
Attenzione selettiva
Attenzione selettiva
Durante la vita quotidiana siamo in contatto con molteplici oggetti, alcuni dei quali possono essere utili per i nostri scopi comportamentali; abbiamo perciò bisogno di meccanismi di attenzione selettiva per rapportarci correttamente agli stimoli che risultano rilevanti. Studi neurofisiologici suggeriscono che i meccanismi dell'attenzione sono necessari sia per modulare l'attività di scarica dei neuroni e fare in modo che essi reagiscano in modo ottimale alla stimolazione, sia per combinare fra loro le caratteristiche fisiche degli oggetti e consentirne una percezione corretta. In questo saggio passeremo in rassegna i dati acquisiti sulle basi fisiologiche dell'attenzione visiva, in parallelo agli studi psicologici dell'attenzione in soggetti normali. A questi studi faremo anche riferimento per comprendere i disturbi neuropsicologici dell'attenzione, in particolare la sindrome della negligenza visiva unilaterale. Nel complesso, i risultati suggeriscono l'esistenza di diversi meccanismi cerebrali di elaborazione selettiva dell'informazione che, agendo di concerto, guidano l'attenzione selettiva verso gli oggetti.
Basi neuronali dell'attenzione selettiva
Gli studi neurofisiologici sull'elaborazione visiva nel cervello rivelano che l'attività di scarica delle cellule si adegua allo stato di attenzione dell'animale. Le cellule non si attivano in modo passivo in seguito a stimoli visivi, ma le loro reazioni vengono piuttosto modulate dall'attenzione.

J. Moran e R. Desimone (1985) sono stati i primi a indicare gli effetti della modulazione dell'attenzione sulle cellule neuronali che rispondono a segnali visivi. Questi autori hanno registrato dall'area V4 del cervello di scimmie, un'area visiva della corteccia situata lungo la linea di separazione tra le cortecce occipitale e temporale, e che notoriamente contiene cellule che reagiscono in maniera selettiva al colore (per esempio, una cellula può eccitarsi per una striscia rossa ma non per una verde). Ogni cellula possiede un campo recettivo sulla retina: se lo stimolo a cui la cellula risponde preferenzialmente si presenta all'interno del campo recettivo, la cellula si attiva, ma cessa di fado quando lo stimolo cade al di fuori del suo campo recettivo. Moran e Desimone hanno presentato dunque due stimoli all'interno del campo recettivo delle cellule dell'area V4, uno stimolo appropriato per una data cellula, per esempio una striscia rossa, e uno non appropriato, per esempio una striscia verde. Generalmente una cellula non si attiva fortemente in seguito a tali stimoli composti, in quanto produce una risposta quantitativamente intermedia tra quelle che emetterebbe per ciascuno di essi. Tuttavia, questi ricercatori hanno addestrato gli animali a prestare attenzione a uno degli stimoli, cioè mantenendo gli occhi fissi da un'altra parte. È interessante notare che quando l'animale è attento alla posizione dello stimolo appropriato, la cellula si attiva fortemente, come se questo fosse presentato da solo; quando invece presta attenzione alla posizione dello stimolo non appropriato la cellula non si attiva, come se quello appropriato non fosse più presente. In altre parole, il campo recettivo della cellula sembra 'restringersi' per adattarsi alle dimensioni dell' area del campo visivo su cui si concentra l'attenzione. Bisogna notare che la configurazione di stimoli mostrata all'animale era sempre la stessa, indipendentemente dal fatto che l'attenzione venisse concentrata sullo stimolo preferito o su quello non preferito: perciò le risposte delle cellule non venivano modulate esclusivamente dal segnale in entrata. Il lavoro di Moran e Desimone (1985) suggerisce, quindi, un meccanismo di attenzione selettiva basato sull'elaborazione preferenziale della posizione dello stimolo: l' attenzione selettiva modifica i campi recettivi delle cellule affinché essi si conformino alla posizione del segnale su cui si concentra l'attenzione. In uno studio successivo, L. Chelazzi e collaboratori (1993) hanno analizzato un secondo meccanismo neuronale dell'attenzione selettiva, che coinvolge stavolta anche le proprietà dello stimolo oltre alla sua posizione. Questi autori hanno sottoposto alcune scimmie a un compito chiamato delayed matching to sample (corrispondenza differita con lo stimolo campione). In questo paradigma sperimentale (fig. 1), al soggetto viene dapprima mostrato uno stimolo (per esempio, un triangolo); quindi lo stimolo campione viene ritirato e l'animale deve attendere un periodo di durata variabile. Successivamente, lo stimolo campione viene ripresentato insieme a uno o più stimoli irrilevanti (per esempio, un quadrato e un cerchio), chiamati distrattori, e il compito del soggetto è di emettere una risposta verso lo stimolo campione ignorando i distrattori. In questo caso, la risposta corretta consisteva nel compiere un movimento degli occhi verso lo stimolo bersaglio. Le scimmie non potevano prevedere quale sarebbe stata la posizione nel campo visivo in cui sarebbero stati presentati gli stimoli, quindi dovevano ricordare le proprietà percettive dello stimolo corretto (cioè forma, colore, ecc.) per distinguerlo dal distrattore.
Chelazzi e collaboratori (1993) hanno studiato in questa situazione sperimentale le risposte delle cellule della corteccia inferotemporale, immediatamente al di sotto dell'area V4 studiata da Moran e Desimone (1985). Questi autori hanno scoperto che le cellule, la cui attività era modulata dalle proprietà dello stimolo bersaglio, mantenevano un tasso di attivazione relativamente alto durante l'intervallo precedente l'apparizione dello stimolo bersaglio e del distrattore. In sostanza, esse dopo essere state esposte allo stimolo bersaglio prima dell' intervallo, durante l'intervallo rimanevano in uno stato di preattivazione o di sensibilità elevata alle proprietà percettive del bersaglio. Quando venivano presentati il bersaglio e il distrattore, le cellule già preattivate sensibili al bersaglio aumentavano ulteriormente la propria attivazione; al contrario, le cellule sensibili al distrattore, e quindi non preattivate durante l'intervallo, diventavano inizialmente attive per poi diminuire rapidamente il proprio ritmo di scarica. Il punto importante è che la prevalenza delle cellule facilitate, che reagivano al bersaglio, sulle cellule non facilitate, che reagivano al distrattore, si verificava molto prima che l'animale spostasse lo sguardo verso il bersaglio. Chelazzi e collaboratori hanno concluso perciò che l'attenzione selettiva per il bersaglio si verificava grazie a un processo di feature priming (facilitazione dipendente dalle caratteristiche dell'oggetto). Le cellule sensibili alle proprietà dello stimolo bersaglio erano attivate già prima dell'apparizione del bersaglio e del distrattore. Questo dava alle cellule sensibili al bersaglio un vantaggio su quelle sensibili al distrattore nella produzione della risposta corretta, come accadrebbe se le diverse cellule fossero in competizione per rappresentare lo stimolo mostrato all'animale. Mediante un processo di inibizione selettiva di gruppi di cellule sensibili agli stimoli irrilevanti, i neuroni nel lobo inferotemporale possono rappresentare dunque solo un oggetto, cioè il bersaglio selezionato.
Questi studi indicano che esistono almeno due modi alquanto diversi grazie ai quali il cervello realizza ciò che si intende per attenzione selettiva: facilitare l'elaborazione di una certa posizione del campo visivo in cui uno stimolo atteso può comparire (Moran e Desimone, 1985), oppure facilitare l'elaborazione delle caratteristiche percettive di un oggetto indipendetemente dalla sua posizione (Chelazzi et al., 1993). Come vedremo più avanti, tale distinzione tra forma di attenzione 'spaziale' e forma di attenzione 'centrata sull'oggetto' sembra essere anche importante per comprendere i disturbi di attenzione selettiva di cui soffrono alcuni pazienti con lesioni cerebrali (v. oltre).
Altri studi neurofisiologici hanno dimostrato che i neuroni possono reagire in maniera selettiva a diverse caratteristiche dell'informazione visiva: al colore, al movimento, alla forma, e così via (Livingstone e Rubel, 1988). Nonostante ciò, nella nostra percezione, i colori sono integrati con la forma e il movimento. Questo processo di integrazione rappresenta per il cervello ciò che potremmo chiamare un 'problema di accoppiamento', che consiste nel calcolare come una certa forma vada associata a un colore per riconoscere un oggetto nella sua intrinseca unità, soprattutto quando ci sono molti oggetti presenti nello spazio visivo. In parole povere, se abbiamo quattro cellule che reagiscono rispettivamente al colore rosso, al colore verde, al quadrato e al cerchio, come farà il sistema cognitivo a rendersi conto del fatto di essere di fronte a un cerchio rosso e un quadrato verde anziché, per esempio, un quadrato rosso e un cerchio verde, visto che verranno attivate tutte le quattro cellule? Un modo per accoppiare insieme gli attributi potrebbe essere l'attenzione selettiva spaziale. Se quest'ultima attiva le cellule sensibili a tutti gli stimoli che si presentano in una determinata posizione, allora solamente queste raggiungeranno livelli elevati di eccitazione (Treisman, 1998). In tal modo, la rappresentazione neuronale del colore rosso e quella della forma del cerchio saranno associate, poiché il rosso e il cerchio si presentano congiuntamente in una certa posizione del campo visivo e quindi l'osservatore 'vedrà' proprio un cerchio rosso. Inoltre, le cellule che si trovano a livelli gerarchici più alti del sistema visivo, come appunto il lobo inferotemporale, possiedono campi recettivi molto ampi e, pertanto, rispondono ai segnali indipendentemente dal punto esatto in cui gli stimoli si presentano (Desimone e Ungerleider, 1989). Questo fatto è importante, probabilmente, per il riconoscimento degli oggetti, in quanto permette che essi vengano riconosciuti dalle stesse cellule anche quando si presentano in punti diversi della retina; questa forma di flessibilità è definita invarianza del punto di vista (Humphreys e Heinke, 1998). Tuttavia, quando nel mondo percettivo dell'osservatore sono presenti molteplici oggetti, il campo recettivo di una certa cellula potrà contenere probabilmente stimoli appropriati e non appropriati e la cellula, quindi, potrebbe non essere mai fortemente attivata. Ancora una volta, l'attenzione potrebbe essere necessaria per assicurare che i campi recettivi siano sensibili in modo selettivo alla posizione degli stimoli appropriati, oppure per assicurare che le cellule relative agli stimoli bersaglio vengano preattivate e possano così vincere la competizione contro le cellule che codificano il distrattore, anche quando altri stimoli irrilevanti si presentano all'interno del campo recettivo. In questo saggio verrà ribadito ulteriormente che è necessario comprendere i processi dell'attenzione nel cervello per poter risolvere un problema centrale nella visione: come impedire che le cellule corticali reagiscano in maniera non ottimale quando nel campo recettivo ci sono oggetti diversi, e come far sì che le informazioni codificate da cellule diverse vengano associate.
Dati provenienti dalla psicologia cognitiva
Attenzione per lo spazio
Nell'uomo, gli studi sull'attenzione visiva indicano anche che essa può essere rivolta allo spazio oppure agli oggetti. M. 1. Posner (1980) ha dimostrato che il tempo impiegato per individuare uno stimolo visivo si riduce se gli osservatori vengono indotti a prestare attenzione alla posizione del bersaglio prima che questo venga presentato. Posner suggerisce dunque che, quando si concentra l'attenzione sulla posizione del campo visivo in cui comparirà uno stimolo, l'elaborazione di quella posizione viene potenziata e la selezione dello stimolo che occupa tale posizione viene facilitata. Gli effetti benefici dell'attenzione selettiva si verificano anche quando al soggetto dell'esperimento viene impedito il movimento degli occhi: questo dimostra che esiste la possibilità di aumentare l'efficacia dell'elaborazione visiva anche indipendentemente da movimenti espliciti della fissazione oculare. Lavori successivi hanno dimostrato che gli stessi processi coinvolti ne II 'orientare l'attenzione verso lo spazio possono essere dissociati. L'attenzione viene orientata piuttosto automaticamente verso gli stimoli che forniscono alla visione periferica forti segnali transitori, ma può essere anche orientata in maniera più controllata dietro istruzioni precise (Miiller e Rabbit, 1989).
All'interno di una regione dello spazio visivo a cui è rivolta l'attenzione, l'elaborazione degli stimoli sembra essere approfondita, e ciò può produrre una competizione tra le risposte ai vari stimoli se esse sono incompatibili l'una con l'altra. Per esempio, nel cosiddetto compito di Eirksen, i soggetti devono rispondere a una lettera bersaglio, presentata centralmente, che può essere affiancata da lettere con funzione di distrattore, le quali richiedono la stessa risposta al bersaglio o una differente. l tempi di reazione sono rallentati quando le risposte ai distrattori sono incompatibili con la risposta al bersaglio (Eirksen, 1995). Tuttavia, separando spazialmente gli stimoli fra loro, si riducono gli effetti di tale competizione (Eriksen e Eriksen, 1974).
Attenzione per gli oggetti
La distanza tra gli elementi di una configurazione di stimoli ha un chiaro effetto sull' attenzione visiva, ma non è affatto l'unico fattore importante. Per esempio, J. Driver e G. C. Baylis (1989) hanno mostrato che effetti di interferenza tra le risposte si possono produrre con coppie di stimoli lontani nello spazio, quando gli stimoli si spostano contemporaneamente. In questo caso, la percezione del movimento indotta dal movimento comune sopprime gli effetti della separazione degli stimoli nello spazio sull'attenzione selettiva.
Altri studiosi hanno usato tempi brevi di presentazione degli stimoli per analizzare i limiti del processo di attenzione selettiva. In genere si riscontra che le persone, difficilmente, riescono a selezionare più di un oggetto alla volta se il ritmo di presentazione è molto rapido; infatti, l'individuazione di un secondo oggetto risulta difficoltosa. Ciò si verifica quando il compito richiede che i soggetti individuino due attributi o caratteristiche di oggetti diversi. È interessante notare che se il compito richiede di individuare due attributi dello stesso stimolo, il livello di prestazione dei soggetti è leggermente inferiore a quello ottimale (Duncan, 1984; Baylis e Driver, 1993). Sembra infatti che l'essere umano sia in grado di prestare attenzione contemporaneamente a tutti gli attributi di un singolo oggetto, ma non a quelli di oggetti diversi. Questa tendenza a selezionare gli attributi di un unico oggetto si riscontra anche quando la distanza tra gli attributi è comparabile in condizioni che prevedono uno o molteplici oggetti.
Questi studi dimostrano che le persone possono rivolgere l'attenzione sia agli oggetti sia alla loro localizzazione spaziale. Una questione aperta è il modo in cui interagiscono queste due forme di selezione. Un'ipotesi è che esse agiscano in parallelo. L'attenzione prestata a una posizione nello spazio potenzia l'elaborazione degli oggetti che si trovano in quella regione, influenzando la selezione basata sugli oggetti. Similmente, l'attivazione comune di parti degli oggetti obbliga l'attenzione spaziale a concentrarsi su quelle regioni del campo visivo in cui si trovano tali attributi, influenzando la selezione basata sullo spazio (Farah et al., 1993; Humphreys e Riddoch, 1993; Schneider, 1995). Questa idea di sinergia tra il sistema di selezione basato sullo spazio e quello basato sugli oggetti è sostenuta da prove sperimentali, le quali indicano che viene facilitata l'identificazione di stimoli presentati nella posizione spaziale verso cui è diretta un'azione di risposta (per esempio, il movimento degli occhi o della mano), anche se i soggetti non sono stati allertati preliminarmente a prestare attenzione a tale posizione (Deubel e Schneider, 1996; Deubel et al., 1998). In questo caso, l'azione diretta verso una certa posizione nello spazio sembra avere effetti enormi sui processi di identificazione che presumibilmente hanno luogo all'interno di un sistema di codifica basato sugli oggetti.
È interessante notare che una delle conseguenze derivanti dagli errori che commettiamo quando non prestiamo attenzione agli oggetti è che potremmo non accoppiare correttamente le loro proprietà. A. Treisman e H. Schmidt (1982) hanno evidenziato che quando i soggetti non prestano completa attenzione alle lettere colorate che vengono loro presentate per breve tempo, spesso nei resoconti di risposta essi attribuiscono a una certa lettera il colore sbagliato. Inoltre, i soggetti spesso sono convinti di aver visto un accoppiamento del tutto illusorio di lettera e colore, e ciò suggerisce che tali errori siano di origine percettiva. Tali accoppiamenti illusori sarebbero ipotizzabili se le cellule che rispondono a lettere familiari avessero larghi campi recettivi in cui possono ricadere numerosi colori, quando al soggetto sono presentate molte lettere. Questi errori possono essere evitati se i soggetti prestano attenzione alla posizione della lettera bersaglio; ciò può aiutare a ridurre l'ampiezza del campo recettivo per quella lettera, in modo tale che soltanto il colore appropriato venga associato alla forma (Luck et al., 1997). Quest'ultima ipotesi è avvalorata dai risultati indicanti che l'accoppiamento illusorio si verifica in maniera più frequente quando gli stimoli sono vicini nello spazio rispetto a quando sono distanti (Cohen e lvry, 1989).
Bisognerebbe sollevare un'ultima questione sul problema della connessione tra gli attributi visivi: le ricerche svolte sull' attenzione selettiva centrata sugli oggetti suggeriscono che le parti possono essere percepite in connessione (raggruppate) anche senza l'attenzione. lnfatti, se le parti non venissero percepite in connessione per formare gli oggetti, sarebbe difficile spiegare come possano verificarsi gli effetti sull'attenzione dipendenti dall'oggetto. Tuttavia la sintesi percettiva senza attenzione potrebbe essere piuttosto instabile e, in tal caso, si verificherebbero alcuni accoppiamenti illusorio Inoltre, la sintesi operata dall'attenzione potrebbe anche essere particolarmente importante quando il compito richiedesse di unire fra loro connotati diversi di uno stesso oggetto, come il colore e la forma.
Dati provenienti dalla neuropsicologia
In seguito a danni cerebrali, le persone possono avere alcuni problemi nel prestare attenzione agli stimoli in maniera selettiva. Ciò appare particolarmente evidente nella sindrome della cosiddetta negligenza unilaterale; i pazienti affetti da questo disturbo cognitivo possono non reagire agli stimoli presentati sul lato dello spazio contro laterale rispetto alla lesione: per esempio, stimoli presentati alla sinistra dei pazienti con danni all'emisfero cerebrale destro non danno luogo ad alcuna risposta evidente. La negligenza non è causata semplicemente dalla cecità locale di una parte del campo visivo. lnfatti, i pazienti di questo tipo generalmente non trascurano gli stimoli presentati nella porzione cieca del campo quando le condizioni di osservazione non richiedono un'ispezione rapida, o se essi possono muovere gli occhi (Halligan et al., 1990). Al contrario, i pazienti affetti dalla sindrome della negligenza unilaterale falliscono anche quando non devono riferire di oggetti presentati realmente ma soltanto immaginare una scena (Bisiach e Luzzatti, 1978), così come tendono a presentare problemi particolari nel rispondere a stimoli contro laterali, se la loro attenzione è già impegnata con uno stimolo visivo nel campo ipsilaterale (Posner et al., 1984). La negligenza può essere interpretata come perdita di una qualche forma di rappresentazione interna dello spazio (Bisiach e Luzzatti, 1978; Caramazza e Hillis, 1990), e la gravità di questo disturbo può senz'altro essere alleviata allenando i pazienti a concentrare l'attenzione sul lato colpito (Riddoch e Humphreys, 1983). Gli studi su tale sindrome possono quindi far luce sui meccanismi neuronali che sono alla base dell'attenzione selettiva basata sullo spazio. Tali studi forniscono prove a sostegno del fatto che l'attenzione per gli oggetti e quella per lo spazio possono essere colpite l'una indipendentemente dall'altra; ciò indicherebbe non solo che queste due forme di attenzione esistono, ma che esse possono risiedere in strutture neuronali distinte. Tale dissociazione funzionale tra i due disturbi appare particolarmente evidente in pazienti che mostrano negligenza per lo spazio oppure negligenza per gli oggetti, a seconda della prova a cui sono sottoposti.
G.W. Humphreys e M.I Riddoch (1994; 1995) hanno descritto un paziente, JR, che manifestava negligenza a sinistra o a destra a seconda del fatto che questi concentrasse l'attenzione sugli oggetti singoli o sulle posizioni di oggetti distinti nello spazio: si verificava negligenza del lato sinistro per i singoli oggetti, ma negligenza del lato destro per lo spazio. Il contrasto poteva emergere persino con lo stesso stimolo. Quando si chiedeva al paziente di leggere parole e stringhe di lettere per intero, cioè concentrarsi sulle lettere come se fossero parti di un unico oggetto, si verificava negligenza a sinistra: perciò la parola "part" (parte) veniva letta "dart" (dardo). Quando invece si chiedeva al paziente di leggere ogni lettera a voce alta, cioè concentrarsi sulle singole lettere come se fossero oggetti indipendenti, si verificava negligenza a destra: JR quindi leggeva in maniera corretta le lettere che aveva trascurato nella prova precedente e ometteva quelle che aveva letto prima. Quindi, la parola "part" veniva letta "p, a, n>. JR aveva lesioni bilaterali che interessavano le regioni occipitoparietale sinistra e frontoparietale destra, e così la sua negligenza a sinistra, nel caso degli oggetti singoli, poteva essere causata dalla lesione frontoparietale destra, mentre quella a destra poteva dipendere dalla lesione occipitoparietale sinistra.
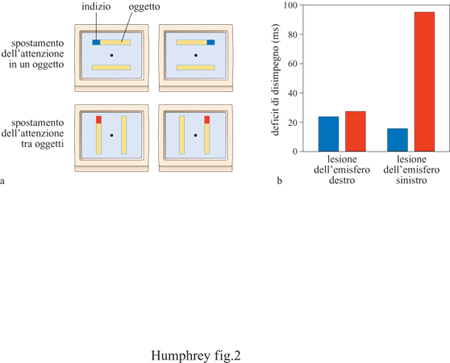
R. Egly e collaboratori (1994) hanno studiato un campione di pazienti con lesioni parietali e hanno dimostrato che quelli con danni al lobo parietale destro hanno difficoltà a prestare attenzione agli oggetti singoli, mentre quelli con danni al lobo parietale sinistro hanno difficoltà a spostare l'attenzione tra oggetti diversi nello spazio (fig. 2). Humphreys e D. Heinke (1998) hanno anche mostrato che possono esistere dissociazioni doppie tra pazienti che mostrano negligenza per gli oggetti singoli e quelli con negligenza tra gli oggetti distinti nello spazio. Questi autori hanno sottoposto i pazienti a una serie di prove che richiedevano o di concentrarsi sulle parti di alcuni oggetti come, per esempio, leggere parole o riconoscere visi, oppure di spostare l'attenzione da un oggetto all'altro, cioè da una parola all'altra o da un viso a un altro. Alcuni pazienti presentavano negligenza per gli oggetti singoli in tutte le prove, mentre questa era lieve quando dovevano spostare l'attenzione da un oggetto a un altro, anche se il secondo si trovava nel campo visivo compromesso, ovvero quello contro laterale alla lesione. Per esempio, questi pazienti potevano non riuscire a leggere la lettera più a sinistra in ognuna delle due parole che venivano loro presentate, ma riuscivano comunque a spostare l'attenzione dalla parola posta a destra a quella posta a sinistra. Il problema in questo caso consisteva nel selezionare le informazioni presenti sul lato di un oggetto contro laterale alla lesione, indipendentemente dalla posizione di questo rispetto al corpo del paziente (v. anche Behrmann e Tipper, 1994; Tipper e Behrmann, 1996). Al contrario, altri pazienti non mostravano negligenza per gli oggetti singoli, ma trascuravano quelli posti nell' emicampo visivo contro laterale alla lesione, per cui riuscivano a leggere in maniera corretta, tra due parole, quella più a destra, ma non individuavano la parola a sinistra. Queste doppie dissociazioni suggeriscono che i meccanismi che guidano l'attenzione per gli oggetti possono essere separati funzionalmente da quelli per gli oggetti distinti nello spazio. Nel cervello umano, tali meccanismi possono anche essere localizzati funzionalmente all'interno di ogni emisfero.
Altre osservazioni neuropsicologiche, provenienti dallo studio del fenomeno della estinzione spaziale, confermano che la modulazione dell' attenzione centrata sullo spazio e quella sugli oggetti possono essere dissociate in seguito a un danno cerebrale. L'estinzione spaziale consiste nel fatto che un paziente può individuare in maniera corretta uno stimolo unico presentato nell' emicampo visivo controlaterale rispetto alla lesione, ma non riesce a fado quando un altro stimolo viene presentato contemporaneamente nell'emicampo visivo normale (ipsilaterale), come se lo stimolo 'forte' nell' emicampo intatto inibisse l' individuazione dello stimolo 'debole' nell'emicampo danneggiato. Diversi studi hanno dimostrato che l'estinzione può essere ridotta raggruppando gli stimoli presentati nei campi contro laterale e ipsilaterale del paziente. L'oggetto 'estinto' può essere recuperato se il paziente ne rileva qualche somiglianza con lo stimolo ipsilaterale (Gilchrist et al., 1996; Mattingley et al., 1997a; Ward et al., 1994). Questo risultato è interessante perché suggerisce che l'informazione visiva relativa agli stimoli estinti è comunque elaborata a qualche livello e consente l'associazione con l'oggetto ipsilaterale.
Siccome i pazienti non riescono a individuare stimoli presentati simultaneamente, sembra dunque che l'informazione visiva venga elaborata inconsciamente. I. Gilchrist e collaboratori (1996), per esempio, hanno presentato a un paziente, con estinzione a sinistra, coppie di stimoli, differenti per la forza dell' associazione, nell' emicampo destro o in quello sinistro: gli stimoli potevano formare un gruppo singolo oppure potevano non essere associati in alcun modo e, perciò, venivano percepiti come due oggetti separati. In generale, la prestazione risultava migliore quando gli stimoli venivano associati rispetto a quando ciò non si verificava. Inoltre il paziente otteneva risultati migliori con gli stimoli presentati nell' emicampo destro rispetto a quelli presentati nell' emicampo sinistro. Tuttavia, l'effetto dell' associazione era dello stesso ordine di grandezza in ogni campo. In questo caso sembra che l'associazione sia ugualmente forte e che influenzi allo stesso modo la selezione su entrambi i lati dello spazio. Inoltre, sembra esserci un'inclinazione spaziale a favore degli stimoli presentati nel campo destro. Pertanto il disturbo dell' attenzione selettiva spaziale (che colpisce un campo visivo più dell'altro) sembra essere indipendente dagli effetti di associazione che, invece, non siano polarizzati spazialmente.
Gli studi clinici sulla negligenza e sull'estinzione di cui abbiamo riferito sopra convergono con i risultati della ricerca neurofisiologica e psicologica nel suggerire che esistono due meccanismi dell'attenzione distinti, uno riguardante la selezione delle posizioni degli oggetti nello spazio, l'altro la selezione delle parti che appartengono a un oggetto. Sebbene, generalmente, questi due meccanismi agiscano insieme, essi possono essere colpiti indipendentemente l'uno dall'altro in seguito a danni cerebrali.
Un'ulteriore conseguenza della lesione cerebrale, per cui i sistemi di codifica basati sullo spazio e sull'oggetto vengono dissociati, è che i pazienti hanno difficoltà ad accoppiare tra loro le proprietà degli oggetti. Per esempio, A. Cohen e R. Rafal (1991) hanno scoperto che l'accoppiamento illusorio di colori e forme, di solito, colpisce i pazienti con danno parietale unilaterale quando gli stimoli vengono presentati nell'emicampo visivo contro laterale rispetto alla lesione. Effetti ancor più imponenti si riscontrano quando i pazienti riportano danni parietali bilaterali: in tal caso essi possono commettere molti errori di accoppiamento illusorio anche in condizioni di osservazione libera e indipendentemente dalla posizione degli stimoli nel campo visivo (Friedman-Hill et al., 1995; Treisman, 1998). Questi pazienti sembrano avere difficoltà a concentrare l'attenzione sulla posizione dello stimolo, e quindi non riescono ad accoppiare le proprietà degli oggetti quando nel campo visivo viene presentato più di uno stimolo. L'attenzione prestata alla posizione spaziale può facilitare l'elaborazione delle caratteristiche che si trovano in tale posizione e che verranno accoppiate fra loro, e non la percezione di altre caratteristiche non rilevanti e che, quindi, sono attivate in maniera più debole. Questa ipotesi tuttavia, come nel caso degli studi psicologici dell'attenzione per gli oggetti in persone normali, viene resa meno credibile dal fatto che alcune forme di accoppiamento di caratteristiche possono ancora intervenire senza attenzione.
Come abbiamo notato, il fenomeno dell'estinzione viene influenzato dall'associazione degli elementi visivi e ciò sembra aver luogo prima che l'attenzione volontaria inizi a operare, consentendo ai pazienti di selezionare le informazioni rilevanti a cui prestare attenzione. Inoltre, persino in pazienti con lesioni parietali bilaterali e attenzione spaziale danneggiata, il riconoscimento degli oggetti può rimanere relativamente intatto: ciò suggerisce che tali pazienti conservano la capacità di accoppiare le parti degli oggetti ai fini del riconoscimento. Di fatto, in tali casi, l'entità dell'estinzione può dipendere dalla forza dell' associazione tra gli stimoli che formano il gruppo: se vengono presentati stimoli che formano un gruppo 'forte' e stimoli che formano un gruppo meno 'forte', i pazienti riportano lo stimolo che fa parte del gruppo 'forte' (Hamphreys et al., 1994). Ancora una volta potrebbero essere vere due ipotesi: l'attenzione non è necessaria per tutte le forme di accoppiamento, ma senza l'attenzione spaziale qualsiasi forma di accoppiamento tra proprietà è instabile, e perciò i pazienti tendono a fare accoppiamenti illusori più spesso del normale; in alternativa solo alcune forme di accoppiamento richiedono l'attenzione spaziale, mentre altre no. Per esempio, l'accoppiamento tra colore e forma può richiedere l'attenzione, laddove l'accoppiamento tra forme può esserne indipendente e questo fa sì che il riconoscimento di oggetti possa essere preservato in pazienti con lesioni parietali bilaterali.
Attenzione selettiva in altre modalità sensoriali
La discussione sull'attenzione selettiva spaziale si è concentrata interamente sulla visione e ciò per un'ottima ragione, in quanto sono stati raccolti molti più dati neurofisiologici e neuropsicologici sull'attenzione visiva di quanti ne siano disponibili per altri canali sensoriali.
Ciononostante, studi recenti hanno mostrato l'esistenza di associazioni interessanti tra canali sensoriali diversi che influenzano i meccanismi dell'attenzione selettiva. Per esempio, un evento rilevante in una data modalità (per esempio un suono) può allertare l'attenzione in altre modalità (per esempio la visione) verso la sua direzione di provenienza, anche se il suono non predice la posizione dello spazio in cui apparirà uno stimolo visivo (Driver e Spence, 1999). A questo punto occorre domandarsi se le varie informazioni sensoriali in entrata vengono integrate in un unico sistema sopramodale che le organizza per guidare l'attenzione spaziale. La risposta sembra essere negativa per diversi motivi. lnnanzitutto, gli effetti della facilitazione di un canale sensoriale sull' altro nella guida dell' attenzione sono asimmetrici: indizi uditivi producono uno spostamento dell'attenzione visiva verso la posizione indicata, ma non è vero il contrario (Spence e Driver, 1997). Inoltre, l'efficacia di un certo stimolo sensoriale nel modulare l'attenzione è quasi sempre maggiore nel canale sensoriale attivato anche quando gli effetti abbracciano anche altre modalità; per esempio, l'effetto facilitante di un indizio preliminare uditivo è più forte nel rilevare un secondo stimolo uditivo piuttosto che visivo, sebbene in quest'ultimo caso un certo effetto sia comunque riscontrabile. lnfine, a riprova del fatto che gli effetti della facilitazione tra modalità sono complessi, le persone possono dirigere in modo consapevole la propria attenzione verso il lato dello spazio in cui si aspettano la comparsa di un bersaglio visivo, e contemporaneamente verso un altro lato dal quale si aspettano l'arrivo di uno stimolo uditivo (Driver e Spence, 1998).
Pertanto sembrano esistere effetti di orientamento dell'attenzione verso lo spazio all'interno di particolari modalità, cioè orientamento visivo verso indizi visivi, e orientamento uditivo verso indizi uditivi. Tuttavia esistono effetti asimmetrici di facilitazione tra modalità, cosicché indizi preliminari forniti in un canale sensoriale facilitano l'orientamento in un altro canale, come si verifica con indizi uditivi preliminari che facilitano la percezione di bersagli visivi. Tali associazioni possono essere più forti tra alcune modalità rispetto ad altre. Dividere l'attenzione verso lati opposti dello spazio è più difficile nei compiti visivo-tattili, in cui i soggetti aspettano un bersaglio visivo su un lato e uno tattile sull'altro, rispetto ai compiti visivo-uditivi. L'orientamento simultaneo dell'attenzione verso segnali visivi e tattili sembra invece essere più forte rispetto a quello verso segnali visivi e uditivi. Tale legame tra visione e tatto può spiegare perché, in seguito a danni parietali, alcuni pazienti presentino il fenomeno dell'estinzione in queste due modalità: tali pazienti non riescono a individuare uno stimolo visivo controlaterale alla lesione quando vengono toccati contemporaneamente sul lato ipsilaterale del corpo (Mattingly et al., 1997b). Conclusioni l dati sperimentali che abbiamo passato in rassegna indicano che l'attenzione selettiva costituisce una fase cruciale del processo di elaborazione delle informazioni percettive da parte del cervello. Essa è necessaria per far sì che solo le informazioni rilevanti siano disponibili per programmare le azioni comportamentali appropriate. Nel caso della visione, essa può anche essere necessaria per modulare la funzionalità delle cellule neuronali che si trovano ad alti livelli gerarchici del sistema di riconoscimento degli oggetti, al fine di ridurre l'interferenza causata dalla presenza simultanea di molti oggetti nel campo visivo. L'attenzione selettiva interviene anche nella sintesi delle diverse caratteristiche fisiche di un oggetto nella percezione corretta e stabile di questo. Le osservazioni sperimentali neurofisiologiche, psicologiche e neuropsicologiche concordano nel suggerire che esistono diversi meccanismi di attenzione selettiva, alcuni dei quali sono coinvolti nel guidare l'attenzione verso la posizione degli stimoli (selezione basata sullo spazio) e altri nel selezionare interi oggetti (selezione basata sugli oggetti). Generalmente questi meccanismi possono operare in modo congiunto, sebbene questa sinergia possa venir meno in seguito a danni cerebrali. L'orientamento dell'attenzione verso lo spazio visivo può anche essere legato all'orientamento dell'attenzione all'interno di altre modalità (udito, tatto), cosicché le azioni vengono dirette in maniera coerente verso oggetti nello spazio.
Ringraziamenti
Questo lavoro è stato reso possibile dai fondi concessi all'autore dal MedicaI Research Institute del Regno Unito e dal Human Frontier Science Program.
Bibliografia citata
BAYLIS, O.C., DRIVER, J. (1993) Visual attention and objects: evidence for hierarchical coding of location. J. Exp. Psychol. Rum. Percept. Perform. 19, 451-470.
BEHRMANN, M., TIPPER, S.P., (1994) Object-based attentional mechanisms: evidence from patients with unilateral neglect. In Attention and performance, XV, a c. di Umilta C., Moscovitch M., Cambridge, Mass., MIT Press, pp. 351-375.
BISIACH, E., LUZZATTI, C. (1978) Unilateral neglect of representational space. Cortex, 14, 129-133.
CARAMAZZA, A., HILLIS, A.E., (1990) Levels of representation, coordinate frame s, and unilateral neglect. Cognit. Neuropsychol., 7, 391-445.
CHELAZZI, L., MILLER, E.K., DUNCAN, J., DESIMONE, R. (1993) A neural basis for visual search in inferior temporal cortex. Nature, 363, 345-347.
COHEN, A., IVRY, R. (1989) Illusory conjunctions inside and outside the focus of attention. J. Exp. Psychol. Rum. Percept. Perform., 15, 650-663.
COHEN, A., RAFAL, R. (1991) Attention and feature integration: illusory conjunctions in a patient with a parietal lobe lesion. Psychol. Sci., 2, 106-109.
DESIMONE, R., UNGERLEIDER, L.O. (1989) Neural mechanisms of visual processing in monkeys. In Randbook of neurophysiology, voI. 2, a c. di Boller F., Grafrman J., New York, Elsevier, pp. 267-299.
DEUBEL, H., SCHNEIDER, W.X. (1996) Saccade target selection and object recognition: evidence for a common attentional mechanism. Vision Res., 36, 1827-1837.
DEUBEL, H., SCHNEIDER, W.X., PAPROTTA, I. (1998) Selective dorsal and ventral processing: evidence for a common attentional mechanism in reaching and perception. Visual Cognit., 5, 81-108.
DRIVER, J., BAYLIS, O.C. (1989) Movement and visual attention: the spotlight metaphor breaks down. J. Exp. Psychol. Rum. Percept. Perform., 15, 448-456.
DRIVER, J., SPENCE, C. (1998) Cross modallinks in spatial attention. Philos. Trans. R. Soc. Lond. B Biol. Sci., 353, 1319-1331.
DRIVER, J., SPENCE, C. (1999) Crossmodallinks in spatial attention. InAttention, space and action: the cognitive neuroscience of selective perception and action, a c. di Humphreys O.W., Duncan J., Treisman A., Oxford-New York, Oxford University Press.
DUNCAN, J. (1984) Selective attention and the organization of visual information. J. Exp. Psychol. Gen., 113, 501-517.
EGLY, R., DRIVER, J., RAFAL, R.D. (1994) Shifting visual attention between objects and locations: evidence from normal and parietallesion subjects. J. Exp. Psychol. Gen., 123, 161-177.
ERIKSEN, B.A., ERIKSEN, C.W. (1974) Effects of noise letters upon the identification of a target letter in a nonsearch task. Percept. Psychophysi., 16, 143-149.
ERIKSEN, C.W. (1995) The flankers task and response competition: a useful tool for investigating a variety of cognitive problems. Visual Cognit., 2, 101-118.
FARAH, M.J., WALLACE, M.A., VECERA, S.P. (1993) What and where in visual attention: evidence from the neglect syndrome. In Unilateral neglect: clinical and experimental studies, a c. di Robertson I., Marshall J.C., Londra, Lawrence Erlbaum Associates, pp. 123-137.
FRIEDMAN-HILL, S.R., ROBERTSON, L.C., TREISMAN, A. (1995) Parietal contributions to visual feature binding: evidence from a patient with bilaterallesions. Science, 269, 853-855.
GILCHRIST, I., HUMPHREYS, G.W., RIDDOCH, M.J. (1996) Grouping and extinction. Cognit. Neuropsychol., 13, 1223-1256.
HALLIGAN, P.W., MARSHALL, J.C., WADE, D.T. (1990) Do visual field deficits exacerbate visuo-spatial neglect? J. Neurol. Neurosurg. Psychiatry, 53, 487-491.
HUMPHREYS, G.W., HEINKE, D. (1998) Spatial representation and selection in the brain: neuropsychological and computational constraints. Visual Cognit., 5, 9-48.
HUMPHREYS, G.W., RIDDOCH, M.J. (1993). Interactions between object and space systems revealed through neuropsychology. In Attention and performance, XIV, a c. di Meyer D.E., Komblum S., Hillsdale, Lawrence Erlbaum Associates, pp. 143-162.
HUMPHREYS, G.W., RIDDOCH, M.J. (1994) Attention to withinobject and between-object spatial representations: multiple sites for visual selection. Cognit. Neuropsychol., 11, 207-242.
HUMPHREYS, G.W., RIDDOCH, M.J. (1995) Separate coding of space within and between perceptual objects: evidence from unilateral visual neglect. Cognit. Neuropsychol., 12, 283-311.
HUMPHREYS, G.W., ROMANI, C., OLSON, A., RIDDOCH, M.J., DUNCAN, J. (1994). Non-spatial extinction following lesions ofthe parietallobe in humans. Nature, 372, 357-359.
LIVINGSTONE, M., HUBEL, D. (1988) Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science, 240, 740-749.
LUCK, S.J., GIRELLI, M., McDERMOTT, M.T., FORD, M.A. (1997) Bridging the gap between monkey neurophysiology and human perception: an ambignity resolution theory of visual selective attention. Cognit. Psychol., 33, 64-87.
MATTINGLEY, J.B., DAVIS, G., DRIVER, J. (1997a) Preattentive filling-in of visual surfaces in parietal extinction. Science, 275, 671-674.
MATTINGLEY, J.B., DRIVER, J., BESCHIN, N., ROBERTSON, I.H. (1997b) Attentional competition between modalities: extinction between touch and vision after right hemisphere damage. Neuropsychologia, 35, 867-880.
MORAN, J., DESIMONE, R. (1985) Selective attention gates visual processing in the extrastriate cortex. Science, 229, 782-784.
MULLER, H.J., RABBITT, P.M. (1989) Reflexive and voluntary orienting of visual attention: time course of activation and resistance to interruption. J. Exp. Psychol. Rum. Percept. Perform., 15, 315-330.
POSNER, M.1. (1980) Orienting of attention. Q. J. Exp. Psychol., 32, 3-25.
POSNER, M.I., WALKER, J.A., FRIEDRICH, P.J., RAFAL, R.D. (1984) Effects of parietal injury on covert orienting of attention. J. Neurosci., 4, 1863-1874.
RIDDOCH, M.J., HUMPHREYS, G.W. (1983) The effects of cueing on unilateral neglect. Neuropsychologia, 21, 589-599.
SCHNEIDER, WX. (1995) V AM: a neuro-cognitive model for visual attention control of segmentation, object recognition and space-based motor action. Visual Cognit., 2, 331-375.
SPENCE, C., DRIVER, J. (1997) Audiovisual links in exogenous covert spatial orienting. Percept. Psychophys., 59, 1-22.
TIPPER, S.P., BEHRMANN, M. (1996) Object-centered not scenebased visual neglect. J. Exp. Psychol. Rum. Percept. Perform., 22, 1261-1278.
TREISMAN, A. (1998) Feature binding, attention and object perception. Philos. Trans. R. Soc. Lond. B Biol. Sci., 353, 1295-1306.
TREISMAN, A., SCHMIDT, H. (1982) Illusory conjunctions in the perception of objects. Cognit. Psychol., 14, 107-141.
WARD, R., GOODRICH, S., DRIVER, J. (1994) Grouping reduces visual extinction: neuropsychological evidence for weight-linkage in visual selection. Visual Cognit., l, 101-130.