
Cooperazione
Cooperazione
La cooperazione si verifica quando due organismi traggono beneficio da un'azione comune. Essa pone un dilemma evolutivo nel caso in cui un individuo si trovi nella possibilità di 'imbrogliare', ottenendo così i benefici senza cooperare. Analisi teoriche e studi dettagliati condotti in molti sistemi biologici suggeriscono che la parentela e la fedeltà al partner costituiscono i meccanismi più importanti per spiegare l'evoluzione della cooperazione negli animali. Anche le pari opportunità riproduttive, la possibilità di scegliere il partner con cui cooperare e il controllo reciproco possono favorire la cooperazione. Comprendere i fenomeni della cooperazione può aiutare a gettare luce sulla virulenza dei parassiti, sulle transizioni chiave nell'evoluzione della vita, dall'origine delle molecole autoreplicanti alla meiosi, e sul comportamento umano.
La cooperazione e il dilemma evolutivo
Alcuni organismi si comportano spesso in modo da aiutarsi reciprocamente per sopravvivere e riprodursi. Molte piante in fiore, per esempio, producono nettare come fonte di cibo per animali che a loro volta trasportano da un fiore all'altro il polline consentendo la fecondazione. Sebbene tale meccanismo sia a prima vista efficiente per garantire la sopravvivenza di queste specie, un semplice ragionamento illustra il problema evolutivo che si pone nel caso di una cooperazione di questo tipo. Data una popolazione di piante che producono nettare, per quale motivo non esiste una pianta 'imbrogliona' che tenta di produrre meno nettare e più polline? Finché gli impollinatori visiteranno un mutante di questo tipo la sua frequenza nella generazione successiva aumenterà, in quanto esso sarà in grado di produrre più polline e quindi di garantire la formazione di più semi. Chiaramente la cooperazione tra piante e impollinatori dovrebbe persistere solo se gli individui che 'imbrogliano', in questo caso quelli che non producono l'appropriato quantitativo di nettare, fossero esclusi o eliminati.
Esistono parecchi meccanismi in grado di limitare la presenza di 'imbroglioni' nei sistemi cooperativi. Un meccanismo ampiamente riconosciuto è la fedeltà al partner; in altre parole, la cooperazione è limitata solo a quegli individui che interagiscono ripetutamente tra loro. Queste interazioni possono verificarsi perché gli animali vivono in società chiuse o perché possono riconoscersi reciprocamente e ricordare le interazioni passate. Quando le interazioni non sono ripetute, una possibile alternativa è che in alcune situazioni un animale aiuti solo il partner più cooperativo. Nell'esempio dell'impollinazione ciò potrebbe verificarsi se l'impollinatore fosse in grado di discriminare tra piante diverse e non trasportasse il polline di quei fiori che non producono abbastanza nettare. In realtà un certo grado di 'imbroglio' può essere tollerato. È ragionevole attender si che ciò avvenga solo quando gli individui che cooperano sono geneticamente imparentati e la scelta del partner è impossibile. La parentela permette a un individuo di trarre vantaggio anche dall'azione più egoistica del partner per 'procura' genetica.


Il grado dell'inganno in un qualunque sistema cooperativo dipende da quale è il momento in cui i partner beneficiano della cooperazione; inoltre varia a seconda che i reciproci benefici richiedano o meno un'azione collettiva. l mutualismi che coinvolgono due individui della medesima specie o di specie differenti sono caratterizzati da situazioni nelle quali tutti ottengono un beneficio immediato come diretta conseguenza della loro azione collettiva. l licaoni (Lycaon pictus), per esempio, inseguono e uccidono prede molto più grandi di loro cacciando in gruppo (fig. 1). Il successo della caccia, misurato come quantità di prede uccise da ciascun licaone per chilometro percorso, è più elevato per quelli che vivono in branchi numerosi (Creel e Creel, 1995). Quando la partecipazione alla caccia di gruppo è necessaria per catturare e uccidere la preda, l'inganno non paga. l benefici ricevuti simultaneamente all'azione quasi certamente caratterizzano altri esempi di caccia cooperativa tra cui quello di alcune formiche tropicali che in gruppo catturano prede molto più grandi di loro (fig. 2).
Se si considera la cooperazione semplicemente come un mutuo beneficio derivante da un'azione qualsiasi, allora moltissimi comportamenti egoisti possono essere considerati cooperativi quando, involontariamente, favoriscono altri individui. Qualunque atto egoistico finalizzato a incrementare la capacità di alimentarsi o a mettere in guardia e difendere se stesso o la propria prole può nel contempo aiutare altri organismi mostrando loro dove alimentarsi, avvertendo li del pericolo o mettendo in fuga i predatori (West Eberhard, 1975). Questi casi rappresentano esempi di mutualismo involontario perché sono conseguenza di un atto egoistico. In questi casi gli 'imbroglioni' non possono 'imbrogliare' perché non esiste un comportamento alternativo che fornisca all'individuo egoista un successo riproduttivo o una probabilità di sopravvivenza maggiori.
Le opportunità per l'inganno si originano ogni volta che le azioni da cui trarre beneficio richiedono uno sforzo collettivo o sono separate nel tempo. Scambi che implicano donazioni di qualche tipo, per esempio di cibo, sono chiamati mutualismi con beneficio di ritorno quando il costo della donazione è piccolo o inesistente e altruismi reciproci quando la donazione è costosa (Trivers, 1971). Anche se gli scambi reciproci che implicano lunghi ritardi temporali caratterizzano una grande varietà di pratiche culturali umane, molti biologi evoluzionisti ritengono che la reciprocità sia rara tra gli animali, in quanto tenere fuori gli 'imbroglioni' da questo sistema probabilmente è difficile. Affinché tutti gli individui traggano profitto dalla reciprocità è necessario che ricordino il valore della merce scambiata in passato e l'identità del partner con cui lo scambio è avvenuto. Gli 'imbroglioni' che solo di rado pagano i loro debiti sarebbero difficili da scoprire senza un considerevole controllo da parte dei partecipanti.
Anche se tuttora vi sono controversie sulla diffusione e sull'evoluzione della cooperazione in natura, elaborazioni teoriche e prove sperimentali consistenti indicano che sia le interazioni fedeli e ripetute sia la parentela tra i partner costituiscono due meccanismi determinanti per l'evoluzione della cooperazione. In assenza della fedeltà al partner o della parentela sono necessari altri meccanismi sociali per controllare la presenza e il comportamento di individui che tentano di ottenere dalla cooperazione più della loro onesta quota di benefici.
Recentemente sono stati presi in considerazione diversi meccanismi possibili in grado di favorire la cooperazione: le pari opportunità riproduttive, la scelta del partner con cui cooperare e la vigilanza reciproca. Sebbene vi siano prove a sostegno di ciascuna di queste ipotesi, nuovi studi teorici e sperimentali potranno senza dubbio migliorare le nostre conoscenze sul modo in cui viene mantenuta la cooperazione nei diversi sistemi.
Molti esempi riportati in questo saggio illustrano forme e meccanismi di cooperazione tra animali interagenti. Nonostante ciò la maggior parte della teoria ha una rilevanza diretta per le interazioni tra le cellule all'interno di organismi, tra i differenti elementi genetici in una cellula, e persino tra le molecole in un brodo primordiale. Comprendere l'evoluzione della cooperazione può aiutare a capire la virulenza dei parassiti, le malattie come il cancro, in cui le linee cellulari competono anziché cooperare, e alcune transizioni chiave nell'evoluzione della vita, dalle origini delle molecole replicanti agli organismi multicellulari (Maynard Smith e Szathmary, 1995).
Cooperazione e fedeltà al partner
Reciprocità
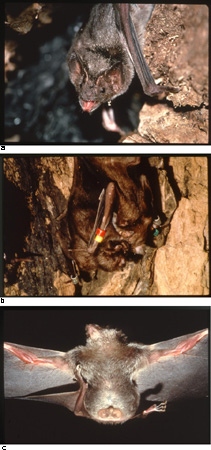
La fedeltà al partner costituisce la base per la reciprocità della cooperazione. R.L. Trivers (1971) fu il primo a mettere in evidenza che fino a quando due individui si aiutano l'un l'altro ripetutamente, sopravvivono e si riproducono più di due individui che non cooperano. Trivers chiamò questa forma di cooperazione altruismo reciproco, per cercare di spiegare che in questi casi la cooperazione avviene a dispetto di un costo immediato per la riproduzione o la sopravvivenza. Se i benefici superano i costi ciascun partecipante trarrà nel tempo profitto dall'interazione. Tuttavia, gli individui che non restituiscono il beneficio trarranno un profitto ancora maggiore. Affinché la reciprocità rimanga vantaggiosa, gli individui che cooperano devono apprendere e ricordare le precedenti interazioni per identificare gli 'imbroglioni' e applicare ritorsioni contro di loro. La condivisione del cibo nel vampiro vero di Azara (Desmodus rotundus) illustra i potenziali vantaggi della reciprocità (fig. 3). Le femmine rigurgitano il sangue per darlo alla loro prole e ad altri membri della colonia che non siano riusciti a procurarselo (Wilkinson, 1984). Ogni notte il 7% degli adulti e il 33% dei giovani non riesce ad alimentarsi in quanto le prede, costituite in gran parte dell'America latina da bovini o cavalli, spesso li riconoscono e li allontanano. Sembra che i giovani, meno esperti, abbiano maggiori difficoltà a evitare di essere scoperti rispetto agli individui adulti. Esperimenti con individui in cattività indicano che i pipistrelli vampiri non solo identificano e alimentano di preferenza gli individui della stessa colonia, ma rigurgitano più facilmente il sangue a beneficio di quegli individui che in precedenza avevano offerto loro del cibo. Quest'aiuto può essere di enorme importanza dato che un individuo che non riesce ad alimentarsi per tre notti consecutive è destinato a morire. In assenza della condivisione reciproca del sangue la mortalità annuale supererebbe l'80% mentre, di fatto, le femmine selvatiche possono vivere più di 20 anni.
Il meccanismo attraverso cui tali pipistrelli vampiri riconoscono ed escludono gli 'imbroglioni', ossia quegli individui che ricevono il sangue senza offrirlo mai, resta sconosciuto. Tuttavia, i pipistrelli vampiri che riescono ad alimentarsi con successo possono ingerire una quantità di sangue maggiore di 30 g, equivalente alloro peso corporeo (v. figura 3c). Probabilmente, durante le operazioni di pulizia reciproca gli altri pipistrelli vampiri si accorgono dello stomaco rigonfio dovuto all'ingestione di tanto sangue (v. figura 3b). La pulizia reciproca avviene frequentemente appena prima di rigurgitare il sangue, come è, in effetti, da attendersi se durante tale operazione avviene una sorta di valutazione della storia alimentare recente degli altri membri della colonia. Poiché in natura sia la pulizia reciproca che la condivisione del sangue avvengono solo tra individui di una stessa colonia che abbiano trovato dimora nello stesso albero cavo per mesi o anni, la fedeltà del partner sembra essere essenziale per mantenere questo sistema cooperativo.
Il dilemma del prigioniero
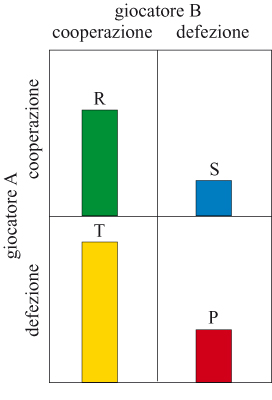
L'effetto della fedeltà al partner sull' evoluzione della cooperazione è stato esaminato attentamente da un punto di vista teorico. Gran parte di questa teoria si basa sull'analisi di un gioco ripetuto da due partecipanti che prende il nome di dilemma iterato del prigioniero. Anziché soldi i giocatori biologici acquistano o perdono punti che riflettono differenze nel loro successo riproduttivo complessivo, o fitness (la 'moneta' dell'evoluzione). In una mano del gioco ciascun giocatore può ottenere un premio (punteggio R, Reward, premio) se entrambi cooperano, e una punizione (punteggio P, Punishment, punizione) se entrambi non cooperano. La tentazione di non cooperare con un partner che coopera comporta un punteggio più alto (punteggio T, Temptation, tentazione), mentre, cooperare con un partner che non coopera comporta il punteggio più basso (punteggio S, Sucker, ingenuo). In termini algebrici il dilemma del prigioniero esiste se T> R > P > S e R> (T + S) /2 (fig. 4). In una sola mano del gioco la migliore strategia è non cooperare, anche se i giocatori otterrebbero un punteggio migliore cooperando entrambi. Questo si verifica perché un giocatore che non coopera evita di ottenere S se l'altro non coopera oppure ottiene T se l'altro coopera. Cioè per il giocatore A non cooperare è la miglior risposta a entrambe le possibili mosse del giocatore B in una singola mano del gioco.
Se si giocano più mani e i giocatori basano le loro decisioni sulle interazioni precedenti può emergere la cooperazione. In una serie di simulazioni al computer R. Axelrod (1984) scoprì che la semplice strategia TFT (Tit For Tat, occhio per occhio) aveva un rendimento superiore a tutte le altre strategie possibili. La strategia TFT incarna la regola aurea che consiste nell'iniziare sempre cooperando e nel copiare semplicemente la precedente decisione del partner nelle interazioni successive. Di conseguenza chi applica la strategia TFT è gentile, perché non è mai in difetto per primo, ed è disposto a perdonare, perché è in difetto solo una volta se il partner riprende a cooperare. Questa strategia ha successo se la probabilità che due individui si incontrino di nuovo è abbastanza grande; di conseguenza la fedeltà al partner consente di escludere gli 'imbroglioni'.
Una teoria più recente indica che la strategia TFT potrebbe rappresentare solo il primo stadio di evoluzione della cooperazione. Sebbene questa sia la migliore strategia in un ambiente uniforme, è vulnerabile a errori involontari. Una defezione accidentale tra due individui che adottano TFT porterà a cicli di mutua defezione frnché un secondo errore non correggerà l'incomprensione. La strategia GTFT (Generous Tit For Tat, occhio per occhio generoso) prevede di rispondere a una defezione con un'altra defezione solo in due terzi dei casi: questa strategia perdona di più di quella TFT e quindi è immune da questi errori occasionali. Simulazioni al computer effettuate da M. Nowak e K. Sigmund (1993) mostrano che spesso dalla strategia GTFT si origina una strategia chiamata PAVLOV. Quest'ultima è così definita perché stabilisce la mossa successiva basandosi sul punteggio ottenuto in quella precedente come se rispondesse a un condizionamento: ripete la mossa se è stata premiata con T o R, cambia se è stata punita con S o P. In altre parole PAVLOV obbedisce alla regola "se vinci continua così, se perdi cambia". La strategia PAVLOV funziona meglio di quella TFT in un ambiente in cui si verificano errori perché è in grado di correggerli e perché sfrutta un individuo che coopera in maniera incondizionata.
Diversi esempi di cooperazione tra gli animali sembrano implicare la reciprocità e sono compatibili con le strategie che implicano una ritorsione, come la strategia TFT e quella PAVLOV (Dugatkin, 1997). La parula dal cappuccio (Wilsonia citrina), per esempio, mostra un aumento di aggressività in risposta a registrazioni che simulano incursioni territoriali da parte dei maschi di territori adiacenti, ma non a quelle di individui sconosciuti, come ci si aspetterebbe in caso di ritorsione contro i vicini che non rispettano i confini. In alcuni pesci serranidi ermafroditi e in alcuni policheti, le uova e gli spermatozoi sono scambiati in sequenze di piccole aliquote, presumibilmente per minimizzare il possibile guadagno che si potrebbe ottenere con l'inganno, trasferendo solamente gli spermatozoi, che sono piccoli e quindi richiedono poco dispendio energetico, e non le uova. Una serie di esperimenti sugli spinarelli (Gasterosteus aculeatus) e sul guppy (Poecilia reticulata) hanno dimostrato che questi pesci spesso lasciano il branco e si avvicinano in coppia a predatori molto più grandi di loro. Si ritiene che il controllo del predatore sia utile alle coppie di pesci in avvicinamento perché in tal modo ciascuno ottiene informazioni sul tipo di predatore che si trova nelle vicinanze e sulla probabilità di essere attaccato. Le coppie di individui interagiscono ripetutamente nel tempo e si muovono a turno verso il predatore, come potremmo prevedere immaginando che ogni coppia di movimenti rappresenti una mano nel gioco del dilemma del prigioniero.
L'importanza del dilemma del prigioniero per spiegare la cooperazione tra gli animali resta tuttavia controversa perché, anche se ognuno di questi esempi è generalmente compatibile con la strategia TFT, per la maggior parte di essi esistono spiegazioni alternative, alcune delle quali non assegnano i punteggi secondo i criteri del dilemma del prigioniero. In particolare, il dilemma del prigioniero assume che entrambi i giocatori prendano decisioni simultanee sulla mossa successiva. Se invece gli animali alternano le mosse, come nel caso della condivisione del cibo, del controllo del predatore e della difesa del territorio, le simulazioni al computer dimostrano che un'altra strategia, denominata FBF (Firm But Fair, risoluto ma onesto), alla fine elimina le strategie GTFT e PAVLOV (Frean, 1994). Questa nuova strategia è risoluta perché l'individuo che la adotta applica una ritorsione, defezionando dopo aver cooperato con un 'imbroglione', ed è onesta perché esso non applica ritorsioni contro un 'imbroglione' se ha ingannato esso stesso e coopera con gli ingenui piuttosto che continuare a sfruttarli. Così l'ordine in cui si gioca non influenza il risultato di una singola mano, ma il comportamento quando i giocatori si scambiano interazioni non simultanee.
La difesa territoriale nel leone (Panthera leo) è un'eccezione istruttiva a gran parte di queste teorie perché alcuni individui sembrano attuare una cooperazione incondizionata anche in risposta a una defezione. I leoni maschi si riuniscono in coalizioni che competono tra loro per il controllo di un gruppo di femmine. Piccole coalizioni sono composte generalmente da individui non imparentati mentre quelle più grandi sono caratterizzate dalla presenza di fratelli. Sebbene le sostituzioni di gruppi di maschi nel controllo delle femmine siano rare (solo una ogni due o tre anni) la difesa territoriale è stata studiata approfonditamente diffondendo ruggiti con degli altoparlanti per simulare intrusioni da parte di una coalizione concorrente. Tutti i maschi di una coalizione che in un certo momento controlla un gruppo di femmine si avvicinano agli altoparlanti indipendentemente dal comportamento o dal grado di parentela degli altri componenti. La mancanza di qualsiasi apparente defezione tra maschi di una coalizione ha senso perché il successo riproduttivo di ognuno dipende dal mantenere il controllo sul branco di femmine. Dato che tutti i membri di una coalizione si accoppiano e condividono la paternità, ciascun maschio ottiene un beneficio aiutando a cacciare la coalizione nemica.
Contrariamente alla difesa incondizionata dei maschi, le leonesse nella stessa situazione si precipitano più difficilmente a difendere il branco (Heinsohn e Packer, 1995). Le femmine si avvicinano agli altoparlanti solo quando il numero di individui che ruggisce è inferiore a quello delle leonesse del gruppo, in altre parole quando il pronostico è a loro favore. Inoltre, le singole femmine differiscono sensibilmente nella motivazione a guidare la carica. Alcune agiscono sempre spavaldamente e si muovono verso gli altoparlanti, altre si mostrano spaventate e si attardano dietro alle prime. Anche se le femmine leader spesso si voltano a guardare le loro compagne indecise, esse continuano ad avvicinarsi agli altoparlanti senza mostrare alcuna forma di ritorsione. Le leonesse sembrano quindi tollerare le compagne 'imbroglione' che non cooperano, almeno nel contesto della difesa territoriale. Sebbene la cooperazione ripetuta in risposta a un'apparente mancanza di cooperazione sia incompatibile persino con la strategia più incline al perdono tra quelle di successo nel dilemma del prigioniero, sono stati osservati altri casi (sempre tra parenti) in cui vi è una disuguaglianza nei benefici della cooperazione. La parentela, perciò, fornisce un'importante ragione per cooperare in queste situazioni (v. oltre).
Cooperazione e affinità genetica
Selezione di parentela
Essere imparentati vuoI dire condividere geni. La riproduzione implica la duplicazione e la trasmissione di geni alla prole. La probabilità che una copia esatta di un gene si trovi in un altro individuo è chiamata grado di parentela ed è indicata con il simbolo r. Nella specie umana e negli altri organismi diploidi, cioè quelli che possiedono due copie dei geni su cromosomi appaiati, il grado di parentela della madre con il figlio è pari a 1/2 perché il figlio riceve metà dei suoi geni dalla madre e metà dal padre. Lo stesso vale per il grado di parentela di un figlio con la madre. Qualunque coppia di individui con un antenato comune è in qualche modo imparentata, sebbene r possa essere molto piccolo se l'antenato risale a molte generazioni precedenti. In ogni caso r è misurato come la probabilità che un gene tipico di un individuo abbia una copia esatta in un altro a causa della discendenza diretta da un antenato comune.
L'affinità genetica è importante perché ingannare in un'interazione cooperativa potrebbe non essere vantaggioso se il partner è un parente. Va ricordato che la selezione naturale favorisce quei caratteri che aumentano il numero di figli prodotti da un individuo nell' arco della sua vita e la loro sopravvivenza. La ragione è che i genitori sono imparentati con i propri figli e condividono con essi una parte dei geni. Dal punto di vista di un gene, qualunque comportamento che aumenti la sua frequenza nella generazione successiva sarà favorito. Di conseguenza la selezione naturale dovrebbe anche favorire i caratteri che aumentano il numero di altri parenti come i fratelli o i cugini e la loro sopravvivenza. Tuttavia, dato che i fratelli hanno un grado di affinità genetica più elevato dei cugini, si prevede che gli animali siano in grado di discriminare e di conseguenza interagire di preferenza con i parenti più stretti.
Se due animali hanno un grado sufficiente di affinità genetica anche il comportamento altruistico può essere favorito. Un gene che influenzi il comportamento altruistico si diffonderà quando i benefici per l'altruista (B) moltiplicati per il grado di parentela (r), sono maggiori del costo per l'individuo che dà l'avvio al comportamento (C); in altre parole, quando rB - C> O (Hamilton, 1964). Questa semplice disequazione, nota come regola di Hamilton, è il fondamento di un'importante teoria denominata selezione di parentela (Maynard Smith e Szathmary, 1995). La selezione di parentela influenza l'evoluzione del comportamento altruistico e di molte altre forme di comportamento sociale in una grande varietà di animali.
Per illustrare in che modo la selezione di parentela influenzi il comportamento cooperativo consideriamo la costruzione del nido e l'allevamento della prole nel topolino delle case (Mus domestieus). Spesso due o tre femmine di questa specie costruiscono il nido, partoriscono insieme e ciascuna accudisce indifferentemente tutti i piccoli presenti nel nido comune. Un vantaggio importante nel costruire un nido comune potrebbe consistere nel fatto che due o tre femmine proteggono meglio la prole dai maschi, potenzialmente infanticidi. Tuttavia, dato che l'allattamento è in genere il periodo energeticamente più dispendioso nella vita di ogni femmina di mammifero, per quale motivo le femmine di topo non evitano di accudire i piccoli nel nido comune? La risposta sembra essere che in genere ciascuna femmina è imparentata con tutti i piccoli. I topo lini delle case, infatti, costruiscono i nidi comuni di preferenza con parenti stretti come sorelle e figlie (Manning et al., 1992). Le femmine che forniscono latte agli altri piccoli oltre che ai propri lasciano alle generazioni successive più copie dei propri geni rispetto alle femmine meno cooperative.
Riconoscimento dell'affinità genetica
Affinché la selezione di parentela possa funzionare, le femmine di topo devono essere capaci di riconoscere quelle più affrni geneticamente e nidificare di preferenza insieme a esse. In altre parole, devono saper distinguere i parenti dai non parenti. In alcuni animali questo tipo di riconoscimento è basato su un'associazione verificatasi in precedenza. Per esempio, un odore comune acquisito da individui cresciuti insieme, cibandosi degli stessi alimenti, può impartire caratteristiche olfattive riconoscibili. Un odore specifico del nido può permettere a un animale di effettuare discriminazioni semplici distinguendo tra fratelli e individui non imparentati, ma non è molto utile per prevedere l'affinità genetica negli individui di una nidiata che presentano gradi di parentela diversi, come accade se due o più maschi ne condividono la paternità. In questi casi sarebbe meglio se gli animali potessero in qualche modo confrontare direttamente i geni per determinare il grado di affinità.
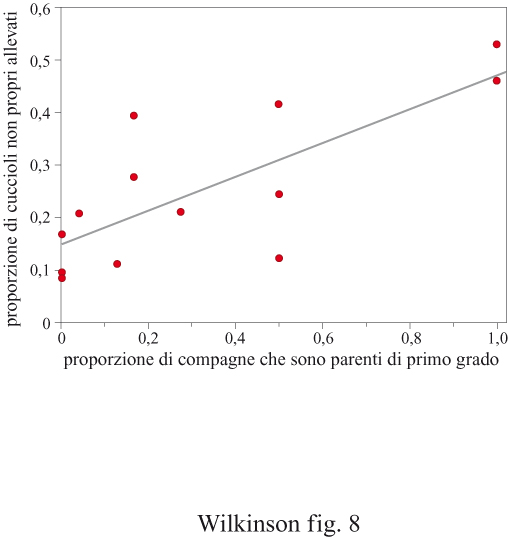
Il riconoscimento dell'affinità genetica richiede la presenza di regioni variabili nel genoma e la capacità sensoriale di riconoscere e discriminare gli individui sulla base di queste differenze. Altrimenti, in presenza di geni identici, l'effetto di un antenato comune non può essere discriminato da quello dovuto al caso. Studi su topi congenici opportunamente incrociati al fine di risultare geneticamente identici per gli alleli nel complesso dei loei multipli di istocompatibilità, o MHC (Major Histoeompatibility Complex), hanno dimostrato che le femmine di topo formano nidi comuni se condividono il medesimo aplotipo MHC (Manning et al., 1992). In tutti i vertebrati il complesso MHC è caratterizzato da un'enorme variabilità genetica per facilitare il riconoscimento cellula-cellula allo scopo di difendere l'organismo dalle invasioni di cellule parassite da parte di organismi portatori di malattie. Di conseguenza i loei MHC sono ideali come indicatori genetici del grado di parentela e potrebbero svolgere questa funzione in molti altri vertebrati, dai tunicati all'uomo. Come nei topo lini delle case, le leonesse africane che condividono un territorio accudiscono i piccoli in cooperazione. Una delle principali cause di mortalità dei cuccioli è l'infanticidio che avviene in seguito all'avvicendamento tra coalizioni di maschi. Come nei topolini, il numero di femmine determina il grado di protezione dei piccoli dai maschi infanticidi. Ma i leoni discriminano più accuratamente rispetto ai topo lini. Le femmine che allattano permettono ai cuccioli geneticamente più affini di prendere più latte (fig. 8) rispetto a quelli meno affini (Pusey e Packer, 1994). In che modo le leonesse riconoscano i livelli di parentela con i piccoli è ancora da chiarire, ma l'età, un'eventuale associazione precedente e il grado di somiglianza dei loei MHC sono tutti meccanismi plausibili.
Controllo riproduttivo in individui imparentati

Quando in una struttura sociale gli individui imparentati appartengono a diversi livelli di una scala gerarchica e hanno scarse opportunità di vivere al di fuori di un gruppo sociale, la cooperazione può avvenire anche se i partecipanti ne traggono benefici di entità differente. Un esempio è costituito dall'allevamento in cooperazione nell'ani beccosolcato (Crotophaga suleirostris). Questi cuculi neri e lucenti vivono in gruppi sociali costituiti da una a quattro coppie di uccelli che condividono un nido comune. Le femmine depongono le loro uova secondo un ordine temporale: la più bassa nella gerarchia depone per prima, la dominante per ultima. Ciascuna femmina prima di cominciare a deporre rimuove alcune uova già presenti nel nido. Come conseguenza di questo comportamento di rimozione delle uova la femmina dominante contribuisce alla covata finale più delle altre (fig. 9). Quando le uova si schiudono, tutti i membri del gruppo partecipano alla nutrizione e alla protezione dei pulcini. Perché le femmine subordinate non abbandonano il gruppo per nidificare per conto loro? La ragione sembra essere la generale mancanza di un habitat di nidificazione di alta qualità nelle zone dell'America Centrale dove vivono questi uccelli. Di conseguenza una femmina subordinata produce più piccoli in un gruppo cooperativo di quanti ne potrebbe produrre da sola. Se poi è anche imparentata con la dominante può ricevere un ulteriore beneficio incrementando le probabilità di sopravvivenza degli altri piccoli (Vehrencamp, 1983).
Il controllo riproduttivo in individui imparentati può avvenire anche nei mammiferi come la mangusta nana meridionale (Helogale parvula), il licaone, il tamarino dai ciuffetti bianchi (Saguinus oedipus), e l'eterocefalo glabro (Heterocephalus glaber). In tutte queste specie, in genere, solo una singola femmina alleva i piccoli con successo. La riproduzione di altre femmine nel gruppo sociale è inibita dal rilascio di feromoni specifici oppure i loro piccoli sono uccisi alla nascita. Nonostante questo trattamento gli individui non riproduttori, che tipicamente sono imparentati con i dominanti, rimangono nel gruppo, raccolgono cibo e aiutano la coppia dominante nella cura della prole. L'eterocefalo costituisce un esempio estremo: vive in colonie di grandi dimensioni in aree deserte dell' Africa orientale e meridionale dove si ciba di grandi tuberi di poche specie di piante. Un adeguato sfruttamento di questa risorsa molto dispersa sul territorio richiede da parte della colonia lo sviluppo di un sistema di cunicoli esteso e in continua espansione. Una volta rinvenuto un tubero gli operai lo trasportano a pezzi fino alla tana comune e lo dividono con gli altri. Gli operai comunicano la loro scoperta con vocalizzazioni che richiamano i membri della colonia alla fonte di cibo. Anche se gli operai non potranno mai riprodursi, ottengono benefici indiretti aiutando la femmina riproduttiva perché il grado di affinità genetica all'interno di una colonia è in genere molto elevato a causa dell'inincrocio (inbreeding), quasi certamente derivante da accoppiamenti incestuosi precedenti. Contrariamente a quanto avviene nella maggior parte degli animali, gli eterocefali si allontanano raramente dalla colonia. L'esistenza di un controllo riproduttivo è stata riportata anche per le vespe del genere Polistes (West Eberhard, 1975). In molte specie le sorelle si allontanano insieme e costruiscono un nuovo nido. A causa di una particolare modalità di determinazione del sesso nota come aplodiploidia, nella quale i maschi portano una copia dei cromosomi mentre le femmine ne portano due, le sorelle nate da una madre fecondata da un solo maschio hanno il 75% di probabilità di condividere un gene identico per discendenza. Sebbene le sorelle possano inizialmente cooperare per costruire un nido, solo una (talora due) svilupperà alla fine gli ovari e deporrà le uova. Le rimanenti sorelle spesso diventano operaie che continuano a costruire il nido e a prendersi cura delle uova e delle larve. Da un punto di vista evolutivo una vespa subordinata dovrebbe essere disposta ad aiutare la regina, se così facendo trasmette più copie dei propri geni di quante ne potrebbe portare la sua progenie se invece decidesse di abbandonare il nido e tentare di costruirne uno nuovo.
Il problema della selezione di gruppo
Fino alla metà degli anni Sessanta la cooperazione era in genere ritenuta un meccanismo di regolazione della popolazione. Questo punto di vista è ben rappresentato dalla frase "la cooperazione avviene perché è cosa utile per la specie". Una critica dettagliata di G.C. Williams (1966) mise in evidenza i punti deboli di questa interpretazione. A meno che la migrazione tra popolazioni di conspecifici sia trascurabile e le estinzioni di queste popolazioni frequenti, gli individui che egoisticamente massimizzano la propria riproduzione escludono quelli che la limitano per ridurre la competizione. La principale eccezione a questa interpretazione si verifica quando gli animali sono imparentati tra loro. La scoperta che il controllo riproduttivo si riscontra tipicamente in animali che formano gruppi costituiti da parenti stretti, è perciò compatibile con l'idea che sia la selezione di parentela, piuttosto che quella di gruppo, a favorire la cooperazione.
Dalla metà degli anni Ottanta, alcuni biologi evoluzionisti hanno ipotizzato l'esistenza di una forma alternativa di selezione naturale a livello di gruppo. Piuttosto che da un'estinzione differenziale delle popolazioni, questo nuovo tipo di selezione a livello di gruppi contraddistinti da un carattere (trait-group selection) dipende da una produttività differenziale dei gruppi. Studi teorici hanno dimostrato che questa forma di selezione può favorire il comportamento cooperativo tra organismi se prima della dispersione essi si assortiscono in modo casuale in gruppi sulla base della loro predisposizione ad agire secondo uno stile cooperativo. In teoria questi gruppi potrebbero essere composti da animali non parenti, ma il meccanismo più semplice in grado di generare un assortimento non casuale in gruppi è l'associazione basata sulla parentela. Quando tali gruppi sono composti da parenti stretti la selezione di parentela, come menzionato in precedenza, è sufficiente per spiegare l'esistenza della cooperazione. Sebbene la selezione di gruppo sia stata dimostrata in laboratorio (Wade, 1977), pochi esempi (o forse nessuno) sono stati riscontrati in natura. Al momento, il caso più plausibile di trait-group selection riguarda la cooperazione tra gruppi umani non imparentati (Wilson e Sober, 1994).
Meccanismi che favoriscono la cooperazione
Pari opportunità riproduttive
Il comportamento dell'ani beccosolcato, dell'eterocefalo e delle vespe del genere Polistes potrebbe far prevedere che tutti i casi di cooperazione nell'allevamento della prole debbano riguardare animali strettamente affini. Tuttavia, in diverse specie di formiche e termiti le colonie possono essere fondate da più femmine riproduttive imparentate solo lontanamente o per nulla. I dati a disposizione mostrano che le colonie fondate da più regine tendono a crescere più rapidamente e hanno una maggiore probabilità di sopravvivere e di produrre individui riproduttivi rispetto a quelle fondate da una sola femmina. Di conseguenza la riproduzione pro capite è spesso più elevata in queste colonie con più regine, o poliginiche. Le termiti e le formiche differiscono dalle vespe, dagli uccelli e dagli eterocefali per il fatto che il mancato sviluppo riproduttivo è irreversibile. Mentre le operaie degli altri taxa sono capaci di sviluppare in qualunque momento gli organi riproduttivi, la maggior parte delle formiche e delle termiti non ha questa flessibilità fisiologica. Le regine di queste ultime specie devono sviluppare gli ovari e ingrandire l'addome prima di poter deporre le uova. Inoltre, mentre nelle vespe le regine in sovrannumero sono invariabilmente eliminate o finiscono con l'aiutare la regina senza riprodursi, entro pochi mesi dalla formazione della colonia, alcune regine di termiti e formiche possono vivere insieme e continuare a riprodursi per periodi molto più lunghi. Nonostante ciò la riproduzione cooperativa da parte di più fondatrici sembra rappresentare un periodo di transizione tipico soltanto della fase precoce di qualunque colonia di insetti sociali. Nella maggior parte dei casi alla fine insorgono dei conflitti e solo una regina sopravvive. Un'ipotesi plausibile in grado di spiegare l'esistenza di una cooperazione temporanea è che le fondatrici siano incapaci di determinare chi alla fine assumerà il controllo riproduttivo. La cooperazione può mantenersi per tempi prolungati se ciascuna fondatrice ha la stessa possibilità di assumere il dominio riproduttivo della colonia.

Un altro interessante esempio di cooperazione assicurata dalle pari opportunità riproduttive è quello del mixomicete Dictyostelium discoideum. Il ciclo vitale di questi organismi (fig. 12) passa da uno stadio a cellule libere e disperse a un aggregato multicellulare detto plasmodio, capace di compiere movimenti coordinati. Quando il plasmodio raggiunge un sito ecologicamente idoneo produce un corpo fruttifero contenente spore in cima a uno stelo. Sebbene la selezione di parentela possa spiegare perché le cellule dello stelo si sacrifichino per le loro spore sorelle, bisogna sottolineare il fatto che alcuni plasmodi si formano da più di 100.000 cellule aggregate. Nel plasmodio in via di formazione la determinazione casuale del destino delle cellule può favorire la cooperazione senza l'intervento della selezione di parentela. La distribuzione spaziale casuale dei diversi tipi cellulari al momento di stabilire il destino delle cellule sostiene quest'ipotesi (Frank, 1995).
Pari opportunità riproduttive caratterizzano anche una forma di cooperazione biologica di tipo molto diverso rispetto a quelle considerate finora: il passaggio dei cromosomi da una cellula diploide a gameti aploidi. Attraverso il processo della meiosi ciascun cromosoma ha il 50% di probabilità di fmire in un gamete. La presenza di cromosomi 'imbroglioni' dimostra che la meiosi rappresenta un esempio di cooperazione in buona fede. In una varietà di organismi può verificarsi la divisione meiotica guidata, cioè un cromosoma può finire in più della metà dei gameti. Un cromosoma sessuale che disobbedisce alle leggi della segregazione mendeliana provoca un'alterazione del rapporto primario tra i sessi. Sebbene questo comportamento egoista possa essere vantaggioso per i geni localizzati sul cromosoma 'imbroglione', il risultato della divisione meiotica guidata per un cromosoma sessuale è l'estinzione quando il sesso limitante scompare, a meno che qualche forza intervenga a bilanciare nuovamente le frequenze di trasmissione. Così la meiosi può essere considerata come un elaborato meccanismo per assicurare la cooperazione tra cromosomi (Maynard Smith e Szathmary, 1995).
Scelta del partner
Le interazioni mutualistiche che caratterizzano un gran numero di specie forniscono buoni esempi di come un meccanismo alternativo al precedente permetta di limitare gli 'imbroglioni' e mantenere la cooperazione. Tale meccanismo si basa sulla scelta del partner con cui cooperare (Bull e Rice, 1991). Molte specie di omotteri, come gli afidi e i membracidi, le larve di molte farfalle licenidi e riodinidi e alcune piante, come le acacie delle formiche, forniscono alle formiche soluzioni ricche di zuccheri in cambio della protezione dai predatori o dai competitori. Le singole piante o gli insetti potrebbero imbrogliare non producendo il premio per le formiche. La cooperazione, perciò, dovrebbe persistere finché le formiche non riescono a trovare piante o insetti alternativi che forniscano premi più consistenti. Allo stesso tempo insetti e piante potrebbero negare la ricompensa se le formiche assistenti non assicurassero un'adeguata protezione. Così la scelta del partner potrebbe regolare la cooperazione come avviene in qualunque economia di mercato (Noe e Hammerstein, 1994).
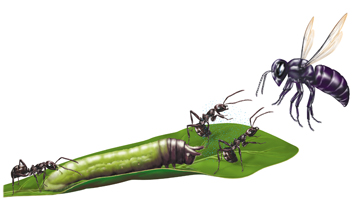
L'importanza della scelta del partner è stata dimostrata in studi sulle associazioni mutualisti che tra alcuni bruchi e formiche. I bruchi dei licenidi e dei riodinidi secernono soluzioni zuccherine da una ghiandola dorsale la cui unica funzione è quella di produrre una ricompensa per le formiche (fig. 14). Inoltre alcuni bruchi attraggono le formiche generando vibrazioni acustiche di richiamo e possono emettere segnali chimici che le stimolano ad attaccare i predatori in avvicinamento (De Vries, 1992). Si è osservato che bruchi isolati producono una maggiore quantità di soluzione zuccherina quando il numero delle formiche dedite alle loro cure viene aumentato da uno a due. Nelle specie di licenidi che formano grandi raggruppamenti spesso le formiche uccidono i singoli bruchi che non producono nettare. Mentre le pupe attraggono le formiche continuando a produrre la secrezione zuccherina, le farfalle che emergono dopo la schiusa sono soggette a predazione da parte delle formiche stesse, dimostrando ulteriormente che la cooperazione in questo sistema si basa sul valore di mercato del nettare (Noe e Hammerstein, 1994).
La scelta selettiva del partner può anche rafforzare il comportamento mutualistico intraspecifico. Per esempio, in certi primati e delfrni i maschi cooperano formando alleanze per attrarre le femmine sessualmente recettive. Le alleanze dei primati sembrano però differire da quelle dei delfini per almeno due aspetti. Le prime tendono a essere temporanee e danno luogo a benefici ineguali per i partecipanti in modo molto simile a quanto accade nel caso del controllo riproduttivo su individui imparentati. Per esempio, un babbuino maschio (Papio anubis) ne recluta un altro nel suo gruppo solo se sta tentando di accoppiarsi con una femmina che sia in quel momento legata a un maschio di rango superiore. Se l'alleanza ha successo l'individuo dominante nell'alleanza ha la priorità nell'accoppiamento. Al contrario, alcune alleanze di delfini con naso a bottiglia, o tursiopi (Tursiops truncatus), persistono per lungo tempo, con due o tre maschi che cooperano in più occasioni per corteggiare e richiamare le femmine e non esistono prove di un successo riproduttivo asimmetrico. Tutti i maschi di un'alleanza spesso si accoppiano con le femmine che hanno radunato (Connor et al., 1992). Rispetto ai babbuini la cooperazione più stretta che si riscontra fra i delfini maschi è quindi compatibile con la casualità del successo riproduttivo dei singoli partecipanti, agendo probabilmente in aggiunta alla scelta del partner come meccanismo per prevenire l'imbroglio e promuovere la cooperazione.
Vigilanza reciproca
Un'alternativa alla scelta del partner per rafforzare la cooperazione è reprimere la competizione tra individui appartenenti a un gruppo potenzialmente cooperativo. Il modo migliore per reprimere la competizione è conservare un livello sufficientemente elevato di parentela tra tutti i componenti del gruppo in modo da far coincidere i loro interessi riproduttivi. Nell'ape domestica (Apis mellifera), per esempio, l'esistenza di un controllo reciproco dipende dal grado di parentela. Le operaie femmine si riproducono deponendo uova non fecondate da cui si sviluppano maschi aploidi. Se, come avviene di solito, la regina si è accoppiata più volte, le operaie sono meno imparentate con la prole maschile aploide delle loro sorelle di quanto non lo siano con la prole della loro madre (Maynard Smith e Szathmary, 1995). Come è prevedibile, le operaie ispezionano continuamente le celle dell' alveare alla ricerca di uova deposte dalle sorelle. Quando trovano un uovo deposto da un'operaia lo distruggono immediatamente. Il comportamento di vigilanza si interrompe se la regina muore (Ratnieks e Visscher, 1989). Una situazione analoga si verifica durante la trasmissione degli elementi genetici citoplasmatici come gli organelli simbionti. Negli eucarioti le linee genetiche di elementi simbionti citoplasmatici non sono mescolate durante la trasmissione e ciò limita le opportunità per un conflitto genetico. Per esempio, vi sono prove importanti a sostegno dell'ipotesi che i mitocondri e i cloroplasti si siano sviluppati da batteri che formarono un'associazione simbiotica con le cellule eucariotiche primitive. Questi organelli sono ora trasmessi dalla madre a tutti i figli. L'eredità uniparentale assicura che uno zigote contenga organelli derivanti da una singola linea genetica. Al contrario, l'ereditarietà biparentale di organelli ridurrebbe la parentela tra gli organelli all'interno di una cellula e tra cellule diverse. La diversità genetica tra mitocondri potrebbe influenzarne le capacità metaboliche e generare competizione tra linee cellulari per la trasmissione alla generazione successiva. Come ogni vittima del cancro può testimoniare, la competizione intercellulare è molto pericolosa per l'organismo. L'eredità uniparentale agisce quindi per mantenere un elevato grado di parentela tra gli organelli. La prova meno confutabile a sostegno dell'importanza della trasmissione verticale uniparentale nel favorire la cooperazione si ottiene esaminando la variazione del grado di pericolosità dei parassiti per i loro ospiti in funzione della modalità di trasmissione. La virulenza dei parassiti eucarioti e procarioti è associata alle forme di trasmissione che permettono l'infezione da parte di più linee genetiche (Maynard Smith e Szathmary, 1995). Le infezioni multiple sono più frequenti in caso di trasmissione orizzontale piuttosto che verticale, in particolare se la trasmissione verticale è uniparentale. Prove sperimentali a sostegno di una relazione tra virulenza e trasmissione orizzontale sono state ottenute per il batterio Escherichia coli infettato da un fago a DNA filamentoso (Bull et al., 1991). Quando le condizioni di crescita permettevano solo la trasmissione verticale l'effetto deleterio dei fagi sul tasso di crescita dei batteri era minimo. Quando le condizioni di crescita dei batteri favorivano la coniugazione, permettendo quindi la trasmissione orizzontale, apparivano fagi con effetti più pericolosi. Anche dati comparativi riguardanti gli effetti dei fagi sui batteri supportano l'esistenza di una relazione fra trasmissione orizzontale e virulenza (Maynard Smith e Szathmary, 1995).
Diversi eventi nella storia della vita possono essere interpretati come modi per assicurare elevati livelli di parentela tra entità genetiche potenzialmente variabili che devono cooperare per funzionare correttamente (Maynard Smith e Szathmary, 1995). Per esempio, negli organismi multicellulari l'ubiquità della riproduzione a partire da una singola cellula fa sì che tutte le cellule di un embrione in via di sviluppo siano geneticamente identiche.
La separazione precoce della linea germinale dalle cellule da cui si sviluppa il resto del corpo può ulteriormente favorire il controllo di qualunque eventuale competizione tra linee cellulari. Inoltre, la separazione precoce delle cellule della linea germinale riduce anche il numero di divisioni cui queste devono andare incontro e la probabilità che le mutazioni somatiche portino a una diminuzione del grado di affinità genetica tra le cellule che producono gameti. Un caso parallelo di separazione di linee cellulari in potenziale competizione riguarda molti organismi simbionti che sono spesso alloggiati in organi speciali nel corpo dell'ospite (Frank, 1996).
La cooperazione nell'uomo
Quale dei meccanismi citati può aver avuto un ruolo importante nel favorire la cooperazione tra i nostri antenati? Quasi certamente tutti. Le interazioni cooperative in vertebrati non umani e nelle famiglie umane sono spesso coerenti con le previsioni formulate in base alle teorie della selezione di parentela e del controllo riproduttivo (Emlen, 1995). Per esempio, l'assistenza più onerosa nell'allevare i bambini è fornita dai parenti più stretti. È meno probabile, però, che la selezione di parentela sia in grado di spiegare le interazioni tra famiglie. Quando le famiglie si raggruppavano per formare unità sociali più grandi la ripetizione delle interazioni deve aver creato le opportunità per alleanze sia a carattere temporaneo sia stabili, basate sulla scelta dei partner con cui cooperare e sulla reciprocità. Le circostanze ecologiche che favorivano relazioni a lungo termine tra individui in piccoli gruppi avrebbero posto le basi per la cooperazione. La longevità e la capacità di ricordare l'identità dei singoli individui oltre all'esito delle interazioni passate forniscono tutti gli ingredienti necessari per una reciprocità stabile. Finché gruppi di individui possono anticipare molte opportunità future di scambi proficui, la cooperazione appare inevitabile.
Né la reciprocità né la selezione di parentela possono spiegare adeguatamente i casi di cooperazione etnica in cui grandi gruppi di individui convergono su obiettivi comuni e arrivano spesso al punto di difendere questi obiettivi con la guerra. Tra gli animali non umani non imparentati non esistono casi analoghi di cooperazione su larga scala. Questo suggerisce che la cooperazione etnica possa essere il risultato di un qualche sistema di eredità culturale in cui il gruppo, piuttosto che l'individuo, è oggetto di selezione. La selezione a livello di gruppo culturale è stata proposta per spiegare perché gli uomini, a differenza di tutti gli altri animali, cooperino, spesso contro i propri stessI mteressi, con individui con cui non condividono geni (Boyd e Richerson, 1991).
Bibliografia citata
AXELROD, R. (1984) The evolution of cooperation. New York, Basic Books.
BOYD, R., RICHERSON, P.J. (1991) Culture and cooperation. In Cooperation and prosocial behaviour, a c. di Hinde R.A., Groebel J., New York-Cambridge, Cambridge University Press, pp. 27-48.
BULL, J.J., MOLINEAUX, I.L., RICE, W.R. (1991) Selection of benevolence in a host-parasite system. Evolution, 45, 875-882.
BULL, J., RICE, W.R. (1991) Distinguishing mechanisms for the evolution of cooperation. J. Theor. Biol., 149, 63-74.
CONNOR, R.C., SMOLKER, R.A., RICHARDS, A.F. (1992) Two levels of alliance formation among male bottlenose dolphins (Tursiops truncatus). Proc. Natl. Acad. Sci. USA, 89, 987-990.
CREEL, S., CREEL, N.M. (1995) Communal hunting and pack size in African wild dogs, Lycaon pictus. Anim. Behav., 50, 1325-1339.
DE VRIES, P. (1992) Singing caterpillars, ants and symbiosis. Sci. Am., 267, 76-82.
DUGATKIN, L.A. (1997) Cooperation among animals: a modern perspective. New York, Oxford University Press.
EMLEN, S.T. (1995) An evolutionary theory of the family. Proc. Natl. Acad. Sci. USA, 92, 8092-8099.
FRANK, S .A. (1995) MutuaI policing and repression of competition in the evolution of cooperative groups. Nature, 377, 520-522.
FRANK, S.A. (1996) Host control of symbiont transmission: the separation of symbionts into germ and soma. Am. Nat., 148, 1113-1124.
FREAN, M.R. (1994) The prisoner's dilemma without synchrony. Proc. R. Soc. Lond. B, 257, 75-79.
HAMILTON, W.D. (1964) The genetic evolution of social behavior. J. Theor. Biol., 7, 1-51.
HEINSOHN, R., PACKER, C. (1995) Complex cooperative strategies in group-territorial African lions. Science, 269, 1260-1262.
MANNING, C.J., W AKELAND, E.K., POTTS, W.K. (1992) Communal nesting patterns in mice implicate MHC genes in kin recognition. Nature, 360, 581-583.
MAYNARD SMITH, J., SZATHMARY, E. (1995) The major transitions in evolution. San Francisco, W.H. Freeman.
NOE, R., HAMMERSTEIN, P. (1994) Biological markets: supply and demand determine the effect of partner choice in cooperation, mutualism and mating. Behav. Ecol. Sociobiol., 35, 1-11.
NOWAK, M., SIGMUND, K. (1993) A strategy of win-stay, lose shift that outperforms tit-for-tat in the Prisoner's Dilemma game. Nature, 364, 56-58.
PUSEY, A.E., PACKER, C. (1994) Non-offspring nursing in social camivores: minimizing the costs. Behav. Ecol., 5, 362-374.
RATNIEKS, F.L.W., VISSCHER, P.K. (1989) Worker policing in the honeybee. Nature, 342, 796-797.
TRIVERS, R.L. (1971) The evolution of reciprocal altruism. Q. Rev. Biol., 46, 35-57.
VEHRENCAMP, S.L. (1983) A model for the evolution of despotic versus egalitarian societies. Anim. Behav., 31, 667-682.
WADE, M.J. (1977) An experimental study of group selection. Evolution, 31, 134-153.
WEST EBERHARD, M.J. (1975) The evolution of social behavior by kin selection. Q. Rev. Biol., 50, 1-33.
WILKINSON, G.S. (1984) Reciprocal food sharing in vampire bats. Nature, 309, 181-184.
WILLIAMS, G.C. (1966) Adaptation and natural selection. Princeton, Princeton University Press.
WILSON, D.S., SOBER, E. (1994) Reintroducing group selection to the human behavioral sciences. Behav. Brain Sci., 17, 585-654.
Bibliografia generale
AXELROD, R. The evolution of cooperation. New York, Basic Books, 1984.
DUGATKIN, L.A. Cooperation among animals: a modern perspective. New York, Oxford University Press, 1997. MAYNARD SMITH, J., SZATHMARY, E. The major transitions in evolution. San Francisco, W.H. Freeman, 1995.