
Botanica
Botanica
Introduzione
La botanica è la scienza che studia le piante. L'unità di questa scienza sta nell'oggetto trattato e non negli scopi che si prefigge chi ad essa si dedica, nè nel modo di affrontare lo studio o nei metodi impiegati. È dunque una scienza di notevole complessità, che abbraccia varie branche. Questo articolo considera la botanica nel senso più ampio del termine, trattando tutta la sfera di quei campi che vengono definiti talvolta come scienze delle piante. La parte principale è dedicata a una panoramica delle varie branche e in essa viene dato particolare rilievo allo stato attuale delle conoscenze e ai principali progressi che sono stati compiuti all'incirca negli ultimi cento anni; aspetti più remoti da un punto di vista storico sono stati affrontati solo quando avevano un particolare significato per il progresso della conoscenza o per l'evoluzione delle idee. La parte finale considera l'importanza dello studio delle piante per l'umanità e comprende una discussione sul probabile sviluppo della botanica nel prossimo futuro, alla luce delle tecniche ora disponibili per l'indagine biologica.
La botanica e le sue branche
Per poter meglio valutare le finalità della botanica, è necessaria una certa conoscenza dei tipi di organismi classificati come piante. Le piante verdi hanno un tipo di nutrizione, noto come autotrofico, basato sull'utilizzazione diretta dell'energia luminosa e sull'assunzione di molecole sempllci dall'ambiente. Questa modalltà di nutrizione non basta, tuttavia, a definire una pianta, poiché alcuni organismi per altro simili a piante - fra cui un gruppo importante, quello dei Funghi - possiedono un tipo di nutrizione saprofitico o parassitario. Altre caratteristiche abitualmente presenti nelle piante sono la forma sessile o provvista di radici, e la mancanza di quegli attributi che sono caratteristici degli animali superiori, cioè un sistema nervoso e gli organi di senso ad esso collegati. Tutte queste caratteristiche, considerate insieme, permettono di distinguere sufficientemente bene i membri superiori del regno vegetale da quelli del regno animale, ma in certi gruppi di organismi - soprattutto organismi di piccole dimensioni e considerati primitivi in senso evoluzionistico per la loro semplicità di forma e di metabolismo - non può essere effettuata una netta divisione in ‛piante' e ‛animali'. Una classificazione di questi organismi è, tuttavia, assolutamente necessaria, e la loro assegnazione ai campi di studio della botanica o della zoologia si basa in genere sulla pratica tradizionale o su arbitrarie considerazioni di convenienza.
Nella tab. I è riportata una classificazione degli organismi inclusi generalmente nel regno vegetale. La disposizione adottata è tradizionale, con una suddivisione che ha il merito di essere familiare e di poter essere messa in relazione con quella di altri sistemi. I Batteri, considerati talvolta come parte del regno vegetale, non sono compresi nella tab. I e non vengono trattati in questo articolo.
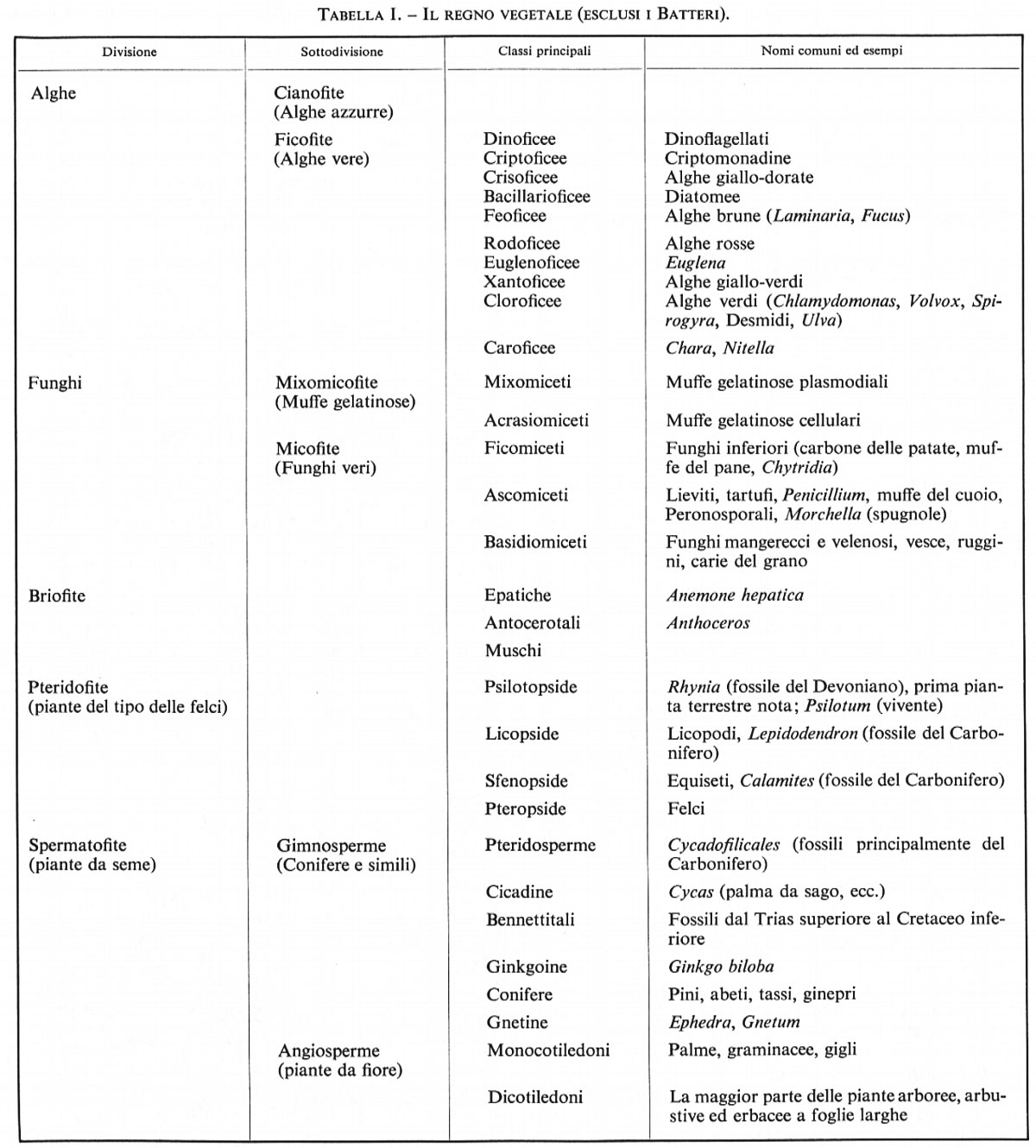
La stima circa il numero delle specie di piante presenti oggi sulla Terra varia notevolmente. Gli studiosi più competenti concordano sul fatto che vi siano tra 250.000 e 300.000 specie di piante da seme. Ai rimanenti gruppi appartengono, probabilmente, altre 100.000-150.000 specie, con un totale compreso fra 350.000 e 450.000.
La botanica è suddivisa in diverse branche. Tra le principali vi sono quelle di seguito elencate.
Tassonomia
La tassonomia è lo studio dei principi e dei metodi pratici usati per la classificazione delle piante; comprende la sistematica, che può essere definita come l'uso di denominare e classificare le piante in un sistema generale. La classificazione si ottiene raggruppando le piante nelle varie categorie di un sistema secondo le loro somiglianze e le loro differenze.
Il sistema correntemente in uso, con i suoi schemi di nomenclatura, si basa essenzialmente sui principi stabiliti da Linneo; è stato codificato internazionalmente, e le regole concordate sono raccolte nel Codice internazionale di nomenclatura botanica. Il codice viene riesaminato periodicamente dai Congressi internazionali di botanica. Solamente nel caso che la descrizione e la denominazione di una pianta soddisfino le richieste del codice, il nome ha validità internazionale. Alla nomenclatura delle piante coltivate si applicano regole un po' differenti rispetto a quelle concordate per le specie selvatiche.
Fino dai primi tempi della botanica scientifica i caratteri usati per la classificazione delle piante (conosciuti come criteri tassonomici) sono stati tratti essenzialmente dalla struttura esterna, o morfologia. Lo studio della struttura interna, o anatomia, ha contribuito fornendo per un lungo periodo informazioni importanti per la classificazione. Attualmente la morfologia e l'anatomia esistono come branche della botanica indipendenti dalla tassonomia, e sono studiate per molte altre ragioni, diverse da quella di fornire criteri tassonomici.
Durante gli ultimi due secoli, gli studiosi di sistematica tentarono uno studio completo del regno vegetale. Sebbene rimanga ancora molto da fare, esiste attualmente una vasta raccolta di conoscenze sulla flora mondiale delle piante vascolari (Pteridofite e Spermatofite); i gruppi inferiori, in particolare i Funghi, sono meno conosciuti. I risultati degli studi tassonomici sono pubblicati in trattati sistematici, monografie e articoli di ricerca. Per alcuni gruppi si conservano esaurienti cataloghi di nomi di specie, come l'Index Kewensis, e collezioni di consultazione sono raccolte nei grandi erbari, i più ricchi dei quali si trovano nei Kew Gardens di Londra (circa 4.500.000 esemplari), a Lemngrado (circa 4.000.000 di esemplari) e a Parigi (circa 3.750.000 esemplari).
Negli ultimi anni ci sono stati molti sviluppi, sia teorici che pratici, nella tassonomia vegetale. Il consolidamento dell'idea di evoluzione organica ha avuto importanti implicazioni; ma nel periodo immediatamente postdarwiniano gli studiosi di sistematica hanno riconosciuto le idee evoluzionistiche soprattutto nel trattare i raggruppamenti superiori del sistema tassonomico, raggruppamenti che venivano riordinati in maniere differenti per adattarli a mano a mano ai concetti in via di sviluppo di correlazione e di posizione nella scala evolutiva. Sono state invece per lo più ignorate le implicazioni della teoria darwiniana riguardo all'applicazione del sistema in sé, e in particolare riguardo al significato dell'idea di non fissità delle specie per l'applicazione della sistematica descrittiva. Oggi, il sistema di Linneo è visto come un buon espediente per codificare le variazioni degli organismi, ma spesso troppo rigido per adattarsi a tutto ciò che riguarda l'evoluzione delle popolazioni vegetali. I progressi nella comprensione dell'origine e della natura delle variazioni (progressi conseguiti dalla genetica, dalla citologia e dalla fisiologia dello sviluppo) hanno portato a una maggiore consapevolezza di queste limitazioni della sistematica classica, e hanno favorito lo sviluppo di un metodo di studio sperimentale chiamato biosistematica o tassonomia sperimentale. Nonostante che l'applicazione della tassonomia sperimentale porti spesso a risultati in contrasto con la sistematica classica, le discrepanze non sono state così gravi da richiedere una ricostruzione di vasta portata del sistema linneano. La maggior parte delle nuove osservazioni riguarda variazioni a livello delle specie di Linneo o a livello inferiore, in relazione ad argomenti come l'interfertilità delle popolazioni e le variazioni nel numero di cromosomi. Tali dati non possono essere facilmente assimilati nella tassonomia per quel che riguarda la nomenclatura; per utilizzarli, sono stati proposti molti metodi che comportano vari tipi di classificazione per scopi speciali.
Indipendentemente dal contributo dei metodi sperimentali, la gamma dell'informazione strutturale e chimica disponibile per la tassonomia si è enormemente allargata nell'ultimo cinquantennio con l'introduzione di svariate nuove tecniche di studio. Il miglioramento dei mezzi ottici ed elettronici per studiare la morfologia e l'anatomia (v. sotto, § b) ha aumentato in maniera sostanziale l'ampiezza dei criteri disponibili e gli effetti di ciò non si sono ancora sentiti appieno, dal momento che i nuovi metodi sono stati finora applicati a pochissimi gruppi. Dati chimici comparativi di molte specie sono ora raccolti per parecchi dei gruppi principali; fin dal 1950 la chemiotassonomia si è sviluppata come campo di ricerca e i suoi risultati sono esposti in molte pubblicazioni specifiche. Alcuni dati chimici sono di particolare interesse perché sono in relazione con le molecole polimeriche che recano l'informazione nella sequenza dei monomeri. Le componenti proteiche di riserva dei semi, dei tuberi, ecc. hanno grande valore tassonomico per famiglie, generi e perfino specie. Si possono tracciare anche relazioni tra le specie tramite rapporti di somiglianza del materiale genetico, l'acido desossiribonucleico, saggiando in provetta l'affinità molecolare.
La tassonomia classica era legata in gran parte alla valutazione intuitiva delle variazioni da parte di esperti studiosi di sistematica; la descrizione era spesso inadeguata, imprecisa e di rado accuratamente quantificata. In molti gruppi studiati a fondo, la tassonomia si basa attualmente soprattutto sulla biometria. Valutazioni di somiglianza o di differenza si basano su metodi statistici, compresa l'analisi delle variazioni plurime, che lascia poco spazio al giudizio soggettivo. Questo metodo di studio è conosciuto come tassonomia quantitativa o numerica. Nelle applicazioni più complesse l'uso del calcolatore è essenziale, e sono ora disponibili programmi per molti usi, compresa la creazione di ‛chiavi' per l'identificazione.
A causa della grande quantità di organismi diversi da descrivere e da classificare, e della quantità di fatti - molti di tipo radicalmente nuovo - ora disponibili che devono essere introdotti nel sistema, nella tassonomia vegetale ci sono oggi seri problemi di manipolazione dei dati. L'aumentata meccanizzazione li sta ora risolvendo; sistemi memorizzati dai calcolatori per immagazzinare e recuperare i dati tassonomici sono già in uso, almeno in parte, e se ne può senz'altro prevedere una rapida espansione per il prossimo futuro.
Strettamente legato alla tassonomia è lo studio della storia evoluzionistica delle piante, o filogenesi. Due sono i concetti base della teoria di Darwin: l'uno che gli organismi viventi si sono evoluti da una stirpe ancestrale comune, e l'altro che, in generale, questa stirpe ha dato origine a diverse discendenze, divenute progressivamente più adatte ai particolari habitat o ai particolari modi di vita sotto la pressione della selezione naturale. Le implicazioni per la tassonomia sono evidenti: un sistema ‛naturale' di classificazione - vale a dire basato sulla maggior correlazione possibile di attributi - deve riflettere in qualche modo relazioni evolutive, dal momento che gli organismi che divergono meno avranno maggiori caratteristiche comuni, e si troveranno quindi più vicini l'un l'altro nel sistema di classificazione. È interessante notare che questo metodo è stato facilmente frainteso, dal momento che ancora oggi si sente dire spesso che la classificazione naturale è ‛basata' sulla filogenesi. Alcune classificazioni, elaborate mettendo insieme organismi con il maggior numero di tratti comuni, possono essere ‛interpretate' filogeneticamente, cioè come una forma di rappresentazione degli eventi evolutivi, ma ovviamente non possono ‛esprimere' di per sé la filogenesi a causa della complessità dei processi evolutivi, inclusi i tassi di variazione nelle differenti popolazioni e la possibilità che si verifichino fenomeni di convergenza.
Morfologia e anatomia
La morfologia e l'anatomia trattano degli attributi esterni e interni della forma delle piante a livelli di organizzazione superiori a quello della cellula, mentre la citologia si riferisce generalmente allo studio delle strutture interne alla cellula. Vi sono, naturalmente, molte ragioni per studiare la struttura delle piante e molte tecniche che possono essere applicate per fornire osservazioni con diverso grado di approfondimento. I tratti morfologici e anatomici sono stati studiati, durante il XVIII secolo, soprattutto per il loro valore tassonomico, nonostante il rilievo che aveva avuto, in precedenza, la funzione nei lavori di M. Malpighi, N. Grew e S. Hales. In seguito, durante il periodo di J. M. Schleiden e K. von Nageli, furono sempre più adottati parametri di giudizio basati sullo sviluppo, e durante gli ultimi cinquanta anni è stato posto in sempre maggior rilievo il significato fisiologico della struttura delle piante, in relazione sia alla funzione interna sia all'adattamento ecologico.
È evidente l'importanza degli studi morfologici comparativi per la classificazione. La forma esterna può essere valutata a occhio nudo, con la lente d'ingrandimento e con il microscopio composto. Le nuove tecniche di microscopia ottica, come il contrasto di fase e di interferenza, forniscono ora i mezzi per scoprire i particolari di strutture trasparenti o traslucide altrimenti non discernibili. Tuttavia, i limiti di risoluzione dei metodi ottici sono determinati dalla lunghezza d'onda della luce: la migliore risoluzione è dell'ordine di 0,2 micrometri. Recentemente, questi limiti sono stati ampiamente superati con l'introduzione del microscopio elettronico a scansione, capace di dare una risoluzione di 10 nanometri. Con la nascita di questo settore di morfologia submicroscopica si sono già ottenute nuove e sorprendenti informazioni sulla struttura delle superfici e delle appendici delle superfici delle piante e delle loro spore. Parallelamente, nello studio dell'anatomia delle piante i limiti del microscopio ottico sono stati superati con l'uso del microscopio elettronico a trasmissione, che offre una risoluzione di 1,0 nanometri o anche più. Ciò ha reso possibile lo studio dettagliato delle strutture molecolari delle pareti cellulari.
È improbabile che l'assimilazione dei nuovi dati morfologici e anatomici nella tassonomia richieda qualche radicale mutamento concettuale, tuttavia determina un problema pratico a causa della quantità di informazioni che attualmente ci sono fornite. Per le piante superiori, questo problema si sta risolvendo con pubblicazioni di sistematica specifica, che trattano particolari rami dell'argomento.
Lo studio della morfologia e dell'anatomia comprende gli adattamenti strutturali nei fenomeni della riproduzione delle piante: campo in cui non esistono limiti tra queste discipline e la citologia. Dai tempi delle fondamentali ricerche di W. Hofmeister sulla sessualità e sulla riproduzione nelle piante, nel 1851, le conoscenze si sono enormemente accresciute, particolarmente per le piante inferiori, le Crittogame di Linneo. Vi sono stati grandi cambiamenti nella trattazione sistematica di alcuni di questi gruppi, da quando si è tenuto conto della micromorfologia e della biologia della riproduzione; ognuno dei maggiori gruppi di Crittogame ha ora la sua parte di trattati sistematici, monografie e cataloghi.
I sistemi riproduttivi delle Gimnosperme e delle Angiosperme sono stati studiati nei dettagli durante il XX secolo, ma, l'influenza delle nuove conoscenze sulla tassonomia è stata per questi gruppi minore che per le Crittogame. Per le piante da fiore, il termine ‛embriologia' è generalmente esteso a comprendere anche i gametofiti maschili e femminili (granuli pollinici e sacchi embrionali). L'aumentata mole di conoscenze è basata sulla ricerca microscopica e sulla sperimentazione; allo stesso tempo sono stati condotti studi comparativi diretti a sistematizzare l'informazione. I vari tipi di sviluppo embrionale sono stati sistematicamente trattati in monografie speciali, ed è in corso di preparazione un completo trattato mondiale dei pollini e delle spore.
I sistemi riproduttivi delle piante comportano maggiori varietà rispetto a quelli degli animali, e inoltre presentano un maggior numero di aberrazioni riproduttive. Dal punto di vista della struttura e della fisiologia, i vari tipi di anomalie riproduttive rappresentano modificazioni o deviazioni dei modelli di sviluppo che, in base ai'risultati di studi comparativi, costituiscono il comportamento modale di un particolare gruppo. Possono essere classificati in questo modo i diversi tipi di sviluppo del sacco embrionale, come pure le differenti forme di poliembrionia (formazione di più embrioni) osservate nelle Conifere, e le varie forme di apomissi (sistemi riproduttivi in cui è eliminato il processo sessuale) trovate in molte famiglie di piante da fiore.
I problemi associati allo sviluppo delle piante ed all'origine della forma (morfogenesi) sono al centro di molte ricerche attuali. Qui si fondono i contributi della morfologia, dell'anatomia, della citologia, della fisiologia, della biochimica e della genetica; l'insieme delle conoscenze che ne deriva viene spesso denominato fisiologia dello sviluppo; tale argomento sarà trattato in seguito (v. sotto, § f). I libri di testo di morfologia e anatomia vegetale, basati in passato sulla struttura degli organi maturi e caratterizzati da una forte tendenza allo studio comparativo, tendono ora a mettere in rilievo gli aspetti che riguardano lo sviluppo. Per le piante superiori ciò ha significato una sempre maggiore valorizzazione dell'importanza funzionale dei punti di accrescimento del fusto e della radice, parti della pianta che possono essere considerate come permanentemente embriogeniche. Teorie dell'organizzazione dei punti di accrescimento possono essere basate sulla suddivisione in aree morfologiche, sulla funzione nello sviluppo o sulle caratteristiche biochimiche dei vari tessuti. Molte nuove tecniche, compresi i metodi di osservazione come i procedimenti di accelerazione dell'immagine cinematografica, sono attualmente in uso per questo tipo di studio.
Oggi, molti studiosi di morfologia e di anatomia si occupano intensamente degli aspetti fisiologici delle loro ricerche. Scienziati dei primi anni del XX secolo, e in particolar modo G. Haberlandt, cercarono di interpretare le caratteristiche strutturali delle piante alla luce della loro funzione e aprirono il campo dell'anatomia fisiologica vegetale. Attualmente, molti dei principali problemi ancora insoluti di fisiologia vegetale, e particolarmente quelli relativi al trasporto delle sostanze, richiedono raffinati studi strutturali. Di particolare interesse è il lavoro degli ultimi dieci anni sulla microstruttura del floema. L'adattamento ecologico delle piante è sempre espresso, almeno in parte, nella morfologia, dove i più recenti metodi di osservazione stanno fornendo nuovi importanti dati. Attualmente è in grande sviluppo quel campo della biofisica delle superfici vegetali che considera gli effetti della forma delle foglie e della micromorfologia delle superfici sugli equilibri energetici e sugli scambi di gas. Studi di questo tipo sono di grande significato per la fisiologia pratica delle coltivazioni, come anche per l'ecologia.
Citologia
Il principio che le cellule formano le unità strutturali degli organismi viventi fu stabilito durante la prima metà del XIX secolo, e da allora le cellule sono state studiate sotto vari aspetti. Per diversi anni l'attenzione è stata focalizzata sul nucleo cellulare (scoperto nel 1831 da R. Brown) e in particolare sul comportamento dei cromosomi durante la divisione cellulare e la riproduzione. Gli ultimi decenni del secolo segnarono la scoperta del fenomeno della mitosi (divisione della cellula somatica) e della meiosi (divisione riduttiva che precede la formazione delle cellule sessuali o gameti) grazie al contributo fondamentale degli studiosi di citologia vegetale, in particolare di E. Strasburger. In seguito, le piante hanno costituito il materiale di studio più adatto per le ricerche sulla poliploidia e sulla citologia degli ibridi. Studi riguardanti l'effetto della perdita o dell'aggiunta di singoli cromosomi sulla struttura e la fisiologia delle Angiosperme hanno costituito la base per ricerche successive su fenomeni simili negli animali, compreso l'uomo.
Per quel che riguarda le tecniche di osservazione in citologia, il principale progresso compiuto negli ultimi anni è stato l'avvento del microscopio elettronico a trasmissione. Questo ha esteso il campo degli studi strutturali a livelli molecolari, rendendo possibile la visualizzazione dell'acido desossiribonucleico (DNA), la sostanza costituente i cromosomi che trasporta l'informazione genetica. Il microscopio elettronico, tuttavia, non ha chiarito bene l'architettura del cromosoma stesso, del quale si conosce la complessa organizzazione a elica non facilmente interpretabile a causa delle sezioni sottili che devono essere usate negli strumenti ordinari. La disponibilità di microscopi elettronici con un'accelerazione di 1.000.000 di volt rende oggi possibile esaminare con un alto potere risolutivo cromosomi interi e chiarire così particolari della loro organizzazione e del processo di divisione.
L'analisi della struttura e delle funzioni del citoplasma, cioè del protoplasma della cellula che si trova al di fuori del nucleo, non compì progressi notevoli nel XIX secolo. Gli studi compiuti nei primi anni del XX secolo si basavano soprattutto sull'uso di metodi fisici per l'analisi di proprietà quali la viscosità, e i trattati pubblicati verso il 1950 potevano fornire poco più che considerazioni speculative circa la natura della sostanza fondamentale. Più tardi si poterono ottenere maggiori conoscenze sui corpi particolati (organelli) del citoplasma, che si dimostrarono centri metabolici specializzati: il mitocondrio, interessato al processo della respirazione, e il doroplasto, presente nei tessuti delle piante verdi, implicato nella fotosintesi; ma la loro ultrastruttura rimase in gran parte sconosciuta. La situazione fu rivoluzionata dalla microscopia elettronica e dallo sviluppo di una tecnica per ottenere sezioni ultrasottili. Fra le scoperte più importanti che siano state realizzate vi è quella del sistema di membrane del citoplasma, noto come reticolo endoplasmatico, e delle particelle ad esso associate composte da proteine e acido ribonucleico (RNA), i ribosomi, che partecipano alla sintesi proteica sia nelle cellule vegetali sia in quelle animali. L'identità di struttura fra cellule vegetali e animali si estende anche a organelli quali il mitocondrio e l'apparato di Golgi (un organulo cellulare che compie varie sintesi, compresa quella dei polisaccaridi nelle cellule vegetali). Si conosce oggi abbastanza dettagliatamente la struttura dell'organello presente esclusivamente nelle cellule vegetali, il cloroplasto, e le varie teorie generalmente accettate cercano di collegare le sue caratteristiche strutturali con la sua funzione nell'assorbimento dell'energia luminosa e nelle reazioni fondamentali della fotosintesi. È stato dimostrato che cloroplasti e mitocondri contengono materiale ereditario sotto forma di DNA, e che sono dotati di una certa indipendenza genetica dalla cellula in cui si trovano (v. cellula: Fisiologia della cellula).
Un particolare componente strutturale del citoplasma, il microtubulo, svolge un'importante funzione nel movimento all'interno della cellula, benché non si sappia con precisione in quale maniera. I microtubuli formano il fuso mitotico, struttura collegata al movimento dei cromosomi durante la divisione cellulare. Sembra che nelle piante essi determinino alcuni aspetti della struttura molecolare delle pareti cellulari, e che possano rappresentare uno dei fattori mediante i quali il protoplasma vivente dirige la morfogenesi della parete. Nella biologia cellulare delle piante, che ha rapporti stretti con la biochimica, gli studi speri- mentali dipendono oggi in gran parte dall'utilizzazione di nuove tecniche che sono state messe a punto negli ultimi due decenni. Le vie anaboliche e cataboliche possono essere studiate mediante l'uso di molecole ‛traccianti' contenenti isotopi radioattivi, e i vari processi metabolici possono essere associati a particolari componenti cellulari con il procedimento dell'autoradiografia, che consente di localizzare con grande precisione all'interno della cellula i siti radioattivi, grazie all'effetto determinato su un'emulsione fotografica posta a contatto con il preparato: in questa maniera fu dimostrato per la prima volta in cellule vegetali il rapporto esistente fra sintesi di DNA e divisione cellulare. Tali metodi sono complementari alle tecniche citochimiche, che permettono la localizzazione di sostanze e funzioni all'interno della cellula mediante l'utilizzazione di reazioni ultramicrochimiche in grado di dar luogo a prodotti di reazione distinguibili col microscopio ottico, oppure, con un potere risolutivo molto maggiore, col microscopio elettronico. Studi di questo tipo permettono, insieme ai dati ottenibili mediante analisi biochimiche, di correlare la complessità morfologica della cellula alle varie funzioni metaboliche. Il risultato principale è stata la dimostrazione di quanto importante sia, nelle funzioni della vita, la compartimentazione della cellula; enzimi particolari e sistemi enzimatici sono associati con organelli di diversi tipi, e i movimenti dei substrati e i prodotti di reazione sono strettamente controllati. In tale controllo, le membrane delle cellule svolgono una funzione fondamentale. Per questo importante motivo, si studia oggi con particolare attenzione la struttura molecolare, la permeabilità e le altre proprietà delle membrane delle cellule vegetali. È stato ipotizzato che gli ormoni interessati al controllo della crescita, dello sviluppo e del movimento possano compiere la loro funzione agendo sulle membrane.
Genetica
Oggi la genetica interessa l'intera biologia, fornendo concetti unificatori e principi ormai indispensabili per campi apparentemente assai diversi, come la sistematica e la biochimica cellulare. Questa scienza è stata fondata da Mendel più di un secolo fa con uno studio sui caratteri ereditari del pisello coltivato, e una parte piuttosto notevole della genetica sperimentale continua a basarsi su materiale vegetale, dai Batteri e Funghi filamentosi alle piante superiori.
Vi è uno stretto legame fra la genetica e la citologia, e la combinazione delle due discipline viene talvolta chiamata citogenetica. La teoria cromosomica dell'eredità, secondo la quale i fattori mendeliani (chiamati ora geni) sono trasportati nel cromosoma, venne dimostrata nel primo decennio del XX secolo (v. genetica).
Un grandissimo successo scientifico degli anni intorno alla metà di questo secolo consistette nell'identificazione del gene con un filamento di acido desossiribonucleico (DNA) e nella scoperta dei meccanismi mediante i quali ha luogo la duplicazione del DNA e viene utilizzata l'informazione codificata, trasportata dalle molecole, per le funzioni vitali delle cellule e degli organismi. Ciò implica, sostanzialmente, la trascrizione dell'informazione contenuta nel DNA in un composto intermedio, l'acido ribonucleico del tipo noto come RNA ‛messaggero' (mRNA), e la successiva traduzione di questa informazione nella struttura di numerose proteine specifiche attive nel metabolismo e nell'accrescimento. In questo tipo di ricerca, generalmente, il materiale sperimentale era costituito da muffe e Batteri. Era naturale, quindi, che la prima dimostrazione completa dell'intero processo per un organismo multicellulare, utilizzando componenti isolati e non dipendenti da cellule viventi, dovesse essere realizzata a partire dal DNA dei nuclei della pianta di pisello, il materiale degli esperimenti di Mendel, ottenendo come prodotto terminale la proteina di deposito caratteristica del seme di pisello (v. acidi nucleici; v. proteine).
Nello studio dell'azione del gene, la genetica è correlata con la biochimica e la fisiologia; altri legami, altrettanto importanti, collegano la genetica biometrica e la genetica delle popolazioni con lo studio della coltivazione, dell'ecologia e della sistematica delle piante. L'inizio degli studi biometrici sull'eredità precedette la riscoperta dell'opera di Mendel nell'anno 1900, e ci volle del tempo prima che le ricerche mendeliane venissero messe in relazione con i risultati biometrici. Una vera pietra miliare fu rappresentata dall'analisi dell'eredità quantitativa in piante quali il grano, nelle quali un'interpretazione soddisfacente di ciò che appariva come una variazione graduale e continua nella progenie ibrida si basò sull'ipotesi che l'espressione dei caratteri fosse controllata da numerosi fattori mendeliani interagenti, l'ipotesi dei ‛fattori multipli'. Oggi, i fattori multipli vengono chiamati poligeni, e sono usati complicati metodi matematici per analizzare i caratteri ereditari poligenici e valutare l'effetto dell'ambiente nel controllo dell'espressione di caratteri quantitativi; metodi, questi, che hanno una grande importanza nelle coltivazioni dei cereali e di altre piante.
La genetica delle popolazioni studia la variazione interna di popolazioni naturali di organismi e come tale variazione sia generata e regolata. L'andamento della variazione genetica in una popolazione dipende dal sistema di incrocio, dai tipi di ricombinazioni, mediante i quali possono variare le combinazioni dei geni trasportati dai cromosomi, e dalla frequenza di mutazione genica. Le piante hanno tipi di incrocio più vari degli animali e dimostrano un più ampio spettro di variazioni cromosomiche, di numero, dimensione e struttura, e anche di comportamento, durante il particolare processo di divisioni cellulari che precede la formazione delle cellule sessuali, quando i geni che derivano dai cromosomi provenienti dai genitori vengono ricombinati prima di essere trasmessi alla prole.
Gli studi sulle popolazioni di piante hanno notevolmente contribuito alla moderna interpretazione della variazione e dell'adattamento all'interno delle specie. Le ricerche in questo campo sono legate soprattutto al nome di G. W. Turesson, che introdusse il termine di ‛genecologia' per indicare la disciplina formata dalla combinazione della genetica, dell'ecologia e della tassonomia, branche che comprendono gli studi sulla microevoluzione delle piante. L'adattamento locale delle popolazioni di una data specie dipende dalla selezione naturale, che opera sull'insieme delle varianti genetiche, dovute alla ricombinazione e alla mutazione dei geni. Quando una specie incontra differenti condizioni ecologiche nelle diverse parti dell'area in cui è presente, ci si può aspettare un differenziamento in seguito ai vari tipi di pressione selettiva che agiscono in ciascuna area. Il Turesson chiamò ‛ecotipi' le popolazioni locali adattatesi in questa maniera. Dall'inizio della genecologia, si è raccolto un gran numero di informazioni sulla variabilità interna delle specie vegetali, e si sono modificati e precisati alcuni dei concetti originari. Si sa, oggi, che un aspetto comune della variazione adattativa implica non il differenziamento di ecotipi distinti, ma piuttosto il cambiamento graduale e continuo di caratteristiche strutturali o fisiologiche lungo un tratto continuo di territorio, spesso correlato a una variabilità ambientale ugualmente graduale, come per esempio la stagione di crescita. Gradienti di questo tipo all'interno delle specie vengono chiamati ‛clini', ed ‛ecoclini' quando sono chiaramente adattativi.
L'orientamento attuale nella genecologia delle piante è caratterizzato dal tentativo di interpretare l'adattamento all'ambiente in termini fisiologici. Lo scopo è difficile da raggiungere a causa della grande difficoltà nell'identificare e misurare i fattori ambientali che contribuiscono attualmente alle pressioni selettive nei differenti habitat. Nonostante ciò, la strumentazione fisica, oggi disponibile, permette di compiere qualche progresso. Questi problemi saranno ripresi in paragrafi successivi in connessione con la fisiologia e l'ecologia (v. sotto, §§ f-g).
Biochimica
La biochimica cerca di descrivere i processi vitali delle piante in termini chimici precisi. Le sue tecniche fondamentali sono state sviluppate tutte durante il XX secolo, e argomento centrale delle sue ricerche sono gli studi sulle proprietà degli enzimi, catalizzatori del metabolismo biologico. Mediante lo studio di enzimi e di sistemi enzimatici isolati, si possono ricostruire i processi metabolici e chiarire le interazioni fra i differenti processi. La nozione che i vari sistemi metabolici cellulari siano situati in particolari tipi di organelli è stata quasi completamente sviluppata negli ultimi trent'anni. La localizzazione di questi sistemi associati a particelle (o ‛particolati') è stata consentita dalla possibilità di frazionare le cellule in condizioni tali da non distruggere gli enzimi; gli strumenti più importanti che vengono utilizzati a tale scopo sono i vari tipi di centrifughe. L'uso della centrifugazione ad alta velocità permette di separare i vari componenti della cellula e di isolare organelli, quali i mitocondri, in modo da poterne studiare le proprietà in sistemi in vitro. La centrifugazione in gradiente di densità può essere utilizzata per separare differenti classi di molecole presenti nella cellula come, per esempio, i vari tipi di RNA - permettendo in questa maniera un potere di risoluzione fisica analogo a quello ottenuto col microscopio elettronico.
La conclusione forse più importante che si può trarre dalle scoperte della biochimica moderna è che tutti gli organismi formati da cellule hanno in comune la maggior parte dei loro processi metabolici fondamentali. Il codice genetico contenuto nel DNA del nucleo cellulare viene letto sempre nella medesima maniera: l'informazione viene dapprima trasferita all'mRNA, e quindi tradotta nella struttura di proteine specifiche, come accennato nei precedenti paragrafi. Molte delle reazioni catalizzate da proteine enzimatiche sono comuni a piante, animali e microrganismi: un esempio evidente è quello del processo chimico, mediante il quale in organismi aerobi viene liberata energia nel corso della respirazione per essere poi immagazzinata o trasferita ad altre reazioni. Nelle piante, le molecole degli zuccheri vengono dapprima scisse da enzimi presenti nel citoplasma, e quindi ossidate da un gruppo di enzimi presenti nei mitocondri: è un processo ciclico, al quale partecipano numerosi enzimi. L'energia prodotta viene in parte immagazzinata mediante la formazione di legami fosforici in particolari molecole ‛ad alta energia', le quali agiscono da intermediari nel trasferimento dell'energia ad altre reazioni di sintesi: per esempio, gli ‛scheletri' di atomi di carbonio utilizzati nella sintesi degli amminoacidi sono ottenuti in questa maniera.
Fra i processi metabolici caratteristici delle cellule vegetali vi sono quelli che riguardano la fotosintesi, il processo che sta alla base della vita autotrofica delle piante verdi e di alcuni tipi di Batteri. Forse il maggior risultato della biochimica vegetale è stata la scoperta delle reazioni metaboliche attraverso le quali, nei vari passaggi della fotosintesi, il diossido di carbonio (o anidride carbonica) dell'atmosfera viene ridotto a carboidrati grazie all'utilizzazione dell'energia solare. Nei cloroplasti, l'energia luminosa immagazzinata nei granuli dai pigmenti di clorofilla viene utilizzata per scindere le molecole di acqua, liberando così ossigeno, e riducendo un composto intermedio: questo prende parte a reazioni successive, che non richiedono la presenza di luce e che permettono la fissazione del diossido di carbonio. Una delle principali vie metaboliche del carbonio fu scoperta, negli anni immediatamente seguenti la seconda guerra mondiale, da M. Calvin e A. A. Benson mediante l'utilizzazione dell'isotopo radioattivo 14C. Anche in questo caso il processo è ciclico: dapprima il diossido di carbonio viene fissato da una molecola accettrice a cinque atomi di carbonio, questa poi si scinde dando molecole a tre atomi di carbonio, le quali si uniscono formando i carboidrati. In questo processo viene ripristinata la molecola accettrice, mentre vengono formati anche zuccheri a sei atomi di carbonio, che seguono altre vie metaboliche. Si conosce ora un'altra differente sequenza di reazioni per la fissazione del diossido di carbonio: recenti ricerche hanno dimostrato che, oltre alla reazione che produce composti con attività riducente, esiste un'altra reazione luminosa della fotosintesi, la quale genera legami fosforici ad alta energia; in questa maniera l'energia luminosa è resa direttamente disponibile per consentire altre reazioni, comprese alcune di quelle che permettono la fissazione del diossido di carbonio.
Una più completa trattazione della respirazione e della fotosintesi si può trovare in altri articoli (v. bioenergetica; v. fotosintesi).
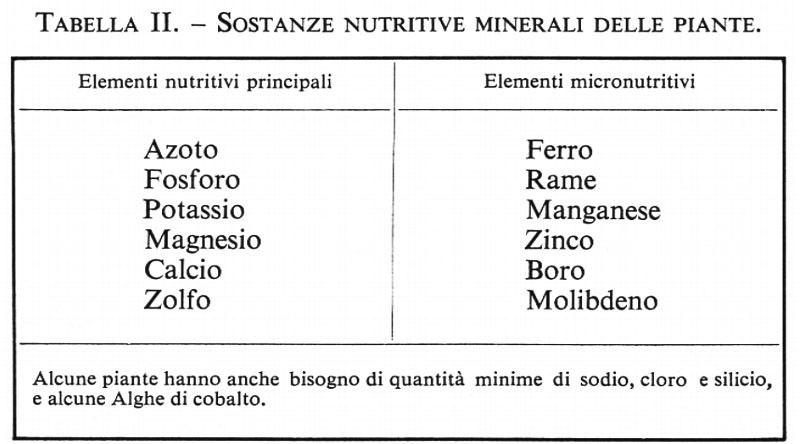
La fotosintesi è certamente uno dei processi essenziali per l'esistenza della vita sulla terra, ma di uguale importanza è un'altra serie di reazioni che interessa un gas atmosferico: la fissazione dell'azoto, che dipende ugualmente dalle piante. Benché questo elemento sia indispensabile per tutti gli esseri viventi, gli organismi superiori sono privi della capacità di utilizzare l'azoto direttamente, capacità posseduta invece da particolari Alghe e Batteri; alcuni microrganismi, generalmente simbionti, arricchiscono direttamente le riserve di azoto del suolo e rendono questo elemento disponibile per le piante con radice e quindi per gli animali. Verso la fine del XIX secolo si poté mettere in evidenza, con classici esperimenti sul terreno, l'importante caratteristica delle piante leguminose - piselli, fagioli, trifoglio, ecc. - di arricchire il terreno di azoto; tale capacità si dimostrò dovuta alla presenza di Batteri simbionti che vivono nei noduli delle radici; negli ultimi anni, è stata dimostrata la capacità di fissazione simbiontica dell'azoto anche da parte di specie differenti dai legumi. È stata scoperta una forma di emoglobina, la leghemoglobina - come è stata chiamata da A. I. Virtanen -, nei noduli radicali delle leguminose: la leghemoglobina viene formata dalle cellule ospiti in risposta all'invasione batterica, ma ancora oggi non si hanno dati definitivi circa la possibile funzione svolta da questo pigmento nel processo stesso di fissazione dell'azoto. Per la sintesi della leghemoglobina è necessario il ferro e probabilmente anche il cobalto: per tale motivo, il cobalto è essenziale perché i legumi possano sviluppare la loro capacità di fissare l'azoto, benché questo elemento non sia considerato un vero e proprio nutrimento per le piante (v. tab. II).
Le piante superiori, e soprattutto le piante con fiori, mostrano una straordinaria abilità biosintetica, riuscendo a formare un grandissimo numero di composti organici, molti dei quali non sembrano di particolare importanza per il metabolismo fondamentale della cellula. Questi prodotti cosiddetti ‛secondari' comprendono gli alcaloidi, i pigmenti, i terpeni e sostanze simili; tali composti, molti dei quali servono all'uomo nella medicina o nell'industria, possono fornire per quel che riguarda la stessa botanica, utili criteri di classificazione.
Gran parte delle ricerche di biochimica vegetale riguarda lo studio dei processi biosintetici di prodotti secondari.
Fisiologia
La fisiologia è collegata in vari modi alla biochimica, ma mentre il biochimico studia i processi cellulari e subcellulari, il fisiologo rivolge la sua attenzione alla funzione degli organi, ai sistemi di organi e agli interi organismi. Nelle piante, fenomeni di particolare interesse sono costituiti dalla fissazione, dal movimento, dal deposito e dall'utilizzazione dell'acqua e delle sostanze nutritive; dalla regolazione e dal controllo degli scambi gassosi necessari alla respirazione e alla fotosintesi; dalla fisiologia dell'accrescimento, dello sviluppo e del movimento; dai meccanismi di percezione e di risposta a stimoli ambientali; e dai vari eventi connessi con la riproduzione.
Le principali esigenze nutritive delle piante furono accertate nel XIX secolo, ma solo con la possibilità di avere dei composti spettroscopicamente ‛puri' si riuscì a dimostrare l'indispensabilità, per l'accrescimento delle piante, di elementi quali lo zinco, il rame, il boro, il molibdeno e il cloro. L'elenco delle sostanze essenziali per le piante comprende ora sedici elementi, sei come macronutritivi e gli altri come micronutritivi (v. tab. Il). Deficienze nella disponibilità delle diverse sostanze nutritizie essenziali producono sintomi caratteristici di alterazioni fisiologiche, che nelle coltivazioni possono essere facilmente diagnosticate e corrette aggiungendo al terreno le sostanze necessarie. Lo studio, compiuto in quest'ultimo secolo, sul valore nutritivo dei minerali per le piante ha avuto un'enorme importanzà economica, poiché su di esso si basa la maggior parte dell'odierna produzione mondiale di alimenti.
Collegato alla ricerca sul valore nutritivo dei minerali è il già ricordato studio sulla fissazione dell'azoto. Sempre in questo settore, un altro importante campo di studio è quello dei processi attraverso i quali le sostanze nutritive entrano nella pianta. L'assunzione, da parte di piante provviste di radice, di sali dal terreno o dalla soluzione nella quale sono immerse nel caso di specie acquatiche, comporta il passaggio di ioni attraverso le membrane e il loro deposito nell'interno della cellula vivente: ciò implica generalmente un movimento contro un gradiente di concentrazione, processo che richiede energia ed è abbinato, come è oggi ben noto, con la respirazione. Nello studio sul movimento degli ioni sono state utilizzate soprattutto alghe con cellule di grandi dimensioni e provviste di numerosi vacuoli, particolarmente adatte per l'applicazione di metodi biofisici. Alcune teorie abbastanza complesse per spiegare il movimento degli ioni nelle piante sono già state elaborate, ma l'esatto meccanismo delle ‛pompe ioniche', di cui si è immaginata l'esistenza a livello delle membrane cellulari, non è stato ancora completamente chiarito.
La fisiologia dei rapporti fra l'acqua e le piante è un argomento di grande importanza poiché, generalmente, la vegetazione sulla terra si trova per la maggior parte del tempo in uno stato di mancanza totale, o quasi, d'acqua. Si ammette generalmente che l'acqua non venga mai trasportata attivamente nella pianta - cioè, spinta dall'energia respiratoria - ma piuttosto che si muova secondo i propri gradienti di potenziale (ovvero la differenza di pressione di diffusione), i quali possono essere innalzati dalle pressioni reali o dai potenziali osmotici dipendenti dalle concentrazioni del solvente nei vacuoli della cellula. Su questa base si può dare una spiegazione teorica ragionevolmente accettabile dell'economia dell'acqua nelle piante terrestri.
La traspirazione è la fuoriuscita di acqua sotto forma di vapore nell'atmosfera dalle parti aeree delle piante, e il flusso di traspirazione è il movimento dell'acqua attraverso la pianta a partire dalla radice, dove viene assunta dal terreno. Non considerando la complessità dei passaggi intermedi, l'acqua passa da una condizione di potenziale maggiore a livello del terreno a una di potenziale minore nell'atmosfera, e il gradiente costituisce la forza che ne rende possibile il movimento. L'ascesa della linfa nei grandi alberi può essere spiegata con l'ipotesi che la tensione venga trasmessa attraverso il tessuto di conduzione (lo ‛xilema') lungo colonne d'acqua dalle dimensioni capillari senza soluzione di continuità: è questa la teoria della coesione della linfa ascendente, esposta nella sua prima enunciazione da H. H. Dixon e J. Joly negli ultimi anni del XIX secolo; per convalidarla, tuttavia, gli studi sulla struttura dei presunti tessuti di conduzione dell'acqua hanno incontrato alcune difficoltà, così da renderla non universalmente accettata come completamente idonea a spiegare questi fenomeni.
La regolazione della perdita di acqua dalle strutture aeree delle piante terrestri deve essere considerata come parte del controllo generale degli spostamenti dei gas, dal momento che gli scambi che avvengono durante il processo di fotosintesi seguono gli stessi meccanismi, avvengono cioè principalmente attraverso gli stomi della superficie della foglia.
Uno stoma si apre o si chiude grazie alla flessione della coppia di cellule situate attorno ad esso: la forza che causa il movimento di apertura è l'entrata dell'acqua con meccanismo osmotico, mentre la chiusura avviene per perdita di acqua e quindi di turgidità. Non è chiaro l'esatto meccanismo di controllo, ma si conosce l'influenza di un gran numero di fattori ambientali. A causa dell'importanza che ha la regolazione della traspirazione per la fisiologia delle piante coltivate, soprattutto nelle terre aride, su tali fenomeni si sono concentrate molte ricerche. Fra i problemi che vengono attivamente affrontati, vi sono quelli microclimatici, riguardanti gli effetti che ha la vegetazione sul clima in prossimità del terreno, in particolare le modalità con le quali la geometria della vegetazione influisce sul movimento del vapore acqueo. Per quel che riguarda le singole piante, numerosi fattori interagiscono per regolare gli scambi di gas. È stato già osservato in un precedente paragrafo (v. sopra, § b) che sono importanti soprattutto la morfologia della foglia e la micromorfologia delle sue superfici: infatti, come era stato previsto dai primi studiosi della morfologia delle piante, alcune caratteristiche strutturali, quali peli e stomi infossati, sono connessi soprattutto con la regolazione del movimento del vapore acqueo, creando turbolenza o aumentando le vie di diffusione. La forma della foglia è, per via indiretta, di grande importanza nel regolare gli equilibri energetici, poiché la traspirazione è il mezzo più efficace per il passaggio d'energia dalla pianta al suo ambiente. In questi campi, la fisiologia vegetale è legata con la biofisica.
Forse, i progressi più significativi per la fisiologia vegetale in questo secolo sono stati compiuti nello studio dell'accrescimento e dello sviluppo. L'accrescimento delle piante superiori è differente, per diverse importanti caratteristiche, da quello degli animali, Soprattutto perché esse non presentano un vero stato embrionale, mentre dimostrano continuamente, nel corso della vita, fenomeni di crescita e di formazione di organi; inoltre, l'accrescimento delle piante dipende in gran parte dalle caratteristiche della parete cellulare, fattore che non influisce sull'accrescimento degli animali. Generalmente, l'aumento di dimensioni di un organo vegetale dipende dall'aumento del numero delle cellule in esso presenti e dall'ingrandimento di ciascuna singola cellula; il controllo dell'accrescimento deve quindi implicare almeno due sistemi subordinati: uno preposto al controllo della divisione cellulare e l'altro a quello della crescita cellulare. Da un punto di vista storico, fu studiata prima dettagliatamente la fisiologia dell'aumento delle dimensioni della cellula: F. A. F. C. Went e F. W. Went riuscirono a dimostrare a Utrecht negli anni venti, lavorando con il cotiledone o coleottile delle Graminacee (organo che cresce per l'aumento delle dimensioni delle cellule e senza divisioni cellulari) che l'aumento della cellula era regolato in quest'organo da un agente diffusibile che essi chiamarono auxina. Questi studi portarono all'estrazione del primo ormone delle piante (v. tab. III). Nello stesso periodo, lo studio compiuto a Formosa da un fitopatologo, E. Kurasawa, su di una malattia della pianta del riso aprì una nuova via alla ricerca, che portò infine all'identificazione di una seconda classe di sostanze attive nella crescita, le gibberelline. La scoperta di fattori attivi sulla divisione delle cellule vegetali fu dovuta all'utilizzazione di colture di tessuti di piante, in cui gruppi di cellule staccate dalla pianta venivano fatti crescere in un mezzo nutritivo in condizioni di sterilità. Seguendo un'indicazione fornita da O. Haberlandt nei primi anni del secolo, F. Skoog e collaboratori identificarono finalmente dei composti, capaci di accelerare la proliferazione di cellule in coltura, costituenti una classe di sostanze stimolanti la crescita, chiamate ora citochinine. I primissimi esperimenti furono compiuti con sostanze sintetiche non presenti in natura nelle piante; ora sono stati scoperti i corrispondenti composti naturali (v. tab. III). L'ultima fase della ricerca sugli ormoni vegetali si è rivolta a quelle sostanze che possono agire nella regolazione del letargo e della caduta delle foglie e ha portato all'isolamento e all'identificazione dell'acido abscissico, fattore interessato probabilmente nella regolazione del periodo di riposo dei germogli e dei semi. Si conoscono anche altri tipi di inibitori che possono essere interessati nella regolazione dell'accrescimento.

La scoperta del modo di azione dei vari tipi di sostanze che agiscono sull'accrescimento delle piante costituisce una impresa assai impegnativa. Una difficoltà è costituita dal fatto che la maggior parte delle risposte che si osservano nelle piante intere sono dovute all'interazione di varie molecole fisiologicamente attive fra loro e con fattori nutritivi. Benché le proprietà di ciascuna delle classi elencate nella tab. III possano essere specificate in particolari sistemi parziali, raramente si può predire da questi dati la risposta che vi sarà nella pianta intera. Nonostante ciò, vi sono numerosi campi d'indagine che evolvono rapidamente. La parete cellulare svolge un ruolo importante nella regolazione dell'accrescimento, e le caratteristiche della parete vengono certamente modificate dalle sostanze che agiscono sull'aumento in estensione: risposte di questo tipo sono senz'altro indirette, essendo l'effetto mediato da sistemi enzimatici presenti nel citoplasma. È stato dimostrato che auxine, gibberelline e citochinine modificano alcune caratteristiche del metabolismo fondamentale della cellula, fra cui l'attività enzimatica e in alcuni casi la sintesi proteica. Una serie di osservazioni assai importanti sono state fatte in studi riguardanti gli effetti delle gibberelline sulla sintesi di enzimi che scindono l'amido e di altri enzimi presenti in alcuni tessuti della cariosside di orzo: sembra che in questo caso l'ormone stimoli la sintesi enzimatica agendo sul gene, implicando una funzione che avviene a uno stadio assai precoce. Si riscontra un comportamento simile in alcuni casi in cui agisce un'auxina. Ricerche recenti sulle citochinine hanno fatto ipotizzare - ma si tratta di una mera ipotesi - che queste svolgano una funzione a livello della sintesi proteica, quando gli amminoacidi si uniscono per formare una catena polipeptidica secondo l'ordine determinato dalla sequenza di basi presente nell'mRNA. In opposizione all'ipotesi che le sostanze che stimolano l'accrescimento, come le auxine e le gibberelline, agiscano in maniera specifica, regolando la funzione del gene, si deve tener conto della molteplicità degli effetti che esse provocano e del fatto che le risposte variano da tessuto a tessuto, anzi in ogni particolare tessuto, a seconda dell'età e dei processi che ha subito precedentemente. Ciò suggerisce l'ipotesi che gli ormoni possano agire come attivatori piuttosto aspecifici, e si possono immaginare vari meccanismi per questo tipo di azione: la loro attività potrebbe essere mediata da molecole ‛recettrici' specifiche presenti in certi stadi della vita di una cellula; o, come è stato già osservato, il loro bersaglio potrebbe essere costituito dai sistemi di membrane che regolano il passaggio di sostanze all'interno del citoplasma e fra i diversi organelli.
Le ricerche sullo sviluppo delle piante hanno proceduto di pari passo con quelle sulle sostanze necessarie all'accrescimento. Sviluppo è il termine col quale viene definita la successione dei cambiamenti che subisce un organismo dallo stato di uovo fecondato sino alla maturità; esso è l'insieme di molti cambiamenti singoli a partire da quelli che avvengono a livello cellulare, nei quali una cellula o un gruppo di cellule subisce un cambiamento di forma, struttura, o proprietà chimiche, in quanto facente parte di un'area specializzata per una particolare funzione. Per molte piante inferiori lo sviluppo e la differenziazione cellulare si equivalgono. Risultati assai indicativi sono stati ottenuti da studi compiuti da J. Himmerling su di un'alga gigante unicellulare, l'Acetabularia. Mediante il trapianto è stato messo in evidenza il ruolo del nucleo cellulare nella determinazione di caratteri specifici, e il fatto che l'espressione dei geni nucleari dipende dalla sintesi di differenti tipi di mRNA è stato dimostrato grazie a esperimenti in cui venivano utilizzati degli inibitori chimici, e a ricerche di altro tipo. Un'importante scoperta è stata la dimostrazione che alcuni aspetti del differenziamento di questi organismi possono essere regolati da fattori ambientali in una fase successiva alla ‛lettura' dell'informazione genetica: tale risultato comporta che non è necessario supporre che il controllo dello sviluppo nelle piante debba essere sempre esercitato al livello fondamentale del gene.
Gran parte delle possibilità che ha l'uomo di utilizzare le piante dipende dalla sua capacità di farle crescere e di manipolarne i processi di sviluppo secondo i propri fini, così che non desta sorpresa il fatto che un tema continuamente ricorrente nella fitofisiologia sia quello dell'effetto dell'ambiente sui processi di crescita. Sono già stati ricordati gli importanti fattori nutritivi minerali e i rapporti con l'acqua. Altri fattori ambientali variabili sono i regimi di luce e di temperatura, dei quali alcuni effetti sono direttamente prevedibili dalla natura dell'economia di base delle piante: per la maggior parte di queste, la luce, essendo l'unica fonte di energia, svolge sempre un ruolo fondamentale per l'accrescimento; la temperatura, invece, per l'influenza esercitata sulle velocità delle reazioni chimiche, interviene necessariamente in una qualche maniera in tutti i processi vitali. Tuttavia, in un senso più specifico, la temperatura e la luce possono partecipare al particolare controllo dello sviluppo fornendo dei ‛segnali' dall'ambiente che inducono la comparsa di determinate modificazioni. Ciò è soprattutto evidente nei climi molto legati alla stagione, nei quali la sopravvivenza dipende da una precisa sincronizzazione dello sviluppo con il ritmo stagionale: così la fioritura, la produzione di frutti, la dispersione dei semi e il letargo devono essere regolati a seconda del periodo dell'anno in quei climi, nei quali il periodo dell'accrescimento è limitato da un inverno freddo o da un'estate calda e secca. Nel 1911 fu scoperta da Tournois l'importanza della lunghezza del giorno come ‛orologio' per il controllo dello sviluppo delle piante, e da allora l'effetto della durata del giorno, noto come fotoperiodismo, è stato oggetto di una gran quantità di studi: uno dei risultati è stato il riconoscimento di diversi tipi di risposte fra le piante (v. tab. IV). Un'altra scoperta è stata quella dell'esistenza di un legame fra il ruolo esercitato dalla luce nel regolare lo sviluppo delle piante nella loro fase di accrescimento e il suo effetto sulla germinazione di certi tipi di semi. Ricerche compiute negli Stati Uniti sulla sensibilità delle diverse reazioni alla luce di varie lunghezze d'onda hanno dimostrato che era interessato lo stesso pigmento recettore: questo, successivamente isolato e chiamato fitocromo, ha la caratteristica peculiare di essere presente in due forme metastabili che assorbono luce di diversa lunghezza d'onda; le due forme sono interconvertibili mediante l'irradiazione con luce di un colore adatto. È noto oggi che il fitocromo è interessato in diversi fenomeni dello sviluppo sia nelle piante inferiori sia in quelle superiori, fenomeni che vengono definiti talvolta fotomorfogenici, poiché comportano un cambiamento della forma in risposta alla luce. Non vi è dubbio che il fitocromo sia il pigmento recettore della luce, anche se deve essere ancora chiarito il suo meccanismo fondamentale d'azione; probabilmente è significativo il fatto che nella cellula il fitocromo è associato alle membrane.

Effetti specifici della temperatura sullo sviluppo delle piante erano già noti da più di un secolo e mezzo, ma lo studio scientifico di questi effetti ha avuto inizio solamente con le ricerche di G. Gassner in Germania sulla fioritura dei cereali. Alcune specie, cereali e altre, hanno un ciclo di accrescimento biennale, con la fioritura nel secondo anno; in queste specie di norma si è trovato che il fotoperiodo e l'effetto della temperatura interagiscono nel processo di regolazione del tempo, essendo di solito necessarie, per uno sviluppo successivo dopo l'accrescimento del primo anno, temperature fredde tipiche dell'inverno. Il nome del fitofisiologo russo T. D. Lysenko è legato al trattamento dei cereali con una temperatura artificiale per stimolarne la fioritura prematura: tale trattamento viene chiamato ‛vernalizzazione'. Effetti della temperatura si hanno anche in molte piante nella regolazione del letargo dei germogli.
Gli effetti sullo sviluppo dovuti sia al fotoperiodismo sia alla temperatura sono probabilmente determinati, almeno in parte, da diversi tipi di ormoni vegetali: in numerosi casi è stata dimostrata una spiccata influenza di queste variabili sul metabolismo delle auxine e delle gibberelline. Nonostante ciò, benché vi sia oggi una gran quantità di dati empirici, raramente si possono fare generalizzazioni. Alcuni fra i più importanti e complessi problemi sono quelli riguardanti i sistemi ‛interni' di controllo del tempo nelle piante, i cosiddetti ‛biologici', che consentono una sensibilità alle variazioni di durata del giorno. Il fenomeno dell'esistenza, in piante di varie classi, di fenomeni ritmici innati fu posto in evidenza nel 1930 da E. Bünning, e vi sono oggi buone prove sperimentali che dimostrano come la risposta fotoperiodica sia associata a ritmi metabolici interni, denominati ‛circadiani' poiché hanno un ciclo di circa 24 ore.
Le piante non solo rispondono a fattori ambientali, adattandovisi col loro programma di sviluppo, ma anche modificando le proprie caratteristiche di accrescimento, di forma, di posizione e di orientamento. Nelle piante inferiori dotate di motilità, si riscontra una sensibilità alla luce, alla temperatura e alle sostanze chimiche simile a quella presente negli animali meno evoluti, e le cellule si muovono tutte insieme verso ambienti adatti. I movimenti adattativi nelle piante sessili dipendono dalla flessibilità dei tessuti, che è dovuta a sua volta a cambiamenti nel contenuto d'acqua o a curvature derivanti da diverse velocità di accrescimento in due lati di un organo. Le curvature dovute alla luce (fototropismo) e alla gravità (geotropismo) furono studiate per la prima volta da Charles Darwin un secolo fa, e su questo fenomeno continuano a concentrarsi tuttora molte ricerche. Non vi è dubbio che gli ormoni della crescita partecipino ai processi di sviluppo e differenziamento; sono state formulate varie teorie che cercano di spiegare i fenomeni del fototropismo e del geotropismo sulla base della ridistribuzione degli ormoni formatisi in seguito a stimoli ambientali. Negli ultimi anni si è cercato di dimostrare queste ipotesi usando sostanze sintetiche stimolatrici dell'accrescimento marcate con isotopi radioattivi, ma non si può dare ancora una spiegazione del tutto esauriente di tutti i vari tipi di effetti tropici conosciuti.
Gli studi odierni sui fenomeni connessi con la riproduzione delle piante dimostrano assai bene quanto il progresso di tali conoscenze dipenda dall'applicazione simultanea delle tecniche di numerose discipline. L'induzione dell'attività riproduttiva dipende da risposte ormonali di differenti tipi, che sono state studiate con metodi biochimici sia negli organismi inferiori, compresi i Funghi e le Alghe, sia nelle piante da fiore. Spesso sono necessari anche stimoli ambientali, e talvolta l'interazione fra singole piante di una specie aventi differenti funzioni sessuali. Lo studio della differenziazione della cellula sessuale, della sua fecondazione e dello sviluppo dell'embrione costituisce un campo di studio comune alla biologia cellulare, alla biochimica e alla fisiologia. Benché dalla fine del XIX secolo in poi la citologia della riproduzione delle Angiosperme sia stata studiata in maniera assai approfondita, i suoi aspetti fisiologici sono stati, al contrario, trascurati fino agli ultimi decenni. Fra i campi attualmente di particolare interesse, possono essere ricordati lo studio della fisiologia dei gametofiti, o generazioni che producono gameti, dei sacchi embrionali e dei granuli di polline.
Ecologia e fitosociologia
L'ecologia cerca di mettere in relazione il comportamento di singole piante, di specie di piante e di comunità di piante con i loro ambienti viventi e non viventi (v. associazioni biologiche). L'ecologia descrittiva, che riguarda la struttura e le relazioni interne delle comunità di piante, è spesso definita sociologia vegetale o fitosociologia. L'ecologia è strettamente collegata alla geografia delle piante, tanto che per i botanici del secolo scorso non vi era distinzione tra queste due discipline. La geografia floristica delle piante è in ogni caso strettamente connessa con gli aspetti storici della vegetazione, mentre l'ecologia si occupa in primo luogo degli eventi contemporanei (v. ecologia).
È stato da lungo tempo riconosciuto che le piante formano delle comunità con una struttura caratteristica, e l'uomo sa che la natura di queste comunità e la loro distribuzione sulla superficie della terra dipendono in massima parte da fattori secolari, come il clima e la configurazione topografica. Da una concezione generale del tipo di vegetazione è derivato il concetto più specifico dell'associazione di piante, caratterizzata, questa, dalle specie che la compongono e dalla loro importanza relativa per quel che riguarda il numero e le dimensioni, o l'estensione dell'area ricoperta. Un importante progresso negli anni trenta fu la crescita in Europa di una scuola fitosociologica che tendeva a una classificazione la più ampia possibile delle associazioni vegetali, particolarmente quelle delle regioni temperate, sulla base della composizione delle specie, della dominanza e del grado di purezza delle singole specie. In questo modo si è realizzata una mappa della vegetazione di gran parte dell'Europa.
Il metodo fitosociologico non è stato accettato da tutti gli ecologi, dal momento che molti hanno trovato difficile ammettere che la vegetazione possa essere classificata in un modo troppo rigido. La classificazione fitosociologica dipende in una certa misura da valutazioni soggettive sulla composizione della vegetazione, e negli ultimi due decenni sono stati sempre più utilizzati i metodi quantitativi nell'ecologia descrittiva delle piante.
Le tecniche per valutare la composizione della vegetazione sulla base di una campionatura obiettiva si sono evolute, e sono ora utilizzabili procedimenti statistici per paragonare e classificare comunità di piante. Per la sua stessa natura, l'ecologia quantitativa delle piante genera una grande quantità di dati, e la tecnologia dei calcolatori è ora un essenziale ausilio per esami più approfonditi.
La struttura delle associazioni vegetali è determinata da molti fattori, tra i quali non solo le principali influenze climatiche come la temperatura, le precipitazioni, l'illuminazione e il vento, ma anche i fattori geologici e topografici, gli effetti prodotti dagli animali compreso l'uomo, l'interazione fra le stesse piante e naturalmente tutta la storia precedente dei viventi della zona. Per la complessità che le interazioni comportano, l'ecologia interpretativa delle piante, che cerca di rintracciare le cause remote dei tipi di distribuzione delle piante e delle strutture della comunità, è una disciplina difficile. I primi tentativi di mettere in relazione tipi di vegetazione con vari parametri climatici hanno incontrato qualche successo solo quando i fattori limitanti in singole zone erano ben definiti, come si verifica allorché la piovosità o la temperatura sono le principali determinanti, mentre gli sforzi per spiegare dettagliatamente la struttura e i tipi di distribuzione delle associazioni vegetali sulla base dei singoli fattori climatici o di semplici combinazioni di questi fattori, generalmente non hanno approdato a buoni risultati.
Per semplificare i problemi, sono stati condotti studi approfonditi sulle singole specie (autoecologia) in opposizione allo studio delle comunità, o sinecologia. Tali studi hanno mostrato che i fattori dominanti che determinano il comportamento di una specie possono variare completamente nell'area ove questa è presente, cosicché i suoi confini di distribuzione possono essere regolati da diverse circostanze - ad esempio un valore di temperatura in una zona del campo, la piovosità in un'altra e il tipo del suolo in un'altra ancora. Un'importante caratteristica della recente ricerca ecologica sulle piante, soprattutto degli studi autoecologici, è stato l'aumento dei metodi sperimentali. Utilizzando le tecniche di laboratorio in questo campo, si possono misurare la fotosintesi e la respirazione, valutare la fissazione e l'utilizzazione delle sostanze minerali, studiare molti aspetti dell'interazione fra le piante, compresi gli effetti della competizione e delle malattie. Un'informazione quantitativa di questo tipo è necessaria negli studi sulla produttività della vegetazione, sia per le comunità naturali sia per quelle regolate dall'uomo.
Anche se i metodi sperimentali possono essere applicati per chiarire molti aspetti delle caratteristiche delle piante in ambienti naturali, le possibilità di variare i fattori dell'habitat in esperienze all'aperto sono limitate. In ogni modo, con l'uso di ambienti artificiali in serre, si può studiare con buona precisione la risposta delle specie vegetali a una vasta gamma di condizioni. Gli aspetti tecnici per fornire in ambienti chiusi condizioni climatiche adatte alla crescita delle piante sono complessi, e le installazioni necessarie per consentire un controllo soddisfacente sono spesso ingenti. Un lavoro pionieristico su questi problemi fu compiuto in California negli anni quaranta, e da allora le installazioni di ambienti artificiali di grandi dimensioni per farvi crescere piante sono chiamate ‛fitotroni'.
Gruppi di ambienti controllati sono stati usati ampiamente per studi fisiologici sul comportamento delle piante e le reazioni a importanti variazioni ambientali, come la lunghezza del giorno e la temperatura, sono state rilevate in modo che non sarebbe stato possibile con la semplice osservazione sul campo. Una notevole messe di informazioni di questo tipo è stata raccolta sia per le specie selvatiche, sia per le piante coltivate e per quelle economicamente importanti, quali gli alberi delle foreste.
Geografia delle piante, paleobotanica e storia della flora
La geografia delle piante riguarda la distribuzione delle specie vegetali e della flora sulla superficie della terra; essa, come abbiamo visto, è strettamente collegata all'ecologia delle piante come una branca della botanica. La scienza ebbe inizio ai primi del XIX secolo soprattutto in seguito alle idee di A. von Humboldt, il quale riconobbe che l'attuale distribuzione delle specie vegetali deriva non solo dall'influenza di fattori simultanei - climatici, geologici e topografici - ma anche dall'influenza delle precedenti migrazioni, alcune assai antiche. Le relazioni tassonomiche della flora regionale furono in seguito delineate dettagliatamente da vari botanici e specilmente da A.-P. De Candolle e J. D. Hooker. Quest'ultimo, che nel periodo finale delle sue ricerche subì l'influenza dell'opera di Darwin, si rese conto che la distribuzione attuale della flora spesso non poteva essere spiegata senza presupporre più vasti cambiamenti geologici in epoche primitive. Negli anni più recenti, i dati sulla distribuzione delle piante hanno portato molti botanici ad accogliere la teoria di A. Wegener sulla deriva dei continenti, secondo la quale i continenti esistenti erano anteriormente uniti in un unico grande continente che, nell'era mesozoica, fu separato in segmenti lungo linee di rottura, le quali, allorché i componenti si separarono, divennero gli oceani Atlantico e Indiano. Un'ipotesi di questo genere può essere usata per spiegare le affinità floristiche e faunistiche fra gli odierni continenti senza ricorrere al postulato dei ponti di terra o di una casuale diffusione a grande distanza. Benché inizialmente la teoria della deriva dei continenti fosse sostenuta principalmente dai biologi, negli ultimi decenni anche i dati geologici l'hanno convalidata e l'opinione dei geologi si è spostata in questa direzione.
Naturalmente lo studio dei fossili vegetali è importante per conoscere la storia della vegetazione del mondo. La moderna paleontologia è strettamente connessa al concetto di evoluzione organica, quantunque i fossili siano stati descritti, pur senza una conoscenza reale, in epoca predarwiniana. I più antichi fossili erano le Alghe dell'era precambriana. Per la vegetazione della superficie terrestre, il principale dato delle ricerche paleobotaniche è stata la scoperta di una successione dei vari gruppi principali, l'uno dopo l'altro, in seguito alla prima colonizzazione della terraferma, che deve essere avvenuta più di 400 milioni di anni fa. La più antica flora terrestre risale all'Alto Devoniano ed era composta di piante erbacee, che tuttavia avevano sistemi vascolari e ciclo riproduttivo molto simili a quelli delle moderne Felci. In seguito, sempre nel periodo Devoniano, sono apparse grandi forme legnose, e nel Carbonifero moltissime parti del mondo furono coperte da flora arborea, che ha poi prodotto giacimenti di carbone. Nelle piante simili alle Felci del Carbonifero ha avuto luogo un'evoluzione delle caratteristiche del seme e ciò ha segnato un importante progresso. Questo fu consolidato dalle Gimnosperme, che hanno formato la vegetazione dominante dell'era mesozoica. Le Angiosperme, piante fiorifere, probabilmente registrarono la maggior parte della loro evoluzione in epoca precretacea, ma è nel Cretaceo che esse divennero uno dei maggiori elementi della flora terrestre. Durante l'epoca terziaria le piante che producono fiori hanno costituito incontestatamente la flora dominante della maggior parte della vegetazione terrestre.
Si è avuta perciò una forma di progressione prodotta dall'evoluzione, poiché ogni gruppo successivo rappresentava nell'adattamento biologico un progresso rispetto ai precedenti. Varie spiegazioni sono state avanzate per individuare le cause delle grandi linee evolutive tra le piante; ma senza dubbio i cambiamenti ecologici, causati dalle variazioni climatiche, dai movimenti dei continenti e dai processi di formazione delle montagne, hanno avuto una parte importante. Per l'attuale classe dominante, le Angiosperme, l'evoluzione coordinata con quella degli animali è stata importante nello sviluppo dei meccanismi di impollinazione, dei mezzi di dispersione dei semi e di altre caratteristiche.
La paleobotanica fornisce parte dei dati per ricostruire la filogenesi dei diversi gruppi di piante, cui contribuiscono anche la tassonomia e la morfologia comparata degli esemplari viventi. In qualche gruppo, soprattutto le Felci e le Gimnosperme, la ricchezza dei reperti fossili ha reso possibile, per certe stirpi, plausibili ricostruzioni di una probabile linea ancestrale. Benché vi sia una ricca testimonianza fossile per le piante da fiore durante l'epoca terziaria, la loro antica storia non è ben conosciuta, e non vi è un sistema filogenetico valido per tutto il gruppo. È stata avanzata l'ipotesi che la mancanza di antiche Angiosperme fossili, dalle quali si potrebbe dedurre una significativa linea evolutiva, sia dovuta alle loro origini in aree montuose relativamente aride, ove scarse erano le possibilità di fossilizzazione. È interessante notare che le piante da fiore mostrano oggi una grande diversità ecologica, molto maggiore di ogni altro gruppo di piante esistente: non soltanto esse formano la maggior parte delle foreste del mondo fuori dalle zone boreali, ma si estendono in habitat caratterizzati da condizioni estreme di siccità e di freddo e, in un secondo momento, hanno invaso l'acqua dolce e gli habitat marini.
La storia più recente della vegetazione del mondo è stata tracciata studiando i resti di piante parzialmente fossilizzati in ligniti e torbe. Poiché la maggior parte dei generi importanti che contribuirono alla costituzione delle flore del tardo Terziario e del Quaternario esistono ancora oggi, è stato possibile collegare la composizione delle foreste e delle comunità vegetali di quei periodi con i moderni tipi di vegetazione, e conseguire un importante risultato nella comprensione delle condizioni ecologiche prevalenti. Risultati particolarmente importanti si sono ottenuti per la glaciazione del Pleistocene e per il periodo seguente fino ai nostri giorni, poiché molte testimonianze di vegetazione sono state conservate nelle torbiere. I microfossili e, soprattutto, le spore ben conservate e i granuli di polline sono stati di grande importanza in questa ricerca. Lo studio di questi ultimi tipi di resti costituisce il campo della moderna palinologia. I granelli di polline hanno spesso caratteristiche strutturali così spiccate da rendere possibile l'identificazione del genere e della specie. Poiché la configurazione di un deposito di torba a ciascun livello dipende necessariamente dal numero delle piante genitrici che crescevano nelle vicinanze nel periodo in cui il deposito si formò, gli studi statistici possono offrire un'indicazione abbastanza accurata della composizione della vegetazione in quel periodo. Con questi mezzi è stato delineato un quadro esauriente dei cambiamenti della vegetazione postglaciale e delle tendenze climatiche per l'Europa nord-occidentale e per l'America settentrionale. Le parti delle piante che hanno contribuito ai depositi sono state datate con precisione analizzando il loro contenuto e valutando l'isotopo radioattivo del carbonio 14C. Poiché durante la vita le piante assumono un contenuto di 14C proporzionale al rapporto di questo isotopo nell'atmosfera rispetto all'isotopo non radioattivo 12C, la sua perdita per decadimento radioattivo dopo la morte fornisce la misura del tempo passato da quando iniziò la fossilizzazione. Se ogni strato di un deposito può essere datato con precisione in tale modo, si può stabilire la cronologia di tutto il sistema. Si possono anche stabilire relazioni con le culture umane, quando negli strati si trovano manufatti. In questo modo è stata chiarita l'ecologia degli antichi insediamenti umani.
La botanica e le altre discipline scientifiche
Le piante, come gli altri orgarnsmi viventi, sono costituite da materia a un alto livello di organizzazione. L'esame delle branche della botanica compiuto nei precedenti paragrafi mostra che i botanici da soli adempiono non soltanto al compito di descrivere e classificare le piante di tutti i tipi, ma anche di definirne la costituzione materiale in termini chimici, fisici e matematici, e di caratterizzarne in tale modo i processi vitali e tutte le molteplici interazioni che esse hanno con l'ambiente.
La botanica è perciò strettamente dipendente dalla matematica e dalle scienze fisiche. La matematica fornisce i mezzi per una precisa descrizione e per la manipolazione dei dati e, infine, per formulare espressioni con le quali sono meglio caratterizzati molti fenomeni biologici. La fisica fornisce una gran parte dell'ossatura concettuale per la comprensione dei processi vitali delle piante, incluso l'acquisto di energia e di materiali dall'ambiente, il trasporto interno di acqua e materiali nutritivi, l'accrescimento e lo sviluppo, la percezione e la risposta a stimoli esterni, e le interazioni con gli altri organismi. La chimica, in sostanza, è la scienza fisica che costituisce la base della biochimica, fornendo il linguaggio in cui vengono espressi tutti i processi metabolici e descritti tutti i prodotti sintetizzati dalle piante.
Le relazioni con le scienze geologiche sono di natura differente: mentre le scienze fisiche forniscono i concetti e i termini per descrivere la costituzione e i processi vitali delle piante, le scienze geologiche forniscono informazioni sulle caratteristiche fisiche degli ambienti nei quali esse vivono, diventando così fondamentali per l'interpretazione della loro ecologia e per la comprensione della loro distribuzione.
Ovviamente, esistono intime connessioni tra le scienze delle piante e quelle degli animali: per quanto concerne l'ecologia, animali e piante verdi sono in larga misura complementari, come devono necessariamente essere gli abitanti eterotrofi e autotrofi della biosfera se deve essere mantenuto l'equilibrio biologico. Nello stesso tempo, piante e animali hanno in comune molti dei loro processi biochimici fondamentali, comprese, come abbiamo visto, le basi chimiche dell'ereditarietà. Di conseguenza, la botanica e la zoologia hanno in comune vasti settori nei quali i rispettivi campi di ricerca si sovrappongono o si intrecciano. Infatti, a certi livelli la distinzione tra i due regni ha poco significato.
La botanica e i problemi umani
Le piante forniscono la base per la vita umana sulla terra, essendo contemporaneamente la fonte primaria del cibo dell'uomo e i produttori dell'ossigeno che questo respira. Allo stesso tempo, le piante e i loro prodotti formano una gran parte dell'ambiente in cui l'uomo vive.
Alcuni dei principali contributi dati dalla botanica applicata in questo secolo sono delineati nei paragrafi seguenti.
Fisiologia delle coltivazioni
Questo ramo della botanica applicata interessa l'analisi scientifica della produttività delle piante. Il suo scopo principale è di identificare i fattori ambientali e interni che determinano il rendimento delle coltivazioni, cosicché le pratiche di coltivazione possano essere pianificate per elevare al massimo la resa di prodotto economicamente utile in relazione al terreno, al lavoro e all'utilizzazione delle risorse. I principali progressi della prima metà del XX secolo sono risultati dalla migliore conoscenza del meccanismo di nutrizione delle piante.
Negli ultimi decenni si è avuto un crescente contributo da parte della biochimica vegetale e della ricerca sull'accrescimento e sullo sviluppo. Troviamo qui un importante legame con i recenti progressi nella coltivazione delle piante, che verrà trattata in uno dei paragrafi successivi (v. sotto, § b).
Durante un lungo periodo della storia dell'agricoltura si era pensato che la sostanza che costituisce il corpo delle piante derivasse dal suolo. Solo verso la fine del XVI secolo fu dimostrato da J. B. van Helmont che non si possono spiegare in questo modo gli incrementi di peso che si hanno durante la crescita, e dovette ancora trascorrere più di un secolo e mezzo prima che J. Priestley e J. Ingenhousz scoprissero la fotosintesi. La natura del contributo del suolo alla crescita delle piante fu chiarita durante il XIX secolo, principalmente da J. von Liebig, J.-B. Boussingault e J. Sachs. Come abbiamo visto precedentemente (v. tab. Il), sappiamo che sono circa 16 gli elementi necessari alle piante verdi, che, oltre l'acqua, vengono assorbiti dal suolo. L'importanza agricola della nutrizione inorganica è stata chiarita da esperimenti rigorosamente controllati, a lungo termine, condotti su appezzamenti di terreno, come quelli di J. B. Lawes e J. H. Gilbert della Rothamsted Experimental Station alla fine del XIX secolo, che cercavano di mettere in relazione, in modo quantitativo, il rendimento economico con le principali sostanze nutritizie minerali somministrate come fertilizzanti del suolo. Più tardi, la scoperta di fabbisogni micronutrizionali delle piante ha portato alla scoperta di malattie da carenza e reso possibile l'uso di semplici pratiche diagnostiche e di trattamento al fine di curarle.
Oggi, la maggior parte dei paesi progrediti ha appositi servizi per sottoporre a controllo sistematico i terreni agricoli, che forniscono informazioni analitiche dalle quali è possibile dedurre quali sostanze siano necessarie per poter ottenere un aumento del prodotto in ogni coltivazione. Sfortunatamente, in molte parti del mondo la produzione risulta scarsa non tanto per una mancanza di conoscenza delle condizioni del suolo necessarie a incrementare il raccolto, ma a causa della mancanza di risorse economiche necessarie a fornirle.
I miglioramenti nella nutrizione delle colture hanno messo frequentemente in risalto altri fattori responsabili della limitazione del prodotto, fra i quali la capacità genetica delle varietà che vengono usate, di rispondere ai trattamenti fatti. Una tendenza moderna è quella di abbinare le ricerche nutrizionali e genetiche, in modo da unire le nuove occasioni offerte da migliori metodi di coltivazione con le varietà in grado di sfruttarli.
Finora, nessun altro campo di ricerca di fisiologia vegetale ha contribuito all'incremento dei raccolti quanto gli studi sulle possibilità nutritive del suolo. Tuttavia, oggi, una migliorata economia del raccolto è il risultato dell'applicazione di ricerche provenienti da altri campi di indagini fisiologiche, fra cui la circolazione dell'acqua, l'accrescimento e lo sviluppo, e la biochimica metabolica.
La maggior parte delle piante terrestri si trova in uno stato di carenza di acqua, almeno per una parte della vita, e in alcune parti del mondo il prodotto economico delle coltivazioni è legato più alla disponibilità di acqua che a qualsiasi altro fattore. Questo fatto era conosciuto fin dai tempi della preistoria nella pratica dell'irrigazione. L'analisi scientifica della carenza d'acqua nelle piante ha avuto inizio verso la fine del XIX secolo, e con il lavoro di fisiologi vegetali come C. J. Maksimovič si è arrivati a una stima del fenomeno in termini quantitativi e a una certa comprensione degli adattamenti trovati in specie e varietà resistenti alla siccità. Più tardi i risultati di questo tipo di ricerca sono stati usati per progettare metodi di irrigazione migliori e per sviluppare migliori regimi per le piante - usando per esempio sistemi di copertura delle coltivazioni - così da fornire microclimi ottimali per determinate specie. Nuove possibilità si sono schiuse per il controllo artificiale del tasso di traspirazione, usando sostanze chimiche che influenzano l'apertura degli stomi delle foglie, sebbene questi metodi siano ancora in fase sperimentale. Vi sono stati tentativi positivi per produrre piante resistenti alla siccità, mediante un precondizionamento dei semi con soluzioni saline; anche se simili effetti sono soltanto marginali, possono tuttavia essere significativi per la produzione. La conoscenza delle caratteristiche fisiologiche e strutturali delle piante che contribuiscono alla loro resistenza è importante nella selezione per la tolleranza alla siccità.
Senza dubbio i più importanti contributi per la produttività delle piante, dalle ricerche sull'accrescimento e sullo sviluppo, si sono avuti con lo sviluppo di erbicidi selettivi, un' applicazione che deriva dai primi lavori condotti a Utrecht sulle auxine, che verranno descritti in seguito (v. sotto, § c). Altri usi economici degli ormoni di crescita delle piante sono stati trovati in agricoltura e in orticoltura. L'effetto delle auxine sull'inizio della formazione delle radici è sfruttato nella propagazione, particolarmente nel promuovere lo sviluppo delle talee da specie legnose. In alcune specie le auxine sono state usate per controllare la fioritura e per ottenere frutti senza impollinazione.
Inibitori della crescita sono usati per controllare la forma di alcune piante ornamentali, come il crisantemo, dove la stazione eretta può essere modificata causando l'aborto della gemma apicale.
Ulteriori applicazioni pratiche risultano da ricerche su altri aspetti dello sviluppo delle piante, soprattutto sul controllo della produzione di foglie, della crescita del fusto, della fioritura e della produzione di frutti. La scoperta della sensibilità fotoperiodica delle piante ha portato a una intensa ricerca sulle risposte di molte specie coltivate, fra le quali i cereali; studi simili sono stati condotti sugli effetti della temperatura, in particolare sulla necessità di trattamenti a bassa temperatura per la fioritura di cereali invernali e di molte coltivazioni biennali. L'applicazione di alcune scoperte in questo campo non è facile da realizzare per l'impossibilità di controllare le condizioni climatiche naturali. Ciò nonostante il trattamento artificiale preventivo con basse temperature dei cereali (vernalizzazione) ha acquisito, in passato, una certa importanza in Russia come mezzo per abbreviare il periodo di fioritura nelle varietà invernali, e permettere così una durata maggiore del periodo durante il quale si potevano far crescere. Più recentemente una migliore conoscenza dei parametri ambientali circa la determinazione del prodotto è stata usata per una migliore pianificazione dei sistemi di semina e di coltivazione.
Le più notevoli applicazioni di questo tipo si sono avute con la realizzazione di coltivazioni a elevata produttività, come ananas, canna da zucchero, vari ortaggi e piante da fiore. Sono stati tracciati modelli di sviluppo tali da poter fare previsioni molto precise sulla crescita che ci si può attendere in date condizioni; in base a queste previsioni si possono pianificare programmi di coltivazione intensiva.
Usando i calcolatori per paragonare il rendimento realmente ottenuto con quello teorico, si possono programmare dei trattamenti correttivi e portare all'optimum i tempi del raccolto. Il successo di tali sistemi dipende da una buona coordinazione di tutti i fattori variabili, come il genotipo delle varietà usate e i pretrattamenti dei semi, bulbi o altri propaguli, nonché le modalità di coltivazione dopo la semina. Gli ultimi sviluppi in questa direzione si possono riconoscere nei moderni metodi di produzione di alcune coltivazioni di serra, fra cui quelle ornamentali. Nella serra è possibile controllare le condizioni del suolo, la disponibilità di acqua, la temperatura, l'intensità e la durata della luce e perfino la composizione dell'atmosfera, fornendo una maggiore concentrazione di diossido di carbonio, quando questa è favorevole per le piante. Con un'appropriata conoscenza della fisiologia dello sviluppo delle piante usate si possono ottenere produzioni abbondanti, che giustificano dal punto di vista economico le spese di installazione e di manutenzione degli impianti.
Nonostante i notevoli risultati finora ottenuti, l'applicazione di una gran parte delle ricerche sulla fisiologia delle coltivazioni resta oggetto di studio futuro. Recentemente molte ricerche sono state indirizzate verso l'analisi della natura del prodotto delle coltivazioni, applicando tecniche paragonabili a quelle impiegate nell'analisi dei sistemi usati nell'industria. Parimenti a un'unità produttiva, una pianta può essere considerata come un sistema per acquistare materiali ed energia, convertirli in forma adatta, e immagazzinarli per l'uso da parte dell'uomo. Con appropriati metodi analitici, è possibile misurare l'entità dell'aumento di materiale secco a differenti stadi di crescita, determinare il percorso delle sostanze assimilate nella pianta e seguire lo sviluppo degli organi di riserva - semi, frutti, radici o altre parti - usati dall'uomo. Si può valutare l'‛efficienza' di ogni stadio e si possono identificare i fattori che limitano la resa per ogni tipo di coltivazione e per ogni ambiente. Si sono avuti molti risultati sorprendenti con analisi di questo tipo. Per esempio, si sa ora che nel grano i carboidrati che vengono portati alle cariossidi sono sintetizzati praticamente tutti nella foglia più alta dello stelo e nelle parti verdi della stessa infiorescenza. Questo significa che, a parte la funzione della radice nell'assorbire sostanze nutritive dal suolo, il resto della pianta è poco più di un sistema di sostegno autosufficiente. Inoltre, studi a livello biochimico hanno mostrato che alcune specie - soprattutto Graminacee tropicali come il granoturco - raggiungono livelli molto elevati di accumulo di materiale secco, usando una via foto sintetica diversa da quella della maggior parte delle piante dei climi temperati per la sua maggiore efficienza a concentrazioni più basse di diossido di carbonio, e anche usando una minore quantità dei prodotti fotosintetici nella respirazione, specialmente durante il giorno. Molte di queste scoperte hanno una grande importanza per la coltivazione delle piante, dato che mostrano come possono essere migliorate le rese nelle nuove varietà.
Coltivazione delle piante
Durante la maggior parte dell'attività agricola dell'uomo, il miglioramento delle coltivazioni è dipeso da una selezione per quantità e qualità del prodotto. È soltanto da un secolo e mezzo che viene praticata una selezione sistematica delle piante e solo da pochi decenni la genetica vegetale ha dato un contributo significativo. Il fatto che attualmente in molte parti del mondo non esista uno stato di carestia grave è dovuto al successo degli agricoltori, soprattutto in questi ultimi anni con lo sviluppo di varietà di cereali ad alta produttività, responsabili della cosiddetta ‛rivoluzione verde'.
Le coltivazioni in generale si basano su incroci controllati di ceppi parentali per produrre ibridi, seguiti da selezione e da ulteriori incroci e reincroci per produrre le combinazioni geniche necessarie a dare le caratteristiche richieste, come la resistenza alle malattie, la robustezza, ecc. Nelle coltivazioni di piante che si riproducono sessualmente è spesso necessario che il prodotto sia una linea pura, ma ciò non è richiesto quando la specie si riproduce vegetativamente, dato che linee fortemente eterozigoti possono essere propagate senza variazioni. Per ottenere i vari incroci e manipolarne la progenie, è necessaria una profonda conoscenza della biologia della riproduzione delle specie coltivate.
I più elaborati sistemi odierni di coltivazione si basano in gran parte sulla manipolazione citogenetica, spesso di notevole complessità. In alcune coltivazioni viene utilizzato il fenomeno della eterosi, o vigore degli ibridi: in questi casi l'agricoltore mantiene le linee pure che spesso mancano di vigore e di fertilità e ad ogni stagione produce, incrociandole con appropriate combinazioni, un seme ibrido che, una volta cresciuto, dà origine a piante caratterizzate dal vigore e dalla produttività degli ibridi di prima generazione.
L'uso di questo metodo e di altri più complicati che ne derivano, dalla metà degli anni trenta fino a ora, ha portato a un grande aumento di prodotto nelle coltivazioni del granoturco negli Stati Uniti e la stessa tecnica viene ora applicata ad altre colture.
Talvolta si possono ottenere prodotti di qualità migliore inducendo cambiamenti nel numero di cromosomi: ciò può essere realizzato usando sostanze del tipo della coichicina, che impedisce la divisione cellulare permettendo però la divisione dei cromosomi. Nella coltivazione della barbabietola da zucchero, i semi commerciali vengono prodotti incrociando piante a numero normale (diploide) di cromosomi con altre aventi un numero doppio (tetraploide); il seme che ne risulta è triploide e dà piante con un maggior contenuto di zucchero. Spesso il successo di una coltura dipende dal superamento di impedimenti all'incrocio; anche in questi casi la manipolazione del numero di cromosomi può offrire un mezzo valido sia per portare i genitori allo stesso livello, così da rendere possibile l'incrocio, sia per ridare la fertilità a un ibrido sterile.
Il contributo della fisiologia vegetale alle coltivazioni è aumentato costantemente dalla metà del XX secolo, e tutto fa supporre che continuerà ad aumentare nei prossimi decenni. A livello tecnico, una buona conoscenza della fisiologia dell'accrescimento può facilitare i processi di coltura, incrementando il numero di generazioni che si possono far crescere in un dato periodo di tempo e permettendo di ottenere la fioritura in tempo utile per rendere possibile l'incrocio di specie o varietà che normalmente non fioriscono contemporaneamente. I metodi più validi di propagazione hanno già facilitato i programmi di coltura, soprattutto delle specie legnose. I metodi di coltura delle cellule e dei tessuti possono rivoluzionare la coltivazione di alcune specie, permettendo una propagazione di massa di ceppi geneticamente uniformi di piante selezionate.
Questi contributi tecnologici sono piuttosto importanti, ma i progressi più significativi si stanno realizzando in seguito all'abbinamento di programmi di coltura con ricerche di fisiologia dell'accrescimento. Abbiamo visto che i limiti alla produzione in diversi ambienti risultano spesso evidenti quando si analizzano i fattori fisiologici che contribuiscono al prodotto. L'economia del raccolto tende a superare le limitazioni correggendo le deficienze ambientali, ma di solito queste sono insite nella stessa pianta, e in questi casi sono necessari miglioramenti nel genotipo, prima di poter utilizzare un ambiente migliore. Conoscendo quali siano le caratteristiche della pianta che ne diminuiscono realmente la resa, un coltivatore può valutare meglio il proprio progetto, e nel programmare la pianta può mettere assieme quei tratti caratteristici che, singolarmente non promettenti, forniscono nel complesso la qualità desiderata.
Egli può tener conto anche di altri importanti fattori economici, come la facilità di coltivazione della pianta e la sua adattabilità a essere trattata con mezzi meccanici. I primi risultati di questo tipo di indagine si sono visti nella coltura del grano e del riso, programmi patrocinati dalle Fondazioni Rockefeller e Ford nel Messico e nelle Filippine. Il grande pregio del nuovo grano messicano a spiga corta è quello di poter sfruttare un'intensa fertilizzazione del suolo, cosa che non avviene con le varietà precedenti. Con l'uso di questo grano, la produzione in Messico è aumentata di circa il 4oo%.
La coltura delle piante è basata, al giorno d'oggi, anche sugli studi biochimici delle qualità nutritive, del sapore, del colore e di altre proprietà. Alcune proteine delle piante non sono perfette per la nutrizione umana in quanto mancano di certi amminoacidi; le proteine del granoturco, per esempio, tendono ad avere una scarsa quantità di lisina. La produzione di granoturco ricco in lisina, realizzata dagli agricoltori alla Purdue University negli Stati Uniti, ha recato un importante contributo alla nutrizione in quei paesi dove le piante forniscono la fonte principale di proteine per la popolazione umana. La selezione basata su criteri biochimici ha avuto successo anche per l'eliminazione dei componenti tossici dai foraggi e per migliorare il loro valore nutritivo per gli animali; metodi simili sono stati usati per migliorare il sapore dei cibi umani.
Controllo delle piante infestanti e delle malattie
Un buon raccolto richiede non solo che le piante desiderate crescano in maniera efficiente, ma anche che le altre piante - competitrici o parassite - vengano eliminate. Si sono fatti, recentemente, grandi progressi per il controllo delle erbe infestanti e degli agenti patogeni delle piante; molto è dipeso dall'applicazione dei risultati di ricerche originariamente condotte con scopi completamente diversi.
Un esempio è lo sviluppo di un diserbante, simile agli ormoni, scoperto nel corso di ricerche compiute in Olanda sulle auxine. Durante la seconda guerra mondiale, W. G. Templeman scoprì che un composto sintetico, l'acido 2,4 diclorofenossiacetico (2,4-D), già conosciuto per le sue proprietà simili alle auxine, se veniva spruzzato in concentrazione appropriata, poteva uccidere le erbe infestanti a foglie larghe in un campo di cereali, senza rovinare i cereali stessi. Furono elaborati in seguito altri erbicidi selettivi, con cui è stato possibile avere dei raccolti tpuliti' e ottenere i vantaggi di una maggiore produzione eliminando la competizione per la luce, l'acqua e le sostanze nutritive del suolo. I meccanismi fondamentali della selettività d'azione degli erbicidi tipo auxine sono ancora poco conosciuti.
La più grande superficie di esposizione delle erbe a foglia larga rispetto a quelle delle erbe a foglia stretta determina differenti gradi di penetrazione delle sostanze chimiche somministrate, e ciò deve contribuire in qualche maniera alla selettività; ma allo stesso tempo si sa che piante resistenti inattivano più rapidamente le sostanze chimiche come il 2,4-D. L'azione letale in piante sensibili è dovuta probabilmente allo squilibrio causato da una stimolazione eccessiva del metabolismo in alcuni tessuti e al conseguente anormale trasferimento ed esaurimento delle riserve. Diversi erbicidi non appartenenti alla classe generale delle auxine sono stati elaborati durante questi ultimi due o tre decenni; molti hanno avuto applicazioni importanti in agricoltura e in orticoltura. Il modo di agire di alcuni di questi è stato compreso piuttosto bene: ad esempio gli erbicidi appartenenti alla classe chimica delle triazine sono inibitori della fotosintesi e la loro azione letale dipende in parte dalla capacità di distruggere questa funzione nelle foglie. La ricerca sul controllo chimico delle erbe infestanti è principalmente basata sull'effetto di trattamenti con differenti dosi di erbicidi a tempi diversi, che permettono l'eliminazione di un tipo di pianta, ma non interferiscono con la crescita di un altro. L'agricoltura più progredita e l'orticoltura ad alta produzione, sia all'aperto sia in serra, fanno oggi molto uso di questi programmi di trattamento, la cui elaborazione fino a pochi anni fa sarebbe stata considerata troppo complessa. Gran parte della ricerca è volta anche ad assicurare che questi composti chimici non siano tossici per l'uomo e per gli animali se usati in maniera appropriata, e che si tratti di sostanze che vengano facilmente degradate nel suolo e non raggiungano quindi livelli che potrebbero essere pericolosi per specie utili.
Ancora più importanti delle perdite causate all'agricoltura e all'orticoltura dall'infestazione sono gli effetti delle malattie delle piante: stime a livello mondiale rivelano che circa un quinto di tutta la produzione viene perduto in questo modo. Il fatto che la situazione non sia peggiore dipende in massima parte dal lavoro svolto dagli agricoltori per produrre varietà immuni e dai fitopatologi per individuare pratiche di coltura meno favorevoli allo sviluppo degli agenti patogeni e per approntare sistemi di controllo chimici o di altra natura.
Per un agricoltore la preoccupazione principale per certe colture è in realtà la ricerca della resistenza alle malattie. Si possono adottare due principi: uno consistente nell'introduzione in una varietà di uno o pochi geni in grado di dare un'immunità completa o parziale, l'altro nella creazione di una resistenza più o meno generalizzata con numerosi geni, lasciando spesso nel ceppo un certo grado di variabilità nella resistenza alle infezioni. Spesso la difficoltà maggiore si ha nel cercare le fonti di questa resistenza, e l'agricoltore deve talvolta risalire ai ceppi primitivi o al tipo selvatico della specie che coltiva per trovare le proprietà richieste. Dato però che anche gli agenti patogeni si evolvono, le varietà resistenti sono spesso vittime di nuove varianti di malattie, cosicché l'agricoltore deve affrontare di nuovo il problema. La tendenza moderna ad avere delle vaste colture geneticamente uniformi, al fine di ottenere produzioni molto efficienti, aumenta la probabilità di comparsa di malattie, e vi sono stati esempi di catastrofici insuccessi nella produzione dovuti a questa pratica.
Nonostante il successo dei metodi di coltura per quanto riguarda la resistenza alle malattie, spesso il controllo è basato sull'uso di sostanze chimiche: grandi progressi sono stati realizzati in questo campo da quando, nel XIX secolo, si è introdotto l'uso dei composti dello zolfo e del rame inorganico per combattere le malattie da funghi. I moderni fungicidi comprendono i tipi che agiscono in superficie, come la famosa poltiglia bordolese, contenente solfato di rame, che proteggono dall'invasione degli agenti patogeni, e i tipi sistemici che agiscono all'interno della pianta sia proteggendola da una possibile infezione, sia combattendo un'infezione già in atto. In alcune parti del mondo il carbone e altri funghi parassiti dei cereali, una volta molto diffusi, sono stati praticamente eliminati trattando i semi con composti organometallici a base di mercurio. Allo stesso modo, le misure profilattiche basate sulle applicazioni in superficie di altri fungicidi organici hanno consentito di avere sotto controllo la ruggine delle patate e altre malattie in grado di provocare una carestia. Le ricerche attuali sono dirette verso lo sviluppo di fungicidi di tipo sistemico - sostanze chimiche che vengono assorbite dalla pianta e distribuite in tutti i tessuti - assicurando così una protezione generale. Sono disponibili composti che permettono un controllo della ruggine polverosa delle coltivazioni erbacee e sono stati ottenuti risultati promettenti con altri composti per combattere la temibile ruggine del grano. Tuttavia meno positivi sono stati gli sforzi per ottenere fungicidi sistemici per combattere le malattie delle piante legnose; e le epidemie come quella che ha colpito recentemente gli olmi, eliminando alcune specie del genere in gran parte del loro territorio, sono ancora al di fuori di un efficace controllo umano.
La botanica nella medicina e nell'industria
L'interesse dei medici per le proprietà farmacologiche reali o immaginarie delle piante ha dato inizio al loro studio scientifico, e gli autori dei primi erbari sono stati per la maggior parte cultori di medicina. Oggi, malgrado la grande importanza farmacologica della chimica organica delle sostanze sintetiche, la maggior parte dei medicinali sono prodotti delle piante o loro derivati. Molti medicinali importanti provengono, come si è sempre verificato, dalla raccolta di materiale vegetale in natura, ma ormai la coltivazione di piante medicinali è diffusa in molte parti del mondo. Alcuni degli esempi più positivi di introduzione di piante nel secolo scorso si riferiscono a piante medicinali, come nel caso del chinino, introdotto nel subcontinente indiano e nelle Indie Orientali dal Sudamerica. Attualmente molti studi sulle piante sono diretti all'identificazione e alla raccolta di specie selvatiche caratterizzate da interessanti proprietà farmacologiche. Per questi lavori, le pratiche di medicina tribale hanno spesso fornito preziose indicazioni.
I progressi recenti più notevoli nell'uso medico di prodotti vegetali si sono avuti con la scoperta degli antibiotici. Il primo e più conosciuto esempio è la penicillina, un prodotto di una muffa del genere Penicillium, introdotta in medicina durante la seconda guerra mondiale. Molti lavori successivi si sono rivolti all'esame di altre muffe e di Batteri per la produzione di composti attivi e alla selezione di ceppi ad alta produttività. Più di 500 differenti antibiotici sono stati isolati da piante inferiori, per lo più da un gruppo di funghi del terreno, gli Actinomiceti, che possiedono alcune caratteristiche simili a quelle dei Batteri. Una trentina degli antibiotici conosciuti sono largamente usati come agenti terapeutici; la loro produzione mondiale si avvicina a mille tonnellate annue.
La produzione industriale di antibiotici comporta processi fermentativi di tipo simile a quelli usati per la preparazione di molti altri prodotti. La fermentazione industriale ha le sue radici nella produzione della birra, una delle più antiche utilizzazioni delle piante inferiori. Gran parte dello sviluppo tecnologico in questo campo ha comportato nuovi metodi nella coltura dei funghi, compreso l'uso dei processi continui in contrapposizione ai metodi di colture separate. La coltura dei microrganismi fornisce all'industria chimica un modo economico e concreto per la produzione di molte molecole complesse, fra cui alcune vitamine e altri composti, quali l'ormone vegetale acido gibberellico.
La silvicoltura e il miglioramento dell'ambiente umano
Probabilmente l'uomo ha trascorso la maggior parte della sua evoluzione come un animale della foresta, ma per molto tempo durante il suo periodo civilizzato ha considerato le foreste come ambienti ostili e ha dedicato molte delle sue energie a distruggerle. Nel Medio e nell'Estremo Oriente la lotta alle foreste è continuata per migliaia di anni, e in epoca storica la distruzione delle foreste è stata rapida in Europa e in altre parti del mondo con l'aumentare della popolazione. ‛Sviluppo' nelle regioni scoperte da poco dall'uomo ha generalmente significato prima di tutto la rimozione degli alberi. Solo in epoca relativamente recente l'uomo civilizzato ha riconosciuto che le regioni boscose del mondo costituiscono una risorsa importante che potrebbe alla lunga esaurirsi. Sebbene coltivazioni di alberi siano state fatte in Cina per molti secoli, è solo dal XVIII secolo che si pratica in Europa la creazione artificiale di nuove regioni boscose. Oggi la silvicoltura è un importante obiettivo in molti paesi, ma si deve riconoscere che in tutto il mondo sono state distrutte più foreste di quante ne siano state create.
Il contributo della botanica scientifica alla silvicoltura presenta molti aspetti. I boschi sono entità eterogenee, e in particolare nei tropici sono costituiti da molte specie di alberi; la tassonomia è perciò di importanza fondamentale. Lo sfruttamento razionale delle foreste come risorse di legname dipende dalla conoscenza delle proprietà del legno, e in questo caso le ricerche sull'anatomia delle piante sono risultate indispensabili. Il rimboschimento richiede, nella selezione e nella coltivazione delle piante e nella cura di appropriate condizioni di coltura, accorgimenti simili a quelli usati per l'agricoltura; in questo settore hanno recato il loro contributo la fisiologia e la genetica degli alberi.
In base alle attuali tendenze si prevede che vi saranno deficit notevoli di legname nei paesi sviluppati nei prossimi tre decenni del XX secolo. In Europa, per esempio, è probabile che la distruzione farà precipitare la produzione di circa 60.000.000 m3 nel 1975. Mentre gran parte di queste richieste saranno soddisfatte con la distruzione di foreste non ancora sfruttate di altri continenti, è evidente la necessità di trattare, a lungo termine, la silvicoltura come una produzione industriale al pari dell'agricoltura, per assicurare la continuità dei prodotti almeno nelle parti del mondo adatte a una crescita redditizia di alberi. Dato che il ciclo di coltura è così lungo, la silvicoltura di produzione comporta molti problemi particolari, alcuni dei quali di natura ecologica. La ricerca attuale affronta problemi come il vantaggio relativo di alcune piantagioni miste in contrapposizione a quelle di un'unica specie, tenendo conto delle richieste di nutrimento e di acqua, e dell'eventualità di malattie.
I botanici sono stati a lungo consapevoli dei gravi danni provocati dall'uomo, in ogni continente, alle comunità naturali, e particolarmente delle esigenze della conservazione. Negli ultimi anni le popolazioni dei paesi industrializzati sono diventate sempre più sensibili alla degradazione dell'ambiente nel mondo, e istituzioni governative e internazionali rispondono alle pressioni ora crescenti promuovendo progetti sempre più estesi di ricostituzione e di risanamento delle regioni devastate. In questi, svolge un ruolo importante la silvicoltura, non solo come mezzo di produzione del legname, ma anche come mezzo per il miglioramento dell'ambiente umano. La silvicoltura di ‛ricostituzione' presenta problemi alquanto differenti da quelli della silvicoltura di produzione, in quanto le considerazioni estetiche possono superare in importanza le richieste di una produzione redditizia con spesa non eccessiva. Spesso le migliori foreste di ‛ricostituzione' sono quelle che si avvicinano di più all'ecosistema naturale della zona considerata, proprio perché in esse si cerca di stabilire un equilibrio tra le piante e gli animali presenti. I dati ecologici sulle piante sono essenziali per proteggere e ricostruire tutti gli ecosistemi; punto questo, trattato ancora nel capitolo che segue.
La botanica: prospettive future
Molti campi della botanica, per i quali si prevede un progresso nei prossimi anni, sono stati ricordati nei capitoli precedenti. Esamineremo ora insieme alcuni dei differenti aspetti e cercheremo di prevedere quali saranno i probabili principali progressi nel prossimo futuro sia nella botanica teorica sia in quella applicata.
Abbiamo visto che la ricerca fondamentale in botanica è strettamente dipendente dall'applicazione delle nuove tecnologie, consistente principalmente nell'uso di perfezionati strumenti fisici e delle più raffinate tecniche chimiche, ma anche nell'adozione di potenti metodi di elaborazione dei dati che richiedono spesso l'impiego dei calcolatori. Nello stesso tempo è quanto mai chiaro che il progresso dipenderà dal libero interscambio tra le differenti branche.
Ciò è certamente vero per il vasto campo della biologia cellulare vegetale. L'intenso lavoro sulla biochimica metabolica svolto negli ultimi cinquant'anni ha dato origine a una complessa raffigurazione fatta di molti sistemi parziali che tengono conto di tutte le sintesi più importanti della cellula e forniscono un riassunto esauriente delle vie di flusso dell'energia. In questa raffigurazione esistono certamente delle lacune; ma una volta che esse siano colmate, è improbabile che si possano acquisire conoscenze radicalmente nuove sull'insieme delle operazioni che si svolgono nella cellula. In molti laboratori l'attenzione si è spostata dalle vie metaboliche studiate in sistemi isolati al modo in cui esse sono organizzate e controllate nella cellula vivente: tecnologicamente parlando, questo è un tipo di ricerca molto più difficile, che presenta problemi teorici estremamente gravi, perché le interazioni fra i differenti sistemi sono tali da mettere a dura prova le facoltà concettuali dell'uomo.
Il differenziamento e lo sviluppo sono tra gli argomenti più difficili di cui si è intrapreso lo studio; questi problemi sono, ovviamente, comuni a tutta la biologia. Gli importanti lavori sui microrganismi compiuti negli ultimi venti anni hanno condotto alla comprensione della biochimica dell'azione genica; si tratta ora di estendere le ricerche in maniera altrettanto rigorosa alle piante e agli animali superiori e di scoprire come l'attività genica è regolata nei sistemi molto più complicati degli organismi pluricellulari.
Vi sono stati recentemente numerosi tentativi di ideare un modello di sistemi di controllo per tali organismi, ma nessuno è stato suffragato da dati sperimentali. Una caratteristica delle piante e degli animali superiori è che il loro materiale genetico è contenuto nei cromosomi, che a loro volta sono contenuti nel nucleo: ciò rappresenta un livello di organizzazione ben più elevato rispetto a quello che si osserva nei Batteri. Inoltre le piante e gli animali superiori posseggono quantità molto maggiori di materiale genetico (DNA) nei loro nuclei rispetto a quelle presenti nelle cellule batteriche. Molti studiosi di biologia cellulare hanno fatto notare che l'eccesso di DNA non può essere completamente spiegato da una maggiore complessità metabolica, che richiede un maggior numero di enzimi e quindi di geni specifici per le proteine, perché la quantità di DNA presente è molto maggiore di quella necessaria a codificare tutti gli enzimi conosciuti nella biochimica metabolica. Secondo un'ipotesi corrente l'eccesso di DNA sarebbe in rapporto alla regolazione dell'azione genica, per la complessità dei circuiti di controllo necessari a programmare gli intimi dettagli del differenziamento e dello sviluppo osservati negli organismi multicellulari. Lo studio delle piante ha chiarito molti aspetti della struttura, della chimica e della funzione dei cromosomi, ed è ormai chiaro che in questi nuovi campi il contributo del materiale vegetale non sarà meno significativo.
A mano a mano che il punto focale si trasferisce dai sistemi biochimici isolati ai modi in cui essi sono organizzati all'interno della cellula, maggiore attenzione sarà necessariamente diretta all'architettura subcellulare, e in particolar modo agli organelli - mitocondri, cloroplasti, ecc. - e alle membrane citoplasmatiche. La cellula è un'entità suddivisa in molti compartimenti, con numerosi centri funzionali fisicamente separati. Una migliore comprensione di come le vie biochimiche sono integrate dipenderà da una migliore valutazione di come sono regolati i movimenti delle sostanze nutritive, l'energia e le sostanze di controllo di carattere ormonale, e in questo campo sono inevitabili alcune connessioni tra gli studi di biochimica, di biofisica e delle ultrastrutture.
Un gruppo centrale di problemi concerne la struttura dei vari tipi di membrane cellulari e il modo in cui essi consentono il passaggio selettivo delle sostanze. Ormai sembra chiaro che nella funzione della membrana può trovarsi la chiave di alcuni aspetti della morfogenesi cellulare, e forse anche dell'azione degli ormoni. Nelle piante, il meccanismo d'azione degli ormoni rimane uno dei campi più controversi; per questo non si può ancora dire che la funzione di ognuna delle classi di ormoni sia perfettamente compresa. Si tende a supporre che agiscano a un livello di base, forse nella regolazione dell'azione genica, o della sintesi delle proteine. Alcune molecole, di cui è nota l'attività simile a quella delle citochinine, sono presenti in una specie di RNA che agisce in un determinato stadio della sintesi proteica, e ciò può dare qualche indicazione riguardo alla funzione di questa classe di regolatori della crescita delle piante; possibili siti d'azione devono ancora essere specificati per altre classi. È certo che il chiarimento del meccanismo d'azione degli ormoni avrebbe un profondo effetto sull'intera fisiologia vegetale, e nei prossimi dieci anni ci si deve attendere una ricerca approfondita su questo argomento.
Così come è accaduto per gli aspetti fondamentali di questa scienza, è probabile che nei prossimi anni il progresso nelle applicazioni della botanica dipenda da una cooperazione interdisciplinare. Nell'agronomia ci si attende un crescente apporto di idee provenienti dalla fisiologia dell'accrescimento e dello sviluppo delle piante, specialmente per quanto riguarda l'esatto controllo della produzione del raccolto. In qual misura le tecniche già applicate in serra e nelle coltivazioni di campo in certe parti del mondo potranno estendersi ad altri tipi di operazioni agricole, dipenderà dal grado di precisione con cui potranno essere predette, e perfino controllate, alcune variabili ambientali, come il tempo. È certo che il controllo della produzione altamente automatizzato con i calcolatori offre una prospettiva attraente per soddisfare le enormi richieste di cibo e di altri prodotti vegetali prevedibili per la fine del XX secolo.
Un'area di sicuro progresso nel prossimo futuro è nell'uso di prodotti chimici, oltre quelli che forniscono il nutrimento fondamentale, per la regolazione dei processi di accrescimento nelle coltivazioni. In alcune circostanze può essere vantaggioso poter ritardare la crescita e lo sviluppo, in altre può essere vero il contrario, e sono già disponibili i mezzi per produrre questi effetti, almeno nei terreni di prova. Così, sostanze con attività ‛antigibberellinica' sono già state usate su piccola scala per ridurre l'allungamento del fusto nei cereali in cui si voleva aumentare la forza meccanica della pianta, mentre le auxine sintetiche sono state usate per causare l'inizio controllato della fioritura nell'ananas. È da notare che il crescente uso di sostanze chimiche nelle coltivazioni introduce nuovi problemi particolari a causa dei rischi dovuti alla persistenza dei residui chimici. Contemporaneamente allo sviluppo di nuovi metodi per combattere agenti nocivi e malattie, e di vie chimiche per il controllo della crescita, sarà necessario introdurre sistemi di indicatori tossicologici sia per proteggere le popolazioni umane sia per prevenire danni accidentali ad altri organismi oltre a quelli che sono obiettivo del trattamento. Ricerche ecotossicologiche sono in via di attuazione in molti paesi e sono dirette a stabilire adeguate misure di sicurezza.
Abbiamo notato precedentemente (v. cap. 4, § b) che un'efficiente produzione di piante richiede che il genotipo sia adatto ai metodi di coltivazione e ai trattamenti. La coltivazione delle piante del futuro rifletterà inevitabilmente questa filosofia di ‛sistema totale'. Per molte colture la forma tradizionale delle piante risulta non necessariamente la migliore, sia dal punto di vista dell'efficienza fisiologica, sia da quello della coltivazione e del mercato. La pianta del granoturco ne fornisce un esempio: la tendenza attuale è diretta a cambiare la posizione delle foglie per migliorare la ricezione della luce solare nell'intera coltura, così da aumentare la resa in materia secca. Il prossimo stadio può essere quello di cambiare l'architettura della pianta, ottenendo numerose spighe al posto di una sola per migliorare la distribuzione dei prodotti fotosintetici. Dobbiamo attenderci sempre maggiori richieste di ingegneri esperti di macchinario agricolo per la coltivazione di quelle piante che si trattano più facilmente con sistemi meccanici. Anche questo potrebbe dare come risultato dei cambiamenti radicali nella forma delle piante, per esempio la trasformazione in varietà coltivabili in campo dei frutti attualmente prodotti sugli alberi o in cespugli. In pochi decenni alcune piante comuni potrebbero facilmente diventare simili nell'aspetto a specie completamente differenti.
Sono da attendersi cambiamenti anche più radicali nell'utilizzazione delle piante da parte dell'uomo con l'introduzione di specie non ancora sfruttate in coltivazione. È già noto che spesso la fonte tradizionale di un genere di consumo vegetale non è necessariamente la migliore. La produzione di cellulosa dipende attualmente soprattutto dagli alberi; con l'esaurimento di foreste a legno tenero, un'alternativa sta nell'uso di varie specie di canne palustri che hanno un tasso di produzione annuale molto elevato. Queste possono essere coltivate nelle acque poco profonde lungo i margini dei laghi, e raccolte annualmente come messi. Per un altro importante prodotto vegetale, le proteine commestibili, esperimenti-pilota hanno già dimostrato che si possono estrarre direttamente dalle foglie di numerose specie perenni precedentemente non considerate di valore economico. Per certi paesi del mondo ciò potrebbe offrire un mezzo economico e concreto per la produzione di questo importante componente della razione alimentare.
Torniamo, in conclusione, al problema della degradazione dell'ambiente naturale provocata dall'aumento della popolazione umana e dal diffuso benessere. Vi sono molti aspetti di questo problema, ma dal punto di vista della botanica sono di grande interesse i cambiamenti che hanno luogo nelle associazioni vegetali in tutto il mondo. Tra i principali fattori che li causano vi sono la sterilizzazione dei terreni da parte dell'uomo per l'edificazione, la creazione di strade e l'estrazione di minerali, le pressioni risultanti dalle operazioni agricole e dallo sfruttamento delle foreste, l'inquinamento industriale e urbano, e gli effetti del turismo divenuti sempre maggiori in parti del mondo precedentemente inaccessibili e ora aperte ai viaggi. L'influenza di questi diversi fattori sta cambiando l'ecologia della vegetazione in una maniera senza precedenti, e sta causando l'estinzione di piante in tutti i continenti. Le stime basate sulle perizie patrocinate dall'Unione Internazionale per la Conservazione della Natura e delle Risorse Naturali (IUCN) fanno pensare che per le piante da fiore, su un totale di circa 25.000 specie nel mondo, una specie su dieci è attualmente esposta a qualche pericolo.
Nel 1972 si è tenuta a Stoccolma una conferenza sull'ambiente umano, organizzata dalle Nazioni Unite, nella quale si è raggiunto un accordo tra tutte le nazioni partecipanti su di una serie di importanti raccomandazioni per affrontare il problema. Queste comprendevano provvedi- menti necessari a preservare dei campioni adeguati di vegetazione naturale in riserve, e la creazione o lo sviluppo di collezioni di specie viventi e di banche dei semi per mantenere in vita specie in via di estinzione nel loro habitat naturale. Sono di particolare importanza i relativi ceppi selvatici e le razze primitive delle piante coltivate, che sono probabilmente fondamentali per il successo della coltivazione delle piante negli anni a venire. La Food and Agriculture Organisation delle Nazioni Unite (FAO) è impegnata attivamente nel tentativo di conservare queste risorse di piante selvatiche, in collaborazione con stazioni di ricerca e laboratori universitari di botanica in numerosi paesi. Con la collaborazione delle Nazioni Unite sono state create a Smirne, in Turchia, e a Bari, in Italia, delle banche di genetica delle piante. Se verranno seguite le raccomandazioni della conferenza di Stoccolma, queste installazioni saranno di modello per istituzioni simili da far sorgere altrove.
Abbiamo visto prima che la silvicoltura svolge un ruolo speciale nella politica di conservazione in molte parti del mondo, e ciò perché la vegetazione naturale dominante su gran parte della superficie terrestre è costituita da boschi. Molti studiosi di ecologia delle piante credono attualmente che sarà necessario cercare di ricostruire le foreste naturali e altri ecosistemi, se si vuole assicurare la conservazione anche solo di una parte della fauna e della flora mondiale. Queste idee ci possono fornire una prospettiva sui modi con cui l'uomo userà le piante verso la metà del XXI secolo. La produzione di cibo e di derrate sarà concentrata in aree determinate, con una meccanizzazione intensiva e con tutti gli altri accessori che serviranno a rendere massimo il prodotto e minimo il dispendio di lavoro umano. Gran parte del restante territorio verrebbe riportato ad uno stato seminaturale, con ecosistemi autosufficienti o con minimi contributi esterni che fornirebbero non soltanto le condizioni di sopravvivenza della flora e della fauna naturali, ma anche un ambiente ricreativo di grande importanza per la sopravvivenza dell'uomo stesso. La scienza botanica avrà da svolgere una parte fondamentale nell'avvento di tale mondo.
Bibliografia
Alexopoulos, C. J., Bold, H. C., Algae and Fungi, New York 1966.
Bach, P. de (a cura di), Biological control of Insect pests and weeds, London 1964.
Baker, R. St. B., Green glory, London 1958.
Bierhorst, D., Morphology of vascular plants, New York 1971.
Bold, H. C., Morphology of plants, New York 1957.
Bold, H. C., The plant kingdom, Englewood Cliffs, N. J., 1964.
Bonner, J., Varner, J. E., Plant biochemistry, New York-London 1965.
Bower, F. O., The origin of a land flora, London 1908.
Brachet, J., Mirsky, A. E. (a cura di), The cell, 6 voll., New York-London 1959-1964.
Braun-Blanquet, J., Pflanzensoziologie, Wien-New York 1964.
Cain, A. J., Foundations of plant geography, New York 1961.
Chang, J. H., Climate and agriculture. An ecological survey, Chicago 1968.
Clarke, G. L., The elements of ecology, London-New York 1965.
Clowes, F. A. L., Apical meristems, Oxford 1961.
Clowes, F. A. L., Juniper, B. E., Plant cells, Oxford 1968.
Cutter, E. G. (a cura di), Trends in plant morphogenesis, London 1966.
Darlington, C. D., The evolution of genetic systems, Edinburgh 19582.
Darlington, C. D., Chromosome botany and the origins of cultivated plants, London 19632.
Daubenmire, R., Plant communities. A textbook of synecology, London 1968.
Davis, P. H., Heywood, V. H., Principles of Angiosperm taxonomy, Edinburgh-London 1963.
Delevoryas, T., Plant diversification, New York 1966.
Dittmer, H. J., Phylogeny and form in the plant kingdom, Princeton 1964.
Edlin, H. L., Man and plants, London 1967.
Esau, K., Anatomy of seed plants, New York-London 1961.
Esau, K., Plant anatomy, New York-London 1963.
Evans, L. T. (a cura di), The induction of flowering, Melbourne 1969.
Eyre, S. R., Vegetation and soils. A world picture, London 1964.
Eyre, S. R., World vegetation types, London 1972.
Fahn, A., Plant anatomy, Oxford 1967.
Fogg, G. E., The growth of plants, London 1966.
Foster, A. S., Gifford, E. M., Comparative morphology of vascular plants, San Francisco 1959.
Frey-Wyssling, A. W., Mühlethaler, K., Ultrastructural plant cytology, Amsterdam 1960.
Gleason, H. A., Cronquist, A., The natural geography of plants, New York-London 1964.
Godwin, E., The history of the Britishflora, Cambridge 1956.
Good, R., The geography of the flowering plants, London 1964.
Gunther, F. A., Jepson, L. R., Modern insecticides and world food production, London 1960.
Hall, T., World crops. International directory and handbook, London 1967.
Heslop-Harrison, J., New concepts in flowering-plant taxonomy, London 1961.
Hillman, W. S., The physiology of flowering, New York 1965.
Hutchinson, J. (a cura di), Essays on crop plant evolution, Cambridge 1965.
Hutchinson, J. (a cura di), Population and food supply, Cambridge 1969.
Jacks, G. V., Whyte, R. O., The rape of the earth. A world survey of soil erosion, London 1940.
King, L. J., Weeds of the world. Physiology and control, London 1966.
Kreig, M., Green medicine, New York 1964.
Ledbetter, M. C., Porter, K., Introduction to the fine structure of plant cells, Berlin-New York 1970.
Leopold, C., Plant growth and development, New York 1961.
Linskens, H. F. e altri (a cura di), Encyclopaedia of plant physiology, 18 voll., Berlin-New York 1951-1967
Löve, A., Löve, D., North Atlantic biota and their history, London 1963.
Nobel, W., Plant cell physiology, San Francisco 1972.
Oosting, H. J., The study of plant communities. An introduction to plant ecology, San Francisco 1958.
Phillips, I. D. J., Introduction to the biochemistry and physiology of plant growth hormones, New York 1971.
Polunin, N., Introduction to plant geography, London 1960.
Robards, A. W., Electron microscopy and plant ultrastructure, New York 1970.
Russell, E. W., Soil conditions and plant growth, London 19639.
Scagel, R. F., Bandoni, R. J., Rouse, G. E., Schofield, W. B., Stern, J. R., Taylor, T. M. C., An evolutionary survey of the plant kingdom, London-Glasgow 1965.
Schery, R. W., Plants for man, London 1954.
Seward, A. C., Plant life through the ages, Cambridge 1941.
Singleton, W. R., Elementary genetics, Princeton 19672.
Sinnott, E. W., Plant morphogenesis, New York 1960.
Sporne, K. R., The morphology of Gymnosperms, London 1965.
Sporne, K. R., The morphology of Pteridophytes, London 1966.
Spurr, S. H., Forest ecology, New York 1964.
Srb, A. M., Owen, R. D., Edgar, R. S., General genetics, San Francisco 19652.
Stackman, E. C., Bradfield, R., Mangelsdorf, P. C., Campaigns against hunger, Cambridge, Mass., 1967.
Stebbins, G. L., Chromosomal evolution in higher plants, Reading, Mass., 1971.
Steward, F. C., Plant physiology. A treatise, 6 voll., New York-London 1961-1972.
Steward, F. C., Growth and organisation in plants, Reading, Mass., 1968.
Street, H. E., Plant metabolism, Oxford 1966.
Swain, T. (a cura di), Plantsin the development of modern medicine, Cambridge, Mass., 1972.
Tachtadžjan, A., On the origin of the temperate flora of Eurasia, in ‟Botanical journal", 1957, XLII, pp. 1635-1653.
Taylor, N., Drug plants that changed the world, London 1966.
Thomas, W. L. (a cura di), man's role in changing the face of the earth, Chicago 1965.
Torrey, J., Development in flowering plants, New York 1967.
Walter, H., Ecology of tropical and subtropical vegetation, Edinburgh 1971.
Wardlaw, C. W., Essays of form in plants, Manchester 1968.
Wardlaw, C. W., Morphogenesis in plants, London 1968.
Wareing, P. F., Philipps, I. D. J., The control of growth and differentiation in plants, Oxford 1970.
Weaver, J. E., Clements, F. E., Plant ecology, New York 1929.
Weevers, T., Fifty years of plant physiology, Waltham, Mass., 1949.
Whittingham, C. P., The chemistry of plant processes, London 1964.
Whyte, R. O., Crop production and environment, London 1946.
Wilkins, M. B. (a curadi), Physiology of plant growth and developpement, London 1969.
Wulff, E., Historical plant geography, Waltham, Mass., 1945.
Zeuner, F. E., Dating the past, London 1952.