
Coevoluzione uomo-vegetazione nell'ecosistema mediterraneo
Coevoluzione uomo-vegetazione nell'ecosistema mediterraneo
La prima domesticazione di piante commestibili a scopo agricolo è avvenuta intorno al 9000 a. C. nel bacino del Mediterraneo orientale. Qui, in condizioni naturali, era presente una vegetazione di foresta sempreverde relativamente povera di specie, mentre la flora erbacea era limitata ad ambienti estremi in habitat marginali (rupi, alte montagne, ambienti costieri). L'uomo ha interferito su questa situazione, soprattutto attraverso gli incendi, già in epoca molto remota. Dove la foresta veniva incendiata si formavano ampie aree libere, che rappresentavano un'occasione di espansione per le specie degli ambienti estremi. L'intensa speciazione così originatasi ha dato origine a una flora interamente nuova, propria degli ambienti disturbati dall'azione dell'uomo. In questa flora si trovavano numerose specie commestibili, tra le quali i precursori di molte piante coltivate: frumento, orzo, segale, avena, veccia, lenticchia. Tra l'azione di disturbo dovuta alla popolazione umana e l'espansione della vegetazione ricca di piante alimentari si è stabilito un sinergismo. In questo modo si è avviato un processo di coevoluzione pianta-uomo essenziale per lo sviluppo della cultura mediterranea.
Domesticazione delle piante agrarie
Il processo di domesticazione delle piante agrarie ha avuto inizio in molte zone della Terra in maniera del tutto indipendente: nel Mediterraneo, in India, in Iran, in Cina, nell'Africa subsahariana, nella Mesoamerica, nelle Ande. Le tracce più antiche dell'agricoltura sono state trovate nel Mediterraneo orientale, nel sito di Gerico risalente al 9000 a. C. e in altri siti di poco più recenti localizzati nella mezzaluna fertile (area compresa tra Siria, Iraq e Giordania) e in Egitto.
La domesticazione delle piante agrarie nel Mediterraneo orientale (Renfrew, 1973) ha avuto indubbiamente un'importanza primaria, non solo perché è stata la prima, ma anche perché da quest'area sono derivate alcune tra le piante che tuttora hanno la massima diffusione su scala mondiale e che forniscono la maggior parte degli alimenti in tutto il mondo: il frumento, l'orzo, la segale, l'avena (Diamond, 1997). Qui è avvenuta anche, quasi contemporaneamente, la domesticazione di alcuni tra i più importanti animali d'allevamento come la vacca, la pecora, la capra, il cane e il suino (questi ultimi addomesticati anche in altre aree). Per quanto tutto ciò sia indubbiamente legato alle particolari condizioni climatiche della zona, è noto che climi di tipo mediterraneo caratterizzano anche altre quattro parti del mondo dove tuttavia non sono avvenute importanti vicende di domesticazione: Califomia, Cile centrale, Sud-Africa e Australia Occidentale (Di Castri e Mooney, 1977). È dunque interessante chiedersi perché la domesticazione si sia sviluppata nell'ambiente mediterraneo prima che, per esempio, ai tropici, e perché tra le cinque aree possibili, proprio nel Mediterraneo e non nelle altre dove vi era un clima quasi identico. La risposta va cercata nella storia evolutiva della flora mediterranea, profondamente differente da quella delle flore degli altri ecosistemi di tipo mediterraneo, e nella storia della presenza umana nelle varie aree. La flora mediterranea costituisce dunque un interessante esempio di come la bio diversità di un gruppo di piante possa condizionare lo sviluppo socio-culturale e contemporaneamente venime condizionata.
Un ambiente in rapido cambiamento e la biodiversità
Il bacino mediterraneo possiede una flora costituita da circa 20.000 specie (Greuter et al., 1984-89), e pertanto è uno dei territori del globo ad alta densità floristica (Malyshev, 1975). Sull'evoluzione di questa flora sappiamo ancora molto poco, soprattutto perché l'approccio scientifico largamente prevalente (non solo nella biologia evoluzionistica) è quello riduzionistico, che porta a considerare l'evoluzione come un fenomeno che riguarda solo il vivente, trascurando le retroazioni che si stabiliscono a livello di popolazione, di specie, di comunità e di eco sistema. Per approfondire il problema della domesticazione delle piante agrarie bisogna anzitutto esaminare le caratteristiche climatiche dell'ambiente mediterraneo e l'evoluzione della flora in quest' area. È possibile anche indicare alcuni eventi che hanno avuto conseguenze di carattere generale (Pignatti, 1978; 1979; 1995).
Bioclima mediterraneo
Si tratta di un clima temperato-caldo, talora quasi subtropicale, con estati calde e aride e inverni relativamente miti (Di Castri e Mooney, 1977; Pignatti, 1986). La temperatura media annua è compresa tra 14°C e l8÷19°C, le precipitazioni tra 600 e 1200 mm annui, che possono scendere a 350÷400 mm nelle aree subdesertiche. Le gelate e le precipitazioni nevose sono rare e in genere si esauriscono in pochi giorni.
Per le condizioni fisiologiche della pianta questo clima determina un periodo di stress durante i mesi estivi, quando le alte temperature provocano una elevata traspirazione e il suolo si secca, almeno in superficie. A queste condizioni negative per la vita vegetale, si aggiunge anche una decisa riduzione delle piogge, talora vera e propria siccità, che può durare da due a quattro mesi. Tali condizioni, in genere, costringono i vegetali a sospendere completamente le proprie attività vitali e, quindi, l'estate rappresenta un periodo di pausa vegetativa quasi completa. Le specie legnose della flora mediterranea sono per lo più sempreverdi, cioè possono mantenere le foglie d'inverno, non essendoci le gelate prolungate che potrebbero danneggiarle; esse durante l'estate mantengono le foglie, ma la fotosintesi in generale scende a valori molto bassi oppure cessa del tutto. Le specie erbacee invece, durante il periodo estivo di solito perdono completamente le foglie, mantenendo soltanto organi sotterranei come bulbi o rizomi, oppure passano il periodo arido sotto forma di seme. Il clima ad aridità estiva è proprio sia delle aree che circondano il Mar Mediterraneo, sia, con differenze di scarso significato, degli altri eco sistemi di tipo mediterraneo.
Derivazione da due ceppi floristici
Nella flora vascolare dell'area mediterranea si possono distinguere due componenti: i gruppi ad affinità prevalentemente occidentale (Pignatti e Pignatti, 1989), che comprendono soprattutto specie della macchia acido fila e dei terreni poveri, e i gruppi ad affinità prevalentemente orientale, che includono gran parte delle specie arboree caducifoglie e la vegetazione degli ambienti salati e costieri. A queste due componenti, in generale prevalenti, si aggiungono altre correnti floristiche, in particolare i gruppi con affinità paleotropicale, atlantico-temperata e nordica, questi ultimi probabilmente come conseguenza delle glaciazioni.
Ambiente desertico durante il Messiniano
Circa 7 milioni di anni or sono si verificò un generale sollevamento della soglia di Gibilterra e il collegamento tra il Mar Mediterraneo e l'Oceano Atlantico venne interrotto. Il Mar Mediterraneo divenne un bacino chiuso e la forte evaporazione determinò la formazione di deserti salati. In questo periodo va probabilmente collocata l'espansione di gruppi di alofite come Limonium, Salicornia, Arthrocnemum, Salsola, Artemisia gr. maritima, in gran parte con affinità tetisiana. L'emersione dei fondi marini consentì anche l'espansione di gruppi di specie montane, come gli Astragalus spinosi, che dal loro centro caucasico-iraniano si diffusero sulle montagne mediterranee fino alla Sierra Nevada. Il Messiniano ebbe una durata ridotta, inferiore a l milione di anni, quindi il collegamento con l'Atlantico fu ripristinato e il bacino nuovamente riempito. Questa fase fu importante per la comparsa di specie di ambienti marginali, con condizioni di vita estreme, come gli ambienti di vetta, le creste, le rupi e gli stillicidi. Queste specie sono una componente importante della biodiversità della flora mediterranea. Si tratta di piante di piccole dimensioni, defmibili come opportuniste in quanto hanno maggiore capacità di adattamento in condizioni particolarmente severe. Esse si diffondono in ambienti diversificati (specie euriece), e molte di esse avranno un'ulteriore evoluzione. Alla fine del Pliocene il clima divenne di tipo subtropicale con piogge abbastanza abbondanti, dall'autunno alla primavera, e aridità estiva: si stabilì così quella che sarebbe diventata la caratteristica climatica più importante della regione mediterranea. In seguito si verificarono le glaciazioni, che nel bacino mediterraneo determinarono un abbassamento dei limiti forestali, e alcune fasi aride che interessarono le aree costiere.
Variazioni del livello marino
Nelle ultime fasi del Pliocene e durante il Pleistocene si verificarono ripetuti cambiamenti del livello marino, che modificarono profondamente l'andamento delle coste nella penisola e nelle isole. Questo ha portato a una sommersione della superficie terrestre fino al livello di 270÷280 m s.m. durante il Selinuntiano e, al contrario, a un abbassamento del livello marino fino a quasi 100 m sotto quello attuale durante le glaciazioni. Le sommersioni hanno avuto conseguenze per la flora soprattutto per la distribuzione delle specie endemiche o comunque specializzate nell'ambiente costiero e insulare, che vennero eliminate dalla flora delle isole meno elevate. Le isole che nel Selinuntiano vennero completamente sommerse (Vulcano, Stromboli, Lampedusa, Zannone, Linosa) presentano un numero di specie significativamente meno elevato, in media 260, rispetto alle altre (Pantelleria, Favignana, Ponza, Marettimo, Levanzo) caratterizzate in media da 460 specie (Pignatti, 1995). Nella distribuzione del genere Limonium, tipico di ambienti costieri, abbiamo la situazione seguente: nelle isole del primo gruppo si trova soltanto l specie endemica, mentre in quelle del secondo gruppo si trovano 7 specie endemiche. Anche in questo caso si tratta di un gruppo a elevata biodiversità, la cui evoluzione è centrata su un ambiente marginale con caratteristiche estreme come rupi e scogliere lungo la costa.
Poliploidia
l fenomeni di speciazione si accompagnano spesso a un aumento del livello di ploidia in quanto la mutazione responsabile della formazione di nuove specie può essere seguita dal raddoppio del numero cromosomico. Alternativamente il nuovo genotipo si può formare per ibridazione di due genotipi preesistenti affini e in seguito venire raddoppiato. Si ritiene che ciò avvenga nella nostra flora in circa la metà dei casi, con gruppi che presentano frequentemente poliploidia (pteridofite, Rosa, Leucanthemum, Festuca, Carex) e altri nei quali la poliploidia è praticamente inesistente (gimnosperme, fagacee). L'ambiente mediterraneo, da questo punto di vista, presenta condizioni del tutto particolari; la vegetazione climax offre solo limitate occasioni di speciazione, poiché si tratta di un ambiente nel quale quasi tutte le nicchie sono saturate, mentre la vegetazione sottoposta a disturbo si trova in condizioni di elevata instabilità: si vengono così a determinare nuove nicchie ecologiche, alle quali nuovi genotipi si possono eventualmente adattare.
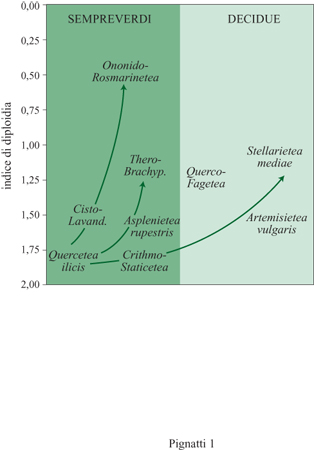
Queste condizioni si riflettono sulla frequenza dei poliploidi, la cui ripartizione ecologica è estremamente diversificata (Pignatti, 1978): essi infatti risultano rari nella vegetazione stabile, come le ceno si climax a carattere forestale, e più frequenti nei tipi di vegetazione sottoposti ad azioni di disturbo (fig. 1). Si può quindi affermare che un'azione di disturbo rende possibile l'evoluzione e la stabilizzazione di nuove specie, eventualmente con processi di poliploidizzazione. l poliploidi risultano quindi più frequenti negli ambienti disturbati.
La selva sempreverde di tipo mediterraneo
Alla fine del Pliocene l'ambiente mediterraneo era caratterizzato da una vegetazione climatogena (vegetazione naturale che si sviluppa in dipendenza dei fattori generali del clima) in un clima di tipo mediterraneo caldo: si trattava di una vegetazione forestale sempreverde o caducifoglia oppure di savana. Le associazioni climatogene sono floristicamente molto stabili e povere (nella lecceta attualmente vi sono soltanto 10÷ 25 specie su una superficie standard di 100 m²). Esse rappresentano un ambiente conservativo, che non favorisce l'evoluzione delle specie vegetali. Prevalgono piante appartenenti alle fagace e, alle oleacee e alle ericacee, ed elementi ad affinità tropicale come i generi Laurus, Myrtus, Smilax, Rubia, mentre le famiglie che forniscono il massimo contingente di specie nella flora mediterranea (cariofillacee, crucifere, papilionacee, composite, e anche le stesse graminacee) sono poco rappresentate nella vegetazione climatogena.
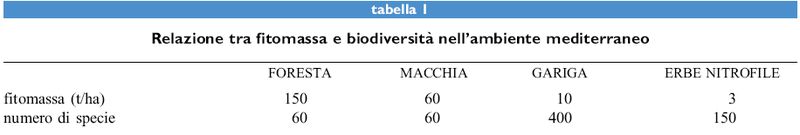
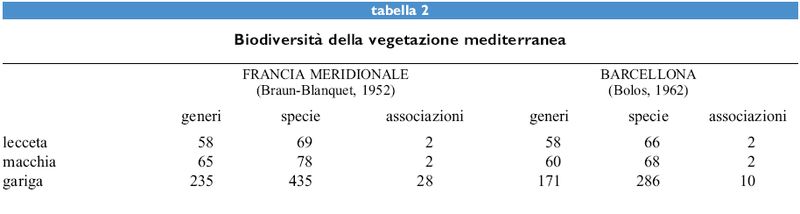
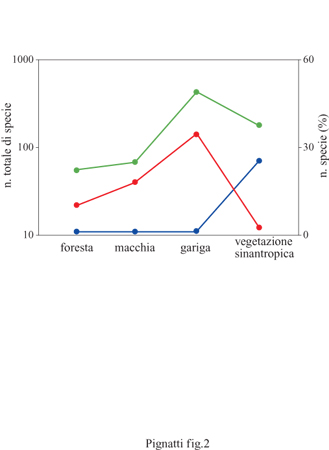
La flora della foresta è, come già detto, abbastanza povera; secondo lo studio di J. Braun-Blanquet (1937), che abbraccia tutta la Francia mediterranea, vi sono appena una sessantina di specie legate a questo tipo di ambiente (tab. I). La situazione non cambia nella macchia, che si differenzia soprattutto per il leccio, con portamento arbustivo anziché arboreo, ma per il resto ha una composizione specifica abbastanza simile. La gariga (tab. 2), invece, cui si arriva per un'ulteriore degradazione da incendio e pascolo (fig. 2), presenta una flora molto più ricca (Naveh e Whittaker, 1979). In base a un principio attualistico, si può ritenere che in questa situazione la maggiore diversificazione della flora sia avvenuta al di fuori della vegetazione climatogena, cioè negli ambienti extrasilvatici, come vette, creste, rupi, brecciai e sulla costa nelle scogliere.
Evoluzione per riduzione
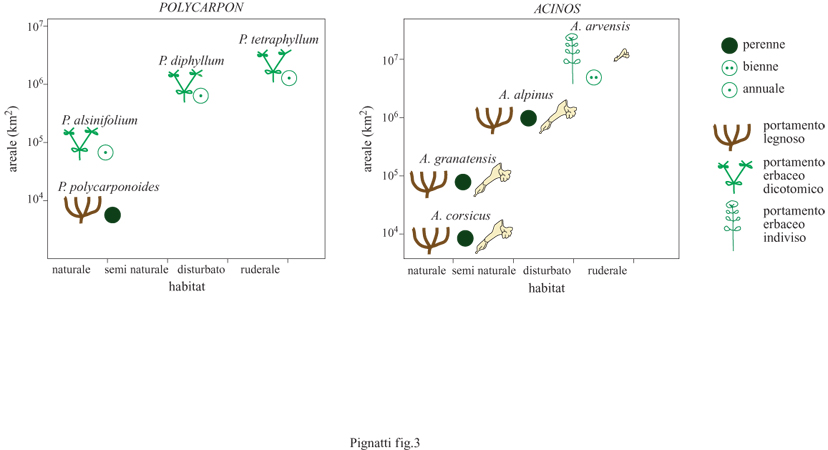
La flora d'Italia, composta da circa 6000 specie appartenenti a più di 1100 generi, mostra relazioni non casuali tra morfologia, biologia, ecologia e distribuzione delle singole specie; i risultati sembra possano venire generalizzati all'intera flora mediterranea. Nei gruppi più diversi si possono riconoscere specie che consideriamo ancestrali oppure derivate (fig. 3), che nei casi estremi presentano i differenti caratteri. Le specie ancestrali hanno un apparato vegetativo ben sviluppato, sono piante legnose arbustive o fruticose, o comunque perenni, diploidi, hanno un areale ristretto, spesso puntiforme, e un habitat naturale in ambienti marginali come vette, rupi, brecciai, scogliere. Le specie derivate, al contrario, hanno un apparato vegetativo ridotto, sono erbe annuali, poliploidi, presentano un areale amplissimo, spesso panmediterraneo, e un habitat in ambienti creati dall'uomo.
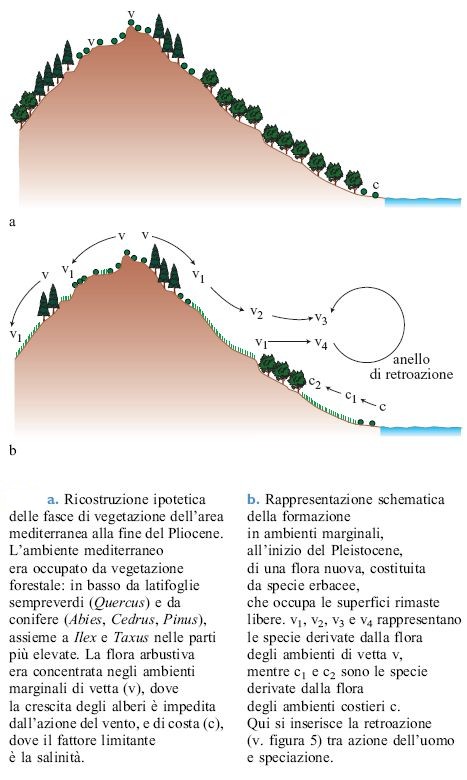
Tra i due estremi esiste una miriade di casi intermedi. È stato descritto un modello (Pignatti, 1979) nel quale si ipotizza che le specie ancestrali alla fine del Pliocene fossero limitate agli habitat marginali, in quanto le formazioni climatogene forestali occupavano le stazioni più favorite con ceno si a bassa diversità (fig. 4). Negli habitat marginali si costituì così un serbatoio genetico dal quale ebbe inizio un intenso processo di speciazione durante il Pleistocene. Questo processo dipese dalla formazione di nuove nicchie aperte, in grado di venire colonizzate da genotipi del tutto nuovi, e portò a un frequente passaggio alla poliploidia.

Le nuove nicchie si formarono attraverso alcuni meccanismi. Le variazioni climatiche connesse alle glaciazioni determinarono una contrazione della vegetazione forestale, l'espansione di fasce steppiche, variazioni del livello marino e fenomeni di glacialismo sia pure limitati a pochi gruppi di alte montagne. Tuttavia, a partire dall'interglaciale Riss-Würm cominciarono a farsi sentire anche le conseguenze dell'azione dell'uomo sulla vegetazione naturale. Si avviò in questo modo un imponente processo di speciazione (v. figura 4b), probabilmente con tempi relativamente rapidi, che ha certamente avuto conseguenze molto importanti sull'intera flora mediterranea. Anche in questo caso si verificò l'invasione di ambienti sgombri da concorrenza. Le riserve genetiche erano localizzate negli ambienti marginali dove, come si è visto prima, si era specializzata una flora arbustiva ed erbacea. Piante degli ambienti di vetta, delle creste, rupi, scogliere, brecciai diedero origine a una flora completamente nuova che colonizzò gli ambienti disturbati per opera dell'uomo (tab. 3). In questa flora prevalevano le specie opportuniste che, sfruttando il ciclo breve (pochi mesi) e l'intensa produzione di semi, furono in grado di propagarsi con grande efficienza. Le specie annuali costituiscono oggi più del 30% della flora mediterranea ammontando a circa 5000÷7000 specie, e una parte consistente di queste potrebbe aver avuto origine attraverso il meccanismo sopra indicato. Al contrario l'ambiente forestale, nel quale tutte le nicchie risultavano saturate, rimase privo di queste specie opportuniste e non diede origine a particolari processi di speciazione.
Ecologia del fuoco e origine della gariga
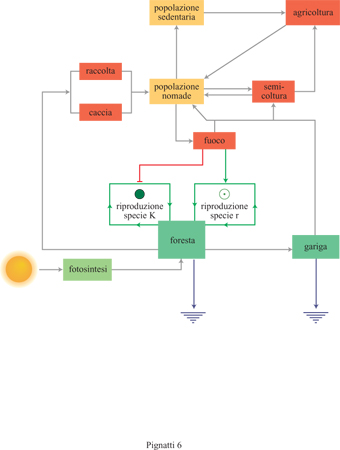
La foresta sempreverde con dominanza di leccio è soggetta, in condizioni naturali, alla degradazione per opera del fuoco (Trabaud e Lepart, 1980). Nell'ecosistema mediterraneo tale fattore può essere considerato naturale, e poteva agire anche prima che l'uomo si insediasse in questa regione. In effetti ancora oggi si può osservare l'azione di incendi spontanei in aree a vegetazione di tipo mediterraneo dell'Australia Occidentale e del Sud-Africa. Da quando l'uomo ha occupato il bacino mediterraneo, tuttavia, agli incendi per cause naturali, relativamente rari, sono venuti ad aggiungersi anche gli incendi provocati, che sono certamente più frequenti. L'uomo usava il fuoco in vari modi, in relazione alla caccia: dallo studio delle ultime comunità di cacciatori-raccoglitori ancora esistenti (aborigeni australiani, Khoi-San sudafricani) si è compreso che l'uomo incendiava la vegetazione per stanare la selvaggina provocando incendi capaci di propagarsi per decine di chilometri; oppure bruciava la sterpaglia in modo da far sviluppare in poche settimane una vegetazione erbacea effimera in grado di attirare gli erbivori, concentrando li e agevolandone la caccia. Si può dunque ammettere che nel bacino mediterraneo ampie superfici di foresta venissero incendiate e quindi trasformate in un habitat vuoto; questo risultava quindi disponibile per i nuovi genotipi che casualmente si fossero formati attraverso mutazione, ibridazione e poliploidia. Con l'avvento della cultura neolitica, che nel Mediterraneo si sviluppò probabilmente attorno al 9000÷6000 a. C., cominciò l'espansione della pastorizia, che provocò ulteriori danni al bosco, soprattutto per il diffondersi della capra. Con l'età del Ferro, circa 3000 anni fa, l'uomo fu in grado di distruggere il bosco mediante l'anellatura degli alberi più longevi e mediante il taglio. In tutti questi casi il risultato fu sostanzialmente lo stesso: la foresta sempreverde venne sostituita da altri tipi di vegetazione arbustiva ed erbacea (v. figura 4).
La gariga è la vegetazione che segue l'incendio, ha l'aspetto di una macchia bassa, con singole specie arbustive di piccola taglia (come cisti, eriche e rosmarino) alternate a piccoli prati e copre spesso una superficie di pochi metri quadrati. Il termine gariga si ritrova in tutta l'area dell'idioma occitanico, dalla Catalogna al sud della Francia e alle vallate alpine del Cuneese in Piemonte. Le condizioni ecologiche sono molto variabili, per il maggiore o minore spessore del suolo, la presenza di pietre, la maggiore o minore acidità del substrato, la possibilità di formazione di pozze o acquitrini durante la stagione umida; si hanno così differenti associazioni e combinazioni specifiche sempre diverse. L'ambiente di gariga è costituito dunque da un mosaico di molte associazioni, e questo ne spiega l'elevata densità floristica. Nel Mediterraneo orientale si hanno formazioni simili, come nel caso di Phrygana in Grecia e Batha in Israele, altrettanto ricche, ma entrambe caratterizzate dalla prevalenza di arbusti spinosi. L'Italia, per la sua posizione intermedia, si trova ad avere esempi sia di gariga (soprattutto in Liguria, Toscana, Sardegna) che di Phrygana (in Puglia, Sicilia e anche in Sardegna, presso Cagliari). Nella gariga si hanno pratelli effimeri con graminacee (Avena, Briza, Catapodium, Cynosurus, Dactylis, Lolium, Vulpia), leguminose (Lathyrus, Lupinus, Medicago, Pisum, Trifolium, Vicia), composite (Crepis, Geropogon, Hypochoeris) e altri generi come Capsella, Draba, Erophila, Echium, Erodium, Geranium, Hutchinsia, Sherardia. Quando l'incendio si ripete frequentemente il suolo viene lisciviato fino a costituire una sorta di sabbia silicea sulla quale diventano prevalenti le cistacee (Tuberaria, Helianthemum). Tra gli arbusti, su terreno ricco in calcare, si hanno timo e rosmarino, e talora la Phrygana a Sarcopoterium spinosum.
Quando l'azione dell'uomo sull'ambiente e sulla vegetazione si fa ancora più intensa, si passa agli aggruppamenti di erbe, che si possono osservare vicino agli abitati, negli stazzi, nei luoghi di raccolta e riposo del bestiame: sono tutti ambienti nei quali si accumula materia organica, sotto forma di scarti, rifiuti, escrementi dell 'uomo e degli animali, che arricchiscono il suolo in nitrati. Ne deriva una flora di erbe nitrofile con Atriplex, Borago, Chenopodium, Cichorium, Diplotaxis, Parietaria, Raphanus, Rumex, Scorzonera, Sonchus, Tanacetum, Taraxacum, Urtica; molte delle quali sono commestibili, almeno dopo la cottura.
Per meglio approfondire le relazioni tra l'uomo, come autore dell'impatto e in tal modo inconsapevole generatore di nuove nicchie, e la flora, sarà ora necessario considerare quelle piante che possono essere ritenute utili per l'uomo nella cultura del Paleolitico. In questo stadio della cultura umana il mondo vegetale rappresentava la massima risorsa (Hehn, 1911): l'uomo da esso ricavava utensili, medicine, cibo e molte altre cose, anche se l'utilizzazione a scopo alimentare era quantitativamente prevalente.
Nell'ambiente della foresta sempreverde non vi sono molte piante alimentari: si possono ricordare il corbezzolo e nelle condizioni più aride l'olivo e il carrubo; vi si trovano inoltre i funghi che però trattandosi di ambiente arido sono relativamente scarsi. Le ghiande delle querce sempreverdi come leccio e sughera sono ricche di tannino e non risultano commestibili. Al contrario le ghiande di Quercus tenoreana, che vive in ambiente collinare a clima submediterraneo, sono dolci e in epoca romana costituivano un cibo di base per la popolazione più povera; esse venivano ancora raccolte e commerciate fino ai primi anni del 20° secolo. Anche il castagno ha avuto grande importanza alimentare. Queste specie arboree, tuttavia, vivono nella fascia del bosco caducifoglio e non si presentano in ambiente mediterraneo.
La flora della gariga possiede numerose specie alimentari, nessuna delle quali, presa singolarmente, è in grado di mantenere una popolazione, ma nel loro insieme, certamente tali da fornire un'alimentazione variata ed equilibrata. Inoltre, nella maggior parte dei casi si tratta di frutti secchi, facilmente conservabili. Vi troviamo (Pignatti, 1995) piante amilacee: le graminacee come orzo, avena, segale e forme primitive di frumento quali Aegylops e Dasypyrum; bulbi e tuberi amilacei come asfodelo, lampagione, cipolle selvatiche, Orchis; semi ricchi di proteine quali leguminose come veccia, lenticchia, cece, lupino, fava; semi oleaginosi come lino, colza; verdure quali valerianella, barba di becco, radicchio. A questi cibi di base si può poi aggiungere una relativa abbondanza di aromi, come l'aglio, il rosmarino, l'origano, la menta, la mentuccia, la salvia, il timo, la lavanda, il mirto, il ginepro, il finocchio, il prezzemolo, molti dei quali hanno anche azione medicinale.
Per quanto riguarda la flora nitrofila (Pignatti, 1983; 1987) che vive attorno agli insediamenti, si può notare una serie quasi infinita di verdure come Urtica, Atriplex, Chenopodium, Silene, Crepis, Taraxacum dalle quali derivano insalate, radicchio, spinaci selvatici, cicoria, scorzonera e tante altre; mancano invece cibi di base ricchi di proteine o carboidrati assimilabili.
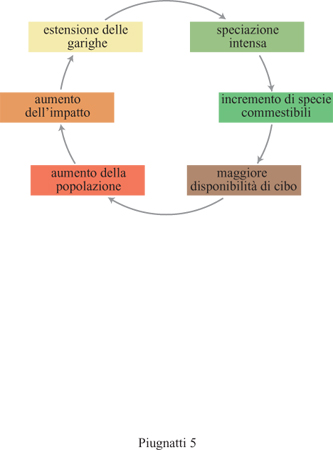
Si viene così a stabilire un anello di retro azione (fig. 5), in quanto l'estendersi della gariga determina un aumento nella disponibilità di cibo, questo a sua volta determina un aumento della popolazione, si hanno così incendi della vegetazione più frequenti e di conseguenza di nuovo si estende la gariga. Il sistema lavora in ciclo e il processo assume un carattere autocatalitico.
Questa vegetazione, che ebbe un ruolo fondamentale per sostenere la popolazione umana prima e durante il passaggio dalla condizione di cacciatore-raccoglitore alla domesticazione delle piante commestibili, è una flora effimera, così come effimero è l'ambiente che segue l'incendio nel quale essa vegeta. Le specie che la costituiscono hanno vita breve e spesso completano il loro ciclo vitale in 1÷3 mesi soltanto. Si tratta comunque di una presenza imponente: le specie sono migliaia, alcune delle quali molto frequenti. Questo fenomeno è stato paragonato (Pignatti, 1978) alla comparsa di strutture dissipative secondo la termodinamica dei processi irreversibili elaborata da I. Prigogine (fig. 6).
L'agricoltura rappresenta il più antico mezzo ideato dall'uomo per concentrare l'energia e gestirla al fine di sopperire alle necessità del sistema. Infatti, per tutto lo sviluppo dell'umanità, fino alla rivoluzione industriale, l'uomo è stato per le sue necessità interamente dipendente, in maniera diretta o indiretta, dall'energia solare. Il Sole però eroga energia in una forma diffusa che non può venire conservata o trasferita. La coltivazione di piante utili permette invece di ottenere prodotti ricchi di energia (come le granaglie), che possono venire utilizzati come fonte energetica al momento e nel luogo nei quali risultano maggiormente necessari. Diviene così possibile la costituzione di città e di società complesse.
L'orto mediterraneo
Nelle prime fasi dell'agricoltura le terre venivano considerate m bene comune, ed è verosimile che con l'utilizzo di attrezzi :udimentali in legno e pietra lavorata si sia sviluppata una sorta di agricoltura itinerante. Con l'avvento della metallurgia (già nel 3000÷5000 a. C.) divenne possibile la costruzione di attrezzi più efficienti (zappa, vanga, falcetto) e soprattutto dell'aratro, che consentì di cominciare a sfruttare la forza degli animali da lavoro (soprattutto bovini, ma anche cavallo, asino, mulo, cammello), permettendo un'ulteriore concentrazione dell'energia. La conseguenza di questa maggiore efficienza fu la sedentarizzazione della popolazione; per la produzione agricola questo comportò un'ulteriore evoluzione, soprattutto con l'irrigazione e la concimazione. L'uomo cominciò cioè a plasmare il paesaggio (Naveh e Lieberman, 1984).
La produzione venne concentrata su superfici sempre meglio attrezzate, e ciò si ottiene con l'introduzione di piante legnose. Il contadino affrontò il problema in maniera creativa con una imitazione della lecceta, usando piante utili (Hehn, 1911) come olivo, cappero, corbezzolo, vite, fico (tutte spontanee nella vegetazione forestale mediterranea). Si arrivò così al modello dell'orto mediterraneo (Sereni, 1961), sviluppato si soprattutto in Grecia e nelle isole.
A questo punto è fondamentale considerare la possibilità che il sistema che andava differenziandosi potesse ricevere un input di ordine dall'esterno (Pignatti, 1991). Questo poteva essere di natura diversa, purché si trattasse di un vincolo che abituasse la popolazione a determinati comportamenti, ma non fosse troppo intensivo, altrimenti lo sviluppo della società ne risulterebbe bloccato. Per le culture fluviali (Egitto, Mesopotamia) tale vincolo fu la dipendenza dalle regolari piene dei fiumi, per le città costiere greche fu la scarsità di terre alluvionali coltivabili che costrinse a costituire colonie oltremare, per i popoli italici fu l'insediamento su colline scoscese, quasi fortezze naturali. In queste condizioni sorse e si consolidò la civiltà mediterranea, da principio legata a piccole città-stato (la cultura della polis, Pignatti, 1994).
Diffusione transcontinentale
La storia successiva è nota (Cavalli-Sforza e Cavalli-Sforza, 1993; Diamond, 1997). Le colture iniziate nel Mediterraneo già durante l'evo Antico si diffusero verso nord (Europa settentrionale), sud (Africa subtropicale) e soprattutto est (fino alla valle dell'lndo e del Gange). Dopo le grandi scoperte geografiche alla fine del Medioevo esse vennero introdotte anche nelle Americhe e successivamente in Sud-Africa e in Australia. Oggi il frumento (in generale con la segale, l'orzo e l'avena) è coltivato in tutte le zone temperate del globo e da qui irradia nella fascia temperato-fredda, in Europa fino a 65° lat. N, e nella fascia subtropicale e tropicale; autentici granai si hanno in USA, Canada, Argentina e Australia. Tra le piante che non sono originarie del bacino del Mediterraneo, soltanto due (riso e mais) raggiungono una diffusione comparabile.
La ragione principale del successo delle colture originatesi nel clima mediterraneo è probabilmente collegata alle caratteristiche eco fisiologiche delle specie mediterranee, che sono soprattutto erbe annuali a ciclo breve che germinano in autunno (oppure all'inizio della primavera), fioriscono in primavera e maturano i frutti in maggio-luglio. In questo clima esse si adattano egregiamente alle condizioni di vita del campo, che appunto ospita le specie coltivate soltanto per un periodo di alcuni mesi, durante il quale esse devono arrivare a compiere interamente il loro ciclo vitale; dopo il raccolto il terreno rimane a riposo.
Possiamo a questo punto tentare di rispondere alla domanda che ci siamo posti all'inizio di questo saggio: perché la domesticazione delle piante agrarie sia avvenuta proprio nel bacino del Mediterraneo e non negli altri eco sistemi a clima mediterraneo. La risposta deve tener conto di due fattori: i tempi evolutivi e i caratteri biologici dei diversi stock floristici.
Per quanto riguarda i tempi, va tenuto presente che il processo di domesticazione è il risultato di una stretta interazione tra uomo e vegetali: quanto più lungo è il periodo nel quale ciò è avvenuto tanto più è probabile il successo. Da questo punto di vista il Mediterraneo si trova in una posizione singolare rispetto agli altri eco sistemi di tipo mediterraneo: l'uomo qui era certamente già presente come Homo erectus e in epoca assai antica, come è dimostrato da diversi siti (Monte Carmelo, Ternifine, Atapuerca, Isernia) che vengono datati a 0,5÷1 milione di anni fa. Da allora la presenza dell'uomo nel Mediterraneo è stata costante, prima come Homo erectus e successivamente come uomo di Neandertal e come Homo sapiens, fino dalla comparsa della nostra specie. L'uso del fuoco è documentato da almeno mezzo milione di anni; forse all'inizio l'uomo utilizzava l'accensione spontanea d'incendi, ma già nel sito di Terra Amata (200.000 anni fa) sembra esser stato in grado di accendere il fuoco per le proprie necessità. Al confronto, l'occupazione umana nelle altre aree a clima mediterraneo è molto più breve: 40.000 anni in Australia, 12.000 in Califomia, 11.000 in Sudamerica; in Africa l'uomo è presente frn dalle ultime fasi del Pliocene, però sempre in aree climaticamente non mediterranee, come la savana in Tanzania ed Etiopia e la prateria d'altitudine nel Transvaal; l'occupazione della Provincia del Capo, che ha clima mediterraneo, è molto recente, probabilmente non risale a molto più di 10.000 anni fa (Diamond, 1997). In sostanza, l'uomo nel bacino del Mediterraneo ha potuto convivere con le piante commestibili per un periodo 25÷100 volte più lungo rispetto ai suoi confratelli nelle altre aree a clima mediterraneo, e questo indubbiamente ha rappresentato un decisivo vantaggio per la domesticazione.
Anche le caratteristiche biologiche intrinseche della flora mediterranea non hanno minore significato. Essa deriva da uno stock antico (atlantico e tetisiano), ma le vicende successive l'hanno modificata permettendo l'emergenza di tipi opportunisti, eurieci e in grado di evolversi rapidamente per ibridazione e poliploidia. In seguito all'impatto dell'uomo, soprattutto attraverso il fuoco, questa flora reagì con un imponente processo di speciazione. Niente di simile è avvenuto negli altri eco sistemi mediterranei: la flora dell'Australia Occidentale è quasi priva di specie erbacee e le specie annuali sono limitate alle effimere fioriture dopo le piogge negli ambienti desertici; in Sud-Africa si ha la specializzazione di piante bulbo se; in California e Cile non si ha una simile abbondanza di specie in grado di colonizzare gli ambienti creati dall'uomo. Speciazione per riduzione e coevoluzione uomo-vegetazione sono fenomeni del tutto peculiari dell'ambiente mediterraneo.
Conclusione
La flora e la vegetazione della regione mediterranea presentano una biodiversità particolarmente elevata. Ciò è la conseguenza di avvenimenti antichi e recenti che hanno influenzato profondamente la biogeografia di quest'area; anche l'Italia, situata in posizione centrale, ne è stata interessata. In particolare, si è avuto un intenso processo di speciazione collegato alla diffusione dell'uomo. La comparsa di nuove specie causata dall'aumento di popolazione ha portato non soltanto a un aumento numerico della flora mediterranea, ma anche alla sua diversificazione, attraverso la selezione di specie annuali o comunque a ciclo breve e un significativo aumento della poliploidia. Tra lo sviluppo della popolazione umana e l'evoluzione della flora si è stabilito un legame di retro azione che ha impresso al sistema un andamento ciclico. La coevoluzione uomo-vegetazione ha costituito un fattore decisivo per lo sviluppo della civiltà in ambiente mediterraneo e rappresenta un avvenimento probabilmente unico nella storia dell'umanità.
Bibliografia citata
BRAUN-BLANQUET, J. (1937) Les groupements végétaux de la France méditerranéenne. Parigi, CNRS.
CAVALLI-SFORZA, L., CAVALLI-SFORZA, F. (1993) Chi siamo. La storia della diversità umana. Milano, Mondadori.
DIAMOND, J. (1997) Guns, germs and steel: the fates of human society. New York, W.W. Norton & Co.
DI CASTRI, F., MOONEY, H. (1977) Mediterranean type ecosystems, origin and structure. Berlino-New York, Springer-Verlag.
GREUTER, W., BURDET, H.M., LONG, G. (1984-89) Med-Checklist, volI. I, 3, 4, Ginevra, Conserv. Jard. Bot.
HEHN, V. (1911) Kulturpflanzen und Haustiere in ihrem Übergang aus Asien nach Griechenland und ltalien sowie in das übrige Europa. Berlino, BomtraegeI.
MALYSHEV, L.I. (1975) The quantitative analysis of flora: spatial diversity, level of specific richness and representativity of sampIe areas. Botan. Zhurn., 7, 1537-1550.
NAVEH, Z., LIEBERMAN, A.S. (1984) Landscape ecology, theory and application. New York, Springer-Verlag. NAVEH, Z., WHITTAKER, R. (1979) Structural and floristical diversity of shrublands and woodlands in northem Israel and other mediterranean areas. Vegetatio, 41, 171-190.
PIGNATTI, S. (1978) Evolutionary trends in mediterranean flora and vegetation. Vegetatio, 37,175-185.
PIGNATTI, S. (1979) Plant geographical and morphological evidences in the evolution of the mediterranean flora (with particular reference to the italian representatives). Webbia, 34, 243- 255.
PIGNATTI, S. (1983) Human impact in the vegetation of the mediterranean basin. In Man 's impact on vegetation, a c. di Holzner W., Werger M.J.A., Ikusima I., Boston-Hague, Dr. W. Junk PubI., pp. 151-161.
PIGNATTI, S. (1986) The consequence of climate on the mediterranean vegetation. Annali di Bot., 42, 123-130.
PIGNATTI, S. (1987) The relationships between natural vegetation and social system in the mediterranean basin. In Vegetation ecology and creation of new environments, a c. di Miyawaki A. et al., Tokyo, Tokai University Press, pp. 35-45.
PIGNATTI, S. (1991) Man-enviroument relationships in the earliest phases of civilization. Biology Forum, 84, 83-90.
PIGNATTI, S. (1994) Ecologia del paesaggio. Il paesaggio vegetale italiano. Torino, UTET.
PIGNATTI, S. (1995) Biodiversità della vegetazione mediterranea. Atti dei Convegni Lincei, 115, 7-31.
PIGNATTI, E., PIGNATTI, S. (1989) Life forms distribution and phytogeographical affinities of the canarian flora. Flora, 183, 87-95.
RENFREW, J.M. (1973) Paleoethnobotany. The prehistoric food plants of the Near East and Europe. New York, Columbia University Press.
SERENI, E. (1961) Storia del paesaggio agrario italiano. Bari, Laterza.
TRABAUD, L., LEPART, J. (1980) Diversity and stability in garrigue ecosystems after fire. Vegetatio, 43, 49-57.