
Evoluzione, coevoluzione e biodiversita
Evoluzione, coevoluzione e biodiversità
La speciazione prodotta da due popolazioni conspecifiche e dal loro conseguente isolamento genetico è un fattore evolutivo di particolare rilevanza. Essa può anche verificarsi all'interno di una stessa area geografica e di una stessa popolazione. La coevoluzione è un processo per cui l'evoluzione di più partner (animali, piante, batteri o funghi), dipende dalla loro interazione reciproca. Entrambi i partner acquisiscono adattamenti specifici a seguito di pressioni mutualmente selettive. I processi evolutivi producono diversità biologica, cioè biodiversità; in questo saggio verranno definiti il concetto di biodiversità e i livelli gerarchici delle varie forme di organizzazione biologica che vanno incontro al differenziamento. Si considereranno, inoltre, le diverse componenti della diversità, lo varietà delle interazioni tra organismi, i fattori biologico-evolutivi ed ecologici che causano lo diversità, e i processi funzionali della diversità. Una particolare attenzione sarà dedicata alla diversità tra le biocenosi e alloro interno e verrà evidenziato come lo preservazione della biodiversità sia un prerequisito essenziale per lo sopravvivenza dell'uomo sulla Terra.
Principi fondamentali
La teoria dell'evoluzione biologica ha influenzato in modo radicale e duraturo la nota concezione del mondo anche a livello umanistico, sociale e religioso. La concezione antropocentrica del mondo è stata turbata per la prima volta da Nicola Copernico nel 16° secolo, il quale spostò la Terra - e quindi l'uomo - fuori dal centro dell'Universo. La teoria dell'evoluzione del naturalista inglese Charles Darwin sconvolse ulteriormente la visione antropocentrica, asserendo che la specie Homo sapiens è in stretto rapporto evolutivo con tutti gli altri organismi viventi. L'evoluzione biologica, la coevoluzione tra le specie e il fenomeno della biodiversità (che è il risultato dei processi evolutivi) sono strettamente correlati da un punto di vista causale. Qui di seguito verranno esposti i fattori che rappresentano prerequisiti essenziali per i processi evolutivi.
Energia. - La vita non sarebbe possibile senza il Sole come fonte extraterrestre di energia. Lo sviluppo di composti chimici di più alta complessità determinati dalla fotosintesi, ovvero la produzione primaria degli organismi autotrofi (produttori) - senza i quali i consumatori e i decompositori (animali, batteri, funghi) non si sarebbero potuti evolvere - e la temperatura favorevole per i processi metabolici degli organismi viventi sono prerequisiti essenziali per la vita. Anche la varietà ordinata delle diverse forme di vita (biodiversità) all'interno delle biocenosi e degli eco sistemi, rispecchiata dalle numerose specie animali e vegetali che li compongono, si basa su un rifornimento costante di energia.
Le leggi della termodinamica si applicano sia alla materia inorganica che organica. L'ordine, espresso in termini di negentropia, all'interno di un sistema (in questo caso un bio sistema) è possibile solo quando vi sia energia disponibile, ovvero vi sia rifornimento di entalpia libera. Senza rifornimento di energia le diverse componenti del sistema, ovvero i suoi compartimenti, non possono assicurare alcun trasferimento di materia o informazione. Questo trasferimento è necessario per il funzionamento dell'intero sistema, che si auto organizza grazie alle caratteristiche intrinseche al sistema stesso, ovvero grazie alle interazioni esistenti all'interno dei suoi compartimenti. Una minore disponibilità di energia libera conduce a un aumento di entropia interna al sistema e alla sua disorganizzazione.
Gli organismi e lo loro informazione genetica. - Il codice genetico (DNA) degli organismi contiene informazioni che consentono la preservazione dei processi vitali specifici di ciascuna specie od organismo, e l'organizzazione generale del metabolismo costruttivo ed energetico. Il codice inoltre permette all'organismo di rispondere alle sollecitazioni dell'ambiente, nonché la trasmissione dell'informazione genetica attraverso le generazioni.
Strutture e funzioni degli organismi. - Gli organismi sono caratterizzati da strutture specifiche. Per riuscire a sopravvivere in un determinato ambiente abiotico e biotico gli organismi acquisiscono specifici adattamenti morfologici, fisiologici, biochimici ed etologici. La grande varietà e diversità degli ambienti terrestri comporta l'esistenza di una grande diversità di strutture. Determinate strutture degli organismi sono di regola in rapporto con specifiche funzioni (adattamento), e viceversa.
Mutazione. - Le mutazioni sono la base dell'evoluzione. Una mutazione consiste in un cambiamento nel genotipo, sia esso spontaneo o indotto da particolari agenti mutageni (sostanze che provocano mutazioni, come le alte temperature o i raggi ultravioletti).
Selezione. - Le mutazioni determinano l'esistenza di un' ampia varietà di fenomeni e forme possibili, tra le quali la selezione 'sceglie' quelle che sono meglio conformate alle condizioni ambientali prevalenti. Queste forme si riproducono meglio (hanno una fitness maggiore) e quindi passano una maggiore quantità di alleli alla generazione successiva.
Tempo. - La storicità dei sistemi tassonomici è una delle caratteristiche fondamentali della biologia. La biodiversità è il prodotto dell'evoluzione e quindi il risultato dell'effettiva filogenesi attraverso milioni di anni. La legge di Dollo sull'irreversibilità dei processi storico-evolutivi implica che una specie possa avere origine solo una volta. Strutture complicate che si sono perse nel corso dell'evoluzione non possono più riapparire nella loro forma originale, dato che le combinazioni di mutazioni casuali e selezione direzionale sono per definizione non riproducibili. Ci sono comunque esempi che dimostrano come vie metaboliche complesse, che si osservano per esempio in gruppi filo geneticamente vicini, siano state in molti casi acquisite attraverso stadi più semplici di mutazione e selezione, come avviene nel caso del metabolismo acido delle crassulacee (CAM) nelle piante.
Illustreremo ora alcuni principi basilari dell'evoluzione biologica. Il processo evolutivo può essere considerevolmente accelerato, e il grado degli adattamenti coordinati può essere particolarmente pronunciato, quando le specie o i gruppi di specie esercitano una pressione selettiva significativa una rispetto all'altra (coevoluzione). Alcuni fenomeni particolarmente spettacolari sono il risultato di una coevoluzione. Le forme della diversità verranno descritte in rapporto alle gerarchie dei sistemi biologici (specie, bioceno si, eco sistema, paesaggio).
Evoluzione
Teoria dell'evoluzione
La teoria dello sviluppo filogenetico degli organismi, delineata per la prima volta da Darwin nella sua opera fondamentale del 1859 The origin of species (L'origine delle specie), è nei suoi tratti essenziali valida ancora oggi, ed è stata corroborata da dati provenienti da quasi tutti i campi della biologia.
La teoria si fonda su due assiomi fondamentali: tutti gli organismi sono filogeneticamente correlati tra loro; l'evoluzione è determinata da cambiamenti non direzionali del materiale genetico (le mutazioni), che conducono a trasformazioni di forma, funzione e modalità di vita degli organismi (le specie), e dalla selezione direzionale operata dalle componenti biotiche e abiotiche dell'ambiente.
La teoria della selezione di Darwin (che nel caso ci si riferisca alla filo genesi viene designata come teoria dell'evoluzione o teoria della discendenza), contraddice la tesi dello zoologo francese J.-E. de Lamarck, definita nel 1809, secondo la quale l'evoluzione è determinata da cambiamenti nel fenotipo che vengono trasmessi alle generazioni successive. La teoria della selezione naturale implica invece che i processi evolutivi siano generati da una riproduzione differenziale di differenti genotipi. In modo totalmente indipendente da Darwin, nello stesso periodo, il biogeografo A.R. Wallace defrnÌ gli stessi meccanismi principali che sono alla base dei processi evolutivi.
La genetica moderna, e in particolare la genetica di popolazione, ha contribuito a una migliore comprensione dei processi di mutazione e di variazione genetica, e molti dati hanno confermato e so stanziato la teoria dell'evoluzione. Discipline come l'ecologia, l'etologia, la biologia dello sviluppo, la sistematica, la zoologia e la botanica, la paleontologia hanno compiuto notevoli progressi, sviluppando nel contempo procedure matematiche appropriate e specifiche. Di conseguenza, la biologia evolutiva si è potuta affermare come scienza sintetica (Huxley, 1974).
Uno degli aspetti più importanti delle attuali ricerche è costituito dalla ricostruzione dei processi storici filo genetici e dallo studio dei fattori che li hanno indotti.
Evidenze dell'evoluzione
Importanti compiti delle ricerche in campo evolutivo sono l'analisi dei cambiamenti all'interno delle specie, la ricostruzione degli eventi che hanno caratterizzato la filo genesi (studi degli alberi filogenetici) e la dimostrazione dei fattori che determinano l'evoluzione (studi di causalità evolutiva). Verranno ora descritte le conoscenze ormai consolidate attraverso le quali è stata dimostrata la validità della teoria evolutiva.
Ricerca delle omologie. - Gli organismi possono essere distinti gli uni dagli altri in base alla presenza di determinati organi e strutture. Si definiscono omologhi quegli organi e quelle strutture le cui somiglianze sono dovute alla condivisione delle stesse informazioni ereditarie. Dato che gli organi omologhi cambiano nel corso dell'evoluzione, la loro identificazione e il loro studio devono essere affrontati seguendo precisi criteri (Osche, 1966), quali il criterio della posizione, il criterio della continuità o della stabilità e il criterio della qualità specifica. Il primo stabilisce che organi e strutture sono omologhi quando occupano la stessa posizione all'interno dell'organismo (organi omotopici), come, per esempio, le diverse parti (labium, labrum, mandibola e mascella) della bocca degli insetti. Il secondo stabilisce che l'omologia di organi e strutture che non sono simili né occupano la stessa posizione nell'organismo (organi eteromorfi o eterotopici) può essere identificata attraverso lo studio di forme intermedie o di transizione, ovvero:
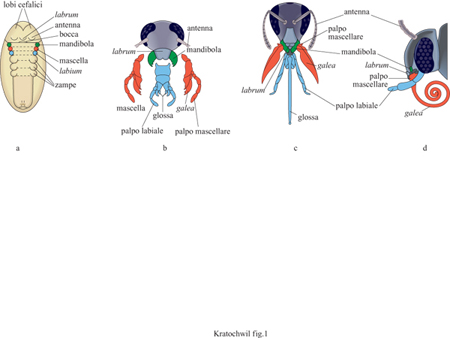
l) studiando lo sviluppo embrionale. Per esempio, la conversione delle appendici omologhe seriali disposte secondo un andamento segmentale (metamerico) in parti boccali e arti nell'embrione di insetto (fig. 1).
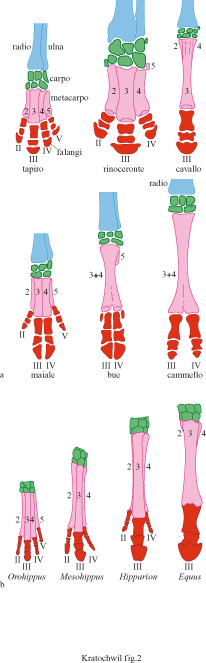
2) Comparando forme viventi o fossili strettamente imparentate. Ne è un esempio la riduzione delle dita che si osserva attraverso il tempo nelle serie fossili dei mammiferi ungulati (fig. 2).
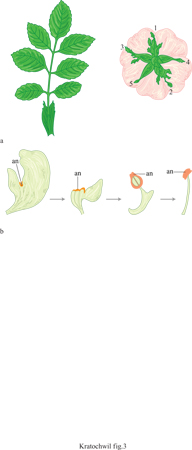
3) Comparando organi che ricorrono serialmente in uno stesso individuo. È il caso della trasformazione da foglie normali a sepali che si verifica per una progressiva riduzione della foglia superiore, oppure della transizione da petali a stami (fig. 3).
Il terzo criterio, quello della qualità specifica, afferma che, indipendentemente dalla loro posizione all'interno dell'organismo, l'omologia di alcune strutture complesse può essere riconosciuta quando siano in entrambe riconoscibili le stesse singole parti che la compongono (organi omomorfi), come, per esempio, i denti dei vertebrati.
Oltre che ai caratteri morfologici, il concetto di omologia può anche riferirsi ad aspetti biochimici (confronto di enzimi, ormoni, DNA, ecc.) ed etologici (quale il confronto tra le vocalizzazioni innate nelle varie specie animali).
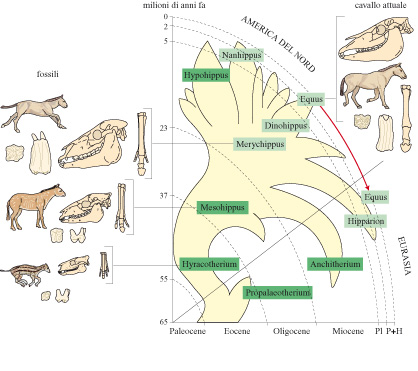
Dati paleontologici. - I fossili costituiscono importanti testimonianze che consentono di ricostruire l'evoluzione di specifici gruppi di organismi. In alcuni casi è stato addirittura possibile tracciare in modo preciso la sequenza di adattamenti e di fenomeni speciativi, come nel caso dell'evoluzione degli equidi durante i periodi Terziario e Quaternario (fig. 4).

Le forme di transizione giocano un ruolo particolarmente importante quando si confrontano livelli più elevati di organizzazione gerarchica. Ne sono un esempio il fossile /chthyostega del Devoniano, che presenta caratteristiche comuni ai pesci e agli anfibi (fig. 5), o Archaeopteryx, fossile del Giurassico, che possiede caratteri comuni ai rettili e agli uccelli.
Dati biogeografici. - Molti gruppi di organismi sono attualmente confinati in determinate aree geografiche. Probabilmente si sono originati nelle aree in cui attualmente vivono, e barriere naturali, come oceani e montagne, ne hanno impedito un'ulteriore dispersione. Questo spiegherebbe la loro assenza in altre regioni.

Specie che si ritrovano solo in aree limitate sono defrnite endemiche. Le isole continentali sono particolarmente ricche di endemismi, come per esempio l'Australia con la sua fauna di mammiferi marsupiali. Lo stesso fenomeno si verifica nelle isole più antiche, spesso di origine vulcanica, come le isole Galapagos, con la loro fauna (fig. 7) di fringuelli di Darwin (sotto famiglia geospizini; Lack, 1947), o le isole Hawaii con i suoi uccelli drepanidi (famiglia drepanididi; Mayr, 1943).
Si deve però distinguere tra endemismi originari, per i quali il centro di origine e la distribuzione attuale coincidono (per esempio, i fringuelli di Darwin), ed endemismi relitti, la cui attuale distribuzione rappresenta un frammento di un'area anticamente molto più estesa.
Organi rudimentali. - Le continue modificazioni di organi e strutture che si verificano nel corso dell'evoluzione sono spesso accompagnate da cambiamenti nella loro funzione. Gli ossicini dell'orecchio medio dei mammiferi, per esempio, derivano da ossa che facevano (e fanno tuttora) parte dell'articolazione della mandibola degli anfibi, dei rettili e degli uccelli. Esistono oltre alle testimonianze di tali cambiamenti casi di organi che non possiedono più una funzione, e sono ormai ridotti a rudimenti (organi rudimentali o vestigiali). Questi altro non sono che una testimonianza di 'adattamenti del passato'.
Esempi di organi rudimentali nella fauna attuale sono rappresentati dalle piccole ossa residuali dell'apparato appendicolare dei rettili striscianti, che non sporgono più all'esterno. Un pitone, per esempio, possiede ancora un rudimento di cinto pelvico in corrispondenza delle zampe posteriori, anche se queste non sono più presenti.

Le balene (misticeti) mancano completamente delle parti esterne degli arti posteriori, ma internamente al corpo possono riscontrarsi rudimenti del femore e del cinto pelvico (fig. 8). Altri esempi sono gli occhi rudimentali degli animali troglobi che vivono perennemente nelle grotte (è il caso del pesce Anoptichthys jordani, della famiglia caracidi) o di quelli che vivono in ambienti sotterranei (è il caso del ratto talpa Spalax typhlus, roditore della famiglia spalacidi).
Esempi di organi rudimentali nelle piante sono la permanenza all'interno del fiore dei cinque stami superiori, che non sporgono più all'esterno, nelle specie del genere Scrophularia (famiglia scrofulariacee), o la presenza di residui di stami nei fiori diclini (che si sono evoluti a partire da fiori originariamente completi), o anche la presenza di stami non più necessari - in alcune piante acquatiche.
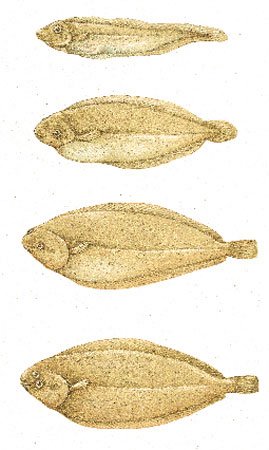
Testimonianze fornite dallo studio dello sviluppo embrionale. - Nel corso dello sviluppo embrionale (ontogenesi) in molti organismi si formano organi e strutture che poi regrediscono completamente nel corso dello sviluppo (sviluppo indiretto), e molti di questi corrispondono a organi e strutture tipiche dei loro progenitori ancestrali. Per esempio, nelle sogliole (pesci appartenenti all'ordine dei pleuronettiformi, come i generi Scophthalmus, della famiglia scoftalmidi, e Pleuronectes, della famiglia pleuronettidi) le larve appena uscite dall'uovo possiedono una simmetria bilaterale, e solo successivamente divengono asimmetriche (fig. 9). Le balene prive di denti (sottordine misticeti) catturano il cibo, rappresentato da piccolissimi crostacei, intrappolandolo in una struttura filtrante particolare, i fanoni. Tuttavia durante lo sviluppo embrionale si sviluppano dei rudimenti di denti, che successivamente regrediscono. Ciò è in relazione con il fatto che il loro antenato possedeva denti come le attuali balene fornite di denti (sottordine odontoceti), quali per esempio il delfrno (famiglia delfinidi).
Tra le piante, l'albero della vita (genere Thuja, famiglia cupressacee) ha foglie piccole e a forma di scaglia, ma negli stadi giovanili si sviluppano prima i lunghi aghi tipici delle specie originarie delle conifere (Coniferophytina).
Adattamenti all'ambiente. - L'evoluzione documenta la varietà dei possibili adattamenti degli organismi a determinati fattori ambientali, sia nello spazio sia nel tempo. Gli adattamenti sono essenziali all'organismo per sopravvivere e riprodursi in un ambiente specifico per assicurare la conservazione della specie.
L'ambiente viene in questo contesto recepito come un complesso di fattori esterni (eco fattori) che influenzano un organismo, direttamente e indirettamente. Degli ecofattori fanno parte sia i fattori essenziali per la sua sopravvivenza (ambiente minimo) sia le influenze ambientali aggiuntive (ambiente ecologico). L'ambiente di un organismo comprende quindi tutti i fattori biotici e abiotici che hanno su di esso un influenza negativa o positiva all'interno dell 'habitat che questo ha colonizzato.
Fattori ambientali abiotici (fisiografici). - L'ambiente abiotico è caratterizzato da alcuni fattori: i fattori climatici (temperatura, luce, umidità, precipitazioni, vento, correnti, ecc.), quelli edafici (proprietà chimiche e fisiche del suolo), quelli orografici (geomorfologia e paesaggio), inclusi l'esposizione (posizione relativa a un punto cardinale) e l'inclinazione (pendenza).
Fattori ambientali biotici. - L'ambiente biotico di un organismo è determinato dal tipo di interazioni tra l'organismo stesso e gli organismi appartenenti ad altre specie, caratteristiche della comunità in cui vive. Tali interazioni comprendono il mutualismo (simbiosi nel senso più ampio), la predazione, il parassitismo, la competizione intraspecifica (competizione all'interno di una specie) e la competizione interspecifica (competizione tra specie diverse).
Nicchie ecologiche. - La nicchia ecologica di una specie caratterizza le relazioni tra una specie e il suo ambiente specifico. Non è concepita come una unità spaziale, ma piuttosto come un sistema dinamico di relazioni tra le potenzialità di una specie e le caratteristiche dell'ambiente in cui queste potenzialità si possono esprimere (Hutchinson, 1965). La nicchia ecologica include quindi una dimensione autofitica/autozoica e una dimensione ambientale. La dimensione autofitica/autozoica comprende le caratteristiche morfologiche e fisiologiche che sono state acquisite nel corso dell'evoluzione (filogenetiche), mentre la dimensione ambientale rappresenta la somma di tutti i fattori ecologici effettivi all'interno di un habitat specifico. La nicchia ecologica di una specie si realizza nel punto di sovrapposizione di queste due componenti (Schmitt, 1987).
Fattori evolutivi: mutazione, ricombinazione e selezione
Come già in parte discusso, la mutazione e la selezione sono importanti fattori evolutivi. Per la loro funzione cruciale è opportuno entrare in alcuni dettagli sul loro funzionamento e accennare a un altro fattore essenziale dell'evoluzione, la ricombinazione.
La mutagenesi. - l geni situati sui cromosomi determinano i caratteri ereditari degli organismi e nel loro insieme costituiscono il genoma. Secondo il grado di ploidia del suo portatore, un gene può avere molti alleli. ln un assetto diploide di cromosomi, due alle li determinano la qualità di un carattere. Una caratteristica di un organismo può essere determinata dall'azione combinata di numerosi geni (poligenia), ma è anche possibile che un gene influenzi lo sviluppo di molti caratteri (pleiotropia).
Cambiamenti nel genotipo possono essere causati da mutazioni spontanee (mutazioni casuali) o provocate da agenti mutageni (quali le alte temperature, le radiazioni a onde corte, o alcuni composti chimici). Le mutazioni possono essere cito logicamente suddivise in quattro gruppi: mutazioni genomiche, cromosomiche, geniche e punti formi; quest'ultima consiste nel cambiamento di una sola base nelle catene degli acidi nucleici che compongono il genoma. Si definisce mutagenesi il cambiamento dei geni attraverso le mutazioni. Dato l'enorme numero di geni di un organismo (da centomila fino a oltre un milione), la probabilità che avvenga una mutazione è piuttosto elevata, anche a tassi medi di mutazione di 10-⁴ ÷10-⁶ mutazioni/gene per ciascuna generazione. Per esempio, il 2 ÷ 3% degli individui di ciascuna generazione del moscerino della frutta (Drosophila) è rappresentato da forme mutate. Negli esseri umani, si verifica una mutazione ogni 100.000÷200.000 duplicazioni geniche. Questo significa che ogni essere umano possiede in media 1 ÷ 2 alleli mutati rispetto alla generazione parentale.

Le mutazioni sono responsabili del mantenimento di una certa variabilità genetica all'interno del genoma di una popolazione (pool genico). Una popolazione è definita come un gruppo di individui della stessa specie che formano, nello stesso momento (sincronicamente) e nella stessa unità spaziale (sintopicamente), una potenziale comunità riproduttiva. Le mutazioni forniscono cioè il 'materiale grezzo' per l'evoluzione (fig. 10).
La ricombinazione. - Per ricombinazione si intende lo scambio di geni tra gli alleli che si verifica nel corso della riproduzione sessuale (meiosi e fusione di gameti, ovvero singamia). l nuovi genotipi che ne derivano estendono significativamente la variabilità genetica di una popolazione. La ricombinazione è un processo relativamente casuale.
I fattori selettivi. - La selezione 'assicura le carriere' (il successo) dei genotipi, nel senso che quelli meno adatti sono ostacolati. l genotipi più adatti possono passare un' alta percentuale dei loro alleli e geni alla generazione successiva (possiedono cioè una fitness elevata). La selezione stabilizza o altera la frequenza di determinati geni all'interno di una popolazione (selezione stabilizzante e selezione direzionale). I fattori selettivi includono le condizioni climatiche (quali le basse temperature o la siccità), la competizione per il cibo, la competizione per lo spazio, i nemici e i parassiti, alcuni nutrienti per le piante (per esempio, l'azoto) e altri ancora.
Un brillante esempio di pressione selettiva dovuta all'attività antropica è dato dal caso della falena Biston betularia (famiglia geometridi) Nella sua forma normale, questa falena è difficilmente individuata quando riposa con le ali distese sulla corteccia di un tronco rivestito di licheni. Attraverso questo meccanismo di camuffamento con il paesaggio circostante (mimetismo) riesce a sfuggire ai suoi nemici, gli uccelli insettivori. A causa dell'inquinamento dell'aria dovuto all'emissione di composti solforosi nelle aree industriali della Gran Bretagna, la vegetazione di licheni è andata via via scomparendo, e la corteccia dei tronchi si è ricoperta di fuliggine scura. Nel 1848 a Manchester, in Gran Bretagna, fu scoperta per la prima volta una forma di colore scuro (melanica) della falena (Biston betularia carbonarius). Il vantaggio selettivo (dovuto a una migliore protezione dai predatori sul fondo ormai scurito dei tronchi) ha condotto a un chiaro aumento di questa forma rispetto a quella chiara: 50 anni più tardi, il 95% di tutta la popolazione (dal 1952 al 1956 fino al 98%) consisteva di forme carbonarius (Kettlewell, 1972). Miglioramenti nel campo delle tecnologie di protezione ambientale hanno nel recente passato favorito la rigenerazione della vegetazione dei licheni e quindi un rinnovato vantaggio selettivo della forma chiara della farfalla.
Organi e strutture appartenenti a gruppi di organismi che non hanno una stretta relazione filo genetica possono trovarsi nella condizione di dover realizzare le stesse funzioni, e di conseguenza mostrano adattamenti simili. Il fenomeno per cui strutture e organi che hanno origini diverse divengono via via più simili nel corso dell'evoluzione è definito convergenza. Un esempio di convergenza nelle piante è rappresentato dai fusti succulenti (sviluppatisi per rispondere alla necessità di creare riserve d'acqua che assicurino la sopravvivenza in ambienti aridi), che si ritrovano nelle asclepiadacee, nelle composite, nelle euforbiacee, nelle cactacee e nelle didieracee.

Negli animali, un perfetto esempio di convergenza è rappresentato dalla forma affusolata del corpo di diversi vertebrati che vivono nell'acqua: elasmobranchi (squali), pesci ossei (pescispada), rettili fossili (ittiosauri), uccelli (pinguini) e mammiferi (delfini) (fig. 12). Particolarmente interessanti sono i fenomeni di convergenza che si osservano tra i mammiferi placentati e i marsupiali, la cui distribuzione è principalmente limitata all'Australia.
Speciazione per separazione e isolamento
Cos'è una specie? - Esistono diverse definizioni di una specie biologica: quella che si fonda su un concetto morfofisiologico (morfospecie) che definisce una specie come il complesso di individui (inclusi i loro discendenti) le cui caratteristiche essenziali (morfologiche, fisiologiche, ma anche etologiche) sono identiche. Questa definizione è l'unica applicabile ai fossili.
Un'altra definizione, che si fonda su un concetto genetico (biospecie), intende una specie come costituita da un insieme di popolazioni effettivamente o potenzialmente interfeconde, che sono riproduttivamente isolate da altri gruppi di popolazioni, ovvero tra i quali non esiste scambio di geni (comunità riproduttiva).
Un'ultima definizione, che si fonda su un concetto ecologico (ecospecie), parte dalla considerazione che ciascuna specie esprime richieste specifiche all'ambiente in cui vive, in parte rappresentate dalla sua nicchia ecologica. Specie che vivono sintopicamente e sincronicamente non possono appartenere alla stessa nicchia ecologica.
Speciazione omogenetica e cladogenetica. - A parte la speciazione allocronica che si realizza lungo una dimensione storica con la progressiva trasformazione di una specie attraverso un certo numero di generazioni (anagenesi), esiste un processo di speciazione sincronica che corrisponde alla separazione di una specie in due specie sorelle (cladogenesi).
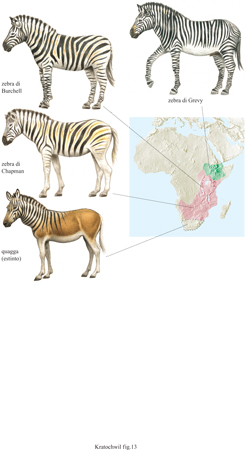
Prima fase della speciazione allopatrica: la separazione. - Il primo stadio nella formazione allopatrica di una specie consiste nella separazione (geografica) di una popolazione dalla popolazione parentale. Ciò può essere dovuto a una molteplicità di cause: alcuni individui che superano determinate barriere geografiche (montagne, deserti, mari); cambiamenti macroclimatici nel corso dei periodi geologici (per esempio, le ere glaciali) che forzano le specie a rifugiarsi in aree sparse dove le condizioni climatiche sono più tollerabili (aree rifugio); un aumento dellivello del mare che comporta la separazione delle terre continentali in aree frammentate (formazione di isole continentali). La variabilità limitata dei genotipi (dovuta al fatto che nelle popolazioni così isolate si ritrova solo una porzione della popolazione di origine), nuove mutazioni, e diverse condizioni ambientali (fattori selettivi) possono successivamente condurre alla speciazione. L'importanza della separazione geografica è so stanziata dal fatto che specie con un'ampia distribuzione formano sotto specie geografiche. Le sottospecie differiscono leggermente nella loro composizione genica e fenotipica, e ciascuna occupa una certa sottoregione geografica all'interno dell'areale di distribuzione della specie. Valgano come esempio le diverse specie e sotto specie di zebra (fig. 13).
Seconda fase della speciazione allopatrica: lo sviluppo dei meccanismi di isolamento riproduttivo. - In seguito alla separazione geografica di una popolazione precedentemente continua in due o più popolazioni isolate, si sviluppano meccanismi che impediscono il successivo reincrocio e ibridazione delle popolazioni separate, che potrebbe avvenire lungo una zona di contatto secondario in seguito a una reintrogressione.
Si possono così sviluppare: meccanismi di isolamento postzigotici, che rendono incompatibili i genomi successivamente all'accoppiamento o meccanismi di isolamento prezigotici, che invece prevengono l'accoppiamento stesso.
Esempi di meccanismi di isolamento prezigotici sono: l'isolamento ciclico o stagionale, che consiste nella differenziazione dei periodi riproduttivi (come avviene per esempio per le farfalle), o dei periodi di fioritura delle piante; l'isolamento meccanico, che comporta una diversità nelle strutture degli organi copulatori; l'isolamento etologico in cui i cambiamenti dei segnali visivi, acustici e olfattivi permettono agli animali di riconoscere il proprio partner per l'accoppiamento (come avviene per esempio tra scimmie, uccelli, locuste, farfalle) e, nel caso della zoogamia, permettono alle piante di scegliere i loro impollinatori.
Speciazione simpatrica. - La formazione simpatrica di specie si verifica tra gli individui di una stessa popolazione all'interno della stessa area di distribuzione, in assenza cioè di barriere geografiche. Questo tipo di speciazione è particolarmente frequente nelle piante. Un corredo di cromosomi viene duplicato (autopoliploidia), cosicché l'individuo poliploide neoformato resta isolato dal resto degli individui diploidi (come per esempio molte felci). Circa un terzo di tutte le specie di piante si sono originate per poliploidia. Specie poliploidi strettamente imparentate possono in taluni casi incrociarsi e dare origine a nuove specie allopoliploidi che, a loro volta, sono geneticamente isolate dalla specie originaria. Molte delle nostre piante coltivate (il cotone, del genere Gossypium, o molti cereali) sono state create per allo- o autopoliploidia. Al contrario, tra gli animali la poliploidia è un fenomeno raro.
Macroevoluzione
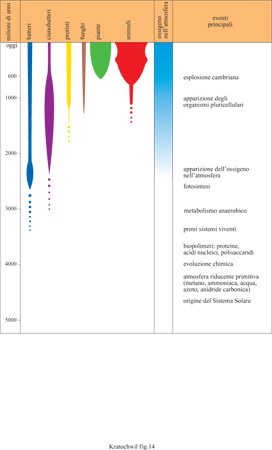
La Terra ha avuto origine circa 5 miliardi di anni fa e le prime forme di vita circa 3,8 ÷ 3,5 miliardi di anni fa. Lo sviluppo dei procarioti (cianobatteri e batteri) è iniziato circa 3,8 miliardi di anni fa. Le prime spugne sono comparse circa 570 milioni di anni fa, i pesci circa 480 milioni, i rettili circa 280 milioni, i mammiferi circa 200 milioni, gli uccelli circa 150 milioni, e i primati circa 60 ÷ 70 milioni di anni fa (fig. 14).
Mentre i processi evolutivi che si verificano all'interno delle popolazioni sono definiti microevolutivi, e consistono nel cambiamento delle frequenze alleliche all'interno delle popolazioni stesse, il termine macroevoluzione si riferisce alla formazione di nuove specie, famiglie, ordini, classi o tipi di fauna e di flora. L'evoluzione di queste forme di organizzazione più elevata non avviene rapidamente, ma attraverso piccoli passi successivi (tipogenesi additiva). È il caso, per esempio, dell'evoluzione degli equidi. l processi evolutivi (speciazione) possono essere accelerati quando gli organismi sono in grado di realizzare una nicchia ecologica molto ampia. Nel corso della filo genesi ciò si è verificato, per esempio, con la conquista della terra da parte dei pesci crossopterigi, con la conquista dell'aria da parte degli uccelli, e con l'apparizione delle prime piante terrestri nel Devoniano. La realizzazione di una nicchia ampia spesso comporta una rapida frammentazione in numerose specie (fenomeno noto come radiazione adattativa). Ne sono un esempio i fringuelli di Darwin delle isole Galápagos, i drepanidi delle Hawaii, o i marsupiali australiani.
Coevoluzione
Il termine coevoluzione indica un'evoluzione interdipendente di due partner o due sistemi di partner (piante, animali, funghi o batteri) che acquisiscono adattamenti specifici in conseguenza di mutue pressioni selettive. Come regola si distingue tra una coevoluzione accoppiata o reciproca (Janzen, 1980), in cui esiste una relazione tra due specie e l'una esercita una continua pressione selettiva sull'altra, e una coevoluzione diffusa o reticolata (Gilbert, 1975) alla quale partecipano molte specie, come nel caso delle piante con fiore, che sono visitate e impollinate da molte specie di insetti, e che quindi nel corso dell'evoluzione sviluppano, come adattamento verso i loro impollinatori, determinate caratteristiche comuni (forma, colore, profumo).
Il grado di dipendenza dei partner può variare significativamente. Nei casi estremi il successo riproduttivo di una specie dipende da un'altra specie, come per esempio l'impollinazione del fico da parte degli agaonidi (v. oltre).
Esistono tre diverse forme di interazioni coevolutive: la coevoluzione gene contro gene, la coevoluzione tra specie (coevoluzione specifica) e la coevoluzione tra gruppi di specie (coevoluzione diffusa).
Coevoluzione gene contro gene
Rientra in questa categoria il fenomeno per cui un determinato gene di un parassita che ne codifica la virulenza, possiede un suo antagonista nell'ospite che codifica la resistenza al parassita stesso. Questo tipo di interazioni si verifica spesso tra piante e funghi patogeni.
In una popolazione di Glycine canescens (famiglia fabacee) possono essere identificati Il diversi modelli fenotipici di resistenza, riconducibili ad almeno 12 fattori genetici di resistenza, che possono essere interpretati come risposta a 9 diversi genotipi del fungo della ruggine Phakopsora pachyrhizi (Burdon, 1987).
Coevoluzione tra specie (coevoluzione specifica)
Una coevoluzione stretta può essere particolarmente evidente in alcuni sistemi mutualistici o simbiotici. Saranno esposti ora alcuni esempi che si riferiscono ai sistemi costituiti dai fiori e dai loro impollinatori, e quelli costituiti dalle piante e dagli animali deputati alla loro dispersione.
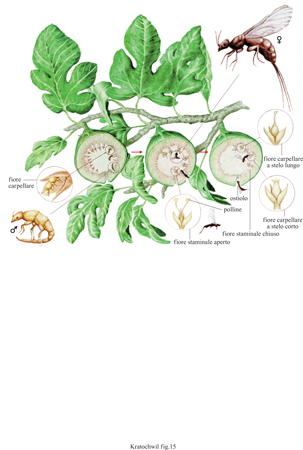
I sistemi fiore-impollinatore. - L'impollinazione del fico (genere Ficus, famiglia moracee) è molto complicata (Wiebes, 1979). Le infiorescenze a urna di un albero di fichi (siconi) nelle quali i fiori, estremamente ridotti, giacciono nella cavità centrale, attraggono migliaia di agaonidi (generi Ceratosoles, Blastophaga, Sycophaga, famiglia agaonidi), imenotteri della dimensione di appena pochi millimetri. Ciascuna specie di fico (ne esistono più di mille in tutto il mondo) è impollinata dalla 'sua' specie di insetti agaonidi. Le femmine degli agaonidi, che trasportano il polline in due borse toraciche, si insinuano attraverso la strettissima apertura rivestita di scaglie dell'infiorescenza (ostiolo), con gli stigmi rivestiti di polline. Quindi conficcano il loro ovopositore nello stelo di un fiore e vi depongono un uovo. In questo punto dello stelo si sviluppa una galla, come conseguenza della puntura e di alcune sostanze secrete dall'imenottero. La larva vive all'interno di questa galla nutrendosi dei suoi tessuti cellulari. Durante questo periodo il livello di diossido di carbonio nell'aria, all'interno del siconio, aumenta, e alla fine dello sviluppo larvale ammonta a circa il 10%. A queste concentrazioni sono attivi soltanto i maschi dell'agaonide, i quali fertilizzano le femmine all'interno delle galle, e quindi l'abbandonano perforando le pareti del siconio. Con il diminuire della concentrazione di anidride carbonica le femmine ritornano attive, lasciano le galle, riempiono le borse toraciche con il polline, che nel contempo è stato secreto dai carpelli dei fiori maschili, e abbandonano il siconio. Il processo quindi ricomincia da capo. Per evitare che le uova siano deposte in tutti i fiori esistono due tipi di fiori femminili: a stelo lungo e a stelo breve. Le uova sono deposte solo nei fiori con stelo breve, dato che l'ovopositore non è abbastanza lungo da penetrare nei fiori con stelo lungo (fig. 15).
Dispersione di piante da parte di animali (zoocoria). - Una stretta relazione lega la nocciolaia (Nucifraga caryocatactes caryocatactes) al pino cembro (Pinus cembra) (Mattes, 1978). La nocciolaia è diffusa negli ambienti montani europei, nei boschi ricchi di abeti, larici e pini cembri. Si nutre prevalentemente dei semi dei coni dei pini cembri, che riesce ad aprire con il suo becco a scalpello. In previsione dell'inverno la nocciolaia sotterra i semi del pino in vari nascondigli (2 ÷ 11 semi per ciascun nascondiglio, con una media di 3,5). Un esemplare può possedere più di 10.000 nascondigli, e quelli che vengono 'dimenticati' (circa il 20%) sono essenziali per la dispersione dei pini cembri. La nocciolaia ha una preferenza per la creazione di nascondigli lungo il margine del bosco, favorendo quindi la ricrescita preferenziale in queste aree. Relazioni simili a questa sono state osservate anche in Giappone tra la nocciolaia giapponese (Nucifraga caryocatactes japonicus) e il pino Pinus pumila.
Coevoluzione tra gruppi di specie (coevoluzione diffusa)
Esempi di coevoluzione tra gruppi di specie possono essere particolarmente frequenti in situazioni in cui nella struttura ecologica sono coinvolte associazioni tra le specie (coevoluzione associata). Un'associazione è definita come un gruppo di specie che utilizza la stessa classe di risorse ambientali in un modo simile, per esempio le specie che si nutrono del floema (afidi o pidocchi delle piante, sottofamiglia afidini), gli impollinatori e i predatori. In questo contesto è importante che molte specie diverse possano rappresentare vettori evolutivi.
Nel mutualismo ci sono numerosi esempi per questo tipo particolare di coevoluzione: interazioni tra le piante e i loro impollinatori, tra le piante e le specie animali che le disperdono, e tra le piante e le specie animali che le 'proteggono'.
Le piante e i loro impollinatori animali. - Molte piante hanno sviluppato strategie per sfruttare la mobilità degli animali al fine di conseguire l'impollinazione incrociata (mobilità in prestito). Grazie al fenomeno della zoofilia il polline - con opportune modalità - viene trasferito per assicurare lo scambio genico. Nelle regioni temperate il polline è trasportato dagli insetti (entomofilia), nelle regioni tropicali e subtropicali, questa funzione è assolta anche da uccelli (omitofilia), pipistrelli (chirotterofilia) e rettili (saurofilia), e in Australia anche dai mammiferi marsupiali della famiglia dei falangeridi (Faegri e van der Pijl, 1979). I visitatori dei fiori sono attratti dal nettare o dal polline, da oli, e in parte anche dai tessuti della pianta.
Lo spettro dei visitatori spazia dai generalisti (visitatori dei fiori euriantici) come il bombo (genere Bombus), agli specialisti (visitatori dei fiori stenoantici), che possono utilizzare solo determinate specie, generi o famiglie di piante. Esempi di quest'ultimo caso sono l'ape Colletes cunicularius, che visita esclusivamente i salici piangenti (genere Salix, famiglia salicacee), l'insetto andrenide Andrena florea, che si ritrova solo in due specie di brionia, Bryonia alba e B. dioica (famiglia cucurbitacee).

Tra le piante e i visitatori dei fiori si sono evoluti molti coadattamenti, con la naturale conseguenza che alcune sostanze attrattive primarie delle piante (pollini, nettare, oli) sono correlate con l'evoluzione delle strutture delle parti boccali degli animali adibite alla raccolta e al trasporto del polline (tab. I).
Così i granuli pollinici possiedono specifiche strutture superficiali che sono in relazione con gli apparati delle api deputati alla loro raccolta. L'ape Lasioglossum lineare (famiglia alictidi), uno dei principali impollinatori della pulsatilla (Pulsatilla vulgaris, famiglia ranuncolacee), possiede specifici apparati sulle zampe posteriori, costituiti da peli sottili. Questi peli coincidono in modo preciso con le suture dei granuli pollinici della pulsatilla (Kratochwil, 1988). L'intera morfologia del fiore delle piante zoo file è adattata alla morfologia dei rispettivi impollinatori, così come anche alle loro capacità sensitivo- fisiologiche (riconoscimento dei colori e degli odori) e alloro comportamento ('manipolazione' del fiore).
Alcune sindromi adattative si sono evo Iute tra diversi gruppi di animali (insetti, uccelli) e le piante che essi impollinano. La sindrome dell'ornitofilia (impollinazione a opera di uccelli) è comparabile con la chirotterofilia (impollinazione a opera di pipistrelli).
Le piante e gli animali deputati alla loro dispersione: l'esempio della mirmecocoria. - Viene definito mirmecocoria il fenomeno per cui le formiche disperdono i semi delle piante e possono in cambio nutrirsi degli annessi lipoidi da gialli a giallo-bianchi della diaspora (elaiosoma del seme). Piante mirmecocore sono, per esempio, l'aglio orsino (Allium ursinum, famiglia liliacee), le viole (genere Viola, famiglia violacee), la celidonia (Chelidonium majus) e la Corydalis cava (fam. papaveracee e fumaracee). Nel mondo esistono 70 famiglie di piante nelle quali è presente la mirmecocoria (Beattie, 1983). Sia nelle regioni boscose sia nelle aree aperte più aride la mirmecocoria può svolgere un ruolo importante; in particolare molte specie mirmecocore si possono trovare nel fynbos della Provincia del Capo in Sudafrica, una vegetazione a sclerofille simile alla macchia mediterranea.
Le piante e i loro protettori animali. - Le piante chiamate piante delle formiche (mirmecofite) forniscono un 'ambiente confortevole' alle formiche. Queste ultime in cambio proteggono la pianta dai fitofagi e contribuiscono alla loro nutrizione (mirmecotrofia).
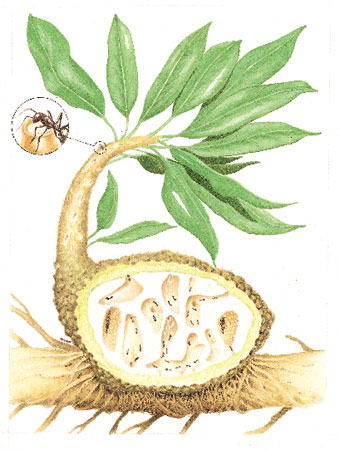
Di grande interesse sono alcune piante delle regioni tropicali e subtropicali, che forniscono uno 'spazio vitale' alle formiche (mirmecodomatia), e in cambio possono utilizzare come nutrienti (attraverso il trasporto di ioni) le sostanze di rifiuto della colonia di formiche, o anche venire da queste liberate dalle liane (fig. 21).
Coevoluzione tra piante e insetti fitofagi. - Animali erbivori filogeneticamente affini spesso si nutrono di piante filogeneticamente affini. Tra le farfalle, i bruchi delle cavolaie (famiglia pieridi) preferiscono le crucifere. Queste possiedono come prodotti secondari del loro metabolismo degli oli glicosidici attraverso i quali i pieridi riconoscono la pianta larvale. Queste sostanze hanno invece effetti tossici o repulsivi su mammiferi, funghi, batteri e altri insetti (Feeny, 1977).
I tessuti di molte specie di piante contengono sostanze secondarie (vegetoalcaloidi, furanocumarine e altre ancora) che le proteggono contro i loro potenziali consumatori (difese chimiche).
Fenomeni di coevoluzione si ritrovano anche nell'ambito delle relazioni preda-predatore, e tra ospiti e parassiti.
Biodiversità
Nella sua accezione originaria, diversità indica variazione, differenziazione, diversificazione, in contrasto con uniformità. La diversità può essere recepita come qualcosa di statico: l'eterogeneità denota le irregolarità, la varietà le differenze. La variabilità ha significato dinamico. Sistemi diversi possono essere semplici ma anche molto complicati; come regola, la complessità sta sicuramente a designare la diversità dei sistemi e può essere intesa nel senso di intricato, complicato. La complessità comprende la profondità strutturale dei sistemi, la diversità la loro ampiezza. Se riferita ai sistemi biologici, la diversità può essere anche vista come ricchezza.
Per biodiversità si intende la diversità biologica, ovvero la totalità della differenziazione, della variazione, della complessità, e della ricchezza della vita sulla Terra. La Convenzione sulla diversità biologica, stipulata a Rio de Janeiro nel 1992, ha stabilito che "la 'diversità biologica' significa diversità (in accordo con i significati di differenziazione, variazione, variabilità, complessità e ricchezza) negli organismi viventi, da qualsiasi fonte essa derivi, inclusi tra gli altri gli eco sistemi terrestri, marini e acquatici in genere, nonché i complessi ecologici di cui questi fanno parte". Ciò significa che include "la diversità all'interno di una specie, tra specie diverse e degli ecosistemi" (Bisby, 1995).
Ambiti di validità della biodiversità

La diversità è una qualità fondamentale che si manifesta nei diversi livelli di organizzazione della materia e dell'energia. È un elemento caratteristico di tutti i livelli gerarchici, biologici e non biologici (diversità gerarchiche). Esiste, perciò, una diversità a ogni singolo livello (fig. 22). l livelli della vita sono molto diversi, e qui generalmente distinguiamo tra diversità strutturali e diversità funzionali. l dati sulla diversità possono essere studiati a ciascun livello della struttura gerarchica utilizzando due diversi approcci (Solbrig, 1991): un approccio descrittivo, quale l'identificazione, la determinazione, la descrizione e la differenziazione degli elementi e dei loro componenti, un approccio funzionale, quale l'analisi causale delle combinazioni degli elementi e dei loro componenti, così come l'assorbimento, la trasformazione e l'elaborazione dell'energia e della materia.
Oggetto di questa analisi è il livello degli ecosistemi nel senso più ampio: le loro componenti biotiche (biocenosi) e i loro habitat (biotopi). Verrà qui trattato inoltre il livello dei complessi di eco sistemi (unità paesaggistiche). Questi complessi sono formati da numerosi eco sistemi, la cui relazione segue regole definite. Fin dal Neolitico, e in continuo aumento negli ultimi 150 anni, l'uomo ha considerevolmente influenzato gli eco sistemi e i complessi di eco sistemi in molte parti del mondo. Uno studio sulla biodiversità deve quindi includere i sistemi uomo - ambiente. Cause antropogeniche possono essere alla base di aumenti o diminuzioni della biodiversità.
Forme di biodiversità
Le diverse forme di biodiversità possono essere classificate utilizzando quattro differenti tipologie che verranno descritte brevemente di seguito.
Diversità degli elementi (modello degli elementi). - È possibile discutere questa forma di biodiversità sotto vari aspetti:
l) diversità tassonomica e sintassonomica, di specie e di cenosi. I livelli della diversità di varie specie e la diversità delle cenosi possono essere distinti in diverse unità spaziali: diversità α, diversità β, diversità γ, e diversità δ (Goetze e Schwabe, in stampa). La diversità α, che viene definita anche come 'diversità all'interno di una comunità', si riferisce alla diversità di specie in una determinata area si può descrivere utilizzando indici numerici o diversi metodi di calcolo matematico. I gradienti ambientali che caratterizzano la transizione tra diversi biotopi possono essere analizzati attraverso la diversità β, definita anche 'gradiente di diversità tra comunità'; questo procedimento è particolarmente adatto per regioni che presentano gradienti ecologici (ecoclini), per esempio zone di contatto tra foresta e terreni aperti e complesse zonazioni costiere, ma meno utile nel caso di aree con discontinuità molto pronunciate. La diversità γ, definita anche 'diversità dei complessi di comunità', caratterizza la diversità dei paesaggi: la porzione di un paesaggio (fisiotopo) è composta di diverse comunità, la cui totalità forma un complesso vegetazionale; all'interno di un fisiotopo si possono ritrovare determinate combinazioni di fattori uniformi (substrato geologico, condizioni del suolo, bilancio di nutrienti, bilancio idrico). Le unità rilevanti per lo studio della diversità γ sono rappresentate dagli ecosistemi e dai complessi di ecosistemi. La diversità δ caratterizza le variazioni nel numero di tipi di vegetazione che si ritrovano lungo un gradiente ecologico, in modo analogo alla diversità β, nella quale vengono prese in esame le variazioni nel numero di specie lungo un gradiente ecologico.
2) Diversità delle forme di vita. Il concetto di forme di vita comprende l'intero complesso di qualità specie-specifiche di un organismo (caratteristiche morfologiche, fisiologiche ed etologiche), che si sono evo Iute come adattamento alle particolari condizioni offerte da un determinato ambiente. Queste forme di vita possono essere caratterizzate tipologicamente: un tipo di forma di vita caratterizza un gruppo di specie. Queste specie possono appartenere a diverse categorie sistematiche (generi, famiglie, ordini), ma nel corso dell'evoluzione hanno acquisito, in risposta alle condizioni specifiche di un habitat, modi di vita e caratteri morfologici, fisiologici e etologici analoghi; di conseguenza possiedono la stessa forma di vita. Per gli animali, i tipi di forme di vita possono essere classificati in accordo con i loro regimi alimentari (fitofagi, zoofagi, parassiti, filtratori, consumatori di substrato, erbivori, consumatori di linfa, succhiatori, raccoglitori, predatori, trappolatori), in base alle modalità di locomozione (scavatori, striscianti, arrampicatori, saltatori, volatori, corridori), o tenendo conto dei luoghi di residenza (edafici, atmobiotici, erbicoli, fillobici, lignicoli, epizoici, endozoici). Per quanto riguarda le piante, le diverse forme di vita si possono distinguere sulla base delle modalità di sopravvivenza durante le stagioni sfavorevoli (seguendo la classificazione di Raunkiaer), o degli adattamenti all'equilibrio idrico (xerofile, mesofile, igrofile, idrofile), o anche delle esigenze luminose, dei fattori del suolo (edafici) e della dieta.
3) Diversità delle strutture spaziali. Un ambiente può essere diviso in tre diverse strutture spaziali: stratotopo, coriotopo e merotopo. Questa differenziazione è essenziale per il rilevamento e l'analisi delle sinusie all'interno di una biocenosi.
4) Diversità trofica. Prevede la classificazione degli organismi nei livelli di produttori, di consumatori e di decompositori, e nelle ulteriori suddivisioni di questi livelli.
5) Diversità fenologica. Indica la classificazione delle strutture temporali, quali le variazioni diurne e stagionali, i fenomeni periodici annuali.
6) Diversità genetica e specifica delle popolazioni. Prevede la caratterizzazione della variabilità genetica e dello spettro dei genotipi, dei fenomeni di omozigosi, eterozigosi e deriva genetica, dei tassi di mutazione delle singole popolazioni, e di altri parametri simili (v. anche il saggio di A. Piazza, nel l volume dell'opera).
7) Diversità biochimica. Indica la defrnizione dei costituenti delle piante (per esempio, gli alcaloidi), che sono in una certa misura importanti come 'difese biochimiche' contro i fitofagi o che costituiscono essenze attrattive per gli animali che visitano i fiori (Feeny, 1977).
Diversità delle interazioni. - Le specie creano fra di loro polisistemi formando le cosiddette connessioni biocenotiche. Queste interazioni tra gli organismi inducono l'apparizione di caratteri che possono contribuire alla stabilizzazione del sistema (la quasi stabilità nella composizione della specie). Queste interazioni possono essere suddivise in interazioni probiotiche (mutualismo, simbiosi, commensalismo) e antibiotiche (predazione, parassitismo, ecc.).
Meccanismi che generano la diversità (modello causale). - I processi fondamentali che generano la biodiversità possono essere distinti in processi che producono effetti su un scala temporale evolutiva (separazione, speciazione, radiazione) e processi che producono effetti su una scala temporale ecologica.
l) Effetti su scala evolutiva. Nei periodi di tempo che caratterizzano l'evoluzione, la biodiversità è raggiunta attraverso la speciazione (allopatrica o simpatrica). In questo caso assumono una grande importanza la separazione di popolazioni originariamente collegate, la loro conseguente differenziazione, lo sviluppo di meccanismi di isolamento, e la formazione di nicchie ecologiche differenziate. Un esiguo numero di estinzioni rappresenta un fattore decisivo per raggiungere alti tassi di differenziazione. Una diversità di specie particolarmente elevata è conseguente alle radiazioni; ne sono un esempio i fringuelli di Darwin (geospizini) nelle isole Galapagos (Lack, 1947), o i drepanidi e i moscerini della frutta delle Hawaii (Mayr, 1943).
2) Effetti nei tempi ecologici. Nella scala temporale ecologica, una biocenosi ricca di specie può svilupparsi solo quando si ha un'immigrazione di comunità, e queste sono di nuova formazione. In questo contesto il numero di nicchie ecologiche da realizzare gioca un ruolo decisivo. La nicchia ha una dimensione autofitica/autozoica e una dimensione ambientale: la dimensione autofitica/autozoica include tutti i caratteri morfologici e fisiologici (per gli animali, anche i caratteri etologici) di una specie che sono stati acquisiti nel corso della filogenesi, mentre la dimensione ambientale rappresenta la somma di tutti i fattori ecologici effettivi. La nicchia ecologica di una specie si realizza nella sovrapposizione delle due dimensioni (Schmitt, 1987). L'ampiezza di una nicchia dipende dal grado di specializzazione delle nicchie ecologiche che la realizzano. Sovrapposizioni di nicchia sono tollerate soltanto da specie caratterizzate da una maggiore ampiezza di nicchia.
Processi di funzionamento (modello funziona/e). - Una questione al centro di notevoli controversie riguarda la valutazione del contributo della biodiversità al funzionamento delle biocenosi. Non vi è dubbio che molte specie sono costantemente collegate da determinate interazioni, e che queste relazioni possono essere obbligate. Una struttura di interazioni di questa natura assume caratteristiche di sistema solo quando può essere distinta da altri sistemi, e quando può essere determinato un flusso indipendente di materia. La differenziazione di biocenosi ed eco sistemi ha un carattere meramente ipotetico; di conseguenza, a questioni quali la ridondanza che può tollerare una biocenosi o un eco sistema senza minare il mantenimento del bilancio funzionale, o l'esistenza di limiti superiori e inferiori alla biodiversità, si può rispondere solo per via teorica.
Più un sistema è diverso, più deve essere diversa la struttura funzionale che lo stabilizza. La diversità degli elementi e la diversità delle interazioni contribuiscono in maniera fondamentale alla sua stabilizzazione. La materia (i nutrienti) e il flusso di energia sono necessari per mantenere il sistema e raggiungere la quasi-stabilità. l processi di stabilizzazione includono l'assorbimento, la trasformazione e il trasferimento di materia e nutrienti.
Diversità intrabiocenotica
Una biocenosi è composta dalla comunità di piante (fitoceno si) che colonizza un fitotopo, e dalla comunità di animali (zoocenosi) che vive in uno zootopo. Dato che le piante superiori sono fisiognomicamente dominanti, le comunità vegetali possono essere più facilmente analizzate e caratterizzate rispetto alle zoocenosi.
Esistono diversi approcci pragmatici allo studio delle biocenosi e della loro diversità: l'investigazione dei gruppi tassonomici (zootassocenosi), per la classificazione della biodiversità; l'investigazione dei gruppi funzionali o delle associazioni (sottosistemi, unità più piccole, gruppi funzionali di specie coesistenti che utilizzano le stesse risorse in modo simile), per la biodiversità funzionale, l'analisi di determinate relazioni (per esempio, i complessi piante-insetti, le catene e le reti alimentari), per la biodiversità delle interazioni; l'esame dei microhabitat (sinusia): per la classificazione delle biodiversità nei microhabitat.
Oltre il 90% delle specie animali che vivono sulla Terra è legato ad ambienti caratterizzati da una propria vegetazione. Il primo passo nello studio di una comunità animale può essere costituito dalla caratterizzazione dell'ambiente, dato che le comunità di piante o i complessi vegetazionali costituiscono unità caratterizzabili da un punto di vista ecologico, strutturale, dinamico, corologico e sinergetico. Questa caratterizzazione di un ambiente attraverso la sua comunità di piante e i complessi di comunità di piante rappresenta il punto di partenza per la catalogazione e l'analisi della diversità biocenotica.

La seconda fase è costituita dalla classificazione dei microhabitat, che dovrebbe basarsi su tre diversi tipi di struttura spaziale: stratotopo, coriotopo e merotopo. l diversi strati, per esempio di una foresta, vengono definiti stratotopi; in questo caso possono essere distinti lo strato degli alberi, dei tronchi, dell'erba, ciascuno colonizzato dalla sua stratocenosi. l coriotopi invece rappresentano strutture verticali indipendenti dall'intera unità spaziale o dalle parti dello stratotopo, le cosiddette coriocenosi, quali per esempio la comunità di insetti di un albero o di un cespuglio. Infine, in un ambiente strutturalmente ricco, si possono identificare i merotopi, ovvero elementi strutturali che si trovano all'interno di uno stratotopo o di un coriotopo, come gli organismi che vivono sulle foglie, sulla corteccia o i visitatori di fiori (fig. 23).
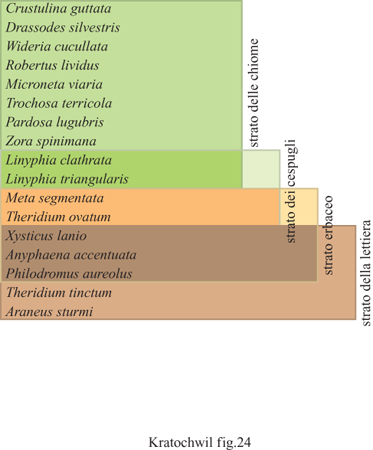
Stratocenosi. - Le analisi della biodiversità tassonomica dimostrano che ciascuno strato possiede il suo inventario di specie animali, come per esempio lo stratocenosi di ragni nei boschi di querce-betulle dell'Europa centrale. Il confronto tra gli strati in diverse comunità vegetali, per esempio tra gli strati delle foglie e del suolo di un prato a erba melica - foresta di faggio (Melico-Fagetum) e di un querceto-carpineto (Querco-Carpinetum) mostra nette differenze nella composizione di specie di lombrichi nelle stratocenosi e, in particolare, nello strato della lettiera (fig. 24) (Rabeler, 1957).
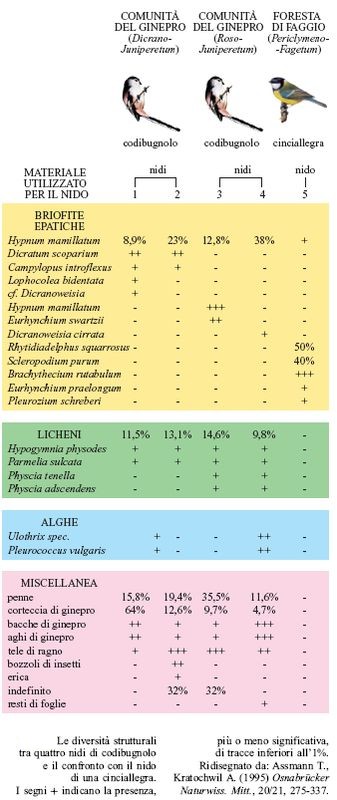
Coriocenosi. - Altri elementi strutturali includono elementi speciali, chiaramente differenziabili, i cosiddetti coriotopio La diversità di un coriotopo verrà illustrata utilizzando come esempio un nido di uccello (Assmann e Kratochwil, 1995). Le diverse specie di uccelli necessitano di speciali requisiti per costruire il loro nido: nella Germania settentrionale il codibugnolo (Aegithalos caudatus) costruisce caratteristici nidi in alto sui ginepri (Juniperus communis). L'analisi del materiale utilizzato per il nido ha rivelato la presenza di materiali specifici, costituiti da determinate specie di muschi, di licheni, di alghe. La composizione dipende dalla comunità di piante nella quale viene costruito il nido. È questo un esempio ordinario di diversità strutturale tipica di un determinato ambiente. Il nido della cinciallegra (Parus major) è costruito in un altro modo, e inoltre questa specie si ritrova per lo più in ambienti abbastanza diversi (fig. 25). La diversità di specie comporta perciò diversità nelle piccole strutture da esse create.
Merocenosi. - l merotopi sono parte degli stratotopi e dei coriotopi. Strato-, corio- e merotopi combinano in un certo grado la diversità strutturale e funzionale. Verranno qui analizzate in particolare le nicchie ecologiche, i livelli di interazione e le strutture di relazioni tra gli organismi.
La comunità dei visitatori dei fiori corrisponde a una merocenosi, in cui i fiori rappresentano i merotopi. lnnanzitutto troviamo una 'bio diversità sistematica' di gruppi animali molto diversi: imenotteri aculeati, lepidotteri, coleotteri, ecc. All'interno di questo sistema formato dal fiore e dal suo visitatore esiste una 'diversità funzionale' introdotta dal visitatore: le relazioni alimentari (polline, nettare, oli), o relazioni legate ad altre risorse, come l'uso dei fiori come luoghi di riscaldamento (dovuto alla loro forma parabolica che simula uno specchio), come luoghi di incontro, come risorse di cibo per predatori e parassiti, come rifugi notturni (per esempio, per le api) o come fornitori di materiali per il nido. l fiori possono anche fornire essenze odorose utilizzate per marcare i percorsi di sciamatura, come accade nelle api raccoglitrici sciamanti neotropiche (sottofamiglia euglossine). Solo per le piante che producono oli, circa 1400 specie appartenenti a lO famiglie note in tutto il mondo, esistono 300 specie di api specializzate nello sfruttarle (Vogel, 1988).
Nelle piante il livello di simbiosi varia dal parassitismo al mutualismo, che può essere ritrovato nella sua forma più distintiva negli esemplari del genere Ophrys: i fiori imitano la femmina dell'ape e si insinuano attraverso stimoli ottici, odorosi e tattili nel comportamento istintivo dell'ape maschio per assicurarsi il trasferimento del polline. La diversità di specie e la diversità funzionale sono sempre correlate con la diversità strutturale. Si è già discusso l'esempio della correlazione tra le strutture deputate alla raccolta del polline di un'ape e la forma di certi granuli pollinici.
La diversità strutturale di una merocenosi di un sistema formato da un fiore e dal suo visitatore è immensa, e include: diversità visiva (i colori dei fiori nello spettro del visibile e dell'ultravioletto); olfattiva (la moltitudine dei profumi dei fiori); etologica (la varietà dei comportamenti dei visitatori dei fiori); fenologica (la variazione diurna e stagionale della presenza dei fiori e dei loro impollinatori).
Ciascuna comunità di piante possiede la sua comunità di animali a diversi livelli. Spesso gli animali hanno bisogno di interi complessi di vegetazione. A livello di un ecosistema, i livelli di diversità strutturale e funzionale dei diversi gruppi di organismi sono correlati con il loro specifico ambiente abiotico. Le biocenosi o i complessi di biocenosi sono caratterizzati da determinate specie caratteristiche e ciascuna biocenosi possiede il suo ambito di tipi e modelli di diversità. Maggiore è la diversità di specie, più vari sono gli altri tipi di diversità: genetica, spazio-strutturale, fisiognomica, biochimica, fenologica, ecc.
Diversità interbiocenotica
Di regola i paesaggi non sono composti da singole bioceno si, ma piuttosto da complessi di biocenosi e da mosaici di diversi ecosistemi. Lo sviluppo, per esempio, di singole unità di vegetazione in associazioni, non è arbitrario, ma segue determinate regole. È interessante il fatto che le regolarità a livello dei complessi specie/biocenosi (legge di Thienemann) e dei complessi biocenosi/biocenosi seguono le stesse leggi naturali.
Nel primo caso: "Più sono variabili le condizioni di vita in un determinato luogo, maggiore è il numero di specie della corrispondente comunità" e "più le condizioni di vita di un biotopo deviano dalle condizioni ottimali per la maggior parte degli organismi che ci vivono, più la biocenosi diverrà povera di specie e caratteristica e maggiore sarà il numero di individui di una specie in esse presenti".
Nel secondo caso: "Più sono variabili le condizioni ambientali di un complesso di habitat, più elevato è il numero delle sue biocenosi" e "più le condizioni ambientali di un complesso di habitat deviano dalle condizioni ottimali per la maggior parte delle biocenosi, più povero diverrà in bioceno si, le quali saranno più caratteristiche e più grandi" (Schwabe e Kratochwil, 1994).
Diversa importanza di una specie in sistemi diversi
È un dato ormai acquisito che spesso l'importanza funzionale di una specie non sia conosciuta e che sovente non esistano indicazioni su come le specie reagiscono a fattori di stress in determinate biocenosi. Ci sono specie di importanza centrale in un eco sistema, le cosiddette specie portanti, senza le quali l'intero sistema crollerebbe. Queste specie chiave sono, per esempio, nel mondo animale, il castoro (Castor fiber e Castor canadensis canadensis) che modella l'intero paesaggio fluviale nelle regioni boreali. La funzione di molte specie all'interno di un eco sistema non è al momento scientificamente documentata. P.R. Ehrlich e A.H. Ehrlich (1981) hanno formulato un'ipotesi secondo la quale ciascuna specie svolge la stessa funzione del rivetto di un aeroplano. La sua importanza non può essere predetta per qualsiasi situazione. Ci sono molti sottosistemi ridondanti e nessuno sa - utilizzando ancora la metafora dell'aeroplano - se la perdita di un rivetto possa essere compensata o se l'assenza di uno solo di essi possa avere gravI conseguenze.
Aspetti applicativi della biodiversità
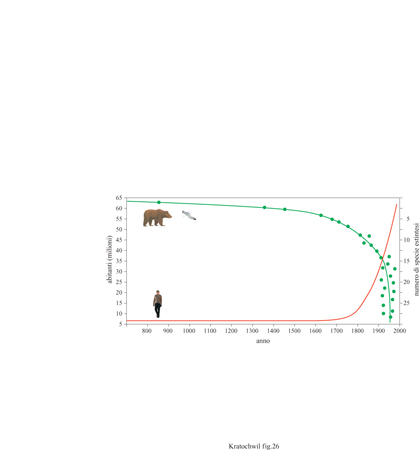
Fino a oggi sono state descritte circa un milione e mezzo di specie e di animali viventi sulla Terra, ma il loro numero attuale effettivo potrebbe variare tra i 5 e i 30 milioni. Per l'ultimo quarto del 20° secolo gli scienziati hanno predetto l'estinzione di circa un milione di specie. Dal 1600 al 1900 l'uomo ha provocato l'estinzione di una specie ogni quattro anni; dopo il 1900, scompare una specie ogni anno; attualmente si va estinguendo più di una specie al giorno: si suppone che scompaia una specie ogni ora. Entro la fine del 20° secolo il 20 ÷ 50% di tutte le specie viventi sarà scomparso (fig. 26). In condizioni naturali, il tasso netto di crescita del numero di specie è pari a 0,37% ogni milione di anni: un valore estremamente basso. Il tasso naturale di estinzione è invece, a causa dell'uomo, aumentato di 10.000 volte; il declino delle specie attuali è 100 volte maggiore rispetto alla perdita di specie negli ultimi 65 milioni di anni. Il tasso di perdita di diversità genetica a livello di popolazione accresce di molto questo valore.
Le zone a biodiversità particolarmente elevata sono concentrate ai tropici, e principalmente nella aree montane tropicali. In pochi ettari di foresta del sudest asiatico o della regione amazzonica si trovano più specie di alberi che nell'intera Europa. Nella foresta sempreverde del Venezuela ci sono almeno 90 specie di alberi per ettaro.
In particolari regioni la perdita di biodiversità è estremamente significativa: a livello mondiale numerosi eco sistemi sono particolarmente a rischio di estinzione, e tra questi le foreste pluviali tropicali, alcuni eco sistemi marini, isole marine, alte fasce montane, ambienti artici e subartici, savane, steppe, semideserti, grandi sistemi fluviali, foreste a mangrovie, molti laghi, ma anche i paesaggi delle campagne in cui viviamo.
Una perdita di biodiversità non può essere tollerata, per ragioni ecologiche, etiche, religiose, estetiche e culturali, soprattutto perché la distruzione della bio diversità è un fenomeno irreversibile.
Mantenere la biodiversità, stabilire i principi teorici e tradurli in misure pratiche rappresentano alcuni dei principali compiti che l'umanità avrà di fronte nei prossimi anni. La preservazione della biodiversità è strettamente legata alla sopravvivenza dell'uomo sulla Terra, e questo principio deve quindi essere incorporato nel concetto stesso di sviluppo sostenibile.
Bibliografia citata
ASSMANN, T., KRATOCHWIL, A. (1995) Biozönotische Untersuchungen in Hudelandschaften Nordwestdeutschlands Grundlagen und erste Ergebnisse. Osnabriicker Naturwiss. Mitt., 20/21: 275-337.
BEATTlE, A. (1983) Distribution of ant dispersal plants. Sonderbande des Naturwissenschafilichen Vereins in H., 7, 249-270.
BISBY, F.A. (1995) Characterization of biodiversity. In Global biodiversity assessment, a c. di Heywood V.H., Watson R.T., Cambridge, Cambridge University Press, pp. 21-57.
BURDON, J.J. (1987) Phenotypic and genetic pattem of resistance to the pathogen Phakopsora pachyrhizi in populations of Glycine canescens. Oecologia, 73, 257-267.
DARWIN, CH. (1859) On the origin of species. Londra, Murray.
EHRLlCH, P.R., EHRLlCH, A.H. (1981) Extinction. The causes and consequences of the disappearance of species. New York, Random House.
FAEGRI, K., VAN DER PUL, L. (1979) The principles olpollination ecology, 3' ed., Oxford, Pergamon Press.
FEENY, P. (1977) Defense ecology of the Cruciferae. Ann. Missouri Bot. Gard., 64, 221-234.
GILBERT, L.E. (1975) Ecological consequences of a coevolved mutualism between butterflies and plants. In Coevolution of animals and plants, a c. di Gilbert L.E., Raven P.H., Austin, University of Texas Press, pp. 210-240.
GOETZE, D., SCHWABE, A. (in stampa) Linking ecosystem mosaics and biodiversity concepts by means of sigmasociology. ColI. Phytosoc. 27. Berlino.
HUTCHINSON, G.E. (1965) The niche, an abstractly inhabitat hypervolume. In the ecological theatre and the evolutionary play. New Haven.
HUXLEY, J.S. (1974) Evolution. The modern synthesis, 3a ed., Londra, G. Allen & Unwin Ltd.
JANZEN,D.H. (1980) When is it coevolution? Evolution, 34, 611-612.
KETTLEWELL, H.B.D. (1972) The evolution of melanism. Oxford, Clarendon Press.
KRATOCHWIL, A. (1988) Zur Bestäubungsstrategie von Pulsatilla vulgaris Mill. Flora, 181, 261-325.
LAcK, D.L. (1947) Darwin'sfinches. Cambridge, Cambridge University Press.
MATTES, H. (1978) Der Tannenhäher im Engadin. Münster Geogr. Arbeiten, 2, 1-87.
MAYR, E. (1943) The zoogeographic position of the Hawaiian Island. Condor, 45, 45-48.
OSCHE, G. (1966) Grundzuge einer allgemeinen Phylogenetik. In Handbuch der Biologie, voI. 3, a c. di von Bertalanffy L., Costanza, Akademishe Verlagsgesellschaft Athenaion pp. 817 -906.
RABELER, W. (1957) Die Tiergesellschaft eines Eichen-Birkenwaldes im nordwestdeutschen Altmoränengebiet. Mitt. flor. soz. Arbeitsgem., 6/7, 297-319.
SCHMITT, M. (1987) Ecological ruche sensu Glinther and ecological licenses sensu Osche - two valuable but poorly appreciated explanatory concepts. Zoo!. Beitr., 31, 49-60. SCHWABE, A., KRATOCHWIL, A (1994) Gelten die biozönologischen Grundprinzipien auch für die landschaftsökologische Dimension? Einige Uberlegungen mit Beispielen aus den Inneralpen. Phytocoenologia, 24, 1-22.
SOLBRIG, O.T. (1991) Biodiversity. Scientific issues and collaborative research proposals. Parigi, UNESCO.
VOGEL, S. (1988) Die Olblumensymbiosen - Parallelismus und andere Aspekte ihrer Entwicklung in Raum und Zeit. Z. zool. Syst. Evolut.-Forsch., 26, 341-362.
WIEBES, J.T. (1979) Coevolution of figs and their insect pollinators. Ann. Rev. Ecol. Syst., 10, 1-12.
Bibliografia generale
FUTUYMA, D.J. Evolutionary biology. Sunderland, Sinauer Associates, 1986.
GILBERT, L.E., RAVEN, P.H. Coevolution of animals and plants. Austin, University of Texas Press, 1980.
HEYWOOD, V.H., WATSON, R.T., a c. di, Global biodiversity assesment. Cambridge, Cambridge University Press, 1995.
MAYR, E. Populations, spedes and evolution. Cambridge, Harvard University Press, 1970.
SCHULZE, E.-D., MOONEY, H.A., a c. di, Biodiversity and ecosystem function. Berlino, Springer, 1993.
WILSON, R.H. The diversity of life. Cambridge, Harvard University Press, 1972.