corteccia cerebrale
corteccia cerebrale
La corteccia cerebrale è considerata la struttura più evoluta e complessa tra tutti i sistemi viventi: si stima che nell’uomo la superficie corticale sia ca. 2.600 cm2, i neuroni ca. 30 miliardi, le sinapsi ca. 1015 e addirittura 10 elevato a 1 milione i circuiti che si potrebbero formare. È all’enorme complessità di questa struttura che si devono le nostre capacità sensoriali, motorie e percettive, la memoria e le funzioni che chiamiamo superiori (linguaggio, coscienza, capacità logica, previsione delle conseguenze delle azioni, creatività e così via), capacità e funzioni che permettono la miglior adattabilità della nostra specie all’ambiente e rappresentano la base della cultura dell’uomo. Qui vengono descritte dapprima le caratteristiche anatomiche e citologiche della corteccia; successivamente viene presentata una breve sintesi delle sue principali connessioni afferenti ed efferenti e della sua organizzazione intrinseca; infine vengono discussi i principali meccanismi attraverso i quali la corteccia è capace di integrare le informazioni. [➔ cervello, struttura e funzione del; connessionismo; corteccia motoria; funzioni cerebrali superiori; neurone; neurotrasmettitori; sensazione e percezione] La c. c. include strutture tra loro diverse per caratteristiche morfofunzionali e filogenetiche: archicorteccia, paleocorteccia (che insieme formano l’allocorteccia) e neocorteccia (o isocorteccia). L’archicorteccia, costituita dall’ippocampo, dal giro dentato e dal subicolo, presenta 3÷4 strati, si ritrova già negli anfibi ed è una forma primitiva di corteccia. La paleocorteccia comprende il trigono olfattivo, le circonvoluzioni olfattorie, la c. c. piriforme e presenta 3÷4 strati. La neocorteccia compare nei mammiferi e ha un’estensione imponente nei primati e soprattutto nell’uomo; l’aumento del volume del cervello che si è verificato nell’evoluzione dei mammiferi è determinato soprattutto dall’aumento della neocorteccia. Quest’aumento ha determinato una caratteristica fondamentale della neocorteccia nei primati e soprattutto nell’uomo: la girificazione, un meccanismo evolutivo che ha permesso di aumentare l’estensione della superficie corticale senza aumentare il volume cerebrale. È così imponente lo sviluppo della neocorteccia nei primati che il termine è diventato sinonimo stesso di corteccia. Qui ci riferiamo alla neocorteccia e tutti i termini relativi alla c. c. si riferiscono alla neocorteccia.
Laminazione e architettonica
La c. c. è caratterizzata dalla presenza di sei strati: I strato, ricco di fibre, con poche cellule; II strato, costituito da cellule piccole e addensate, alcune piramidali, altre rotondeggianti o stellate; III strato, per lo più con cellule piramidali di medie dimensioni; IV strato, formato da cellule piccole e fittamente addensate; V strato, costituito da cellule piramidali di medie o grosse dimensioni con ampie arborizzazioni dendritiche basali e dendriti apicali Esiche ascendono verso il I strato; VI strato, con cellule di varie forme. Esistono differenze nella struttura della c. c.: regioni diverse hanno spessore ed estensione degli strati diversi e variano per densità e tipi di cellule. Sulla base di queste differenze, si distinguono numerose (40÷107) aree citoarchitettoniche, raggruppabili in cinque tipi: c. c. agranulare, c. c. di tipo frontale, c. c. di tipo parietale, c. c. di tipo polare, c. c. granulare (o koniocorteccia). I tipi corticali frontale, parietale e polare presentano i sei tipici strati e sono definiti c. c. omotipica. I tipi agranulare e granulare costituiscono le c. c. eterotipiche, nelle quali non sono distinguibili sei strati. La c. c. agranulare è caratterizzata dall’assenza (o dalla scarsissima rilevanza) degli strati II e IV e da cospicui strati III e V ed è tipica delle c. c. motorie. La c. c. granulare è caratterizzata dalla ricchezza di cellule rotondeggianti, da strati II e IV ben sviluppati e da strati III e V scarsamente sviluppati; queste c. sono caratteristiche delle aree sensoriali: l’esempio paradigmatico di c. c. granulare è l’area 17, in cui il IV strato è distinto in tre sottostrati, uno dei quali, il IVb, è così sviluppato da essere visibile a occhio nudo (stria di Gennari).
Tipi cellulari
I neuroni corticali sono distinti in due famiglie: piramidali e non piramidali. I neuroni piramidali costituiscono circa i 3/4 dei neuroni corticali e sono localizzati negli strati II÷III e V÷VI; possiedono un corpo triangolare a base inferiore, un dendrite apicale di lunghezza variabile che spesso raggiunge gli strati superficiali, e dendriti basali a sviluppo orizzontale. I dendriti dei neuroni piramidali possiedono numerose spine che rappresentano siti di contatti sinaptici. I neuroni piramidali rispondono a brevi stimoli depolarizzanti generando potenziali d’azione di durata compresa tra 0,5 e 1 ms. Gli assoni dei neuroni piramidali rappresentano il canale efferente della c. c.; prima di uscire dalla sostanza grigia emettono collaterali che proiettano ad altri strati e a regioni corticali relativamente lontane (connessioni orizzontali), che rivestono una non trascurabile importanza nell’elaborazione delle informazioni. I terminali dei neuroni piramidali formano sinapsi asimmetriche e utilizzano il glutammato come neurotrasmettitore, esercitando un’azione eccitatoria sugli elementi postsinaptici. I neuroni non piramidali si suddividono in due gruppi, quelli provvisti di spine (cellule stellate spinose) e quelli che ne sono sprovvisti (neuroni non piramidali senza spine). Le cellule stellate spinose hanno assoni che terminano nelle vicinanze del corpo e sono pertanto interneuroni (➔); per il resto, presentano le caratteristiche dei neuroni piramidali. I neuroni non piramidali senza spine rappresentano il 20÷30% dei neuroni corticali e costituiscono un gruppo eterogeneo di cellule che condivide alcune fondamentali caratteristiche: i dendriti sono praticamente sprovvisti di spine; salvo rarissime eccezioni, gli assoni non escono dalla sostanza grigia, formano sinapsi simmetriche e liberano GABA come neurotrasmettitore. I neuroni non piramida li sono perciò interneuroni inibitori; in molti casi producono e liberano anche un neuropeptide (➔ cotrasmettitore) e rispondono a brevi stimoli depolarizzanti con treni di potenziali d’azione di durata variabile tra 0,2 e 0,3 ms.
Connessioni della corteccia
Ogni area della c. c. comunica con altre regioni del cervello per mezzo di connessioni afferenti ed efferenti.
Afferenze corticali. Alla c. c. giungono afferenze dal talamo e da strutture sottocorticali; inoltre, la c. c. riceve proiezioni da altre regioni corticali. Le afferenze talamo-corticali sono la principale sorgente di informazioni sottocorticali. I nuclei talamici più rilevanti sono i nuclei specifici, che proiettano densamente a un’area, e i nuclei intralaminari, che proiettano diffusamente a varie aree. Le proiezioni dai nuclei specifici terminano soprattutto nel IV strato; gli assoni talamo-corticali formano sinapsi asimmetriche prevalentemente asso-spinose e utilizzano il glutammato. Le proiezioni dai nuclei intralaminari terminano nel I strato e in parte negli strati III e V÷VI. Le afferenze sottocorticali extratalamiche originano da varie strutture: i nuclei del rafe danno origine a dense proiezioni dirette alla c. c. e la maggior parte di questi neuroni libera serotonina; il locus coeruleus contiene poche migliaia di neuroni noradrenergici che proiettano densamente alla c. c.; dal nucleo basale di Meynert, il più importante dei gruppi cellulari del telencefalo basale, emergono due fasci di fibre colinergiche che innervano la c. c.; l’area tegmentale ventrale dà origine a fibre che innervano la c. c. e liberano dopammina; il nucleo tuberomammillare dà origine a fibre che innervano la c. c. e liberano istamina. Le proiezioni extratalamiche condividono tra loro alcune caratteristiche: limitato numero di neuroni d’origine, estesa innervazione di ampie regioni ed elevata frequenza di sinapsi non convenzionali. Gran parte dei loro effetti sull’eccitabilità dei neuroni è mediata dall’azione su canali dello ione potassio; il loro effetto suggerisce che essi modulino i processi intracorticali e quindi le efferenze corticali, soprattutto in risposta a stimoli che rivestono un significato fisiologico primario.
Efferenze corticali. I neuroni d’origine delle proiezioni efferenti dalla c. c. sono quasi esclusivamente neuroni piramidali; sono localizzati negli strati II÷III e V÷VI, ma la loro distribuzione non è casuale. Esiste infatti una segregazione laminare dei neuroni efferenti in relazione alle strutture bersaglio. I neuroni del II strato proiettano soprattutto alle regioni corticali dello stesso lato; quelli del III danno origine a proiezioni associative e callosali (verso il corpo calloso); i neuroni del V strato proiettano al midollo, al ponte, al bulbo, ai collicoli, al nucleo rosso e al corpo striato; quelli del VI strato proiettano soprattutto al talamo e al claustro. Alcuni neuroni infragranulari proiettano anche ad altre aree della c. c. e alcuni neuroni del V strato proiettano al talamo (nuclei intralaminari). Il neurotrasmettitore utilizzato dalle fibre corticifughe è il glutammato.
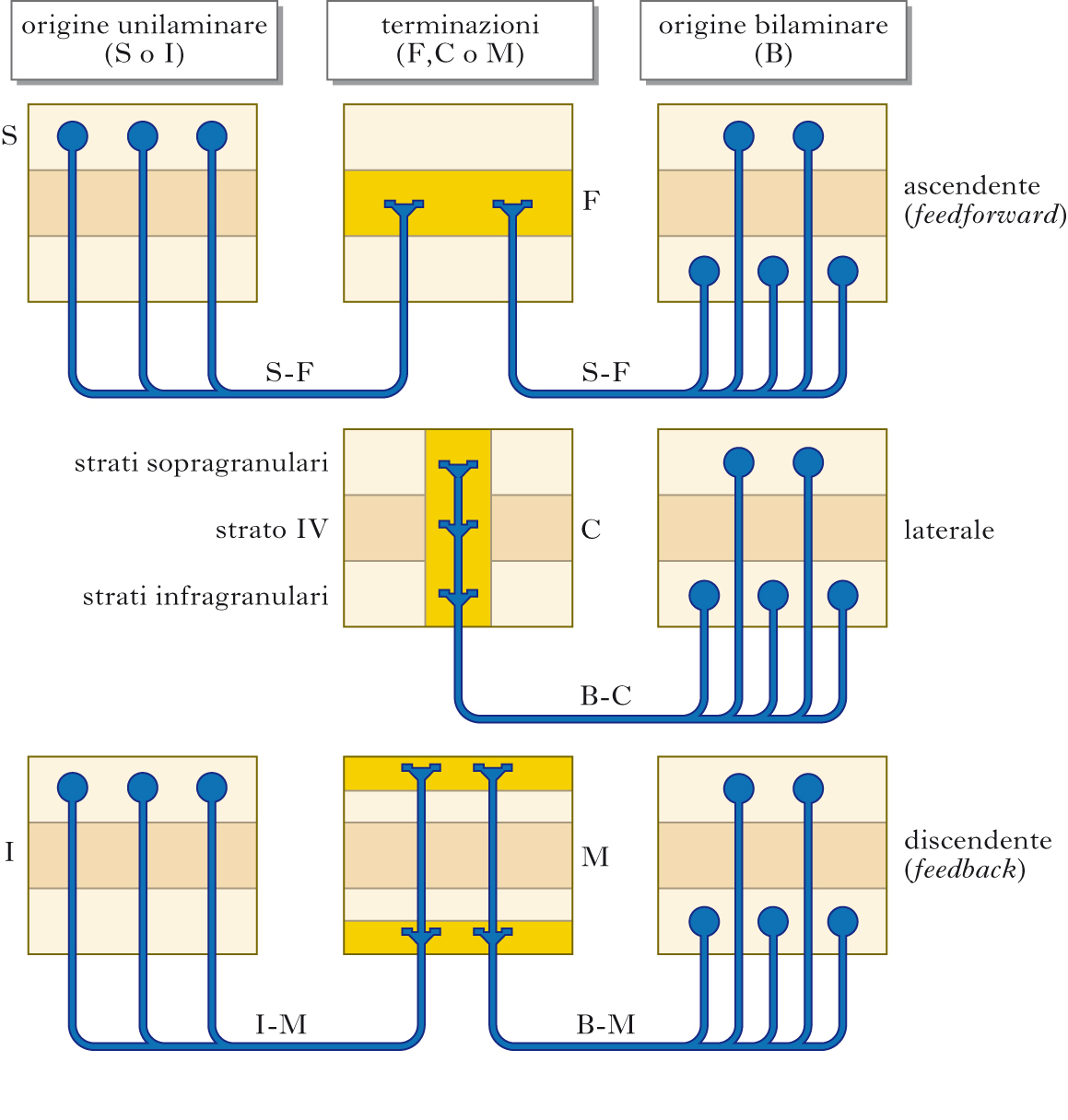
Connessioni corticocorticali. Tutte le aree della c. c. comunicano con altre aree, sia dello stesso emisfero (connessioni associative) sia dell’emisfero controlaterale (connessioni callosali). Salvo rarissime eccezioni, i neuroni d’origine delle proiezioni corticocorticali sono piramidali, i loro assoni formano sinapsi asimmetriche e liberano glutammato. La maggior parte delle proiezioni corticocorticali è reciproca. Esistono diversi tipi di proiezioni corticocorticali, chiamate F, M e C. Nelle proiezioni di tipo F i terminali assonici si distribuiscono nel IV strato dell’area ricevente e in parte nel III; i neuroni d’origine sono localizzati prevalentemente negli strati II÷III, anche se a volte hanno una distribuzione bilaminare, sopra- e infragranulare. Nelle proiezioni di tipo M i terminali si distribuiscono agli strati I÷III e V÷VI, evitando il IV; i neuroni d’origine sono localizzati negli strati V÷VI (a volte negli strati II÷III e V÷VI). Le proiezioni di tipo C sono caratterizzate dal fatto che i terminali si distribuiscono a tutti gli strati della c. in maniera colonnare; i neuroni d’origine si trovano negli strati II÷III e V÷VI.

Organizzazione intrinseca della corteccia
Organizzazione colonnare. Un microelettrodo fatto scendere verticalmente nella c. somestesica (l’area corticale deputata all’elaborazione delle informazioni originate dal corpo) prima registra l’attività elettrica di neuroni che hanno lo stesso campo recettivo e rispondono alla stessa submodalità. Questo fenomeno si ripete con una periodicità di circa 300÷500 μm. Vernon B. Mountcastle ipotizzò perciò nel 1957 che la c. c. fosse costituita da una serie di unità operative elementari modulari, che definì colonne. Le osservazioni di Mountcastle sono state confermate in tutti i tipi di c. c. esaminati e in diverse condizioni sperimentali. Un notevole contributo alla definizione del concetto di organizzazione colonnare è stato fornito dagli studi neuroanatomici. Dapprima, David H. Hubel e Torsten Wiesel hanno dimostrato che le fibre talamocorticali sono organizzate in fascicoli funzionalmente omogenei che terminano in maniera colonnare; successivamente, l’organizzazione colonnare è stata confermata nelle proiezioni corticocorticali e mediante studi funzionali. Anche lo studio degli eventi che si verificano durante lo sviluppo della c. c. ha fornito importanti dati in favore dell’organizzazione colonnare. L’ipotesi colonnare postula che l’unità elementare della c. sia la minicolonna, una stretta catena di neuroni che si estende verticalmente attraverso gli strati II÷VI; ogni minicolonna contiene ca. 80÷100 neuroni (con l’eccezione dell’area 17, in cui il numero è maggiore) ed è prodotta dalla divisione iterativa di un piccolo gruppo di cellule progenitrici del neuroepitelio. In ogni colonna sono presenti i principali tipi cellulari, densamente connessi lungo l’asse verticale. Secondo quest’ipotesi una minicolonna rappresenta la più piccola unità di elaborazione della corteccia. Questa versione dell’ipotesi colonnare è leggermente diversa da quella originaria, se non altro per le dimensioni delle colonne. Questa considerazione offre lo spunto per sottolineare che l’organizzazione colonnare basata sulle minicolonne è ancora un’ipotesi, anche se molto probabile e di grande valenza euristica. Il largo seguito che essa ha avuto è da spiegare con il fatto che risulta utile sia in chiave filogenetica, per spiegare l’aumento dello sviluppo della c. c. e delle capacità cerebrali, sia in termini operativi, proponendo un modello semplificato che si presta a studiare il flusso di informazioni nella corteccia. Letture rigide e schematiche trascurano tuttavia innegabili specificità di area e di specie, che possono sottendere importanti differenze funzionali.
Circuito canonico. È possibile individuare il circuito sinaptico essenziale all’elaborazione dell’informazione in una minicolonna? Anche se provvisoriamente, si può affermare che un circuito canonico è costituito da tre popolazioni di neuroni che interagiscono tra loro: una è rappresentata da interneuroni inibitori GABAergici, le altre due (una negli strati II÷III e una negli strati V÷VI) da neuroni eccitatori. Le cellule non piramidali provviste di spine sono omologate ai neuroni eccitatori degli strati II e III. Le principali connessioni interlaminari sono quelle esistenti tra i neuroni eccitatori degli strati superficiali e profondi; viceversa, i neuroni inibitori esercitano la loro azione su entrambe le popolazioni di neuroni eccitatori. I tre gruppi di neuroni ricevono input diretti dalle fibre talamiche, ma poiché l’afferenza talamica rende ragione solo del 10% dell’input eccitatorio, il 90% dell’eccitazione è generato dalle connessioni intracorticali che legano i neuroni piramidali. Questa fortissima eccitazione ricorrente è necessaria per fornire la corrente sinaptica indispensabile al mantenimento di una ragionevole frequenza di scarica. L’inibizione, che previene l’eccitazione ricorrente, è così potente da essere efficace anche quando è relativamente debole. La più nota modalità di flusso delle informazioni in un circuito intracorticale è la seguente: l’arrivo di potenziali talamici attiva i neuroni del IV strato, questi i neuroni degli strati II e III che, a loro volta, attivano quelli degli strati V÷VI; questi ultimi riattivano poi gli strati superiori.
Integrazione corticale: organizzazione gerarchica e sistemi distribuiti
Uno dei problemi della fisiologia della c. c. è quello di cercare di comprendere se una data funzione, per es. la percezione, dipenda dalla convergenza d’informazioni originate da diversi canali sensoriali attraverso specifiche vie o se sia il risultato dell’interazione tra diverse regioni corticali. Risposte a questa domanda possono essere cercate riprendendo una distinzione fondamentale delle aree corticali e studiando la funzione delle connessioni corticocorticali.
Aree associative. La distinzione tra aree sensorimotorie e associative e il concetto che le seconde svolgano un ruolo di particolare rilievo nella genesi delle funzioni superiori risale alla seconda metà del 19° secolo. Lo studio della localizzazione delle funzioni corticali aveva evidenziato che mentre la stimolazione elettrica o la lesione di alcune regioni della c. c. era associata a movimenti o a sintomi neurologici (paralisi, cecità o altri disturbi sensoriali), la stimolazione o la lesione di altre regioni non era accompagnata da questi fenomeni. Queste regioni, definite ineccitabili o latenti, furono meglio definite negli anni Venti del 20° sec. da Paul E. Flechsig, che studiò lo sviluppo della mielinizzazione nella c. c. umana. Nacque così la distinzione tra centri sensoriali e motori (nei quali prevalgono le fibre corticifughe non dirette alla c. c.) che mielinizzano prima, e centri associativi (nei quali prevalgono le fibre corticocorticali) che mielinizzano tardivamente. A rafforzare la validità di queste osservazioni, le zone associative risultavano comprendere tutta la superficie ineccitabile della corteccia. Da allora, quelle regioni della c. c. furono definite associative e considerate la sede delle funzioni nervose superiori (➔ funzioni cerebrali superiori). Le aree associative possono essere divise in due grandi famiglie: unimodali e multimodali. Le prime sono suddivise in vari sottotipi, in relazione al grado di dipendenza delle caratteristiche funzionali dei loro neuroni dalle afferenze che giungono dalle aree di livello inferiore. Le aree multimodali invece sono regioni situate all’interfaccia tra campi corticali costituiti da aree unimodali; sono sostanzialmente rappresentate dalla regione associativa posteriore (parti dei lobi parietale, occipitale e temporale), dalla regione associativa frontale (in pratica la c. c. prefrontale) e dalla regione associativa limbica (soprattutto la c. c. paraippocampale). Il concetto sotteso a questo tipo d’organizzazione è che esista una sequenza di connessioni che dalle aree sensoriali porta alle aree unimodali di livello inferiore e, successivamente, a quelle di livello superiore, e che nel passaggio da un livello all’altro si verifichi una convergenza di afferenze da canali paralleli e quindi un aumento della complessità delle caratteristiche di scarica dei neuroni. Le aree unimodali di livello superiore convergono nelle aree polimodali, nelle quali si verificherebbe l’integrazione delle informazioni afferenti; queste aree infine proiettano sia alle strutture profonde sia alle stesse aree dalle quali hanno ricevuto informazioni. Numerosi studi, a partire da quelli pioneristici di Edward G. Jones e Thomas Philip S. Powell negli anni Sessanta del 20° sec., hanno documentato che è possibile seguire la progressione sequenziale delle connessioni dalle aree sensoriali fino alle aree multimodali, suggerendo che esistano regioni nelle quali si verifica un elevato livello d’integrazione delle afferenze. Poiché si ritiene che un’area di livello inferiore fornisca le informazioni necessarie all’elaborazione che si verifica in un’area di livello superiore, le proiezioni di tipo F, che vanno da un’area di livello inferiore a una di livello superiore, vengono anche dette ascendenti (feedforward); le proiezioni di tipo M, che vanno da un’area gerarchicamente superiore a una inferiore sono dette discendenti (feedback); le proiezioni di tipo C non sono inquadrabili nello schema gerarchico e vengono dette laterali.
Integrazione corticale. Lo studio dell’organizzazione delle proiezioni corticocorticali ha rafforzato l’idea che esista una gerarchia tra aree della c., ma non dimostra che questo sia in essa l’unico principio operante; i risultati che sostengono l’ipotesi gerarchica sono stati ottenuti in regioni che mediano i primi stadi dell’elaborazione corticale. Viceversa, le connessioni esistenti tra aree meno vicine alle afferenze sensoriali assomigliano spesso a quelle laterali, formando una complessa rete di comunicazione bidirezionale nella quale le gerarchie sono sfumate o scomparse. L’integrazione corticale dipende quindi da meccanismi diversi che operano in regioni e in tempi diversi. Esiste infatti un livello relativamente semplice di elaborazione nel quale è possibile dimostrare l’esistenza di un’organizzazione gerarchica del flusso d’informazioni e nel quale le connessioni corticocorticali esibiscono chiare differenze (feedforward o feedback); questo livello include le aree sensoriali e parte delle aree associative unimodali. Esiste tuttavia un altro livello d’elaborazione che include le aree associative multimodali e una parte di quelle unimodali per le quali non è possibile dimostrare un’organizzazione gerarchica e in cui le connessioni corticocorticali non sono funzionalmente polarizzate. La complessa rete di comunicazione bidirezionale che ne risulta sembra essere il substrato del secondo, e forse più potente, meccanismo d’integrazione: è l’attivazione – totale o parziale, più o meno temporalmente coerente e variamente modulata dalle afferenze extralamiche – di questi sistemi distribuiti in tutta la c. c. che determina la generazione di nuove proprietà (proprietà emergenti) che rappresentano ben più della somma delle proprietà dei loro elementi costituenti. Fiorenzo Conti