
Apprendimento sociale e imitazione
Apprendimento sociale e imitazione
A partire dalla fine degli anni Ottanta, i ricercatori che lavorano nei diversi settori dell'antropologia, della psicologia e della biologia hanno compiuto importanti passi in avanti nello studio delle forme di apprendimento sociale, sia di tipo imitativo sia di tipo non imitativo. È infatti ormai noto che in molte specie di vertebrati l'apprendimento sociale non imitativo svolge un ruolo importante per quanto riguarda la riproduzione, l'acquisizione di cibo e i comportamenti necessari per evitare i predatori. Sono stati anche compiuti alcuni studi i quali dimostrano come non solo le grandi scimmie antropomorfe possono apprendere per imitazione; alcuni uccelli e taluni mammiferi non appartenenti all'ordine dei primati sembrano infatti in grado di imitare semplici atti motori dopo averli osservati in altri individui.
Perché studiare l'apprendimento sociale
La maggior parte degli scienziati che si dedica all'apprendimento sociale negli animali si interessa a uno dei due seguenti problemi. l primatologi e gli psicologi spesso cercano di scoprire se gli animali diversi dall 'uomo possono imitare dei comportamenti che hanno visto eseguire da altri. Per questi ricercatori, quando un animale impara una particolare azione, semplicemente osservando un altro animale che la esegue, in quel momento offre la stimolante opportunità di studiare le capacità cognitive di specie diverse dalla nostra. Per altri ricercatori lo studio dell'apprendimento sociale negli animali riflette un interesse più generale nell'ecologia comportamentale. Ciò li porta a studiare l'apprendimento sociale per capire in che modo le informazioni acquisite da altri individui contribuiscono allo sviluppo di sequenze comportamentali adattive in animali che vivono in natura. Questi ricercatori sono interessati alle interazioni sociali che guidano il comportamento in direzioni utili. Per costoro non è quindi importante se la trasmissione di comportamenti da un animale all'altro avvenga per mezzo dell' imitazione o di qualche processo di apprendimento sociale di tipo non imitativo e meno impegnativo dal punto di vista cognitivo. Entrambi questi approcci allo studio dell'apprendimento sociale sono utili ed entrambi hanno una tradizione lunga e autorevole nelle scienze della vita.
Dalla fine del 19° secolo sono stati compiuti vari tentativi per defmire con precisione i diversi tipi di apprendimento sociale e per classificarne vari esempi sulla base dei processi di apprendimento dai quali dipendono. L'apprendimento sociale a volte sembra essere prodotto da un processo di vera e propria imitazione (imparare a compiere un particolare atto motorio dopo averlo visto effettuare da altri); altre volte sembra invece dipendere da un processo di emulazione (imparare le cose necessarie per riuscire a risolvere un compito attraverso l'osservazione del comportamento di un modello, senza tuttavia imparare il comportamento del modello in sé e per sé; Tomasello, 1996), di incentivazione dell'interesse (imitazione apparente che risulta dal dirigere l'attenzione dell' animale verso un particolare oggetto o parte dell'ambiente; Thorpe, 1963), o da altri processi comportamentali rigorosamente definiti. Esistono vari esempi di tassonomia dei processi implicati nell'apprendimento sociale (Galef, 1988b; Whiten e Ham, 1992). Probabilmente questi diversi tipi di apprendimento sociale richiedono gradi diversi di sofisticazione dei meccanismi cognitivi. Per questo motivo, scoprire quale tipo di apprendimento sociale è coinvolto nella trasmissione di comportamenti da un individuo all'altro ha implicazioni importanti per lo studio delle capacità cognitive di specie diverse dalla nostra.
Dal punto di vista storico, la distinzione più importante, e l'unica che sarà menzionata in questo saggio, riguarda la possibilità di determinare, in ogni situazione specifica, se un osservatore impara o meno direttamente qualcosa riguardo al comportamento del modello che osserva. Nei vari tipi di apprendimento sociale non imitativo, un animale che guarda un altro animale comportarsi in un certo modo e osserva il risultato prodotto da quel comportamento, impara qualcosa di utile riguardo agli aspetti dell' ambiente che sono rilevanti per il compito in questione, oppure impara quali cambiamenti dell'ambiente siano causati dal comportamento dell'animale che agisce (Heyes, 1996; Tomasello, 1996). Per esempio, uno scimpanzé che osserva un altro scimpanzé andare a caccia di larve di insetti capovolgendo ceppi di legno con la mano sinistra potrebbe imparare che i ceppi sono oggetti interessanti (imparare qualcosa riguardo al proprio ambiente), oppure che, quando un ceppo viene capovolto, compare del cibo (imparare qualcosa sui cambiamenti nell'ambiente causati dal comportamento), oppure che infilare la mano sinistra sotto un ceppo e muoverla rapidamente verso l'alto è un comportamento che è ricompensato con il cibo (imparare qualcosa riguardo al comportamento di chi agisce). Se si utilizza la definizione più attuale di imitazione (Heyes, 1996; Tomasello, 1996), soltanto quest'ultimo tipo di apprendimento, in cui lo scimpanzé che osserva impara direttamente qualcosa riguardo al comportamento dello scimpanzé che agisce, implicherebbe la presenza di un processo di imitazione.
Questo 'vero' apprendimento imitativo sembra richiedere che un animale, o un essere umano, immagazzini una rappresentazione visiva della configurazione dei movimenti eseguiti da un altro. Dal momento che colui che imita è raramente in grado di vedere i propri movimenti in modo da poterli confrontare con la rappresentazione visiva dell' atto da imitare immagazzinata in memoria, l'imitatore deve utilizzare le proprie sensazioni propriocettive e cinestesiche per produrre movimenti sulla base dell'informazione contenuta in memoria. Questo tipo di confronto di informazioni provenienti da diverse modalità sensoriali è considerato un compito cognitivo difficile da risolvere. È per questo che la presenza di un tale processo consente di distinguere l'apprendimento imitativo vero e proprio da altre forme potenzialmente meno complesse di apprendimento sociale non imitativo.
Apprendimento sociale non imitativo
Forse il modo più semplice per presentare gli ultimi progressi compiuti nello studio dell'apprendimento sociale non lmltativo consiste nel descrivere, anche solo brevemente, qualche esempio di studi recenti sui comportamenti appresi socialmente che sono potenzialmente in grado di contribuire alla sopravvivenza o al successo riproduttivo. Gli esempi che seguono sono stati scelti per mostrare sia la gamma di comportamenti che subisce l'influenza di forme non imitative di apprendimento sociale, sia le diverse modalità con cui l'informazione acquisita dagli altri può facilitare l'acquisizione di sequenze adattive di comportamenti.
Imparare che cosa mangiare: come Rattus norvegicus seleziona il cibo ed evita le sostanze tossiche
Alcuni decenni fa uno studioso di ecologia applicata, cercando di migliorare l'efficacia dei metodi di disinfestazione dai roditori, scoprì che l'uso ripetuto di un solo tipo di esca avvelenata in una popolazione di ratti (Rattus norvegicus) faceva fallire miseramente qualsiasi tentativo di controllarne il numero di individui. Subito dopo l'introduzione delle esche avvelenate nell' area in cui vivevano le popolazioni sotto esame, si poteva osservare una diminuzione nel numero di ratti. Tuttavia, alcune popolazioni, scelte come popolazioni focali e ripetutamente esposte a una stessa esca, ripristinavano presto le proprie dimensioni originali. l motivi per cui l'operazione falliva in seguito all'uso ripetuto della stessa esca in una stessa area risultarono abbastanza semplici da capire. lnnanzitutto, nonostante subito dopo l'introduzione delle esche nel territorio della colonia la maggior parte dei ratti ne mangiasse dosi letali, in molte colonie accadeva che alcuni membri, alloro primo incontro con l'esca, ne mangiavano solo una piccola quantità che bastava a farli ammalare ma non a ucciderli. l ratti che sopravvivevano imparavano in una singola occasione ad associare il sapore dell' esca ai suoi effetti tossici e quindi a evitarla in seguito. Avveniva inoltre qualcosa di ancora più scoraggiante: i giovani ratti che raggiungevano la fase di svezzamento in colonie i cui membri erano sopravvissuti al loro primo incontro con l'esca avvelenata rifiutavano anch'essi l'esca che gli altri membri della colonia avevano imparato a evitare e mangiavano solo i cibi che i sopravvissuti stavano mangiando. Il comportamento di evitare la sostanza tossica, appreso dagli individui che sopravvivevano, era in qualche modo trasmesso ai giovani della colonia.
Evitare l'esca avvelenata, comportamento appreso socialmente dai ratti durante lo svezzamento, è un fenomeno rilevante e facile da studiare in laboratorio (Galef, 1988a). Per questo motivo è stato possibile studiare il processo di apprendimento sociale che causa la trasmissione delle scelte alimentari dagli adulti ai giovani della propria specie.
Potenziali influenze prenatali nelle preferenze alimentari. - Anche prima della nascita un giovane ratto può acquisire informazioni riguardo al cibo mangiato dalla madre, almeno per alcuni tipi di cibo. P.G. Hepper (1988) nutriva con aglio femmine di ratto in stato avanzato di gestazione e meno di un'ora dopo il parto assegnava i piccoli a madri adottive che non avevano mai mangiato aglio. Quando i piccoli allevati dalle madri adottive avevano 12 giorni, e ancora si nutrivano esclusivamente di latte materno, Hepper offriva loro due piatti tra cui scegliere, l'uno contenente aglio e l'altro cipolla; lo studioso ha scoperto così che i piccoli nati da femmine che avevano ingerito aglio mentre erano incinte si mantenevano nelle vicinanze del piatto con l'aglio, mentre i piccoli nati da femmine che non avevavo mangiato aglio durante la gravidanza non mostravano una preferenza per l'aglio rispetto alla cipolla.
Informazioni sul sapore contenute nel latte materno. - Il tipo di cibo che una femmina di ratto mangia mentre allatta può modificare il sapore del proprio latte e il sapore del latte con cui un piccolo è stato nutrito influenza le sue preferenze alimentari quando è svezzato. Per esempio, i piccoli di ratto, una volta svezzati, mostrano una preferenza verso il tipo di cibo di cui si nutre una femmina che li allatta per parecchie ore, mentre non la mostrano quando interagiscono, per lo stesso numero di ore, con una femmina che si nutre di quel tipo di cibo e che si comporta in modo materno verso di loro, senza però allattarli (Galef, 1977).
Effetti osservati nel periodo dello svezzamento. - B.G.Galef Jr. e D. Clark hanno utilizzato telecamere a circuito chiuso e videoregistrazioni accelerate per effettuare delle osservazioni su nove ratti selvatici, appartenenti a tre diverse lettiere, durante il loro primo pasto con cibo solido. Tutti i piccoli venivano osservati esattamente nelle stesse circostanze mentre mangiavano per la prima volta. Ciascuno mangiava in prossimità di un adulto che mangiava a sua volta; nessuno di essi mangiava in posti dove non era presente un adulto nelle vicinanze. Apparentemente, la presenza di un ratto adulto rendeva una particolare postazione di foraggiamento attraente per i piccoli e faceva aumentare notevolmente la probabilità di imparare a mangiare il tipo di cibo che vi si trovava (Galef, 1977). Anche se si anestetizzava un ratto adulto e lo si collocava in stato di incoscienza in prossimità di una postazione di foraggiamento, quest'ultima diventava significativamente più attraente per i piccoli nella fase di svezzamento rispetto ad altre postazioni che non avevano un adulto nelle vicinanze. Tuttavia, questo non significa necessariamente che i ratti adulti devono essere fisicamente presenti nelle vicinanze per guidare i piccoli verso una postazione di foraggiamento. Mentre mangiano, i ratti depositano infatti tracce olfattive sia nelle vicinanze di una fonte di cibo, sia sul cibo che mangiano, sia lungo il percorso effettuato per tornare alla tana dopo avere mangiato. Tutti questi odori risultano attraenti per i giovani ratti e li inducono ad avvicinarsi e a preferire le postazioni di foraggiamento utilizzate dagli adulti (Galef, 1977).

Effetti dopo lo svezzamento. - Galef, con i suoi studenti, ha anche scoperto che un giovane ratto (l'osservatore), dopo aver interagito per qualche minuto con un conspecifico che ha appena mangiato del cibo (il dimostratore), mostra un sostanziale incremento della preferenza verso il tipo di cibo mangiato dal conspecifico (Galef, 1988a; 1996). Lo studio dei processi responsabili che inducono socialmente queste preferenze alimentari è iniziato con la seguente scoperta: quando il ratto osservatore veniva esposto a un dimostratore anestetizzato, la cui testa era stata cosparsa con un leggero strato di cibo in polvere (fig. 1), esso aumentava la propria preferenza per il cibo che trovava sulla testa del ratto che agiva da dimostratore. D'altro canto, l'esposizione a batuffoli di ovatta impolverata con frammenti di cibo non faceva sviluppare una preferenza alimentare in questi osservatori (Galef, 1988a). Chiaramente, qualcosa che si riferisce alla presenza di un ratto dimostratore insieme a un certo tipo di cibo è in grado di modificare le preferenze alimentari di un ratto osservatore.
Sono stati compiuti esperimenti per scoprire quali sono gli stimoli emessi dai ratti dimostratori che determinano le preferenze alimentari negli osservatori. Si è scoperto che la presenza congiunta di uno dei costituenti dell'alito del ratto, il disolfuro di carbonio, e di un particolare tipo di cibo produce lo stesso effetto sulle preferenze alimentari che assaggiare il cibo in presenza di un ratto che respira (Galef, 1996).
Imparare a nutrirsi: lo sfruttamento delle pigne nei ratti neri
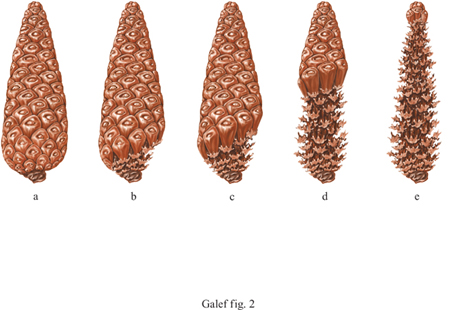
Le pinete israeliane sono abitate da colonie di ratti neri (Rattus rattus) che hanno una dieta composta quasi esclusivamente di pinoli e acqua (Terkel, 1995). L'estrazione dei pinoli dalle pigne è un comportamento stabile in queste colonie di ratti e permette loro di sopravvivere in aree dove i pinoli sono l'unico tipo di cibo presente in quantità sufficiente a nutrire una popolazione di mammiferi. Osservazioni di laboratorio sono state condotte sul comportamento alimentare di ratti prelevati da colonie che vivono nelle pinete. Queste osservazioni mostrano che il metodo di foraggiamento che permette ai ratti di ricavare più energia dai pinoli di quanta non ne perdano per rimuovere le squame dure e non commestibili della pigna è basato sullo sfruttamento della struttura della pigna. Le squame alla base della pigna vanno rimosse per prime, poi vanno rimosse le altre, seguendo la disposizione a spirale fino a raggiungere la sommità della pigna (fig. 2).

Osservazioni condotte in cattività su ratti prelevati da aree diverse da quelle delle pinete e ai quali erano state offerte delle pigne, hanno rivelato che solo 6 ratti adulti su 222, sebbene affamati, apprendevano autonomamente a utilizzare il percorso a spirale necessario a rimuovere le squame delle pigne, comportamento che consente un guadagno netto di energia nutrendosi di pinoli. D'altro canto, praticamente tutti i ratti allevati da adulti capaci di estrarre efficientemente i pinoli dalle pigne hanno acquisito la tecnica appropriata. Alcuni aspetti dell'interazione tra le madri capaci di estrarre i pinoli dalle pigne e i piccoli da loro allevati sembrano essere importanti per la trasmissione di questa tecnica da una generazione all'altra. Altri esperimenti, condotti successivamente, hanno dimostrato che, quando giovani ratti fanno esperienza completando l'operazione di rimuovere le squame da pigne già parzialmente utilizzate da un ratto adulto (fig. 3), più del 70% di essi apprende il modo più efficace di intaccare le pigne (Terkel, 1995).
La tradizione dello sfruttamento delle pigne che si osserva nei ratti neri che abitano le pinete è particolarmente interessante perché permette loro di prosperare in un ambiente relativamente sterile, che altrimenti gli sarebbe precluso. In questo caso particolare, e forse anche in altri, l'apprendimento sociale apre una nuova nicchia ecologica ad alcuni membri di specie selvatiche.
Imparare che cosa temere: come le scimmie evitano i serpenti
È difficile capire come gli animali apprendano a evitare i predatori. Innanzitutto, se si considera l'ampia gamma di predatori a cui molti animali sono vulnerabili, sembra quasi impossibile capire come possa essersi sviluppato un sistema innato per distinguere tutti gli animali innocui da quelli potenzialmente pericolosi. In secondo luogo, imparare a evitare i predatori attraverso l'esperienza individuale non può funzionare bene. I predatori devono infatti essere evitati fin dalla prima occasione in cui li si incontra. Non vi è tempo di apprendere per prove ed errori. È noto ormai dagli anni Cinquanta che, al contrario delle scimmie antropomorfe e non antropomorfe nate in libertà e trasferite in laboratorio, le quali evitano vigorosamente il contatto con i serpenti e con gli oggetti che possono sembrare serpenti, le scimmie nate e allevate in cattività sono relativamente indifferenti a quegli stessi oggetti.
S. Mineka e M. Cook (1988) hanno studiato le risposte ai serpenti in scimmie allevate in cattività e l'influenza derivata dall' osservazione di scimmie nate in libertà che mostrino di avere paura dei serpenti. I due ricercatori hanno riscontrato che le scimmie allevate in laboratorio, che all'inizio non reagiscono, o reagiscono poco, alla presenza di serpenti, dopo avere osservato le risposte esibite dalle scimmie nate in libertà imparano a reagire vigorosamente alla vista di un serpente. Questo effetto di reazione ai serpenti delle scimmie allevate in laboratorio non sembra attenuarsi neanche dopo tre mesi.
È significativo notare che le scimmie non apprendono risposte emotive per nessun altro oggetto verso cui abbiano osservato un conspecifico reagire. Cook e Mineka hanno mostrato a scimmie allevate in laboratorio videoregistrazioni di altre scimmie che esibivano il tipo di risposta suscitato dalla vista di un serpente; l'esperimento è stato compiuto dopo aver effettuato un montaggio in modo da far sembrare che le scimmie a volte mostrassero paura nei confronti dei serpenti, a volte nei confronti dei fiori. Le scimmie allevate in laboratorio imparavano ad avere paura dei serpenti, ma non dei fiori. Sembra dunque che le scimmie imparino selettivamente ad associare oggetti che sembrano serpenti con le risposte di paura esibite da altri membri della propria specie.
Imparare con chi accoppiarsi: la scelta del partner nel pesce guppy
È noto già dagli anni Settanta che l'apprendimento sociale gioca un ruolo importante nello sviluppo delle configurazioni adattive di risposte nell'ambito delle scelte alimentari e nell'evitare i potenziali predatori. Tuttavia, sono stati raccolti più recentemente risultati a favore dell'idea che l'apprendimento sociale influenzi direttamente anche la scelta del partner sessuale. L.A. Dugatkin e i suoi collaboratori (1996) hanno esaminato il ruolo giocato dall'apprendimento sociale nella scelta del compagno effettuata dai parenti selvatici del guppy da acquario (Poecilia reticulata). Questa specie di pesci è stata scelta come oggetto di studio per due ragioni: innanzitutto, le condizioni in cui questi pesci si accoppiano nei corsi d'acqua di Trinidad (dove i pesci utilizzati in questo esperimento erano stati catturati) rendono possibile un effetto delle influenze sociali nella scelta del compagno. In secondo luogo, questi pesci sono relativamente facili da allevare, mantenere e osservare in laboratorio. Negli esperimenti di Dugatkin ai lati di un acquario erano collocati due compartimenti stagni e ciascun membro di una coppia di pesci maschi, simili per dimensione e vivacità dei colori (qualità che attraggono le femmine di questa specie), era collocato in uno dei compartimenti. Una femmina (il soggetto) era quindi collocata in un contenitore trasparente al centro dell'acquario e una seconda femmina (il modello) era segregata, per mezzo di una barriera di vetro, a un'estremità dell' acquario, in vicinanza di uno dei maschi. Una volta messa a punto la situazione sperimentale, il soggetto femmina poteva osservare il maschio corteggiare la femmina modello vicina per 10 minuti. Alla fine del periodo, la femmina modello era rimossa dell'acquario, la femmina soggetto veniva rilasciata dalla porzione dell'acquario in cui era confinata ed era lasciata libera di scegliere tra uno dei due maschi che si trovavano nei loro compartimenti alle estremità dell'acquario. Dugatkin ha osservato 20 femmine mentre sceglievano un maschio in questa situazione. Di queste, 17 sceglievano di stare nelle vicinanze del maschio che avevano visto corteggiare la femmina modello.
I risultati di questo primo esperimento sono compatibili con l'ipotesi che le femmine di guppy copiano vicendevolmente la scelta di un partner. Tuttavia, esistono varie spiegazioni alternative che vanno esaminate prima di poter accettare l'ipotesi. Dal momento che questa specie di pesci, in natura, tende a formare branchi, le femmine potrebbero mostrare una preferenza verso un'area dove precedentemente hanno visto due pesci piuttosto che verso quella dove hanno visto un pesce soltanto. Tuttavia, in un esperimento del tutto identico a quello appena descritto, tranne per il fatto che i compartimenti posti alle estremità dell'acquario contenevano altre femmine invece di maschi, le femmine soggetto hanno scelto le femmine nel compartimento vicino a quello in cui era stato confinato il modello femmina soltanto 10 volte su 20, effettuando cioè le proprie scelte in modo casuale (Dugatkin, 1996).
I dati di Dugatkin dimostrano che i fattori sociali esercitano una forte influenza sulla scelta del compagno da parte delle femmine di guppy. I risultati di Dugatkin sono anche compatibili con le spiegazioni avanzate per alcuni interessanti fenomeni comportamentali osservati in circostanze naturali. Per esempio, in specie come il gallo della salvia, in cui i maschi competono direttamente per le femmine su territori comuni (lek), ogni giorno solo pochi maschi possono avere accesso a tutte le femmine. Informazioni dettagliate riguardo alle circostanze nelle quali i galli della salvia si accoppiano in natura sono compatibili con l'ipotesi che le femmine osservano e copiano le scelte del maschio effettuate dalle altre, provocando ogni giorno una quasi totale unanimità nella scelta di un maschio da parte di tutte le femmine (Gibson et al., 1991).
Ancora più sorprendenti sono le osservazioni secondo le quali i maschi di alcune specie di pesci corteggiano e si accoppiano rapidamente con le femmine di altre specie, anche se con l'accoppiamento essi non contribuiscono in nessun modo al materiale genetico della prole della femmina. I. Schlupp, C. Marler e M.J. Ryan (1994), basandosi sui risultati di Dugatkin sull'influenza sociale nella scelta del compagno da parte delle femmine di guppy, hanno effettuato osservazioni su altre specie di pesci e hanno riscontrato che i maschi di una specie della famiglia dei pecilidi (Poecilia latipinna), che si accoppiano frequentemente con le femmine di un'altra specie (Poecilia formosa), di fatto aumentano il proprio successo riproduttivo con le femmine della propria specie per mezzo di questi accoppiamenti interspecifici. Le femmine di Poecilia latipinna tendono infatti a preferire i maschi che hanno visto accoppiarsi, anche se questi si accoppiano con femmine di Poecilia formosa.
Imparare a corteggiare: l'apprendimento del canto nel molotro nero
Il molotro nero (Molothrus ater) è un uccello particolarmente interessante per chi studia le influenze sociali sullo sviluppo del comportamento. Le femmine di questa specie depongono le uova in nidi costruiti da membri di altre specie; infatti il molotro nero è un parassita di covata. Per questo motivo, i giovani molotri sono sempre allevati da genitori adottivi, membri di altre specie. I piccoli non entrano in contatto con gli adulti della propria specie durante lo sviluppo. A causa dell'isolamento dagli adulti dei giovani molotri, per molto tempo si è pensato che l'apprendimento del canto nei maschi di questa specie non fosse influenzato dall'interazione sociale. Tuttavia, grazie alle intuizioni e agli sforzi di M.J. West e A.P. King (1996), adesso è noto che i giovani maschi di molotro nero, come i piccoli di molte altre specie di uccelli canori, imparano i dettagli del proprio canto per mezzo delle interazioni sociali.
I maschi di molotro appartenenti a differenti aree geografiche esibiscono diverse varianti del canto e questi 'dialetti' sono conservati grazie alla differenza nelle risposte prodotte da femmine provenienti da diverse aree. Gli stessi canti che durante la stagione degli accoppiamenti inducono con maggiore probabilità la femmina ad assumere la postura copulatoria che permette l'accoppiamento, in altri periodi dell'anno suscitano nelle femmine la risposta del 'colpo d'ala'. Dopo che la femmina ha dato un colpo d'ala in risposta a un canto, il maschio che lo ha emesso lo ripete tre o quattro volte consecutivamente, violando la regola elementare nel canto dei molotri maschi che normalmente emettono uno dopo l'altro e senza ripetizioni i vari canti del proprio repertorio. La vista di una femmina che dà un colpo d'ala fa aumentare non solo la frequenza con cui un maschio emette un canto fuori dalla stagione degli accoppiamenti, ma anche la frequenza con cui il canto viene emesso durante la stagione degli accoppiamenti. Questo fa sì che il maschio aumenti il proprio potenziale successo riproduttivo (West e King, 1996).
Ciò che sorprende di più è il fatto che il canto del molotro maschio è anche influenzato dalle interazioni con altri maschi. Le varianti del canto che sono più efficaci nel provocare posture di copulazione nella femmina sono anche molto efficaci nel provocare attacchi da parte di altri maschi di molotro. Di conseguenza, soltanto i maschi dominanti in un gruppo possono continuare a emettere quei canti che sono più efficaci nel suscitare posture di copulazione nelle femmine senza rimanere gravemente feriti negli attacchi da parte di altri maschi.
Considerazioni generali sull'apprendimento sociale non imitativo
Dagli esempi di apprendimento sociale non imitativo negli animali presentati si possono trarre due lezioni di fondo. La prima è che l'apprendimento sociale può giocare un ruolo nello sviluppo e nel mantenimento di molte sequenze di comportamento che contribuiscono al successo riproduttivo. La seconda è che alla base dell'apprendimento sociale vi sono vari processi comportamentali essenzialmente diversi tra loro. Le preferenze di cibo nel ratto sono influenzate dal fatto che il ratto è esposto all'odore del cibo combinato con le sostanze chimiche presenti nell'alito di un altro ratto. I ratti neri apprendono tecniche complesse per ottenere il cibo se hanno l'opportunità di estrarre pinoli da pigne già parzialmente sfruttate da adulti esperti. Le femmine di guppy si accoppiano con i maschi che poco prima hanno corteggiato altre femmine e i maschi di molotro imparano quali sono i canti più efficaci in un particolare contesto sulla base delle lusinghe che ricevono dalle femmine e delle punizioni che ricevono dagli altri maschi.
Sebbene i processi comportamentali alla base dei vari esempi di apprendimento sociale discussi in precedenza differiscano considerevolmente tra loro, nessuno di essi è prodotto da una forma di apprendimento dove si impara un'azione particolare vedendola effettuare da un altro; ovvero, nessuno di quei processi è un esempio di apprendimento per imitazione. L'apprendimento per imitazione sarà perciò l'argomento della seconda parte di questo saggio.
L'imitazione
La letteratura descrittiva è ricca di osservazioni informali su animali, dagli scimpanzé (che secondo alcune descrizioni sarebbero capaci di imparare ad aprire barattoli di vernice, a temperare matite e a utilizzare la carta vetrata) ai delfini (un resoconto riporta che avrebbero copiato un sommozzatore che puliva i vetri del loro acquario), che imparano complesse sequenze motorie per mezzo dell'imitazione (Moore, 1992). Anche se le descrizioni di questi comportamenti, simili a quelli dell'uomo ma eseguiti da animali, sono molto probabilmente accurate, esistono due problemi di base nell'accettarle come dati importanti per capire le capacità di specie diverse dalla nostra.
In primo luogo, in mancanza di osservazioni sistematiche è molto difficile sapere con quale frequenza gli animali eseguano questi comportamenti apparentemente imitativi. Di conseguenza, non si può determinare se si tratti soltanto di corrispondenze casuali tra il comportamento dell'animale e di chi si prende cura di esso, osservate in un'unica occasione tra milioni di volte in cui le persone osservano gli animali ogni giorno. Decine di milioni di persone hanno animali domestici e possono osservare il loro comportamento per diverse ore al giorno, eppure è soltanto in quelle rare occasioni in cui un animale sembra avere spontaneamente copiato un comportamento mostrato da chi si prende cura di lui, che l'osservazione è riportata. Nessuno riferisce degli svariati milioni di volte in cui gli animali da compagnia vedono il proprio padrone eseguire un comportamento che sarebbe utile imparare, ma non riescono a copiarlo (Visalberghi e Fragaszy, 1990).
In secondo luogo, è difficile determinare sulla base di osservazioni effettuate in ambienti non controllati, come questi comportamenti simili a quelli umani si siano sviluppati negli animali che li esibiscono. Il presunto imitatore ha di fatto osservato un comportamento e poi successivamente lo ha riprodotto? O piuttosto il comportamento del padrone ha fatto sì che la sua attenzione si focalizzasse su qualche oggetto, per esempio una porta, che poi ha provato ad aprire decine o centinaia di volte prima di riuscirvi?
Per evitare i problemi connessi con l'utilizzazione di queste osservazioni casuali di comportamenti potenzialmente interessanti, nello studio dell'imitazione si è cercato di trovare situazioni in cui i soggetti che imitano sono animali che vivono in laboratorio, dove le osservazioni sono effettuate in condizioni controllate. Per moltissimi anni gli studi sull'imitazione sono stati quasi sempre fallimentari nel tentativo di far apprendere agli animali per imitazione quando altre forme di apprendimento sociale erano impedite. Non sorprende quindi che, dopo tanti insuccessi, molti ricercatori abbiano concluso che, nonostante i resoconti informali sostengano il contrario, gli animali probabilmente non sono in grado di apprendere attraverso l'imitazione.
Tuttavia, alcuni scienziati hanno continuato a preoccuparsi della discordanza tra i risultati formali e i resoconti informali riguardo a questo problema. È quest'ultimo gruppo di studiosi a essere oggi in fase ascendente. Infatti, nel corso degli anni Novanta è stata progressivamente raccolta una grande quantità di prove sperimentali che, nel loro insieme, danno un supporto considerevole all'idea che un' ampia gamma di animali sia in grado di apprendere per imitazione.
Trasformare gli aneddoti in osservazioni: studi sull'orango in natura
A.E. Russon e B.M.F. Galdikas (1993) lavoravano in un campo nel Borneo dove un gruppo di conservazionisti si occupava di preparare alla re introduzione in natura oranghi catturati dai cacciatori di frodo e allevati in cattività. Questi ricercatori hanno effettuato osservazioni sistematiche, e a volte videoregistrazioni, di ogni comportamento apparentemente imitativo esibito dalle scimmie mentre queste andavano in giro e interagivano con le persone residenti nel campo. Russon e Galdikas hanno descritto dozzine di comportamenti umani complessi che le scimmie sembravano copiare. Per esempio, il personale del campo travasava regolarmente il carburante dai bidoni alle lattine. L'operazione consisteva nel sollevare il coperchio dei bidoni e delle lattine, nell'inserire un'estremità di un tubo di gomma nel bidone e nel succhiare dall' altra estremità per iniziare a far scorrere il carburante. Infine, l'estremità del tubo veniva rapidamente inserita in una lattina. Una femmina adulta di orango è stata vista in un capannone dove si conservavano i bidoni di carburante mentre svitava il coperchio sia di un bidone sia di una lattina per inserire un'estremità del tubo nel bidone di carburante. La scimmia ha poi inserito l'altra estremità del tubo in bocca e ha formato delle fosse nelle guance come avviene quando si succhia qualcosa. Dopo molte manipolazioni del tubo e del bidone, l'orango ha inserito l'estremità del tubo che aveva preso in bocca nella lattina aperta. Nonostante il fatto che il bidone selezionato dall'orango fosse in effetti vuoto, e che l'intervallo di tempo tra succhiare a un'estremità del tubo e inserirlo nella lattina fosse inappropriato, la sequenza comportamentale era simile a quella di un essere umano che travasa del carburante.
Anche se non è possibile sapere, in circostanze non controllate, come la scimmia possa essere giunta a strutturare una tale sequenza di comportamenti, i tanti esempi descritti da Russon e Galdikas sembrano suggerire che gli oranghi in circostanze appropriate siano capaci di imitare. In effetti le osservazioni di Russon e Galdikas offrono indicazioni riguardo alle situazioni che fanno aumentare la probabilità che un orango imiti spontaneamente un essere umano. Le analisi delle osservazioni effettuate nella giungla del Borneo hanno mostrato che, quando esiste una stretta relazione sociale tra un modello umano e una scimmia, aumenta la probabilità che questa lo imiti. Considerando questo risultato, non sorprende il fatto che alcuni tra i più promettenti e attuali studi di laboratorio sulla capacità delle scimmie antropomorfe di imitare gli esseri umani abbiano utilizzato come soggetti scimmie antropomorfe allevate in stretto contatto con esseri umani.
L'imitazione negli scimpanzé addestrati al linguaggio e allevati in interazione con esseri umani
M. Tomasello e colleghi hanno studiato l'apprendimento imitativo di scimpanzé 'acculturati', allevati da umani e addestrati a utilizzare un linguaggio simile al nostro, confrontandolo con quello di bambini di due anni e di altri scimpanzé allevati dalle proprie madri. Tutti i soggetti osservavano un essere umano che svolgeva la funzione di modello e dimostrava una serie di azioni nuove dirette verso un oggetto. Per esempio, i soggetti vedevano un modello umano sistemare una palla sulla propria testa e utilizzare una leva per sollevare il coperchio di un barattolo di vernice. Ai bambini era detto "fai questo" e le scimmie venivano preaddestrate a riprodurre azioni a loro familiari eseguite da un dimostratore umano (Tomasello, 1996). I risultati dell'esperimento di Tomasello sono chiari. Le scimmie allevate dalle proprie madri naturali erano quasi completamente incapaci di imitare le azioni nuove dimostrate dallo sperimentatore. I bambini di due anni e le scimmie allevate dagli umani e addestrate al linguaggio imitavano le azioni dello sperimentatore con la stessa, notevole, frequenza. L'esperimento di Tomasello è solo uno dei tanti che permettono di giungere alle medesime conclusioni. Risultati compatibili con l'idea che gli scimpanzé allevati dagli uomini siano capaci di imitare azioni nuove dimostrate da un modello umano sono stati ottenuti recentemente in varie situazioni di laboratorio (Whiten e Custance, 1996).
Imitazione in animali diversi dalle scimmie antropomorfe
I risultati di altri esperimenti recenti indicano che non soltanto le scimmie antropomorfe, ma anche vari animali a cui non sembrerebbe plausibile attribuire la capacità di imitare sembrano invece in grado di riprodurre semplici atti motori dopo averli osservati in altri individui.
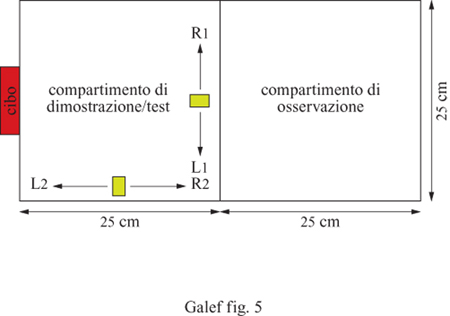
In una serie di studi condotti da C.M. Heyes e collaboratori, alcuni ratti guardavano attraverso una finestra di uno schermo un altro ratto, precedentemente addestrato a muovere verso destra o verso sinistra una leva sospesa al soffitto della gabbia. Il ratto dimostratore ha premuto per cinquanta volte di seguito la leva verso la corretta direzione, ricevendo cibo come ricompensa per ogni spostamento della leva. A questo punto, il ratto dimostratore era rimosso dal dispositivo e un ratto osservatore era sistemato al suo posto nel compartimento della gabbia contenente la leva. Il topo osservatore poteva premere la leva in qualsiasi direzione per cinquanta volte, ottenendo una ricompensa a ogni spostamento. Per alcuni osservatori, la leva rimaneva nella stessa posizione durante la fase di addestramento e in quella di test; per altri, la leva era spostata prima del test da una posizione vicina allo schermo a una posizione vicina alla parete frontale del dispositivo (fig. 5). In entrambe le condizioni, ciascuno dei ratti osservatori ha mostrato la tendenza a spingere la leva nella direzione in cui la muoveva il dimostratore precedentemente osservato (Heyes, 1996). T.R. Zentall e i suoi studenti hanno riportato risultati simili nei piccioni e nelle quaglie (Akins e Zentall, 1996; Kaiser et al., 1997).
Infine, B.R. Moore (1992) ha fornito prove piuttosto sorprendenti di imitazione di esseri umani da parte di un pappagallo cinerino (Psittacus erithacus) chiamato Okichoro. Moore aveva sistemato Okichoro in una voliera che conteneva, oltre ai soliti trespoli e giochi, un microfono e una video camera che permettevano di osservarne il comportamento mentre era solo e indisturbato. Per un periodo di cinque anni, Moore visitava la voliera diverse volte al giorno. Durante ciascuna delle sue visite, effettuava ripetutamente movimenti ben distinti, ciascuno accompagnato da una parola o una frase diversa. Per esempio, ogni giorno Moore, allontanandosi dalla voliera di Okichoro, lo salutava agitando la mano e diceva "ciao". Okichoro ha imparato presto a dire "ciao" e, alla fine del primo anno, è stato visto pronunciare la parola "ciao" e allo stesso tempo agitare la propria zampa mentre era da solo nella voliera. Quando si trovava nella voliera di Okichoro, Moore diceva anche "guarda la mia lingua", poi apriva la bocca e tirava fuori la lingua. Dopo un certo tempo che Okichoro aveva imparato a dire "ciao" e ad agitare la zampa, è stato visto sullo schermo televisivo dire anche "guarda la mia lingua", poi aprire la bocca e tirar fuori la lingua. Okichoro raramente ha aperto la bocca e tirato fuori la lingua dopo avere detto "ciao" o ha agitato la zampa dopo avere detto "guarda la mia lingua". Nel corso di cinque anni di studio, Okichoro è stato osservato mentre copiava molte azioni diverse che coinvolgevano sei diverse parti del corpo. A ciascuna delle azioni Okichoro associava una frase appropriata.
Alcuni scienziati considerano questi risultati sull' apprendimento sociale in soggetti non appartenenti all'ordine dei primati come esempi di imitazione vera e propria. Altri, al contrario, si rifiutano di considerarli vera imitazione perché è risultato difficile accertare se le sequenze motorie utilizzate dai soggetti per riprodurre il comportamento di un modello fossero effettivamente nuove. È infatti possibile che l'azione copiata dall'animale facesse già parte del proprio repertorio comportamentale. In questo caso osservare il comportamento di un altro agente gli ha soltanto permesso di imparare in quali circostanze eseguire tale comportamento. Non è chiaro se la produzione di un particolare comportamento, che fa già parte del repertorio di un animale, in seguito all'osservazione di un modello, vada considerata come un esempio di vera imitazione. Vi è ancora da svolgere un arduo lavoro teorico prima di giungere in modo unanime a un'esatta definizione di imitazione e di stabilire cosa si può inferire dalle capacità cognitive di un animale in base al fatto che mostra di sapere imitare comportamenti nuovi o comportamenti che già gli sono familiari.
Conclusioni
Gli studiosi dell'apprendimento sociale imitativo e non imitativo negli animali si trovano in una fase emozionante della propria ricerca. Dopo un secolo di tentativi, forse oggi ci troviamo frnalmente in una posizione che ci consente di iniziare a esplorare i processi cognitivi basilari dell'apprendimento imitativo negli animali e di compararli con quelli dell'apprendimento imitativo nell'uomo. Dopotutto, anche se un essere umano e una scimmia antropomorfa sono in grado di imitare, potrebbero comunque non farlo nello stesso modo. Siamo ormai anche in possesso di prove valide sull'importanza dell' apprendimento sociale nello sviluppo di sequenze comportamentali che facilitano la sopravvivenza e la riproduzione di animali che vivono in habitat naturali. Questi risultati costituiscono un contributo importante alla comprensione delle fonti di informazione usate dagli animali per rispondere in modo adattivo alle sfide poste dal proprio ambiente. Due decenni di duro lavoro da parte di ricercatori impegnati in diverse discipline iniziano a dare i propri frutti e nel prossimo decennio la ricerca sull'apprendimento sociale e sull'imitazione promette di essere altrettanto produttiva di quanto lo è stata nel decennio scorso.
Bibliografia citata
AKINS, C.K., ZENTALL, T.R. (1996) Imitative learning in male Japanese quail (Coturnix japonica) using the two-action method. J. Comp. Psychol., 110, 3116-3120.
DUGATKIN, L.A. (1996) Copying and mate choice. In Social learning in animals: the roots of culture, a c. di Heyes C.M., Galef B.G. Jr., New York, Academic Press, pp. 85-106.
GALEF, B.G. Jr. (1977) Mechanisms for the social transmission of acquired food preferences from adult to weanling rats. In Learning mechanisms in food selection, a c. di Barker L.M., Best M.R., Domjam M., Waco, Baylor University Press, pp. 123-150.
GALEF, B.G. Jr. (1988a) Communication of information conceming distant diets in a social, centraI-pIace foraging species: Rattus norvegicus. In Social learning: psychological and biological perspectives, a c. di Zentall T.R., Galef B.G. Jr., Hillsdale, Lawrence Erlbaum Associates, pp. 119-140.
GALEF, B.G. Jr. (1988b) Imitation in animals: history, definition and interpretation of data from the psychological laboratory. In Social learning: psychological and biological perspectives, a c. di Zentall T.R., Galef B.G. Jr., Hillsdale, Lawrence Erlbaum Associates. pp. 3-28.
GALEF, B.G. Jr. (1996) Social enhancement of food preferences in Norway rats: a brief review. In Social learning in animals: the roots of culture, a c. di Heyes C.M., Galef B.G. Jr., San Diego, Academic Press, pp. 49-64.
GIBSON, R.M., BRADBURY, J.W., VEHRENCAMP, S.L. (1991) Mate choice in lekking sagegrouse: the role of vocal display, female site fidelity and copying. Behav. Ecol., 2, 165-180.
HEPPER, P.G. (1988) Adaptive fetal learning: prenatal exposure to garlic affects postnatal preference. Anim. Behav., 36, 935-936.
HEYES, C.M. (1996) Genuine imitation? In Social learning in animaIs: the roots of culture, a c. di Heyes C.M., Galef B.G. Jr., San Diego, Academic Press, pp. 371-390.
KAISER, D.H., ZENTALL, T.R., GALEF, B.G. Jr. (1997) Can imitation in pigeons be explained by local enhancement together with trial and error leaming? Psychol. Sci., 8, 459-460.
MINEKA, S., COOK, M. (1988) Social leaming and the acquisition of snake fear in monkeys. In Social learning: psychological and biological perspectives, a c. di Zentall T.R., Galef B.G. Jr., Hillsdale, Lawrence Erlbaum Associates, pp. 3-28.
MOORE, B.R. (1992) Avian movement imitation and a new form of mimicry: tracing the evolution of a complex form of leaming. Behaviour, 122, 231-263.
RUSSON, A.E., GALDIKAS, B.M.F. (1993) Imitation in free-ranging rehabilitant orangutans (Pongo pygmaeus). Comp. Psychol., 107, 147-161.
SCHLUPP, I., MARLER, C., RYAN, M.J. (1994) Benefit to male sailfin mollies of mating with heterospecific females. Science, 263, 373-374.
TERKEL, J. (1995) Cultural transmission in the black rat: cone feeding. In Advances in the study of behavior, voI. 24, a c. di Slater P.J.B., Rosenblatt J.S., Snowdon C.T., Milinski M., San Diego, Academic Press, pp. 119-154.
THORPE, W.H. (1963) Learning and instinct in animals, 2a ed., Londra, Methuen.
TOMASELLO, M. (1996) Do apes ape? In Social learning in animaIs: the roots of culture, a c. di Heyes C.M., Galef B.G. Jr., San Diego, Academic Press, pp. 319-344.
VISALBERGHI, E., FRAGASZY, D.M. (1990) Do monkeys ape? In 'Language' and intelligence in monkeys and apes: comparative developmental perspectives, a c. di Parker S.T., Gibson K.R., Cambridge, Mass., Cambridge University Press, pp. 247-273.
WEST, M.J., KING, A.P. (1996) Social leaming: synergy and song birds. In Social learning in animals: the roots of culture, a c. di Heyes C.M., Galef B.G. Jr., San Diego, Academic Press, pp. 155-178.
WHITEN, A., CUSTANCE, D. (1996) Studies of imitation in chimpanzees and children. In Social learning in animals: the roots of culture, a c. di Heyes C.M., Galef B.G. Jr., San Diego, Academic Press, pp. 291-318.
WHITEN, A., HAM, R. (1992) On the nature and evolution of imitation in the animaI kingdom: reappraisal of a century of research. In Advances in the study of behavior, a c. di Slater P.J.B., Rosenblatt J.S., Beer C., New York, Academic Press, pp. 239-283.
Bibliografia generale
GALEF, B.G. Jr. Social influences on food preferences and feeding behaviours of vertebrates. In Why we eat what we eat, a c. di Capaldi B., Washington, American Psychological Association, in corso di stampa.
HEYES, C.M., GALEF, B.G. Jr. Social learning in animals: the roots of culture. San Diego, Academic Press, 1996.
ZENTALL, T.R., GALEF, B.G. Jr. Social learning: psychological and biological perspectives. Hillsdale, Lawrence Erlbaum Associates, 1988.