apprendimento
apprendimento
L’apprendimento è un processo che si manifesta sotto forma di cambiamenti adattativi durevoli del comportamento indotti dall’esperienza individuale. Le capacità di apprendere, ossia di stabilire relazioni causali tra eventi e di modificare il proprio comportamento in base a tali esperienze, e di memorizzare tali relazioni (ovvero immagazzinarle e successivamente richiamarle) sono rese possibili dalla particolare organizzazione funzionale del sistema nervoso. Gli studi condotti sugli animali hanno dimostrato come il cervello possegga uno o più meccanismi neurali attraverso i quali gli stimoli e le azioni possono essere associati tra loro. I meccanismi di base, biochimici e fisiologici, che regolano la trasmissione dell’impulso nervoso, si sono conservati nel corso dell’evoluzione e sono sostanzialmente gli stessi negli invertebrati e nell’uomo. I risultati ottenuti in laboratorio e sul campo in diverse specie animali hanno quindi rappresentato una solida base teorica per la comprensione dei processi cognitivi complessi (e dei relativi substrati biologici) caratteristici della specie umana. [➔ cognizione spaziale; comportamento sociale; memoria; neurone, Neuroni specchio; recettore nervoso] Nell’a. di tipo non associativo, che viene solitamente considerato la forma più semplice di a. e che risulta ampiamente diffuso in tutto il regno animale, è presente un singolo stimolo. Si può distinguere tra abituazione e sensibilizzazione. Nel caso dell’abituazione (➔), si osserva una diminuzione della risposta emessa quando gli stimoli vengono presentati ripetutamente. Tale diminuita reattività (che non è riconducibile ad adattamento sensoriale o a fenomeni di fatica muscolare) non comporta, a differenza delle altre forme di a., un’acquisizione di nuove risposte, quanto piuttosto perdita di quelle acquisite perché lo stimolo viene considerato non rilevante. L’abituazione gioca quindi un ruolo fondamentale, durante lo sviluppo, in individui giovani che devono selezionare gli stimoli a cui reagire, tralasciando quelli irrilevanti. Nel caso invece della sensibilizzazione, la risposta a uno stimolo risulta aumentata rispetto ai valori attesi dopo presentazione a un singolo stimolo di intensità molto elevata. I dettagli molecolari di entrambe queste forme di a. sono stati studiati nel mollusco Aplysia californica, una lumaca di mare.
Apprendimento associativo
L’a. associativo riguarda le relazioni tra due eventi (due stimoli, uno stimolo e una risposta, una risposta emessa e un evento successivo). Nel condizionamento classico si apprende che uno stimolo fa prevedere un evento successivo, nel condizionamento operante vengono apprese le conseguenze del proprio comportamento. Il condizionamento classico. Nel condizionamento classico (o pavloviano), uno stimolo inizialmente a carattere neutro (stimolo condizionato), per es. il suono di un campanello, viene ripetutamente associata alla presenza della carne liofilizzata (stimolo incondizionato) e si osserva che questo finisce per evocare la risposta prima incondizionata e ora condizionata di salivazione nel cane. Il fisiologo russo Ivan Petrovic Pavlov, dopo aver osservato sporadicamente questo fenomeno durante i suoi studi sulla digestione nei cani (la risposta di salivazione dei cani risultava associata al camice bianco dell’operatore che portava il cibo), condusse un’analisi sistematica del fenomeno, definendolo riflesso condizionato (associazione tra stimolo condizionato e risposta condizionata) e descrivendone successivamente varie caratteristiche: l’andamento nel tempo, con una diminuzione della latenza alla risposta condizionata, l’estinzione dopo ripetute presentazioni dello stimolo condizionato in assenza dello stimolo incondizionato. Gli studi di Pavlov costituirono un passaggio fondamentale nello studio dell’a., che, da allora, divenne un’analisi obiettiva di stimoli e di risposte misurabili in laboratorio. Un caso particolare di condizionamento classico pavloviano è l’a. di un’avversione condizionata alimentare. Si tratta di una forma di a. associativo, in cui il malessere gastrointestinale provocato da un alimento (gusto specifico del cibo ingerito) induce un’avversione per quell’alimento, che verrà quindi successivamente evitato. L’intensità della risposta di evitamento dipende dall’intensità del malessere, dal grado di familiarità del soggetto per quell’alimento, dall’intensità della stimolazione gustativa associata. L’avversione condizionata di tipo alimentare, consente di riassumere alcuni aspetti importanti del condizionamento classico: il valore adattativo di tale a., soprattutto per le specie con grande capacita di adattamento, il cui successo dipende da un equilibrio tra esplorazione alimentare ed evitamento di potenziali tossine presenti nell’ambiente naturale; le significative limitazione biologiche nella natura sensoriale degli stimoli presenti nel paradigma: nella maggior parte delle specie (compresa quella umana) l’avversione condizionata si stabilisce solo verso stimoli di natura gustativa e a fronte di una successiva insorgenza di uno stato di malessere interno; a differenza di altre forme di a., il condizionamento classico richiede un numero ristretto di esperienze (anche una singola) e si instaura nonostante lo stato di malessere si presenti a distanza di ore dall’ingestione del cibo.
Il condizionamento operante. Nella seconda forma di a. associativo, il condizionamento operante (detto anche strumentale o a. per prove ed errori) un’attività volontaria (per es., un movimento) viene associata con un rinforzo. Negli stessi anni degli studi sui cani di Pavlov in Russia, lo psicologo sperimentale statunitense Edward L. Thorndike condusse i primi esperimenti per valutare in animali da laboratorio il progressivo stabilirsi di un’associazione tra una risposta motoria emessa e un evento. Successivamente Burrhus F. Skinner mise a punto una gabbia sperimentale per il ratto contenente un meccanismo (operandum, da cui il termine operante) che, quando azionata, forniva un rinforzo positivo (in genere del cibo). Dopo la prima pressione casuale della leva, l’animale impara (per tentativi ed errori) ad associare la sua attività (la pressione sulla leva) con il rinforzo positivo; la presenza del rinforzo aumenta la probabilità che il comportamento che lo ha preceduto sia ripetuto. In possesso di queste informazioni, tutte le volte che il ratto e affamato e si ritrova nella stessa gabbia e probabile che fornisca la risposta appropriata. A differenza del condizionamento classico, che risulta limitato ad alcune risposte riflesse specifiche che vengono evocate da stimoli particolari, il condizionamento operante coinvolge comportamenti spontanei la cui frequenza aumenta se questi inducono variazioni favorevoli.

Utilizzazione terapeutica del condizionamento. Alcune forme di condizionamento associativo sono state utilizzate nell’uomo a scopo terapeutico. Tecniche di desensibilizzazione sistematica per contrastare disturbi d’ansia sono basate sulla associazione tra stimolo ansiogeno e induzione di una risposta antagonista all’ansia (per es., uno stato di rilassamento muscolare) per inibire reazioni emotive spiacevoli. Anche il biofeedback (utilizzato in casi di dolore cronico, pressione sanguigna elevata) è una forma di condizionamento operante utilizzato in clinica per monitorare alcune funzioni corporee e meglio controllarle.
Apprendimento sociale
In molte specie animali, dagli insetti ai primati, l’a. individuale può essere influenzato dal contesto sociale. La presenza di altri individui conspecifici e di interazioni sociali può sia aumentare le probabilità che si manifestino comportamenti già presenti nel proprio repertorio, sia promuovere l’acquisizione di comportamenti del tutto nuovi. Attraverso il ruolo di osservatore di un comportamento altrui, si possono acquisire informazioni che riguardano l’ambiente o gli effetti sull’ambiente dei comportamenti osservati (a. sociale non imitativo), oppure informazioni che concernono direttamente il comportamento osservato (a. sociale imitativo). L’a. sociale non imitativo è un fenomeno piuttosto comune nel mondo animale, attraverso il quale si stabiliscono preferenze alimentari in specie onnivore, si identificano luoghi specifici di foraggiamento, si acquisiscono particolari modalità di manipolazione per accedere a risorse alimentari altrimenti non disponibili. L’a. sociale imitativo, quello in cui si impara a eseguire un’azione osservandone la sua esecuzione da parte di altri, è invece meno diffuso nel mondo animale e limitato ad alcune specie, come l’uomo, le scimmie antropomorfe, alcuni uccelli canori. Neuroni specializzati a rispondere a particolari stimoli, che sono coinvolti in fenomeni plastici, sono stati evidenziati nella corteccia visuomotoria di primati non umani. Queste neuroni (➔), detti neuroni a specchio (mirror neurons), aumentano la loro attività elettrica quando, per es., la scimmia osserva un’altra scimmia compiere un’azione, come maneggiare uno strumento. L’attivazione dei neuroni a specchio potrebbe corrispondere a fenomeni plastici associati all’a. sociale.
Il caso dell’imprinting
L’imprinting (➔), un processo attraverso il quale le preferenze sociali dell’individuo immaturo si restringono a una particolare classe di stimoli, può risultare determinante per lo stabilirsi di un legame con il genitore (imprinting filiale) o nella formazione delle preferenze sessuali dell’individuo adulto (imprinting sessuale). Inizialmente osservato dall’ornitologo Oskar Heinroth e dall’etologo Konrad Lorenz nella prima metà del Novecento, è stato successivamente studiato soprattutto in alcune specie di uccelli, dove si verifica subito dopo la schiusa, ma è stato descritto anche in alcuni mammiferi, dove risulta comunque meno diffuso, poiché nelle prime fasi dello sviluppo postnatale dei mammiferi sono soprattutto gli stimoli olfattivi materni (ai quali la prole risulta passivamente esposta durante l’allattamento) a determinare le successive preferenze sociali. L’imprinting è considerato una forma particolare di a. con tre caratteristiche distintive: è limitato a una breve fase sensibile, il periodo perinatale, la cui durata varia da specie a specie; si verifica in totale assenza di stimoli di rinforzo convenzionali (è sufficiente l’esposizione del soggetto a un determinato stimolo in movimento durante la fase sensibile); produce modificazioni comportamentali a breve e a lungo termine, non irreversibili, ma difficilmente estinguibili. Dati sperimentali indicano come alcune sottoregioni dell’iperstriato ventrale (una regione presente negli emisferi telencefalici degli uccelli) costituiscano la sede principale dove vengono immagazzinate le informazioni rilevanti per il riconoscimento dello stimolo. A fenomeni di imprinting sono stati inoltre associate variazioni strutturali a livello sia pre- che postsinaptico in alcuni circuiti dell’iperstriato ventrale sinistro (aumento dei recettori NMDA, della dimensione della densità postsinaptica, della disponibilità di vescicole sinaptiche).
Apprendimento spaziale
Nell’a. latente, così definito dallo psicologo sperimentale Edward C. Tolman, che si occupò del fenomeno negli anni Trenta del 20° secolo, si costituisce un’associazione tra stimoli in assenza di un rinforzo apparente. Analizzando le modalità di esplorazione di ambienti sperimentali complessi (labirinti) in ratti di laboratorio, Tolman dedusse che l’acquisizione delle informazioni spaziali avveniva attraverso la costruzione di una mappa spaziale e non da una successione a catena di associazioni stimolo-rinforzo. L’ipotesi di Tolman venne confermata quando nel 1971 furono per la prima volta identificate nel ratto le cosiddette cellule di posizione (place-cells): singoli neuroni ippocampali che si attivano quando l’animale occupa un punto preciso dell’ambiente in cui si trova. L’attività coordinata di diversi neuroni genera una rappresentazione interna dello spazio occupato dall’animale (place field). Sulla base di queste osservazioni John O’Keefe e Lynn Nadel ipotizzarono che la funzione primaria della regione ippocampale sia la formazione di una mappa cognitiva dell’ambiente esterno. Studi successivi hanno poi confermato la presenza di cellule di posizione anche nell’uomo. Nel ratto la rappresentazione è plastica: quando un animale entra in un nuovo ambiente si formano nuovi place fields che poi rimangono stabili per settimane o mesi. Lo sviluppo delle tecniche elettrofisiologiche ha consentito di misurare variazioni dei place fields nell’animale in seguito a modificazioni della configurazione spaziale (dalla forma quadrata a quella circolare di uno spazio chiuso). La regione ippocampale non è la sola deputata alla codificazione delle informazioni spaziali: nella corteccia entorinale, che rappresenta la maggiore afferenza all’ippocampo, sono state infatti identificate nel 2005 le cellule a griglia (grid cells) che si attivano anch’esse in corrispondenza di una particolare posizione nell’ambiente ma, a differenza delle cellule di posizione, possiedono ognuna molteplici campi di attivazione che codificano l’intero ambiente esterno, secondo una spaziatura modulare esagonale. Negli stessi anni in cui sono state identificate le cellule di posizionamento, il neurofisiologo britannico Tim Bliss e il norvegese Terje Lømo, allora studente di psicologia, descrissero il fenomeno del potenziamento a lungo termine (➔) (LTP) nell’ippocampo del coniglio, che consiste nell’aumento stabile e duraturo dell’intensità della risposta neuronale in seguito a stimolazione elettrica ad alta frequenza delle cellule afferenti; il neurone postsinaptico permane in una condizione attivata per intervalli di tempo molto più lunghi di quanto avvenga nella trasmissione dell’impulso nervoso. L’LTP presenta caratteristiche che lo rendono adatto a rappresentare una base fisiologica dei processi di a. associativo. Si tratta infatti di un fenomeno persistente, specifico per i neuroni attivati da stimolazione presinaptica ad alta frequenza, e associativo, in quanto inducibile nel neurone postsinaptico anche per sommazione di diverse attività presinaptiche sotto soglia. Dalla metà degli anni Ottanta del 20° secolo, molti gruppi di ricerca hanno dimostrato che l’induzione dell’LTP richiede l’attività concomitante sia nel neurone presinaptico che postsinaptico; solitamente l’LTP è indotto dall’attivazione di un particolare tipo di recettore (NMDA) del neurotrasmettitore glutammato, mentre i recettori di tipo AMPA per lo stesso neurotrasmettitore modulano l’ingresso dello ione Ca2+ e quindi l’intensità e la durata della risposta elettrica del neurone alla stimolazione. Il coinvolgimento dell’LTP nell’a. associativo è stato dimostrato da Richard Morris e collaboratori che hanno osservato come il blocco farmacologico della funzionalità del recettore NMDA (in seguito all’infusione cronica di un antagonista del recettore stesso nei ventricoli cerebrali di ratti) prevenga il fenomeno dell’LTP e interferisca selettivamente con l’a. spaziale nel labirinto acquatico. Studi più recenti in topi geneticamente manipolati, con mutazioni o ablazioni nei geni codificanti le proteine coinvolte nella plasticità sinaptica, hanno confermato l’associazione tra deficit nell’a. spaziale e alterata risposta elettrica dei neuroni postsinaptici ippocampali.
La basi biologiche dell’apprendimento
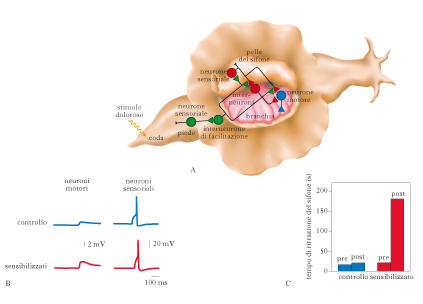
Gli studi sul sistema nervoso degli invertebrati, in partic. gli esperimenti condotti dal Eric Kandel e dai suoi collaboratori sulla lumaca di mare Aplysia californica, hanno fornito un contributo fondamentale alla conoscenza delle basi neurali dell’a. e della memoria. Questo invertebrato possiede un sistema nervoso relativamente semplice (formato da soli 20.000 grandi neuroni, raggruppati in 10 gangli principali), che risulta quindi facile da analizzare con tecniche di elettrofisiologia e biochimica. Il comportamento di difesa della lumaca di mare di fronte a nuove stimolazioni è di ritrarre energicamente la branchia (l’organo della respirazione) e il sifone (un piccolo canale carnoso posto sopra la branchia per l’espulsione dell’acqua). Quando l’animale viene sfiorato, i neuroni sensoriali che ricevono la stimolazione tattile inviano l’informazione ai motoneuroni e la branchia e il sifone si ritraggono. L’analisi della risposta difensiva di Aplysia ha rivelato che nel caso di abituazione (ripetute presentazioni dello stimolo tattile) l’informazione trasmessa dai neuroni sensoriali a quelli motori diminuisce progressivamente di intensità fino a raggiungere un livello non più in grado di evocare la risposta dei motoneuroni. Si osserva una diminuzione persistente dell’efficacia delle connessioni sinaptiche fra i neuroni sensoriali e i motoneuroni dovuta al rilascio di sempre minori quantità di neurotrasmettitore. Questo fenomeno di modificazione plastica si accompagna alla diminuzione dei potenziali elettrici postsinaptici dei motoneuroni e dipende, almeno in parte, dal diminuito accesso di ioni calcio (Ca2+) nella terminazione sinaptica del neurone sensoriale. Nella risposta difensiva di Aplysia possono anche osservarsi fenomeni di sensibilizzazione: se si stimola in modo potenzialmente nocivo la cute di Aplysia, l’animale ritrarrà la branchia, sensibilizzandosi al punto che poi basterà sfiorare appena la cute del sifone o la testa, per provocare una reazione di analoga intensità. Anche la sensibilizzazione comporta una modificazione dell’efficacia delle sinapsi che collegano il neurone sensoriale al neurone motore. La modificazione consiste nell’incremento della durata e dell’intensità del potenziale d’azione dei motoneuroni. Quest’effetto – opposto all’abituazione – viene innescato da un aumento dei livelli di serotonina rilasciata dagli interneuroni in risposta allo stimolo doloroso. La serotonina innesca una cascata di reazioni nei neuroni sensoriali che si traduce in un aumento della permeabilità agli ioni potassio, in un maggiore rilascio di neurotrasmettitore, e quindi in un allungamento sia del potenziale d’azione nel tempo sia dell’eccitabilità presinaptica. In caso di una nuova stimolazione, anche di bassa intensità, la comunicazione fra i neuroni sensoriali e motori risulta rafforzata. A queste elementari forme di a. si possono associare variazioni strutturali a livello pre e postsinaptico, con una diminuzione dei contatti sinaptici nel caso di abituazione e un aumento nella sensibilizzazione. Si è inoltre osservato che, con il ripetersi dello stimolo, la serotonina modula non solo la permeabilità di membrana, ma anche l’espressione genica. In partic., il passaggio da memoria a breve termine a memoria a lungo termine coinvolge l’attivazione di proteine chinasi intracellulari e, a livello del nucleo, del fattore di trascrizione CREB, che promuove la trascrizione di geni e la sintesi di proteine necessarie per la formazione di nuovi contatti sinaptici tra i neuroni. Sempre in Aplysia, è stato anche possibile misurare gli eventi molecolari che sottendono il condizionamento classico utilizzando un paradigma sperimentale in cui la stimolazione tattile del mantello è seguita da una stimolazione avversiva (un lieve shock elettrico) sulla coda del mollusco. Dopo alcuni accoppiamenti dei due stimoli, la sola stimolazione tattile è in grado di provocare la risposta di retrazione del mantello. I meccanismi molecolari dell’a. associativo sembrano gli stessi coinvolti nel processo di sensibilizzazione: in seguito alla presentazione dello stimolo condizionato si osserva attivazione delle proteine chinasi, aumento di ioni potassio, ingresso massiccio di ioni Ca2+ nel neurone presinaptico sensoriale, facilitazione del rilascio di neurotrasmettitore, con il conseguente aumento della durata e dell’intensità del potenziale d’azione postsinaptico. Analoghi meccanismi sono stati osservati anche in altri invertebrati, per es. nel moscerino della frutta Drosophila melanogaster. Laura Ricceri