
depressione a lungo termine
depressione a lungo termine
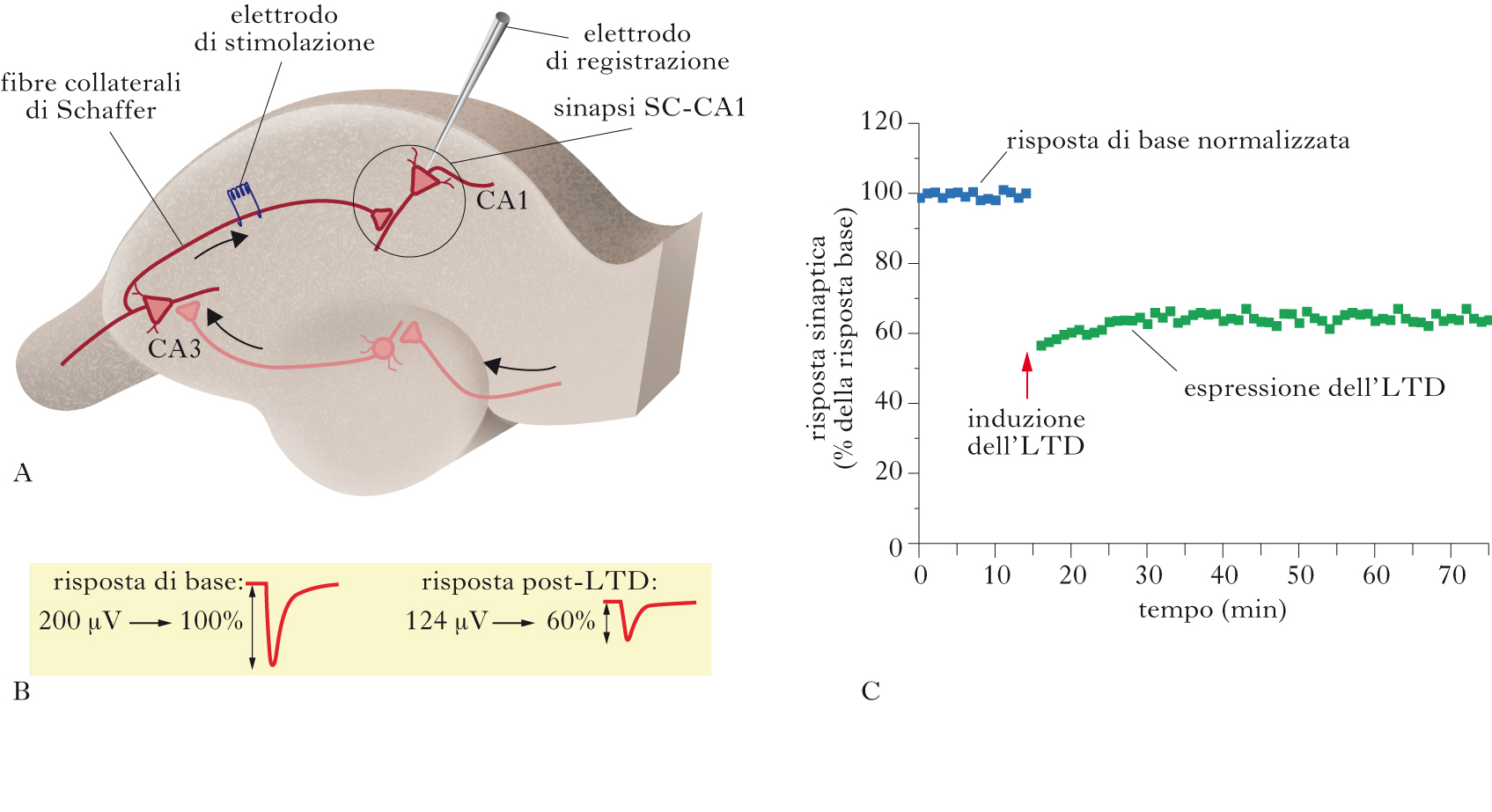
Fenomeno di plasticità sinaptica (in sigla LTD, dall’ingl. Long Term Depression) che consiste nella diminuzione dell’efficacia di una sinapsi come conseguenza di un particolare tipo di stimolazione o di attività endogena. Essa è complementare al potenziamento a lungo termine (➔) (LTP, Long Term Potentiation) nel regolare l’efficacia delle sinapsi. In condizioni stabili la risposta di una sinapsi sottoposta a una serie di stimoli costanti, di bassa intensità e sufficientemente distanziati fra loro (stimoli di controllo) non varia significativamente (risposta di base). Quando invece la sinapsi è sottoposta a un determinato tipo di stimolazione (induzione), al suo interno avvengono modificazioni tali (espressione) che, successivamente, la risposta allo stimolo di controllo sarà ridotta rispetto alla risposta di base. La riduzione è duratura nel tempo (dell’ordine di un’ora, quando le registrazioni avvengono in vitro, o giorni o settimane se si osserva in vivo) e si distingue da fenomeni di plasticità a breve termine (che agiscono su scale di millisecondi fino a minuti) e da una generica degradazione del segnale (run-down).
Induzione ed espressione
L’LTD è stata studiata soprattutto a livello delle sinapsi eccitatorie, dove presenta modalità di induzione ed espressione diverse a seconda dell’area del cervello. L’LTD è definita omosinaptica se segue la regola di Hebb, ossia se, durante l’induzione, i terminali pre- e postsinaptici sono attivati contemporaneamente. L’LTD eterosinaptica è invece osservata quando all’interno di uno stesso neurone una sinapsi viene stimolata con un protocollo di potenziamento di tipo ltp e, contemporaneamente, si osserva depressione a una sinapsi adiacente non stimolata. Per indurre LTD omosinaptica, per es. alle sinapsi Schaffer Collateral (SC)-CA1 dell’ippocampo (un prototipo per lo studio della plasticità), viene generalmente utilizzato uno stimolo elettrico a bassa frequenza (1 Hz) per un tempo prolungato (5÷15 minuti). Tale treno di stimoli provoca l’apertura dei canali NMDA (➔), permettendo il passaggio di ioni Ca2+. L’aumento di Ca2+ intracellulare è fondamentale per l’induzione di LTD (così come lo è per l’LTP), anche se rimane da determinare la sua esatta quantificazione al fine di indurre il fenomeno. Il segnale di Ca2+ agisce poi su una cascata biochimica che attiva proteine quali la calcineurina e la proteina-fosfatasi 1. L’aumento di Ca2+ può anche venire da altre fonti, come esempio per es. i canali al Ca2+ voltaggio-attivati, il reticolo endoplasmatico o l’attivazione di recettori metabotropici del glutammato mGluR1. Nel caso dell’ltd nmda-dipendente (alla sinapsi SC-CA1, ma non solo), il sito di espressione della depressione è prevalentemente postsinaptico e comporta cambiamenti nelle proprietà dei recettori AMPA; per es., la defosforilazione del sito Ser-845 della subunità GluR1 diminuisce la probabilità di apertura del canale e quindi riduce la trasmissione mediata dal recettore AMPA. Un meccanismo alternativo è l’endocitosi degli stessi recettori, un evento mediato da variazioni dello stato di fosforilazione della subunità GluR2.Nei primi anni del Ventunesimo secolo è stato scoperto un diverso tipo di LTD, indotta dai neuromediatori endocannabinoidi ed espressa presinapticamente. È stata studiata nell’ippocampo, e trovata anche nel cervelletto, nello striato e nella corteccia. In questo caso l’influsso di Ca2+ postsinaptico provoca il rilascio di endocannabinoidi (➔), che diffondono verso il terminale presinaptico attivando i recettori CB1. L’attivazione di tali recettori provoca una inibizione prolungata del rilascio di glutammato e quindi una diminuzione della risposta sinaptica.
Rilevanza fisiologica dell’LTD
Nonostante sia estremamente difficile dimostrare una relazione causale tra apprendimento e plasticità sinaptica (tra le difficoltà c’è il divario tra la bassa percentuale di sinapsi modificate da un evento di apprendimento e le misure di intere popolazioni di sinapsi fatte in vivo), esistono molti studi che ne evidenziano una forte correlazione. Nell’ippocampo (una delle strutture del cervello coinvolte nella memoria) esperimenti condotti su animali in vivo hanno dimostrato che l’LTD è facilitata durante l’esplorazione di un ambiente con oggetti nuovi o con una nuova configurazione spaziale (più che con un generico nuovo ambiente, correlato invece all’LTP). Analogamente lo stress comportamentale facilita l’induzione di LTD nell’ippocampo in vivo (mentre rende più difficile l’induzione di LTP), probabilmente attraverso l’azione di corticosteroidi. Un diverso tipo di apprendimento è quello motorio, controllato principalmente dal cervelletto. Qui, l’LTD è indotta dall’attivazione di recettori mGluRI e viene espressa, come nell’LTD NMDA-dipendente, tramite variazioni delle proprietà dei recettori AMPA. L’LTD è stata studiata in vivo nei riflessi condizionati, ma la sua rilevanza in questo fenomeno è ancora oggi (2010) oggetto di dibattito. È stata invece ben caratterizzata l’LTD nella corteccia visiva in relazione alla dominanza oculare (➔ ambliopia) durante il periodo critico (➔). Se in tale periodo l’animale è sottoposto a visione monoculare, le risposte sinaptiche nella parte di corteccia visiva corrispondente all’occhio privato di vista risultano depresse con meccanismi simili all’LTD. Infine, è stato trovato che l’LTD è alterata in soggetti con malattie neurologiche quali la malattia di Alzheimer o la sindrome dell’X fragile.