
Competizione spermatica
Competizione spermatica
Quando una femmina si accoppia con molti maschi gli spermi rivali entrano in competizione per fecondare le uova disponibili e la femmina può essere in grado di esercitare un controllo a livello dell'apparato riproduttore. Le differenze nella morfologia e nella fisiologia dell'apparato riproduttore femminile determinano quali sono le caratteristiche importanti dell'eiaculato per il successo dei maschi. La conoscenza dei meccanismi fisiologici coinvolti è essenziale per capire quanto le strategie maschili e femminili possano riflettersi nel successo dei maschi. La competizione spermatica è stata un fattore di selezione per l'evoluzione di una maggiore produzione di spermi, di eiaculati più effìcienti, di tassi più alti di copulazione e del controllo della compagna nei maschi delle specie poliandriche. Anche all'interno delle popolazioni vi potrebbero essere differenze nell'investimento nell'eiaculato a seconda dei livelli di competizione spermatica incontrati da maschi che seguono differenti tattiche riproduttive. Poiché maschi e femmine utilizzano strategie differenti per rendere massimo il loro successo riproduttivo in taluni casi può sorgere un conflitto tra sessi.
Che cosa è la competizione spermatica
Charles Darwin sviluppò il concetto di selezione sessuale per spiegare l'evoluzione di caratteri maschili cospicui, come ornamenti e armature, che non trovavano una giustificazione con la teoria della selezione naturale per la sopravvivenza del più adatto. Tali caratteri conferiscono ai maschi vantaggi in termini di competizione per il partner, sia perché migliorano la competitività negli incontri agonistici tra i maschi, sia in quanto rendono i maschi attraenti per le femmine. Dunque, la selezione sessuale comprende due differenti processi: la competizione tra maschi e la scelta da parte della femmina. Tradizionalmente, gli ecologi del comportamento hanno studiato la selezione sessuale con l'assunto che il successo nel trovare un partner fosse equivalente al successo riproduttivo. Così si riteneva che i maschi competessero per accoppiarsi con quante più femmine possibile e le femmine fossero in grado di scegliere quali maschi potessero accoppiarsi con esse solo a livello comportamentale. La copulazione era considerata il punto finale di entrambi i processi. Questa visione è stata notevolmente modificata quando è divenuto chiaro che le femmine di molte specie si accoppiano con più di un maschio durante ciascun ciclo sessuale. Ciò ha implicazioni importanti in quanto consente di ipotizzare che sia la competizione tra i maschi sia la scelta delle femmine proseguano a livello fisiologico dopo la copulazione, e ciò significa che l'accesso sessuale alle femmine non assicura la fecondazione. Quando le femmine si accoppiano con più maschi gli eiaculati dei maschi rivali competono per fecondare le uova a disposizione e d'altra parte le femmine nel loro apparato riproduttore potrebbero esercitare la scelta di quale sperma opererà la fecondazione. In termini stretti la competizione spermatica si riferisce solo alla competizione tra eiaculati rivali ponendo enfasi sugli adattamenti del maschio. Tuttavia, si sta cominciando a prestare più attenzione al punto di vista della femmina.
La competizione spermatica sembra essere largamente diffusa tra gli animali (Smith, 1984) così come tra le piante esiste la competizione tra pollini. Le femmine degli insetti immagazzinano gli spermi in organi specializzati, le spermateche, in cui essi rimangono vitali per periodi prolungati di tempo (anche anni) rendendo la competizione spermatica possibile anche se la femmina si riaccoppia dopo un lungo intervallo di tempo (Parker, 1970). Per queste ragioni gli insetti furono per un certo periodo il principale soggetto di studio sulla competizione spermatica.
Si pensava che gli uccelli non fossero buoni candidati per studiare la competizione spermatica in quanto gran parte delle specie sono socialmente monogame e il maschio e la femmina formano coppie che cooperano nell'allevamento della prole. Tuttavia, questo quadro è completamente cambiato da quando l'applicazione di tecniche molecolari per determinare la paternità (per esempio, DNA fingerprinting, impronte digitali del DNA) ha rivelato che sia i maschi sia le femmine sono spesso sessualmente infedeli e s'impegnano in copulazioni extracoppia (EPCs, Extra Pair Copulations) col risultato di nidiate a paternità mista (Birkhead e Møller, 1992). Queste scoperte hanno rivoluzionato lo studio dei sistemi riproduttivi degli uccelli. Anche tra i mammiferi ci si attendeva che la competizione spermatica fosse rara, perché in gran parte delle specie le femmine non possiedono organi specializzati di immagazzinamento di spermi e, di conseguenza, gli spermi hanno una vita breve. Tuttavia le osservazioni comportamentali suggeriscono che in molte specie di mammiferi gli accoppiamenti da parte di maschi rivali avvengono in un intervallo di tempo talmente breve che gli spermi rivali possono competere; la competizione spermatica avviene persino tra alcune specie in cui l'evidenza osservazionale suggeriva la monoandria, in quanto le femmine producono figliate provenienti da più genitori o, in specie con figli unici, prole di maschi diversi da quello che si osserva in accoppiamento con la femmina nel periodo dell'ovulazione (Gomendio et al., 1998).
La notevole discrepanza tra le attese previste, sulla base dei livelli di competizione spermatica basati sulle osservazioni comportamentali, e la paternità genetica osservata è in parte una conseguenza della nostra ignoranza sui meccanismi fisiologici che regolano questi fenomeni. È necessario chiarire questi meccanismi per capire come i pattern di accoppiamento si riflettano nell' effettiva paternità.
Meccanismi di competizione spermatica
I meccanismi fisiologici alla base della competizione spermatica differiscono molto tra i gruppi tassonomici in quanto 'l'arena' in cui gli eiaculati rivali dovrebbero competere, cioè l'apparato riproduttore femminile, mostra un sorprendente grado di variabilità. Un'analisi comparativa effettuata negli uccelli (Birkhead, 1996) e nei mammiferi (Gomendio et al., 1998) illustrerà tale diversità.
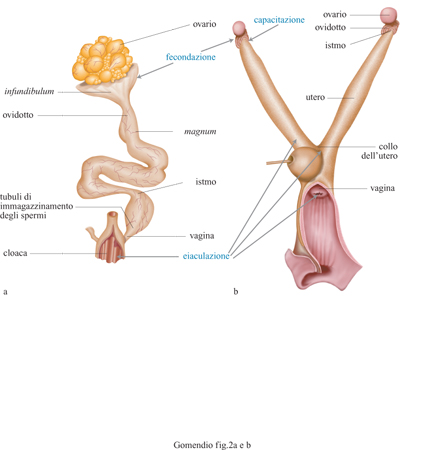
Le femmine degli uccelli possiedono tubuli di immagazzinamento degli spermi, localizzati a livello della giunzione uterovaginale (fig. 2). Solo una piccola frazione dell' eiaculato entra nei tubuli (circa 0,1%); dunque vi è una drastica riduzione del numero di spermatozoi nella vagina. Immagazzinando gli spermi nei tubuli le femmine assicurano un' ade guata riserva di spermi che dura molti giorni durante i quali avvengono ovulazione, fecondazione, deposizione delle uova. Un'analisi condotta in specie diverse ha mostrato che la durata di immagazzinamento degli spermi è correlata positivamente alla durata della deposizione delle uova. I tubuli di immagazzinamento degli spermi potrebbero anche proteggere lo sperma dall'espulsione dall'apparato riproduttore femminile ogni volta che un uovo viene deposto. I maschi rivali sembrerebbero competere per ottenere che una quantità maggiore dei loro spermi rimanga nei tubuli di immagazzinamento. Per un certo periodo si è pensato che gli spermi fossero in competizione nei tubuli o per stratificazione (un sistema detto last-in first-out, per cui l'ultimo arrivato è il primo a uscire) o per rimozione (l'ultimo eiaculato rimuove parte del precedente eiaculato dal tubulo). Tuttavia, osservazioni recenti suggeriscono che, dopo essere entrato nei tubuli di immagazzinamento, nel tempo lo sperma viene passivamente perduto. Così, all'aumentare del periodo di tempo dall 'ultimo accoppiamento, nei tubuli di immagazzinamento sono presenti sempre meno spermi. Ciò spiega perché quando due maschi si accoppiano con una femmina con un intervallo di tempo maggiore di quattro ore l'ultimo maschio sarà il padre genetico di gran parte della prole, poiché in quel momento nei tubuli della femmina la quantità di sperma del primo maschio è diminuita a tal punto da assicurare una predominanza dello sperma appartenente all'ultimo maschio. Per spiegare l'elevato successo delle copulazioni extracoppia è necessario anche comprendere come gli eiaculati sono prodotti e immagazzinati dal maschio. A causa delle modalità con cui gli spermi sono immagazzinati negli organi riproduttori maschili, un maschio a riposo produrrà un eiaculato con spermatozoi più mobili e in quantità maggiori rispetto a un maschio che si è accoppiato recentemente. Il successo sproporzionato delle copulazioni extracoppia può essere spiegato tenendo in considerazione due fattori. In primo luogo, le femmine smettono di accettare offerte di copulazione dal maschio della coppia all'incirca quando comincia la deposizione; così, al procedere della deposizione delle uova lo sperma di questo maschio viene gradualmente perduto dai tubuli di immagazzinamento femminili. In secondo luogo, i maschi tendono a effettuare copulazioni extracoppia solo dopo che il periodo fertile del proprio partner è terminato. Dunque, un maschio che compie una copulazione extracoppia lo fa dopo un periodo di bassa frequenza di accoppiamenti (durante la deposizione delle uova) e dunque produrrà un eiaculato molto competitivo che probabilmente avrà la meglio su quello che rimane dell'eiaculato del partner precedente.
Al contrario, le femmine dei mammiferi non possiedono organi specializzati di immagazzinamento dello sperma e di conseguenza lo sperma rimane fertile solo per circa 48 ore una volta immesso nel tratto femminile, mentre le uova rimangono fertili solo per circa 24 ore dopo l'ovulazione (v. figura 2). Le femmine dei mammiferi potrebbero non aver bisogno di immagazzinare gli spermi in quanto nell'ambito di ciascun ciclo sessuale tutte le uova vengono rilasciate e fecondate simultaneamente poco dopo l'ovulazione. Gli spermatozoi eiaculati sono trasportati passivamente verso le sezioni superiori dell'apparato femminile da movimenti muscolari e dall'attività delle ciglia che ricoprono il lume. Nelle fasi successive lo spermatozoo deve nuotare per attraversare barriere fisiche, come la giunzione uterotubarica, e percorrere le ultime sezioni dell' apparato femminile. Dopo esser passato attraverso la giunzione uterotubarica, lo spermatozoo si attacca alla mucosa della parete dell' ovidotto a livello dell' istmo inferiore e quegli spermatozoi che non ci riescono muoiono o perdono la loro fertilità. Intorno al periodo dell' ovulazione una piccola frazione degli spermatozoi presente nell 'istmo completa la capacitazione (un processo di maturazione che rende lo spermatozoo capace di fecondare) e si stacca dalla parete. In questo stadio gli spermatozoi acquisiscono un tipo peculiare di motilità, nota come iperattivazione, che costituisce una via efficiente di propulsione nei fluidi viscosi di questa sezione dell'apparato femminile.
Il primo spermatozoo che giunge in prossimità dell'uovo sembra essere quello con la più alta probabilità di fecondarlo. Al contrario di quanto in genere si pensi, in vivo il rapporto tra spermatozoi e uova è circa 1:1. Quando lo spermatozoo raggiunge l'uovo, e dopo che ha subito il processo di capacitazione, inizia a penetrare nel cumulo ooforo. La reazione dell' acrosoma (cioè il rilascio degli enzimi contenuti nel cappuccio acrosomiale) inizia probabilmente quando lo spermatozoo penetra nel cumulo ooforo; la sua azione si estende fino alla superficie della zona pellucida che viene penetrata tramite un'azione combinata di enzimi acrosomiali e movimenti della coda.
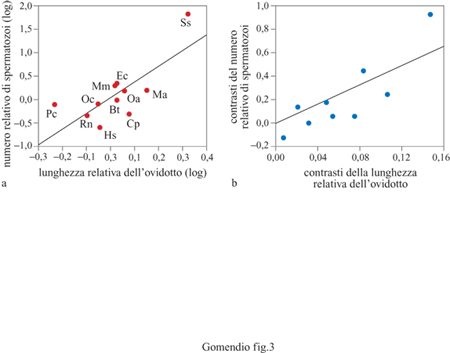
Nel percorso lungo l'apparato femminile l'eiaculato va incontro a una drastica riduzione del numero di spermatozoi e ciò suggerisce che l'apparato femminile rappresenti una formidabile barriera per lo sperma. La natura di questa barriera determina quali caratteristiche dello sperma sono selezionate dall'apparato femminile. Nei mammiferi le caratteristiche più importanti dell'eiaculato che determinano il successo della fecondazione sono: il numero, la motilità e la morfologia degli spermatozoi (strettamente correlate tra loro), e l'abilità a penetrare la membrana dell 'uovo (cioè la fase acrosomiale). La rilevanza delle barriere femminili è la grandezza che meglio rappresenta i differenti livelli di intensità di selezione sugli eiaculati che portano a una coevoluzione dei caratteri maschili e femminili. Un'analisi comparativa in diverse specie di mammiferi ha mostrato che quando le femmine possiedono ovidotti lunghi i maschi tendono a produrre eiaculati che contengono più spermi (Gomendio e Roldan, 1993a). Come vedremo, la competizione spermatica esacerba la selezione per le stesse caratteristiche dell' eiaculato selezionate dall' apparato femminile (fig. 3). Ancora non sappiamo se le femmine evolvono barriere più efficaci quando prevale la competizione spermatica al fine di aumentare la competizione tra eiaculati o di facilitare la discriminazione tra essi. Per esempio, le femmine hanno ovidotti più lunghi quando c'è competizione spermatica? Non vi sono dati a disposizione per valutare questa ipotesi.
Nei mammiferi gli spermi rivali sembrano competere per raggiungere le uova per primi in quanto ciò assicura la fecondazione, ammesso che lo spermatozoo sia in grado di penetrare la membrana dell'uovo; dunque nei mammiferi la competizione spermatica assomiglia a una gara. Una visione alternativa sostiene che nei mammiferi gli spermi rivali competono direttamente bloccando o attaccando altri eiaculati. Tuttavia i dati presentati per sostenere questa ipotesi sono piuttosto deboli e provengono soprattutto da studi condotti nella specie umana, sotto tutti gli aspetti considerata monoandrica (Gomendio et al., 1998).
Le femmine dei mammiferi sono sessualmente recettive durante periodi limitati di tempo neII'ambito di ciascun ciclo sessuale noto come estro, variabile in durata a seconda della specie. Poiché lo sperma dei mammiferi ha una vita breve i maschi corrono il rischio di non lasciare spermatozoi vivi proprio nel momento in cui una femmina va incontro all'ovulazione. Questa limitazione temporale ha portato all' evoluzione di spermi più longevi nelle specie in cui tra l'inizio dell' estro delle femmine e l'ovulazione intercorrono intervalli di tempo più lunghi (Gomendio e Roldan, 1993a). Tuttavia, quando i periodi dell'estro durano più di pochi giorni la vita fertile dello sperma non dura per l'intero periodo estrale. In questo caso, il momento in cui avviene l'accoppiamento è cruciale per il successo dell'eiaculato. Se un maschio si accoppia troppo presto gran parte del suo sperma sarà morto nel momento in cui la femmina ovula. Se la copulazione è temporalmente troppo prossima all'ovulazione non ci sarà tempo per gli spermatozoi di maturare nell'apparato femminile e anche in questo caso essi saranno incapaci di fecondare quando si verificherà l'ovulazione. Infine, le copulazioni che avvengono dopo l'ovulazione non hanno possibilità di fecondazione. Poiché la sincronizzazione tra copulazione e ovulazione è così importante, non ci sono effetti di sequenza di partner successivi nei mammiferi. Il maschio che si accoppia nel momento giusto avrà un netto vantaggio e se molti maschi si accoppiano con la stessa femmina nel momento giusto, quello in grado di trasferire un eiaculato più competitivo avrà probabilmente un successo di fecondazione più elevato.
Le tattiche riproduttive del maschio
G.A. Parker (1970; 1984) suggerì che la competizione spermatica somiglierebbe a una lotteria in cui il maschio che possieda un numero più elevato di biglietti (cioè spermi trasferiti nell'apparato femminile) avrebbe le maggiori possibilità di vittoria. Pressioni selettive per un' aumentata produzione di spermatozoi dovuta alla competizione spermatica sono state osservate in un gran numero di gruppi tassonomici che comprendono insetti, anfibi, uccelli e mammiferi. Dunque l'aumentata produzione di spermatozoi in risposta alla competizione spermatica sembrerebbe essere stata favorita in gruppi con differenti meccanismi di base, suggerendo che il suo valore è quasi universale. Un'aumentata produzione di spermatozoi potrebbe risultare in eiaculati contenenti più spermatozoi o in maschi in grado di eiaculare più spesso. Sia negli uccelli sia nei mammiferi i maschi delle specie in cui la competizione spermatica è comune hanno testicoli più grandi in relazione alla loro dimensione corporea, tendono ad accoppiarsi più spesso, e producono eiaculati con un maggior numero di spermatozoi (Birkhead e Møller, 1992; Gomendio et al., 1998).
La competizione spermatica sembrerebbe aver selezionato non solo in ragione del numero maggiore di spermatozoi, ma anche in ragione del miglioramento della loro qualità. Così i maschi dei primati poliandrici (in cui le femmine si accoppiano con più di un maschio in ogni ciclo sessuale) producono spermatozoi più mobili per eiaculato rispetto ai maschi dei primati monoandrici (Møller, 1988). Inoltre, la competizione spermatica dovrebbe selezionare in ragione di spermi che siano in grado di avere la meglio sugli spermi rivali. M. Gomendio ed E.R.S. Roldan (1991) hanno suggerito che tra i mammiferi la competizione dovrebbe favorire spermatozoi più lunghi in quanto, nuotando più velocemente, avrebbero maggiori possibilità di aggiudicarsi la vittoria, ossia la fecondazione dell'uovo. La loro analisi comparativa mostra che le specie poliandriche hanno spermatozoi più lunghi di quelle monoandriche sia nei primati sia nei roditori. In seguito la stessa relazione tra sistemi riproduttivi femminili e lunghezza degli spermatozoi è stata riscontrata nelle farfalle e negli uccelli. Non è ancora noto se in tutti questi gruppi tassonomici il vantaggio di produrre spermatozoi più lunghi a causa della competizione consiste nella capacità di nuotare più velocemente.
Vi sono gruppi di animali, come le farfalle, in cui i maschi trasferiscono alla femmina i nutrienti utilizzati per la produzione delle uova e per il proprio mantenimento incrementando così la fecondità e la longevità della femmina. In questi casi i maschi poliandrici trasferiscono una quantità di nutrienti maggiore rispetto ai maschi monoandrici (Bissondath e Wiklund, 1995).
Quando il rischio di competizione spermatica è elevato i maschi possono provare a migliorare le loro possibilità di fecondare le uova controllando la compagna, prevenendo in tal modo l'accoppiamento da parte di altri maschi o perlomeno ritardando qualsiasi altro ulteriore accoppiamento. Il controllo della compagna potrebbe assumere diverse forme, dall' esclusione aggressiva di altri maschi a copulazioni prolungate che prevengono fisicamente altri maschi dall'accoppiarsi, o alla formazione di tappi copulatori. A seconda che ci sia o no un vantaggio del primo maschio, il vantaggio dell'ultimo maschio o alcun vantaggio dipendente dalla sequenza, ai maschi conviene controllare le femmine in differenti periodi. Quando vi è il vantaggio per il primo maschio i maschi competeranno per essere in grado di localizzare le femmine per primi. Ciò ha portato in molti gruppi tassonomici all'evoluzione di organi sensori più elaborati nei maschi piuttosto che nelle femmine. Negli scoiattoli terricoli della specie Spermophilus tridecemlineatus l'ovulazione è indotta dalla copulazione e vi è un vantaggio per il primo maschio; di conseguenza i maschi non effettuano il controllo della compagna dopo l'accoppiamento. l benefici di localizzare una femmina in estro prima di altri maschi sono dunque notevoli e ciò ha consentito, durante la stagione degli accoppiamenti, l'evoluzione di capacità di movimento e ricerca molto elevate oltre a complesse abilità cognitive che permettono ai maschi di anticipare l'inizio dell'estro nelle femmine da un giorno all'altro (Schwagmeyer, 1995). Il controllo o meno delle femmine dipenderà non solo dal rischio di competizione spermatica, ma anche dai costi associati alla perdita di opportunità di accoppiarsi con altre femmine. Paragonando due specie di scoiattoli, P.W. Sherman (1989) osservò che quando altre femmine recettive sono difficili da reperire i maschi provano ad assicurarsi la fecondazione della femmina con cui si sono già accoppiati controllando la compagna, mentre quando altre femmine recettive sono prontamente disponibili, ai maschi non conviene rimanere.
Modelli teorici recenti suggeriscono che vi sono due contesti in cui maschi appartenenti alla stessa popolazione dovrebbero differire nel grado in cui investono negli eiaculati (Parker, 1990a; 1990b). In un caso, se ai maschi si assegnano i ruoli di favoriti e sfavoriti in maniera non casuale, il maschio nel ruolo sfavorito dovrebbe investire di più negli eiaculati. In un altro caso, se vi sono individui che 'si insinuano' (sneakers) e sorveglianti, gli accoppiamenti furtivi sono rari, quelli che si insinuano dovrebbero investire di più negli eiaculati e, qualora i sorveglianti siano abili ad accorgersi degli accoppiamenti furtivi, dovrebbero aumentare il numero degli spermatozoi che trasferiscono nell'apparato riproduttore femminile.
Nei pesci a fecondazione esterna le opportunità di fecondazioni furtive sono evidenti. In certe specie, come il salmone dell'Atlantico (Salmo salar), i maschi seguono due strategie alternative: possono maturare come grandi maschi anadromi che dominano i siti di deposizione delle uova e difendono le femmine, oppure maturare come minuscoli giovani salmoni precoci che ottengono fecondazioni insinuandosi mentre i maschi anadromi e le femmine sono impegnati nel controllo e nella deposizione. A supporto delle suddette affermazioni i salmoni giovani hanno un indice gonadosomatico più elevato, producono un numero relativamente maggiore di spermatozoi, una maggior proporzione di spermatozoi è mobile e i loro spermatozoi vivono più a lungo (Gage et al., 1995). In altre specie i maschi seguono differenti strategie in fasi differenti del loro sviluppo.
Nei pesci della barriera corallina appartenenti alla specie Thalassoma bifasciatum, i maschi adulti occupano e difendono i siti di deposizione mentre i maschi più giovani aggiungono il loro sperma a quello di altri maschi oppure si riuniscono nei siti di accoppiamento e si accoppiano in gruppi. Così i giovani maschi devono affrontare livelli di competizione spermatica più elevati rispetto ai maschi territoriali più anziani e, di conseguenza, hanno testicoli relativamente più grandi. Questa differenza è reversibile poiché alla fine i giovani maschi invecchiano diventando maschi territoriali e a quel punto i loro testicoli si riducono in dimensioni e la fuoriuscita totale di sperma diminuisce. Per tale motivo, essi cambiano il loro comportamento e investono nel controllo della compagna piuttosto che negli eiaculati (Warner et al., 1995).
In una popolazione di toporagni (Sorex araneus) furono osservate due strategie alternative: alcuni maschi stabilivano aree familiari (home range) che si sovrapponevano in zone a elevata densità femminile, inseminavano più femmine ed erano padri di una prole più vasta, mentre altri maschi occupavano piccole aree familiari lontane dalle femmine e dovevano percorrere lunghe distanze durante la stagione riproduttiva per visitare le aree delle femmine; questi maschi inseminavano meno femmine e producevano meno prole. L'analisi dei testicoli rivelò che nonostante le dimensioni testicolari fossero simili, il secondo tipo di maschi aveva più riserve di spermi. Secondo alcuni ciò implica che i maschi che si accoppiano nel ruolo di sfavoriti sarebbero capaci di produrre più spermi (Stockley et al., 1994). Tuttavia, ciò sembra improbabile data l'assenza di differenze nella dimensione dei testicoli. Sembrerebbe più logico pensare che il secondo tipo di maschi abbia più riserve di spermi in quanto si è accoppiato meno frequentemente durante la stagione riproduttiva. Ciò suggerisce un interessante meccanismo: i maschi che si accoppiano nel ruolo di sfavoriti potrebbero avere più spermi da risparmiare anche se si accoppiano meno frequentemente; di conseguenza, essi potrebbero eiaculare più sperma quando hanno l'opportunità di accoppiarsi. Ciò potrebbe essere valido per maschi giovani o subordinati che riescono ad accoppiarsi raramente. In accordo con le predizioni di G.A. Parker vi sarebbero differenze nella qualità dell'eiaculato, tuttavia non tali da implicare differenze nella produzione di sperma in maschi che occupano ruoli di favoriti e sfavoriti.
Una variazione dell'ipotesi di Parker prevede che un individuo maschio dovrebbe essere in grado di modificare opportunamente la qualità dell'eiaculato in base al rischio della competizione spermatica. Gli uccelli forniscono un esempio interessante poiché lo stesso maschio può incontrare un basso rischio di competizione spermatica quando si accoppia con la sua femmina, ma un rischio elevato quando compie copulazioni extracoppia. Al momento i risultati disponibili suggeriscono che i maschi non sono in grado di modificare da un punto di vista fisiologico il contenuto dell'eiaculato (Birkhead, 1996). Tuttavia, come descritto precedentemente, il fatto che i maschi tendono a compiere copulazioni extracoppia dopo la deposizione delle uova da parte della loro partner significa che, dopo un periodo di riposo, i loro eiaculati contengono spermatozoi più veloci e più numerosi. Non è chiaro se il momento in cui si verificano le copulazioni extracoppia venga scelto in base al controllo della compagna o al successo della copulazione stessa. Dunque, anche se i maschi non possiedono la capacità fisiologica di modificare gli eiaculati, gestendo il periodo in cui compiono gli accoppiamenti potrebbero raggiungere lo stesso scopo.
Le tattiche riproduttive della femmina
Adottare le prospettive del sesso femminile comporta innanzitutto chiedersi perché le femmine dovrebbero accoppiarsi con più di un maschio. Per le femmine accoppiarsi comporta dei costi perché devono dedicarvi tempo ed energie che altrimenti potrebbero essere utilizzate in altre attività; quando si accoppiano le femmine tendono a essere più facilmente individuate dai predatori e sono meno mobili, ciò rende più difficile la fuga in caso di minaccia di predazione; inoltre la mobilità ridotta potrebbe risolversi in una minore efficienza nel comportamento alimentare. Infine, i contatti ravvicinati con i maschi comportano il rischio di infezioni parassitarie o di malattie veneree. Le femmine sessualmente recettive potrebbero anche essere oggetto di molestie da parte dei maschi e ferirsi nel cercare di resistere ai tentativi di accoppiamento o durante i combattimenti maschili per accedere sessualmente alle femmine.
Quando le femmine sono controllate da un partner maschile l'accoppiamento con altri maschi può presentare ulteriori rischi se la femmina viene scoperta in quanto il partner potrebbe reagire o allontanandosi, diminuendo così le cure parentali, oppure punendo la femmina. La poliandria potrebbe anche avere conseguenze per la prole, riducendo la fitness della femmina, per esempio nel caso dell' aumento dei conflitti tra compagni di nidiata provenienti da differenti padri.
Poiché il costo dell'accoppiamento con più maschi sembrerebbe considerevole, si prospettano due ipotesi che potrebbero spiegare l'evoluzione degli accoppiamenti multipli.
Secondo la prima ipotesi le femmine non otterrebbero benefici dall'accoppiarsi con più maschi, ma incorrerebbero in costi elevati se rifiutassero di farlo. Questi costi sarebbero principalmente il risultato di molestie e danni fisici da parte dei maschi. In molti primati sociali che vivono in gruppi con più maschi, questi attaccano le femmine che rifiutano i loro tentativi di copulazione. Tali attacchi possono provocare ferite gravi e possono addirittura uccidere una femmina. Inoltre, spesso il maschio reagisce al rifiuto femminile persistendo nel corteggiamento e ciò può essere pericoloso per la femmina, in particolare nelle specie dimorfiche come alcuni mammiferi marini in cui la femmina può essere schiacciata dal maschio durante il corteggiamento. Nei gerridi, sembra che le femmine si accoppino più frequentemente di quanto sia nei loro interessi in quanto rifiutare maschi con la conseguenza di molestie ripetute può risultare più costoso che accoppiarsi, poiché tra l'altro comporta lotte per divincolarsi dai maschi e renderli meno motivati, e perché le femmine dovrebbero allontanarsi dalle aree migliori di alimentazione per evitarli (Rowe et al., 1994). Questo comportamento femminile è stato chiamato poliandria per convenienza. Sebbene vi siano contesti in cui molestie sessuali e punizioni da parte dei maschi indubbiamente determinano nettamente il grado di poliandria delle femmine, è improbabile che questi fattori siano in grado di spiegare tutti i casi che si osservano.
Secondo un'altra ipotesi le femmine traggono vantaggio dall'accoppiarsi con più maschi e i benefici prevalgono sui costi. Sebbene per un certo periodo l'opinione prevalente sia stata che la femmina sarebbe forzata ad accettare accoppiamenti con diversi maschi, molte osservazioni rivelano come la femmina spesso solleciti la copulazione di altri maschi o ricerchi attivamente altri partner sessuali. Queste osservazioni suggeriscono che in questi casi la femmina dovrebbe trarre vantaggio da accoppiamenti multipli. I possibili benefici dell' accoppiamento con diversi maschi possono essere diretti e indiretti.
Benefici diretti
Nutrienti provenienti dal maschio. - Nelle farfalle i maschi trasferiscono una spermatofora contenente nutrienti utilizzati dalla femmina per la produzione delle uova e per il proprio mantenimento. In tali circostanze la femmina trae beneficio da accoppiamenti multipli poiché ottiene più nutrienti e ciò determina una migliore prestazione riproduttiva e un'aumentata longevità (Bissondath e Wiklund, 1995). Poiché alcuni nutrienti che provengono dal maschio sono poco abbondanti nelle femmine esse si accoppiano frequentemente al fine di ottenere questi nutrienti difficili da reperire. Il numero ottimale di accoppiamenti per una femmina dipende in parte dalla qualità delle spermatofore che riceve. Produrre spermatofore è costoso per il maschio, di conseguenza maschi accoppiatisi di recente possono trasferire solo piccoli eiaculati. Le femmine che si accoppiano con questi maschi compensano aumentando il numero di accoppiamenti.
Cure paterne. - La femmina potrebbe accoppiarsi con molti maschi per aumentare il numero di padri potenziali e ottenere così più aiuto nell'allevamento della prole. Il paragone tra due popolazioni di sordo ne (Prunella collaris) e passera scopaiola (Prunella modularis) illustra elegantemente questo punto (Davies et al., 1996). Entrambe le specie nidificano in gruppi composti da due o più maschi non imparentati e due o più femmine, in cui vi è un maschio alfa che tenta di monopolizzare le femmine per avere la paternità di gran parte o di tutta la prole. Tuttavia, le femmine sfuggono ai tentativi di controllo attuati dai maschi alfa poiché è nel loro interesse accoppiarsi con altri maschi per ottenere il loro aiuto nell'allevamento della prole. Gli altri maschi non forniranno il loro aiuto a meno che non siano sicuri di esser padri di almeno una parte della prole. Le femmine sono molto attive nel sollecitare copulazioni sia da parte dei maschi alfa sia degli altri beta e ciò comporta elevati tassi di accoppiamento, talora più di un migliaio per covata (mediamente circa cento o duecento per covata). Questi tassi di accoppiamento sono ovviamente molto più elevati di quanto la femmina avrebbe bisogno per assicurare la fecondazione di tutte le uova. Il tipo di sollecitazione femminile in entrambe le specie suggerisce che le femmine tentano di rendere massimo l'aiuto dei maschi nell'alimentazione dei nidiacei. Le femmine tendono a sollecitare accoppiamenti con i maschi con cui si sono accoppiate meno volte. Nei sordoni che vivono nei Pirenei francesi i maschi alfa riducono la quantità di cure parentali al diminuire del grado di condivisione dei partner, mentre ciò non avviene per gli altri maschi. In accordo con ciò, in questa popolazione le femmine preferiscono concedere più accoppiamenti ai maschi alfa. Al contrario i maschi alfa e gli altri maschi delle passere scopaiole che vivono nel giardino botanico di Cambridge non riducono il loro aiuto al di sotto di una certa soglia critica. In questa popolazione le femmine scelgono indifferentemente i maschi alfa e gli altri maschi e sembra che tentino di concedere un numero simile di accoppiamenti a entrambi i tipi di maschi. È probabile che i maschi di sordone richiedano maggiori garanzie di paternità per prendersi cura della prole in quanto questa popolazione vive in gruppi più grandi nei quali il maschio alfa ha la possibilità di accoppiarsi con altre femmine, un'opportunità che non vale la pena di sacrificare a meno che la paternità dei nidiacei non sia certa. In altri contesti le femmine potrebbero accoppiarsi con molti maschi non per ottenere le cure paterne, ma per ridurre le possibilità che la loro prole sia danneggiata o uccisa da questi maschi. Nei primati che vivono in gruppi con molti maschi è stato ipotizzato che le femmine si accoppierebbero con diversi maschi per diminuire il rischio di infanticidi.
Garanzia di fecondazione. - Le femmine copulano con più di un maschio per evitare la possibilità di non essere fecondate se si accoppiano con maschi sterili. Fino a ora non si hanno informazioni sufficienti sulla diffusione della sterilità maschile per valutare rigorosamente questa ipotesi, ma l'evidenza sperimentale ottenuta con diamanti mandarino (Taenyopigia guttata) vasectomizzati non fornisce prove per questa ipotesi (Birkhead, 1996). Non vi sono nemmeno prove che i maschi più attraenti avrebbero eiaculati migliori, cosa che implicherebbe benefici diretti sulla fecondazione per le femmine.
Competizione tra femmine. - È stato anche suggerito che le femmine potrebbero accoppiarsi con più maschi per monopolizzare l'accesso sessuale ai maschi (e ai loro spermi) e prevenire altre femmine dall'essere fecondate o dall'essere fecondate da maschi di alta qualità. Finora non è chiaro se si possa essere davvero evo Iuta questa forma di comportamento 'dispettoso'.
Benefici indiretti
Aumentata diversità genetica della prole. - In alcuni insetti eusociali, come la formica tagliafoglie (Acromyrmex versicolor), gli accoppiamenti poliandrici sono comuni. In questa specie le femmine sembrano avere il controllo totale sugli accoppiamenti, si accoppiano in media con tre maschi, e tutti i maschi sono padri di un numero simile di figli (Reichart e Wheeler, 1996). Negli insetti eusociali le regine producono prole geneticamente mista e ciò potrebbe risolversi in una più marcata divisione del lavoro, una ripartizione dei sessi più in accordo con gli interessi della regina, un'aumentata resistenza contro patogeni e parassiti e costi ridotti di produzione di maschi diploidi.
Genetic bet-hedging. - Le femmine si accoppiano con molti maschi per ridurre gli effetti negativi dell'accoppiamento occasionale con maschi di bassa qualità. È stato suggerito (Watson, 1991) che il comportamento di ricerca del partner nei ragni femmina della specie Linyphia litigiosa si potrebbe spiegare con il genetic bet-hedging ('cautela' genetica). Il primo partner sessuale della femmina è in genere un maschio che ha scacciato altri maschi. Così le femmine utilizzano la competizione tra maschi come filtro principale per i loro primi partner sessuali. Al contrario, gli accoppiamenti successivi sono apparentemente il frutto di una scelta femminile e non necessitano di lotte fra altri maschi. Le femmine potrebbero riaccoppiarsi per compensare l'incertezza di essersi accoppiate con un maschio di bassa qualità. Dato che vi è precedenza per lo sperma del primo maschio, è probabile che esso, avendo già superato gli ostacoli della competizione intrasessuale, divenga il padre genetico di gran parte della prole.
Aumento della qualità genetica della prole. - Le femmine potrebbero inizialmente accettare maschi geneticamente inferiori sia se la ricerca di maschi comporta costi sia per evitare il rischio di fallire nella riproduzione. Tuttavia, se in seguito vi è l'opportunità di accoppiarsi con maschi geneticamente migliori, le femmine potrebbero riaccoppiarsi. Nelle specie con legame di coppia, come molti uccelli, non tutte le femmine hanno l'opportunità di formare coppie con maschi di alta qualità. Numerosi studi condotti sugli uccelli hanno mostrato che le femmine accoppiate con maschi di bassa qualità cercano più frequentemente accoppiamenti extracoppia, e che nella gran parte dei casi questi accoppiamenti avvengono con maschi di qualità migliore del loro partner. Ovviamente le femmine ottengono da questi accoppiamenti solo sperma e dunque è chiaro che le femmine agendo in tal modo ottengono benefici genetici. Uno studio recente ha mostrato piuttosto chiaramente che le cose stanno proprio così. Le femmine di cannareccione (Acrocephalus arundinaceus) cercano di avere copulazioni extracoppia con maschi che possiedono un repertorio canoro migliore dei loro partner ed è stata osservata una correlazione positiva tra l'ampiezza del repertorio canoro del maschio e la sopravvivenza della prole genetica dei suddetti maschi dopo l'involo (Hasselquist et al., 1996). Non è ancora chiaro perché un vasto repertorio canoro sia un buon indicatore di geni maschili in grado di produrre una prole con una sopravvivenza migliore. Una possibilità è che i maschi anziani, possedendo vasti repertori canori e utilizzando il canto come segnale, permettano alle femmine di scegliere maschi la cui capacità di sopravvivenza è provata dalla loro età.
Le femmine di lucertola non sarebbero in grado di rilevare la qualità genetica del maschio dal suo fenotipo. Con questa limitazione le femmine potrebbero accoppiarsi con molti maschi per aumentare le opportunità di essere fecondate da maschi geneticamente superiori. Questa ipotesi assume che i maschi geneticamente superiori abbiano prestazioni migliori sotto la pressione della competizione spermatica. In alcune popolazioni di vipere e lucertole europee il numero dei partner sessuali delle femmine è correlato alla sopravvivenza della prole; tuttavia in altre popolazioni tale correlazione non è stata osservata. Dunque non sono chiari né le relazioni né i meccanismi alla base di tali comportamenti.
Nelle popolazioni in cui la possibilità di inincrocio è elevata le femmine possono evitarne gli effetti deleteri sulla prole con accoppiamenti multipli. Nei maschi, elevati livelli di inincrocio sono correlati a scarsa qualità degli eiaculati. Così, quando le femmine si accoppiano con molti maschi, quello col più basso livello di inincrocio potrebbe essere il maschio che con più probabilità feconda le uova. Se così fosse, la prole sarebbe meno inincrociata e più vitale e i figli maschi avrebbero eiaculati più competitivi. Questo è uno dei casi in cui la selezione tramite competizione spermatica può essere più efficiente della selezione a livello fenotipico. Quest'ultima, infatti, potrebbe non mostrare così chiaramente gli effetti dell'inincrocio. La competizione spermatica può anche essere un modo per evitare l'inincrocio in altri contesti. Per alcuni mammiferi, che vivono in gruppi familiari ristretti, è stato suggerito che le femmine cerchino di accoppiarsi con maschi extragruppo (o extracoppia) per evitare l'inincrocio. Queste femmine possono anche essere limitate nella scelta del partner. Quando le femmine vivono in gruppi sociali ristretti gli accoppiamenti con individui estranei costituiscono probabilmente il modo migliore per ridurre gli effetti dell'inincrocio sulla prole, mentre evitare di accoppiarsi con maschi del gruppo probabilmente comporta dei costi.
Vantaggio nella sollecitazione della competizione spermatica. - È stato anche suggerito che le femmine trarrebbero benefici dal promuovere la competizione spermatica se, consentendo ai maschi che producono gli eiaculati più competitivi la fecondazione, potessero trasmettere tali caratteri ai loro figli. Questa ipotesi può spiegare quanto le femmine traggano vantaggio dall'essere fecondate da maschi con eiaculati competitivi in un contesto di competizione spermatica. Tuttavia, essa non spiega perché l'accoppiamento con molti maschi si sia evoluto per primo. Alcuni modelli hanno mostrato che gli accoppiamenti multipli possono diffondersi rapidamente a condizione che vi sia una piccola frazione di femmine che copulano in tal modo dall'origine (sia perché ci sono vantaggi per le femmine, sia perché le copulazioni sono forzate dai maschi) e vi siano differenze ereditabili tra maschi in uno o in molti dei seguenti caratteri: quantità di spermi; successo dello sperma nella fecondazione; abilità di rimuovere lo sperma di copulazioni precedenti; abilità di prevenire ulteriori copulazioni. In questa situazione i figli erediteranno un'elevata efficienza di fecondazione e tale carattere si diffonderà. Tuttavia, l'evidenza empirica diretta a favore di queste ipotesi è ancora carente.
Il conflitto tra i sessi
l maschi e le femmine utilizzano differenti modalità per rendere massimo il loro successo riproduttivo. Talora ciò che è vantaggioso per un sesso risulta svantaggioso per il successo riproduttivo dell'altro. Perciò ciascun sesso proverà a manipolare l'altro nel tentativo di rendere massimo il proprio successo riproduttivo, mentre l'altro sesso proverà a resistere a tale manipolazione. Questa situazione può portare a una gara evolutiva tra sessi con interessanti implicazioni.
Il conflitto tra i sessi può avvenire in una varietà di forme. Si può avere conflitto sull'accoppiarsi o meno, sulla frequenza delle copulazioni, sull 'utilizzo di sperma di maschi differenti per la fecondazione, su esistenza e durata del comportamento di controllo della compagna, su periodi degli accoppiamenti e sul contenuto dell'eiaculato. Iniziamo a esaminare alcuni casi di possibile conflitto riguardante la presenza o meno di competizione spermatica.
Il primo caso è quello in cui è nell'interesse della femmina essere selettiva sul suo (o sui suoi) partner sessuale o impegnarsi in pochi accoppiamenti mentre i maschi provano ad aumentare il loro successo riproduttivo accoppiandosi con più femmine possibile. l gerridi forniscono un esempio ben studiato (Rowe et al., 1994). Le femmine immagazzinano lo sperma per almeno 10 giorni senza ottenere benefici nutritivi dall'accoppiamento. La strategia ottimale per le femmine sarebbe quella di accoppiarsi solo poche volte al fine di ottenere tutto lo sperma di cui hanno bisogno per ottimizzare al meglio il loro potenziale di fertilità. Ulteriori accoppiamenti non sarebbero vantaggiosi per le femmine in quanto accoppiarsi comporta costi. Infatti, quando le femmine si accoppiano devono trasportare il maschio e ciò rallenta i movimenti e riduce l'efficienza nel foraggiamento. Inoltre, durante la copula, le femmine sono più visibili e meno capaci di sfuggire ai predatori. D'altra parte i meccanismi di competizione spermatica sono tali che l'ultimo maschio che si è accoppiato ha più possibilità di diventare il padre della prole. Così per un maschio ogni nuova femmina incontrata rappresenta una buona opportunità di produrre più prole. Questi interessi divergenti portano a strategie conflittuali: i maschi tenteranno di accoppiarsi con le femmine che incontrano, mentre le femmine che si sono già accoppiate saranno riluttanti ad accoppiarsi. Tuttavia, la persistenza del maschio impone alla femmina anche costi che potrebbero far diminuire la sua riluttanza. Per evitare la copula le femmine devono lottare per togliersi di tomo i maschi e ciò oltre a essere energeticamente costoso aumenta il rischio di predazione; inoltre vi sono costi dovuti alle molestie maschili, come discusso precedentemente. Molti fattori influiscono sui benefici e i costi di ciascuna alternativa per maschi e femmine: un rapporto deviato a favore dei maschi aumenta il tasso di molestie facendo diminuire la riluttanza femminile e aumentare le frequenze di accoppiamento. D'altra parte elevati rischi di predazione causano una riduzione nel tasso di molestie e un aumento della riluttanza delle femmine facendo diminuire la frequenza di accoppiamento.
In altri contesti i maschi non preferiti tentano copulazioni forzate e possono sviluppare strutture morfologiche che rendono difficile per le femmine evitare gli accoppiamenti. Nei grilli della specie Cyphoderris strepitans la femmina si alimenta delle ali posteriori polpose del maschio e delle secrezioni corporee che colano dalle ferite durante l'accoppiamento. Il maschio permette questo comportamento in quanto mantiene la femmina occupata per il tempo necessario a completare il trasferimento delle spermatofore. Quando i maschi si accoppiano ripetutamente le loro ali posteriori finiscono per essere quasi completamente mangiate e le nuove femmine tendono a evitare questi maschi. l maschi con ali posteriori ridotte utilizzano una struttura morfologica detta gin trap (trappola) per intrappolare le femmine riluttanti e forzare le copulazioni (Sakaluk et al., 1995). In certe specie, per contrastare questa strategia maschile, le femmine hanno evoluto strutture morfologiche che aumentano il controllo femminile della copulazione, come le spine addominali dei gerridi.
Il secondo caso di conflitto è quello in cui è nell'interesse della femmina accoppiarsi con molti maschi e questi tentano di prevenire che ciò avvenga. Questo è chiaramente il caso delle specie con legame di coppia, come molti uccelli, in cui le femmine accoppiate con maschi di bassa qualità traggono benefici dall' accoppiarsi con maschi di qualità migliore. l maschi tenteranno di controllare le loro femmine e le femmine tenteranno di ingannare i maschi. Quando le femmine si basano sull' aiuto del maschio per allevare la prole, i costi di essere scoperte in attività sessuali extracoppia saranno particolarmente elevati dato che i maschi potrebbero rispondere a una diminuzione della certezza di paternità riducendo la quantità di cure parentali da essi fornite. Tuttavia, quando le femmine non hanno bisogno dell' assistenza del maschio per allevare la prole esprimono una scelta del partner senza limitazioni e compiono più spesso copulazioni extracoppia. Ciò avviene in una specie con allevamento in comune, lo scricciolo azzurro (Malurus cyaneus), in cui ci sono aiutanti (helpers) che cooperano nell'alimentazione e nella difesa della prole. Quando le femmine nidificano in coppie permettono ai loro partner di ottenere una maggiore condivisione di paternità per assicurarsi la loro assistenza nell'allevamento dei nidiacei. Tuttavia, quando le femmine nidificano in gruppi in cui altri aiutanti compensano le riduzioni nelle cure parentali da parte dei maschi dominanti, le femmine producono nidiate contenenti più nidiacei con padri genetici esterni al gruppo (Mulder et al., 1994).
In specie che vivono in gruppi dominati da un maschio, il padrone dell'harem reagirà aggressivamente a qualsiasi tentativo di una sua femmina di avvicinarsi o accoppiarsi con maschi non appartenenti al gruppo. Nelle amadriadi (scimmie appartenenti alla famiglia dei cercopitecidi) le tattiche di controllo dei maschi includono un tipo di raggruppamento aggressivo in cui i maschi mordono vigorosamente le femmine sul collo. Più specificamente, quando il rischio di competizione spermatica è elevato i maschi possono mostrare adattamenti per prevenire o avere successo nella competizione spermatica, e ciò può essere dannoso per le femmine. l maschi possono tentare di controllare le femmine, cosa che potrebbe non solo limitare la loro scelta del partner, ma anche diminuire la loro efficienza di foraggiamento e aumentare la loro possibilità di essere predate. Inoltre, possono tentare di rimuovere gli spermi di partner precedenti dall'apparato femminile, potrebbero tentare di accoppiarsi frequentemente o impegnarsi in copulazioni prolungate al fine di sostituire o diluire i precedenti eiaculati. Tali comportamenti possono anche andare contro gli interessi delle femmine qualora queste preferissero maschi precedenti come padri della loro prole, e ciò anche a causa dei costi inerenti copulazioni prolungate o accoppiamenti ripetuti. Le femmine potrebbero anche incorrere in costi riguardanti la fertilità qualora i maschi rispondessero al rischio di competizione spermatica dedicando energie al controllo della compagna piuttosto che all'investimento nell'eiaculato. In Thalassoma bifasciatum i maschi territoriali difendono i siti di deposizione delle uova da maschi non territoriali più piccoli. l maschi territoriali con elevati potenziali riproduttivi sembrerebbero distribuire gli eiaculati tra femmine successive, ciò porta a un minor numero di spermatozoi per deposizione e a un tasso inferiore di fecondazione per le femmine (Warner et al., 1995). Non sembrano esserci limitazioni fisiologiche alla produzione di spermatozoi da parte di questi maschi, ma si instaura un conflitto tra produzione di gameti e controllo della compagna.
Un recente esperimento effettuato su Drosophila illustra in modo evidente quanto i sessi siano coinvolti in una 'corsa agli armamenti' e quanto, nel momento in cui uno dei sessi la rallenta, l'altro acquisisce rapidamente un vantaggio. In Drosophila melanogaster il fluido seminale riduce la competitività dello sperma di altri maschi, riduce il desiderio della femmina di riaccoppiarsi e aumenta il tasso di deposizione delle uova. Inoltre, esso esercita notevoli effetti tossici sulle femmine, diminuendone la sopravvivenza. In un esperimento originale, ai maschi furono concesse possibilità di evoluzione, mentre alle femmine non fu permesso di sviluppare meccanismi di 'ritorsione'. Dopo 41 generazioni i maschi che si adattavano erano migliori dei maschi di controllo: producevano più progenie maschile, ottenevano più accoppiamenti con femmine già accoppiate ed erano più capaci di prevenire che le femmine con cui si erano accoppiati producessero prole da maschi giunti successivamente. Tuttavia, questi adattamenti maschili condussero a una riduzione della sopravvivenza femminile, dovuta a un aumentato tasso di riaccoppiamento e un'aumentata tossicità del fluido seminale (Rice, 1996).
È spesso difficile determinare quale dei due sessi vince il conflitto o ottiene un vantaggio. Si è a lungo ritenuto che l'intensità delle forze selettive operanti sul maschio per rendere massimo il numero di prole prodotta fosse maggiore di quelle operanti sulla femmina per favorire certi maschi con lo scopo di migliorare la qualità della prole (Parker, 1984). Così si è assunto che siano gli adattamenti del maschio a prevalere. Tuttavia, nelle specie a fecondazione interna, indipendentemente dalle strategie comportamentali e fisiologiche seguite dal maschio, l'eiaculato perviene infine nell'apparato femminile, lontano da qualsiasi controllo maschile. Ciò concede alle femmine la possibilità di sviluppare, con costi minimi, meccanismi fisiologici di controllo della paternità durante questi stadi finali. Se così fosse, l'intensa selezione sui maschi per ottenere il massimo numero di fecondazioni potrebbe essere contrastata dai bassi costi del controllo femminile e si potrebbe giungere a un compromesso. Per esempio, è stato suggerito che l'ostilità dell'apparato riproduttore femminile potrebbe costituire un mezzo per scegliere gli eiaculati più competitivi e che alcune complesse interazioni tra spermi e apparato femminile potrebbero rendere le femmine in grado di favorire gli eiaculati di certi maschi (Roldan et al., 1992). Questa interessante possibilità non è stata finora valutata e sono ancora controverse le evidenze che suggeriscono un controllo femminile della paternità a livello fisiologico. Soltanto in pochi studi è stato suggerito che il fatto che i maschi più grandi ottengano maggiori quote di paternità in esperimenti di accoppiamenti controllati indicherebbe una selezione, da parte delle femmine, a favore degli spermi dei maschi di dimensioni maggiori. Un'interpretazione alternativa è che i maschi più grandi avrebbero la meglio nella competizione spermatica in quanto produrrebbero eiaculati più competitivi e non perché le femmine li favorirebbero. Sono necessari ancora molti studi per chiarire questi punti.
Conseguenze della competizione spermatica
l sistemi riproduttivi sono il prodotto delle strategie riproduttive di tutti gli individui che vivono in una popolazione. Come già illustrato, vi sono conflitti di interessi tra maschio e femmina, tra maschi in competizione e anche tra femmine in competizione.
Tradizionalmente i sistemi riproduttivi sono stati considerati il risultato delle tattiche di acquisizione del partner da parte del maschio, mirate all'accoppiamento con più femmine possibile. In questo contesto si è assunto che il successo riproduttivo maschile fosse correlato al numero di femmine con cui un maschio si accoppia. Gli studi sulla competizione spermatica hanno rivelato che questa è solo una parte della storia: nelle specie poliandriche gli spermi di maschi rivali sono in competizione all'interno dell'apparato riproduttore femminile e le femmine potrebbero esercitare qualche tipo di scelta anche a questo livello. Questo nuovo scenario comporta molte implicazioni. In primo luogo, il successo riproduttivo di un maschio non può essere identificato nel suo successo di accoppiamento. In secondo luogo, le strategie riproduttive del maschio e della femmina sono anche modulate in un certo grado dai meccanismi di competizione spermatica, che alla fine determineranno la paternità. Nelle specie monoandriche alcuni aspetti delle strategie maschili di accoppiamento possono essere compresi meglio se visti nell' ottica del tentativo di evitare la competizione spermatica durante il periodo in cui gli eiaculati sono più vulnerabili.
In tale contesto le strategie maschili di accoppiamento possono essere considerate come una commistione di tattiche di acquisizione del partner e tattiche di controllo della compagna. Dunque, in caso di competizione spermatica, vi è un conflitto tra numero di femmine accoppiate e impegno nel proteggere gli eiaculati. Ciò implica che, oltre ai fattori tradizionali che si ritiene influenzino le tipologie di accoppiamento (cioè distribuzione delle femmine nello spazio e nel tempo), vi sono altri fattori importanti correlati alle modalità di azione della competizione spermatica. Gomendio e Roldan (1993b) hanno suggerito che le differenze nei meccanismi di competizione spermatica tra uccelli e mammiferi potrebbero aver influenzato in modo determinante l'evoluzione di differenti sistemi riproduttivi.
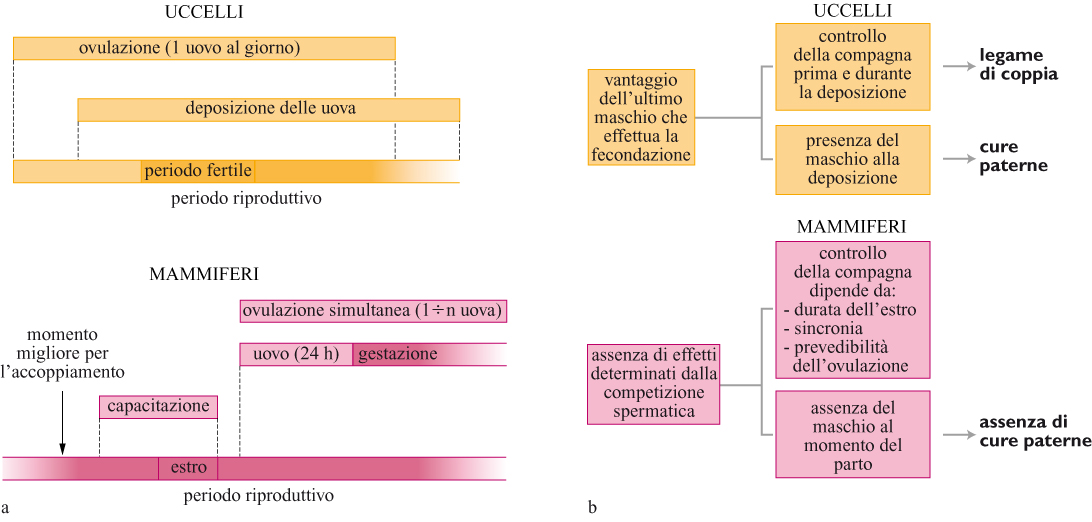
Negli uccelli la femmina ovula, feconda e depone le uova in una sequenza cronologica che comprende un certo numero di giorni (un tasso di produzione di circa un uovo al giorno) e può immagazzinare sperma prima e durante la deposizione nei tubuli di immagazzinamento degli spermi (fig. 4). l meccanismi di competizione spermatica determinano un vantaggio per l'ultimo maschio. La femmina accetta le copulazioni dal suo partner alcuni giorni prima che il primo uovo sia deposto fino all'inizio della deposizione assicurando al partner maschile un grado di certezza di paternità che garantirà la sua cooperazione. In questo modo la femmina assicura anche che sia immagazzinato sperma a sufficienza per fecondare tutte le uova. Le femmine necessitano l'assistenza dei maschi per allevare la prole e traggono vantaggio dall'aiuto del partner, ma anche dal produrre prole di maschi di alta qualità. Così, la femmina cessa di accettare accoppiamenti quando inizia la deposizione delle uova per rendere massime le opportunità di avere, nel caso trovasse un maschio migliore, una copulazione extracoppia con esito favorevole.
Dalla prospettiva del partner maschile le cose appaiono molto diverse. Un maschio beneficerebbe dal continuare ad accoppiarsi durante la deposizione delle uova al fine di rendere massime le sue possibilità di avere prole genetica. La femmina non gli permette di fado, tuttavia, esso potrebbe effettuare copulazioni 'di vendetta' se si accorgesse di una copulazione extracoppia da parte della femmina. Le potenziali copulazioni extracoppia sono molto rischiose per i partner maschili a causa del vantaggio fecondativo dell'ultimo maschio. Così i maschi devono controllare la compagna per tutto il periodo della sequenza di deposizione (cioè giorni) per avere una paternità certa. l vantaggi del controllo della compagna hanno probabilmente contribuito all'evoluzione del legame di coppia così comune negli uccelli.
Infine, andrebbe considerato il punto di vista dei potenziali partner extracoppia. l maschi probabilmente non si impegnano in copulazioni extracoppia durante il periodo della deposizione in quanto danno la priorità al controllo della compagna. Tale imposizione al riposo sessuale implica che i partner maschili possiedono eiaculati potenzialmente molto competitivi verso la fine della deposizione. Ciò, insieme al vantaggio fecondativo dell'ultimo maschio, significa che è relativamente facile per i maschi ottenere prole extracoppia dal momento che per loro è sufficiente accoppiarsi solo una volta nel momento giusto per avere la meglio sul partner della femmina. Gli elevati benefici associati alle copulazioni extracoppia, sia per i maschi sia per le femmine, hanno probabilmente favorito l' evoluzione di tale comportamento negli uccelli.
Nei mammiferi lo scenario è radicalmente diverso. Le femmine sono fertili per periodi molto limitati di tempo perché tutte le uova vengono prodotte nello stesso momento rimanendo vitali per circa 24 ore e le femmine non possiedono organi specializzati di immagazzinamento degli spermi (v. figura 4). Di conseguenza gli spermi dei mammiferi hanno vita breve. l meccanismi di competizione spermatica sono tali che non vi è un effetto nella sequenza di partner successivi. Gran parte della paternità è piuttosto determinata dal momento dell' accoppiamento rispetto all'ovulazione, dal numero di spermatozoi (sia in uno che in diversi eiaculati) e dalla qualità. Per i maschi la migliore strategia è dunque accoppiarsi in prossimità dell' ovulazione, lasciando tuttavia il tempo per il processo della capacitazione. Rispetto agli uccelli i maschi dei mammiferi hanno meno bisogno di difendere le femmine dopo l'ovulazione, e poiché queste ultime vanno incontro a gestazione e lattazione i maschi non traggono vantaggi dal rimanere con loro. Dato che è la femmina a occuparsi della prole, per i maschi vi è poca opportunità di migliorame la sopravvivenza e conviene loro assai più accoppiarsi con altre femmine. Considerando questi fattori è comprensibile come la poliginia sia il sistema nuziale più comune nei mammiferi.
Tra i mammiferi, gran parte della variabilità nel comportamento sessuale maschile è stata attribuita a quattro caratteristiche delle femmine: il grado a cui il tasso riproduttivo femminile può essere aumentato dall' assistenza maschile nell'allevamento della prole; la dimensione dell'area familiare della femmina e del suo centro (core area); la dimensione e la stabilità dei gruppi femminili; la densità e la distribuzione delle femmine nello spazio. È probabile che alcuni aspetti della fisiologia riproduttiva femminile uniti a meccanismi di competizione spermatica determinino in maniera decisiva le strategie riproduttive. Questi fattori comprendono la lunghezza del ciclo estrale, l'ovulazione spontanea o indotta, la prevedibilità dell'ovulazione e il grado della sincronia femminile nella recettività sessuale.
Nelle specie a ovulazione indotta è il primo maschio che si accoppia con la femmina a indurre l'ovulazione. Sembra probabile che i maschi abbiano evoluto periodi di fertilità degli spermatozoi (normalmente l'ovulazione si verifica poche ore dopo la copulazione) e tempi di capacitazione che sincronizzano questi eventi in modo che lo sperma del primo maschio sarà pronto a effettuare la fecondazione nel momento in cui si verifica l'ovulazione. Se così fosse, allora il primo maschio potrebbe avere un vantaggio rispetto ai maschi che si accoppiano più tardi nel ciclo femminile. Se la sincronia fosse così perfetta da consentire scarsissime possibilità di fecondazione agli eventuali maschi che si accoppiassero in seguito, allora il maschio che si accoppia per primo non dovrebbe controllare la femmina. Tuttavia, se gli eventuali maschi che si accoppiano successivamente avessero possibilità di fecondazione, allora il primo maschio dovrebbe controllare la compagna fino a ovulazione avvenuta per impedime la fecondazione da parte di altri maschi. Dato che l'ovulazione tende a verificarsi poche ore dopo la copulazione, il periodo di controllo della compagna dovrebbe essere comunque di breve durata. Gli scoiattoli terricoli della specie Spermophilus tridecemlineatus hanno un'ovulazione indotta 8 ÷ 36 ore dopo la copulazione; il primo maschio ad accoppiarsi è il padre di gran parte della prole e i maschi si accoppiano molte volte con una femmina, ma non controllano la compagna (Schwagmeyer, 1995). Ciò suggerisce che il primo modello proposto sia quello valido. In questa specie i maschi lasciano la propria partner dopo che hanno avuto una copulazione più lunga di 9 minuti. È possibile che una copulazione così prolungata sia necessaria per indurre l'ovulazione. Studi di laboratorio hanno mostrato che, come atteso, i ritardi tra le copulazioni di due maschi successivi favoriscono il primo. In accordo con queste osservazioni, i dati ottenuti studiando questa specie in natura hanno mostrato che i maschi che trovano una femmina in estro che si è accoppiata da più di quattro ore, tendono a non accoppiarsi con essa. Infatti quando il ritardo supera le quattro ore, il secondo maschio non è più favorito nell' accoppiarsi con la femmina perché le sue possibilità di fecondarla sono molto scarse. Il vantaggio del primo maschio è stato anche osservato nei conigli, anch'essi specie a ovulazione indotta.
Per le specie a ovulazione spontanea occorre considerare la lunghezza dell'estro. Nelle specie in cui l'estro è breve gran parte dei maschi che si accoppia all'interno del periodo estrale avrà possibilità di fecondare le uova. Se la recettività sessuale delle femmine è molto sincronizzata e queste sono spazialmente vicine, i maschi hanno un gran numero di opportunità di accoppiarsi con altre femmine mentre potrebbero essere incapaci di difendere un grande gruppo di femmine. In questo caso non si verifica il controllo della compagna. Nello scoiattolo della specie Spermophilus beldingi le femmine recettive si riuniscono in gruppi e i maschi non controllano le femmine presumibilmente perché le femmine possono essere facilmente localizzate e i maschi possono ottenere un maggior successo andando a cercare altre femmine in estro dopo la copulazione (Sherman, 1989).
Se invece la recettività sessuale femminile non è sincronizzata o le femmine sono spazialmente disperse, i maschi potrebbero essere in grado di difendere una femmina in estro per volta e sarebbero certamente incapaci di cercarne altre in estro. Dunque, si dovrebbero osservare brevi periodi di controllo della compagna dato che non vi sarebbe alcuna perdita di opportunità di accoppiamento. Ciò sembra verificarsi in un'altra specie di scoiattolo (Spermophilus brunneus): le femmine vanno in estro per un pomeriggio all'anno, i maschi le controllano per circa tre ore dopo la copulazione e in tal modo migliorano le proprie opportunità di esser padri della prole (Sherman, 1989).
Nelle specie in cui l'estro è prolungato solo i maschi che si accoppiano intorno al periodo dell' ovulazione avranno possibilità di fecondare le uova. Nel caso in cui le femmine segnalino l'ovulazione i maschi dovrebbero tentare di accoppiarsi intorno a tale periodo. Nei babbuini, i maschi competono per le copulazioni quando le protuberanze sessuali raggiungono le massime dimensioni (segnalando l'ovulazione nel giro di uno o due giorni). Al di fuori di questo periodo i maschi dominanti tendono a ignorare le femmine e quelli subordinati o i giovani si avvantaggiano dell'opportunità di accoppiarsi con le femmine. Se vi sia o meno controllo della compagna dipende dalla eventuale perdita di opportunità di accoppiarsi con altre femmine e così vale per il grado di sincronia femminile. Per questa ragione le associazioni di coppie sono più ricorrenti nei babbuini (riproduttori non stagionali) che nei macachi (riproduttori stagionali).
Nel caso in cui le femmine non segnalino l'ovulazione i maschi sono incapaci di prevedere quando avverrà l'ovulazione e così sono incapaci di valutare il valore delle copulazioni in differenti momenti della durata del ciclo. Mentre le femmine sono sempre sincrone in un certo grado perché sono recettive per gran parte del tempo, i maschi non sono in grado di localizzare il momento dell' ovulazione. Questa situazione favorisce accoppiamenti a brevi intervalli per lunghi periodi di tempo (per assicurare la fecondazione) e, potenzialmente, prolungati controlli della compagna. E questo sembra essere il caso della specie umana.
Bibliografia citata
BIRKHEAD, T.R. (1996) Sperm competition: evolution and mechanisms. In Current topics in developmental biology, voI. 33, Londra, Academic Press, pp. 103-158.
BIRKHEAD, T.R., M0LLER, A.P. (1992) Sperm competition in birds. Londra, Academic Press.
BISSONDATH, C.J., WIKLUND, C. (1995) Protein content of spermatophores in relation to monandry/polyandry in butterflies. Behav. Ecol. Sociobiol., 37, 365-371.
DAVIES, N.B., HARTLEY, I.R., HATCHWELL, B.J., LANGMORE, N.E. (1996) Female control of copulations to maximize male help: a comparison of polygynandrous alpine accentors, Prunella collaris, and dunnocks, P. modularis. Anim. Behav., 51, 27-47.
GAGE, M.J.G., STOCKLEY, P., PARKER, G.A. (1995) Effects of alternative male mating strategies on characteristics of sperm production in the Atlantic salmon (Salmo salar): theoretical and empirical investigations. Phil. Trans. R. Soc. Lond. B, 350, 391-399.
GOMENDIO, M., HARCOURT, A.H., ROLDAN, E.R.S. (1998) Sperm competition in mammals. In Sperm competition and sexual selection, a c. di Birkhead T.R., Möller A.P., Londra, Academic Press, pp. 667-751.
GOMENDIO, M., ROLDAN, E.R.S. (1991) Sperm competition influences sperm size in mammals. Proc. R. Soc. Lond. B. Biol. Sci., 243, 181-185.
GOMENDIO, M., ROLDAN, E.R.S. (1993a) Co-evolution between male ejaculates and female reproductive biology in eutherian mammals. Proc. R. Soc. Lond. B., 252, 7-12.
GOMENDIO, M., ROLDAN, E.R.S. (1993b) Mechanisms of sperm competition: linking physiology and behavioural ecology. Trends in Ecology and Evolution, 8,95-100.
HASSELQUlST, D., BENSCH, S., VON SCHANTZ, T. (1996) Correlation between male song repertoire, extra-pair paternity and offspring survival in the great reed warbler. Nature, 381, 229-232.
MØLLER, A.P. (1988) Ejaculate quality, testes size and sperm competition in primates. J. Rum. Evol., 17, 479-488.
MULDER, R.A., DUNN, P.O., COCKBURN, A., LAZENBy-COHEN, K.A., HOWELL, M.J. (1994) Helpers liberate female fairywrens from constraints on extra-pair mate choice. Proc. R. Soc. Lond. B, 255, 223-229.
PARKER, G.A. (1970) Sperm competition and its evolutionary consequences in insects. Biol. Rev. Camb. Philos. Soc., 45, 525-567.
PARKER, G.A. (1984) Sperm competition and the evolution of animaI mating strategies. In Sperm competition and the evolution of animaI mating systems, a c. di Smith R.L., Orlando, Academic Press, pp. 1-60.
PARKER, G.A. (1990a) Sperm competition games: guards and extra-pair copulations. Proc. R. Soc. Lond. B, 242, 127-133.
PARKER, G.A. (1990b) Sperm competition games: raffles and roles. Proc. R. Soc. Lond. B., 242, 120-126.
REICHART, A.K., WHEELER, D.E. (1996) Multiple mating in the ant Acromyrmex versicolor: a case of female control. Behav. Ecol!, Sociobiol., 38, 219-225.
RICE, W.R. (1996) Sexually antagonistic male adaptation triggered by experimental arrest of female evolution. Nature, 381, 232-234.
ROLDAN, E.R.S., GOMENDIO, M., VITULLO, A.D. (1992) The evolution of eutherian spermatozoa and underlying selective forces: female selection and sperm competition. Biol. Rev. Camb. Philos. Soc., 67, 551-593.
ROWE, L., ARNQVIST, G., SIH, A., KRUPA, J. (1994) Sexual conflict and the evolutionary ecology of mating patterns: water striders as amodel system. Trends in Ecology and Evolution, 9, 289-293.
SAKALUK, S.K., BANGERT, P.J., EGGERT, A.-K., GACK, C., SWANSON, L.V. (1995) The gin trap as a device facilitating coercive mating in sagebrush crickets. Proc. R. Soc. Lond. B, 261, 6571.
SCHWAGMEYER, P.L. (1995) Searching today for tomorrow's mates. Anim. Behav., 50, 759-767.
SHERMAN, P.W. (1989) Mate guarding as paternity insurance in Idaho ground squirrels. Nature, 338, 418-420.
SMITH, R.L., a c. di (1984) Sperm competition and the evolution of animaI mating systems. San Diego, Academic Press.
STOCKLEY, P., SEARLE, J.B., MAcDONALD, D.W., JONES, C.S. (1994) Alternative reproductive tactics in male common shrews: relationships between male mate-searching behaviour, sperm production, and reproductive success as revealed by DNA fingerprinting. Behav. Ecol. Sociobiol., 34, 71-78.
WARNER, R.R., SHAPIRO, D.Y., MARCATO, A., PETERSEN, C.W. (1995) Sexual conflict: males with the highest mating success convey the lowest fertilization benefits to females. Proc. R. Soc. Lond. B, 262, 135-139.
WATSON, P.J. (1991) Multiple paternity as genetic bet-hedging in female sierra dome spiders, Linyphia litigiosa (Linyphiidae). Anim. Behav., 41, 343-360.