
Biosintesi
Biosintesi
sommario: 1. Metaboliti primari e secondari. 2. Biogenesi e biosintesi: a) polimerizzazione ‛lineare' dell'acetato; b) ‛ramificazione' dell'acetato: via dell'acido mevalonico; c) via dell'acido scichimico; d) metilazione e alogenazione. 3. Analisi biogenetica: a) acido gibberellico; b) griseofulvina; c) acido lisergico; d) novobiocina; e) tetraciclina; f) penicillina. □ Bibliografia.
Metaboliti primari e secondari
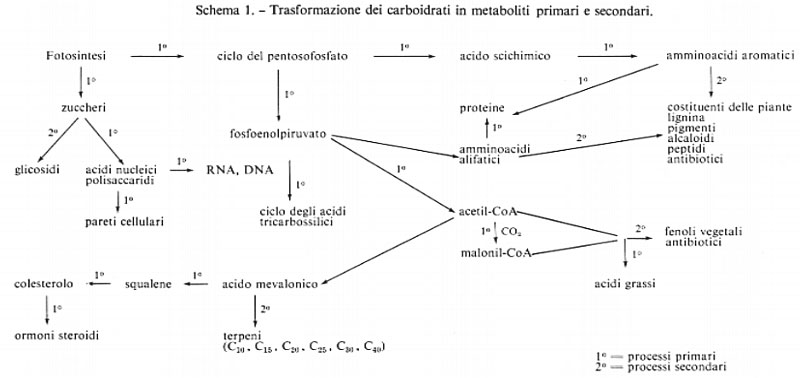
Le molecole di origine naturale che esamineremo vengono sintetizzate nelle piante, nei microrganismi e nei Mammiferi a partire dai carboidrati che, a loro volta, provengono dalla fissazione fotosintetica del diossido di carbonio attraverso il ciclo dei pentosofosfati. Il termine ‛biosintesi' è del tutto generale; esso descrive i processi chimici che stanno alla base della formazione dei metaboliti ‛primari': polisaccaridi, lipidi, proteine, vitamine, ormoni e materiale genetico (RNA e DNA).
In questo contesto tratteremo la formazione di composti, a peso molecolare generalmente inferiore a 2.000, che vengono indicati come metaboliti ‛secondari'. Molti di questi composti (oggi se ne conoscono circa 12.000) costituiscono importanti ‛unità strutturali' per molecole polimeriche più complesse e molti sono, già di per se stessi, di grande interesse. Sebbene ai composti organici di tale gruppo sia stato dato l'attributo di ‛secondari', queste sostanze mostrano spesso una spiccata attività biologica ‛er es., alcaloidi, antibiotici) e il loro studio ha impegnato i chimici organici per più di un secolo. Recentemente, a partire dal 1950 circa, la disponibilità di traccianti radioattivi (carbonio-14, trizio) ha reso possibile lo sviluppo di tecniche che permettono di evidenziare come i carboidrati, gli amminoacidi, l'acido acetico ed analoghe ‛unità strutturali' vengano utilizzati dagli esseri viventi per formare strutture molecolari spesso complesse e strane. La loro complessità varia ampiamente dall'etilene (C2H4) alla vitamina B12 (C6H88N14O14PCo); tuttavia oggi, per un gran numero di casi, si conoscono i principî che stanno alla base della loro struttura molecolare. Lo schema 1, sebbene assai semplificato, illustra come i carboidrati vengano utilizzati per la produzione di metaboliti primari e secondari. Per entrambi i gruppi si hanno gli stessi tipi di reazione; quello che li differenzia è essenzialmente il ‛grado' di polimerizzazione.
Biogenesi e biosintesi
Sin dalla fine del sec. XIX i chimici organici hanno cercato di comprendere i meccanismi delle reazioni che, nei vegetali superiori e nei microrganismi, conducono alla vasta gamma dei prodotti naturali utilizzando, quali composti di partenza, i carboidrati e gli amminoacidi. Il processo di scomporre mentalmente una struttura molecolare complessa nelle sue probabili unità strutturali è noto come ‛biogenesi' o ‛analisi biogenetica'.
La dimostrazione sperimentale o conferma di tale processo divenne possibile solamente quando furono accessibili gli isotopi radioattivi, in particolare il 14C e l' 3H, sebbene tra il 1935 e il 1950 si fossero già conseguiti dei progressi utilizzando isotopi non radioattivi, come l'18O, l'15N, l'2H e il 13C, per i quali è necessaria l'analisi mediante spettrometria di massa. Solamente dopo una tale prova sperimentale si può dire di conoscere la ‛biosintesi'.
Le teorie biogenetiche hanno conferito un grande impulso al successivo sviluppo sperimentale e molti di coloro che hanno contribuito maggiormente a questo studio sono riusciti a vedere le loro idee verificate sperimentalmente, cioè è stata scoperta la vera biosintesi che, in molti casi, corrisponde all'analisi teorica proposta.
Polimerizzazione ‛lineare' dell'acetato
Sperimentalmente si è trovato che l'ipotesi biogenetica di una condensazione testa-coda di unità acetato, suggerita per la prima volta da J. N. Collie nell 897 e riconfermata, indipendentemente, da A. J. Birch nel 1952, costituisce il reale processo biosintetico. Nello schema 2 sono riportati i passaggi mediante i quali l'acido acetico viene legato covalentemente come tioestere al coenzima A, poi condensato con successive unità malonato e da qui trasformato, attraverso una via riduttiva, in acidi grassi ovvero, mediante una via non riduttiva, in chetoni ciclici, da cui possono originarsi dei fenoli.
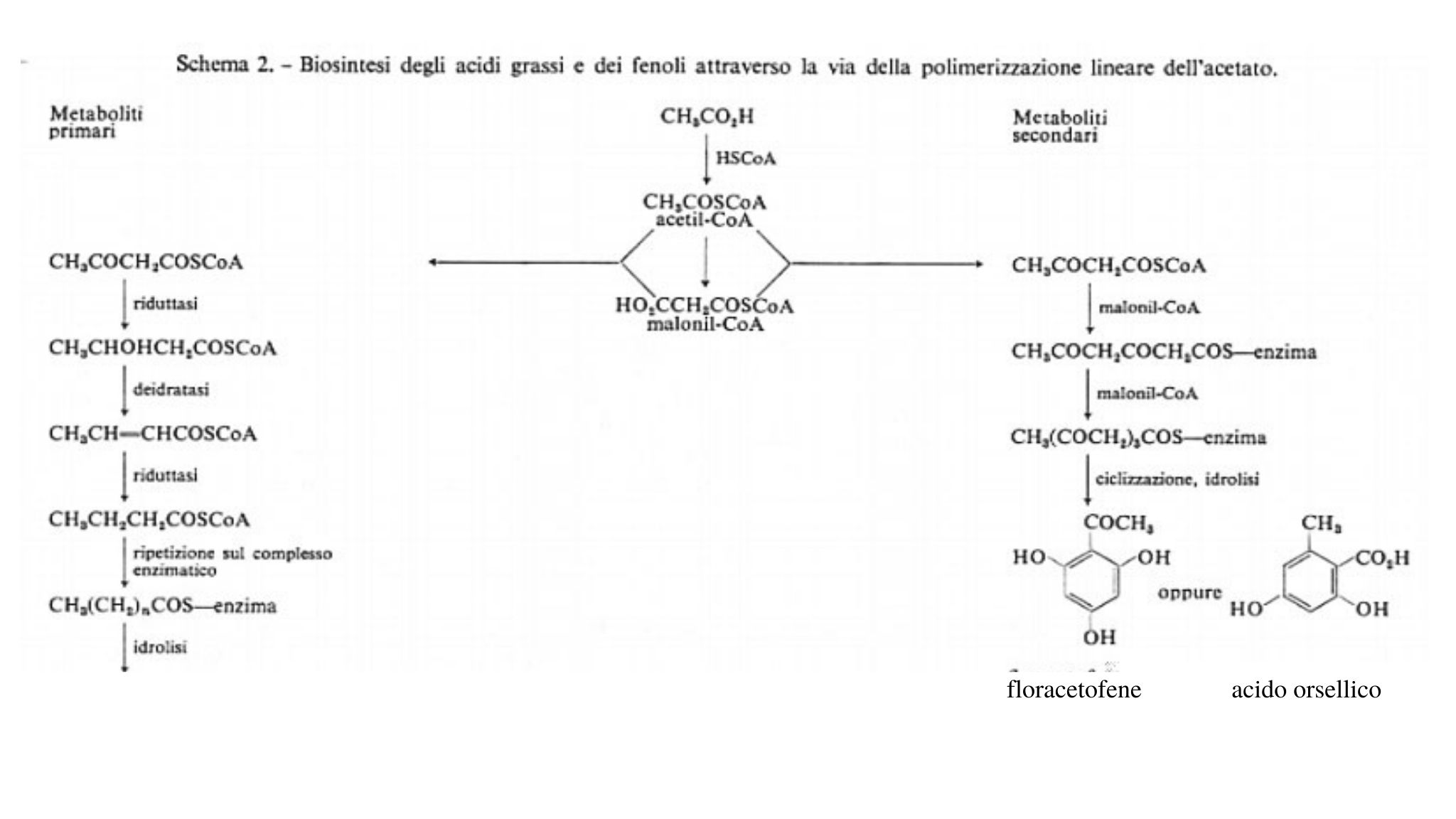
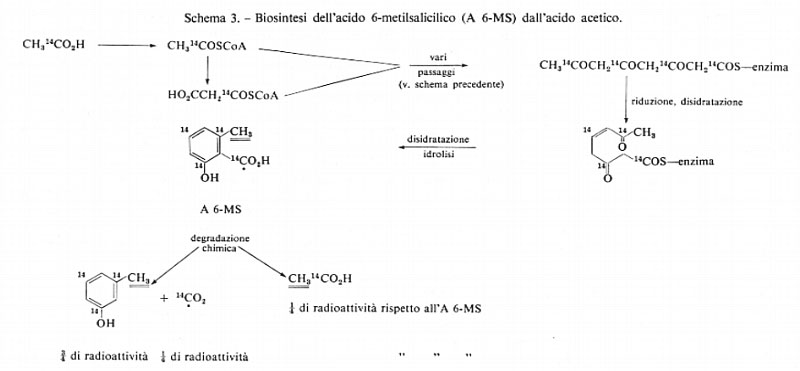
La verifica sperimentale dell'ipotesi dell'acetato' è stata realizzata in batteri (Escherichia coli), lieviti, funghi, vegetali superiori e Mammiferi ed è del tutto generale. Per esempio, la somministrazione di CH314C02H ad un organismo (Penicillium patulum) determina la formazione di acido 6-metilsalicilico marcato, come è mostrato nello schema 3.
La decarbossilazione di questo acido e l'ossidazione della catena laterale metilica forniscono, rispettivamente, diossido di carbonio e acido acetico che presentano una radioattività corrispondente ad 1/4 di quella della molecola di partenza. Contemporaneamente l'organismo produce acidi grassi marcati con 14C, come è rappresentato nei due schemi ora riportati. L'organismo può utilizzare l'acido acetico non soltanto come ‛gruppo di partenza', ma anche, mediante la fissazione di diossido di carbonio, come fonte di acido malonico attivato, secondo lo schema 4.
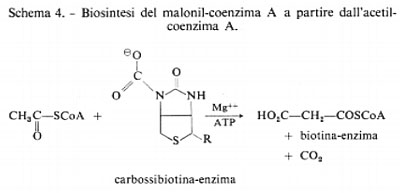
Ognuno degli enzimi responsabili di queste reazioni è stato isolato e ottenuto con un buon grado di purezza. Il lavoro richiesto per la separazione degli enzimi implicati in una intera sequenza di reazioni, ad esempio quella che a partire dall'acido acetico conduce agli acidi grassi e agli steroidi (v. sotto), è molto lungo e complesso. Tuttavia, notevoli progressi sono stati conseguiti in questo campo, in modo particolare dai gruppi di ricerca di F. Lynen (Germania) e K. Bloch (S.U.A.) ai quali, nel 1964, fu assegnato il premio Nobel per la medicina quale riconoscimento per questo interessante lavoro.
‛Ramificazione' dell'acetato: via dell'acido mevalonico
Abbiamo appena descritto la polimerizzazione lineare dell'acido acetico attivato per dare acidi grassi e derivati fenolici. Una seconda grande via metabolica fu prevista dal chimico svizzero (di origine ceca) L. Ružička, il quale notò che molti prodotti naturali sono costituiti (almeno formalmente) da unità isoprenoidi. La ‛regola dell'isoprene' di Ružička è illustrata nello schema 5.
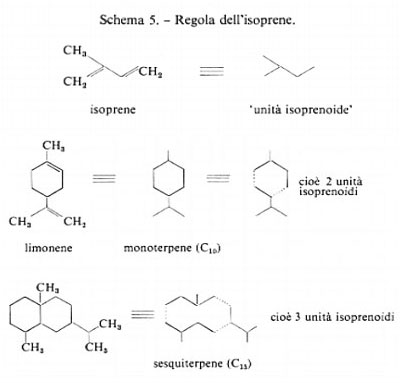
Questa ipotesi biogenetica fu verificata negli anni cinquanta, oltre mezzo secolo dopo la sua formulazione, dalla scoperta, dovuta a K. Folkers, dell'isoprene ‛biologica- mente attivo' o ‛fattore sostitutivo dell'acetato' per i lattobacilli. Era stato trovato che l'acido acetico viene incorporato negli steroidi, come il colesterolo, in modo da far presupporre un qualche precursore ‛ramificato'. Questo composto, che fu infine scoperto nei residui solubili della distillazione alcolica, fu denominato acido mevalonico; nello schema 6 ne è mostrata la sintesi a partire dall'acetato.

È stato sintetizzato acido mevalonico contenente come traccianti, in ogni posizione della molecola, sia il 14C sia l'3H, e il percorso di questa versatile specie monomerica è stato seguito fino alla biosintesi di monoterpeni (composti costituiti da 10 atomi di C), quali il geraniolo (sotto forma di pirofosfato), di sesquiterpeni (per es. il farnesil-pirofosfato) e dello squalene, un idrocarburo triterpenico (v. schema 7, biosintesi degli steroidi a partire dall'AMV). La formazione degli steroidi nell'uomo, negli animali e nei microrganismi segue questo percorso, i cui dettagli meccanicistici e stereochimici sono stati quasi completamente precisati (v. metabolismo: metabolismo dei lipidi).
La trasformazione dello squalene in colesterolo lia inizio con l'epossidazione (mediante O2) di un doppio legame terminale dello squalene e quindi procede mediante una reazione diretta di ciclizzazione, durante la quale nessun protone viene scambiato col mezzo acquoso di reazione, per dare lanosterolo, secondo lo schema 8.
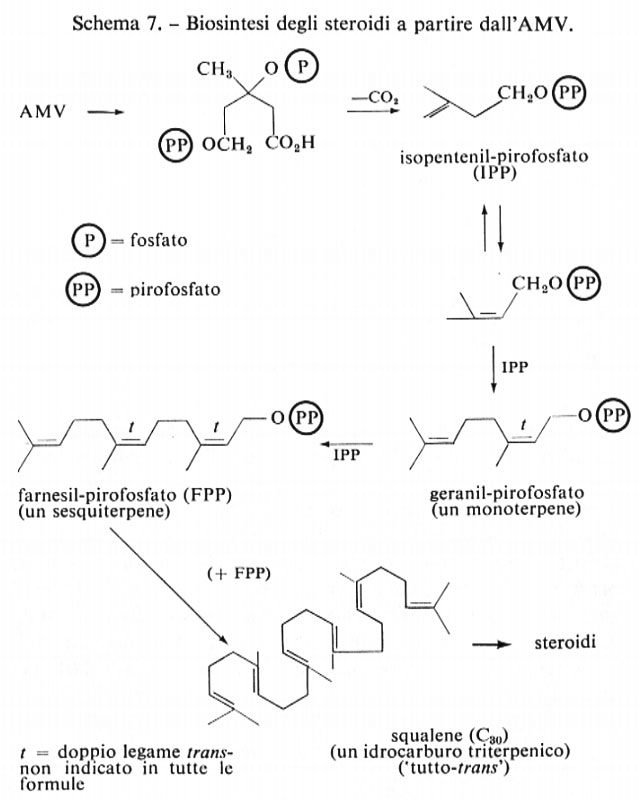
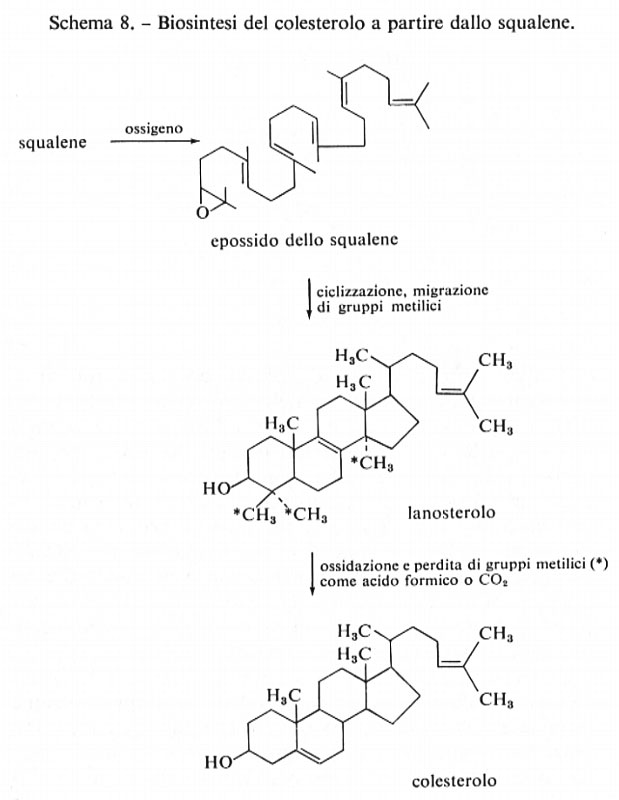
I passaggi finali che portano al colesterolo sono stati quasi completamente chiariti ed implicano la perdita dei gruppi metilici che nello schema sono contrassegnati con un asterisco (*), probabilmente sotto forma di acido formico, e lo spostamento del doppio legame alla posizione 5. Questo schema è operante nella maggior parte dei sistemi studiati, utilizzando sia cellule intatte, sia diverse frazioni surnatanti stabilizzate, ottenute separando materiale cellulare mediante centrifugazione ad elevata velocità (biosintesi in mezzo acellulare).
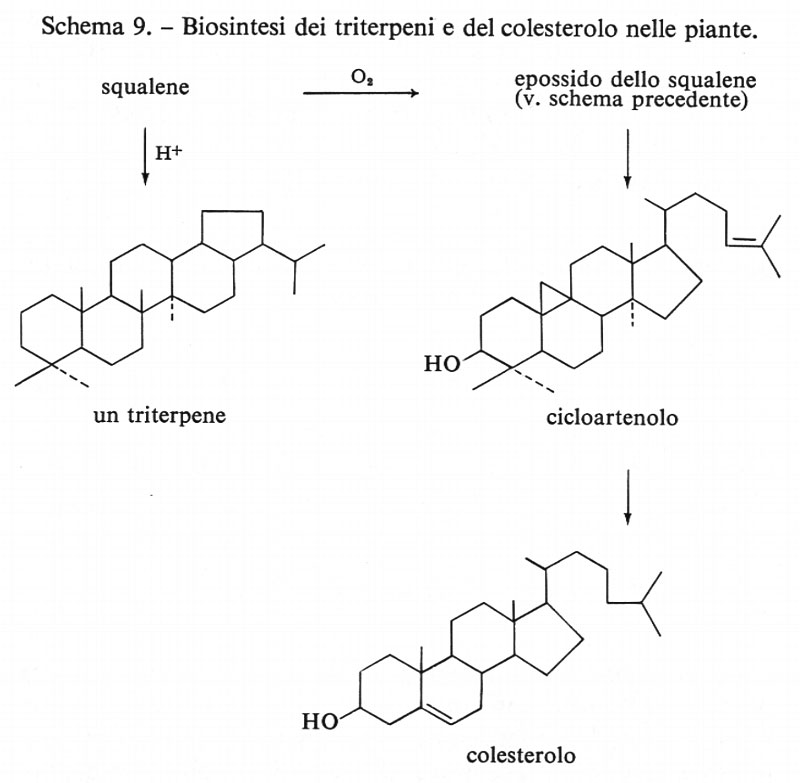
Recentemente sono state scoperte almeno due eccezioni alla formazione del colesterolo secondo questo schema. Negli Insetti, che non hanno gli enzimi necessari per la sintesi del colesterolo, questo composto viene normalmente assunto con la dieta. Anche nei vegetali superiori, attualmente si è propensi a ritenere che il reale precursore del colesterolo non sia il lanosterolo, ma il cicloartenolo (un derivato non comune contenente un anello ciclopropanico), così come è risultato in tutte le piante fino ad ora studiate. Un'altra osservazione generale è che, mentre le piante superiori possono ciclizzare lo squalene in molti modi diversi (v. schema 9), fino a poco tempo fa non erano mai stati riscontrati triterpeni nei Batteri. Di recente è stato invece scoperto che vi si possono trovare, cosicché questo schema di reazioni è possibile anche negli organismi inferiori. Finora non è stata rilevata la presenza, nel metabolismo umano, di triterpeni come quello riportato nello schema suddetto, né sembra che la biochimica dei Mammiferi comporti mai la sintesi di composti aromatici (v. schema 2, biosintesi degli acidi grassi e dei fenoli). D'altra parte, il corpo umano elimina un migliaio di metaboliti urinari. Poiché ne sono stati identificati meno della metà, ci possiamo aspettare ulteriori progressi, nelle biosintesi operate dai Mammiferi, in seguito al riconoscimento di minime quantità di nuovi composti organici.
Via dell'acido scichimico
B. D. Davis e D. B. Sprinson, nell'elaborazione della ‛via dell'acido scichimico', la terza delle grandi vie biosintetiche, descrissero in modo alquanto diverso la biosintesi di un'altra classe di prodotti naturali.
La scoperta di un prodotto intermedio tra i carboidrati e gli amminoacidi aromatici (fenilalanina, tirosina e triptofano) fu possibile impiegando mutanti di Escherichia coli che richiedono questi amminoacidi.
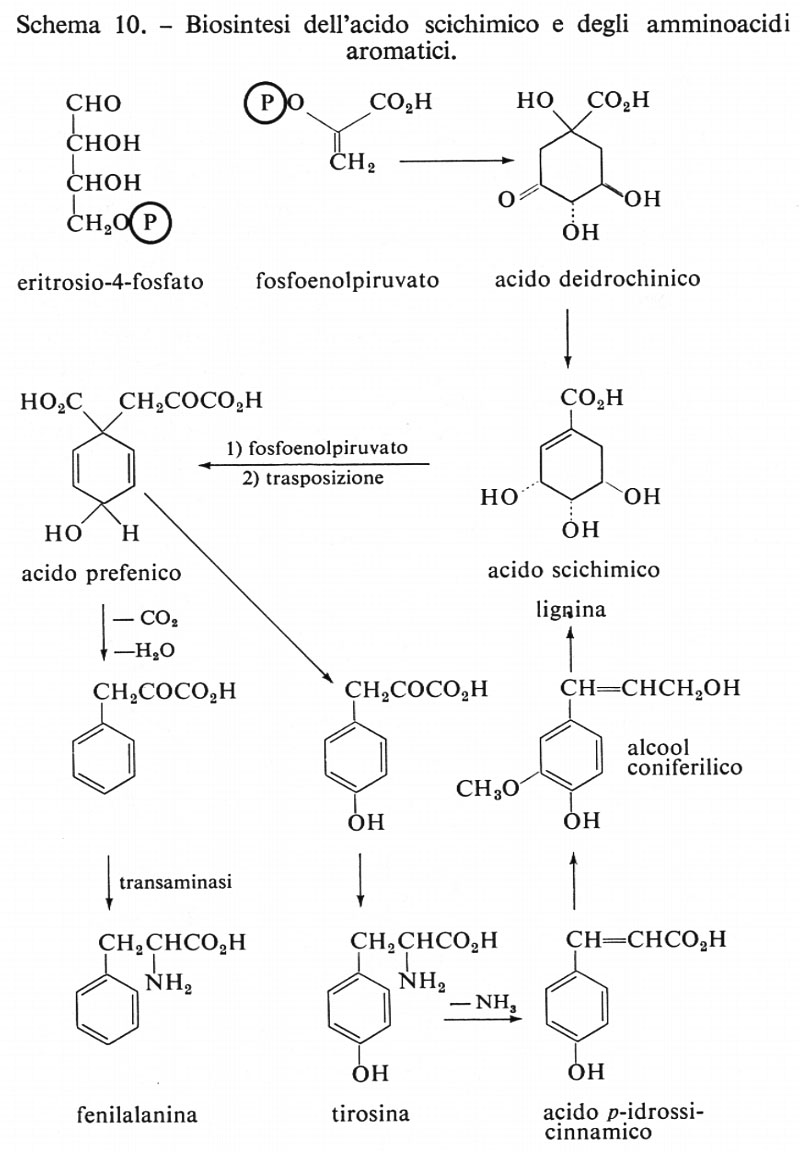
Furono scoperti mutanti nutrizionali che avevano contemporaneamente necessità di diversi acidi aromatici (fenilalanina, tirosina, triptofano, acidi p-amminobenzoico e p-idrossibenzoico). Si scopri che questi mutanti di Escherichia coli crescevano ugualmente bene se veniva loro somministrato acido scichimico (v. schema 10). Altri mutanti venivano bloccati agli ultimi stadi e, mediante lo studio della chimica dei metaboliti da essi isolati, venne infine chiarito lo schema suddetto. Si può vedere che non solo importanti amminoacidi essenziali, quali il triptofano, la fenilalanina e la tirosina, provengono dall'acido scichimico, ma che, per azione di enzimi deaminanti o ‛ammoniaca-liasi', questi composti possono essere trasformati in acido cinnamico e nei suoi derivati, che a loro volta costituiscono le unità strutturali per polimeri quali la lignina, ovvero che combinandosi con chetoni, formano i componenti flavonoidi delle piante e i pigmenti antocianici dei fiori, tanto per citare due esempi comuni.
Metilazione e alogenazione
Gli ultimi due tipi di reazioni biosintetiche sono quelli che avvengono in processi secondari e modificano il modello costruito attraverso le vie dell'acetato, del mevalonato o dello scichimato mediante reazioni di alchilazione all'O e al C, ovvero reazioni di sostituzione elettrofila da parte di alogeni, per esempio Cl+, Br+, 1+. L'S-adenosilmetionina o SAM (v. schema 11), che costituisce una fonte di gruppi metilici elettrofili, è l'agente metilante naturale proveniente dal cosiddetto pool dei C1 (cioè riserva dei frammenti a un atomo di carbonio). Per esempio, l'aggiunta di formiato radioattivo determina frequentemente, ma non sempre, la formazione di metionina marcata e quindi, per mezzo del SAM, possono essere successivamente introdotti in altre molecole gruppi metilici marcati. Spesso i gruppi metilici ‛extra' possono essere scoperti con l'analisi strutturale o biogenetica di un fenolo, come nel caso dell'acido 3-metilorsellico (v. schema 12), che, in base alla regola dell'acetato, contiene un gruppo CH3 ‛extra'. Un caso diverso è quello costituito dall'ormone giovanile degli Insetti che fino a poco tempo fa si riteneva derivasse da una doppia metilazione del farnesolo, come è indicato nella II parte dello schema 12.
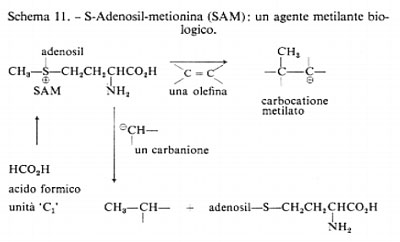
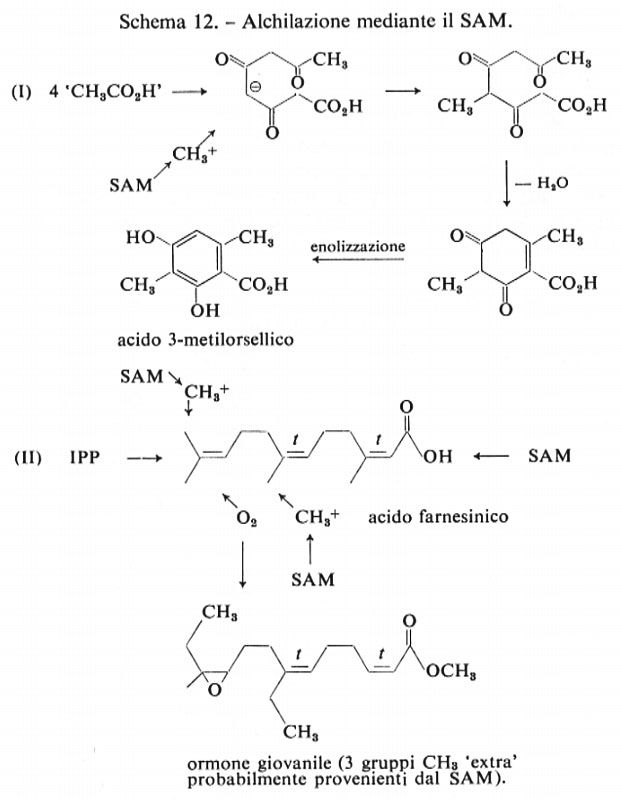
Secondo studi più recenti sembra invece che nella biosintesi dell'ormone giovanile degli Insetti sia implicato l'acido omomevalonico.
Esempi interessanti di alogenazione biologica sono forniti dalla duodotirosina (l'unità strutturale della tiroxina, l'ormone tiroideo) e dalla griseofulvina, un fungicida orale (v. schema 13).
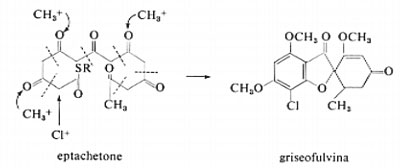
Probabilmente, in questi casi l'alogenuro inorganico viene convertito da un enzima, per esempio l'alogenoperossidasi, nell'alogenocatione Cl+ o 1+ e questa specie effettua la sostituzione elettrofila sull'anello aromatico. È probabile che composti alogenati di questo tipo si ritrovi- no in grande quantità negli organismi marini.
Analisi biogenetica
Dopo aver stabilito quali sono le unità strutturali e le tre principali vie biosintetiche, siamo ora in grado di analizzare parecchi metaboliti complessi dal punto di vista della loro probabile biogenesi. In ognuno dei casi citati è stata studiata anche l'adatta biosintesi, ma ciò è stato possibile forse solo per il 5% del numero totale dei metaboliti conosciuti. Nonostante che i casi studiati siano relativamente pochi, l'analisi biogenetica può essere impiegata per predire la probabile origine di molte classi strutturali chiaramente correlate.
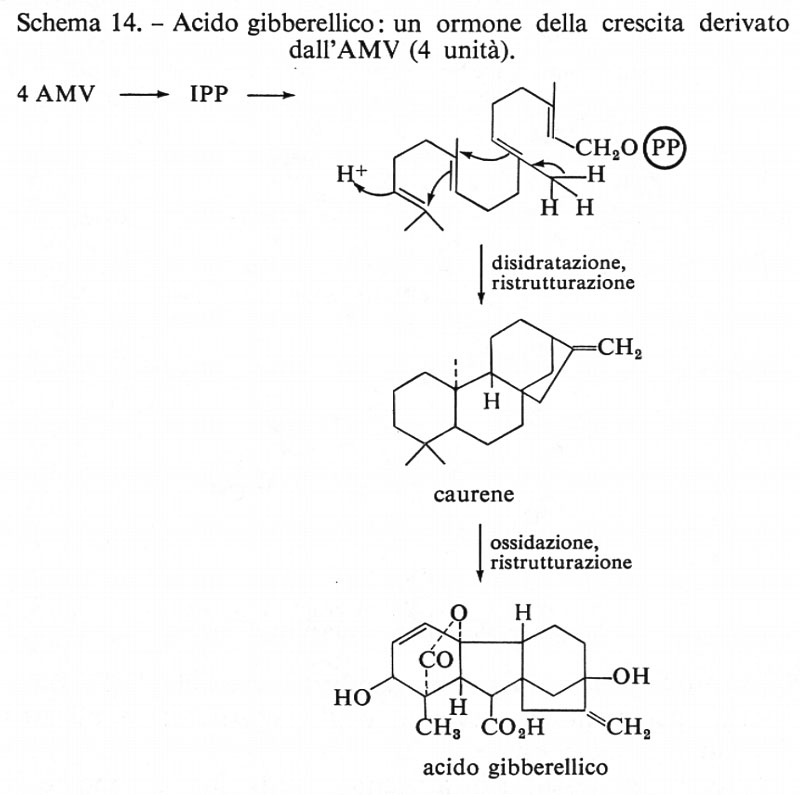
Acido gibberellico
È stato dimostrato che questo interessante ormone dell'accrescimento delle piante proviene dall'acido mevalonico attraverso la via delle unità isoprenoidi. L'acido gibberellico, nonostante la sua struttura complessa, è un derivato diterpenico. Alcuni intermedi della sua biosintesi sono mostrati nello schema 14.
Griseofulvina
Questo antibiotico orale, impiegato nella cura della tigna e del ‛piede d'atleta', è prodotto dalla muffa Penicillium patulum. La sua formazione secondo la via dell' ‛acetato lineare' venne dapprima postulata e quindi comprovata sperimentalmente (v. schema 13, griseofulvina) somministrando alla muffa CH314CO2H (come sale sodico) durante il processo di fermentazione e quindi localizzando gli atomi di 14C per degradazione chimica. La biosintesi è ultimata dalla trasformazione, operata dalla S-adenosilmetionina, di due gruppi idrossilici a carattere fenolico in eteri metilici (OCH3). La molecola subisce anche la clorurazione, sull'anello aromatico, quando nell'ambiente di fermentazione sono presenti ioni cloruro, di origine inorganica. La corrispondente bromogriseofulvina (con Br al posto di Cl) può essere preparata impiegando un mezzo di fermentazione ricco di bromuro di potassio.
I composti biologicamente attivi sopra citati sono esempi del metabolismo dell'acetato lineare e ramificato. Gli esempi successivi illustrano, invece, come si possono combinare varie ‛unità strutturali'.
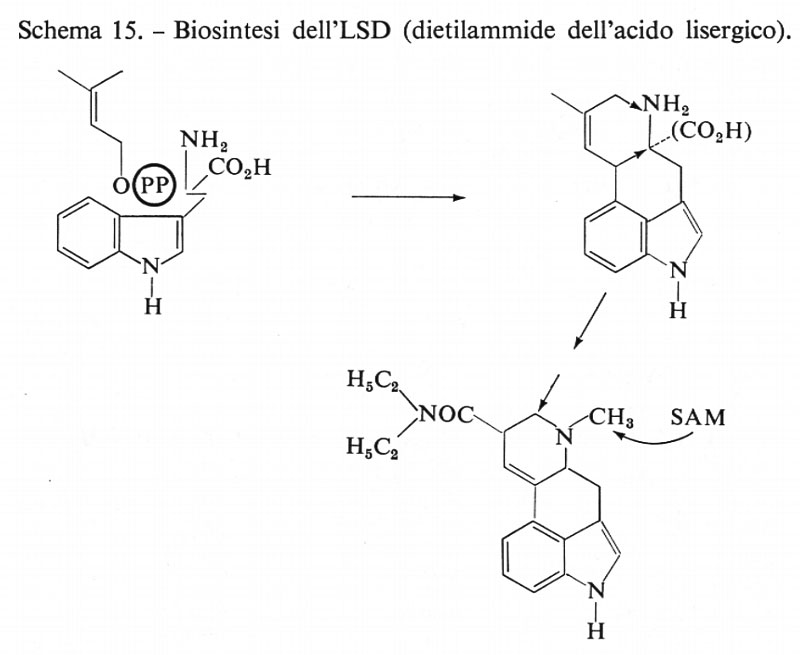
Acido lisergico
La struttura di questo derivato del triptofano si presta all'analisi biogenetica: così si può riconoscere un'unità ramificata C5 (v. schema 15, biosintesi dell'LSD). Infatti, l'acido lisergico e la corrispondente droga LSD (la dietilammide) si producono nella specie Claviceps (fungo della segale cornuta) e in una Convolvulacea (Ipomoea) per combinazione del triptofano con ‛una' unità di isopentenilpirofosfato (a sua volta prodotto dall'acido mevalonico; v. schema 7, biosintesi degli steroidi a partire dall'AMV). Lo schema 15 illustra alcuni processi di ossidazione e di formazione di legami implicati in questa sintesi.
Novobiocina
La struttura di questo antibiotico, che viene prodotto commercialmente dalla fermentazione di Streptomyces sp., è un ottimo esempio di origine biosintetica ‛mista'. Nella novobiocina possono essere riconosciute sette unità biogenetiche formatesi attraverso cinque diverse vie biosintetiche (v. schema 16):
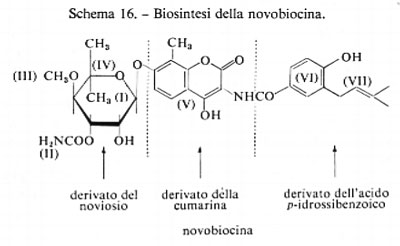
il noviosio (I), che ha la struttura di un glucosio modificato, contenente una funzione ammidica (Il), un gruppo metossilico −OCH3 (III) e gruppi metilici (IV) provenienti dal pool dei C1; l'anello aromatico metilato della cumarina (V), derivata dalla tirosina attraverso l'acido scichimico, che è anche il precursore della parte costituita dal p-idrossibenzoile VI) e, infine, la catena ramificata C5 VII), unita al sistema ciclico (VI) e proveniente dalla sequenza mevalonato-IPP. Nella sintesi della novobiocina è possibile individuare quasi tutti i percorsi biosintetici ad essa interessati.
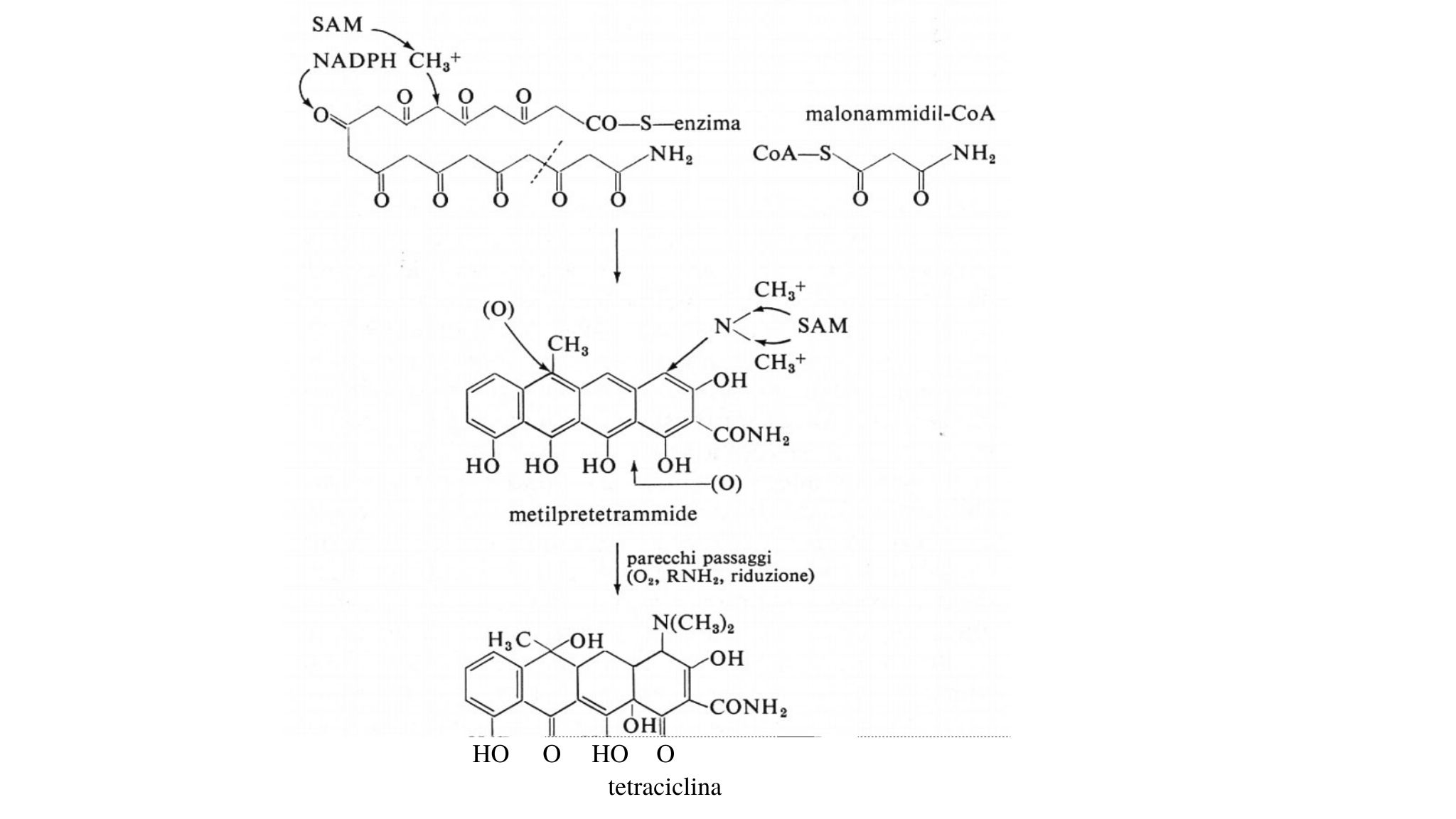
Tetraciclina
Prove effettuate aggiungendo CH314CO2-Na+ al mezzo di coltura di Streptomyces rimosus e studi su mutanti di questo microrganismo hanno ampiamente chiarito i complicati processi di sintesi degli antibiotici del gruppo della tetraciclina per condensazione lineare di unità di acido malonico. Il più probabile punto di partenza o unità capostipite per questo processo è il malonammidil-coenzima A, come indicato nello schema 17.
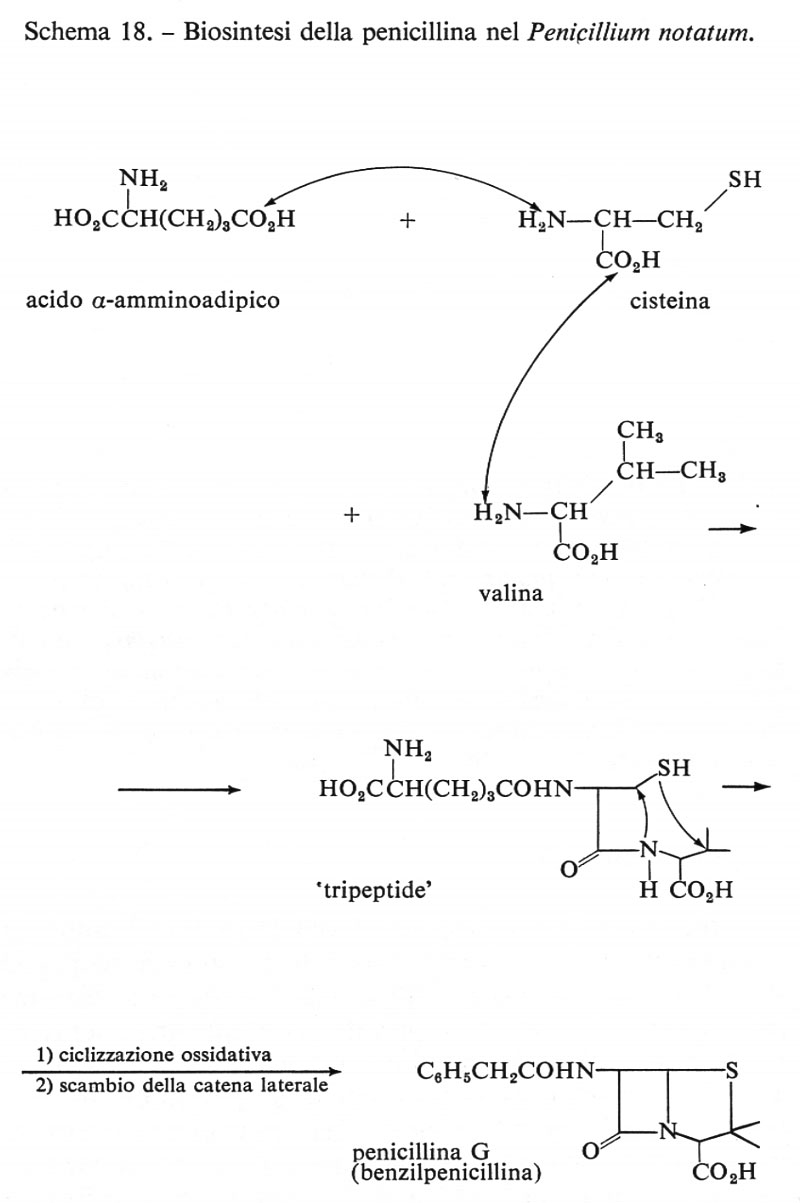
Penicillina
Il nostro ultimo esempio è costituito dal più importante degli antibiotici, la cui ‛presunta' biosintesi è quella mostrata nello schema seguente. Gli amminoacidi costitutivi, l'α-amminoadipico, la cisteina e la valina, si combinano per formare un ‛tripeptide'.
La ciclizzazione, attraverso un meccanismo non ancora conosciuto, produce la penicillina N che, per trasformazione in acido 6-amminopenicillanico, fornisce questo versatile precursore che, unendosi a una delle molte catene laterali che trova nel liquido di fermentazione, dà luogo alle varie penicilline, rappresentate nello schema 18 dalla penicillina G (benzilpenicillina).
Bibliografia
Bu'Lock, J. D.,The biosynthesis of natural products, London 1965.
Lehninger, A. L., Biochemistry, New York 1970.
Richards, J. H., Hendrickson, J. B., Biosynthesis of terpenes, steroids and acetogenins, New York 1964.