
Cooperazione
Cooperazione
C'è cooperazione quando due organismi della stessa specie condividono i benefici di un'azione svolta insieme. Possono costituire buoni esempi di comportamento cooperativo i rapporti tra i partner di una coppia in vista della riproduzione, i comportamenti collegati alla cura della prole, come pure le attività coordinate legate alla costruzione della tana. Tuttavia, la cooperazione manifesta il suo peso nell'evoluzione delle specie con l'emergenza dell'eusocialità. Dal punto di vista della teoria sociobiologica, una società è definibile come 'un gruppo di individui della stessa specie organizzati in modo cooperativo'. È interessante osservare che in certe società, come quelle delle vespe e di altri insetti sociali, a livello individuale si assiste a un comportamento apparentemente conflittuale che però diventa cooperativo a livello globale, per esempio nelle attività collettive legate alla costruzione del nido, alla ricerca del cibo, alla cura delle larve.
Presupposti della cooperazione sono due fattori, l'altruismo basato sulla parentela e il beneficio reciproco (in certi casi accompagnati dall'imposizione a cooperare), che in varia misura agiscono in tutti i tipi di società. La cooperazione intraspecifica di solito coinvolge i legami di parentela, ma vi sono anche casi di cooperazione tra estranei. Nell'altruismo reciproco, per esempio, il vantaggio per uno dei partner è immediato, mentre per l'altro è differito. Da qui l'insorgenza di un problema evolutivo, in quanto l'individuo beneficiato per primo si trova nella possibilità di 'imbrogliare' il partner, ottenendo così un vantaggio senza dare nulla in cambio. Analisi teoriche e studi dettagliati condotti in molti sistemi biologici suggeriscono che la parentela e la fedeltà al partner costituiscono i meccanismi più importanti per spiegare l'evoluzione della cooperazione negli animali. Inoltre, anche le pari opportunità riproduttive, la possibilità di scegliere il partner con cui cooperare e il controllo reciproco possono favorire la cooperazione.
Comportamenti di tipo cooperativo sono osservabili anche tra specie differenti; nelle simbiosi mutualistiche che talora sfociano nella coevoluzione i legami interspecifici sono infatti sostenuti da comportamenti di tipo simil-cooperativo. La cooperazione in senso stretto, di tipo intraspecifico, è collegabile al fenomeno dell'altruismo e alle questioni riguardanti l'esistenza di unità di selezione diverse dall'individuo. Lo studio dei fenomeni della cooperazione, oltre a favorire la comprensione delle origini della socialità, può contribuire a gettare luce sulla virulenza dei parassiti, sulle transizioni chiave nell'evoluzione della vita (dall'origine delle molecole autoreplicanti alla meiosi) e, infine, a chiarire numerosi comportamenti della nostra specie.
Cooperazione e fedeltà al partner
Reciprocità
La fedeltà al partner costituisce la base per la reciprocità della cooperazione. Robert L. Trivers, nel 1971, fu il primo a mettere in evidenza che fino a quando due individui si aiutano l'un l'altro ripetutamente sopravvivono e si riproducono più di due individui che non cooperano. Trivers chiamò questa forma di cooperazione 'altruismo reciproco', per cercare di spiegare che in questi casi la cooperazione avviene a dispetto di un costo immediato per la riproduzione o la sopravvivenza. Se i benefici superano i costi, ciascun partecipante trarrà nel tempo profitto dall'interazione. Tuttavia, gli individui che non restituiscono il beneficio trarranno un profitto ancora maggiore. Affinché la reciprocità rimanga vantaggiosa, gli individui che cooperano devono apprendere e ricordare le precedenti interazioni per identificare gli imbroglioni e applicare ritorsioni contro di loro. La condivisione del cibo nel vampiro vero o di Azara (Desmodus rotundus) illustra i potenziali vantaggi della reciprocità. Le femmine rigurgitano il sangue per darlo alla loro prole e ad altri membri della colonia che non siano riusciti a procurarselo. Ogni notte il 7% degli adulti e il 33% dei giovani non riesce ad alimentarsi, in quanto le prede spesso li riconoscono e li allontanano. I giovani, meno esperti, vengono più facilmente scoperti rispetto agli adulti. Esperimenti con individui in cattività indicano che i pipistrelli vampiri non solo identificano e alimentano di preferenza gli individui della stessa colonia, ma rigurgitano più facilmente il sangue a beneficio di quegli individui che in precedenza avevano offerto loro del cibo. Quest'aiuto può essere di enorme importanza, dato che un individuo che non riesce ad alimentarsi per tre notti consecutive è destinato a morire. In assenza della condivisione reciproca del sangue la mortalità annuale supererebbe l'80% mentre, di fatto, le femmine selvatiche possono vivere più di vent'anni.
Il dilemma del prigioniero
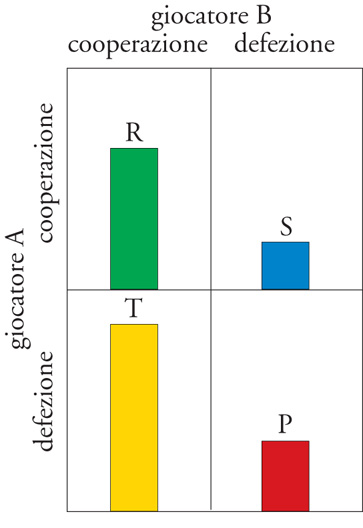
L'effetto della fedeltà al partner sull'evoluzione della cooperazione è stato esaminato attentamente da un punto di vista teorico. Gran parte di questa teoria si basa sull'analisi di un gioco ripetuto da due partecipanti che prende il nome di 'dilemma iterato del prigioniero'. Anziché soldi i giocatori biologici acquistano o perdono punti che riflettono differenze nel loro successo riproduttivo complessivo, o fitness (la 'moneta' dell'evoluzione). In una mano del gioco ciascun giocatore può ottenere un premio (punteggio R, reward) se entrambi cooperano, e una punizione (punteggio P, punishment) se entrambi non cooperano. La tentazione di non cooperare con un partner che coopera comporta un punteggio più alto (punteggio T, temptation), mentre cooperare con un partner che non coopera comporta il punteggio più basso (punteggio S, sucker). In termini algebrici il dilemma del prigioniero esiste se T>R>P>S e R>(T+S)/2 (fig. 4). Se il gioco consiste in una sola mano la migliore strategia è non cooperare, anche se i giocatori otterrebbero un punteggio migliore cooperando entrambi. Questo si verifica perché un giocatore che non coopera evita di ottenere S se l'altro non coopera, oppure ottiene T se l'altro coopera. Cioè per il giocatore A non cooperare è la miglior risposta a entrambe le possibili mosse del giocatore B in una singola mano del gioco.
Se si giocano più mani e i giocatori basano le loro decisioni sulle interazioni precedenti, può emergere la cooperazione. In una serie di simulazioni al computer R. Axelrod scoprì che la semplice strategia TFT (Tit for tat, 'occhio per occhio'), aveva un rendimento superiore a tutte le altre possibili. La strategia TFT incarna la regola aurea che consiste nell'iniziare sempre cooperando e nel copiare semplicemente la precedente decisione del partner nelle interazioni successive. Questa strategia ha successo se la probabilità che due individui si incontrino di nuovo è abbastanza forte; di conseguenza la fedeltà al partner consente di escludere gli imbroglioni.
Una teoria più recente indica che la strategia TFT potrebbe rappresentare solo il primo stadio evolutivo della cooperazione. Sebbene questa sia la migliore strategia in un ambiente uniforme, è soggetta a errori involontari. Una defezione accidentale tra due individui che adottano TFT porterà a cicli di mutua defezione finché un secondo errore non correggerà l'incomprensione. La strategia GTFT (Generous tit for tat, 'occhio per occhio generoso') prevede di rispondere a una defezione con un'altra defezione solo in due terzi dei casi: questa strategia perdona più di quella TFT e quindi è immune da questi errori occasionali.
La difesa territoriale nel leone (Panthera leo) è un'eccezione istruttiva a gran parte di queste teorie, perché alcuni individui sembrano attuare una cooperazione incondizionata anche in risposta a una defezione. I leoni maschi si riuniscono in coalizioni che competono tra loro per il controllo di un gruppo di femmine. Le piccole coalizioni sono composte di solito da individui non imparentati, mentre quelle più grandi sono caratterizzate dalla presenza di fratelli. La difesa territoriale è stata studiata diffondendo ruggiti con altoparlanti per simulare intrusioni da parte di una coalizione concorrente. Tutti i maschi di una coalizione che, in un certo momento, controlla un gruppo di femmine, si avvicinano agli altoparlanti indipendentemente dal comportamento o dal grado di parentela. Questo comportamento si spiega perché il successo riproduttivo di ognuno dipende dal mantenimento del controllo sul branco di femmine. L'importanza del dilemma del prigioniero per spiegare la cooperazione tra gli animali resta tuttavia controversa, perché per la maggior parte degli esempi esistono anche spiegazioni alternative, alcune delle quali non assegnano i punteggi secondo i criteri del dilemma del prigioniero.
Cooperazione e affinità genetica
Selezione di parentela
Essere imparentati vuol dire condividere geni. La riproduzione implica la duplicazione e la trasmissione di geni alla prole. La probabilità che una copia esatta di un gene si trovi in un altro individuo è chiamata grado di parentela ed è indicata con il simbolo r. Nella specie umana e negli altri organismi diploidi, cioè quelli che possiedono due copie dei geni su cromosomi appaiati, il grado di parentela della madre con il figlio è pari a 1/2, perché il figlio riceve metà dei suoi geni dalla madre e metà dal padre. Lo stesso vale per il grado di parentela di un figlio con la madre. Qualunque coppia di individui con un antenato comune è in qualche modo imparentata, sebbene r possa essere molto piccolo se l'antenato risale a molte generazioni precedenti. In ogni caso r è misurato come la probabilità che un gene tipico di un individuo abbia una copia esatta in un altro a causa della discendenza diretta da un antenato comune.
L'affinità genetica è importante, perché ingannare in un'interazione cooperativa potrebbe non essere vantaggioso se il partner è un parente. Va ricordato che la selezione naturale favorisce quei caratteri che aumentano il numero di figli prodotti da un individuo nell'arco della sua vita e la loro sopravvivenza. La ragione è che i genitori sono imparentati con i propri figli e condividono con loro una parte dei geni. Dal punto di vista di un gene, qualunque comportamento che aumenti la sua frequenza nella generazione successiva sarà favorito. Di conseguenza la selezione naturale dovrebbe anche favorire i caratteri che aumentano il numero di altri parenti, come i fratelli o i cugini, e la loro sopravvivenza. Tuttavia, dato che i fratelli hanno un grado di affinità genetica più elevato dei cugini, si prevede che gli animali siano in grado di discriminare e, di conseguenza, interagire di preferenza con i parenti più stretti.
Se due animali hanno un grado sufficiente di affinità genetica, anche il comportamento altruistico può essere favorito. Un gene che influenzi il comportamento altruistico si diffonderà quando i benefici per l'altruista (B) moltiplicati per il grado di parentela (r) sono maggiori del costo per l'individuo che dà l'avvio al comportamento (C); in altre parole, quando rB−C> 0. Questa semplice disequazione, nota come 'regola di Hamilton', è il fondamento di un'importante teoria denominata 'selezione di parentela'. La selezione di parentela influenza l'evoluzione del comportamento altruistico e di molte altre forme di comportamento sociale in una grande varietà di animali. Per illustrare in che modo la selezione di parentela influenzi il comportamento cooperativo, consideriamo la costruzione del nido e l'allevamento della prole nel topolino delle case (Mus domesticus). Spesso due o tre femmine di questa specie costruiscono il nido, partoriscono insieme e ciascuna accudisce indifferentemente tutti i piccoli presenti nel nido comune. Un vantaggio importante nel costruire un nido comune potrebbe consistere nel fatto che due o tre femmine proteggono meglio la prole dai maschi, potenzialmente infanticidi. Tuttavia, dato che l'allattamento in genere è il periodo energeticamente più dispendioso nella vita di ogni femmina di mammifero, per quale motivo le femmine di topo non evitano di accudire i piccoli nel nido comune? La risposta sembra essere che in genere ciascuna femmina è imparentata con tutti i piccoli. I topolini delle case, infatti, costruiscono i nidi comuni di preferenza con parenti stretti come sorelle e figlie. Le femmine che forniscono latte agli altri piccoli oltre che ai propri lasciano alle generazioni successive più copie dei propri geni rispetto alle femmine meno cooperative.
Riconoscimento dell'affinità genetica
Affinché la selezione di parentela possa funzionare, le femmine di topo devono essere capaci di riconoscere quelle più affini geneticamente e nidificare di preferenza insieme a loro. In altre parole, devono saper distinguere i parenti dai non parenti. In alcuni animali questo tipo di riconoscimento è basato su un'associazione verificatasi in precedenza. Per esempio, un odore comune acquisito da individui cresciuti insieme, cibandosi degli stessi alimenti, può impartire caratteristiche olfattive riconoscibili. Un odore specifico del nido può permettere a un animale di effettuare discriminazioni semplici, distinguendo tra fratelli e individui non imparentati, ma non è molto utile per prevedere l'affinità genetica negli individui di una nidiata che presentano diversi gradi di parentela, come accade se due o più maschi ne condividono la paternità. In questi casi sarebbe meglio se gli animali potessero in qualche modo confrontare direttamente i geni per determinare il grado di affinità.
Il riconoscimento dell'affinità genetica richiede la presenza di regioni variabili nel genoma e la capacità sensoriale di riconoscere e discriminare gli individui sulla base di queste differenze. Altrimenti, in presenza di geni identici, l'effetto di un antenato comune non può essere discriminato da quello dovuto al caso. Studi su topi congenici opportunamente incrociati al fine di risultare geneticamente identici per gli alleli nel complesso dei loci multipli di istocompatibilità, o MHC (Major histocompatibility complex), hanno dimostrato che le femmine di topo formano nidi comuni se condividono il medesimo aplotipo MHC. In tutti i Vertebrati il complesso MHC è caratterizzato da un'enorme variabilità genetica per facilitare il riconoscimento cellula-cellula, allo scopo di difendere l'organismo dalle invasioni di cellule parassite da parte di organismi portatori di malattie. Di conseguenza i loci MHC sono ideali come indicatori genetici del grado di parentela e potrebbero svolgere questa funzione in molti altri Vertebrati, uomo compreso.
Come nei topolini delle case, le leonesse africane che condividono un territorio accudiscono i piccoli in cooperazione. Una delle principali cause di mortalità dei cuccioli è l'infanticidio, che avviene in seguito all'avvicendamento tra coalizioni di maschi. Come nei topolini, il numero di femmine determina il grado di protezione dei piccoli dai maschi infanticidi. Ma i leoni discriminano più accuratamente rispetto ai topolini. Le femmine che allattano permettono ai cuccioli geneticamente più affini di prendere più latte rispetto a quelli meno affini. È ancora da chiarire in che modo le leonesse riconoscano i livelli di parentela con i piccoli, ma l'età, un'eventuale associazione precedente e il grado di somiglianza dei loci MHC sono tutti meccanismi plausibili.
Controllo riproduttivo in individui imparentati
Quando in una struttura sociale gli individui imparentati appartengono a diversi livelli di una scala gerarchica e hanno scarse opportunità di vivere al di fuori di un gruppo sociale, la cooperazione può avvenire anche se i partecipanti ne traggono benefici di entità differente. Un esempio è costituito dall'allevamento in cooperazione nell'ani beccosolcato (Crotophaga sulcirostris). Questi cuculi neri e lucenti vivono in gruppi sociali costituiti da una a quattro coppie di uccelli che condividono un nido comune. Le femmine depongono le loro uova secondo un ordine temporale: la più bassa nella gerarchia depone per prima, la dominante per ultima. Ciascuna femmina, prima di cominciare a deporre, rimuove alcune uova già presenti nel nido. Come conseguenza di questo comportamento di rimozione delle uova, la femmina dominante contribuisce alla covata finale più delle altre. Quando le uova si schiudono, tutti i membri del gruppo partecipano alla nutrizione e alla protezione dei pulcini. Perché le femmine subordinate non abbandonano il gruppo per nidificare per conto loro? La ragione sembra essere la generale mancanza di un habitat di nidificazione di alta qualità nelle zone dell'America Centrale dove vivono questi uccelli. Di conseguenza, una femmina subordinata produce più piccoli in un gruppo cooperativo di quanti ne potrebbe produrre da sola. Se poi è anche imparentata con la dominante, può ricevere un ulteriore beneficio incrementando le probabilità di sopravvivenza degli altri piccoli.
Il controllo riproduttivo in individui imparentati può avvenire anche nei Mammiferi come la mangusta nana meridionale (Helogale parvula), il licaone, il tamarino dai ciuffetti bianchi (Saguinus oedipus) e l'eterocefalo glabro (Heterocephalus glaber). In tutte queste specie, in genere, solo una singola femmina alleva i piccoli con successo. La riproduzione di altre femmine nel gruppo sociale è inibita dal rilascio di ferormoni specifici, oppure i loro piccoli sono uccisi alla nascita. Nonostante questo trattamento gli individui non riproduttori, che tipicamente sono imparentati con i dominanti, rimangono nel gruppo, raccolgono cibo e aiutano la coppia dominante nella cura della prole.
Meccanismi che favoriscono la cooperazione
Fino alla metà degli anni Sessanta del Novecento la cooperazione era in genere ritenuta un meccanismo di regolazione demografica della popolazione. Una critica dettagliata di George C. Williams, esposta in un volume del 1966, mise in evidenza i punti deboli di questa interpretazione. A meno che la migrazione tra popolazioni di conspecifici sia trascurabile e le estinzioni di queste popolazioni frequenti, gli individui che egoisticamente massimizzano la propria riproduzione escludono quelli che la limitano per ridurre la competizione. La principale eccezione a questa interpretazione si verifica quando gli animali sono imparentati tra loro. La scoperta che il controllo riproduttivo si riscontra tipicamente in animali che formano gruppi costituiti da parenti stretti è perciò compatibile con l'idea che sia la selezione di parentela, piuttosto che quella di gruppo, a favorire la cooperazione.
Il comportamento dell'ani beccosolcato potrebbe far supporre che tutti i casi di cooperazione nell'allevamento della prole debbano riguardare animali strettamente affini. Tuttavia, in diverse specie di formiche e termiti le colonie possono essere fondate da più femmine riproduttive imparentate solo lontanamente o per nulla. I dati a disposizione mostrano che le colonie fondate da più regine tendono a crescere più rapidamente e hanno una maggiore probabilità di sopravvivere e di produrre individui riproduttivi rispetto a quelle fondate da una sola femmina. Di conseguenza, la riproduzione pro capite è spesso più elevata in queste colonie con più regine, o poliginiche.
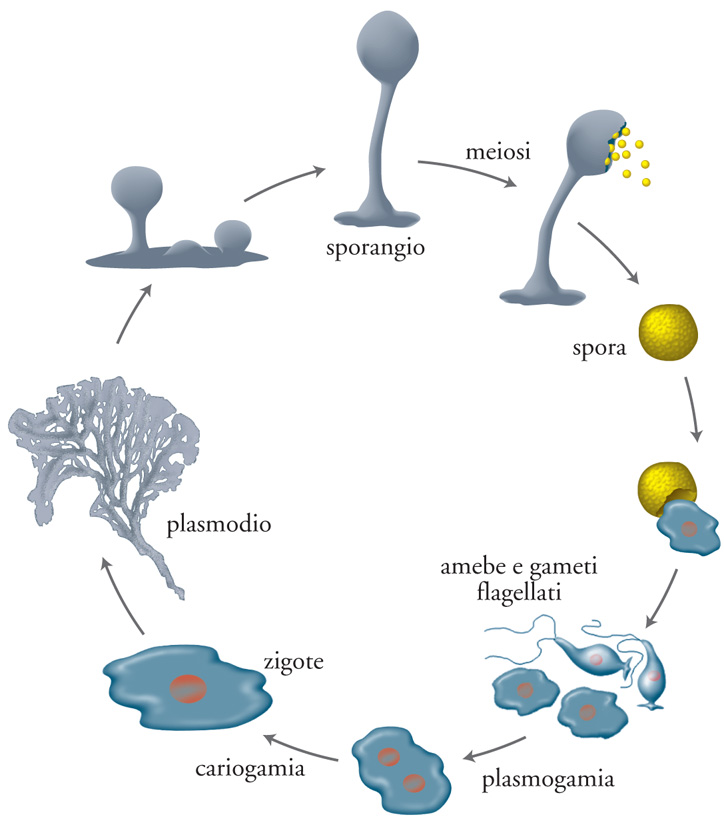
Un altro interessante esempio di cooperazione assicurata dalle pari opportunità riproduttive è quello del mixomicete Dictyostelium discoideum. Il ciclo vitale di questi organismi (fig. 7) passa da uno stadio a cellule libere e disperse a un aggregato multicellulare detto 'plasmodio', capace di compiere movimenti coordinati. Quando il plasmodio raggiunge un sito ecologicamente idoneo, produce un corpo fruttifero contenente spore in cima a uno stelo. Sebbene la selezione di parentela possa spiegare perché le cellule dello stelo si sacrifichino per le loro spore sorelle, bisogna sottolineare il fatto che alcuni plasmodi si formano da più di 100.000 cellule aggregate. Nel plasmodio in via di formazione la determinazione casuale del destino delle cellule può favorire la cooperazione senza l'intervento della selezione di parentela. La distribuzione spaziale casuale dei diversi tipi cellulari al momento di stabilire il destino delle cellule sostiene quest'ipotesi.
Le interazioni mutualistiche che caratterizzano un gran numero di specie forniscono buoni esempi di come un meccanismo alternativo al precedente permetta di limitare gli imbroglioni e di mantenere la cooperazione. Tale meccanismo si basa sulla scelta del partner con cui cooperare. Molte specie di Omotteri, come gli afidi e i Membracidi, le larve di molte farfalle Licenidi e Riodinidi e alcune piante, come le acacie delle formiche, forniscono alle formiche soluzioni ricche di zuccheri in cambio della protezione dai predatori o dai competitori. Le singole piante o gli insetti potrebbero imbrogliare non producendo il premio per le formiche. La cooperazione, perciò, dovrebbe persistere finché le formiche non riescono a trovare piante o insetti alternativi che forniscano premi più consistenti. Allo stesso tempo, insetti e piante potrebbero negare la ricompensa se le formiche assistenti non assicurassero un'adeguata protezione. Così la scelta del partner potrebbe regolare la cooperazione come avviene in qualunque economia di mercato.

L'importanza della scelta del partner è stata dimostrata in studi sulle associazioni mutualistiche tra alcuni bruchi e formiche. I bruchi dei Licenidi e dei Riodinidi secernono soluzioni zuccherine da una ghiandola dorsale, la cui unica funzione è quella di produrre una ricompensa per le formiche (fig. 8). Inoltre alcuni bruchi attraggono le formiche generando vibrazioni acustiche di richiamo e possono emettere segnali chimici che le stimolano ad attaccare i predatori in avvicinamento. Si è osservato che bruchi isolati producono una maggiore quantità di soluzione zuccherina quando il numero delle formiche dedite alle loro cure viene aumentato da uno a due. Nelle specie di Licenidi che formano grandi raggruppamenti spesso le formiche uccidono i singoli bruchi che non producono nettare. Mentre le pupe attraggono le formiche continuando a produrre la secrezione zuccherina, le farfalle che emergono dopo la schiusa sono soggette a predazione da parte delle formiche stesse, dimostrando ulteriormente che la cooperazione in questo sistema si basa sul valore di mercato del nettare.
La scelta selettiva del partner può anche rafforzare il comportamento mutualistico intraspecifico. Per esempio, in certi Primati e delfini i maschi cooperano formando alleanze per attrarre le femmine sessualmente ricettive. Le alleanze dei Primati sembrano però differire da quelle dei delfini per almeno due aspetti. Le prime tendono a essere temporanee e danno luogo a benefici ineguali per i partecipanti, in modo molto simile a quanto accade nel caso del controllo riproduttivo su individui imparentati. Per esempio, un babbuino maschio (Papio anubis) ne recluta un altro nel suo gruppo solo se sta tentando di accoppiarsi con una femmina che in quel momento sia legata a un maschio di rango superiore. Se l'alleanza ha successo, l'individuo dominante nell'alleanza ha la priorità nell'accoppiamento. Al contrario, alcune alleanze di delfini con naso a bottiglia, o tursiopi (Tursiops truncatus), persistono per lungo tempo, con due o tre maschi che cooperano in più occasioni per corteggiare e richiamare le femmine e non esistono prove di un successo riproduttivo asimmetrico. Tutti i maschi di un'alleanza spesso si accoppiano con le femmine che hanno radunato. Rispetto ai babbuini la cooperazione più stretta che si riscontra fra i delfini maschi è quindi compatibile con la casualità del successo riproduttivo dei singoli partecipanti, agendo probabilmente in aggiunta alla scelta del partner come meccanismo per prevenire l''imbroglio' e promuovere la cooperazione.
La cooperazione nell'uomo
Quale dei meccanismi citati può aver avuto un ruolo importante nel favorire la cooperazione tra i nostri antenati? Quasi certamente tutti. Le interazioni cooperative in Vertebrati non umani e nelle famiglie umane sono spesso coerenti con le previsioni formulate in base alle teorie della selezione di parentela e del controllo riproduttivo. Per esempio, l'assistenza più onerosa nell'allevare i bambini è fornita dai parenti più stretti. È meno probabile, però, che la selezione di parentela sia in grado di spiegare le interazioni tra famiglie. Quando le famiglie si raggruppavano per formare unità sociali più grandi la ripetizione delle interazioni deve aver creato le opportunità per alleanze sia a carattere temporaneo sia stabili, basate sulla scelta dei partner con cui cooperare e sulla reciprocità. Le circostanze ecologiche che favorivano relazioni a lungo termine tra individui in piccoli gruppi avrebbero posto le basi per la cooperazione. La longevità e la capacità di ricordare l'identità dei singoli individui oltre all'esito delle interazioni passate forniscono tutti gli ingredienti necessari per una reciprocità stabile. Finché gruppi di individui possono anticipare molte opportunità future di scambi proficui, la cooperazione appare inevitabile.
Né la reciprocità né la selezione di parentela possono spiegare adeguatamente i casi di cooperazione etnica in cui grandi gruppi di individui convergono su obiettivi comuni e arrivano spesso al punto di difendere questi obiettivi con la guerra. Tra gli animali non umani non imparentati non esistono casi analoghi di cooperazione su larga scala. Questo suggerisce che la cooperazione etnica possa essere il risultato di un qualche sistema di eredità culturale in cui il gruppo, piuttosto che l'individuo, è oggetto di selezione. La selezione a livello di gruppo culturale è stata proposta per spiegare perché gli uomini, a differenza di tutti gli altri animali, cooperino, spesso contro i propri stessi interessi, con individui con cui non condividono geni.
Bibliografia
Axelrod 1984: Axelrod, Robert M., The evolution of cooperation, New York, Basic Books, 1984.
Boyd, Richerson 1991: Boyd, Robert - Richerson, Peter J., Culture and cooperation, in: Cooperation and prosocial behaviour, edited by Robert A. Hinde and Jo Groebel, New York-Cambridge, Cambridge University Press, 1991, pp. 27-48.
Dugatkin 1997: Dugatkin, Lee A., Cooperation among animals: a modern perspective, New York, Oxford University Press, 1997.
Emlen 1995: Emlen, Stephen T., An evolutionary theory of the family, "Proceedings of the National Academy of Sciences USA", 92, 1995, pp. 8092-8099.
Hamilton 1964: Hamilton, William D., The genetic evolution of social behavior, "Journal of theoretical biology", 7, 1964, pp. 1-51.
Maynard Smith, Szathmáry 1995: Maynard Smith, John - Szathmáry, Eörs, The major transitions in evolution, Oxford, Freeman, 1995.
Trivers 1971: Trivers, Robert L., The evolution of reciprocal altruism, "Quarterly review of biology", 46, 1971, pp. 35-57.
West Eberhard 1975: West Eberhard, Mary J., The evolution of social behavior by kin selection, "Quarterly review of biology", 50, 1975, pp. 1-33.
Williams 1966: Williams, George C., Adaptation and natural selection, Princeton, Princeton University Press, 1966.