancio idrico giornaliero nell'uomo. b) La conservazione dell'acqua da parte del rene. c) La concentrazione dell'urina: un mezzo per la conservazione dell'acqua. 4. I compartimenti dei liquidi corporei. a) Misura dei compartimenti. b) Composizione dei liquidi corporei e loro contenuto ionico. c) Regolazione del volume dei liquidi corporei. 5. Sopravvivenza in assenza di acqua. a) Disturbi da disidratazione. b) Sopravvivenza in mare. c) Trattamento dell'acqua di mare per renderla potabile. d) L'acqua nello spazio. 6. Conclusione. □ Bibliografia.
1. Introduzione
L'acqua svolge un ruolo fondamentale fra le sostanze che compongono la materia vivente. I materiali di cui gli organismi viventi sono costituiti sono sospesi in acqua, racchiusi all'interno di una membrana vitale e riforniti attraverso un ambiente acquoso. Tutti gli scambi da e verso la cellula, l'unità strutturale della materia vivente, hanno luogo in soluzione acquosa, ed all'interno della cellula i processi energetici vitali avvengono tra elementi idrosolubili. La storia dell'acqua sulla Terra, la sua struttura e le proprietà che la rendono adatta a questo ruolo preminente costituiscono uno degli argomenti più affascinanti non solo della fisica, ma anche della biochimica.
Gli elementi che troviamo ora sulla Terra erano presenti al momento della creazione allo stato atomico, ed è probabile che la maggior parte dell'acqua che ricopre il nostro pianeta si sia formata improvvisamente, in una data epoca geologica, dalla ossidazione di grandi masse di idrogeno gassoso.
Mentre la massa fusa della Terra lentamente solidifica- va, il nostro pianeta doveva essere circondato da una nuvola impenetrabile di vapore acqueo. Ad un osservatore che dallo spazio avesse guardato la luce del sole riflessa da questa bianca capsula, il nuovo pianeta sarebbe apparso più splendente di Venere.
Con il raffreddamento dell'atmosfera il vapore cominciò a condensare e a cadere sulla Terra infuocata per evaporare di nuovo: processo che ha notevolmente accelerato il raffreddamento della superficie terrestre. Quando la temperatura della crosta terrestre scese al di sotto del suo punto di ebollizione, l'acqua cominciò a cadere sulla Terra formando i mari e modificando continuamente l'aspetto del suolo.
La primitiva atmosfera era ricca di diossido di carbonio, un gas poco solubile in acqua, con la quale forma acido carbonico. Quest'ultimo, benché presente solo in tracce, aumentò fortemente il potere solvente dell'acqua e facilitò il dissolversi dei minerali della superficie terrestre, i quali vennero dilavati in enormi quantità verso gli oceani.
Ebbe così inizio il grande ciclo dell'acqua, che prosegue senza interruzione fin dal primo diluvio. Ogni giorno il calore del sole solleva più di 5•1014 litri d'acqua verso l'atmosfera: una quantità incredibile ma che interessa solo un decimilionesimo di tutto il volume d'acqua della Terra. La maggior parte di quest'acqua ritorna sulla Terra come pioggia o neve durante lo stesso giorno in cui è evaporata. Ogni giorno un volume considerevole d'acqua entra inoltre a far parte del mondo vivente per partecipare a qualche processo vitale.
2. Le proprietà dell'acqua e il loro significato biologico
Le particolari proprietà chimico-fisiche dell'acqua (v. tab. I) la rendono l'ambiente più adatto per la vita fra tutte le sostanze conosciute. Ciò per le eccezionali caratteristiche fisiche legate in gran parte alla struttura della molecola dell'acqua e alla sua capacità di formare ‛legami a idrogeno'.
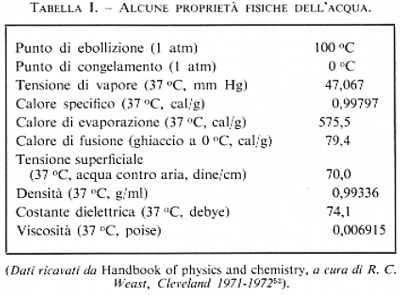
Alla fine del sec. XVIII Cavendish in Inghilterra e Lavoisier in Francia determinarono la composizione dell'acqua come idrogeno ed ossigeno. All'inizio del sec. XIX Gay-Lussac e Humboldt, indipendentemente l'uno dall'altro, scoprirono che un volume di ossigeno si combinava con due volumi di idrogeno. Sappiamo ora che i tre nuclei di una molecola d'acqua sono disposti a formare un triangolo isoscele, con l'atomo di ossigeno in corrispondenza del vertice. L'atomo di ossigeno esercita una maggiore attrazione verso gli elettroni, rendendo la sua posizione relativamente negativa e creando una struttura a dipolo. L'esatta geometria della molecola dell'acqua è ancora oggetto di speculazione. L'approccio teorico al problema è stato di postulare un modello basandosi su dati sperimentali e ipotesi intuitive; quindi di sottoporre il modello ad analisi matematica, e infine di determinare l'aderenza delle espressioni derivate alle proprietà osservate sperimentalmente.
Per mezzo dello spettro di vibrazione-rotazione del vapore d'acqua è stato accuratamente determinato un angolo di legame di 104,5°. Il legame covalente O−H è molto stabile alle temperature fisiologiche. D'altra parte la grande reattività dell'acqua è favorita dal fatto che i legami O−H che uniscono ciascuna molecola d'acqua alle sue vicine sono relativamente deboli. Il concetto di legame O−H rapidamente rotto e ricostituito ha condotto alla teoria dei flickering clusters (‛cumuli tremolanti') delle molecole di acqua nello stato liquido. Questo non significa che le molecole di acqua siano facilmente separabili; al contrario, è ben conosciuta da chi abbia cercato di separare due superfici di vetro bagnate la straordinaria forza di coesione che le lega, e che deve aver origine da questi legami a idrogeno intermolecolari. La proprietà di autoadesione è alla base del fenomeno della capillarità, cioè di quella forza che fa salire i liquidi all'interno di sottili tubi di vetro, ed è importante per il movimento della linfa nelle piante e del sangue nei piccoli vasi.
Secondo un altro modello di acqua allo stato liquido i legami a idrogeno tra le varie molecole verrebbero distorti piuttosto che rotti, e i differenti stati di aggregazione sarebbero spiegati col grado di distorsione dei legami stessi. Secondo ambedue le ipotesi, quindi, un mutamento di forma a livello molecolare contribuirebbe a determinare le proprietà fisiche dell'acqua. In questo caso si parla di contributo ‛configurazionale', che differisce dal contributo ‛vibrazionale', dovuto semplicemente ai movimenti intermolecolari. Le proprietà caloriche dell'acqua offrono un esempio di come le sue caratteristiche fisiche siano collegate alla sua struttura.
Il calore specifico dell'acqua è superiore a quello di qualsiasi sostanza ad eccezione dell'ammoniaca. Questo significa che l'acqua assorbe e libera calore più lentamente di qualsiasi sostanza capace di mantenere la vita. Questa particolare caratteristica fa sì che grandi masse di acqua mitighino la temperatura terrestre e proteggano gli esseri viventi, costituiti in gran parte di acqua, da bruschi sbalzi di temperatura. Quando l'acqua viene riscaldata il calore viene trasformato in movimento molecolare; tuttavia questo spiega solamente la metà circa della grande capacità che ha l'acqua di assorbire il calore. Il rimanente calore deve essere assorbito nel cambiamento della struttura interna, nella rottura o nella distorsione dei legami a idrogeno.
Una proprietà dell'acqua che è stata di grandissima importanza per l'origine della vita riguarda il suo comportamento anomalo tra 4 e 0 °C. La densità di tutte le altre sostanze aumenta regolarmente con il raffreddamento e raggiunge il suo massimo valore alla temperatura di solidificazione. L'acqua invece si dilata nel raffreddamento da 4 a 0 °C; questo fenomeno non solo ha mutato la faccia della Terra per frantumazione delle rocce, ma ha effettivamente reso possibile la vita poiché, se il suo comportamento fosse stato differente, tutta l'acqua nella profondità degli oceani sarebbe stata trasformata in permanenza in ghiaccio; invece questa peculiare proprietà di dilatazione permette all'acqua di salire quando si trasforma in ghiaccio, in modo che questo si forma solamente alla superficie, mentre le zone più profonde vengono protette da una barriera isolante. Riferendoci al nostro modello molecolare, questo fenomeno potrebbe essere spiegato con l'allungamento dei legami a idrogeno tra 4 e 0 °C; tale effetto compenserebbe abbondantemente la modesta componente vibrazionale tendente a contrarre le distanze intermolecolan, in seguito al raffreddamento.
Il corpo umano produce giornalmente una quantità di calore capace di innalzare la temperatura corporea di 40-50 °C. Questo non rappresenta un pericolo perché i meccanismi di dispersione del calore possono eliminare facilmente tale calore. Sebbene l'aumento potenziale della temperatura corporea, per una normale produzione di calore, possa sembrare grande, esso è solo una frazione dell'aumento di temperatura che noi dovremmo sopportare se il nostro corpo fosse principalmente costituito di qualsiasi altra sostanza diversa dall'acqua. È la grande capacità termica dell'acqua del nostro corpo che ci protegge da un innalzamento fatale della temperatura. Se il calore specifico dell'acqua fosse simile a quello di altre comuni sostanze, l'aumento potenziale della nostra temperatura per la stessa produzione di calore sarebbe di 100-200 °C, il che costituirebbe un intollerabile aggravio per i meccanismi adibiti alla dispersione del calore.
L'acqua stessa, essendo il suo calore latente di evaporazione il più elevato che si conosca, provvede efficientemente alla dispersione del calore per evaporazione. Quando la temperatura ambientale è uguale o maggiore di quella corporea, noi possiamo disperdere calore solamente per evaporazione di acqua dalla superficie: il calore latente dell'acqua permette una massima sottrazione di calore per unità di volume evaporato.
L'acqua è chiamata il solvente universale. Oltre la metà degli elementi esistenti sulla Terra sono più o meno solubili in acqua. Per il suo grande potere solvente l'acqua fornisce perciò un mezzo più ricco per i processi vitali di qualsiasi altro solvente. Di grande importanza è inoltre il fatto che l'acqua, come solvente, è inerte; il soluto non è modificato e mantiene le proprie caratteristiche chimiche.
La costante dielettrica dell'acqua è molto elevata. Ciò significa che l'acqua, benché neutra, è in grado di diminuire l'attrazione elettrica tra ioni di carica opposta in essa disciolti; in tal modo le sostanze ionizzabili possono raggiungete il loro massimo grado di ionizzazione in soluzione acquosa. Le proteine, fondamentali per la vita, contengono molti gruppi elettricamente carichi e sono facilmente solubili in acqua. La capacità dell'acqua di favorire la ionizzazione è di particolare importanza per la biologia, poiché molti dei processi operativi della vita avvengono tra specie ioniche.
Recentemente è stato dimostrato (Deryagin) che la struttura e le proprietà dell'acqua subiscono impressionanti mutamenti quando il vapor d'acqua sia fatto condensare in microcapillari di quarzo. Tale forma, chiamata ‛ortoacqua' dai suoi scopritori russi, presenta caratteristiche anomale di calore e di densità, per esempio ha un punto di ebollizione di 500 °C. I recenti tentativi di chiarirne la struttura hanno condotto all'ipotesi che questo liquido sia un polimero superiore e suggeriscono per esso il termine di ‛poliacqua'. È possibile che le superfici intercellulari o i microcanali all'interno delle cellule possano ristrutturare le molecole di acqua in forma polimerica. Se questo fosse vero, le proprietà dell'acqua intracellulare potrebbero essere completamente differenti da quelle familiari fin qui descritte.
3. Assunzione, conservazione e perdita di acqua
a) Bilancio idrico giornaliero nell'uomo
Tutti gli esseri viventi necessitano di acqua. Ciascuna delle migliaia di forme di vita apparse nel corso della storia del nostro pianeta ha utilizzato un modo proprio di assunzione dell'acqua. I Vertebrati a sangue caldo assumono acqua per via orale come cibo e come bevanda. È istruttivo, per familiarizzarsi col metabolismo quantitativo, esaminare il bilancio giornaliero dell'acqua in un individuo. Un uomo di 70 kg la cui dieta giornaliera sia costituita da proteine (100 g), carboidrati (300 g) e grassi (100 g), immette nell'organismo in media 156 meq di sale al giorno. Supponiamo che il peso totale di cibo di una giornata sia di 1,25 kg. L'acqua disponibile in questa dieta consiste di acqua preformata (misurabile per essiccazione del cibo fino a peso costante) e di acqua d'ossidazione. Un valore verosimile di acqua preformata è come minimo il 60% del peso del cibo. A seconda del cibo, il contenuto di acqua può variare dal 60% per la carne magra ad oltre il 90% per certi frutti maturi. Una dieta di 1,25 kg di peso conterrebbe quindi circa 750 ml di acqua preformata (v. tab. II).

Poiché si conosce la composizione della dieta, si può facilmente calcolare l'acqua di ossidazione. I carboidrati, per esempio, vengono completamente ossidati nell'organismo fino a diossido di carbonio ed acqua:
C6H12O6 + 6O2 → 6CO6 + 6H2O;
su base molare,
180 g C6H12O 6 + 192 g O2 → 264 g CO2 + 108 g H2O
cioè
1 g C6H12O6 → 0,6 g H2O.
Un grammo di carboidrati bruciati nel nostro corpo forma 0,6 g o 0,6 ml d'acqua; 300 g formeranno quindi 180 ml di acqua.
Le proteine, che vengono degradate solamente fino ad urea, formano 0,4 ml di acqua per grammo, cioè 40 ml nel nostro esempio; i grassi invece, per il loro elevato contenuto in idrogeno, generano 1 grammo di acqua per grammo di grasso ossidato.
L'acqua totale di ossidazione nel caso del nostro soggetto sarebbe quindi di 320 ml, l'acqua preformata 750 ml. Questi due volumi sono strettamente dipendenti dalla dieta; in questa particolare dieta, 1.070 ml di acqua vengono necessariamente immessi nel corpo. L'altra fonte di acqua, quella bevuta, è molto più variabile nel suo volume, poiché dipende dalle abitudini, usanze, ecc. Vi è comunque un fabbisogno minimo che il soggetto deve bere per mantenere il bilancio idrico: questa quantità può essere calcolata indirettamente tenendo conto delle perdite obbligatorie di acqua.
Viene generalmente chiamata ‛perdita insensibile' di acqua quella che avviene attraverso la cute per evaporazione (escludendo il sudore) ed attraverso i polmoni essendo l'aria espirata satura di vapore acqueo. Tale perdita complessiva di acqua è misurabile per mezzo di una bilancia sensibile, come variazione del peso corporeo. Un valore medio attendibile per la perdita insensibile di acqua è di 0,5 ml/kg di peso corporeo per ora. Nel caso del soggetto considerato la perdita insensibile di acqua a livello dei polmoni e della cute sarebbe di 840 ml/giorno. Di tale quantità si deve quindi tener conto nel valutare la perdita totale di acqua.
Calcoliamo ora il volume minimo di urina dipendente dalle proteine e dai sali ingenti, essendo i sali ed i prodotti finali del metabolismo proteico i soli componenti della dieta che vi compaiono. (Per definire la concentrazione dei soluti presenti nel plasma o nelle urine useremo la ‛osmole', la quale corrisponde al peso molecolare di ciascuna sostanza in grammi diviso per il numero di particelle in cui la singola molecola si dissocia. Per esempio, 60 grammi di urea corrispondono ad una osmole poiché la molecola di urea non si dissocia in soluzione, mentre la molecola di NaCl si dissocia in due particelle, cosicché il suo peso molecolare in grammi corrisponde a due osmoli. Per evitare l'uso di numeri decimali le concentrazioni vengono espresse in milliosmoli essendo 1 osmole 1.000 mosmoli.) In condizioni di equilibrio salino, i 156 meq di NaCl saranno eliminati giornalmente nell'urina partecipando con 312 mosmoli al suo residuo solido totale. La dieta fornisce normalmente circa 50 meq di potassio il quale, con l'anione che l'accompagna, porterà la quantità degli elettroliti urinari a 412 mosmoli. 100 grammi di proteine della dieta contengono 16 grammi di azoto, il 90% del quale deve essere eliminato nelle urine (15 grammi); di questo il 90% è sotto forma di urea. La dieta del soggetto considerato, perciò, comporta l'eliminazione di 14 grammi di azoto ureico. L'urea, con peso molecolare 60, contiene due atomi di azoto per molecola, cioè il 46% (28/60) di azoto. La quantità di urea contenente 14 grammi di azoto ureico è perciò 14/0,46=30 grammi. Poiché questa quantità è metà del peso molecolare espresso in grammi, essa corrisponde a 0,5 moli o 500 mosmoli di urea.
Il carico totale di soluto da eliminare con l'urina è perciò di 912 mosmoli. Il nostro soggetto può eliminare questa quantità ad una qualsiasi concentrazione urinaria compresa tra 50 mosmoli/litro, nel caso di massima diuresi d'acqua, e 1.200 mosmoli/litro, la più elevata concentrazione urinaria raggiungibile nell'uomo. Dal punto di vista dell'economia dell'acqua è interessante conoscere il più piccolo volume possibile di urina; questo sarà 912/1.200=0,760 litri cioè 760 ml. Inoltre circa 100 ml di acqua vengono eliminati ogni giorno nella normale defecazione, cosicché la perdita totale minima di acqua è 840+760+100=1.700 ml per giorno.
La differenza tra questa perdita totale minima di acqua e la quantità di acqua assunta con la dieta (1.700−1.070=630 ml) rappresenta il volume di acqua che deve essere bevuta perché si mantenga l'equilibrio idrico. Le abitudini e le usanze che regolano l'ingestione di sale con la dieta e l'assunzione occasionale di liquidi durante la giornata aumentano il volume di acqua introdotto di un altro litro o più al giorno. Questa ulteriore quantità, a meno che la sua assunzione non sia stata causata dall'alta temperatura ambientale la quale aumenta la perdita di acqua per evaporazione, va ad aggiungersi al volume minimo urinario (v. tab. II). L'acqua ingerita in sovrappiù, essendo per lo più priva di elettroliti, abbassa il valore massimo di osmolarità urinaria scelto nel nostro esempio a circa 600 mosmoli/litro. In condizioni che facciano salire la perdita per evaporazione, il valore minimo di acqua da ingerire può essere definito come la quantità necessaria a provvedere al volume obbligatorio di urina. In ambiente particolarmente caldo e con lavoro pesante, questa quantità può essere anche di altri 20 litri giornalieri. I più alti valori di perdita d'acqua per sudorazione, misurati per brevi periodi di tempo, si avvicinano ai 4 litri per ora.
La sete, il desiderio di bere, regola perfettamente il bisogno di acqua del corpo. La sensazione di sete è probabilmente legata allo stato di idratazione cellulare e sembra essere trasmessa da osmorecettori, microscopiche stazioni di segnalazione, sensibili alle variazioni di concentrazione del sodio. Poiché la perdita di acqua dal corpo, attraverso la cute, i polmoni e l'urina, è un processo continuo, la sensazione di sete deve comparire quando la diminuzione del rapporto acqua corporea/soluto (sale) raggiunge un valore determinato. L'iniezione endovenosa di una soluzione salina molto concentrata provoca un immediata sensazione di sete. Il luogo di ricezione di questo stimolo è stato recentemente localizzato, e con considerevole accuratezza, da B. Andersson nei suoi studi sulle capre. In questi animali l'iniezione di soluzione salina in aree ben circoscritte dell'ipotalamo provoca un forte desiderio di bere. La stimolazione mediante microelettrodi di platino localizzati in questo ‛centro della sete' provoca la stessa risposta. Quando la sete viene provocata mediante limitazione dell'acqua, alcuni animali ingeriscono una ben precisa quantità di acqua, strettamente corrispondente al deficit. Poiché l'assunzione di acqua cessa prima che l'acqua ingerita sia stata assorbita, l'atto del bere deve in qualche modo segnalare il volume bevuto, affinché esso corrisponda al reale bisogno.
Nell'uomo la soddisfazione della sete non è così ben correlata con la reale deficienza d'acqua. Gli individui che lavorano in ambienti caldi non compensano completamente la perdita d'acqua, anche se questa è facilmente disponibile. Al contrario essi si assoggettano ad una ‛disidratazione volontaria', ingerendo solamente i 2/3 circa dell'acqua necessaria. La quantità rimanente viene assunta dopo il periodo di lavoro, generalmente durante i pasti.
La maggior parte dell'acqua ingerita viene assorbita nell'intestino tenue (digiuno e ileo) a causa dell'enorme superficie della sua mucosa. La prima tappa di questo processo consiste nel flusso di ioni e altre specie chimiche dagli spazi tessutali che circondano il lume intestinale verso l'acqua dell'intestino stesso. Questa presenza di ioni nell'intestino facilita molto il successivo assorbimento dell'acqua, processo accoppiato al trasferimento attivo dei sali dal lume intestinale nel sangue. Gli ioni sodio perciò penetrano nel lume intestinale per diffusione lungo un gradiente di concentrazione chimica, e successivamente vengono trasferiti per trasporto attivo ad opera delle cellule epiteliali della mucosa intestinale.
L'economia di questo processo, come meccanismo di riassorbimento dell'acqua, è sorprendente. La soluzione assorbita è isotonica, il che significa che per ogni 0,3 moli di sale viene assorbito un litro di acqua. Poiché il peso molecolare dell'acqua è 18, un litro contiene 55,6 moli. Per ogni molecola di sale quindi 186 molecole d'acqua attraversano la parete intestinale verso il sangue. Contemporaneamente una piccola quantità di acqua esce dal lume intestinale per semplice osmosi poiché, subito dopo l'assunzione di acqua, si viene a creare una forte differenza di concentrazione tra il lume e gli spazi tessutali. È interessante notare che la permeabilità della parete intestinale all'acqua è così bassa da non permettere, se non a una frazione minima di acqua assorbita, di spostarsi in questo modo.
Quando la quantità di acqua ingerita è superiore ai bisogni immediati del corpo, si produce una diluizione del sangue per il trasferimento di ioni dal sangue all'intestino. Come i classici esperimenti di E. B. Verney hanno dimostrato, la diminuzione di concentrazione di sodio nel sangue è il vero stimolo che segnala ai recettori dell'ipotalamo che una quantità di acqua superiore al fabbisogno è penetrata nel corpo. Tale segnale viene quindi inviato, attraverso il tratto neuroipofisario, alla ghiandola pituitaria posteriore, informandola di interrompere la liberazione dell'ormone antidiuretico (ADH). In assenza di tale ormone il rene elimina una grande quantità di urina diluita, la cui concentrazione può raggiungere il valore di 1/6 di quella del sangue.
Il risultato dell'eliminazione di un grande volume di urina diluita è il ritorno della concentrazione dei fluidi corporei verso il suo valore medio normale. Con l'eliminazione di acqua aumenta la concentrazione in sodio del sangue che perfonde i recettori situati nell'ipotalamo, il cui segnale inibitorio alla ghiandola pituitaria posteriore viene gradualmente ridotto. Questo sistema ‛a retroazione' (feedback) per il mantenimento dell'equilibrio idrico è regolato in modo da mantenere un valore medio di sodio piasmatico di circa 142 meq/litro ed è sensibile a variazioni minori del 2%. Poiché il cloruro di sodio rappresenta oltre il 90% degli elettroliti plasmatici, tale sistema di fatto regola la pressione osmotica del sangue, riuscendo a mantenerla entro limiti molto ristretti.
La composizione del liquido assorbito dall'intestino, come quello che penetra nel lume intestinale attraverso la sua parete è caratteristica per ogni, particolare segmento del tubo digerente. Sembra che il duodeno (la prima porzione dell'intestino tenue) serva a mantenere un equilibrio tra il contenuto intestinale e il sangue, effettuando rapidi flussi di liquido nelle due direzioni. L'ileo serve per l'assunzione da parte del corpo di nuovo sodio e acqua. In tutto l'intestino tenue il riassorbimento attivo del sodio è accompagnato da quantità isosmotiche di acqua. Nell'ileo e nel digiuno viene riassorbito attivamente il cloro, il quale viene sostituito da bicarbonato, rendendo il contenuto leggermente alcalino. Il colon riassorbe attivamente il cloro, mentre secerne bicarbonato e potassio.
Il valore di massimo assorbimento di acqua da parte dell'intestino è in grande eccesso rispetto alle normali necessità. Il valore di assorbimento netto è dato dalla differenza tra due flussi molto rapidi e di opposta direzione. Si può dimostrare che il 50% dell'acqua marcata introdotta nell'intestino tenue raggiunge il sangue in 2-3 minuti, mentre per assorbire il 50% di un carico d'acqua occorrono 30 minuti. Recentemente è stato osservato che la velocità di assorbimento dell'acqua in condizioni di equilibrio raggiunge i 15 ml/min, un valore che, prestabilito o casuale, si avvicina al valore di massimo flusso dell'urina.
Il tratto intestinale assorbe, oltre all'acqua ingerita, un grande volume di liquido prodotto dalla secrezione delle ghiandole dell'apparato digerente. Tale volume è più del doppio di quello del plasma da cui questi secreti derivano; ciò dà un'idea della misura in cui i liquidi corporei vengono incessantemente rielaborati e modificati dalle ghiandole dell'apparato digerente.
La composizione elettrolitica dei secreti gastrointestinali diviene importante solamente in caso di perdita anormale di questi liquidi per diarrea, vomito o intervento chirurgico. Poiché sono interessate grandi quantità di liquido, possono rapidamente comparire gravi alterazioni nell'equilibrio acido-base e importanti variazioni nella normale composizione elettrolitica del corpo. Un'accurata misura dei volumi di liquido perduti e l'analisi dei loro costituenti elettrolitici costituiscono i principi basilari di una appropriata sostituzione di liquido.
b) La conservazione dell'acqua da parte del rene
Per meglio apprezzare la capacità del rene di Mammifero di risparmiare l'acqua è utile chiarire l'importanza che il rene riveste nel processo evolutivo. Gli Invertebrati possiedono delle strutture tubulari intercomunicanti, chiamate nefridi, poste tra la loro cavità celomatica e l'ambiente circostante, attraverso le quali passa un fluido. Tale fluido ha già le caratteristiche dell'urina prodotta nei Vertebrati superiori. Si tratta infatti del fluido extracellulare che contiene non solo prodotti di rifiuto, ma anche sostanze importanti per l'organismo (zuccheri, amminoacidi, grassi), le quali devono essere riassorbite nel loro passaggio attraverso questi tubuli. Poiché gli Invertebrati sono in equilibrio osmotico con l'ambiente, essi non hanno bisogno di alcun processo che permetta la conservazione dei sali e dell'acqua. Il sale è presente in abbondanza nel mare circostante, e l'acqua viene prodotta nell'animale stesso per eliminazione attiva del cloruro di sodio.
Nel corso di una serie di processi evolutivi, molte forme di vita marina migrarono dal mare nei laghi d'acqua dolce e nei fiumi. La più elevata pressione osmotica dei fluidi corporei richiese allora una protezione contro un'ingresso di acqua pericoloso per la sopravvivenza di tali animali, se essi non avessero sviluppato dei meccanismi tali da impedire l'ingresso dell'acqua, oppure da eliminarla una volta penetrata nel loro corpo. Furono usati ora l'uno ora l'altro di questi meccanismi. La corazza protettiva dei Crostacei, impermeabile all'acqua, era efficiente, ma allo stesso tempo impediva un loro ulteriore sviluppo. Una seconda soluzione, la quale aprì la strada a più ampi processi evolutivi, consistette nello sviluppo di un efficace apparato filtrante nel rene, per mezzo del quale poteva essere eliminata l'acqua in eccesso. I nefridi, già presenti, furono accoppiati ad un glomerulo formato da un piccolo gomitolo di sottili vasi sanguigni (capillari). Questi furono chiamati ‛corpuscoli' dall'anatomico M. Malpighi di Bologna, che li descrisse per primo. Attraverso di essi, per la forza della pressione del sangue arterioso, il plasma libera un filtrato pressoché privo di proteine, che passa lungo le strutture tubulari.
Poiché in questo filtrato sono presenti tutti i costituenti del plasma a basso peso molecolare, all'apparato filtrante deve essere associato un efficiente sistema di riassorbimento atto alla conservazione di quelle sostanze filtrabili necessarie all'organismo. A tal fine il primo tratto del nefridio (tubulo prossimale) serve al recupero dei materiali nutritivi, mentre il secondo tratto, collegato al primo da un breve e sottile segmento, funziona per la conservazione dei sali. Le cellule di questo tubulo sono impermeabili all'acqua ed hanno la capacità di riassorbire completamente il sodio. Interessante è il fatto che esse usano un sistema di trasporto accoppiato impiegato da forme più evolute: per eliminare gli ioni idrogeno derivati dalla produzione intracellulare di acidi, questi ioni vengono scambiati con ioni sodio.
I pesci d'acqua dolce possiedono dunque dei meccanismi che li proteggono sia contro un eccesso di acqua sia contro un'insufficienza di sodio. I pesci che oggi vivono in acque salate probabilmente rappresentano delle specie che vi ritornarono provenienti dalla vita d'acqua dolce dopo un periodo evolutivo, poiché presentano l'apparato base per la filtrazione e il riassorbimento; tuttavia si trovarono di fronte ad un nuovo dilemma: il rapporto osmotico era invertito; essendo la concentrazione osmolare del mare almeno tre volte superiore a quella del plasma dei pesci d'acqua dolce, il rene dei nuovi immigranti non aveva più alcuna utilità. La protezione contro l'ambiente più concentrato richiedeva ora o l'uso di meccanismi per l'escrezione del sale, come fecero i pesci ossei mediante speciali strutture nelle loro branchie; oppure l'aumento della concentrazione plasmatica al di sopra di quella marina, come nel caso degli Elasmobranchi. A tal fine questi ultimi utilizzarono una sostanza inerte, l'urea, per aumentare la loro concentrazione sanguigna al punto che la concentrazione plasmatica superasse quella dell'ambiente, permettendo loro, così, di assumere acqua dal mare.
I pesci ossei, ritornati al mare, ridussero il numero e la funzionalità dei loro glomeruli, in qualche caso completamente. Questi pesci aglomerulari producono urina per secrezione attraverso quelle medesime cellule che, nelle forme in possesso di capacità filtrante, compiono la funzione del riassorbimento. L'acqua, necessaria alla solubilizzazione e all'escrezione dei prodotti di rifiuto e degli elettroliti, deve attraversare queste cellule per entrare nel lume tubulare. Il meccanismo interessato sembra funzionare analogamente al meccanismo di riassorbimento dell'acqua nelle forme d'acqua dolce. Il trasporto attivo degli elettroliti, in questo caso probabilmente magnesio e calcio, crea un gradiente osmotico lungo il quale si muove l'acqua.
Il passaggio alle forme terrestri, rappresentato oggi dagli Anfibi, portò nuovi problemi per la conservazione dell'acqua. In questi organismi è presente il pericolo della perdita d'acqua per l'attiva azione filtrante del rene e per l'evaporazione attraverso la cute. Avvenne tuttavia che nelle specie di Anfibi sopravvissute, durante i brevi periodi passati sulla terraferma la filtrazione glomerulare si riduceva considerevolmente e cessava quasi completamente la produzione di urine. Poiché la filtrazione a livello del glomerulo dipende dalla pressione del sangue esercitata nei capillari glomerulari, è possibile un controllo mediante la regolazione del grado di vasocostrizione dei vasi afferenti. Incontriamo qui uno dei primi meccanismi di regolazione che coordinano gli scambi idrici, la pressione osmotica e il volume sanguigno mediante l'intervento del sistema nervoso. Inoltre gli Anfibi svilupparono un ormone costituito di otto peptidi (arginina-vasotocina), secreto dall'ipofisi posteriore, il quale aumenta considerevolmente la permeabilità all'acqua della cute di rana e della vescica di rospo, un fattore importante nella regolazione idrica di questi animali.
In un meccanismo regolatore che opera per attività vasomotoria è implicita la presenza di un controllo riflesso del tono vasocostrittore. Il controllo è effettuato per mezzo delle vie simpatiche ed è l'attività di questi nervi che deve essere regolata per via riflessa. Ciò avvenne con lo sviluppo del cosiddetto riflesso ‛depressore', i cui recettori sono situati nell'aorta cefalica e segnalano ogni aumento della pressione del sangue al sistema nervoso centrale, il quale a sua volta inibisce l'attività spontanea del simpatico. Questo meccanismo riflesso serve per proteggere i delicati capillari branchiali da pressioni dannose. Inoltre negli Anfibi i siti recettori sono localizzati nelle vene centrali e negli atri del cuore. Qualunque eccesso di volume in queste zone stimola i recettori con conseguente inibizione del tono simpatico agente sulle arteriole afferenti ai glomeruli, il cui grado di vasocostrizione quindi diminuisce. Come effetto si ha un aumento della pressione nei capillari glomerulari (pressione di filtrazione) e un corrispondente aumento del filtrato e dell'escrezione di acqua da parte del rene. Al contrario, ogni diminuzione del volume sanguigno porta a una riduzione dell'inibizione del simpatico, con conseguente diminuzione della filtrazione e della perdita d'acqua.
Una riduzione della filtrazione glomerulare era tuttavia inconciliabile con l'ulteriore adattamento dei Vertebrati alla vita terrestre. Poiché l'eredità di un rene dotato di potere di filtrazione e di riassorbimento non provvedeva all'eliminazione dei prodotti di rifiuto attraverso la via secretoria, doveva essere trovato un altro meccanismo che non interferisse con la conservazione dei sali e dell'acqua. Ricordiamo che la parte distale dei tubuli renali dei Vertebrati di acqua dolce è impermeabile all'acqua e che questa proprietà permette al rene di eliminare grandi quantità di acqua in caso di necessità.
Nei Vertebrati superiori, invece, lo stesso meccanismo ormonale che aumenta la permeabilità della cute degli Anfibi all'acqua viene utilizzato per aumentare la permeabilità all'acqua della parte distale del nefrone. Sotto controllo ormonale l'acqua filtrata può accompagnare il riassorbimento del sodio e viene reimmessa nel sistema vascolare. Questo meccanismo ormonale permette ai Vertebrati superiori di conservare l'acqua senza ridurre la quantità del filtrato glomerulare, condizione necessaria all'escrezione dei prodotti di rifiuto.
L'ormone antidiuretico (ADH), arginina-vasopressina, viene prodotto nei nuclei di speciali cellule dell'ipotalamo e quindi passa, mediante flusso assonico, lungo il tratto sopraottico-ipofisario, nella parte posteriore dell'ipofisi, dove viene immagazzinato. Questo octapeptide è uno dei più potenti agenti biologici conosciuti, essendo attivo a concentrazioni inferiori a 10-12 M. Lo stimolo adeguato a provocare la liberazione dell'ADH è costituito dalla concentrazione del sodio nel plasma perfondente la regione anteriore dell'ipotalamo. Ogni aumento della concentrazione salina del plasma (causato dalla perdita di acqua) fa aumentare la secrezione di ADH. L'ormone, agendo sulle cellule del tubulo distale e del dotto collettore, aumenta la loro permeabilità all'acqua.
L'esatto meccanismo d'azione dell'ormone antidiuretico è attualmente oggetto di intenso studio. Preparati di cute di rana e di vescica di rospo, oltre a frammenti di dotto collettore perfusi in vitro, sono stati i modelli sperimentali più utili. Se applicato sulla superficie sierosa di queste strutture, l'ormone aumenta selettivamente la permeabilità all'acqua della corrispondente membrana mucosa nel caso del dotto collettore dei Mammiferi, e a particolari soluti organici e al sodio nel caso della cute e della vescica. Un'ipotesi di meccanismo d'azione attualmente accettata postula l'attacco dell'ormone ad un sito recettore sulla membrana sierosa, dove esso reagirebbe con l'adenilciclasi, enzima presente ovunque sulla membrana cellulare. La reazione con questo enzima stimolerebbe la formazione intracellulare di un secondo messaggero, l'adenosin-monofosfato (AMP) ciclico, dall'ATP. Si pensa che questa sostanza diffonda verso la membrana mucosa e ne determini il cambiamento di permeabilità sia per apertura di canali a porosia per variazione dell'orientamento delle strutture della membrana. È interessante notare che la liberazione dell'ormone antidiuretico viene inibita dai riflessi attivati dai recettori dell'atrio sinistro del cuore in seguito all'aumento del volume sanguigno. Come nella rana, in cui un aumento del volume del sangue inibisce l'attività simpatica, così nelle forme superiori vi è un doppio effetto, poiché l'inibizione simpatica produce una vasodilatazione riflessa (con conseguente aumento del filtrato glomerulare) e inoltre un'inibizione della liberazione di ADH. Entrambi gli effetti aumentano l'escrezione di urina e diminuiscono il volume sanguigno. Nei Mammiferi più grossi la regolazione del volume sanguigno si può dimostrare sperimentalmente. L'infusione di sangue porta ad un aumento della pressione venosa e della eliminazione di acqua nelle urine; questo indica chiaramente che vi è un'inibizione nella secrezione di ADH, come è stato recentemente dimostrato col dosaggio dell'ormone nel sangue periferico.
c) La concentrazione dell'urina: un mezzo per la conservazione dell'acqua
L'ormone antidiuretico non potrebbe eseguire da solo il compito della conservazione dell'acqua poiché è anche necessaria una forza che faccia uscire l'acqua dalla porzione distale del nefrone (una volta che l'ormone abbia fatto aumentare la permeabilità all'acqua della sua parete epiteliale) e la faccia ritornare nel sangue. Tale forza viene fornita, nel rene dei Mammiferi, dalla presenza di una nuova struttura nella regione midollare, che permette al rene di restituire acqua al sistema vascolare, formando nel contempo un'urina concentrata. Il rene è formato da una regione esterna detta ‛corticale', e da una regione interna detta ‛midollare'. Il glomerulo (il rene umano ne possiede circa un milione) versa il suo filtrato in un tubulo il quale, dopo parecchie circonvoluzioni (tubulo contorto prossimale) si continua in un'ansa discendente (ansa di Henle), che si ripiega su se stessa ritornando in vicinanza del glomerulo. Vicino al glomerulo esso compie parecchie altre circonvoluzioni (tubulo contorto distale) prima di immettersi in un dotto collettore. I glomeruli situati vicino alla superficie della corticale renale hanno anse discendenti brevi, i cui apici si trovano in corrispondenza della parte esterna della regione midollare; i glomeruli posti più in profondità possiedono anse discendenti molto lunghe, che penetrano nella zona più interna o papillare della midollare. I vasi sanguigni che irrorano questa regione della midollare hanno anch'essi la forma di lunghe anse capillari parallele e strettamente giustapposte alle anse dei nefroni e ai dotti collettori.
Fu la peculiare disposizione anatomica di queste strutture che suggerì l'ipotesi che il meccanismo di concentrazione urinaria operasse per mezzo di un sistema di moltiplicazione ‛a controcorrente', sistema già noto ai fisici. Il modello di tale sistema fu suggerito per primo da W. Kuhn, un chimico-fisico di Basilea.
La possibilità di conservare il calore utilizzando il principio dello scambio di calore a controcorrente era ben conosciuta da molti anni da parte degli ingegneri. Il merito specifico dell'idea di Kuhn fu l'aggiunta di un principio di moltiplicazione ad un modello a controcorrente. In questo modello viene applicata una piccola pressione all'ingresso di un tubo ad U. Una forte resistenza al gomito del tubo abbassa la pressione nel ramo di uscita, mentre mantiene immutata quella nel ramo di entrata. Ad ogni punto lungo il ramo d'ingresso l'acqua è forzata ad attraversare la parete costituita da una membrana semipermeabile, penetrando nel ramo di uscita. La membrana non lascia uscire il soluto, cosicché la soluzione che rimane nel ramo d'ingresso viene leggermente concentrata. Questo effetto singolo si ripete costantemente lungo tutto il ramo d'ingresso del tubo, con il risultato di concentrare più volte la soluzione. Passando nel ramo d'uscita del tubo, il liquido viene ridiluito alla sua concentrazione iniziale dall'acqua filtrata attraverso la membrana semipermeabile lungo il breve percorso.
Si noti che in questo modello semplice il liquido viene concentrato solo temporaneamente al gomito del tubo. Per poter utilizzare questo sistema di concentrazione è necessario aggiungere un altro tubo, il quale sia in contatto con il ramo afferente del tubo ad U attraverso una seconda membrana semipermeabile. Una soluzione che scorra in questo tubo subirà anch'essa un aumento di concentrazione a causa della cessione di acqua per forza osmotica al ramo afferente del tubo ad U. Il vantaggio consiste nel fatto che dal terzo tubo fuoriesce una soluzione concentrata. In questo caso la soluzione nel tubo ad U viene diluita dall'aggiunta di acqua proveniente dal terzo tubo, e per mantenere lo stesso potere di concentrazione sarebbe necessario aumentare l'energia fornita (pressione). L'aggiunta della terza unità fa sì che il liquido che esce dal tubo ad U sia più diluito di quanto lo fosse all'ingresso.
Benché sia improbabile che le differenze di pressione idrostatica giochino un ruolo importante nel meccanismo di concentrazione nel rene di Mammifero, questo modello può servire come guida per chiarire le nostre idee riguardo al suo funzionamento.
Se disponiamo molte di queste unità in ordine longitudinale a somiglianza dei complessi delle anse di Henle e dei dotti collettori nel rene di Mammifero, appare chiaro che: a) ad ogni sezione trasversa perpendicolare all'asse si avranno uguali concentrazioni del liquido ivi contenuto; b) la concentrazione dei liquidi nel ramo d'ingresso aumenterà progressivamente dalla sommità verso il fondo; c) il liquido defluente dal ramo d'uscita dei tubi ad U sarà diluito rispetto a quello d'ingresso.
L'esistenza di un gradiente di concentrazione crescente dalla corteccia renale in direzione della papilla fu dimostrata per la prima volta da H. Wirz in sezioni congelate di rene di ratto. Questa osservazione sta a sostegno delle prime due predizioni derivanti dal nostro modello. La terza predizione (la diluizione del liquido uscente dal sistema) aveva già avuto dimostrazione mediante la tecnica della micropuntura di Walker e altri. Il liquido dei tubuli distali (uscente dalle anse di Henle) venne raccolto con micropipette e all'analisi si dimostrò meno concentrato di quanto lo era il filtrato all'ingresso dell'ansa.
Si può ora affermare con sufficiente certezza che l'urina viene concentrata (con conseguente recupero dell'acqua) mentre scorre nei dotti collettori, i quali passano attraverso i tessuti a concentrazione osmolare crescente della midollare renale. Dato che i dotti collettori hanno le pareti permeabili all'acqua, è evidente che il liquido in essi contenuto perderà acqua per attrazione osmotica lungo il suo percorso dalla corteccia all'apice papillare, equilibrandosi ad ogni livello con il fluido interstiziale della midollare.
Si può vedere che la capacità di concentrare l'urina è proporzionale alla lunghezza dei dotti collettori. Infatti, già nel 1909 J. P. Peters aveva messo in evidenza che le diverse specie animali potevano, con l'allungarsi della papilla urinaria, concentrare sempre più l'urina. Gli animali che vivono nell'acqua dolce, come il castoro e l'ippopotamo, i quali non hanno bisogno di conservare l'acqua, hanno papille molto corte e non possono concentrare l'urina oltre una volta e mezza la concentrazione osmolare del plasma. Alcuni animali del deserto, al contrario, con il loro bisogno vitale di conservare l'acqua, hanno una papilla urinaria di enorme lunghezza che si estende nella pelvi e riescono a produrre un'urina a concentrazione di oltre 15 volte superiore a quella del loro sangue. Questa straordinaria capacità permette ai Roditori del deserto di sopravvivere basandosi solamente sulla loro acqua di ossidazione, potendo infatti alimentarsi di soli semi seccati in stufa.
Quando risultò chiaro che nei tubuli renali non vi erano differenze di pressione idrostatica abbastanza grandi da realizzare il modello a pressione suggerito da B. Hargitay e W. Kuhn, fu necessario pensare ad un'altra fonte di energia. L'analisi del tessuto midollare iperosmotico dimostrò che i suoi principali componenti erano il sale e l'urea. Poiché si sapeva che le cellule renali trasportano attivamente il sodio, si pensò che la forza iniziale capace di creare un gradiente di concentrazione fosse rappresentata appunto dal trasporto attivo del sodio. Kuhn e Ramel proposero quindi un modello teorico di meccanismo di concentrazione basato sul trasporto attivo del sodio.
È stato visto che gli spessi rami ascendenti delle anse di Henle possono trasportare molto attivamente il sodio ed hanno una bassa permeabilità all'acqua, due condizioni assolutamente necessarie perché si venga a creare un interstizio midollare ricco di sodio. Negli adiacenti sottili rami discendenti si ha una perdita di acqua, la quale passa nei tessuti circostanti più concentrati, mentre entra una certa quantità di sodio, così che il liquido tubulare si viene ad equilibrare con l'interstizio che lo circonda.
Il temporaneo stato di concentrazione del liquido tubulare al gomito dell'ansa di Henle sarebbe analogo a quello che si viene a creare al gomito del modello a pressione. In questo modello il liquido che ritorna attraverso il ramo ascendente viene ridiluito dall'acqua che attraversa la membrana. Nel ramo ascendente dell'ansa di Henle, invece, il liquido viene diluito per la rimozione attiva di sale, che va a costituire il principale ingrediente dell'interstizio midollare concentrato.
Benché non ancora definitivamente dimostrato, sembra probabile che il meccanismo attivo che produce il gradiente midollare sia il trasporto del sodio lungo tutto il ramo ascendente dell'ansa (v. Ullrich e altri, 1961). Un fattore essenziale per la riuscita di questo meccanismo concentrante è la bassa permeabilità all'acqua di tutto il ramo ascendente, affinché questa non segua il sodio nel passaggio al tessuto circostante, andandone ad annullare l'effetto concentrante.
Il flusso sanguigno attraverso il rene, per unità di peso tessutale, è tra i più elevati che si conoscano. Il 90% di questo sangue, tuttavia, perfonde la corteccia renale. Il flusso sanguigno attraverso la midollare è eccezionalmente basso. Se non fosse così, il gradiente midollare necessario per il processo di concentrazione urinaria non potrebbe mantenersi, e sarebbe dissipato dall'effetto ‛diluente' del flusso sanguigno. La peculiare configurazione delle anse dei ‛vasi retti', per scambio a controcorrente, serve a mantenere il sale e l'urea negli strati midollari più profondi. Una netta differenza nella disposizione delle anse si ha nel passaggio dalla midollare esterna all'interna: nella esterna i fasci sono abbondanti e molto stipati, cosicché l'effetto trappola esercitato sul soluto di ritorno viene molto aumentato.
I rami discendenti dei ‛vasi retti' sono meno permeabili all'acqua dei rami ascendenti; tale differenza rappresenta la tappa finale nel processo di conservazione dell'acqua. L'acqua uscente dai dotti collettori per attrazione osmotica passa nel plasma più concentrato della parte ascendente dei ‛vasi retti' e ritorna nel circolo venoso.
È così, dunque, che il processo di concentrazione dell'urina opera per conservare l'acqua per le necessità corporee, e che si raggiunge lo scopo per il quale queste particolari strutture della midollare renale sono preordinate.
4. I compartimenti dei liquidi corporei
Una volta immessa nel corpo, l'acqua entra nella dinamica dei liquidi corporei. Questi stanno in parecchi compartimenti più o meno definiti, la cui distribuzione anatomica e la cui composizione elettrolitica sono schematizzate nelle tabb. IV-VI. Nonostante gli scambi costanti tra loro e con l'ambiente esterno, sia il volume sia la composizione di questi differenti compartimenti sono strettamente regolati e costituiscono un sistema aperto in stato di disequilibrio stazionario, caratteristico dei processi vitali.
a) Misura dei compartimenti
Il contenuto totale d'acqua del corpo e la sua distribuzione nei vari compartimenti corporei si possono più convenientemente misurare con la cosiddetta ‛tecnica della diluizione'. Questa consiste semplicemente nell'iniettare nel corpo una quantità conosciuta di una sostanza tracciante e nel determinarne la concentrazione dopo che essa ha raggiunto l'equilibrio nel compartimento sotto studio. Poiché la quantità totale di tracciante presente nel corpo (M) è data dalla sua concentrazione (C) per il volume in cui esso è distribuito (V), cioè:
M =V • C
il volume può essere calcolato una volta che si conoscano la quantità totale e la concentrazione del tracciante:
V = M / C. (1)
Nella determinazione dell'acqua corporea totale, per esempio, si inietta endovena una quantità conosciuta di un tracciante radioattivo, acqua triziata per esempio, e si lascia equilibrare per un periodo di tempo standard. La sua concentrazione nell'acqua plasmatica viene quindi calcolata dall'attività specifica di un campione di plasma (come impulsi di radioattività per minuto). Poiché l'acqua marcata si equilibra con tutta l'acqua del corpo, il suo volume di distribuzione sarà dato dall'equazione (1) sopra riportata. Il tempo necessario all'acqua marcata per equilibrarsi con tutta l'acqua corporea è molto breve, così che non è necessario fare alcuna correzione per l'escrezione dell'isotopo.
La misurazione dei compartimenti dei liquidi corporei per mezzo dei traccianti implica la validità di alcune premesse. La prima è che la sostanza tracciante stessa non alteri le funzioni stabilizzatrici che regolano il compartimento da misurare. Inoltre, perché sia valido il principio della diluizione dell'isotopo, si deve presupporre che il corpo non faccia distinzioni tra molecola marcata e non marcata.
Un altro assunto generalmente ammesso nell'analisi dei sistemi a compartimenti è che la resistenza al flusso tra compartimenti sia localizzata a livello dell'interfacie tra i compartimenti stessi, per esempio a livello della membrana cellulare se il nostro tracciante si dovesse equilibrare con l'acqua intracellulare. Fino ad ora vi è stato accordo nel credere che l'acqua, una volta entrata in un organismo vivente, vada ad equilibrarsi con tutta la rimanente acqua contenuta nell'organismo stesso. Ottanta anni fa Pfeffer postulò che la membrana cellulare costituisse una barriera universale ai movimenti di tutti i soluti e dell'acqua, e che la diffusione all'interno della cellula fosse istantanea. Poiché non si può pensare che alcuna membrana cellulare sia impermeabile all'acqua, è rimasta la convinzione che l'acqua ingerita si distribuisca ovunque rapidamente.
Benché l'errore introdotto nell'accettare questi presupposti per la determinazione dei compartimenti dei liquidi corporei sia piccolo, nessuno di essi si può considerare valido in assoluto.
Così, Ling crede che il fattore limitante la diffusione dell'acqua marcata non sia la permeabilità della membrana cellulare, ma piuttosto la diffusione nella massa di acqua intracellulare. Lo stato fisico in cui l'acqua si trova nella cellula vivente può causarne una distribuzione non omogenea, come già abbiamo suggerito. Cook (v., 1967), parlando dell'acqua non solvente presente negli eritrociti umani, conclude che una frazione significativa di acqua intracellulare è presente in una forma diversa da quella della rimanente acqua, e che la maggior parte di essa è legata alle proteine. Ad un livello più elementare Bentzel e Solomon (v., 1967) affermano che una parte dell'acqua mitocondriale non può partecipare ai fenomeni osmotici. Queste precisazioni porteranno qualche modifica ai valori comunemente accettati per l'acqua intracellulare, ma non saranno da noi ulteriormente approfondite.
L'acqua corporea totale, misurata nell'uomo in vivo con la tecnica della diluizione ed espressa come percentuale del peso corporeo, varia con l'età ed il sesso come mostra la tab. III. L'elevata percentuale d'acqua nell'infanzia è dovuta soprattutto ad un aumentato volume di liquido extracellulare e può essere causata dalla ridotta capacità del neonato di eliminare il sale e dal minore potere concentrante del rene infantile. Il differente contenuto d'acqua nei due sessi dopo l'adolescenza è dovuto principalmente al differente contenuto di grasso. Poiché nel grasso è presente poca acqua, un accumulo di tessuto adiposo abbassa il valore percentuale di acqua riferito al peso corporeo. Quando il contenuto di acqua viene espresso come percentuale della massa corporea magra, queste differenze risultano molto minori.
La prima e più evidente distinzione che si possa fare dell'acqua corporea è in acqua intracellulare ed extracellulare. Il liquido extracellulare comprende il plasma e il liquido interstiziale (che a sua volta include l'ambiente circostante le cellule). All'interno delle cellule, nel liquido intracellulare, si svolgono i processi metabolici dell'organismo. Tutto il movimento di alimenti, sali minerali ed ormoni assunti dall'organismo o messi in circolo deve attraversare il liquido interstiziale per raggiungere le cellule. I prodotti Catabolici del metabolismo cellulare si muovono in direzione opposta per la stessa via. Una piccola parte del liquido interstiziale ritorna al compartimento vascolare attraverso i canali linfatici ed è detta ‛linfa'. La linfa contiene una concentrazione di proteine superiore a quella della più abbondante parte di liquido interstiziale che ritorna nel circolo direttamente attraverso le pareti dei capillari.
La composizione di questi due principali compartimenti mette in evidenza le marcate differenze esistenti tra di loro. A differenza dell'ameba, la cui fase extracellulare, lo stagno, è infinita se paragonata alla sua massa, l'ambiente circostante le cellule dell'organismo ha un volume relativamente piccolo e richiede un controllo costante che ne mantenga stabile la composizione. I principali organi che presiedono a questi meccanismi regolatori sono i reni ed i polmoni; le forze sono quelle della pressione idrostatica, fornita dal cuore, della diffusione e del trasporto attivo.
Il liquido interstiziale viene formato continuamente dalla filtrazione del plasma alle estremità arteriose dei capillari tessutali, dove la pressione idrostatica è di circa 40-45 mm di Hg, e ritorna al circolo in corrispondenza delle estremità venose, dove la pressione oncotica del plasma, tendente a riassorbire il liquido nei capillari, supera la pressione idrostatica del sangue, qui ridottasi a 10-15 mm di Hg. I primi a considerare l'importanza delle forze responsabili dei movimenti del plasma attraverso i capillari, la pressione idrostatica mantenuta dal cuore e l'opposta pressione oncotica causata dalle proteine del plasma, furono Ludwig (filtrazione) e Starling (riassorbimento) alla fine del secolo scorso. Tuttora spesso non ci si rende conto che questo è il tratto più importante del circolo, poiché dopotutto è nelle cellule che noi viviamo, ed esse vengono nutrite e protette dal nostro milieu intérieur, il liquido interstiziale, come Cl. Bernard affermò per primo.
Considerando le caratteristiche di permeabilità dei capillari di Mammifero, le differenze di pressione potrebbero permettere la filtrazione di circa 20 litri di plasma al giorno, in un individuo di media corporatura. Il flusso linfatico totale nell'uomo a riposo è stato calcolato essere di circa 2 litri al giorno, e può aumentare fino a 3-4 litri in seguito alla quotidiana ingestione di cibo. I restanti 16 litri di liquido interstiziale devono venir riassorbiti all'estremità dei capillari, dove vi è una pressione ridotta. Questi 20 litri di liquido interstiziale sarebbero insufficienti a provvedere ai bisogni metabolici delle cellule dell'organismo. La maggior parte del materiale nutritivo raggiunge le cellule per un semplice processo di diffusione attraverso le pareti dei capiliari, e nello stesso modo i prodotti di rifiuto del metabolismo cellulare raggiungono il sangue. L'utilizzazione degli elementi nutritivi da parte della cellula produce quel gradiente di concentrazione necessario al movimento all'interno dell'interstizio, mentre l'accumulo dei prodotti catabolici mantiene un gradiente di concentrazione tale da favorire il loro ritorno nel sangue.
L'equilibrio che si viene a creare tra il plasma ed il liquido interstiziale mediante il meccanismo della diffusione permette di misurare il compartimento extracellulare. Qualsiasi sostanza tracciante capace di attraversare la parete capillare ma non quella cellulare sarebbe teoricamente ideale per misurare il volume del liquido extracellulare. In pratica una sostanza simile non è stata trovata poiché tutti i traccianti finora utilizzati penetrano in parte nelle cellule. Le sostanze comunemente usate per misurare il volume del liquido extracellulare si distribuiscono in volumi che variano dal 16% del peso corporeo, per l'inulina ed il mannitolo, ad oltre il 30%, per gli isotopi instabili del sodio e del cloro. Questa differenza è in parte dovuta alla più lenta diffusione della grossa molecola dell'inulina nell'acqua meno accessibile del tessuto connettivo denso, ed in parte al fatto che gli ioni più piccoli penetrano nelle cellule. Nel caso del sodio, il suo isotopo può anche incorporarsi nella struttura cristallina dell'osso, fenomeno che fa aumentare il suo volume apparente di distribuzione. La differente distribuzione delle sostanze usate per misurare il volume del compartimento extracellulare in toto sta probabilmente ad indicare che esso di fatto esiste come parecchi sottocompartimenti fisiologici. Questi sono: a) il plasma; b) il liquido interstiziale e la linfa; c) il tessuto connettivo denso, la cartilagine e l'osso; d) i liquidi transcellulari.
Il volume plasmatico e del sangue circolante si calcola dalla diluizione subita da sostanze traccianti la cui distribuzione sia limitata al letto vascolare. A tale scopo sono stati usati per parecchi anni il colorante ‛blu Evans', che si lega alle proteine, e la sieroalbumina marcata con iodio131 (RISA). Entrambe le sostanze, se iniettate endovena, hanno la stessa distribuzione della sieroalbumina. Poiché l'uscita di questa grossa molecola dal compartimento vascolare è lenta e relativamente costante, per ottenere il valore della concentrazione dell'albumina marcata prima della sua uscita dal compartimento, si può estrapolare all'indietro fino al momento dell'iniezione la curva della concentrazione in funzione del tempo. Alcuni ricercatori preferiscono far trascorrere dopo l'iniezione un tempo arbitrario per il mescolamento (10 minuti), dopo il quale prelevano un campione di sangue e in base alla concentrazione del tracciante in esso contenuto e alla quantità iniettata, calcolano il volume totale di plasma, supponendo che non vi sia stata alcuna perdita di sostanza. Dal volume plasmatico si può facilmente calcolare il volume di sangue circolante usando l'ematocrito, il quale indica la frazione di globuli rossi per unità di sangue intero.
Il volume del sangue si può anche calcolare misurando la massa totale dei globuli rossi circolanti e quindi, dal valore ematocrito, si può risalire al volume di plasma e di sangue intero. In questo metodo gli eritrociti vengono marcati con un isotopo radioattivo, come il 51Cr o il 32P , o con ossido di carbonio, un gas che possiede una grandissima affinità per l'emoglobina contenuta nei globuli rossi.
È interessante notare che il volume di sangue calcolato con il metodo degli eritrociti marcati è leggermente inferiore a quello ottenuto con i traccianti plasmatici. Questa differenza, dovuta al passaggio più rapido degli eritrociti attraverso i capillari, non è tanto grande da rendere un metodo migliore dell'altro.
Il liquido interstiziale e la linfa non possono essere misurati che per differenza tra il volume extracellulare totale e quello plasmatico. Questa misurazione è soggetta a tutte quelle imprecisioni inerenti alle determinazioni dei compartimenti dei liquidi corporei, sia a causa della correzione per il volume plasmatico, sia per la differente capacità di penetrazione nelle frazioni del liquido extracellulare da parte delle sostanze marcate. La miglior approssimazione si ha probabilmente con i saccaridi, poiché gli zuccheri si equilibrano rapidamente con ogni porzione di liquido extracellulare, ad eccezione del tessuto connettivo denso e dell'osso. Il valore a cui in tal modo si è giunti, il 12% circa del peso corporeo oppure il 20% di tutta l'acqua corporea, comprende anche parte dell'acqua del tessuto connettivo denso, ma è la migliore determinazione finora disponibile.
È evidente che l'osso contiene poca acqua in proporzione al suo peso, ma la sua grande massa spiega come il sistema scheletrico contenga circa l'8% di tutta l'acqua corporea. Il tessuto connettivo denso e la cartilagine hanno una ridotta circolazione sanguigna e pochi elementi cellulari, come appare dagli esami istologici, cosicché la maggior parte della loro acqua appartiene al compartimento interstiziale e si equilibra molto lentamente con le sostanze radioattive usate nella determinazione del fluido extracellulare. Questi due tessuti insieme contengono un altro 8% circa di tutta l'acqua corporea.
I liquidi transcellulari sono prodotti dai sistemi di trasporto attivo delle cellule, e comprendono le secrezioni ghiandolari, il liquido cerebrospinale ed il liquido contenuto nelle camere interne dell'occhio. La quantità maggiore di questi liquidi è di gran lunga quella prodotta dalle ghiandole del tubo digerente e della mucosa intestinale. Ciascuno di essi ha una composizione caratteristica ed appropriata alla speciale funzione cui è destinato. Il loro volume totale è piccolo in condizioni normali, essendo circa il 2,5% di tutta l'acqua corporea.
Nella tab. IV sono schematizzati il contenuto e la distribuzione di tutta l'acqua corporea. Si deve ricordare tuttavia che l'età, il sesso ed il contenuto di grasso sono causa di variazioni significative. I dati ottenuti con le tecniche di diluizione sopra descritte, o con l'analisi diretta di campioni di tessuto o con lo studio di colture di tessuto in vitro, ci consentono di definire con notevole precisione i compartimenti dell'acqua corporea in condizioni normali, e di rilevarne alterazioni dovute a malattia.

b) Composizione dei liquidi corporei e loro contenuto ionico
Tra gli ioni del plasma e quelli del liquido interstiziale si viene a creare un equilibrio determinato dalle forze descritte nell'effetto Gibbs-Donnan, prodotte dalla differente concentrazione proteica dei due compartimenti. Le differenze maggiori esistenti tra l'acqua intracellulare e quella extracellulare sono conseguenza dei processi di trasporto attivo a livello della membrana cellulare, i più importanti dei quali sono l'espulsione attiva del sodio dalle cellule ed il mantenimento di un'elevata concentrazione intracellulare di potassio.
L'acqua si sposta passivamente avanti e indietro attraverso le membrane cellulari lungo i gradienti di concentrazione creati dalla distribuzione ionica. È quindi evidente come il mantenimento delle condizioni caratteristiche di concentrazione ionica sia di fondamentale importanza nel determinare i volumi assoluti e relativi dei compartimenti dei liquidi corporei.
La quantità totale di sodio nell'uomo adulto normale è in media di 60 meq/kg di peso corporeo, cioè 4.200 meq per un individuo di 70 kg. Esso è ripartito circa in ugual misura tra il liquido extracellulare (54,5%) e l'osso (43%), mentre solamente il 2,5% circa si trova nel compartimento intracellulare. La concentrazione media del sodio intracellulare risulta essere, secondo i calcoli, di circa 4,2 meq/l di acqua cellulare, ma alcuni tessuti (per es. la corteccia renale) possono averne quantità molto maggiori. Con l'espressione ‛sodio scambiabile' si indica quella frazione del sodio totale che viene subito diluito dal 22Na o dal 24Na: cioè il sodio dei liquidi sia extracellulare sia intracellulare e circa il 25% del sodio del tessuto osseo. Il rimanente sodio del tessuto osseo, in fase cristallina, non è scambiabile. Tutto il sodio scambiabile si trova in equilibrio con il sodio plasmatico ed è perciò disponibile per rimpiazzare il sodio perduto in condizioni anormali dell'organismo, come un'eccessiva sudorazione, diarrea o acidosi. La tab. V riassume la distribuzione del sodio corporeo in un uomo giovane.
La quantità totale di potassio, calcolata mediante analisi diretta dei tessuti e conteggio dell'isotopo naturale 40K contenuto nel corpo, è di circa 54 meq/kg di peso corporeo. La quantità di potassio scambiabile, calcolata dalla diluizione subita dall'isotopo instabile 42K, è in buon accordo con quella ottenuta coi metodi precedenti; questo indica che, a differenza del sodio, la maggior parte del potassio corporeo è scambiabile. Alcune frazioni del potassio totale del corpo, soprattutto quelle presenti negli eritrociti del sangue, nel cervello e nel tessuto osseo, si equilibrano lentamente. Nonostante ciò, si è trovato che la diluizione del 42K, già dopo 48 ore, dà un valore di potassio corrispondente a circa il 90% di quello corporeo complessivo. La tab. VI riassume i valori medi e la distribuzione del potassio in un uomo giovane.
Studi riguardanti la distribuzione del potassio confermano le differenze riscontrate per il sodio con l'età e il sesso, in gran parte dovute al contenuto corporeo di grasso. Il potassio scambiabile è infatti un buon indice della massa cellulare corporea, sebbene una relazione tra la massa cellulare ed i fenomeni legati all'attività metabolica non possa essere costante a causa delle differenti necessità energetiche dei vari gruppi cellulari.
La piccola frazione di potassio extracellulare, circa il 10% del totale, ha una concentrazione di 5-6 meq/l. Questa concentrazione spesso non riflette la situazione del potassio intracellulare, la quale viene generalmente dedotta dalla storia clinica e dalla sintomatologia. Variazioni in più o in meno del 60% nella concentrazione plasmatica di potassio possono indicare variazioni nello stesso senso delle riserve totali corporee, e sono spesso associate a sintomi gravi a livello cardiovascolare e neuromuscolare.
Il cloro è il più abbondante degli anioni del liquido extracellulare. Per quanto riguarda il cloro intracellulare totale, esso è parecchie volte superiore al sodio, nonostante che il muscolo scheletrico e il tessuto connettivo ne contengano molto poco; ciò è dovuto al fatto che esso è invece presente in quantità apprezzabili nelle cellule del sangue e in quelle della mucosa gastrica, che lo secernono attivamente. Il cloro corporeo totale è stato misurato con il metodo della diluizione del 82Br e del 38Cl in parecchie specie di Mammiferi, e si è calcolato che abbia una concentrazione media di circa 33 meq/kg di peso corporeo. Come per il sodio, il contenuto di cloro è significativamente più elevato nell'uomo che nella donna. Il cloro si comporta nello stesso modo del sodio nell'assunzione e nell'eliminazione dal corpo, poiché è sensibile agli stessi fattori che controllano il bilancio del sodio. Esso è caratterizzato da una relazione inversa con il bicarbonato in molti disturbi dell'equilibrio acido-base.
La quantità di bicarbonato scambiabile è in media di 10-12 meq/kg di peso corporeo ed è suddivisa in ugual misura tra il liquido extracellulare e quello intracellulare. La quantità totale di bicarbonato è molto maggiore a causa della gran massa di carbonato presente nel tessuto osseo, il quale può essere liberato solamente per aggiunta di un acido forte. Nel plasma e nel liquido interstiziale questo anione così versatile è attivo soprattutto nel trasporto del diossido di carbonio dai tessuti ai polmoni. La caratteristica alcalinità del sangue di Mammifero è dovuta all'elevato valore di pK dell'acido carbonico, cosicché la riserva extracellulare di bicarbonato costituisce la prima difesa contro un eventuale eccesso di acido all'interno del corpo.
Quanto al magnesio, il valore che è stato recentemente attribuito al ruolo di questo catione prevalentemente intracellulare ha indotto ad approfondirne lo studio e le analisi delle riserve corporee. Widdowson e McCance hanno calcolato che tutto il magnesio presente nel corpo corrisponde a circa 15 meq/kg di peso corporeo. Come per il calcio, si pensa che solamente la frazione ionizzata abbia importanza dal punto di vista fisiologico. L'intossicazione sperimentale da magnesio produce depressione del sistema nervoso centrale e dell'attività cardiaca. L'ipomagnesiemia, cioè una bassa concentrazione ematica di questo ione, produce una ipereccitabilità neuromuscolare, che può svilupparsi in spasmo o tetania.
c) Regolazione del volume dei liquidi corporei
Il peso di un individuo normale adulto si mantiene relativamente costante da un giorno all'altro, nonostante che l'assunzione giornaliera di acqua e di sali possa variare considerevolmente. Poiché il nostro corpo è costituito, come si è visto, in gran parte di acqua, deve evidentemente esistere una regolazione della quantità totale di acqua. Inoltre, poiché l'acqua in condizioni normali non è mai trattenuta nel corpo senza il sodio, la regolazione idrica implica l'esistenza di una regolazione del sodio corporeo totale. Questo è vero nonostante il fatto che più della metà dell'acqua corporea sia intracellulare (v. tab. IV) e il 98% del sodio sia extracellulare (v. tab. V). Il volume del liquido extracellulare è funzione del contenuto corporeo di sodio; questo liquido costituisce l'ambiente in cui si trovano le cellule; il loro contenuto si trova in equilibrio osmotico con esso. La regolazione del volume idrico è quindi strettamente dipendente dalla regolazione del sodio.
L'organo effettore del sistema di controllo regolatore del sodio è il rene. I problemi di interesse primario riguardano il modo con cui le informazioni sulle riserve corporee di sodio giungono al rene, e i meccanismi con i quali esso ne modifica in modo appropriato l'escrezione. Sappiamo che quando il contenuto di sodio nell'organismo è ottimale, il sale ingerito viene eliminato quantitativamente, mentre una riduzione di sale nella dieta viene controbilanciata dall'eliminazione di urine povere di sodio.
Il rene regola l'eliminazione del sodio modificando i suoi due meccanismi fondamentali: la quantità di plasma filtrato a livello glomerulare e il grado di riassorbimento nel passaggio del filtrato lungo i tubuli. Chi non conosca bene lo sviluppo evolutivo del rene di Mammifero può pensare che il processo di formazione dell'urina sia un inutile sperpero di energia. Un uomo di media corporatura filtra oltre un centinaio di litri di plasma nelle ventiquattr'ore, ma il riassorbimento del filtrato a livello dei tubuli è tale che la quantità di urina che si forma può essere inferiore ad un litro. Il sodio eliminato con le urine costituisce la differenza tra la grande quantità di sodio filtrato e quella gran parte di esso ritornata al plasma circolante con il riassorbimento attraverso i tubuli. Piccole variazioni della filtrazione glomerulare o del riassorbimento tubulare hanno effetti notevolmente diversi sulla escrezione di sodio. Se, rispetto ai normali livelli, la velocità di filtrazione glomerulare dovesse aumentare dell'1%, rimanendo costante il riassorbimento, l'escrezione di sodio aumenterebbe di soli 4 meq al giorno. Al contrario, una variazione dell'1% nel grado di riassorbimento tubulare aumenterebbe l'eliminazione di sodio di 200 meq al giorno. Di fatto questi due processi operano probabilmente in serie e sono interdipendenti l'un l'altro. Sperimentalmente sono state dimostrate alcune delle forze che li controllano.
Il fenomeno della regolazione del sodio implica che l'organo effettore, il rene, debba essere direttamente o indirettamente informato della situazione del sodio nel corpo. In altri sistemi biologici di controllo, i quali operano mediante meccanismi a feedback moderanti, si conosce o si suppone la presenza di elementi sensori, di elementi integratori e di elementi che mandano messaggi. Non è stato finora trovato alcun sistema del genere legato al fenomeno della regolazione salina.
I processi di riassorbimento nei tubuli prossimali e distali funzionano in modo da richiamare più del 98% del sodio normalmente filtrato. Il sodio è il più abbondante componente del filtrato ed esso viene riassorbito tramite un processo di trasporto attivo; ne consegue che questo processo rappresenta la più importante delle operazioni del rene richiedenti energia. Aumenti o diminuzioni della escrezione di sodio sono determinati da aggiustamenti della velocità di filtrazione e del riassorbimento prossimale e distale, variazioni così delicate da essere a stento rivelabili con le comuni tecniche di misura.
Un problema nella regolazione salina, derivante dalle variazioni spontanee della filtrazione glomerulare, è stato risolto mediante un meccanismo intrinseco renale, che è di interesse corrente per i fisiologi. La velocità di filtrazione può variare molto nel corso delle attività quotidiane (aumentando dopo i pasti o col riposo, diminuendo con l'eccitamento o nell'attività), e per ragioni che possono essere del tutto indipendenti dal bilancio del sodio. È evidente che queste variazioni spontanee nel grado di filtrazione, rimanendo uguale la quantità di sodio riassorbito, determinerebbero incongrue variazioni nell'eliminazione dello ione. Ciò che in realtà avviene è che il riassorbimento a livello del tubulo prossimale cambia parallelamente alle variazioni di filtrazione, in modo da lasciar fluire verso il segmento distale una frazione costante di filtrato. È stato osservato che questa attività autoregolatrice, definita ‛equilibrio glomerulo-tubulare', è presente nell'animale integro e nei singoli tubuli prossimali perfusi in situ, mentre manca nei tubuli isolati e perfusi in vitro. Si pensa quindi che essa sia controllata da fattori peritubulari, quali la pressione oncotica delle proteine plasmatiche nei capillari peritubulari, la pressione idrostatica negli spazi peritubulari ed altri fattori extracellulari che possono influenzare, direttamente o indirettamente, il trasporto del sodio.
Il tubulo prossimale, fornendo una percentuale fissa del sodio filtrato al segmento distale del nefrone, lascia a quest'ultimo il compito dell'aggiustamento finale dell'escrezione. Il riassorbimento del sodio nel tubulo distale, a differenza del prossimale, non può superare un valore massimo, il quale è dettato dai limiti della sua capacità funzionale.
Quando la quantità iniziale di sale è grande, la percentuale fornita ai tubuli distali supererà la loro capacità di riassorbimento, per cui l'eccesso viene eliminato. Se la quantità iniziale è piccola, il sodio che giunge ai segmenti distali sarà inferiore alla loro capacità massima, e il riassorbimento sarà completo: in tal caso, nelle urine non vi sarà presenza di sodio.
Un importante aumento del riassorbimento distale del sodio è sotto controllo ormonale. L'aldosterone, secreto dalla corticale surrenale, agisce sulle cellule del tubulo distale aumentandone la capacità di riassorbimento del sodio. Lo stimolo che determina la liberazione dell'aldosterone è la renina, una sostanza prodotta da particolari cellule renali, le quali costituiscono il cosiddetto ‛apparato iuxtaglomerulare'. L'ubicazione di queste cellule, in intimo contatto coi tubuli distali e con le arteriole afferenti ai glomeruli, ha portato a due teorie circa la natura dello stimolo per la liberazione della renina. L'apparato iuxtaglomerulare registrerebbe, secondo la prima teoria, il flusso o la pressione di perfusione del glomerulo; secondo l'altra, invece, la concentrazione del sodio nel liquido del tubulo distale. L'apparato è localizzato in modo conveniente per rilevare entrambi questi dati; esistono risultati sperimentali in favore sia dell'una sia dell'altra funzione, ed entrambe, in effetti, possono svolgere un ruolo nei processi di autoregolazione del rene. Poiché la sostanza intermediaria tra la renina e l'aldosterone è il peptide angiotensina, un attivo agente vasocostrittore che può controllare il flusso sanguigno renale e la pressione di filtrazione, ne consegue che la renina può determinare variazioni nell'eliminazione del sodio per molteplici vie. Una di queste vie potenziali, venuta in luce solo recentemente, è collegata con la distribuzione del flusso sanguigno renale a glomeruli con anse di Henle corte o lunghe. I glomeruli disposti vicino alla superficie del rene sono associati a nefroni contenuti per tutta la loro lunghezza nella corteccia renale o nello strato esterno del rene. Esistono prove che in seguito ad un'elevata ingestione di sale questi nefroni ricevono una parte maggiore di filtrato, mentre con bassa ingestione di sale la distribuzione del filtrato avviene a favore di nefroni le cui lunghe anse discendono in profondità nella regione midollare. Attualmente si sta studiando attivamente il significato di tale differenza in relazione all'economia salina dell'organismo, ed il ruolo del sistema renina-angiotensina nel suo controllo.
5. Sopravvivenza in assenza di acqua
L'uomo sopporta la mancanza di acqua meno della maggior parte degli animali terrestri. La massima perdita di peso nell'uomo prima che sopravvenga la morte per disidratazione è di circa il 20% del peso iniziale. Questo valore sale al 30% nel ratto e nel topo, al 40% nel cane e nel gatto, al 50% negli Uccelli e nei Rettili e ad oltre il 60% nei lombrichi. La velocità di disidratazione è importante quanto il valore assoluto. Se la disidratazione è lenta, la rana, ad esempio, può sopportare una perdita di peso del 40%, ma se questa avviene rapidamente, essa può tollerare solo una perdita del 12%.
Il limite massimo di sopravvivenza per l'uomo in assenza di acqua e nelle condizioni più favorevoli (protezione dal sole, temperatura ambiente mite) è tra gli 11 e i 20 giorni. Camminando sotto il sole con temperature elevate, come nelle zone desertiche, il limite di sopravvivenza diventa una questione di ore, poiché la perdita di acqua in queste condizioni è di circa 1 litro all'ora.
I sintomi da mancanza d'acqua compaiono rapidamente nei climi tropicali, più lentamente nelle zone temperate. E. F. Adolph, nel suo classico studio Physiology of man in the desert (1947), ha trovato che il primo sintomo osservato più frequentemente negli uomini che camminano nel deserto privi di acqua è la debolezza, e non la sete. Con un deficit d'acqua di circa il 3% del peso corporeo incomincia a diminuire la secrezione salivare e la produzione di urina. Una disidratazione superiore al 5% del peso corporeo è tollerata senza che avvengano variazioni fisiologiche serie, sebbene si manifesti un leggero aumento della frequenza cardiaca, compaiano stanchezza e sete, e la temperatura rettale aumenti di 0,5-1 °C. Con un deficit del 10% compaiono alterazioni mentali e la prostrazione fisica è imminente. Col 12% si ha difficoltà ad inghiottire, e tra il 15% ed il 20%, a seconda della velocità di disidratazione e delle condizioni climatiche, sopravviene la morte. La causa della morte per disidratazione non si conosce esattamente. Un fattore importante è certamente lo scompenso circolatorio risultante dalla diminuzione del volume dei liquidi. Kerpel-Fronius attribuisce la morte al danno cellulare prodotto dall'aumento della pressione osmotica dei liquidi tessutali.
a) Disturbi da disidratazione
Sono stati osservati, in condizioni standard di esercizio, gli effetti fisiologici derivanti da situazioni controllate di disidratazione. Una disidratazione acuta pari al 5% del peso corporeo aumentava di 0,5 °C la risposta della temperatura rettale ad un esercizio moderato, e di 30 battiti al minuto la risposta della frequenza del polso. Contemporaneamente il consumo massimo di ossigeno diminuiva di circa 200 ml/minuto. Né la temperatura rettale né la frequenza del polso hanno la normale tendenza a raggiungere un plateau nel periodo finale dell'esercizio. Tale incapacità di mantenere condizioni di stabilità durante un esercizio moderato conferma l'opinione comune circa la gravità di deficit d'acqua anche leggeri in condizioni ambientali sfavorevoli. Lo sforzo fisico diventa impossibile nel giro di poche ore.
b) Sopravvivenza in mare
Una persona che in mare si trovi in una piccola imbarcazione o su una zattera sprovvista di acqua dolce è in una situazione molto simile a quella dell'uomo nel deserto. Il primo è avvantaggiato dal fatto che, nei climi caldi, egli può inumidire periodicamente la pelle e gli abiti con l'acqua di mare in modo da ridurre la perdita di acqua che si avrebbe con l'evaporazione necessaria al raffreddamento del corpo. Scarsa, o nulla, è la quantità di acqua assorbita attraverso la cute, nonostante permanga la credenza comune secondo cui essa potrebbe penetrare per questa via.
L'ingestione di acqua marina non costituisce un'efficace fonte di acqua, come può risultare dalle seguenti semplici considerazioni. L'acqua degli oceani contiene circa il 3,5% di sale, mentre il rene umano non può produrre urina con una salinità superiore al 2%. Si può facilmente calcolare che per eliminare tutto il sale contenuto in 100 ml di acqua marina ingerita, sono necessari 175 ml di urina alla massima concentrazione urinaria di sodio. Si ha quindi una perdita netta di 75 ml d'acqua. Inoltre il contenuto di magnesio dell'acqua marina provoca diarrea, con ulteriore grave perdita d'acqua da parte del corpo.
Sebbene l'esperienza abbia dimostrato che i naufraghi che bevono acqua di mare muoiono prima degli altri, in letteratura si rivendicano di tanto in tanto gli effetti benefici di tale pratica. Vi sono dei motivi per la persistenza di questa credenza. Il primo è che certi mari sono meno salati; il Mar Nero ed il Caspio, per esempio, hanno una concentrazione salina pari alla metà di quella dell'Atlantico, ed il Mar Baltico è pressoché isotonico. I grandi fiumi riversano enormi quantità d'acqua dolce nei mari, diluendone il contenuto salino per una distanza di molte miglia dagli estuari. Inoltre, con climi eccessivamente caldi, come hanno sperimentato i marinai del Kon Tiki, la sudorazione fa perdere tanto sale che può essere realmente benefica la sua reintegrazione mediante piccole quantità d'acqua marina, se con essa si ingerisce abbastanza acqua dolce.
Il pesce, mangiato senza accompagnamento di acqua dolce, non porta alcun vantaggio per l'economia idrica del corpo, poiché tutto il contenuto d'acqua del pesce ingerito è necessario all'eliminazione dei suoi metaboliti proteici e dei sali. Benché si sia spesso sostenuto che il succo estratto dal pesce sia una sorgente d'acqua per i naufraghi, la maggior parte delle prove è a favore della conclusione opposta, che esso sia, cioè, realmente deleterio.
c) Trattamento dell'acqua di mare per renderla potabile
Sono stati ideati parecchi tipi di apparecchi di distillazione, dei quali alcuni utilizzano i raggi del sole, con lo scopo di trasformare l'acqua di mare in acqua dolce potabile da utilizzare nelle scialuppe di salvataggio e nelle zattere. Tali apparecchi sono stati abbandonati, a causa del loro ingombro, della complessità e del basso rendimento, a favore di metodi di purificazione chimica. Questi ultimi sono stati considerevolmente semplificati per la convinzione a cui si è giunti che l'acqua leggermente salata (0,3-0,4% di NaCl) sia in realtà più gradevole e permetta una migliore conservazione dell'acqua di quanto lo sia l'acqua chimicamente pura. Queste considerazioni e la scoperta che il solfato dell'acqua di mare (circa 50 meq/l) non è dannoso per l'organismo, hanno permesso la messa a punto del cosiddetto metodo di dissalazione chimica alla Permutite ad un solo stadio.
L'unità base è costituita da una tavoletta di zeolite d'argento delle dimensioni di un pacchetto di sigarette, e da un sacchetto di plastica pieghevole con un filtro all'interno. La zeolite d'argento è una resina sintetica del tipo del silicato d'alluminio idrato [Ag2Al2Si3O10(H2O)2]. La tavoletta viene immessa nel sacchetto di plastica contenente acqua marina e rotta con le mani. Avviene quindi la precipitazione del cloruro d'argento e la formazione della zeolite di sodio e magnesio, mentre l'acqua divenuta così potabile viene raccolta aspirandola con la bocca attraverso il filtro da un tubo di uscita. Questo sistema fornisce una quantità di acqua potabile sette volte superiore al peso di un singolo apparecchio.
d) L'acqua nello spazio
Con la prospettiva immediata dei voli spaziali dell'uomo verso la luna e oltre, balza in primo piano il problema del rifornimento d'acqua nello spazio. È escluso che si possano trasportare riserve d'acqua, data l'attuale scarsità di spazio disponibile. L'unica soluzione attualmente possibile è la riutilizzazione dell'acqua eliminata nei processi vitali. Infatti costituiscono un'importante fonte di acqua sia quella espirata, e condensata sulle pareti di una navicella spaziale, sia la stessa urina, una volta sottoposta ad un processo di riciclizzazione. Tra i metodi di riciclizzazione sperimentati negli ultimi anni, la liofilizzazione, con successivo passaggio su colonna di carbone attivato, offre le maggiori possibilità di successo. Buone prospettive offre il sistema di distillazione sotto vuoto facendo passare il vapore attraverso un catalizzatore ad alta temperatura per ‛pirolizzare' le impurità volatili. L'assoluta dipendenza dell'uomo da un rifornimento d'acqua costante potrebbe dimostrarsi al limite il più grosso ostacolo per l'esplorazione dello spazio su vasta scala.
6. Conclusione
Abbiamo accennato alla struttura di questa importante sostanza, l'acqua, e discusso brevemente le proprietà responsabili del ruolo peculiare che essa ha avuto nella creazione e nella conservazione della vita. L'acqua fu la sostanza chiave dell'evoluzione e la forza modellatrice della Terra. L'affermazione degli esseri viventi è stata in funzione della loro capacità di provvedersi d'acqua, conservarla per mezzo di complessi strumenti biologici e di regolarne il movimento all'interno dei compartimenti corporei. Il progresso tecnologico dell'ultimo secolo minaccia ora di esaurire le riserve d'acqua potabile, un tempo illimitate. Il compito di conservare e ricostituire l'ambiente idrico costituisce ora una sfida all'inventiva dell'uomo, e dal successo di questo suo tentativo dipende la continuazione della vita.
bibliografia
Adolph, E. F., Measurement of water drinking in dogs, in ‟American journal of physiology", 1939, CXXV, pp. 75-86.
Adolph, E. F., Physiology of man in the desert, New York 1947.
Andersson, B., Thirst and brain control of water balance, in ‟American scientist", 1971, LIX, pp. 408-415.
Andersson, B., Larsson, B., Influence of local temperature changes in the preoptic area and rostral hypothalamus on the regulation of food and water intake, in ‟Acta physiologica scandinavica", 1961, LI, pp. 75-89.
Bentzel, C. J., Solomon, A. K., Osmotic properties of mitochondria, in ‟Journal of general physiology", 1967, L, pp. 1547-1563.
Bernard, Cl., Leçons sur les phénomènes de la vie communs aux animaux et aux végétaux, Paris 1878.
Bombard, A., La survie prolongée en mer. Rapport technique de l'expérience de survie prolongée en mer à bord de l'Hérétique, en 1952, Paris 1954.
Code, Ch. F., Bass, P., MacClary, G. B., Newmum, L. R., Orvis, A. L., Absorption of water, sodium and potassium in small intestine of dogs, in ‟American journal of physiology", 1960, CIC, pp. 281-288.
Cook, J. S., Non-solvent water in human erythrocytes, in ‟Journal of general physiology", 1967, L, pp. 1311-1325.
Diamond, J. M., The coupling of solute and water transport in epithelia, in Renal transport and diuretics. International symposion (a cura di K. Thurau e H. Jahrmärker), Berlin 1969, pp. 77-97.
Edelmann, I. S., James, A. H., Brooks, L., Moore, F. D., Body sodium and potassium. IV. The normal exchangeable sodium, its measurement and magnitude, in ‟Metabolism", 1954, III, pp. 530-538.
Edelmann, I. S., Leibman, J., Anatomy of body water and electrolytes, in ‟American journal of medicine", 1959, XXVII, pp. 256-277.
Fenn, W. O., Noonan, T. R., Mullins, L. J., Haege, L. F., The exchange of radioactive potassium with body potassium, in ‟American journal of physiology", 1941, CXXXV, pp. 149-163.
Gauer, O. H., Henry, J. P., Circulatory basis of fluid volume control, in ‟Physiological reviews, 1963, XLIII, pp. 423-481.
Gregersen, M. I., Blood volume, in ‟Annual review of physiology", 1951, XIII, pp. 397-412.
Keck, W., Brechtelsbauer, H., Kramer, K., Wasser- und Na-Ausscheidung nach isotonischen Kochsalz-Infusionen bei wachen Hunden mit verschiedenem Natriumbestand, in ‟Pflügers Archiv", 1969, CCCXI, pp. 119-130.
Ladell, W. S. S., The effects of water and salt intake upon the performance of men working in hot and humid environments, in ‟Journal of physiology", 1955, CXXVII, pp. 11-46.
Ling, G. N., Ochsenfeld, M. M., Karreman, G., Is the cell membrane a universal rate-limiting barrier to the movement of water between the living cell and its surrounding medium?, in ‟Journal of general physiology", 1967, L, pp. 1807-1820.
Manery, J. F., Danielson, I. S., Hastings, A. B., Connective tissue electrolytes, in ‟Journal of biological chemistry", 1938, CXXIV, pp. 359-375.
Miller, H., Wilson, G. M., The measurement of exchangeable sodium in man using the isotope 24Na, in ‟Clinical science", 1958, XII, pp. 97-111.
Moore, F. D., Determination of total body water and solids with isotopes, in ‟Science", 1946, CIV, pp. 157-160.
Pappenheimer, J. R., Renkin, E. M., Borrero, L. M., Filtration, diffusion and molecular sieving through peripheral capillary membranes, in ‟American journal of physiology", 1951, CLXVII, pp. 13-15.
Pauling, L., The nature of the chemical bond, Ithaca, N. Y., 19603 (tr. it.: La natura del legame chimico, Roma 19612).
Peters, J. P., Water exchange, in ‟Physiological reviews", 1944, XXIV, pp. 491-531.
Pfeffer, W., Osmotische Untersuchungen, Leipzig 19212.
Robinson, E. A., Adolph, E. F., Pattern of normal water drinking in dogs, in ‟American journal of physiology", 1942, CXXXIX, pp. 39-44.
Robinson, S., Robinson, A. H., Chemical composition of sweat, in ‟Physiological reviews", 1954, XXXIV, pp. 202-220.
Smith, D. H., Water movement across the mammalian gut, in ‟Symposium of the Society for Experimental Biology", 1965, XIX, pp. 307-328.
Smith, H. W., From fish to philosopher. The story of our internal environment, Boston 1953.
Steele, J. M., Berger, E. Y., Dunning, M. F., Brodie, B. B., Total body water in men, in ‟American journal of physiology", 1950, CLXII, pp. 313-317.
Ullrich, K. J., Kramer, K., Boylan, J., The countercurrent system in the mammalian kidney, in ‟Progress in cardiovascular diseases", 1961, III, pp. 395-405.
Verney, E. B., Antidiuretic hormone and the factors which determine its release, in ‟Proceedings of the Royal Society" (Series B), 1947, CXXXV, pp. 25-106.
Widdowson, E. M., McCance, R. A., Spray, C. M., The chemical composition of the human body, in ‟Clinical science", 1951, X, pp. 113-125.
Wolf, A. V., Thirst, Springfield, Ill., 1958.