
Biologia
Biologia
di Emanuele Padoa ed Eraldo Antonini
BIOLOGIA
Biologia di Emanuele Padoa
sommario: 1. Introduzione. 2. Caratteri essenziali degli organismi viventi. 3. Le proteine: struttura e funzione come due aspetti dei fenomeni di biologia molecolare. 4. Un gene, una proteina: a) cromatina, cromosomi e geni; b) gli acidi nucleici e il valore genico del DNA; c) la duplicazione del DNA; d) la sintesi proteica e il codice genetico che ne porta l'informazione. 5. La modulazione dell'attività genica: a) DNA genico e ridondante; b) geni attivi e repressi; c) differenziamento cellulare; embriologia e genetica; d) le basi molecolari della mutazione genica. 6. L'evoluzione. 7. Vitalismo e meccanicismo. Finalismo e causalità. Biologia molecolare e organismale. 8. La biologia e l'uomo: a) le scienze biologiche e la concezione del mondo; b) è possibile un'‛etica evoluzionistica'? □ Bibliografia.
1. Introduzione
Nel Dizionario Enciclopedico Italiano la voce biologia si apre con la seguente definizione: composto di bio- e -logia; in senso ampio e più comunemente accettato, l'insieme delle scienze della vita [...]". La biologia studia dunque le proprietà strutturali e funzionali di tutti gli organismi, virus piante e animali, compreso l'uomo, chiaramente abbracciando altre discipline quali l'anatomia, la fisiologia e l'antropologia fisica; non vi sono comprese, invece, le scienze filosofiche, giuridiche e storiche, le discipline, cioè, che dell'uomo studiano l'attività spirituale e che si usano designare come ‛scienze morali', in contrapposizione con le scienze fisiche e naturali, tra le quali la biologia stessa trova posto sicuro e appropriato. L'attributo ‛morale' mette in immediata evidenza come l'uomo, che i dati paleontologici dimostrano oggetto della Natura e prodotto da un'evoluzione determinata dagli stessi fattori che hanno agito sui corsi evolutivi di tutti gli altri organismi, l'Homo sapiens è ormai, rispetto ai bruti, tutt'altra cosa, in quanto è giunto, grazie al peculiare sviluppo del suo sistema nervoso, all'autocoscienza e alla previsione dei risultati della sua attività, quindi alla scelta dei mezzi in vista dei fini. Per questa scelta consapevole, caratterizzata sempre da un germe di libertà anche se caso per caso storicamente determinata, nelle attività spirituali dell'uomo il finalismo è riconoscibile come essenziale realtà. Al contrario, il finalismo, che è altrettanto evidente in tutti gli organismi e nelle loro parti (gli occhi per vedere, i denti per masticare, ecc.), si rivela alla fine come frutto di un'evoluzione guidata da fattori ambientali, l'una e gli altri indagabili col moderno metodo delle scienze naturali, che esclude, in biologia come in fisica, la possibilità di una spiegazione in termini finalistici.
La separazione tra scienze fisico-chimiche e biologiche da una parte, scienze morali dall'altra, sembra essenziale, ma in pratica esistono campi di ricerca nei quali i confini diventano vaghi, come negli studi del comportamento e nelle scienze sociali. In questo articolo considereremo di pertinenza della biologia umana lo studio dell'antropologia fisica, tralasciando completamente, con un'esclusione forse eccessiva, le scienze sociali. Tuttavia, tale limitazione è comprensibile quando si pensi all'enorme importanza che per lo sviluppo della personalità dei singoli rappresenta l'attiva assimilazione dell'eredità culturale, e quando si consideri quanto diversi siano gli uomini dagli altri Primati e dagli Insetti ‛sociali', sicché i legami della convivenza umana risultano necessariamente differenti da quelli che regolano la vita delle Api e dei Cinocefali. Tali differenze sono certamente significative, anche se non v'è dubbio che il comportamento sociale dell'uomo è condizionato da determinate attitudini - per il bene o per il male, quali la collaborazione di gruppo o l'aggressività - frutto di una selezione che ha agito su un fondo ancestrale non soltanto culturale, ma anche ereditario, in senso fisico, genetico, non troppo remoto, addirittura recentissimo, secondo la cronologia paleontologica.
Nelle scienze biologiche, come in quelle matematiche, fisiche e chimiche, la ricerca di base ha trovato ampie applicazioni pratiche, in agricoltura, zootecnia, idrobiologia, ecc.; e come biologia applicata può essere considerata anche la medicina. Tutte queste discipline saranno qui quasi interamente trascurate, per la necessità di limitare un campo già molto vasto. La nostra attenzione sarà rivolta soprattutto ai progressi di una ‛biologia generale', che studia i fenomeni comuni alla totalità degli organismi e che nel sec. XX ha compiuto enormi progressi. Questi, naturalmente, furono preparati dal lungo lavoro precedente, ma hanno proceduto con velocità sempre crescente, nell'ultimo ventennio addirittura impetuosa.
Grandissimi sono stati i progressi delle scienze fisiche nella prima metà del nostro secolo (la teoria della relatività, le leggi della meccanica quantistica, l'analisi della struttura particellare del nucleo atomico), che si rivelarono improvvisamente all'uomo della strada con le esplosioni atomiche che conclusero la seconda guerra mondiale. Ma questi progressi sono stati almeno eguagliati da quelli compiuti dalla biologia negli ultimi trenta-quaranta anni, anche se, per non aver avuto fortunatamente tragiche applicazioni, si sono meno imposti all'attenzione generale.
I biologi della mia età, di circa settant'anni, hanno avuto il privilegio di vivere in un periodo di eccezionale interesse, paragonabile forse soltanto al ventennio 1860-1880 che seguì alla pubblicazione dell'Origine delle specie del Darwin. E vorrei dire che il valore dei recenti progressi della biologia, per determinare la concezione del mondo in coloro che li hanno seguiti almeno nelle loro linee generali e che ne hanno compreso il significato essenziale, è superiore a quello dei progressi delle scienze fisiche: ciò è immediatamente comprensibile, quando si pensi che ogni uomo si sente vivere, e in effetti vive ‛fisicamente', all'incirca come un batterio, e in modo pressoché simile a quello di ogni altro mammifero.
Progressi fondamentali sono stati compiuti dunque in numerose discipline ormai largamente autonome e necessariamente specialistiche. Eppure, proprio in questi ultimi anni, quando la specializzazione sembrava eccessiva, tale da rendere sempre più difficile, e alla fine impossibile, una visione unitaria dei fenomeni della vita, proprio in questi ultimi anni si è arrivati a una sintesi che lascia vedere o intravedere enormi complessità, ma che mette anche in evidenza un disegno perspicuo.
I progressi nelle varie discipline biologiche saranno illustrati nelle voci dedicate a ciascuna di esse e vi faremo continui richiami. Qui cercheremo di mettere in rapporto le acquisizioni compiute nei diversi campi e che ormai si vedono integrarsi, andare a posto come le tessere di un mosaico non ancora finito, ma del quale già si può ammirare l'armoniosa composizione. Questo mosaico non sarà mai completo né definitivo: alcune sue parti forse dovranno essere demolite e rifatte su nuovi disegni, perché in biologia, come in ogni altro ramo del sapere, la soluzione di un problema ne pone altri, in una continua sequenza di cui non è possibile prevedere la fine.
È tuttavia vero che in biologia, come in certi campi della fisica, alcuni approdi recenti sembrano definitivi. Così, definitivo sembra il rigetto di ogni forma di vitalismo che postuli, per poter arrivare alla spiegazione dei fenomeni della vita, l'esistenza di forze e di agenti totalmente estranei al mondo dell'inorganico studiato dalla fisica e dalla chimica... Ma naturalmente ci sono e ci saranno ancora dei vitalisti, come ci sono e ci saranno ancora degli astrologi dopo Galileo, Copernico e Newton.
2. Caratteri essenziali degli organismi viventi
È facile distinguere un organismo da un oggetto del più vasto mondo inorganico: nessuno in genere confonde una pianta o un animale con una pietra e basta avere nozioni minime di botanica per non scambiare un'alga calcarea per un pezzo di scoglio. Il riconoscimento dunque è immediato e sembra intuitivo; ma è difficile, per chi non vi abbia mai pensato, individuare qualche caratteristica, e possibilmente quella essenziale, che differenzi i viventi dagli oggetti, per usare un vecchio aggettivo, inanimati.
Si può dire essenziale degli organismi la capacità di autosintesi, cioè di riprodursi, e di riprodursi uguali a se stessi. Si potrebbe tuttavia obiettare che qualcosa di simile avviene in alcuni fenomeni di cristallizzazione: alcune sostanze, infatti, possono restare liquide, in un cosiddetto equilibrio metastabile, in soluzione soprasatura o al di sotto del loro punto di fusione, e cristallizzare rapidamente, passare cioè a un equilibrio stabile, quando vi arriva un loro germe cristallino. Ma certo non si può confondere una bottiglia di glicerina in frigorifero con una coltura di microrganismi: l'insieme di acidi nucleici e proteine, che è alla base dei fenomeni di riproduzione degli organismi, raggiunge una complessità di gran lunga superiore a quella di una mera struttura cristallina.
Ancora, il concetto di vita è oggi legato all'idea della struttura cellulare: sappiamo infatti che quasi tutti gli organismi sono composti da una o più cellule e che i virus, che soli non hanno struttura cellulare, vivono obbligatoriamente in simbiosi parassitaria con cellule viventi. Ma forse la struttura cellulare non deve essere considerata come carattere peculiare degli organismi: una cellula, infatti, è una struttura così complessa che non è più pensabile - come lo era stata fino ai tempi di R. Virchow e di L. Pasteur - la sua formazione da un substrato non vivo, per generazione spontanea. Conseguentemente, o si postula la comparsa della vita già con la complessità cellulare, per atto sovrannaturale, o si ammette che il raggiungimento ditale livello organizzativo sia il risultato di un lungo periodo evolutivo, a partire da organismi più semplici, all'inizio da composti organici discretamente complessi ma formatisi per fenomeni naturali e già capaci di autosintesi. Quindi le forme di vita, o meglio gli elementi primordiali della vita, precedenti la comparsa della struttura cellulare, esistiti probabilmente sulla Terra in tempi remoti, possono essere considerati come veri organismi viventi, in quanto stati ancestrali agli attuali; e possono forse ancora esistere in altri mondi giovani, come lo era il nostro pianeta due o tre miliardi di anni fa (v. vita, origine della).
Abbiamo già accennato a un'altra caratteristica evidente in tutti gli organismi, il loro finalismo. Essi sono così e così fatti per vivere almeno fino a riprodursi e ciascuna parte dell'organismo ha la sua funzione, il suo fine parziale: al livello molecolare proteico, negli enzimi si riconoscono uno o più ‛siti attivi' destinati ad accogliere gruppi sterici di altre molecole sulle quali devono agire. Tutti avvertono questo finalismo che pervade ogni fenomeno della vita, eppure noi del mestiere, nelle lezioni introduttive o nelle pagine di un testo di una qualsiasi disciplina biologica, il più delle volte facciamo le viste di ignorarlo: in realtà, tale comportamento risponde all'esigenza didattica di celare l'evidente contraddizione tra quella che è una caratteristica degli oggetti del nostro studio e il metodo scientifico, che vogliamo impiegare come l'unico in grado di consentirne un utile studio e che è dominato dal principio di causalità. E d'altra parte è un ingenuo, onesto trucco didattico, in quanto ora sappiamo che la contraddizione è superata, solo che si guardino gli organismi non all'istante, ma come punti di arrivo di una lunga storia evolutiva. J. Monod ha avuto non diciamo il coraggio, ma la franchezza di dichiarare, fin dall'introduzione del suo saggio su Le hasard et la nécessité (1970), che gli organismi sono caratterizzati dalla possibilità di riprodursi uguali a se stessi (proprietà dell'invarianza) e dall'avere, strutture e funzioni, a qualsiasi livello studiate, in vista di un fine: egli ha coniato, per quest'ultima proprietà dei viventi, il termine di ‛teleonomia'. Si potrebbe allora ritenere possibile, se si definiscono gli organismi come invarianti e teleonomici, la costruzione di una macchina, di un superautoma capace di costruire copie di se stesso, e attribuire in tal caso al suo progettatore il merito di aver creato una nuova forma di vita, in plastica e metallo, con nastri magnetici invece di DNA, ecc.; ma a questo punto si arriverebbe veramente alla pedanteria.
Diamo dunque un breve elenco delle principali caratteristiche dei viventi: fondamentale identità ed esclusività a livello chimico dei cosiddetti componenti immediati di tutti gli organismi, in particolare delle proteine e degli acidi nucleici; struttura cellulare, almeno per gli organismi attuali; fenomeni di riproduzione, legati alla proprietà di autosintesi caratteristica di tutti gli organismi, di differenziamento, di accrescimento, di senescenza. E qui possiamo notare che l'invecchiamento è ineluttabile negli organismi pluricellulari, nei quali la morte sembra quasi un attributo essenziale della vita, donde la famosa definizione nell'Enciclopédie di Diderot e d'Alembert, ‟la vita è il contrario della morte", e quella di X. Bichat, ‟la vita è l'insieme delle forze che contrastano la morte"; ma la morte come fenomeno naturale è la conseguenza del differenziamento imposto dalla divisione del lavoro tra i vari tipi cellulari, molti dei quali perdono la capacità di moltiplicazione, si usurano e non sono più sostituibili. Invece negli unicellulari, in continua moltiplicazione, non compaiono mai, se l'ambiente si mantiene costante e favorevole, fenomeni di invecchiamento e morte naturale. Ancora, un'altra caratteristica è l'ereditarietà, cioè fondamentale invarianza, alla quale solo apparentemente contraddice la lunga incessante modifica del patrimonio ereditario, causa di una continua evoluzione creatrice di nuove forme, di una storia della vita svoltasi per un lunghissimo arco di tempo; continuo rapporto di congruenza con l'ambiente; continuo ricambio, per cui gli organismi devono intendersi come sistemi aperti, attraversati da un incessante flusso di materia e di energia attinte senza sosta dall'ambiente, modificate e ad esso restituite. E si può aggiungere la distribuzione dei viventi in gruppi discontinui: meglio definite le specie, raggruppabili in unità sistematiche maggiori in ordine gerarchico, generi, famiglie, ordini, ecc. L'esistenza di gruppi separati è così universale, che si può considerare una caratteristica fondamentale della diversità degli organismi" (Th. Dobzhansky, Genetics and the origin of species, New York 19412, p. 5).
I vari attributi degli organismi sono studiati in diversi capitoli della biologia: biofisica e biochimica; ricambio materiale ed energetico; ecologia; citologia; genetica; evoluzione; riproduzione; embriologia; accrescimento e senescenza; sistematica. Ormai si deve parlare più che di capitoli, addirittura di discipline diverse, e i nostri accenni saranno sempre succinti e numerosi i richiami alle voci particolari. Più ancora, per esigenza di spazio, parecchi aspetti dei fenomeni biologici saranno addirittura taciuti.
3. Le proteine: struttura e funzione come due aspetti dei fenomeni di biologia molecolare
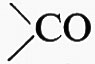
Le proteine erano conosciute già nella prima metà dell'Ottocento come costituenti essenziali di tutti gli organismi e il nome, dovuto a G. J. Mulder (1839), ne indica l'importanza primaria. Per lungo tempo il loro studio progredì lentamente, perché non si riusciva a separarle e purificarle; solo alla fine del secolo A. Kossel dimostrò che ogni molecola proteica deve intendersi come un polimero di amminoacidi e riconobbe il legame peptidico -CO-NH- col quale il gruppo carbossilico di un amminoacido si lega al gruppo amminico del successivo, con la perdita di una molecola di acqua. L'analisi totale di varie specie di proteine, fino ai singoli amminoacidi e al loro elenco pressoché completo, è dovuta al lungo lavoro compiuto nel primo quarto del secolo da E. Fischer, che iniziò anche i tentativi di sintesi, arrivando a catene peptidiche di 18 anelli. Nel grande trattato di biochimica edito da C. Oppenheimer (1924) è riportato un elenco di 20 amminoacidi; il loro numero non è stato, in seguito, praticamente aumentato. Negli anni trenta T. Svedberg applicò il metodo della ultracentrifugazione e dalla velocità di sedimentazione delle diverse proteine calcolò il loro peso molecolare, che ne confermava la complessità, con valori generalmente compresi tra 20.000 e 200.000. Contemporaneamente lo studio dell'orientamento delle molecole in una soluzione in rapido flusso e le immagini di birifrangenza che ne potevano derivare rivelarono l'esistenza di due tipi di proteine, rispettivamente a molecola globulare e allungata; e si vide che le prime sono spesso quelle a funzione enzimatica, mentre le seconde si ritrovano nelle strutture fibrose, quali le fibre di collagene dei connettivi, la cheratina dei peli, una parte delle fibrille contrattili muscolari. Si cominciò così a parlare di proteine enzimatiche e di proteine strutturali e della possibilità che in seno al citoplasma anche il più apparentemente ialino, anisto, esista una trama di molecole filamentose, un ‛citoscheletro', costituente una struttura intima, da cui derivi la polarità di tante cellule. Oggi del citoscheletro si parla meno, perché non si riesce a vedere, nemmeno al microscopio elettronico, questa trama molecolare così esile da venir alterata dalle manipolazioni necessarie per ottenere le sezioni ultrasottili da sottoporre all'osservazione. Eppure ancor oggi non esiste altra spiegazione per tanti fenomeni nei quali si deve ammettere un ordine intimo, per esempio quando un uovo sferico e a citoplasma pressoché ialino si vede dividersi in più cellule secondo un disegno preciso, senza il quale non sarebbe comprensibile il successivo differenziamento dei vari organi.
Negli anni quaranta la tecnica di analisi delle proteine faceva nuovi progressi, coi metodi dell'elettroforesi e della cromatografia, per separare le molecole più o meno mobili delle varie specie proteiche o i loro frammenti ottenuti per digestione parziale o totale. Progredì contemporaneamente lo studio della struttura delle macromolecole, con l'impiego dei metodi cristallografici, soprattutto con la diffrattometria a raggi X. E oggi si è ottenuta l'analisi cristallografica completa della cheratina della lana e di parecchie proteine globulari, fino alla risoluzione di pochi ångström (A), cioè di pochi decimilionesimi di millimetro.
La struttura fondamentale delle molecole proteiche è dunque a catene polipeptidiche, per lo stabilirsi del legame peptidico tra amminoacidi successivi e disposti in serie lineare, senza ramificazioni. Si conoscono una ventina di amminoacidi, che differiscono per il gruppo R (radicale) legato al cosiddetto ‛carbonio alfa'. Gli R sono più o meno complessi: ci limiteremo a dire che possono essere apolari, polari neutri o polari con carica elettrica. I gruppi apolari sono idrofobi e si ritrovano nella glicina (Gly), alanina (Ala), valina (Val), leucina (Leu), isoleucina (Ile), prolina (Pro), fenilalanina (Phe), metionina (Met). Sono idrofili invece i gruppi polari e in soluzione alcuni possono essere ionizzati, a carica negativa o positiva, quindi a funzione acida o basica: hanno un gruppo del primo tipo l'acido aspartico (Asp) e l'acido glutammico (Glu), del secondo, la lisina (Lys), l'arginina (Arg) e l'istidina (His). Hanno gruppi polari neutri o quasi neutri asparagina (Asn), glutammina (Gin), senna (Ser), treonina (Tht), cisteina (Cys). Infine, il triptofano (Trp) è così debolmente idrofilo da potersi collocare tra gli apolari e la tirosina (Tyr), con un idrossile legato a un anello aromatico, è idrofila e con una debolissima funzione acida. Ricordiamo ancora che la metionina è la cisteina contengono nel loro R un atomo di zolfo ed è per questo che quasi tutte le proteine contengono, oltre a C, H, O, N, una piccola percentuale di S, in media un 1% circa. Lo zolfo impartisce una particolarità chimica alla cisteina, dove il gruppo R ha la struttura -CH2-SH: quando due Cys sono ravvicinate, per appartenere a due catene peptidiche che si affrontano, o alla stessa catena ripiegata, si può stabilire per diedrogenazione un cosiddetto ponte di zolfo -CH2-S-S-CH2- piuttosto solido, così da costituire un punto fermo della struttura molecolare.
La composizione in amminoacidi, e più precisamente il loro numero, la loro specie e la loro esatta sequenza, costituisce la cosiddetta ‛struttura primaria' (str. I) della molecola proteica. Una data memorabile della biochimica è stata segnata da F. Sanger, che nel 1953 arrivò all'analisi completa delle sequenze di amminoacidi dell'insulina bovina, una proteina relativamente semplice, di peso molecolare 5.733, con due brevi catene di 21 e 30 amminoacidi, tenute insieme da due ponti S−S. La tecnica di Sanger fu subito estesa al riconoscimento della struttura primaria di altre proteine più complesse; oggi se ne conoscono, sotto questo riguardo, alcune centinaia e tale analisi strutturale è diventata per alcuni laboratori una ricerca quasi abituale.
Raramente le catene peptidiche restano distese: esse hanno tendenza ad accorciarsi, spiralizzandosi per una rotazione delle valenze dei carboni alfa. Una rotazione utile per stabilizzare la catena è quella che allontana il carbonile
dal gruppo R; se l'angolo di rotazione è sempre uguale, la catena si avvolge a elica e si dice che assume una struttura secondaria. L. Pauling e R. B. Corey proposero nel 1951 il modello della ‛alfa elica' come struttura secondaria più stabile e quindi più probabile, perché tra gruppi
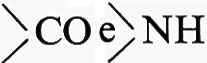
si stabiliscono molti legami a idrogeno paralleli o quasi all'asse dell'elica. M. F. Perutz con la cristallografia a raggi X confermò subito l'esistenza di un'elicatura di tipo alfa, estesa a tratti più o meno lunghi di ogni catena peptidica, a seconda della specie proteica. Una struttura terziaria è data dall'ulteriore ripiegamento di una catena già più o meno elicata e talvolta dall'unione di poche catene diverse tra loro e tenute insieme da legami piuttosto solidi, generalmente da ponti S−S. Infine, la molecola di alcune proteine si comporta in condizioni fisiologiche come una struttura unitaria, ma di fronte a trattamenti piuttosto blandi si rivela come un aggregato di più subunità tenute insieme da legami deboli: si parla allora di struttura quaternaria.
Una ripiegatura delle catene peptidiche fu ipotizzata appena si riconobbe l'esistenza di proteine globulari; e oggi che la cristallografia ha precisato questa struttura terziaria, si è compreso che la configurazione delle proteine non è determinata dall'esterno, ma è assunta spontaneamente come la più stabile configurazione per una data sequenza di amminoacidi" (v. Perutz e altri, 1965, p. 669). Ad esempio, alcuni amminoacidi con appropriati gruppi R possono, diciamo così, cercarsi per stabilire un legame a idrogeno e iniziare l'assunzione della struttura terziaria; o due cisteine anche lontane nella stessa catena, o in due catene peptidiche nell'ambito della stessa molecola, possono affrontarsi e dare un ponte S−S che mette un punto fermo in questa messa in piega. Ancora, molti amminoacidi con catena laterale idrofoba tendono a far gruppo, a sfuggire all'acqua circostante, piegando la catena in modo da portarsi nell'interno della molecola, a costruire una regione tagliata fuori dall'acqua; al contrario, gli amminoacidi con gruppo R idrofilo restano in superficie e danno alla molecola complessiva la sua idrofilia. La regola ‛gruppi apolari dentro e polari fuori' presenta poche eccezioni.
Una diecina di anni fa si è arrivati, con la diffrattometria a raggi X, a riconoscere la completa struttura terziaria di molecole proteiche molto complesse. J. C. Kendrew per primo (1958) studiò così la mioglobina, una proteina immagazzinatrice di ossigeno molecolare, contenuta nelle fibre muscolari dei Vertebrati; essa è composta da una catena di 153 amminoacidi, in buona parte ad alfa elica, poi ripiegata così da assumere una forma ellissoidale di circa 44 × 44 × 25 Å e da porre in vicinanza, con la precisione di frazioni di Å, determinati gruppi R che possono così interagire e assicurare la funzione della molecola, che è di accogliere in una tasca interna un gruppo ‛eme' con un atomo di ferro, che possa legare una molecola di O2. La tasca è formata da una maggioranza di amminoacidi a gruppi idrofobi, per tagliare fuori dall'acqua l'eme, in modo che il suo Fe ferroso non possa ossidarsi a ferrico, incapace di legare l'O2. Due tratti della catena si fanno fronte, costituiscono due pareti della tasca interna e portano ciascuno un'istidina alla quale è coordinato il ferro dell'eme da una parte direttamente, dall'altra con l'intermediario dell'O2.
Nel 1960-1963 Perutz e collaboratori arrivarono a delucidare la completa struttura dell'emoglobina (Hb) di cavallo. L'Hb, contenuta nei globuli rossi del sangue dei Vertebrati, è una trasportatrice di O2; la sua molecola è formata da quattro catene polipeptidiche, ciascuna con la sua tasca idrofoba che accoglie un eme, e uguali a coppie, una coppia di catene alfa, con 141 amminoacidi, e una coppia di beta, con 146 amminoacidi; in tutto una molecola con 574 amminoacidi, di peso molecolare (p. mol.) = 64.500, di forma all'incirca sferoidale, di 64 × 55 × 50 Å. Le quattro catene sono unite con legami non molto solidi (forze di van der Waals, legami a idrogeno), ma capaci d'impartire alla molecola complessiva la sua struttura quaternaria, pur lasciando un gioco di qualche Å nei rapporti di posizione delle quattro catene, a seconda che i loro gruppi eme portino l'O2 o ne siano privi. Le sequenze degli amminoacidi presentano nelle diverse forme di Vertebrati molte corrispondenze ma anche parecchie differenze, che peraltro non modificano sensibilmente la struttura terziaria che resta sempre molto simile non solo tra le catene alfa e beta, ma anche tra queste e l'unica catena della mioglobina. Ciò dipende dal fatto che caratteristica comune è l'esistenza in ogni catena di una trentina di amminoacidi che possono essere diversi, ma appartengono tutti al gruppo degli idrofobi, e mantengono la stessa posizione interna. Gli amminoacidi idrofobi profondi sono dunque largamente intercambiabili e lo stesso si può dire per gli idrofili superficiali; difficili sono invece le sostituzioni tra amminoacidi idrofili e idrofobi, e in certi casi anche una sola sostituzione può avere effetti importanti e danneggiare molto la stabilità e la funzionalità della molecola. Alcuni amminoacidi sono infatti ‛in posizione strategica': risultano così insostituibili le due istidine che coordinano l'eme e mal sostituibili altri amminoacidi nelle loro vicinanze. Su questo argomento torneremo a proposito delle mutazioni geniche, che si traducono in modifiche della struttura primaria e quindi automaticamente della struttura terziaria delle proteine specificate dai geni mutati.
Ormai la struttura terziaria di parecchie altre proteine globulari e di alcune fibrose è conosciuta altrettanto bene come quella della Hb. Ci limiteremo a un cenno sulla struttura degli enzimi. Tutti gli enzimi sono proteici, anche se vi sono associati, come cofattori, metalbioni e molecole organiche quali le vitamine; più precisamente, sono proteine globulari. Sono catalizzatori chimici, servono cioè ad aumentare la velocità di reazioni che avrebbero identico decorso anche senza la loro presenza, ma che, alle temperature compatibili con la vita, procederebbero con estrema lentezza (v. enzimi). Gli enzimi sono strettamente specifici, ciascuno catalizza una particolare reazione, e sono di moltissime specie, perché gli organismi sono macchine chimiche molto complicate. Si calcola che l'Escherichia coli, un batterio, cioè un organismo tra i più semplici, abbia un corredo enzimatico di oltre 1.500 specie. La specificità di azione delle molecole enzimatiche è tale che una piccola modifica della loro struttura terziaria può impedire di accogliere il substrato, cioè la molecola o le molecole sulle quali devono agire. E. Fischer già nel 1894, con mirabile intuizione, aveva prospettato l'analogia ‛della chiave e della serratura', rappresentando la superficie dell'enzima provvista di un'incisura per accogliere il substrato. E oggi per mezzo della cristallografia siamo arrivati a costruire i precisi modelli di molecole enzimatiche e a mettere in evidenza un incavo più o meno profondo, il cosiddetto sito attivo, dove sporgono i gruppi R di particolari amminoacidi, essenziali per ricevere e legare il substrato e per catalizzare le reazioni che lo devono modificare. Esemplificheremo con la molecola del lisozima, studiata nel 1967 da D. C. Phillips e collaboratori, un enzima battericida in quanto in grado di provocare la lisi dei mucopolisaccaridi presenti nella capsula cellulare di molti batteri. I mucopolisaccaridi sono polimeri di zuccheri a sei atomi di C, che portano gruppi laterali azotati; la loro lisi avviene per la rottura dei ponti −O− che uniscono gli zuccheri adiacenti. La molecola del lisozima - formata da un'unica catena polipeptidica di 120 amminoacidi, più volte ripiegata a dare una struttura ovoidale - mostra un'incisura che rappresenta il sito attivo, dove affiorano i gruppi R di una decina di amminoacidi in posizione adatta ad accogliere e tenere fino a sei anelli di zucchero. Due amminoacidi, in particolare un acido glutammico e un acido aspartico in posizione 35 e 52, catalizzano il processo - non tanto semplice da poter essere qui descritto - di rottura dei ponti di ossigeno.
L'emoglobina ci ha fornito un esempio di struttura quaternaria, con quattro ‛protomeri' in due coppie subeguali; in tutti i casi di struttura quaternaria i protomeri sono identici o subeguali. In alcuni casi la polimeria può essere molto elevata: tipico esempio le emocianine di molti Molluschi e Artropodi, trasportatrici di ossigeno, col rame al posto del ferro, e con molecole gigantesche (p. mol. di alcuni milioni), formate da centinaia di protomeri, con una disposizione di significato funzionale per ora mal chiarito. Più frequente, soprattutto negli enzimi, una oligomeria: ad esempio, la gliceraldeide-3-fosfatodeidrogenasi ha p. mol. =144.000 ed è composta da 4 protomeri di p. mol. =36.000. Negli enzimi la oligomeria sembra essere legata alla necessità di arrivare ad una struttura particolarmente complessa, capace di prendere rapporti con due molecole di diverso tipo. Tali sono le ‛molecole allosteriche' di J. Monod, J. P. Changeux e F. Jacob (1963), che possiedono due siti attivi, a uno dei quali è legata la funzione enzimatica specifica, mentre l'altro è capace di accogliere un'altra molecola che ne modula l'attività, perché due siti attivi sono tali che quando uno è occupato l'altro si deforma. Una proteina allosterica potrebbe arrivare alla sua complessità anche se formata da una sola catena polipeptidica; ma ci vorrebbe allora una catena molto lunga e questo sembra difficilmente realizzabile, probabilmente perché vi è un limite all'acquisto automatico di una precisa struttura terziaria: e infatti le singole catene polipeptidiche superano di rado i 200 amminoacidi. La modulazione dell'attività enzimatica da parte di un'altra molecola è fenomeno frequente. Molte volte un enzima che catalizza la produzione di una certa sostanza viene inibito quando nel medium si forma un eccesso del prodotto della sua attività ed è questo uno dei tanti fenomeni di retrocontrollo che regolano ad ogni livello l'attività degli organismi.
Questa introduzione allo studio della struttura proteica vuol mettere in risalto tre punti: primo, quando l'analisi di un fenomeno biologico si fa stringente, scompaiono i pretesti per invocare vitalismi ignoti e indefinibili; secondo, a livello molecolare diviene evidente che struttura e funzione non sono che due aspetti per noi di una medesima realtà; terzo, anche a questo livello si ritrova la precisa aderenza alle leggi fisico-chimiche e la teleonomia.
4. Un gene, una proteina
a) Cromatina, cromosomi e geni
Verso la metà del secolo scorso era stata riconosciuta l'esistenza nel nucleo cellulare di masserelle di una sostanza, che fu chiamata ‛cromatina' perché facilmente tingibile coi colori che si cominciavano a impiegare nella tecnica microscopica. Negli anni ottanta furono ben descritte le modificazioni di questa cromatina durante la divisione cellulare, che si disse avvenire per ‛cariocinesi' o per ‛mitosi', cioè per movimenti nucleari o per formazione di filamenti. Si vide infatti che all'inizio della mitosi la cromatina aumenta e si distribuisce ordinatamente a delineare delle strutture filamentose, i cromosomi; questi si scindono poi longitudinalmente e i due elementi della coppia migrano ai poli opposti della cellula, ove formano due nuclei con corredi cromosomici identici a quello del nucleo da cui hanno preso origine. Successivamente, anche il citoplasma si divide, così che si originano due cellule, ciascuna delle quali potrà scindersi con una nuova mitosi: in tal modo il corredo cromosomico si perpetua invariato. Un meccanismo così complesso doveva avere un significato funzionale preciso e importante, tanto più che i cromosomi si rivelarono costanti e caratteristici per numero e per forma in ogni specie di piante e animali. Si vide inoltre che negli organismi a riproduzione sessuale, che sono la stragrande maggioranza, ogni corredo cromosomico è formato da un certo numero di coppie tra loro uguali (cromosomi ‛omologhi') e che a un dato momento del ciclo riproduttivo molto spesso subito dopo la formazione dei gameti, uova e spermatozoi - ha luogo un processo nucleare alquanto diverso, la ‛meiosi', in seguito al quale il numero dei cromosomi si dimezza, ogni nucleo figlio ricevendo uno dei due omologhi: così, ad esempio, tutte le cellule del nostro organismo hanno 46 cromosomi in 23 coppie e alla spermatogenesi e all'ovogenesi si formano per meiosi spermi e uova a 23 cromosomi; nella fecondazione si uniscono una serie paterna e una materna e il nuovo organismo inizia il suo sviluppo con 46 cromosomi. Si indicano le due condizioni coi termini di aploidia e diploidia: i gameti hanno sempre corredo aploide e nel caso dell'uomo si scriverà n = 23; l'uovo fecondato e tutte le cellule che ne derivano per mitosi sono diploidi, 2n = 46.
Proprio alla fine del secolo, nel 1900, tre botanici, C. Correns, H. de Vries ed E. von Tschermak riscoprirono, contemporaneamente e indipendentemente, le ‛leggi di Mendel', le regole statisticamente valide di trasmissione ereditaria riconosciute 35 anni prima da Gregor Mendel, abate del convento agostiniano di Brno, nella regione ceca dell'allora impero austro-ungarico. Le ricerche, che non è enfatico definire geniali, pubblicate nel 1866, erano rimaste ignorate, perché avevano precorso di troppo i tempi e la scienza ufficiale non era preparata a comprendere e valutare l'opera di questo dilettante. Lo studio dell'eredità ebbe da tale riscoperta, della quale fu compreso finalmente il valore euristico, un impulso decisivo e si affermò, come nuovo capitolo della biologia, la genetica. Tale disciplina registrò nel primo quarto di secolo progressi di estrema importanza, ai quali è legato il nome di Thomas H. Morgan, uno dei maggiori biologi del nostro secolo, che ebbe tra gli altri il merito di circondarsi di eccellenti collaboratori, cui era lasciata ampia iniziativa, e di condurre le ricerche su un moscerino, la Drosophila melanogaster, di facile e rapido allevamento. Sul lavoro di questa équipe ebbe probabilmente influenza anche il fatto che le ricerche furono condotte per molti anni alla Columbia University di New York, dove operava un citologo di grande valore, Edmund B. Wilson: ben presto infatti apparve evidente che le leggi di Mendel e le loro estensioni (caratteri legati al sesso, gruppi di associazione, crossing over, ecc.; v. genetica; v. gene) dimostravano che il materiale ereditario aveva struttura particellare, era costituito da ‛geni' situati nei cromosomi, un determinato gruppo di geni con un preciso ordine lineare in ciascun cromosoma della serie aploide e quindi ogni gene in due esemplari allo stadio diploide. Un libro che segnò una data importante, la Teoria del gene di Th. H. Morgan, esponeva i risultati ai quali era arrivata nel 1926 la genetica e le conseguenti interpretazioni, che ormai erano acquisizioni definitive.
b) Gli acidi nucleici e il valore genico del DNA
Intanto, di pari passo con gli studi sulla morfologia del nucleo, e poi sull'eredità, procedevano le ricerche chimiche sul materiale nucleare. Nel 1874 F. Miescher iniziò il lavoro su masse di cellule con poco citoplasma (lo sperma di salmone, i leucociti del pus settico) e riuscì a separare, come egli scrisse, ‟una nucleina del tutto priva di albumine", ricca di fosforo. Qualche anno dopo R. Altmann precisò l'esistenza di un acido organico con l'8-9% di fosforo e lo chiamò acido nucleico. Fondamentale fu il lungo lavoro, nell'ultimo ventennio del secolo, di A. Kossel che dalle nucleine ottenne fosforo, zuccheri e cinque specie di basi azotate, che riconobbe come basi puriniche (adenina, A; guanina, G) e pirimidiniche (citosina, C; timina, T; uracile, U). Le ricerche furono poi estese al timo, ricco di linfociti, che sono cellule con poco citoplasma, ad alcuni parenchimi ghiandolari, quali il pancreas, e al lievito, cellule tutte con molto citoplasma, ma che si rivelarono ricche di acido nucleico. Verso il 1910 era ormai accertato che gli acidi nucleici sono costituiti da ‛nucleotidi', ognuno dei quali formato da una della cinque basi azotate, A,G,C,T,U, legata a uno zucchero, a sua volta esterificato da acido ortofosforico, H3PO4. Si riconobbe altresì che negli acidi nucleici estratti da tessuto ghiandolare o dal lievito lo zucchero è un pentoso, il d-ribosio di formula C5H10O5. Ma dal timo e dagli spermi si estraeva un acido timonucleico, nel quale si ammetteva la presenza di un composto vicino agli zuccheri, che E. Fischer nel 1914 chiamò ‛glucal' e di cui era data la formula C5H10O4. Ancora nel 1924 questi erano i dati fondamentali riportati nel trattato di biochimica dell'Oppenheimer. Soltanto negli anni trenta la chimica degli acidi nucleici arrivò a un assetto che si può dire, nelle sue grandi linee, definitivo. Si riconobbe che tutti gli acidi nucleici sono polimeri di mononucleotidi disposti in catene lineari più o meno lunghe; che in ogni mononucleotide l'acido fosforico si lega al carbonio in posizione 5' dello zucchero, il cui C in 1' si unisce a sua volta a una delle basi azotate; che nella catena polinucleotidica un'altra valenza del P esterifica il C in posizione 3' dello zucchero del nucleotide successivo. Infine, si stabilì che vi sQno due specie di acidi nucleici: l'acido ribonucleico (RNA), nel quale lo zucchero è il ribosio e le basi azotate sono le due purine adenina e guanina, e le due pirimidine citosina e uracile; e l'acido desossiribonucleico (DNA), nel quale il ribosio è sostituito dal desossiribosio C5H10O4, e delle quattro basi azotate tre sono le stesse presenti nell'RNA, A,G e C, la quarta è invece la timina che sostituisce l'uracile. I biochimici sanno ormai estrarre gli acidi nucleici dagli omogenati cellulari e separare il DNA dall'RNA senza danneggiarne la struttura. Qui basterà ricordare che alcuni facili metodi di colorazione permettono di distinguere nei preparati microscopici le due specie di acidi nucleici; e che l'RNA si ritrova nel nucleo e soprattutto nel citoplasma, mentre il DNA è pressoché esclusivo del nucleo e più precisamente dei cromosomi.
Per parecchi anni, quando la struttura chimica degli acidi nucleici era ormai chiarita, la loro funzione restava enigmatica. In particolare, non si riteneva che fossero di DNA i geni, che pur si sapevano portati dai cromosomi: ciò era dovuto in parte alla scarsa attendibilità dell'ipotesi che l'acido desossiribonucleico con le sue quattro basi azotate potesse avere la molteplice specificità delle proteine coi loro 20 amminoacidi, ma soprattutto al fatto che il DNA si vedeva comparire abbondante all'inizio delle cariocinesi e diminuire al termine della divisione, fin quasi a scomparire durante l'intercinesi. Si pensava quindi al DNA come a una sostanza poco importante, che incrostasse i cromosomi in cinesi per renderli più compatti e per mantenerli raccorciati nelle loro evoluzioni durante la mitosi e la meiosi. Giunse così quasi inaspettata la scoperta, d'importanza fondamentale, che il DNA è il vero materiale ereditario: l'informazione portata dai geni nei cromosomi risiede nelle precise sequenze di nucleotidi del DNA e i singoli geni debbono essere interpretati come molecole o come porzioni di una molecola filamentosa di DNA. Le prove di questo asserto sono irrefutabili e molteplici, e ci limiteremo a ricordare che il carico di DNA rimane costante nel nucleo in tutti gli stadi, in cariocinesi e in intercinesi: esso si raddoppia soltanto in uno stadio dell'intercinesi che precede la mitosi, per passare poi in quantità uguali ai due nuclei figli. Nel 1948, C. Vendrely. A. E. Mirsky e H. Ris ne hanno dato la dimostrazione contemporaneamente e indipendentemente, e con due metodi diversi, per le cellule di vari tessuti di una medesima specie e hanno anche osservato che la quantità di DNA ridotta alla metà negli spermatozoi, i quali infatti sono cellule con nucleo aploide. Durante l'intercinesi, dunque, il DNA non diminuisce, ma si colora poco perché è più diffuso nei cromosomi, che in tale stadio sono rigonfi, più distesi e più imbibiti d'acqua, mentre si concentra nella cariocinesi, quando i cromosomi si contraggono. Cadde così la maggiore obiezione rivolta al valore genico del DNA. A sostegno di tale concezione sta ancora la possibilità di separare dai virus e dai Batteri gli acidi nucleici non contaminati da proteine e ancora provvisti di potere infettante (v. acidi nucleici; v. virus).
La scoperta del valore genico degli acidi nucleici ha dato nuovo impulso alle ricerche sulla loro biofisica e biochimica. Abbiamo parlato di acidi nucleici in genere, perché il DNA ha valore genico in tutti gli organismi cellulari e in molti virus; tuttavia, si conoscono altri virus contenenti soltanto RNA, che deve necessariamente avere valore di materiale ereditario in questi Organismi relativamente semplici, che pur mantengono rigorosamente, nelle successive generazioni, le loro caratteristiche morfologiche e funzionali. La scoperta ha suscitato nuovi problemi: i geni, cui è legata la fondamentale invarianza degli organismi, devono essere capaci di reduplicarsi con grande precisione; e inoltre, se i geni con il loro DNA portano l'istruzione per la costruzione delle strutture caratteristiche degli individui e delle specie, e se le proteine costituiscono il substrato essenziale di queste strutture, si deve concludere che il DNA (o l'RNA in alcuni virus) dirige la sintesi proteica. Ecco i due grandi problemi: duplicazione del DNA genico (RNA in alcuni virus) e traduzione dell'informazione dagli acidi nucleici alle proteine; problemi ormai risolti, e che rappresentano le realizzazioni forse più importanti della moderna biologia molecolare.
c) La duplicazione del DNA
Nel 1951 E. Chargaff, studiando la composizione in basi dei DNA isolati da varie specie di piante e animali, riconobbe che le quantità molari di A e T, di G e C sono uguali (A : T = G : C = 1), mentre può differire ampiamente, con le specie, il rapporto (A + T) : (G + C). Ciò sta a indicare che vi deve essere una certa complementarità tra A e T, G e C, e che in qualche maniera le basi azotate si devono trovare a coppie. Partendo da questo reperto, J. D. Watson e F. H. C. Crick iniziarono lo studio diffrattometrico ai raggi X del DNA, riconoscendovi una struttura elicoidale; e nel 1953 fecero conoscere il ‛modello della doppia elica del DNA', che sul momento rappresentava un'interessante ipotesi di lavoro, ma che fu confermata da tante ricerche ispirate da questo modello, successivamente condotte e da nessuna contraddetta.
Il DNA è rappresentato come formato da due catene polinucleotidiche elicoidali e parallele. In ciascuna catena la continuità è assicurata dagli acidi fosforici che esterificano gli idrossili dei carboni 5′−3′−5′−3′ ... degli zuccheri dei successivi nucleotidi; le basi azotate delle due catene si affrontano in maniera che dove in una catena è un'adenina nell'altra si trova una timina e dove una guanina una citosina. Si stabiliscono così, tra le coppie di una base purinica e di una pirimidinica, precisi rapporti sterici, che permettono all'A di legarsi alla T, alla G di legarsi alla C, rispettivamente con due e tre legami a idrogeno. Il DNA acquista così una struttura secondaria, dove i legami H sono abbastanza solidi da dare stabilità alla doppia elica, ma possono anche sciogliersi uno dopo l'altro e permettere la separazione delle due catene. Il modello rispetta i rapporti equimolecolari messi in evidenza da Chargaff e permette di rappresentare un processo di duplicazione molto verosimile: si può immaginare infatti che un enzima possa a un certo momento iniziare l'apertura della doppia elica, liberando le basi complementari dai legami H, e proseguirla ordinatamente per tutta la lunghezza; ciascuna base diventa allora capace di legarsi a un mononucleotide di base complementare, attingendo da un pool di nucleotidi liberi nel succo nucleare, prodotti di sintesi immediata nella cellula o della demolizione di altro DNA assunto col cibo. Man mano che i singoli nucleotidi vanno a posto, si ristabiliscono i legami H con le basi complementari della catena che funge da stampo e i legami fosforici tra i mononucleotidi nuovi arrivati: in definitiva, da una doppia elica se ne formano due identiche. Tale identità presuppone che non siano avvenuti sbagli nella copiatura; ma, siccome questo processo è molto preciso, gli sbagli sono rari ed è assicurata una trasmissione pressoché invariante del patrimonio genico. Se capita invece un ‛errore di stampa', si avrà una ‛mutazione', cioè una differenza nelle sequenze delle coppie di nucleotidi, e la mutazione diventerà immediatamente ereditaria, perché il DNA mutato comincia a duplicarsi nella nuova versione. Torneremo un poco più a lungo sui fenomeni di mutazione, per i quali l'invarianza viene attenuata e si apre la possibilità di un'evoluzione.
Il modello di Crick e Watson ha resistito alle verifiche sperimentali e si può aggiungere che può essere applicato anche ai virus ad RNA, il quale in questo caso assume una doppia elicatura al momento della moltiplicazione della particella virale nella cellula parassitata. La conferma forse più brillante della teoria della doppia elica fu fornita pochi anni dopo (nel 1957) da A. Kornberg, che ha ottenuto la sintesi del DNA in vitro, riunendo in provetta i mononucleotidi delle quattro basi, una ‛DNA-polimerasi', cioè un enzima o un complesso di enzimi ottenuto da colture di Escherichia coli - batterio che rappresenta oggi per la genetica moderna quello che era la drosofila per la genetica della prima metà del secolo - e un ‛innesco' costituito da una piccola quantità di DNA di qualsiasi provenienza (da un virus, da un batterio o da un mammifero): dopo un certo tempo si ritrova una quantità di DNA molto superiore all'inoculo e l'analisi delle basi dimostra che il DNA di nuova sintesi ha la stessa composizione dell'innesco.
d) La sintesi proteica e il codice genetico che ne porta l'informazione
È un processo che si compie per tappe molteplici ed è conosciuto non solo nelle sue grandi linee, ma anche con una certa ricchezza di particolari. In questo articolo, che vuole essere introduttivo, ci limiteremo a seguire il processo logico che ha guidato la ricerca, senza spiegarne le tecniche (v. acidi nucleici).
Gli acidi nucleici, abbiamo detto, portano l'istruzione per la sintesi proteica: più precisamente, un gene è costituito da un tratto di elica di DNA con una certa sequenza di basi alla quale dovrà corrispondere una certa sequenza di amminoacidi. L'istruzione, si dice, è portata in codice; e si può pensare al codice telegrafico Morse, dove lettere e cifre, per un totale di una quarantina di segni, sono rappresentate da piccoli gruppi di punti e lineette. Un codice che usi quattro segni può specificare 4, 16, 64 oggetti a seconda che i segni siano presi singolarmente o a gruppi di due o di tre. Il codice genetico, per poter specificare con 4 basi azotate 20 amminoacidi, deve essere formato da triplette e soltanto da triplette. Certamente, esse sono esuberanti con le loro 64 combinazioni, ma negli acidi nucleici i nucleotidi sono legati sempre da identici legami fosforici. Ne risulta un codice ‛senza virgole', che deve essere letto a cominciare da un estremo, per gruppi dello stesso numero di segni, nel nostro caso per gruppi di tre. Vedremo, a proposito delle mutazioni, che questa previsione logica è stata verificata sperimentalmente.
Seguiamo ora un altro filo logico: il DNA è il depositario dell'informazione genica ed è capace di duplicarsi; ma quale è il valore dell'RNA? I suoi nucleotidi mostrano piccole differenze rispetto a quelli del DNA: lo zucchero con un ossigeno in più e l'uracile al posto della timina. Ben più importante è invece il rilievo che nell'RNA non si riesce a mettere in evidenza una precisa complementarità di basi e che l'analisi cristallografica non ne dimostra la struttura a doppia elica. L'RNA manca dunque di quella struttura che ne rende possibile la duplicazione, con la sola eccezione dei virus a RNA genico, gli unici nei quali esso assume la struttura a due filamenti. In tutti gli altri organismi, tutto l'RNA è stampato dal DNA: una RNA-polimerasi apre la doppia elica e fa funzionare da stampo una sola delle due eliche, quella che si legge nel senso 5′ → 3′, richiamandovi i nucleotidi con ribosio (l'A chiama l'U, gli altri si comportano come di solito) e saldandoli col legame fosforico. Per comprendere questa scelta tra le due catene, si tenga presente che esse debbono essere considerate, piuttosto che parallele, antiparallele e che la loro lettura, scorrendole a partire da un medesimo livello, si fa in una nel senso C5′ → C3′, nell'altra in senso contrario, grazie a un enzima capace di riconoscere questa differenza; alla fine, risulta stampata una catena semplice di RNA, che si stacca dalla sua matrice e può migrare nel succo nucleare e di là nel citoplasma, mentre il DNA resta ancorato ai cromosomi, che lo trasportano da una mitosi all'altra. Si è compiuta così una prima tappa, che rappresenta la ‛trascrizione' del codice dal DNA all'RNA. Per la seconda tappa, quella della sintesi proteica vera e propria, si parla di una ‛traduzione' RNA → proteina.
In questa seconda tappa si presenta una difficoltà che gli organismi hanno saputo aggirare con un processo di notevole complessità rivelatosi, al solito, perfettamente congruente con le leggi della fisico-chimica. La difficoltà deriva dal fatto che non esistono rapporti sterici tra una tripletta di basi di un acido nucleico e un singolo amminoacido: il meccanismo di traduzione è dunque complesso e qui non possiamo fare altro che accennarvi, sacrificando interamente l'esposizione delle tecniche di ricerca, ingegnose e precise, che consentono di seguire in vivo e in vitro le varie tappe del processo.
Il microscopio elettronico ha rivelato, come componente costante di tutti i protoplasmi, un tipo pressoché uniforme di particelle elementari, rotondeggianti, di circa 150 Å di diametro, i ribosomi, particolarmente abbondanti dove la sintesi proteica è più attiva. La loro separazione, mediante ultracentrifugazione degli omogenati cellulari, ne ha permesso l'analisi e li ha dimostrati molto ricchi in RNA, precisamente in parti pressoché uguali di RNA e di proteine. L'estrazione di tutto l'RNA cellulare permette poi la separazione mediante ultracentrifugazione frazionata di tre tipi di RNA: a) l'RNA dei ribosomi (rRNA), a grosse molecole; b) un RNA a molecola relativamente piccola (p. mol. 27.000), che si può chiamare RNA solubile o di trasferimento (transfer RNA, tRNA) per la sua particolare funzione nella sintesi proteica; infine c) un RNA polidisperso, che alla centrifuga si fraziona in diverse porzioni, perché di p. mol. diverso, l'RNA messaggero (mRNA), quello che porta al luogo di sintesi proteica l'informazione genica e che è formato da un numero di triplette maggiore o minore ed eguale al numero di amminoacidi della catena peptidica, più o meno lunga, da lui codificata. A questo punto si deve notare che tutti gli RNA sono trascritti dal DNA e che vi saranno quindi anche i geni per gli rRNA e per i tRNA; ma questi, indispensabili come strumenti di sintesi, non saranno tradotti a loro volta in proteine, mentre lo saranno gli mRNA.
I tRNA devono essere di almeno 20 specie differenti, affinché ogni tipo di amminoacido possa essere riconosciuto da un suo tRNA, che lo collocherà a un livello preciso del messaggero. Ogni tipo di tRNA possiede infatti, in posizione appropriata, un ‛anticodone', cioè una tripletta di nucleotidi a basi azotate complementari di quelle della tripletta situata nell'mRNA, che rappresenta il ‛codone', la parola di codice a tre segni che simbolizza quell'amminoacido. In effetti, ci sono più di 20 specie di tRNA, perché nel codice ci sono 64 combinazioni di basi per i 20 amminoacidi e a molti amminoacidi corrispondono più codoni. Né questa è una semplice previsione teorica: i tRNA sono ben conosciuti, separabili gli uni dagli altri, e la loro molecola relativamente piccola, di un'ottantina di nucleotidi, ha permesso l'analisi completa di molti di essi. Infine, per ogni tRNA esiste una specie di ‛acilsintetasi', una proteina enzimatica con due siti attivi per il riconoscimento di un amminoacido e del corrispondente tRNA.
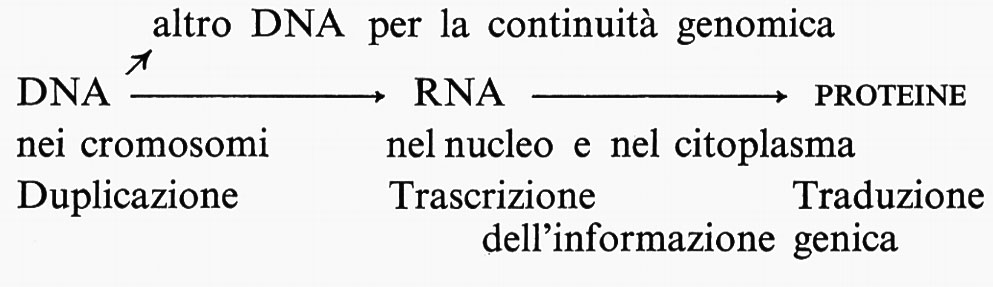
Il processo di traduzione può essere così descritto nel modo più schematico: ogni acilsintetasi riconosce il suo amminoacido e il suo tRNA e li unisce (questa è l'unica fase del processo nella quale una proteina riconosce specificamente una sequenza nucleotidica); intanto, una catena di mRNA ha preso rapporto con un ribosoma e il suo primo codone si mette in una precisa posizione, pronto ad accogliere l'anticodone del tRNA carico del suo amminoacido; successivamente, in qualche maniera non ancora precisata, il ribosoma scorre di un posto rispetto all'mRNA, e si mette così in posizione il secondo codone, arriva il secondo tRNA col suo amminoacido e tra i due amminoacidi ravvicinati si stabilisce il legame peptidico; il ribosoma assume una terza posizione, arriva il terzo tRNA col suo amminoacido, si salda un nuovo legame peptidico e il primo tRNA si libera. E così, passo a passo, è letto tutto l'mRNA, la catena peptidica si allunga sempre più e quando è completa si stacca. Intanto, man mano che un ribosoma si allontana dalla sua posizione iniziale, un altro subentra e poi ancora un altro, così che dopo un certo numero di spostamenti l'mRNA viene a trovarsi adagiato su un certo numero di ribosomi e le catene peptidiche sono montate in serie. Certamente il processo è più complesso di quello che abbiamo così sommariamente descritto; e la ricerca altro DNA per la continuità genomica è ancora attiva, perché molti particolari devono essere chiariti e non per semplice gusto di perfezionismo e di erudizione. I fenomeni, che scherzosamente è stato detto che costituiscono il ‛dogma centrale della biologia molecolare', possono essere così schematizzati:
Il ‛dogma' asserisce che il processo è unidirezionale; e tale deve essere la traduzione, perché la mancanza di adatti rapporti sterici toglie la possibilità che una proteina istruisca la sintesi di un acido nucleico. È invece teoricamente possibile una trascrizione inversa RNA → DNA e recentemente è stata effettivamente dimostrata nel ciclo biologico di alcuni virus cancerogeni: si tratta di virus a RNA genico, il cui ciclo vitale comporta l'insediamento nel DNA della cellula infettata, la quale assume così le caratteristiche di cellula cancerosa e le trasmette alle cellule discendenti. Per spiegare come ciò possa avvenire, si deve ammettere che il virus, penetrato in una cellula, sia in grado di stampare, per trascrizione inversa, un suo DNA capace di ingranare con quello dell'ospite. La previsione, apparentemente azzardata, che H. M. Temin formulò con precisione già nel 1967, è stata confermata da precise ricerche (1970 e tuttora in corso): è ormai dimostrato che il virus del sarcoma di Rous del pollo, così come quello della leucemia del topo, appena penetrato nella cellula infettata dirige la formazione di una DNA-polimerasi, la quale permette poi la trascrizione dell'RNA virale per dare un DNA che si alloga nel genoma della cellula. Per di più, questo tipo di DNA-polimerasi è stato trovato nei globuli bianchi di persone affette da leucemia e mai nelle persone sane, tanto che sembra definitivamente confermato il sospetto dell'origine virale della leucemia umana e appare giustificata la speranza che si possa arrivare a una vaccinoterapia dei tumori da virus.
Termineremo con l'accennare a quello che è stato il risultato se non più significativo certamente più spettacolare delle moderne ricerche di biologia molecolare: la decifrazione completa del codice genetico.
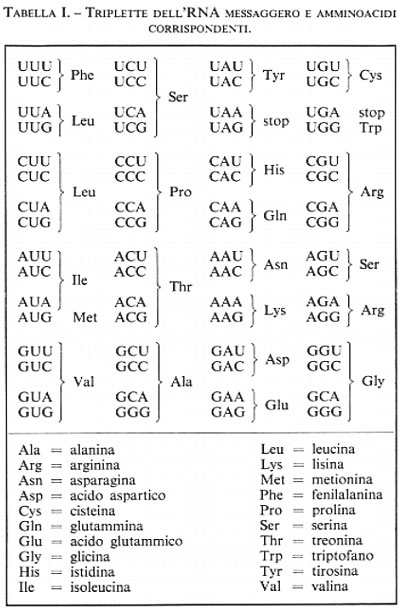
La ricerca, alla quale sono legati particolarmente i nomi di M. Nierenberg e di H. G. Khorana, è stata basata essenzialmente sulla sintesi in vitro e l'analisi degli amminoacidi delle brevi catene peptidiche specificate da molecole di RNA messaggero ottenute per sintesi e costituite da catene oligonucleotidiche di determinata composizione. Nella tab. I sono riportate le 64 combinazioni di basi (A, G, C, U) prese a triplette, vale a dire da ciascun codone dell'mRNA.
Il codice è ‛altamente degenerato', cioè a quasi tutti gli amminoacidi corrispondono più triplette. Tre sole sono le triplette ‛prive di senso', che non codificano alcun amminoacido, ma anche queste hanno un loro significato, sono cioè dei segnali di stop: di regola se ne trova una alla fine di Ogni mRNA (la tripletta complementare deve essere l'ultima di ogni messaggio genico del DNA), la quale indica che a questo punto il messaggio è stato letto fino in fondo e che la catena peptidica è terminata e può staccarsi dal ribosoma. Se per un caso, e se ne conoscono molti esempi, al momento della duplicazione del DNA si verifica un errore che trasforma una tripletta significativa di un amminoacido in una tripletta senza senso, si avrà poi un mRNA con uno stop che interromperà ad un certo punto la sintesi proteica: ne conseguirà la sintesi di una proteina troncata priva di funzione ed una mutazione ad effetto drastico. Si comprende quindi l'utilità di un codice degenerato: infatti, se in un codice soltanto 20 triplette avessero senso, una per ogni specie di amminoacidi, queste mutazioni dannose sarebbero molto più frequenti: in 44 casi, invece che in 3, su 64. Ancora, la tabella dimostra che spesso nelle triplette il terzo elemento non ha significato discriminatorio, o per lo meno che in tale posizione sono intercambiabili le purine, o le pirimidine: in tal modo, in circa il 30% dei casi, uno sbaglio di duplicazione, ripetuto poi nella trascrizione, non altera la scelta dell'amminoacido. La degenerazione, che appesantirebbe la decifrazione di un codice segreto, aumenta la stabilità del codice genetico. Infine, il codice è universale. Vi potranno forse essere delle eccezioni, ma queste non devono essere frequenti; nei casi finora studiati, dai virus all'uomo, le medesime triplette codificano i medesimi amminoacidi. È questa una eccellente prova dell'origine comune della vita, diversificata poi per evoluzione nelle infinite forme di microrganismi, di piante e animali.
5. La modulazione dell'attività genica
a) DNA genico e ridondante
I geni sono dunque formati da DNA (in alcuni virus da RNA), in doppia elica secondo il modello di Watson e Crick, e sono portati dai cromosomi, nei quali, almeno in tutti gli Eucarioti (cioè negli organismi superiori ai Batteri, che sono Procarioti), il DNA è associato a speciali proteine, gli istoni. La quantità di DNA di ciascuna cellula formerebbe una doppia elica molto più lunga dei cromosomi che la portano, ma l'elica vi è raggomitolata, fino a raccorciarsi 500 volte nell'unico cromosoma dei Batteri, fino a più di 10.000 volte nei cromosomi degli organismi superiori. I geni, come ipotizzò Th. H. Morgan più di mezzo secolo fa, in base al fenomeno del crossing over (v. genetica), sono regolarmente allineati nei cromosomi: oggi si dice che essi sono segmenti della doppia elica, ognuno con tante triplette in ciascuna delle due catene complementari, quanti sono gli amminoacidi costituenti la catena polipeptidica da esso codificata. Inoltre, i singoli geni che costituiscono insieme tutta la doppia elica di DNA di un cromosoma devono essere articolati in modo che ciascuno sia capace di aprirsi per stampare il proprio RNA, e collegati da tratti privi di valore genico. Poco sappiamo sull'intima struttura di questi collegamenti, sull'ampiezza della loro estensione e sul loro valore funzionale: probabilmente sono rappresentati da tratti di DNA inerte, incapace cioè di aprirsi per stampare un RNA messaggero. Ormai si può affermare che nei piccoli virus esiste la quantità di acido nucleico genico sufficiente a portare l'istruzione per la sintesi delle loro poche proteine, forse un poco di più per articolare i singoli geni. Ma probabilmente già nei virus più grossi e nei Batteri vi è più DNA di quello che si può prevedere per la sua funzione genica, per codificare tutte le proteine strutturali ed enzimatiche. Negli Eucanoti, poi, vi è sempre molto o moltissimo DNA privo di valore genico, in buona parte riconoscibile citologicamente per la sua caratteristica persistenza in forma contratta anche nei nuclei in interfase, la cosiddetta eterocromatina: nella drosofila, per esempio, da tempo sono ben note in tutti i cromosomi l'esistenza e la localizzazione di estese porzioni eterocromatiniche, prive o povere di geni. Ma il significato di questo ‛DNA ridondante' ancora ci rimane, diciamolo pure, perfettamente ignoto. Per portare un esempio tipico, tra le varie classi dei Vertebrati, e perfino tra certi gruppi minori, il carico di DNA per cellula varia entro limiti amplissimi: poiché molti Pesci ne posseggono una quantità minore rispetto ai Mammiferi, sembrava lecito attribuire alla maggiore complessità di questi ultimi la necessità del funzionamento di un più elevato numero di geni, espresso appunto dalla presenza di più abbondante DNA. Ma questa spiegazione non è valida per gli Anfibi Anuri e Urodeli e tra i Pesci per il gruppo dei Dipnoi, con un carico di DNA superiore a quello dei Mammiferi rispettivamente di circa 4, 10 e 20 volte: certamente una rana, una salamandra e un dipnoo non hanno bisogno di un genoma più complesso di quello del topo o dell'uomo, che hanno su per giù la stessa quantità di DNA per cellula, circa 5 miliardi di coppie di nucleotidi! Molto DNA vuol dire cromosomi e poi nuclei e poi cellule molto grosse; e infatti gli Anfibi, specialmente alcuni Urodeli, e ancor più i Dipnoi, hanno cellule relativamente enormi, che hanno fatto la gioia dei citologi. Probabilmente dimensioni cellulari maggiori sono correlate a un metabolismo basale più lento, mentre il ricambio è più attivo se le cellule sono miniaturizzate, in ragione dell'aumento del rapporto tra superficie cellulare, al cui livello hanno luogo gli scambi con l'ambiente, e massa del citoplasma. La cromatina ridondante, almeno ai limiti estremi ora ricordati, sembra un'ingombrante zavorra legata al metabolismo torpido delle cellule che la possiedono; ma il fatto che tutti gli organismi, anche quelli a ricambio più attivo, hanno una certa quantità di DNA non genico, non ha ancora ricevuto una spiegazione funzionale. Non è un problema assillante; eppure, un fenomeno generale riguardante una sostanza che ha certamente, nella sua parte genica, importanza fondamentale, fino a trent'anni fa insospettata, può riservare grosse sorprese.
b) Geni attivi e repressi
Ogni cellula possiede un corredo completo di cromosomi, ma in nessuna cellula tutti i geni sono contemporaneamente attivi. Così, nelle nostre cellule epidermiche, negli entrobiasti che si differenziano in globuli rossi del sangue, nelle cellule del pancreas, sono attivi i geni per la produzione della cheratina o dell'emoglobina o di parecchi enzimi digestivi; ma nessuna cellula elabora insieme tutte queste proteine. Anche in un batterio, in momenti diversi del suo ciclo vitale, entrano in azione geni diversi; e perfino nei virus si conoscono geni e ‛molto precoci', ‛precoci' e ‛tardivi'. I geni possono dunque trovarsi in stato attivo o represso; e in ogni cellula, in ogni momento funzionale, i geni repressi sono probabilmente più numerosi di quelli attivi, tanto che è invalso l'uso di parlare di geni repressi e ‛derepressi', come se la repressione fosse il loro stato normale. Questi fenomeni di repressione e derepressione genica, sui quali, malgrado le numerose acquisizioni, ancora molto deve essere chiarito, costituiscono un vasto campo della fenogenetica - lo studio cioè dei collegamenti tra geni e caratteri da questi controllati - dove si conducono attualmente ricerche d'avanguardia, certamente tutt'altro che semplici, ma i cui risultati è facile prevedere della massima importanza.
Nei Batteri già molto si conosce grazie alle ricerche pionieristiche di Jacob e Monod (1961), ampiamente confermate e ampliate: i due autori, in base a una penetrante analisi compiuta sull'Escherichia coli, hanno saputo dare un modello generale dei fenomeni di modulazione genica. Basterà qui dire che nel coli, organismo a breve vita, l'attivazione di un gene - nel caso studiato da Jacob e Monod, il gene per la produzione della galattosidasi, l'enzima per la digestione del lattosio, che viene indotto quando si offre al batterio un terreno di coltura ricco di tale zucchero - o il suo ritorno allo stato represso - l'abolita produzione di galattosidasi, divenuta superflua, che si verifica quando si porta la coltura su un substrato con altro zucchero - si compie nel giro di un paio di minuti. Jacob e Monod hanno dimostrato l'esistenza di un ‛gene regolatore' il quale stampa un repressore che, come è stato ora dimostrato, è una proteina allosterica: questa, a seconda che si leghi o no al lattosio presente o assente nel substrato), diventa incapace o capace di riconoscere con il suo secondo sito attivo un tratto di DNA, un ‛operatore' situato immediatamente a monte del gene che codifica la galattosidasi. In conseguenza, a seconda che il repressore possa unirsi o no all'operatore, il gene per la galattosidasi è represso o depresso, incapace o capace di trascrivere il suo RNA messaggero.
Negli Eucarioti, nei quali i cromosomi sono ricchi di istoni e di altre proteine che probabilmente hanno una parte importante nella regolazione dell'attività genica, il modello di Jacob e Monod non sembra immediatamente applicabile. In questi organismi, più complessi e a vita lunga, molte risposte dell'attività genica possono non essere così immediate come nei Batteri, ma devono avere un condizionamento più complicato; ed esistono geni capaci di avvertire stimoli diversi - per esempio ormonali e probabilmente anche di mediatori chimici di impulsi nervosi - atti a pianificare, per così dire, la produzione delle proteine, e con esse la tempestiva messa a punto di eventi morfologici e fisiologici di varia natura. Così in molti Vertebrati - e qualcosa di simile avviene anche in molti altri animali e nelle piante - l'allungamento delle giornate dopo il solstizio d'inverno è avvertito dalla retina: gli stimoli visivi prolungati provocano la secrezione di neuroni ipotalamici, il cui secreto giunge all'ipofisi, dove percuote nelle ‛cellule beta' i geni responsabili della produzione di una proteina a funzione ormonale, la quale a sua volta giunge per via sanguigna ai corpi genitali e vi stimola la spermatogenesi o l'ovogenesi e la produzione, da parte delle cellule endocrine testicolari od ovariche, di ormoni che controllano i caratteri sessuali secondari e il comportamento. In tal modo, per successive attivazioni geniche, tante specie a riproduzione stagionale andranno in amore all'epoca più propizia per il successivo sviluppo della prole; e le piante daranno fiori, frutti e semi alla stagione giusta. In molti casi, dunque, il modello di modulazione genica valido per i Batteri non sembra che si possa ‟estendere all'elefante", come, del resto, avevano già previsto Jacob e Monod.
c) Differenziamento cellulare; embriologia e genetica
Particolarmente complessi devono essere i fenomeni che si svolgono nel corso del differenziamento embrionale e perciò il loro studio si rivela arduo. Si vuole arrivare a comprendere come, negli organismi pluricellulari, le cellule dei vari distretti del germe producano, per l'entrata in attività di particolari geni in momenti precisi dello sviluppo, proteine diverse e specifiche; e come si avviino e si stabilizzino i processi che fanno differenziare cellule epidermiche, ghiandolari, scheletriche, muscolari, nervose, ecc. E si vuole arrivare non soltanto a riconoscere le immagini di questi differenziamenti - molte sono ormai note fino al livello della microscopia elettronica - ma a comprendere gli intimi processi fisico-chimici che presiedono ai fenomeni meravigliosi dello sviluppo di un uovo o di un seme.
I grandi progressi compiuti dalla ricerca embriologica nel nostro secolo e il fervore delle ricerche attuali sono ampiamente esposti in altre voci. Il nostro cenno sarà brevissimo e vuole semplicemente stimolare il lettore alla consultazione delle voci relative (v. embriologia; v. sviluppo embrionale). L'uovo fecondato inizia il suo sviluppo con la cosiddetta segmentazione, cioè con la sua divisione, per successive mitosi precisamente orientate, in più cellule (blastomeri) disposte con un disegno particolare e caratteristico dei vari organismi. In questo periodo il DNA, impegnato in rapide duplicazioni, trascrive poco o punto RNA. All'inizio dello sviluppo embrionale tutti i geni possono essere silenti e la scarsa sintesi proteica procede con gli mRNA prodotti in stadi precedenti nell'ovocita in accrescimento. La sintesi di nuove proteine, per attivazione di un certo numero di geni, si instaura dopo la segmentazione. Ma quando ci si chiede come si compiano questi processi di precise entrate in giuoco di geni diversi, dobbiamo ammettere che ancor oggi poco è da aggiungere a quanto scriveva Morgan in un suo saggio su Embriologia e genetica (1934). Morgan partì dalla constatazione dell'esistenza di una iniziale anisotropia del citoplasma dell'uovo: l'uovo non è infatti uguale in tutte le sue parti, come è ormai sicuramente provato. Così i blastomeri ricevono porzioni diverse dell'ooplasma e i loro nuclei, a contenuto genico equivalente, si trovano a lavorare in substrati diversi; geni diversi saranno in tal modo diversamente attivati in blastomeri diversi e aumenteranno sia le differenze citoplasmatiche sia le differenze nelle possibilità di lavoro del genoma. Così progressivamente, sotto l'azione reciproca del citoplasma e del nucleo, si precisa e si complica il differenziamento dei vari territori embrionali. A un certo momento la situazione si fa ancora più complessa per i fenomeni di induzione embrionale scoperti da Hans Spemann nel 1924 negli Anfibi. Queste ricerche hanno segnato una tappa essenziale dell'embriologia sperimentale; esse sono state ampiamente confermate in altri Vertebrati e, mutatis mutandis, in vari gruppi di Invertebrati: hanno quindi un valore generale. È stato dimostrato che alcune strutture capaci di autodifferenziarsi inducono poi il differenziamento di regioni embrionali ad esse circostanti (induzione primaria); e che una struttura così indotta può successivamente provocare, per induzione secondaria, un ulteriore differenziamento. Secondo le prime ricerche dello Spemann, si autodifferenzia la corda dorsale, che induce poi la comparsa dell'abbozzo del sistema nervoso; più tardi, un'estroflessione dell'encefalo si differenzia come retina e questa a sua volta induce la formazione del cristallino e delle membrane dell'occhio, la conoidea e la sclerotica. Ricerche recenti hanno dimostrato che le sostanze liberate dalla corda dorsale sono proteine, le quali raggiungono per diffusione le cellule ectodermiche ancora indifferenziate, ne modulano l'attività genica e provocano così il loro differenziamento in elementi nervosi.
Quest'ultimo reperto mostra chiaramente l'esistenza di un collegamento tra gli aspetti morfologici e biochimici da un lato e l'entrata in giuoco tempestiva dell'attività genica dall'altro, onde è facile prevedere che l'analisi di tanti altri fenomeni dello sviluppo potrà scendere fino al livello molecolare. Ma molto è ancora da fare.
E a questo punto si può notare che molti problemi di fenogenetica, riguardanti caratteristiche biochimiche metaboliche, sono ormai chiariti al livello della genetica molecolare: così nell'uomo e in tutti i Vertebrati esiste un gene (Cromogeno, C), a cui corrisponde la formazione di un enzima capace di ossidare la diossifenilalanina e dare origine al pigmento melanico, mentre un gene mutato (c) non sa dirigere la sintesi di una molecola enzimatica efficiente ed è perciò causa di albinismo; ancora, esiste un gene (s) per la catena beta dell'emoglobina umana, mentre una sua mutazione (S) provoca la sostituzione di un acido glutammico con una valina cui consegue la formazione di una emoglobina patologica (Hb S), causa di una emopatia, l'anemia falciforme, leggera in eterozigosi (Ss), in omozigosi (SS) gravissima e mortale nell'80% dei casi. E gli esempi si potrebbero moltiplicare, dalla genetica umana a quella dei virus. Ma quando si considerano tanti caratteri morfologici a evidente trasmissione ereditaria, se ne può riconoscere la base mendeliana, ma niente si sa ancora sui meccanismi molecolari che collegano l'attività di un gene, o di un gruppo di geni, con il differenziamento di quella determinata struttura. Nella drosofila si sa che un gene, situato nel cr0mosoma I, in posizione 13,7, è in qualche modo responsabile dello sviluppo dell'ala, tanto che una sua mutazione, crossveinless, determina una leggera modifica della nervatura alare; ma non è ancora possibile spiegare perché una proteina modificata sia in grado di determinare questo effetto morfologico limitato e preciso. Ancora, nell'uomo, l'esadattilia, cioè la comparsa di un dito soprannumerario, rappresenta un carattere ereditario, monogenico, dominante. Allo stesso modo, tanti piccoli caratteri somatici si rivelano ereditari per la loro assomiglianza familiare; e così si rivelano ereditari tutti i caratteri somatici nel loro complesso, per l'enorme assomiglianza tra gemelli monoovulari, tra individui cioè con identico patrimonio genico. Ma perché un gene mutato, che stampa una proteina diversa dalla normale, ha determinato per varie generazioni il labbro inferiore pendulo in vari membri della famiglia degli Asburgo, e perché a un gene mutato corrisponde l'esadattilia? E se si vuole passare da queste cronache familiari ai grandi eventi della storia evolutiva, perché nella famiglia degli Equidi, in parecchie diecine di milioni di anni, il sommarsi di mutazioni geniche selezionate per l'adattamento all'ambiente ha portato, dal piede tridattilo e dai denti molari poco efficienti dello Eohippus che viveva nell'Eocene, al piede unidattilo più agile e ai molari lofodonti, eccellenti per la pastura prativa, del cavallo pleistocenico e attuale? A queste domande non troviamo plausibili risposte. Riconosciamo i fenomeni, ma non ne spieghiamo l'intimo meccanismo. Ancor oggi, per quanto riguarda la mano con un pollice soprannumerario possiamo dire che chi la porta possiede un gene e una proteina dove è una virtus hexadactyla; tuttavia, per il fatto stesso che i problemi sono chiaramente impostati, si può prevedere che in una seconda edizione di questa enciclopedia molto più si potrà scrivere sulla fenogenetica dei caratteri morfologici. Per ora dobbiamo ammettere che molto lavoro è ancora da compiere per riconoscere i collegamenti, che certo esistono, tra biologia organismale e molecolare. E quando si riuscirà a spiegare la genesi di un carattere di dettaglio quale la venatura delle ali di un Insetto, si sarà trovato un modello per comprendere le trasformazioni di strutture importanti e il valore creativo dell'evoluzione basata sulla scelta ambientale di mutazioni geniche.
d) Le basi molecolari della mutazione genica
Abbiamo già detto che la duplicazione del DNA è quasi sempre precisa e che questa precisione spiega la proprietà dell'invarianza degli organismi. Ma talvolta può capitare un errore e da quel momento il DNA comincia a copiarsi nella nuova forma, perché, si potrebbe dire, subito si riafferma l'invarianza. Così un gene ‛muta' e la mutazione diventa ereditaria. Se le duplicazioni avvenissero senza mai uno sbaglio, i genomi rimarrebbero rigorosamente costanti e con essi le successive generazioni. Le singole specie, e quindi le flore e le faune, si perpetuerebbero identiche e veramente si potrebbe affermare oggi, come lo affermavano Linneo e la stragrande maggioranza dei biologi di due secoli fa, che tot sunt species quot ab initio Supremum Ens creavit. Ma ormai la teoria dell'evoluzione è una ragionevole certezza; e la via dell'evoluzione è aperta dalla possibilità di mutazione del patrimonio ereditario, cioè dagli errori nella duplicazione del DNA.
Lo studio della mutabilità è un capitolo della genetica nel quale la ricerca è giunta al livello molecolare; riteniamo di doverne dare almeno un cenno, perché la teoria molecolare della mutazione ha una decisiva importanza per la comprensione dei fattori evolutivi. E poiché studiare l'evoluzione significa vedere i problemi biologici a livello organismale e con mentalità storicistica, accennare ai fenomeni di mutazione significa mostrare, o per lo meno far intravedere, la necessità di intendere i fenomeni biologici dai due punti di vista: riduzionistico, con l'approfondimento permesso dalla moderna biochimica, e composizionistico, dove la biologia diventa storia della vita.
Si può definire come mutazione ogni modifica del genotipo che non sia dovuta a semplice ricombinazione di geni preesistenti. Si possono distinguere mutazioni geniche, per modifiche dell'intima struttura di singoli geni, e mutazioni cromosomiche, dovute ad alterazioni nella struttura e nel numero dei cromosomi. Come esempio di mutazioni cr0mosomiche ricorderemo i fenomeni di poliploidia, cioè l'apparizione spontanea, o provocata da tecniche appropriate, di individui con un numero 4, 6, 8... n di cromosomi, invece dei normali 2n. Le forme poliploidi presentano spesso il fenomeno del gigantismo e molte varietà di piante coltivate per fiori; frutta o semi sono poliploidi. In natura i fenomeni di poliploidia devono avere una certa importanza, specialmente nelle piante.
Per quel che riguarda le mutazioni geniche, dobbiamo ricordare che un gene è un tratto di una doppia elica di DNA (in alcuni virus di RNA) con due catene nucleotidiche a basi azotate complementari e con un numero di coppie di nucleotidi triplo del numero di amminoacidi della catena peptidica codificata dal gene stesso. Le mutazioni geniche possono essere alterazioni di un tratto più o meno esteso della doppia elica, o di una sola coppia di nucleotidi (mutazioni estese e mutazioni punto). Le mutazioni estese consistono generalmente nella perdita (‛delezione') di un numero di coppie di nucleotidi maggiore o minore, comunque superiore all'unità: probabilmente non sono rare e possono avere effetto cospicuo, quando la delezione sia notevolmente ampia e la catena polipeptidica venga poi a mancare di parecchi amminoacidi. Mutazioni estese sono state messe in evidenza nei microrganismi, con una tecnica che comporta la ricerca di fenomeni rari, ricerca possibile soltanto con organismi allevabili in grandissimo numero. Negli organismi superiori, per il limite numerico degli allevamenti, la ricerca è molto ardua. Si può pensare che queste delezioni nell'ambito di un gene siano dovute a deformazioni accidentali dell'elica del DNA, quando si apre per duplicarsi, tali da escludere un certo numero di nucleotidi. Per economia di spazio, siamo costretti a trascurarle.
Più facile lo studio delle mutazioni punto, che interessano una sola coppia di nucleotidi. Se ne possono prevedere di due tipi, per sostituzione di basi (SB), o per inserzione o delezione (ID). Nelle mutazioni ID, al momento della duplicazione s'inserisce un nucleotide in più, o se ne perde uno. Ricordando che il codice è a triplette senza virgole, si comprenderà subito che una inserzione, o una delezione, modifica le letture di tutte le triplette successive e viene codificata una sequenza di nucleotidi del tutto diversa. Le mutazioni ID hanno dunque effetto drastico, ma possono essere riparate completamente o quasi, se una D sopraggiunge in prossimità di una precedente I, o viceversa. In tal modo sono alterate poche triplette e, a seconda del valore dei pochi amminoacidi sostituiti, la riparazione può essere più o meno efficace.
Nelle mutazioni SB è sostituita una coppia di basi: per esempio si passa da A-T a G-C. Queste mutazioni hanno effetto circoscritto, comportano l'alterazione di una sola tripletta e, nella proteina, di un solo amminoacido, ma non sono facilmente riparabili, perché è poco probabile che una nuova sostituzione riparatrice capiti proprio nel punto esatto dove era avvenuta la mutazione. L'effetto di una mutazione SB sarà minore o maggiore a seconda che la sostituzione porti alla codificazione di un nuovo amminoacido poco o molto diverso, per il suo gruppo R, dal precedente; ed è particolarmente grave se la sostituzione riguarda un amminoacido in posizione strategica.
Si comprende così come si producano mutazioni con effetto fenotipico maggiore o minore; e siccome in una catena polipeptidica gli amminoacidi sostituibili sono più numerosi di quelli particolarmente significativi, le mutazioni a effetto leggero saranno le più frequenti. Più difficile invece spiegare il dato empiricamente accertato che esistono geni con frequenza di mutazioni - naturalmente con frequenza statistica - più o meno elevata: così nell'uomo le mutazioni per la corea di Huntington, l'ittiocheratosi congenita e l'emofilia capitano circa una volta su, rispettivamente, un milione, centomila e trentamila gameti; le mutazioni dei loci r, i, pr, su, per caratteri del seme del mais, hanno luogo 500, 100, 10, 2 volte su un milione di semi. Evidentemente, la mutazione sarà tanto più rara quanto più breve la catena nucleotidica e l'effetto sarà tanto più leggero quanto più numerosi gli amminoacidi simili nella proteina codificata dal gene. Ma è difficile giudicare se questa spiegazione sia sufficiente. Comunque, le mutazioni geniche a effetto fenotipico notevole sono sempre eventi rari: un valore di frequenza di mutazione dell'uno su ventimila è già alto.
Le mutazioni possono essere indotte sperimentalmente; fu un evento memorabile nella storia della genetica la dimostrazione, fornita da Hermann J. Muller nel 1927, che se ne poteva aumentare la frequenza, fino a oltre cento volte nella drosofila, mediante l'irradiazione con raggi X. Successivamente fu riconosciuto il potere mutageno di tutte le radiazioni ionizzanti (oltre alle onde elettromagnetiche di alta frequenza, i raggi corpuscolari da atomi radioattivi, le nuvole di neutroni ed elettroni veloci, ecc.). L'effetto mutageno è direttamente proporzionale alla dose di radiazione assorbita, ma è indipendente dal fattore tempo: uguali dosi, cioè dosi che producono un ugual numero di ionizzazioni, applicate massivamente o a concentrazioni diluite per tempi più lunghi o discontinui, inducono la stessa frequenza di mutazione. Ciò significa che la mutazione è un evento irreparabile: una volta che un gene è stato colpito, muta, senza una successiva possibilità di recupero. Per spiegare l'effetto mutageno, si ammette che l'impatto della radiazione, in un certo ambito d'urto dell'edificio atomico del gene, determini una ionizzazione, da cui deriva un rimaneggiamento della sua struttura; oppure, che venga ionizzata una molecola d'acqua immediatamente prossima al gene, con produzione di un radicale libero, un aggressivo chimico di brevissima vita, ma molto energico. Comunque ancor oggi, a più di 40 anni dalla scoperta del Muller, l'analisi dell'azione mutagena delle radiazioni ad alta energia non è ancora arrivata al punto da stabilire quali siano le precise modifiche a livello molecolare. Un dato, tuttavia, è ancora incontrovertibile: per avere l'irreparabilità di un evento mutageno provocato da una sola radiazione ionizzante, non esiste una soglia di azione, così che si deve ammettere che la contaminazione dell'ambiente da isotopi radioattivi creata dalle bombe atomiche 0 da centrali nucleari mal protette può essere causa di alterazioni geniche a qualsiasi diluizione; il pericolo è tanto minore quanto minore è la concentrazione del materiale fissile, ma non diventa mai nullo.
Meglio delle ricerche sui fattori fisici, quelle sui mutageni chimici permettono di arrivare a stabilire modelli molecolari di mutazione di validità generale, non limitati cioè ai fenomeni di induzione sperimentale, ma estendibili alle mutazioni che spontaneamente si producono in natura. Pioniere in questo campo è stata Charlotte Auerbach, che dimostrò nel 1942 l'azione mutagena dell'iprite; successivamente, le ricerche furono estese a molte altre sostanze. Esemplificheremo rapidamente: l'acido nitroso è un mutageno che agisce sul genoma in riposo, in quanto trasforma, per deaminazione, l'adenina e la citosina rispettivamente in ipoxantina (I) ed uracile (U); il successivo accoppiamento avviene tra I e C e tra U e A, e nei due casi si arriva ad una sostituzione di basi. Nello schema sono indicati i passaggi in due successive duplicazioni, quando si parte dalla coppia A-T e si arriva alla G-C,
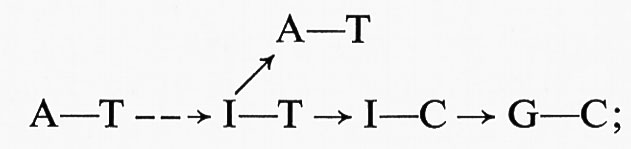
o, viceversa:
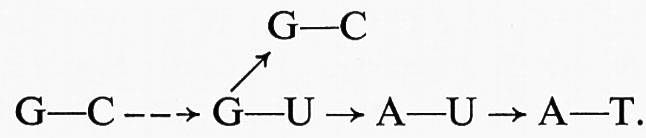
Agiscono invece sul DNA in duplicazione le cosiddette basi analoghe, le quali, aggiunte al terreno di coltura, possono essere incorporate nel DNA. Così, il 5-bromouracile (BU) è abbastanza simile alla timina e vi si sostituisce a far coppia con l'adenina, così che alla prima duplicazione una coppia A−T diventa A−BU; ma talvolta il BU assume una forma enolica (BU*), con perdita di un H, e il BU* alla seconda duplicazione farà coppia non più con l'A ma con la O, finché alla terza duplicazione la O chiama la C. In definitiva, A−T è sostituito da G−C, secondo lo schema:
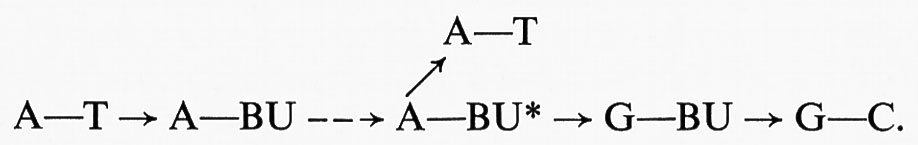
Le acridine hanno affinità per gli acidi nucleici; per questo alcune di esse sono adoperate in citologia come coloranti nucleari: non stupisce quindi che esse abbiano anche azione mutagena. I mutanti da acridina, nuovamente trattati in successive generazioni, ritornano spesso a una forma simile se non identica all'originale e questo rivela le acridine come mutageni per ID; si pensa anche che un'acridina si piazzi tra due basi puriniche e causi una distorsione locale dell'elica di DNA, che si traduce poi in una duplicazione anomala.
Si pone ora il problema, quale sia l'origine delle mutazioni spontanee. I mutageni chimici impiegati in laboratorio non hanno probabilità di arrivare agli organismi in condizioni normali. La radiazione naturale, in rapporto ai raggi cosmici e all'esistenza di una piccola quantità di elementi ‛radioattivi, è molto debole, e non la si può ritenere responsabile che di una piccola frazione della mutabilità spontanea: di un decimillesimo nella drosofila, dell'un per mille nel topo, che vive più a lungo, del 5-10% nell'uomo, con periodo riproduttivo di trenta anni. Una spiegazione attendibile è stata data da Crick, secondo il quale, ammesso che la specificità degli appaiamenti (A−T, G−C) sia dovuta alle caratteristiche steriche che ciascuna base possiede nella sua forma più stabile, si può tuttavia prevedere che una base, per il salto di un idrogeno, possa assumere una forma tautomera meno stabile e quindi più rara, nella quale si appaia a un'altra base: così, la T e la C possono talvolta passare a una forma nella quale un H è saltato dall'azoto in posizione 3′ al carbonio in 5′, e in tali circostanze il loro appaiamento avverrà, rispettivamente, con la G e l'A; alla successiva duplicazione le basi saranno tornate alla forma stabile, ma ormai hanno un nuovo complemento, che darà una nuova coppia, cioè una mutazione SB. Ecco lo schema di una SB da G−C ad A−T, dove la forma tautomera è indicata dall'asterisco:
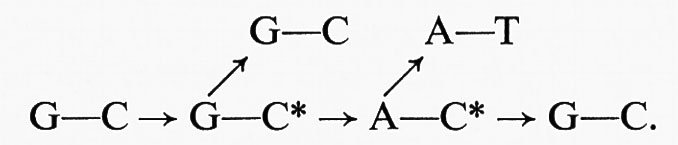
Meno facile spiegare le mutazioni per ID; probabilmente prendono origine da una distorsione dell'elica di DNA: se, per esempio, uno dei due filamenti si scolla per un certo tratto dal complemento, forma un cappio più o meno ampio ed esclude dalla duplicazione uno o più nucleotidi (delezione punto, o estesa); oppure, si spezza un legame fosforico e i due monconi si risaldano dopo aver accolto un nuovo nucleotide (inserzione punto).
Questo breve riassunto delle lunghe ricerche sulle mutazioni, spontanee o indotte sperimentalmente, ci permette di concludere che le mutazioni sono eventi casuali, intendendo con tale aggettivo non solo l'imprevedibilità per noi, ma una fondamentale ‛erraticità' del fenomeno. Una timina a un certo momento assume una forma tautomera meno stabile e quindi più rara di quella nella quale si trova il più delle volte; e in questa nuova forma si stabiliscono le appropriate relazioni steriche e i ponti a idrogeno con la guanina invece che con l'adenina. La mutazione obbedisce alle leggi fisico-chimiche come vi obbediscono tutti e sempre i fenomeni biologici; ma la modificazione tautomera può aver luogo in una qualsiasi delle molte timine dei molti geni che costituiscono il patrimonio ereditario. Muta ora un punto ora un altro, ora in un gene ora in un altro; e tutto quello che sappiamo sulla struttura del DNA come ordinata sequenza di geni, e dei singoli geni come sequenze di nucleotidi, ci fa escludere che un certo gene debba mutare ineluttabilmente a un certo momento e che possa esistere un coordinamento per forze interne nella mutabilità di un complesso genico. Queste conclusioni si possono considerare come definitivamente provate dalla moderna genetica molecolare; nel prossimo capitolo ne vedremo l'importanza per la valutazione degli agenti evolutivi.
6. L'evoluzione
La teoria dell'evoluzione, dapprima geniale intuizione di Lamarck (Phllosophie zoologique, 1809), poi meditata e documentata teoria presentata da Darwin con On the origin of species (1859) e presto diventata ragionevole certezza, rappresenta una delle maggiori affermazioni del pensiero scientifico del sec. XIX. Difficile, e discretamente inutile, cercare la scoperta massima, l'Oscar del pensiero biologico: i morfologi possono vantare la teoria cellulare di J. M. Schleiden e Th. Schwann; i fisiologi l'impetuoso rinnovamento metodologico dovuto a Claude Bernard; e chi guarda alla ricerca di base come origine di importanti applicazioni pratiche penserà a Louis Pasteur; ma è certo che la teoria dell'evoluzione, la splendida affermazione della biologia intesa con spirito naturalistico, ha rinnovato tutto il pensiero biologico, dalla morfologia anatomica, istologica ed embriologica, alla fisiologia e alla biochimica ed è diventata il pensiero dominante del sistematico neontologo e paleontologo, dell'ecologo, del biogeografo, dell'antropologo. Nemmeno è rimasta estranea al pensiero filosofico; e anche gli idealisti più intransigenti nel negare alla ricerca naturalistica il diritto di accesso alla filosofia intesa come scienza propria dello spirito umano, hanno combattuto naturalismo ed evoluzionismo, ma non hanno potuto ignorarla.
La vittoria, dopo la pubblicazione dell'Origine delle specie, fu rapida: nel campo delle scienze biologiche le resistenze furono di pochi e l'opposizione in campo extrascientifico dei vari vescovi Wilberforce, astiosamente contraddittori degli Huxley, fu facilmente travolta. Così, verso la fine del secolo, le prove che vi è stata evoluzione, fornite dall'anatomia, dall'embriologia e dalla fisiologia comparate, dalla sistematica che diventa ricerca di filogenesi, dalla paleontologia, avevano convinto ogni biologo che non volesse chiudere gli occhi per non vedere. Ma sulla dinamica dell'evoluzione, sulle forze che la portano avanti, non si raggiungeva un'intesa. Anzi, mentre si consolidava il convincimento che c'è stata evoluzione, che creazionismo e fissismo sono miti ormai dissolti, s'inasprivano le polemiche sulla valutazione degli agenti evolutivi.
Lamarck e Darwin avevano dato due diverse spiegazioni, ambedue razionali, del processo evolutivo. Lamarck aveva affermato l'eredità dei caratteri acquisiti: l'uso o il non uso di un organo, determinato dalle condizioni di vita, cioè dall'ambiente in cui gli individui si trovano a vivere, è causa a sua volta dello sviluppo maggiore o minore dell'organo; e le modifiche così evocate si trasmettono, almeno in parte, alla prole. Darwin partiva da due dati sicuri: primo, che nell'ambito di ogni specie esiste, per i diversi caratteri, una variabilità individuale, in buona parte ereditaria (e Darwin accettava il dato empirico senza poterlo spiegare), e, secondo, che ogni specie è tanto prolifica che a ogni generazione la maggior parte dei figli muore prima di arrivare a riprodursi; e concludeva che la selezione ambientale favorisce, statisticamente, i meglio adattati, rafforzando così l'adattamento all'ambiente e permettendo nuovi adattamenti in ambienti diversi. Ed ecco che, pur mantenendo il convincimento della realtà dell'evoluzione, si cominciò a criticare del darwinismo l'aspetto casualità, la fortuna o sfortuna di sopravvivere, dimenticando che proprio la selezione operata dall'ambiente rappresenta l'elemento anticaso; e si tornava a considerare la possibilità dei caratteri acquisiti affermata dal Lamarck, di un fattore evolutivo, che se fosse vero (ed è troppo bello per essere vero) spiegherebbe ancor meglio della scelta darwiniana il fenomeno indubitabile dell'adattamento all'ambiente.
Fra le due concezioni, lamarckiana e darwiniana, si insinuava l'affermazione di vari autori di una evoluzione per cause interne agli organismi, che avrebbero un patrimonio ereditario così fatto da doversi modificare ineluttabilmente e in direzioni predeterminate e precise. Perciò si usa dire che all'inizio del nostro secolo l'evoluzionismo era in crisi. E in un certo senso ciò è vero, in quanto la dottrina dell'evoluzione non deve essere soltanto negazione di un fissismo ormai contraddetto da una documentazione imponente e l'affermazione che i fenomeni biologici vanno visti sotto l'aspetto storico; ma deve anche significare riflessione critica sugli agenti evolutivi e raggiungimento di una sintesi attendibile. Ma è anche vero che la certezza sul trasformismo rimase sempre salda e che tutti lavoravano con mentalità evoluzionistica; solo che molti si rifiutavano di entrare nella polemica tra neodarwinisti, neolamarckiani e fautori di un'evoluzione per cause interne. Comunque, la crisi fu superata piuttosto rapidamente, e ormai vorremmo aggiungere definitivamente, soprattutto per l'apporto della ricerca genetica. Un apporto che non poteva essere che decisivo, perché evoluzionismo vuol dire, essenzialmente, processo di incessante modifica del patrimonio ereditario per mutazioni insorgenti come singoli accadimenti nel genoma degli individui e che poi si diffondono e si dilatano a livello di popolazioni.
Importanti per determinare questa svolta sono stati i modelli matematici di popolazioni elaborati da R. Fisher (1929), da S. Wright (1931), da J. B. S. Haldane (1932). Ed è questo uno dei pochi casi, vorrei dire l'unico, in cui un problema di biologia teorica, che si è avvalso di una trattazione matematica piuttosto complessa, ha aperto la strada alla ricerca con metodo induttivo, in questo caso agli studi sugli equilibri di popolazioni osservate in natura o in allevamenti sperimentali. Nel 1937 Th. Dobzhansky, che era entrato nel laboratorio di Morgan con una mentalità e una buona preparazione di naturalista di campagna, pubblicò un libro che ha fatto epoca, Le genetica e l'origine delle specie; e nel 1924 J. Huxley, anch'esso buon naturalista, seppe affermare nel titolo di un'opera ancor oggi interamente valida, Evoluzione: la sintesi moderna, questa decisiva unione dei dati ottenuti con i metodi classici dell'evoluzionismo - paleontologia, sistematica filogenetica, biogeografia, adattamento all'ambiente, ecc. - e col recente apporto della genetica. Ormai in questa sintesi il fattore lamarckiano era trascurato, perché si avvertiva che i fenomeni interpretati come eredità di caratteri acquisiti trovavano una migliore spiegazione in termini di frequenze di geni e di alleli comparsi per mutazioni, che già apparivano casuali e non correlate, e in termini di genetica di popolazioni, dove queste frequenze possono essere modificate dalla scelta ambientale. E alla fine si disse chiaramente che invocare un'evoluzione per sole forze interne significava parlare un linguaggio extrascientifico; giudizio cortese per un almanaccare su insensate analogie, come quando si affermava che le specie nuove sono in potenza nelle ancestrali, come è in potenza il pulcino nell'uovo. Intanto la paleontologia e la sistematica modernamente intese (ricorderemo soltanto due nomi: G. G. Simpson ed E. Mayr) confutavano l'ineluttabilità delle cosiddette leggi empiriche dell'evoluzione, affermanti, ad esempio, la completa irreversibilità degli orientamenti evolutivi che procederebbero per ortoevoluzione, o l'invecchiamento delle linee filetiche e la loro morte naturale, come invecchiano e muoiono gli individui (v. paleontologia; v. sistematica biologica).
Tutto questo fu riconosciuto e affermato quando la genetica molecolare cominciava appena a muovere i primi passi e quando i geni si ritenevano ancora costituiti da proteine. Oggi la biologia molecolare elimina definitivamente il fattore lamarckiano, negando la possibilità di una traduzione nel senso proteine acidi nucleici; ed esclude la comparsa di modifiche del patrimonio ereditario puntuali e coordinate, perché si conoscono ormai le basi molecolari della mutazione e la fondamentale erraticità del fenomeno.
Resta dunque, si può dire, integra la spiegazione darwiniana, solo che si cambi il nome sport con mutazioni; e che non si tradisca il concetto di selezione naturale, intendendola soltanto, sempre e ovunque, come lotta senza quartiere. In realtà, molto più spesso essa va vista come un vantaggio riproduttivo: in una popolazione gli individui che per certe caratteristiche genetiche hanno statisticamente più discendenti tendono dopo un certo numero di generazioni ad affermarsi rispetto al resto della popolazione stessa; e così la selezione, scegliendo i fenotipi, cioè gli individui coi loro caratteri manifesti, sceglie e fa progredire la frequenza di certi geni rispetto ad altri.
Possiamo dunque concordare con Simpson, per il quale ‟è definitivamente stabilito che la selezione naturale è il fattore essenziale per orientare e dirigere l'evoluzione" (v., 1969, p. 121). Ma la selezione naturale è un principio che opera in natura con grande varietà di modi, che non possono essere qui illustrati (v. evoluzione). Ci limiteremo a pochissime notazioni.
L'evoluzione è opportunistica; cioè, una popolazione si adatta al momento a un dato ambiente, perché la selezione, come ogni altro agente naturale, non lavora in vista di un fine. E quando una specie ha raggiunto un buon adattamento rispetto ad un determinato ambiente, lo mantiene: la selezione ha in questo caso valore conservativo. Ma quando l'ambiente cambia, o quando viene tentata l'esplorazione di un nuovo ambiente, la selezione deve cambiare l'adattamento, deve riassumere un valore creativo di novità; e se non vi riesce, le forme ormai disadattate periclitano e si estinguono, o ripiegano nella loro vecchia nicchia ecologica. Perciò un adattamento preciso a un determinato habitat, che rappresenta per una specie, o per un gruppo sistematico maggiore, un momentaneo successo, può ritorcersi, quando il complesso ecologico si altera, in un danno mortale.
Così si comprende anche perché le mutazioni siano dannose molto più spesso che giovevoli: perché un organismo bene adattato ad un certo ambiente ha raccolto e integrato un complesso genico, nel quale una novità turba più spesso che non migliori l'armonia stabilita dalla passata storia evolutiva. Ma se cambia l'ambiente, le mutazioni che prima avrebbero portato danno possono diventare vantaggiose: così un mantello bianco, che distruggerebbe l'adattamento mimetico a un suolo bruno, può essere utile in una popolazione che ha raggiunto un habitat nivale. Per analogia, è difficile portare una modifica in un'opera d'arte valida; ma quando cambiò il gusto, cioè l'ambiente culturale, tante architetture romaniche, gotiche o rinascimentali furono rinnovate, cioè, si diceva, abbellite da apporti barocchi.
7. Vitalismo e meccanicismo. Finalismo e causalità. Biologia molecolare e organismale
Un breviario di biologia, per quanto stringato, richiederebbe molte più pagine di quelle a disposizione per il nostro articolo. Questa vuole in realtà essere la giustificazione per i troppi argomenti taciuti o appena accennati. Abbiamo trascurato del tutto la sistematica, che l'avvento dell'evoluzionismo ha profondamente rinnovato, come ricerca di filogenesi e ricostruzione della storia della vita, per il lavoro comune dei paleontologi e neontologi di campagna e, perché no, degli specialisti museologi. Completamente ignorata si può dire anche l'ecologia; e la sua fondamentale importanza dovrà essere apprezzata dal lettore soltanto in base ai fugaci accenni al valore creativo dell'ambiente, che, determinando la scelta delle mutazioni, porta avanti gli adattamenti e guida i processi evolutivi. D'altra parte l'attenzione ai fenomeni ecologici è oggi continuamente richiamata dal gran scriverne e parlarne sui giornali, alla radio, alla televisione e nei parlamenti, perché alla fine ci si è accorti dei danni che l'uomo ha apportato agli ambienti naturali già da secoli (si pensi al disboscamento dei dossi montani); danni che oggi, con la tecnologia esasperata, sono notevolmente aumentati e possono diventare irrimediabili nel giro di pochi decenni. E si sarebbe dovuto parlare dei processi della biochimica, del riconoscimento esatto delle grandi vie metaboliche e accennare almeno ai rapporti tra fotosintesi e respirazione e ai due grandi processi di sintesi e demolizione: da una parte ai processi chimici endoergonici di costruzione delle grosse molecole organiche a partire da composti inorganici parvimolecolari e con l'apporto dell'energia radiante solare, dall'altra ai processi inversi esoergonici che liberano energia e riportano alle piccole molecole. Si sarebbe visto, così, che per questo continuo flusso di materia e di energia gli organismi, ciascuno e tutti insieme, rappresentano un sistema largamente aperto, il quale, pur mantenendo un grado elevatissimo di ordine, non contraddice al secondo principio della termodinamica, all'ineluttabile aumento di entropia che dovrebbe portare gradualmente all'uniformità, alla degradazione e alla morte. Non vi contraddicono, perché il bilancio entropico è saldato dall'energia del sole, dove aumenta l'entropia: quando il sole si sarà raffreddato, anche la vita si spengerà. Ed è questa una nuova prova che in ogni fenomeno biologico un sufficiente approfondimento della ricerca porta al rifiuto di ogni postulato vitalista, che affermi l'impossibilità di spiegare i fenomeni della vita con le leggi vigenti nel mondo dell'inorganico.
Nelle pagine introduttive è stato accennato al fatto che in ogni organismo e, si può dire, in ogni sua struttura è evidente un finalismo, una teleonomia, per usare un termine coniato dal Monod; e si è messa in evidenza la contraddizione tra questa caratteristica degli oggetti del nostro studio e il metodo scientifico dominato dal principio di causalità, ma subito si è aggiunto che la contraddizione è superata, solo che si guardino gli organismi non all'istante, ma come punto di arrivo di una lunga storia evolutiva. E ora, dopo aver chiarito le basi molecolari della mutazione e dell'evoluzione, crediamo di aver dimostrato questo superamento. Abbiamo infatti mostrato il valore creativo di un'evoluzione portata avanti dalla selezione naturale, cioè dalla scelta di mutazioni che insorgono continuamente nel patrimonio genetico per fenomeni che obbediscono alle leggi fisico-chimiche, ma che sono caratterizzati da una fondamentale erraticità. E questa evoluzione per scelta ambientale spiega i progressivi adattamenti e i finalismi che ritroviamo oggi in ogni struttura vivente, che il paleontologo ritrova nei dinosauri e nelle trilobiti e che certamente si sarebbero riconosciuti - a un livello elementare - nelle antiche associazioni di molecole organiche, non ancora arrivate alla struttura cellulare, ma già capaci di duplicazione. Il finalismo che oggi caratterizza gli organismi è dunque da vedere come una progressiva affermazione, non come una proprietà della vita stabilita fin dall'inizio da una misteriosa entelechia, da un agente che ha il suo fine in sé e che scese e si insediò negli organismi e nelle sostanze di cui sono fatti, fin nelle singole molecole proteiche e di acidi nucleici. E qui si può aprire una parentesi, per osservare che sembra imporsi la scelta del fattore darwiniano, perché l'eredità dei caratteri acquisiti è ormai insostenibile. La spiegazione lamarckiana sarebbe stata anch'essa accettabilissima, come lo sarebbe un altro qualsiasi fattore razionale, se qualcuno sapesse intuirlo e poi dimostrarlo: ma deve essere un fattore razionale, un principio, cioè, che possa essere introdotto in un discorso scientifico, non un postulato vitalistico indimostrabile.
Una veduta coerente dei fenomeni biologici, che escluda postulati vitalistici e superi la contraddizione tra finalismo e causalità, era ricercata da tempo, ma è stata acquisita, ci sembra di poter dire definitivamente, soltanto in tempi recenti per l'integrazione dei contributi forniti dalle ricerche sia in campo molecolare sia in quello organismale. La biologia molecolare ha dimostrato infatti l'aderenza dei fenomeni biologici ai fenomeni fisico-chimici vigenti nel mondo dell'inorganico e ne abbiamo trovato esempi sicuri nelle strutture macromolecolari; ma ci fa ritrovare anche a questo livello le tipiche caratteristiche teleonomiche, i siti attivi delle proteine enzimatiche, le triplette di nucleotidi negli acidi nucleici e il codice genetico, ecc. E questo succede perché la fisica e la chimica studiano fenomeni senza storia, ripetibili e che sono oggi quelli che erano un miliardo di anni fa: e quando li ricerchiamo negli organismi, li ritroviamo al momento e non ci forniscono la possibilità di ricostruirne la storia passata. L'apparente contraddizione tra finalismo e causalità deve essere risolta in termini evoluzionistici, cioè dallo studio della vita come processo storico; e questo studio si avvale bensì degli elementi forniti dalla biologia molecolare per valutare le possibilità di mutazione del patrimonio genico, ma è essenzialmente di pertinenza del biologo organismale. Si conferma così la complementarietà e la uguale dignità dei due metodi di ricerca; e si comprende perché il vecchio dissidio tra vitalismo e meccanicismo, cominciato nel sec. XVII e continuato per tutto il Settecento, in epoca comunque ancora legata a una concezione della biologia fissista e creazionista, non poteva terminare con la vittoria dell'una o dell'altra teoria, ma doveva venir superato finalmente in una visione evoluzionistica, storicistica.
I meccanicisti si ritenevano giustamente più aderenti al pensiero scientifico, in quanto affermavano l'esigenza di spiegare i fenomeni della vita alla stregua di quelli della fisica, fino ad asserire che, almeno per la vita fisica, ‛anche l'uomo è una macchina'. Ma anche questo era un modo di ammettere il finalismo, perché le macchine sono costruite dall'uomo in vista di un fine. I meccanicisti rifiutavano dunque il vitalismo affermando insieme il loro teismo: i viventi obbediscono alle leggi del mondo inorganico, ma la macchina umana e ogni altra macchina vivente sono state costruite da un Dio Creatore in vista di un fine. Ma ora che siamo arrivati ad ammettere che gli organismi hanno dietro a loro una lunga storia evolutiva e che l'evoluzione è portata avanti da cause naturali - molto probabilmente dalla scelta ambientale - diventa lecito rinunciare a un intervento sovrannaturale, pensando agli organismi come venuti su, costruitisi secondo leggi di natura, senza previsione di un fine. Si allargano così le frontiere della scienza anche nel campo dei viventi; e questa posizione non è preclusa necessariamente a chi non sia agnostico o ateo, perché il metodo scientifico non può rispondere e nemmeno formulare la domanda, se sia possibile la conoscenza di Cause prime.
I biologi devono lavorare con mentalità agnostica; e ormai rinunceranno ad alcune credenze, per esempio all'accettazione letterale del libro della Genesi. Ma la loro visione del mondo può essere diversa secondo gli individui. Esistono infatti menti creatrici in campi diversi: poeti, pittori e musicisti spesso negati al ragionamento scientifico e scienziati negati all'intuizione artistica. Similmente, ci sono fisici e biologi che alla scienza non chiedono più di quello che può dare e ne restano appagati; e altri che di fronte al mistero - e sempre rimarrà un fondo alogico della natura - sentono il bisogno di superarlo con un atto di fede che trascenda la ragione. Ci sono e ci saranno sempre biologi agnostici che rifiutano ogni dogma e sempre ci saranno teisti e atei.
8. La biologia e l'uomo
a) Le scienze biologiche e la concezione del mondo
L'uomo, ormai non vi è dubbio, è un prodotto dell'evoluzione, portata avanti da fattori naturali, da una selezione nella quale ha primeggiato l'intelligenza, attraverso lo sviluppo dell'encefalo. I reperti paleontologici non fanno che confermare il posto dell'Uomo nella Natura", per usare il titolo di un libro scritto da Thomas H. Uuxley poco dopo l'apparizione dell'Origine delle specie, nel 1863. Così noi sappiamo oggi che questa evoluzione ha portato dai bruti all'uomo, con un progresso del volume delle masse cerebrali dai 500 cm3 degli Australopitechi ai 680 dell'Homo habilis, agli 800 e 1.100 degli H. erectus di Giava e della Cina, ai 1.250 dell'H. sapiens steinheimensis, ai 1.400 dell'H. sapiens sapiens, quale lo conosciamo dall'Aurignaciano in poi. Ma da alcune diecine di migliaia di anni l'aumento del volume cerebrale e dell'intelligenza è stato trascurabile: basta, per convincersene, visitare le grotte di Altamira e di Lascaux.
Oggi l'evidenza di questa evoluzione è tranquillamente ammessa; il distacco dalla tradizione è avvenuto con una relativa facilità, che l'uomo di scienza deve considerare lusinghiera e stimolante, come prova dell'affermarsi della verità scientifica, del prevalere della ragione sui miti. Eppure, questa verità alla sua prima enunciazione aveva suscitato profonde avversioni: Carlyle rifiutò di entrare in relazione con Huxley, ‟l'uomo che dice che discendiamo dalle scimmie". Ma le prove dell'origine dell'uomo e la teoria generale dell'evoluzione impongono una rinuncia che va ben oltre la riluttanza ad ammettere remoti antenati preumani. All'uomo non si può più assegnare un posto privilegiato, in quanto approdo necessario, ineluttabile, di una evoluzione organica che avrebbe preparato il suo avvento, che si sarebbe svolta in vista di questo fine. Per molti biologi una tale conclusione, che porta a una visione del mondo profondamente diversa da quella animistica e mitica, si impone con la stessa evidenza con la quale si sono imposti i dati meramente naturalistici della paleoantropologia fisica; ma per alcuni biologi, e per moltissime persone di varia cultura, questa rinuncia può essere dolorosa, devastante. Monod, un biologo geniale e orgogliosamente assertore del metodo scientifico, avverte, senza parteciparvi, ma senza cercare di minimizzarla, questa angoscia di tanti; perché l'uomo, se accetta il messaggio scientifico, ‟deve [...] scoprire la sua completa solitudine, la sua assoluta stranezza. Egli ora sa che, come uno zingaro, si trova ai margini di un universo in cui deve vivere. Universo [...] indifferente alle sue speranze, alle sue sofferenze, ai suoi crimini". D'altra parte, già sono numerosi quelli che non ne sono turbati. Per molti, infatti, può essere accettata serenamente, ed essere motivo di impegno, la consapevolezza che la propria vita si compie su questa terra e che l'uomo può dare un giudizio di valori per la consapevole scelta dei mezzi in vista dei fini; e deve darlo anche senza attese ultraterrene, per atto di amore - e se non vogliamo usare parole altisonanti, diciamo per atto di tolleranza - verso gli altri uomini, per porre un argine alla lotta che ha continuato, e ancora continua, con durezza inutile, come lotta intraspecifica, sconosciuta altrove. Durezza veramente inutile perché, ripetiamo, per la nostra specie sembrano ormai trascurabili le possibilità di ulteriori progressi per selezione naturale.
La lotta deve dunque essere definitivamente abbandonata; e, per citare ancora Th. H. Huxley, un autore tra i preferiti dallo scrivente, si può ricordare che questo strenuo sostenitore della teoria darwiniana seppe affermare (1893) che il principio morale è contrario al principio egoistico della lotta per l'esistenza e che la società umana non potrebbe reggersi se tornasse allo stato di natura, di lotta tra individui (e, si può aggiungere oggi, rinunciando all'ottimismo vittoriano e dopo l'esperienza di due guerre mondiali, se persistesse nella lotta tra gruppi). E poi, ci saranno sempre persone impazienti delle parziali verità fornite dalla scienza e capaci di un atto di fede che dia loro la certezza di cose sperate; ed esse vorranno sempre trovare in un filo evolutivo il segno di un Destino e di una Volontà: dagli Invertebrati ai Vertebra- ti, dai Pesci agli Anfibi, ai Rettili, ai Mammiferi, e tra questi ai Primati, agli Ominidi, all'Homo habilis, erectus, sapiens. E saranno numerose: quale che sia il giudizio sull'opera di P. Teilhard de Chardin - lo scrivente condivide il duro giudizio che ne ha dato Monod - non si può ignorare che ha molti lettori. Ci si può ora domandare se è lecito sostenere che chi è convinto della superiorità del metodo scientifico debba augurarsi la dissoluzione di ogni mito irrazionale. La risposta è incerta, perché, anche se sappiamo che oggi per l'uomo molto più che l'evoluzione del patrimonio genico conta l'eredità culturale e la selezione delle idee, e se legittimamente aspiriamo a far conoscere le idee che più per noi valgono e ad affermarle con intransigenza, per persuadere, dobbiamo pure ammettere, magari sottovoce, che il mondo, anche quello delle idee, è bello perché è vario. Un mondo di sola conoscenza ‛vera', scientifica, dove ogni mito fosse bandito, andrebbe meglio? Anche se sembra difficile affermare che andrebbe peggio di quello attuale, nel quale all'irrazionale è offerto tanto spazio, probabilmente nemmeno questo nuovo mondo andrebbe troppo bene; quasi certamente, privo di artisti come la Repubblica di Platone, esso sarebbe mortalmente tedioso.
b) È possiblle un'‛etica evoluzionistica'?
L'uomo dunque è, rispetto ai preominidi suoi antenati, ben altra cosa: in quanto sapiens è ormai anche ethicus, l'unico animale ‛eticizzante'. La sua caratteristica è l'arrivo all'autocoscienza, la libertà del suo spirito. E qui si deve dare per certo che un filosofo di preparazione culturale umanistica, che non ha curiosità per le scienze naturali e ne ha dimenticato il poco studiato nelle scuole preuniversitarie, può giungere a conclusioni validissime studiando l'attività dello spirito umano quale ora è, ignorando come si è fatto. Ma un biologo, che questo non ignora, e sa che dietro a noi c'e una lunga storia evolutiva, una storia naturale, può proporsi il problema di come l'evoluzione abbia portato all'eticità. Su questo punto si è arrivati a una fondamentale concordanza di vedute: l'evoluzione ha portato a un essere capace di stabilire dei valori, che lo hanno guidato nel processo del suo sviluppo psichico e che ancor oggi sono necessari per accrescere il suo patrimonio culturale; e questo antico adattamento si è compiuto sotto il segno della selezione, in un ambiente fin dall'inizio sociale, dove faceva premio la cooperazione. Certamente, fin dai primordi dell'ominazione dovevano esistere gruppi familiari, che poi si sono estesi a clan sempre più ampi: e lo sviluppo del linguaggio sta ad attestare l'importanza della vita di gruppo. In un contesto sociale, una selezione in cui fa premio l'intelligenza diventa una selezione per la cooperazione nell'ambito del gruppo e per l'aggressività di gruppo contro gruppo. Si è arrivati così all'eticità, allo stabilirsi di costumi; e ancor oggi hanno valore positivo collaborazione e altruismo e l'aggressività può essere sublimata, o restare come pesante retaggio in un contesto sociale e razziale dove la lotta ha perduto ormai ogni valore selettivo ed è diventata decisamente dannosa.
Si può pensare a un'evoluzione umana pianificata, perché siamo ormai capaci di intendere l'evoluzione e dirigerla. Se si volessero incrociare per parecchie generazioni uomini e donne, come lo si è fatto con tante piante e animali, si otterrebbero aumenti di statura, di prestanza fisica, di forza muscolare e, perché no, di quozienti di intelligenza, o piuttosto maggiore specializzazione, uomini e donne alfa, beta, fino ad epsilon, come nel Brave new world di Aldous Huxley. Ma questi problemi, per ora, nessuno li pone. Un programma minore, ma attuabile, è quello di diminuire la frequenza di malattie ereditarie, soprattutto degli alleli recessivi che possono essere portati, inavvertiti, in eterozigosi e manifestati all'unione di due portatori. Tutti sono d'accordo sull'opportunità di blande misure eugeniche, di consultori prematrimoniali: un uomo ha il diritto e l'obbligo di sapere che se sposa la sorella di un emofilico, ogni figlio che questa gli darà avrà una probabilità su otto di essere un maschio emofilico; poi potrà esercitare liberamente la sua scelta. In alcuni paesi è obbligatoria, per legge, la sterilizzazione degli affetti da gravi tare genetiche dominanti: è già molto e di più non si può fare. Non si può negare per legge la procreazione alla sorella di un emofilico e poi, quando si comincia con una legislazione eugenica, si apre la strada all'apartheid, e di là al genocidio il passo è breve.
Vogliamo infine affermare che è impossibile credere ad una lontana ingegneria biologica dell'uomo. Per ora è ben remota la possibilità prevista da H. Muller di una banca degli spermi, dove si coltivino frammenti testicolari prelevati da uomini eminenti, per la fecondazione artificiale di donne che volessero aver figli di razza pregiata. E sono nient'altro che farneticazioni le ipotesi sulla possibilità di sintesi in vitro di DNA genico da inserire, portato da un virus innocuo, nel genoma di una persona, per curarne un errore metabolico, o addirittura per farne un fuoriclasse. Si dirà che tante fantasie si sono avverate, il volo di Icaro, il viaggio di Astolfo sulla Luna. Ma è certo che questi sogni di genetica molecolare applicata saranno ancora tali per tutta la durata, supplementi compresi, di questa enciclopedia: molto probabilmente anche per una Enciclopedia del XXI secolo.
Più serio chiedersi se sia possibile un'etica evoluzionistica, intesa come possibilità di trovare in alcuni aspetti della evoluzione organica un'indicazione per stabilire dei valori morali. Bisogna subito ammettere che i fenomeni di evoluzione di altri animali ci possono suggerire ben poco, perché, ripetiamo, l'uomo è un'altra cosa: per lui contano, semmai, i fenomeni di ominazione e la possibilità di un'ulteriore evoluzione culturale. Ricorderemo un solo autore, Julian Huxley, che in molte linee filetiche avverte un progresso, inteso come aumentata complessità di organizzazione per un migliore adattamento all'ambiente; e aggiunge che per l'uomo il progresso evolutivo deve essere visto essenzialmente come acquisto di maggiori capacità di sentire, conoscere, volere e comprendere. Per l'uomo saranno quindi progressi quelli che porteranno a un maggiore affermarsi delle capacità individuali e a un'ulteriore evoluzione sociale, che permetta questa affermazione, sempre più completa, per un numero sempre maggiore di individui. E questa è quasi una verità lapalissiana.
Dobbiamo ancora ripetere che da diecine di migliaia di anni il progresso del volume cerebrale e dell'intelligenza creativa è stato trascurabile. Gli uomini del Magdaleniano erano ormai sapienti e probabilmente capaci di stabilire valori morali quanto lo siamo ora noi. Ma certamente ci sono stati dei progressi nei contenuti etici, perché le idee sono selezionate e il patrimonio culturale è in continua evoluzione. Agamennone scelse liberamente di sacrificare la figlia Ifigenia, per ottenere dagli Dei venti favorevoli per portare i suoi uomini ad Ilio. Egli seguì - e secondo Clitennestra con troppo zelo - i suggerimenti della propria coscienza; ma oggi la vita delle figlie degli ammiragli è più sicura, perché ci si attiene piuttosto ai bollettini meteorologici. E questo ci dice che la conoscenza scientifica porta non a una maggiore libertà, ma ad una migliore libera scelta.
Tante persone, impeccabilmente morali, per millenni hanno accettato - almeno per gli altri, era più comodo - la condizione di schiavi: in America milioni di bianchi, buonissimi cristiani, un secolo fa ricorsero alla guerra per mantenere questa condizione umana. Ed ecco l'ultima verità lapalissiana: non è permesso rinunciare ad una scelta responsabile e l'uomo deve stabilire dei valori, ma la selezione delle idee può modificarne il contenuto. Nessuno, almeno nel mondo occidentale, crede più alla santità delle guerre di religione; molti, in posizioni favorevoli, ancora credono buone alcune forme di schiavitù attenuata, ma nessuno ha ormai il coraggio di dirlo; e questa modifica dei valori etici è altrettanto necessaria - per non fare uomini simili a sepolcri imbiancati, che accettino a fior di labbra cose alle quali non credono, e per accrescere dignità alla condizione umana - quanto è necessaria nell'evoluzione organica una certa plasticità per far fronte alle modifiche ambientali e sopravvivere. Questo può esser detto da un biologo; ed un filosofo idealista avrebbe espresso su per giù la stessa idea, insistendo più sul primo termine ed affermando che per l'uomo verità e bene sono concetti eterni e che poco conta il loro divenire empirico. Ed i biologi ribatteranno che la loro posizione è più aperta.
bibliografia
Crick, F. H. C., Of molecules and man, Seattle 1966 (tr. it.: Uomini e molecole, Bologna 1970).
Dickerson, R. E., Geis, I., The structure and action of proteins, New York 1969.
Dobzhansky, Th., The biology of ultimate concern, New York 1967.
Huxley, Th. H., Huxley, J., Evolution and ethics, 1893-1943, London 1947.
Jacob F., La logique du vivant. Une histoire de l'hérédité, Paris 1970 (tr. it.; La logica del vivente, Torino 1971).
Monod, J., Le hasard et la nécessité. Essai sur la philosophie naturelle de la biologie moderne, Paris 1970 (tr. it.: Il caso e la necessità. Saggio sulla filosofia naturale della biologia contemporanea, Milano 19725).
Montalenti, G., Evoluzione, Torino 1965.
Montalenti, G., Introduzione alla genetica, Torino 1971.
Padoa, E., Biologia generale, Torino 19683.
Padoa, E., Storia della vita sulla Terra, Milano 19712.
Perutz, M. F., Kendrew, J. C., Watson, H. C., Structure and function of haemoglobin. II. Some relations between polypeptide chain configuration and amino acid sequence, in ‟Journal of molecular biology'', 1965, XIII, pp. 669-678.
Petit, C., Prevost, G., Génétique et évolution, Paris 1967 (tr. it.: Genetica ed evoluzione, Milano 1971).
Simpson G. G., This view of life. The world of an evolutionist, New York 1964.
Simpson, G. G., Biology and man, New York 1969.
Simpson, G. G., Beck, W. S., Life. An introduction to biology, New York 19652.
Waddington, C. H., The ethical animal, London 1960.
Watson, J. D., Molecular biology of the gene, New York 19702 (tr. it.: Biologia molecolare del gene, Bologna 19722).
Biologia molecolare di Eraldo Antonini
sommario: 1. Definizione e significato della biologia molecolare. 2. Aspetti teorici della biologia molecolare. 3. Relazioni tra la biologia molecolare e altre discipline. 4. Evoluzione e contenuto attuale della biologia molecolare. □ Bibliografia.
1. Definizione e significato della biologia molecolare
Con l'espressione ‛biologia molecolare' si indica quella parte della scienza che si propone di studiare e interpretare a livello molecolare i fenomeni biologici, prendendo in considerazione la struttura, le proprietà e le reazioni delle molecole chimiche che compongono gli organismi viventi.
Questa nuova branca della biologia, pur discendendo direttamente dalle scienze biologiche classiche, dalla chimica e soprattutto dalla chimica biologica, simboleggia un indirizzo realmente nuovo, malgrado il suo contenuto possa variare con lo sviluppo della materia. Nella prospettiva più ampia, essa comprende infatti la biofisica, la biochimica, la genetica; è quindi una scienza composita che attua il suo programma nel tentativo di spiegare la funzione biologica con la struttura molecolare e le sue modificazioni. Nell'ambito di tale disciplina, sono stati raggiunti negli ultimi vent'anni quei risultati che hanno dato un volto nuovo a tutta la biologia: la determinazione della struttura e della funzione delle grosse molecole che stanno alla base dei fenomeni vitali, quali le proteine e gli acidi nucleici. È stato infatti possibile stabilire qual è la disposizione delle varie migliaia di atomi che le compongono e quali sono le forze che li tengono insieme, nonché i rapporti tra la struttura di queste macromolecole e la loro specifica funzione biologica. Si è individuato in che modo gli enzimi controllano le reazioni chimiche che avvengono nella materia vivente; si è riusciti a capire in che modo il messaggio genetico viene trasmesso attraverso successive generazioni e a individuare la base chimica delle mutazioni.
È sempre difficile stabilire quando nasce una scienza nuova o un nuovo indirizzo scientifico: nel caso della biologia molecolare, la sua genesi va ricondotta alla esigenza, ormai matura dopo la seconda guerra mondiale, di guardare alla biologia in termini quantitativi, secondo le ormai stabilite leggi della chimica e della fisica. La biologia molecolare corrisponde al tentativo di unificare tutti i fenomeni biologici nell'ambito di leggi generali di valore universale. Tale unificazione, è ormai ovvio, può e deve avvenire soprattutto a livello molecolare, e questo rappresenta la naturale evoluzione e in un certo senso il superamento dell'idea cellulare, che, sebbene su basi molto più empiriche, rappresenta il primo tentativo di dare ai fenomeni biologici una base comune.
L'espressione ‛biologia molecolare' sembra aver avuto origine in relazione alle indagini strutturali sulle macromolecole biologiche condotte da cristallografi inglesi rappresentati, intorno al 1930-1940, soprattutto da W. T. Astbury e J. D. Bernai.
Sembra che a usare per primo il termine, ufficialmente, sia stato Astbury nel 1939. Astbury stesso in una famosa ‛Harvey lecture' del 1950, intitolata Avventure in biologia molecolare, così dice a proposito dell'espressione: ‟Il termine ‛biologia molecolare' sembra ora essere diventato di uso piuttosto comune e io sono contento di ciò poiché, sebbene sia improbabile che io lo abbia inventato, a me piace e da tempo ho cercato di propagarlo. Esso implica non tanto una tecnica ma un indirizzo, un indirizzo dal punto di vista delle cosiddette scienze di base con l'idea dominante di ricercare, al di sotto delle manifestazioni su larga scala della biologia classica, il corrispondente piano molecolare. La biologia molecolare si occupa particolarmente delle forme delle molecole biologiche e dell'evoluzione e ramificazione di quelle forme nell'ascesa a livelli di organizzazione sempre più alti. Essa è, in modo predominante, tridimensionale e strutturale - il che non significa, però, che è solo un raffinamento della morfologia. Essa deve necessariamente indagare allo stesso stempo nella genesi e nella funzione".
È stato notato che all'inizio vi erano due gruppi di biologi molecolari: gli ‛informazionisti' e gli ‛strutturisti', che lavoravano indipendentemente gli uni dagli altri, con pochi contatti tra di loro. Gli informazionisti, rappresentati soprattuto dal ‛gruppo del fago' di M. Delbrück e S. E. Luna, si occupavano soprattutto del problema della genetica dei microrganismi e, in modo speciale, della natura della informazione genetica; gli strutturisti, rappresentati specialmente da Astbury, Bernal e loro allievi, erano soprattutto interessati alla definizione della struttura tridimensionale delle macromolecole biologiche, in particolare delle proteine, mediante la cristallografia con raggi X. Le due scuole trovarono sempre più terreno comune con la scoperta del ruolo degli acidi nucleici (v. acidi nucleici) nella trasmissione dell'informazione genetica e nel meccanismo della sintesi proteica; rimane però ancora oggi, nell'ambito della biologia molecolare, un certo dualismo tra gli scienziati che si occupano d'informazione genetica e quelli che lavorano sulla struttura e la funzione delle proteine. Il dualismo si riflette soprattutto nel diverso uso di strumenti tecnici e concettuali da parte dei due gruppi: il primo risente di più dell'eredità della genetica classica e tie usa le tecniche; il secondo fa più uso dei metodi strutturali, della termodinamica e della cinetica chimica, subendo maggiormente l'influsso della biochimica.
2. Aspetti teorici della biologia molecolare
La biologia molecolare rappresenta una nuova branca della scienza (anche per le implicazioni teoriche che ne costituiscono l'essenza dal punto di vista filosofico), alla cui base è la cosciente ammissione che ogni fenomeno biologico può essere analizzato e interpretato in dettaglio nell'ambito delle proprietà chimiche e fisiche delle molecole che compongono la materia vivente. In altre parole, a parte la complessità delle strutture chimiche proprie dei sistemi biologici, la biologia molecolare parte dal presupposto che i fenomeni vitali siano completamente spiegabili secondo le leggi generali derivabili dalla materia non vivente, senza che sia necessario invocare altre leggi speciali per la materia vivente. Questo aspetto concettuale, proprio della biologia molecolare, implica perciò il rifiuto assoluto e definitivo di assunzioni di tipo vitalistico. In biologia il rifiuto del vitalismo è stato per molto tempo solo un atto di fede basato sulla convinzione che non fosse necessario l'intervento di fattori speciali, estranei al mondo fisico, per lo sviluppo e il mantenimento della vita. Attualmente, lo studio quantitativo dei fenomeni biologici apporta continuamente nuovi elementi a sostegno della tesi antivitalistica. Questo è per ora soprattutto evidente nell'ambito di sistemi biologici più semplici, quelli che possono essere definiti esattamente in termini di struttura e funzione. La correlazione tra struttura e funzione, infatti, presuppone la possibilità di associare particolari reattività chimiche a situazioni strutturali ben definite secondo modelli derivati dallo studio di composti semplici non biologici.
Il rifiuto di tesi vitalistiche non implica, però, che non si riconoscano attributi specifici alla materia vivente, che la distinguono anche qualitativamente dal mondo non vivente; anzi, la ricerca di questi attributi e il loro significato rappresenta un altro degli obiettivi della biologia molecolare.
Un altro concetto filosofico importante che nasce dalla biologia molecolare è stato chiamato ‛teleonomia biologica'. Si tratta dell'ammissione che negli esseri viventi le parti (le molecole costitutive) sono integrate nel disegno, nella struttura e nella funzione dell'intero organismo, e le loro proprietà sono correlate alle esigenze di vita di esso.
A livello molecolare si può quindi individuare un preciso determinismo delle parti che costituiscono gli esseri viventi, alle quali sono legate le forme e le funzioni macroscopiche delle specie, la conservazione dell'individuo e la sua riproduzione.
Il fatto di risultare da un piano di organizzazione molecolare, cioè di essere costruito e di riprodursi in modo autonomo, sulla base delle proprietà e delle interazioni delle molecole costituenti, rappresenta un attributo specifico di ogni essere vivente che lo distingue, qualitativamente, da ogni altro oggetto in natura.
Per quanto riguarda i problemi dell'origine della vita e dell'evoluzione, nell'ambito della biologia molecolare essi hanno trovato la possibilità di essere studiati e interpretati razionalmente. Il problema dell'origine della vita può essere ricondotto a quello di una auto-organizzazione della materia a livello molecolare, in condizioni ambientali speciali (v. biofisica; v. vita, origine della). Teorie di autoorganizzazione possono essere costruite, in modo rigoroso, sulle basi di principî fisici noti; queste teorie permettono di proporre modelli molecolari precursori di organismi Viventi e possono spiegare razionalmente i fenomeni generali della selezione e dell'evoluzione biologica (v. evoluzione).
La possibilità di formazione, nelle condizioni prevalenti dell'era primordiale, delle matrici molecolari dei sistemi prebiologici, cioè di composti precursori di proteine e acidi nucleici, è stata ormai dimostrata. È inoltre possibile, dallo studio di differenze e similarità a livello molecolare tra le diverse specie attuali, con processi di estrapolazione all'indietro, ricostruire l'identità chimica di organismi ancestrali. L'evoluzione viene considerata nell'ambito della biologia molecolare come un processo che deriva da e- venti a carico di molecole di acido desossiribonucleico. Essendo stabilito che l'informazione genetica è immagazzinata nella sequenza di basi dell'acido desossiribonucleico, l'evoluzione di ogni specie vivente può essere ricondotta a una serie di variazioni nell'ordine e nella lunghezza di questa sequenza. Le variazioni avvengono apparentemente a caso, per cause varie, e rimangono o scompaiono nei fenotipi per effetto della selezione naturale.
3. Relazioni tra la biologia molecolare e altre discipline
È chiaro che in generale lo studio attuale della biologia non può in ogni caso prescindere dalle proprietà chimiche e chimico-fisiche, cioè molecolari, delle sostanze che compongono gli esseri viventi. Da questo punto di vista si potrebbe acquisire che la biologia si identifica attualmente in gran parte con la biologia molecolare, in quanto tutte le aree della biologia sono o saranno in un prossimo futuro studiabili a livello molecolare. Inoltre potrebbe sembrare che non vi sia una reale distinzione tra biologia molecolare e biochimica. La biologia molecolare rappresenta la naturale evoluzione della biochimica, tendente, più che a identificare le reazioni chimiche che avvengono negli organismi (metabolismo), a individuare le origini di queste reazioni in termini di proprietà strutturali e funzionali delle due preminenti classi di macromolecole biologiche, cioè le proteine e gli acidi nucleici.
Sotto questo aspetto appare predominante in biologia molecolare, rispetto alla biochimica classica, lo studio dell'aspetto macromolecolare dei sistemi biologici e ciò spiega anche perché lo sviluppo della biologia molecolare sia stato così strettamente correlato a quello dei metodi, soprattutto cristallografici, per l'identificazione della struttura tridimensionale e delle proprietà in genere delle macromolecole.
Per quanto riguarda la genetica, la biologia molecolare ha permesso il superamento del concetto puramente ‛biologico' di gene con l'identificazione ‛chimica' di esso in segmenti di acido nucleico. I due fondamentali problemi della genetica, cioè i meccanismi della replicazione e dell'azione dei geni, possono quindi essere ricondotti alle proprietà chimiche degli acidi nucleici.
Alcune aree della biologia potrebbero sembrare al di fuori del contesto della biologia molecolare, ad esempio la biologia a livello dell'intero organismo o la biologia delle popolazioni; eppure anche in queste aree si prevede che sarà decisivo in futuro l'indirizzo molecolare.
In conclusione si può ammettere che la biologia molecolare si è innestata sulle diverse branche della biologia classica come un nuovo indirizzo di studio quantitativo dei fenomeni biologici in cui predomina l'analisi a livello macromolecolare; il fenomeno biologico viene così ricondotto alle proprietà delle molecole biologiche più importanti, cioè proteine e acidi nucleici.
4. Evoluzione e contenuto attuale della biologia molecolare
La nascita della biologia molecolare in parte riflette la storia delle discipline classiche di cui la biologia molecolare rappresenta l'evoluzione. È chiaro che tentativi d'identificazione del piano molecolare che sta alla base degli organismi biologici si possono già individuare fin dalla nascita della chimica fisiologica o chimica biologica. Il concetto, però, che le strutture biologiche sono organizzate su una base molecolare, concetto che sembra ovvio attualmente, si è fatto strada con fatica e lentamente durante i primi quarant'anni di questo secolo, ed è stato completamente accettato solo con il riconoscimento delle proteine e degli acidi nucleici come entità chimiche ben definite. Si arrivò così al superamento del concetto, quasi mistico, di ‛protoplasma', con cui si indicava il contenuto intracellulare come una miscela poco definibile chimicamente.
La nascita, l'evoluzione e il contenuto della biologia molecolare coincidono quindi in gran parte con il processo di identificazione della struttura e funzione delle proteine e degli acidi nucleici. (Per un'ampia trattazione di questi argomenti v. acidi nucleici e proteine).
bibliografia
Antonini, E., Brunori, M., Hemoglobin and myoglobin in their reactions with ligands, in Frontiers of biology (a cura di A. Neuberger e E. L. Tatum), vol. XXI, Amsterdam 1971.
Cohen, S. S., The molecular biology of viruses, New York 1967.
Edsall, J. T., Proteins as macromolecules. An Essay on the development of the macromolecule concept and some of its vicissitudes, in ‟Archives of biochemistry and biophysics", 1962, suppl. 1, pp. 12-20.
Edsall, J. T., Recent advances in protein research, in New perspectives in biology (a cura di M. Sela), vol. IV, Amsterdam 1964, pp. 1-17.
Edsall, J. T., History of biochemistry and molecular biology, in ‟Science'', 1970, CLXX, pp. 349-351.
Eigen, M., Selforganization of matter and the evolution of biological macromolecules, in ‟Die Naturwissenschaften", 1971, LVIII, pp. 465-523.
Fraenkel-Conrat, H., The chemistry and biology of viruses, New York 1969.
Hayes, W., The molecular biology of viruses, in XVIII Symposium of the Society for General Microbiology, Cambridge 1968, pp. 1-4.
Hess, E. L., Origins of molecular biology, in ‟Science", 1970, CLXVIII, pp. 664-669.
Jukes, T. H., Molecules and evolution, New York 1968.
Kendrew, J. C., Concluding remarks, in New perspectives in biology (a cura di M. Sela), vol. IV, Amsterdam 1964, pp. 278-280.
Monod, J., Le hasard et la nécessité. Essai sur la philosophie naturelle de la biologie moderne, Paris 1970 (tr. it.: Il caso e la necessità. Saggio sulla filosofia naturale della biologia contemporanea, Milano 19725).
Smith, C. U. M., Molecular biology. A structural approach, London 1968 (tr. it.: Biologia molecolare, Milano 1971).
Stanley, W. M., Molecular basis of virology, New York 1968.
Steiner, R. F., The chemical foundations of molecular biology, Princeton 1965.
Watson, J. D., Crick, F. H. C., Molecular structure of nucleic acids, in ‟Nature", 1953, CLXXI, pp. 737 ss.
Yčas, M., The biological code, in Frontiers of biology (a cura di A. Neuberger e E. L. Tatum), vol. XII, Amsterdam 1969.