
Endocrinologia
Endocrinologia
SOMMARIO: 1. Che cos'è l'endocrinologia. 2. Gli ormoni circolanti. 3. Funzioni regolate da ormoni. 4. Coordinazione e feedback. 5. La natura chimica degli ormoni: a) i peptidi; b) gli steroidi; c) i derivati degli amminoacidi. 6. Trasmissione del messaggio ormonale nella cellula bersaglio. 7. Malattie endocrine: a) stati di ipofunzione; b) stati di iperfunzione. 8. Altri ormoni. 9. Sostanze ormono-simili. 10. Le molecole ormono-simili della matrice cellulare. 11. I rapporti tra il sistema endocrino e altri sistemi. 12. La nomenclatura degli ormoni. 13. La ricerca e i suoi metodi in endocrinologia. □ Bibliografia.
1. Che cos'è l'endocrinologia
L'endocrinologia è la disciplina scientifica che si occupa degli ormoni (v. tab. I), sia circolanti sia agenti localmente, e dei loro ruoli in condizioni fisiologiche e patologiche. Essa condivide il suo campo di ricerca con molte altre discipline, tra cui l'anatomia, la biochimica, la biologia cellulare, la biologia dello sviluppo/embriologia, la genetica, la medicina, la biologia molecolare, la patologia, la fisiologia e le scienze veterinarie. Gli ormoni sono stati individuati e studiati inizialmente negli esseri umani e in altri vertebrati, ma recentemente è stato dimostrato che ormoni e messaggeri ormono-simili agiscono come regolatori anche negli Insetti, nei vermi, in altri animali pluricellulari, in piante pluricellulari e in microrganismi unicellulari, compreso il lievito. La presente trattazione riguarderà gli ormoni nell'uomo e negli organismi affini, ma va ricordato che tutte le discipline che rientrano nella biologia hanno elementi in comune con l'endocrinologia.
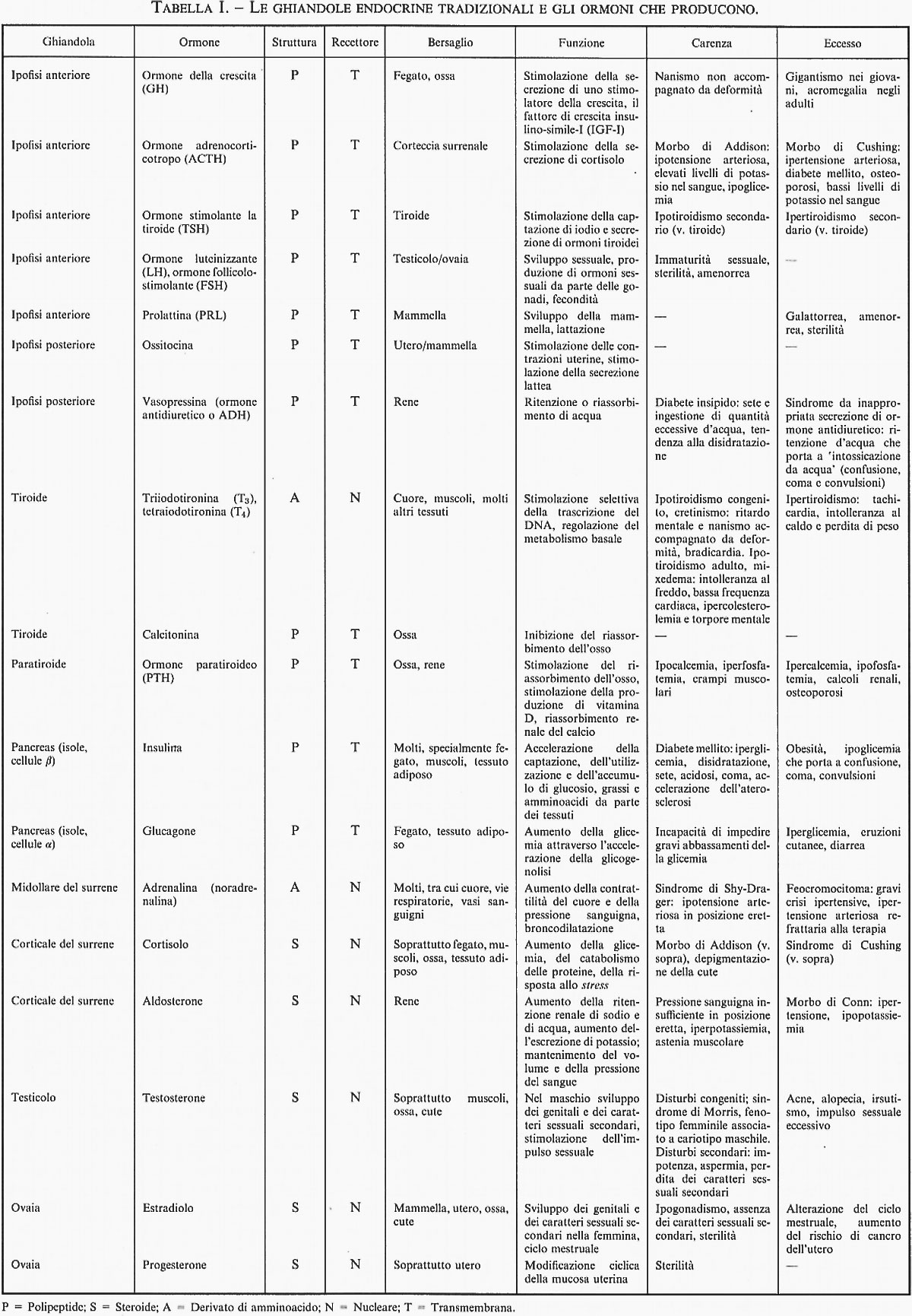
È attualmente noto che praticamente ogni cellula del corpo non solo costituisce una cellula bersaglio per uno o più ormoni o agenti ormono-simili, ma produce inoltre molecole che agiscono sulla cellula stessa, sulle cellule vicine o su cellule distanti come sostanze ormono-simili. La fig. 1 mostra la collocazione delle ghiandole endocrine propriamente dette; esse producono i ben noti ormoni ‛classici', le cui alterate funzioni, in difetto o in eccesso, sono responsabili di una serie di malattie specifiche: i disordini endocrini. L'ipofisi (o ghiandola pituitaria), che è situata immediatamente al di sotto del cervello col quale è collegata, è formata da elementi provenienti sia dall'orofaringe (lobo anteriore o adenoipofisi), sia dal cervello (lobo posteriore o neuroipofisi). La tiroide, che nasce nell'embrione alla base della lingua, durante lo sviluppo migra, attraversando la parte anteriore del collo, fino a raggiungere la sua posizione definitiva, davanti alla laringe. Le paratiroidi, che si sviluppano indipendentemente dalla tiroide, sono quattro strutture distinte, incorporate nella tiroide: in effetti, è necessaria una grande abilità chirurgica per rimuovere la tiroide senza rimuovere le paratiroidi. Le due ghiandole surrenali, una destra e una sinistra, sono organi di forma triangolare, distinti e separati dai reni, situati sulla loro sommità. Ciascuna di esse è costituita da una parte corticale esterna, che produce ormoni steroidei, e da una parte midollare interna, che produce catecolammine: adrenalina e noradrenalina. La parte corticale deriva dal mesoderma dell'embrione, mentre la midollare origina dalla cresta neurale, un precursore embrionale del sistema nervoso simpatico. I testicoli e le ovaie, oltre a produrre le cellule germinali (spermatozoi e oociti), hanno il compito di produrre gli ormoni sessuali, cioè gli androgeni, come il testosterone, e gli estrogeni, come l'estradiolo; nelle ovaie, inoltre, ha luogo la formazione e la regressione del corpo luteo, che secerne il progesterone. Il pancreas consiste di una parte esocrina e di una parte endocrina: la parte endocrina, che costituisce meno del 5% di tutto l'organo, è situata in piccoli ammassi di cellule, noti - dal nome del loro scopritore - come ‛isole di Langerhans' (isolotti pancreatici). I diversi tipi di cellule presenti nelle isole di Langerhans producono insulina (cellule β), glucagone (cellule α), somatostatina e polipeptide pancreatico. La descrizione appena fornita riguarda soltanto le maggiori ghiandole endocrine. Molti dei più importanti organi del corpo (il cervello, il cuore, il fegato, i reni, ecc.), pur svolgendo altre funzioni, contengono anche cellule endocrine. Analogamente, il sistema immunitario e il sangue, producendo cellule, producono anche ormoni e sostanze ormono-simili.
2. Gli ormoni circolanti
Ogni ormone circolante ‛classico' è una molecola unica, sintetizzata, immagazzinata e liberata da un unico tipo di cellula endocrina in una particolare ghiandola endocrina. La molecola di ormone, liberata nel circolo sanguigno dalla cellula in cui è stata sintetizzata, viene diffusa in tutto l'organismo entrando in contatto con gran parte delle sue cellule. Le cellule che rispondono a un determinato ormone sono dette ‛cellule bersaglio' (di quell'ormone). Alcuni ormoni hanno solo un tipo, o pochi tipi, di cellule bersaglio, mentre ad altri rispondono quasi tutti i tipi di cellule (il termine ‛ormone' - derivato dal greco ὁρμάω, che significa ‛suscito' o ‛eccito' - è stato introdotto da E. H. Starling nei primi anni del XX secolo). Il ruolo dell'ormone consiste nel far sì che il programma della cellula bersaglio cambi in modo da soddisfare meglio le necessità della stessa cellula bersaglio e di altre cellule, allo scopo di aiutare l'intero organismo ad agire in maniera coordinata per far fronte alle sfide dell'ambiente esterno (e interno).
In pratica ogni cellula del corpo è una cellula bersaglio per uno o più ormoni o agenti ormono-simili. La fig. 2 mostra come tutti gli ormoni peptidici, che costituiscono la stragrande maggioranza degli ormoni, e amminici - in particolare le catecolammine, come l'adrenalina e la noradrenalina - abbiano i propri recettori sulla superficie cellulare. Legandosi al recettore posto sulla superficie della cellula - cioè all'esterno - l'ormone genera un segnale all'interno della cellula. In molti casi il segnale intracellulare assume la forma di un secondo messaggero solubile (v. cap. 6), come il cAMP (adenosin-3′,5′'-monofosfato ciclico) o lo ione calcio o il diacilglicerolo (DAG). Il secondo messaggero propaga il segnale ormonale all'interno della cellula. Questo può portare a particolari eventi nel nucleo o a modificazioni di proteine nel citoplasma e a mutamenti dell'attività di svariati enzimi presenti nella cellula, che vengono attivati o inattivati. Contrariamente ai peptidi e alle catecolammine, i cui recettori sono situati sulla superficie cellulare, gli ormoni steroidei e tiroidei sono caratterizzati dal fatto di possedere recettori intracellulari. Gli ormoni di questi due gruppi attraversano facilmente la membrana cellulare ed entrano nel citoplasma, dove incontrano i loro recettori. Una volta occupato e attivato da un ormone, il recettore citoplasmatico passa nel nucleo e si lega al DNA. Legandosi al DNA in corrispondenza di diversi siti, il complesso ormone-recettore determina quali RNA messaggeri vadano maggiormente espressi e quali meno; il che, a sua volta, modifica la concentrazione intracellulare di molte proteine, tra cui gli enzimi.
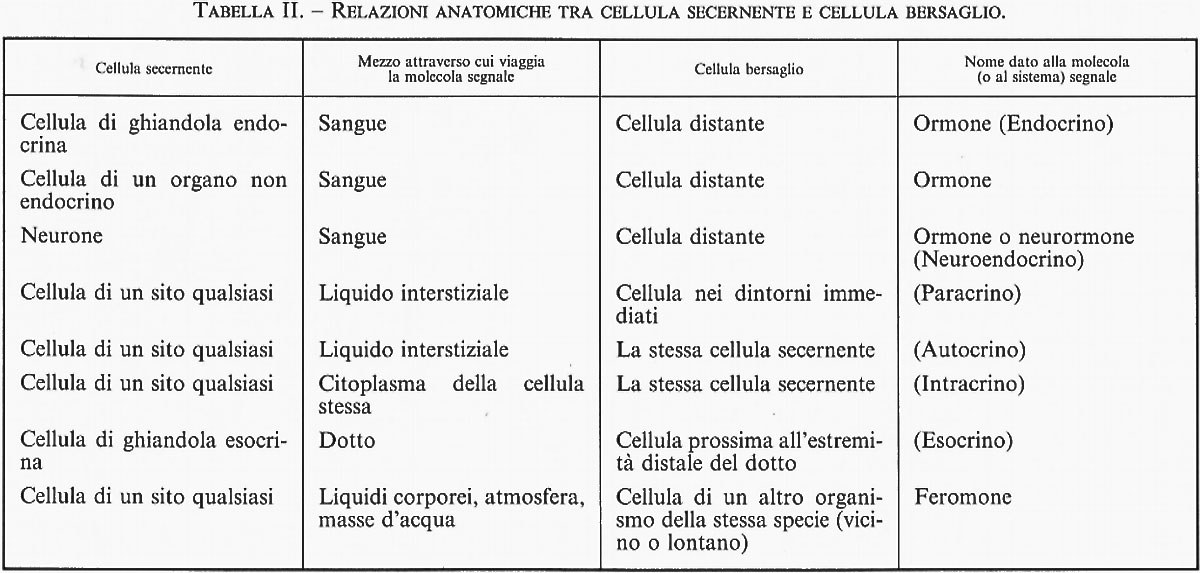
L'insieme costituito dalle ghiandole, dagli ormoni e dalle cellule bersaglio è complessivamente noto come ‛sistema endocrino'. Il termine ‛endocrino' deriva dal greco ἕνδον, ‛all'interno', e κρίνω, ‛secerno', e si riferisce alle ghiandole che liberano gli ormoni prodotti nella corrente sanguigna. Note un tempo come ‛ghiandole prive di dotti' (ductless), le ghiandole endocrine differiscono dalle ghiandole esocrine (come le ghiandole salivari o le ghiandole mammarie) per il fatto che queste ultime riversano i loro prodotti in appositi dotti escretori che sfociano all'esterno dell'organismo o in organi cavi comunicanti con l'esterno (ad es., tratto gastrointestinale).
La fig. 3 mostra e confronta lo sviluppo delle ghiandole esocrine e quello delle ghiandole endocrine. Le cellule epiteliali, che inizialmente formavano uno strato di cellule, cominciano a proliferare e a crescere nel tessuto connettivo sottostante (in alto, nella figura). Nel caso di una ghiandola esocrina (in basso a sinistra), le cellule situate più in profondità (più numerose) diventano le cellule secernenti, mentre quelle che si trovano più vicine alla superficie si trasformano nelle cellule di rivestimento del dotto secretore; le cellule secernenti della ghiandola esocrina riversano i propri secreti nel dotto e, attraverso questo, all'esterno. In basso a destra, nella figura, è illustrata la formazione di una ghiandola endocrina. Anche in questo caso le cellule situate più in profondità (più numerose) diventano cellule secernenti, ma a ridosso delle cellule ghiandolari si forma una fitta rete di capillari destinati a ricevere i prodotti secreti; le cellule che connettevano le cellule secernenti ghiandolari alla superficie scompaiono (mentre nella ghiandola esocrina vanno a formare il dotto secretore). Per una classificazione più generale, v. tab. II.
3. Funzioni regolate da ormoni
Tra i complessi processi organici la cui regolazione è strettamente influenzata dal sistema endocrino (v. tab. I) si annoverano: la riproduzione sessuale (e i fenomeni a essa strettamente collegati, cioè lo sviluppo e il comportamento sessuali, la fecondità, la gravidanza e la lattazione); la crescita e lo sviluppo del feto e del bambino; il mantenimento della composizione biochimica dell'organismo (e in particolare dei livelli dell'acqua, del sodio, del potassio, del calcio, del fosfato e del glucosio); la ‛gestione' del cibo (che comprende la regolazione dell'appetito e del senso di sazietà, la regolazione del movimento del cibo e della sua digestione nel tratto gastrointestinale e il trattamento dei prodotti della digestione). In pratica, ogni cellula del corpo è o è stata soggetta all'azione degli ormoni. Nella sua attività di regolazione e coordinamento delle funzioni corporee il sistema endocrino condivide svariati compiti con il sistema nervoso e con altre unità regolatrici locali.
Dopo aver illustrato il sistema endocrino, verrà mostrato come esso si sovrapponga al sistema nervoso e alle menzionate unità regolatrici locali, sia dal punto di vista funzionale sia sotto il profilo anatomico.
4. Coordinazione e feedback
Spesso parecchi ormoni agiscono in sintonia su svariati tessuti al fine di regolare un processo. Per esempio, quando ci si alza, al mattino, parecchie ore dopo aver mangiato, il livello del glucosio nel sangue risulta normale. Per mantenere questo livello del glucosio, le cellule β del pancreas provvedono a secernere insulina a un livello minimo ma costante. Il livello basale di insulina circolante agisce sul fegato, sulle cellule adipose e sulle cellule muscolari. In risposta al basso livello di glucosio e di insulina, il fegato libera nel sangue dai propri depositi quantità controllate di glucosio; le cellule adipose scindono, a velocità controllata, il grasso accumulato trasformandolo in acidi grassi e glicerolo, sostanze ricche di energia; le cellule muscolari liberano acido lattico e alanina, che vengono trasportati dal sangue nel fegato dove sono convertiti entrambi in glucosio. Inoltre, le cellule muscolari (e quelle adipose) usano gli acidi grassi anziché il glucosio per i propri fabbisogni energetici. Mangiando, le sostanze nutritizie assunte per via alimentare innalzano il livello del glucosio nel sangue, e ciò (in sintonia con segnali emessi dal sistema nervoso e dagli ormoni liberati dalle cellule intestinali) stimola un forte aumento della secrezione di insulina da parte delle cellule β del pancreas. L'elevato livello raggiunto dall'insulina dopo un pasto agisce sul fegato e sulle cellule adipose e muscolari, provocando l'inversione dei processi descritti. Il fegato smette di liberare glucosio e incomincia ad accumularlo sotto forma di glicogeno. Le cellule adipose smettono di scindere il grasso e iniziano ad assumere acidi grassi e glucosio (da usare e da accumulare). Analogamente, le cellule muscolari smettono di liberare alanina e acido lattico e iniziano a consumare molto più glucosio che acidi grassi. L'effetto netto di questi processi è il ritorno a livelli normali di glucosio nel sangue; di pari passo la secrezione di insulina torna a livelli basali. Quando il glucosio scende sotto il livello normale, la midollare del surrene libera adrenalina che agisce sulle cellule β del pancreas, sul fegato, sulle cellule muscolari e adipose. La liberazione di insulina viene bloccata; vengono stimolate la scissione del glicogeno e la liberazione di glucosio da parte del fegato; viene promossa la scissione del grasso. Viene liberato anche il glucagone, un ormone secreto dalle cellule α degli isolotti pancreatici, che agisce in maniera coordinata con l'adrenalina. Questa relazione primaria tra il glucosio e l'insulina, nonché le relazioni secondarie che coinvolgono altre fonti di energia e altri ormoni, costituiscono un archetipo di sistema endocrino.
La fig. 4 illustra il ruolo del sistema ipotalamo-ipofisario nella regolazione di un processo fisiologico; la tab. III indica le associazioni specifiche costituite da un determinato ormone ipotalamico, il relativo ormone ipofisario, la ghiandola bersaglio dell'ormone ipofisario, l'ormone rilasciato, a sua volta, dalla ghiandola bersaglio e i bersagli di quest'ultimo ormone.

Il cervello, integrando segnali ormonali, nervosi e di altra natura, stimola determinate cellule dell'ipotalamo (la parte del cervello localizzata proprio al di sopra dell'ipofisi) a rilasciare un ormone, o fattore, in un sistema locale di vasi sanguigni che raggiunge l'ipofisi; qui l'ormone ipotalamico stimola particolari cellule del lobo anteriore dell'ipofisi (‛adenoipofisi') a rilasciare nella circolazione generale un altro ormone, che a sua volta stimola le cellule endocrine di un organo distante a rilasciare il proprio ormone nella circolazione generale; quest'ultimo ormone stimola svariate cellule in tutto il corpo, soddisfacendo, in tal modo, le necessità fisiologiche del sistema. L'ormone secreto dall'organo bersaglio agisce retroattivamente anche sulle cellule dell'ipofisi e dell'ipotalamo inibendo l'ulteriore liberazione degli ormoni stimolatori (v. fig. 4). L'espressione usata dagli endocrinologi per indicare il processo attraverso cui il prodotto (ormone) finale inibisce l'ulteriore secrezione degli ormoni stimolatori primari è ‛feedback negativo'. Ancora poco conosciuti sono i casi di ‛feedback positivo', in cui il prodotto (ormone) finale stimola la produzione degli ormoni primari. Un ulteriore esempio delle complesse relazioni di feedback tra varie ghiandole e molteplici ormoni è fornito dal normale ciclo mestruale.
Il ciclo mestruale costituisce un esempio eccellente di come un gruppo di ormoni interagisca per portare a compimento un processo biologico complesso (v. fig. 6). Iniziamo col descrivere la serie di eventi biologici che si verificano al livello delle ovaie e dell'utero durante il ciclo mestruale: durante le mestruazioni le cellule endometriali vengono eliminate perché possa aver inizio un nuovo ciclo; nell'ovaia, tra le molte cellule uovo pronte a maturare, una sola progredisce e forma attorno a sé una struttura follicolare (questa parte del ciclo mestruale è detta infatti ‛fase follicolare'); circa a metà del ciclo, il follicolo si rompe e l'uovo ormai maturo ne viene espulso e dall'ovaia passa nella tuba uterina, dove inizia il suo ‛viaggio' verso l'utero; dopo la rottura (deiscenza) il follicolo si trasforma in una struttura secernente endocrina nota come ‛corpo luteo', da cui il nome della seconda metà del ciclo mestruale, la fase luteinica. Questa successione di eventi biologici è guidata da variazioni della concentrazione di diversi ormoni. In prima approssimazione si noti che la prima metà del ciclo mestruale è dominata da livelli crescenti di LH e di estrogeno, mentre la seconda metà del ciclo è dominata da modesti livelli di estrogeno e da livelli molto alti di progesterone. Nella parte centrale della fig. 6 sono riportati i valori relativi ai due ormoni ipofisari, l'ormone follicolostimolante (o FSH) e l'ormone luteinizzante (o LH), noti complessivamente come ‛gonadotropine ipofisarie'. Si tenga presente che l'LH è soggetto a una crescita graduale costante durante la prima metà del ciclo mestruale, fino a raggiungere un picco proprio in prossimità dell'ovulazione, seguito da una rapida caduta ai valori iniziali. D'altra parte, la concentrazione di FSH, moderatamente elevata all'inizio del ciclo mestruale, subisce poi, nell'ordine, un calo graduale, un breve, rapido incremento, fino al raggiungimento di un picco in prossimità dell'ovulazione, e infine una rapida caduta a bassi livelli, dai quali non si scosta fino alla fine del ciclo. Entrambi questi ormoni sono sotto il controllo dell'ipotalamo e il loro bersaglio principale è l'ovaia. La parte centrale della fig. 6 mostra anche i livelli di concentrazione degli ormoni ovarici, la cui secrezione è regolata dalle gonadotropine ipofisarie. Il principale ormone ovarico, l'estrogeno E2β (così denominato in base alla sua formula chimica), è presente in concentrazione relativamente bassa fino alla seconda settimana del ciclo mestruale, quando subisce un rapido aumento fino a raggiungere, anch'esso, un picco in prossimità dell'ovulazione, seguito da un andamento altalenante costituito da un calo, un aumento e un nuovo calo, in successione. Il progesterone, l'altro principale ormone ovarico, subisce un calo progressivo durante la prima settimana del ciclo, seguito da una fase di bassa concentrazione che si protrae fin dopo l'ovulazione; durante le due ultime settimane del ciclo mestruale la concentrazione di progesterone si mantiene ad alti livelli.
5. La natura chimica degli ormoni
a) I peptidi.
In misura preponderante (oltre l'80%) gli ormoni sono oligopeptidi, oppure polipeptidi o piccole proteine, costituiti da sequenze di amminoacidi in numero variabile da alcune unità ad alcune centinaia. Gli ormoni più grandi (il TSH, l'FSH e l'LH dell'ipofisi e le gonadotropine della placenta) hanno pesi molecolari di alcune decine di migliaia di dalton, elevati per un ormone ma bassi se riferiti all'universo delle proteine. Questi ormoni consistono di due catene polipeptidiche, ciascuna delle quali si arricchisce di residui monosaccaridici (catene polisaccaridiche) legati direttamente, al momento della loro sintesi, agli amminoacidi che le compongono. Ciascuna delle due catene deriva da un gene diverso. Con l'eccezione di questi casi, negli ormoni peptidici l'aggiunta di residui monosaccaridici (glicosilazione) è un fatto raro. Se si esclude l'insulina, che ha due catene (entrambe derivate da un solo gene), gli altri ormoni, nella stragrande maggioranza dei casi, sono peptidi a catena singola (senza residui monosaccaridici) derivati da geni singoli. Nel corso della sua biosintesi l'ormone, come nel caso dell'insulina e della vasopressina, può essere prodotto inizialmente come parte di grandi molecole polipeptidiche precursori (chiamate ‛pre-pro-ormoni' e ‛pro-ormoni'), dalla quale viene poi ‛ritagliato' da appositi enzimi: questi lo distaccano dalle proteine più grandi e gli conferiscono la struttura definitiva. I geni che codificano per gli ormoni peptidici e il processo attraverso cui questi ormoni sono sintetizzati sono molto simili a quelli relativi a tutti gli altri peptidi e alle altre proteine.
Più interessante è il fatto che l'attività principale di una cellula secernente riguarda la produzione degli ormoni, e che l'ormone è il prodotto principale dell'apparato biosintetico della cellula. Per esempio, si pensa che l'insulina rappresenti il 10% di tutte le molecole proteiche sintetizzate dalle cellule β del pancreas. Stime simili valgono anche per quel che riguarda la produzione dell'ormone della crescita (GH) da parte delle cellule dell'ipofisi a ciò deputate. Le cellule che secernono ormoni, viste al microscopio, presentano molte caratteristiche strutturali in comune con altre cellule secernenti, sia endocrine sia esocrine, ovvero la capacità di sintetizzare, elaborare e accumulare in granuli grandi quantità di ormoni e di liberare rapidamente e in dosi elevate, quando ciò risulti necessario, tali granuli.
b) Gli steroidi.
Il secondo principale gruppo di ormoni è costituito dagli ormoni steroidei. Questi ormoni sono sintetizzati a partire dal colesterolo, proveniente in parte da fonti locali, ma principalmente da lipoproteine ricche di colesterolo presenti nel sangue. Nelle ghiandole la molecola di colesterolo viene modificata attraverso una serie di passaggi enzimatici altamente organizzati. In questa serie di eventi di biosintesi, un ormone può essere un precursore o pro-ormone del successivo. Per esempio, il progesterone, importante ormone dell'ovaia, è anche un precursore del testosterone; questo, a sua volta, oltre a essere il principale prodotto ormonale dei testicoli, è anche un precursore dell'estradiolo, il più importante ormone sessuale femminile. Di questi ormoni vengono accumulate solamente minime quantità: ciascuno di essi viene in genere sintetizzato su comando, in caso di necessità. Questi ormoni possono essere trasformati in siti extraghiandolari. Per esempio, il testosterone, prodotto in una ghiandola classica, può essere trasformato in estrogeno nelle cellule adipose dall'enzima aromatasi. L'1α,25-diidrossicolecalciferolo è un ormone di origine steroidea derivato dalla vitamina D3: questa viene trasformata nell'ormone attivo attraverso modificazioni chimiche che avvengono nella cute, nel fegato e nel rene.
I più importanti ormoni steroidei secreti dalla corticale del surrene negli esseri umani sono il cortisolo e l'aldosterone (il corticosterone nei Roditori).
c) I derivati degli amminoacidi.
Gli ormoni della midollare del surrene - l'adrenalina (epinefrina) e la noradrenalina (norepinefrina) - sono entrambi derivati, attraverso trasformazioni enzimatiche, dall'amminoacido tirosina. Gli ormoni della tiroide, la tiroxina (T4) e la triiodotironina (T3), sono anch'essi derivati dalla tirosina attraverso un processo che comporta la sintesi di una grande proteina (la tireoglobulina) e trasformazioni enzimatiche (iodurazione e accoppiamento). Il caso della tiroide è particolarmente interessante: la tireoglobulina funge sia da pro-ormone, liberando per scissione proteolitica la tiroxina, sia da scorta di ormone all'interno della ghiandola, nonché da deposito di iodio, componente necessario degli ormoni tiroidei; la tiroxina contribuisce anche alla formazione di triiodotironina al di fuori della tiroide.
6. Trasmissione del messaggio ormonale nella cellula bersaglio
La selettività, o specificità, è uno dei tratti caratteristici dei sistemi endocrini; un ormone attiva un preciso sottoinsieme di cellule e non ha effetto sulle altre. La molecola di ormone è soltanto una delle moltissime molecole (fino a dieci milioni) cui le cellule sono esposte. Una cellula che risponda a un dato ormone, cioè una cellula bersaglio di quell'ormone, possiede un recettore specifico che può riconoscerlo fra tutte le altre molecole presenti nel suo ambiente (v. figg. 2 e 10). Ogni ormone ha il proprio recettore: il recettore e l'ormone si riconoscono reciprocamente legandosi l'uno all'altro. I molteplici legami chimici che uniscono l'ormone al recettore determinano la specificità, cioè il fatto che soltanto l'ormone o una molecola a esso molto simile possono legarsi al recettore; qualsiasi altra sostanza risulta inerte in questo sistema e non si lega al recettore. La maggior parte delle centinaia di differenti recettori rientra in un ristretto numero di famiglie. Nella fig. 10 sono rappresentate schematicamente tre di queste famiglie. I recettori che appartengono a una stessa famiglia sono molto simili. Il primo gruppo, a sinistra dello schema, è costituito dalle proteina-tirosinachinasi. Quando un ormone si lega a questo tipo di recettore, una parte di quest'ultimo si attiva in forma di enzima inducendo la fosforilazione propria e di altre proteine: questa reazione di fosforilazione rappresenta l'attivazione del recettore. Un altro importante tipo di recettore (indicato con R nella fig. 10) è una proteina a catena singola che si snoda attraverso la membrana ripiegandosi su se stessa sette volte così da formare una tasca in grado di legare l'ormone, il quale può essere un peptide, una catecolammina, un derivato di una prostaglandina, ecc. Ogni ormone o molecola ormono-simile ha il proprio recettore specifico; i recettori R, quando vengono attivati, si legano a ‛proteine G', che, a loro volta, si attivano e attivano un enzima, E, il quale porta alla formazione di secondi messaggeri. Un'altra importante famiglia di recettori (a destra, nella fig. 10) è composta da proteine attraversate, per tutta la loro lunghezza - dall'esterno all'interno della cellula - da un canale che consente l'accesso, nella cellula, di particolari molecole, e specialmente di ioni. L'ormone, legandosi sulla parte esterna del recettore, modifica drasticamente il flusso degli ioni.
L'unione dell'ormone con il recettore non è altro (almeno all'inizio) che una reazione reversibile tra due molecole. La forza dell'unione tra l'ormone e il recettore, anch'essa un risultato di comuni legami chimici, ne determina la stabilità e genera la capacità dell'ormone di attivare la cellula bersaglio anche a basse concentrazioni. La forza, o stabilità, dell'unione è detta ‛affinità'; l'affinità e la specificità, entrambe dipendenti dalla molteplicità dei legami chimici tra l'ormone e il recettore, sono due proprietà fondamentali di ogni coppia ormone-recettore. Oltre a istituire il legame, l'unione delle due molecole deve portare all'attivazione della cellula bersaglio. L'unico contributo dell'ormone all'attivazione della cellula consiste nell'attivazione del proprio recettore; il recettore possiede nella propria struttura l'intero programma per l'attivazione della cellula. Ogni recettore possiede due siti fondamentali, uno per l'attacco dell'ormone, l'altro per l'attivazione della cellula. Il grado di attivazione di un recettore da parte di una molecola di ormone a esso legata si chiama ‛attività intrinseca' di quell'ormone. Un ormone che si lega al recettore e lo attiva completamente si chiama ‛agonista' (completo). Una sostanza ormono-simile che si lega al sito d'attacco dell'ormone o in prossimità di esso, ma non attiva il recettore, si chiama ‛antagonista' (completo), se - quando è presente - compete con l'agonista per legarsi al recettore e quindi ne riduce o elimina l'efficacia. Spesso gli scienziati introducono precise modifiche negli ormoni naturali per cambiarne la specificità, l'affinità o l'attività intrinseca nei confronti di un determinato recettore. Questi derivati sintetici, detti ‛analoghi', possono apportare molti vantaggi terapeutici rispetto agli ormoni naturali. L'attivazione del recettore è soltanto il primo di una serie di passaggi attivati dall'interazione ormone-recettore; i processi di rilevanza biologica regolati da questi accoppiamenti ormone-recettore (per esempio la regolazione dell'attività di enzimi lungo le vie metaboliche all'interno della cellula, o di trasportatori sulla superficie cellulare) possono avvenire a molti passaggi di distanza (v. fig. 2). Tra gli agenti biochimici che legano il recettore attivato ai suoi effettori vi sono i cosiddetti ‛secondi messaggeri' (il primo messaggero è l'ormone). I secondi messaggeri sono molecole a basso peso molecolare prodotte nel citoplasma in risposta all'attivazione dei recettori sulla superficie cellulare; queste molecole si diffondono rapidamente e agiscono in vario modo nella cellula attraverso numerosi passaggi successivi (v. fig. 2). Dato che esse sono sintetizzate e degradate rapidamente, le loro concentrazioni mutano velocemente in risposta a cambiamenti dei livelli dell'ormone e del recettore. L'AMP ciclico (cAMP) è il secondo messaggero più conosciuto - nonché il primo a essere stato identificato - ed è utilizzato dall'adrenalina e da molti ormoni peptidici. Esso viene sintetizzato a partire dall'ATP (adenosintrifosfato) intracellulare: il recettore attivato dall'ormone attiva l'adenilatociclasi, un enzima che converte un comune costituente cellulare, l'ATP, in una variante rara, l'AMP ciclico. L'AMP ciclico si lega a un enzima specifico, la proteina-chinasi dipendente dall'AMP ciclico, e lo attiva. Esso è presente solo temporaneamente, essendo rapidamente decomposto da specifici enzimi che lo convertono in AMP (adenosinmonofosfato). La proteina-chinasi prosegue la propria attività fosforilando gli amminoacidi serina e treonina in altre proteine, con ciò attivandole o inattivandole.
Il GMP (guanosinmonofosfato) ciclico, l'ossido d'azoto e lo ione calcio sono altri ben noti secondi messaggeri per ormoni e agenti ormono-simili. La fosforilazione e la defosforilazione di proteine, specialmente enzimi, costituiscono un altro tipo di evento biochimico intermedio attraverso cui i recettori, attivati dagli ormoni, inviano i loro messaggi agli effettori finali (sia l'adrenalina sia l'insulina attivano molte di queste reazioni intermedie entro la cellula). Molte proteine, in risposta all'attivazione di un recettore localizzato sulla superficie cellulare da parte di un ormone, vengono o fosforilate o defosforilate, e di conseguenza il loro stato di attività cambia. La fig. 12 mostra come il gruppo ossidrile della serina, della treonina o della tirosina appartenenti a una proteina possa acquistare uno ione fosfato. L'enzima che aggiunge lo ione fosfato si chiama ‛chinasi'; l'ATP è il donatore del fosfato. La reazione della chinasi che aggiunge lo ione fosfato è strettamente legata a quella di un altro enzima, una fosfatasi, che è capace di rimuovere quello ione e quindi di ripristinare lo stato originario di attività del substrato. L'effetto netto di questo ciclo è la conversione di ATP in ADP (adenosindifosfato), ma questo effetto è del tutto secondario rispetto all'importante cambiamento di attività che si verifica. Nella maggioranza dei casi la fosforilazione determina l'attivazione della proteina, la defosforilazione la sua inattivazione. Tuttavia, in alcune condizioni la forma fosforilata è inattiva e quella defosforilata è attiva. Le fosforilazioni della tirosina sono molto più rare e tendono a regolare ampi dominî dei processi metabolici, cioè in prossimità del recettore attivato, mentre le fosforilazioni della serina sono molto più comuni e generalmente avvengono in corrispondenza delle tappe conclusive delle vie metaboliche.
Gli ormoni steroidei (e gli ormoni tiroidei) attraversano facilmente la membrana cellulare e interagiscono con i propri recettori situati all'interno della cellula, generalmente nel citoplasma (v. fig. 2). Legato al proprio ormone, il recettore, attivato, penetra nel nucleo dove si lega al DNA, attivando o disattivando geni specifici, localizzati presso i siti di legame specifici per tale recettore, presenti nel DNA (v. fig. 2).
7. Malattie endocrine
a) Stati di ipofunzione.
Gli endocrinologi clinici sono medici specializzati nelle malattie del sistema endocrino. Un gran numero di disturbi è associato a una deficiente produzione di ormoni da parte delle ghiandole (v. tab. I), in conseguenza della quale le cellule bersaglio di tali ormoni vengono sottostimolate e quindi varie funzioni organiche risultano alterate, generalmente diminuite. La diagnosi è posta quando si dimostra che i livelli dell'ormone, misurati in un campione di sangue, a riposo o dopo una determinata stimolazione biologica, sono più bassi del normale. I pazienti affetti da questi disturbi spesso possono trarre giovamento dalla somministrazione di un sostituto dell'ormone, per via orale o parenterale. Gli ormoni peptidici, dal momento che vengono degradati dagli enzimi digestivi del tratto gastrointestinale, sono inefficaci quando vengono somministrati oralmente, ma possono essere somministrati con successo per iniezione o per inalazione.
Difetti congeniti, tumori, traumi fisici e infezioni sono alcune delle numerose cause delle carenze ormonali. La cessazione della secrezione di estrogeno da parte delle cellule dell'ovaia a ciò deputate costituisce uno stato di deficienza ormonale che si verifica normalmente all'epoca della menopausa. Un'altra causa comune di deficienza ormonale è l'autoimmunità, un processo patologico a eziologia sconosciuta in cui il sistema immunitario, deputato a difendere l'organismo da materiali estranei, dirige i propri attacchi contro parti del corpo del soggetto stesso. Quando le cellule β del pancreas sono colpite dal processo autoimmune, la secrezione di insulina in risposta al glucosio viene compromessa e alla fine le cellule secernenti insulina vengono distrutte; ciò dà luogo al cosiddetto diabete mellito insulino-dipendente o di tipo I. I pazienti affetti da questa malattia necessitano, per poter vivere, di quotidiane iniezioni di insulina. Le deficienze ormonali della tiroide, della corteccia surrenale e delle cellule dell'ovaia secernenti estrogeno rappresentano altrettante conseguenze, ben note, di processi autoimmuni. Generalmente soltanto un tipo di ghiandola viene colpito, ma in alcuni pazienti l'attacco autoimmune è diretto contro diverse ghiandole; si tratta delle cosiddette sindromi multighiandolari (poliendocrinopatie autoimmuni).
In alcuni pazienti le funzioni organiche appaiono alterate come se essi soffrissero di deficienze ormonali, mentre i livelli ormonali nel sangue non risultano diminuiti. Una possibile spiegazione di questa condizione è che la molecola di ormone sia difettiva, per esempio a causa di una mutazione del gene che codifica per quell'ormone; in questi casi, l'affinità o l'attività intrinseca dell'ormone può essere ridotta, ma l'analisi ormonale standard (basata sulla misurazione di un'altra proprietà dell'ormone) può non essere in grado di rilevare l'alterazione. In alternativa, è possibile che autoanticorpi o altre sostanze si leghino all'ormone nel sangue, riducendo così il suo accesso al recettore.
In un altro gruppo di disturbi i livelli ormonali sono sufficientemente elevati, ma la cellula bersaglio presenta una reattività ridotta. Difetti congeniti a carico del recettore e anticorpi che agiscono contro di esso sono soltanto due dei numerosi esempi di ‛stati di resistenza all'ormone' in cui la ridotta funzionalità del sistema endocrino è dovuta a un difetto nella cellula bersaglio, mentre la quantità e la qualità dell'ormone sono entrambe adeguate. In alcuni casi si rende necessaria una massiccia somministrazione di ormoni per produrre qualche miglioramento, anche se, attualmente, il trattamento di questi disordini in genere non dà risultati soddisfacenti.
b) Stati di iperfunzione.
Un'altra importante categoria di disordini endocrini è quella dei disturbi che si verificano quando il sistema è iperattivo (in alcuni casi il sistema non è iperattivo in termini assoluti: piuttosto, il suo livello di attività non è appropriato alle condizioni fisiologiche). I tumori delle ghiandole endocrine, generalmente benigni, possono secernere quantità troppo elevate di ormoni, spesso perché non rispondono adeguatamente a segnali di feedback negativo (v. figg. 14 e 15). In genere i tumori colpiscono una sola ghiandola, ma alcune famiglie sono predisposte geneticamente a contrarre tumori di diverse ghiandole endocrine. Alcuni tumori delle cellule β del pancreas producono insulina a prescindere dal livello di glucosio nel sangue e quindi possono provocare ricorrenti attacchi di ipoglicemia. I tumori dell'ipofisi, in alcuni rari casi, producono TSH (ormone stimolante la tiroide o tireotropina), a prescindere dal livello degli ormoni tiroidei circolanti che normalmente costituisce il segnale di inibizione retroattiva delle cellule secernenti il TSH. La tiroide, in risposta all'eccesso di TSH, produce ormone tiroideo in eccesso, il che determina un'accelerazione del battito cardiaco, perdita di peso accompagnata da accelerazione del metabolismo, nervosismo, sudorazione, tremore. Una sovrapproduzione di ormone tiroideo, con un insieme di sintomi del tutto simile, può essere causata da alcuni tipi di tumori della tiroide. In caso di malattia autoimmune, inoltre, le cellule immunocompetenti possono produrre anticorpi diretti contro il recettore del TSH; l'anticorpo, legandosi al recettore, può attivarlo proprio come fa il TSH stesso. Infine, anche cellule non endocrine, in conseguenza di tumori, spesso maligni, possono produrre ormoni la cui secrezione è insensibile ai normali segnali di feedback negativo. Queste malattie vengono affrontate con trattamenti chirurgici, chemioterapici o a base di radiazioni, che mirano a distruggere il tessuto iperattivo, oppure con farmaci specifici per bloccare la produzione o l'azione dell'ormone.
8. Altri ormoni
Esistono molti altri ormoni, oltre a quelli principali riportati nella tab. I. L'ipotalamo, localizzato alla base del cervello, possiede cellule nervose che secernono ormoni specifici, i quali regolano l'attività delle cellule secernenti ormoni dell'ipofisi (v. fig. 4). La ghiandola pineale (o epifisi), localizzata al centro dell'encefalo, proprio al di sopra del tronco encefalico (mesencefalo), è il sito di produzione della melatonina, una piccola molecola che contribuisce a regolare i ritmi circadiani dell'organismo e la cui sintesi è regolata dalle variazioni della luce ambientale. Il timo, localizzato nella cavità toracica, è un organo particolarmente importante nei primi anni di vita ed è la fonte di diversi ormoni che regolano lo sviluppo del sistema immunitario. Il tratto gastrointestinale, in particolare le cellule epiteliali che rivestono l'intestino tenue, produce diversi ormoni che, oltre a svolgere altre funzioni, coordinano le funzioni secretorie, digestive e peristaltiche dell'intestino. Questi ormoni sono anche stati tra i primi a essere scoperti e caratterizzati: fra i meglio conosciuti vi sono la secretina, la colecistochinina, la gastrina, l'istamina e l'enteroglucagone. Il rene produce l'eritropoietina, un ormone che stimola l'eritropoiesi da parte del midollo osseo; l'anemia che si riscontra nelle gravi insufficienze renali dipende in gran parte dalla mancanza di tale ormone, la cui somministrazione in questi casi ha un parziale effetto curativo. Recentemente si è scoperto che le cellule adipose secernono leptina, un ormone che segnala al cervello la quantità di grasso contenuta nei relativi depositi. La leptina agisce in modo da ridurre l'assunzione di cibo e da accelerare il tasso metabolico basale, fungendo così da regolatore negativo dell'appetito e del peso corporeo. Ultimamente è stato dimostrato che una razza di topi (topi ob/ob), geneticamente obesa e caratterizzata da estrema voracità, è priva di leptina funzionante. Mutazioni del gene della leptina sono causa di rarissime forme di obesità ereditaria. Un'altra sostanza ormono-simile secreta dalle cellule adipose è il fattore di necrosi tumorale (TNF) α, che viene liberato in risposta all'insulina e agisce riducendo l'effetto dell'insulina sulle cellule adipose e muscolari. Alcune cellule del cuore, specialmente nell'atrio sinistro, producono il fattore natriuretico atriale, una sostanza che modula la regolazione del sodio e dell'acqua da parte del rene. La placenta, oltre a provvedere al trasporto delle sostanze nutritizie dalla madre al feto e delle sostanze di scarto dal feto alla madre, è un importante organo produttore di ormoni, sia peptidici sia steroidei. Gli ormoni secreti dalla placenta costituiscono la base dei più comuni test di gravidanza. La gonadotropina corionica umana (HCG), l'ormone placentare più studiato, ha molte proprietà in comune con l'LH, l'ormone luteinizzante dell'ipofisi. L'HCG è sintetizzato anche nell'ipofisi, benché in quantità molto minori. Anche i derivati della vitamina D e della vitamina A agiscono come ormoni, e assomigliano molto agli ormoni steroidei e tiroidei per quel che riguarda la sintesi, i recettori sulle cellule bersaglio e i siti d'azione nucleari.
9. Sostanze ormono-simili
Oltre agli ormoni menzionati, esistono molte altre sostanze che non rispondono alla definizione tradizionale di ormone, perché sono prodotte da diversi tipi di cellule in varie parti del corpo, ma che svolgono un ruolo importante nelle comunicazioni tra cellule, in modo simile agli ormoni. Un vasto gruppo di queste sostanze è costituito dai ‛fattori di crescita'. Il fattore di crescita insulino-simile I (IGF-I) è secreto da molte cellule, specialmente nel fegato, in risposta all'ormone della crescita. Come suggerisce il suo nome, esso possiede molte proprietà chimiche e fisiologiche simili a quelle dell'insulina. Tra i fattori di crescita meglio studiati vi sono l'IGF-II, il fattore di crescita della cellula nervosa (NGF), il fattore di crescita dell'epidermide (EGF), il fattore di crescita dei fibroblasti (FGF) e il fattore di crescita piastrino-derivato (PDGF). Oltre a essere prodotti in molti siti, questi fattori di crescita agiscono su svariati tipi di cellule modificandone il programma di attività, anche stimolandone o inibendone la divisione e lo sviluppo. Molte di queste sostanze sono state scoperte per la loro capacità di stimolare la crescita di un particolare tessuto, ma è stato successivamente dimostrato che esse svolgono, nei confronti di diversi tipi cellulari, una gamma di attività molto più ampia. Lo sviluppo dei leucociti è regolato da fattori ormono-simili, tra cui il fattore di stimolazione delle colonie di granulociti macrofagi (GM-CSF); quest'ultimo viene usato per stimolare la leucocitopoiesi in pazienti i cui leucociti siano andati distrutti in seguito a radioterapia o chemioterapia.
I neuroni, oltre ai neurormoni, producono molti agenti ormono-simili che trasmettono messaggi da un nervo all'altro o da un nervo ad altre cellule bersaglio e che possono agire come modificatori delle cellule bersaglio. Tra questi agenti, noti collettivamente come ‛sostanze neuroattive', ve ne sono alcuni che funzionano da neurotrasmettitori, altri che funzionano da neuromodulatori e altri ancora che funzionano da fattori neurotrofici; questi ultimi sono simili ai fattori di crescita descritti in precedenza.
Alcune molecole di messaggeri ormono-simili sono prodotte da quasi tutte le cellule. Esse possono agire da sole o di concerto con ormoni e altri agenti ormono-simili. L'ATP e i suoi derivati, che, all'interno della cellula, svolgono un ruolo vitale nel metabolismo cellulare, quando vengono liberati nel liquido extracellulare si legano a specifiche molecole della superficie cellulare, dette ‛recettori purinergici', ed esercitano sulle cellule bersaglio effetti simili a quelli prodotti dagli ormoni. Anche le prostaglandine, derivate dai lipidi della membrana cellulare, e i derivati delle prostaglandine - prostaciclina, endoperossidi, trombossani e leucotrieni - si legano ai recettori della superficie cellulare e influenzano le cellule in modo simile agli ormoni.
Le cellule immunocompetenti sono la fonte di un gran numero di molecole ormono-simili, dette interleuchine, citochine e linfochine, delle quali l'interferone γ, il fattore di necrosi tumorale α e l'interleuchina-2 sono fra le più studiate. Queste molecole regolano la crescita, lo sviluppo e il funzionamento delle cellule immunocompetenti e di altre cellule del sangue. Inoltre, esse esercitano effetti su un gran numero di altre cellule, comprese quelle del sistema nervoso. Per esempio, l'interleuchina-1 agisce, fra l'altro, sul sistema nervoso per produrre la febbre. L'angiotensina e la bradichinina sono due agenti ormono-simili generati nel sangue e nei liquidi dei tessuti a partire da proteine più grandi prodotte nel fegato e in altri siti. I loro compiti principali sono quelli di regolare il calibro dei vasi sanguigni e il funzionamento dei reni.
10. Le molecole ormono-simili della matrice cellulare
In un organismo pluricellulare le cellule sono organizzate in strutture ben definite la cui regolazione è dovuta in buona parte a proteine extracellulari aventi caratteristiche diverse e note col nome di ‛matrice cellulare'. Generalmente le cellule possiedono recettori specifici che possono legarsi a specifici siti d'attacco presenti sulla superficie dei componenti della matrice cellulare. Oltre a consentire un legame fisico tra la cellula e la matrice, i recettori cellulari, quando sono occupati da componenti della matrice provvisti di siti d'attacco, inviano segnali alla cellula per alterarne la funzione. Questi componenti della matrice cellulare, che inducono mutamenti nell'attività e nella funzione della cellula, sono - sotto diversi aspetti - ormono-simili, e i loro effetti completano le attività della cellula regolate da ormoni in senso stretto e da altre molecole ormono-simili.
11. I rapporti tra il sistema endocrino e altri sistemi
Il concetto secondo cui gli organismi pluricellulari sono regolati da due diversi sistemi di coordinazione, distinti e indipendenti - il sistema nervoso e il sistema endocrino - è stato ormai superato; oggi sappiamo, infatti, che i due sistemi si sovrappongono in larga misura. Per esempio, due ghiandole tradizionali quali la parte posteriore dell'ipofisi e la midollare del surrene sono costituite da cellule secretorie nervose o di derivazione nervosa; anche i neuroni del cervello e del tratto gastrointestinale producono altri ormoni. I processi di secrezione, le molecole di segnalazione, i loro recettori e le vie attivate a valle dei recettori sono così simili nei due sistemi che i confini che li separano non sono più nettamente definiti. Il sistema immunitario è un altro sistema ‛ricco di segnali', che si sovrappone in larga misura sia al sistema nervoso, sia al sistema endocrino.
12. La nomenclatura degli ormoni
I nomi degli ormoni e di altre analoghe molecole di segnalazione - così come i nomi di molti personaggi biblici della Genesi - sono stati fatti derivare dalle circostanze in cui ebbe luogo la loro nascita o, per meglio dire, la loro scoperta. Talvolta la funzione scoperta per prima - e che ha dato il nome all'ormone - si è rivelata del tutto secondaria, nel complesso del ruolo fisiologico svolto dall'ormone. Altre volte linee di ricerca diverse e all'apparenza indipendenti hanno portato a coniare altrettanti nomi per quello che alla fine è risultato essere lo stesso ormone (in questi casi si possono continuare a usare i vari sinonimi). L'introduzione di nomi nuovi, nella lodevole intenzione di fare chiarezza, talvolta complica ulteriormente la situazione. Quel che è importante, sia per il lettore, sia per l'endocrinologo, è non lasciare che il nome limiti la comprensione della vasta gamma delle attività biologiche svolte da ciascuno di questi ormoni, o molecole ormono-simili.
13. La ricerca e i suoi metodi in endocrinologia
La ricerca endocrinologica, che ha oggi circa 150 anni (per una schematica cronistoria delle scoperte relative agli ormoni in questo secolo, v. tab. IV), iniziò a svilupparsi lungo due direttrici interrelate: la fisiologia delle ghiandole e le malattie umane. I medici del XIX secolo e dei primi anni del Novecento, osservando pazienti affetti da certe malattie, notarono che determinati cambiamenti anatomici e fisiologici diffusi in tutto il corpo potevano essere collegati a mutamenti macroscopici e microscopici di un singolo organo. Lesioni distruttive e tumori o altre iperplasie a carico di quell'organo erano quanto mai facili da scoprire.
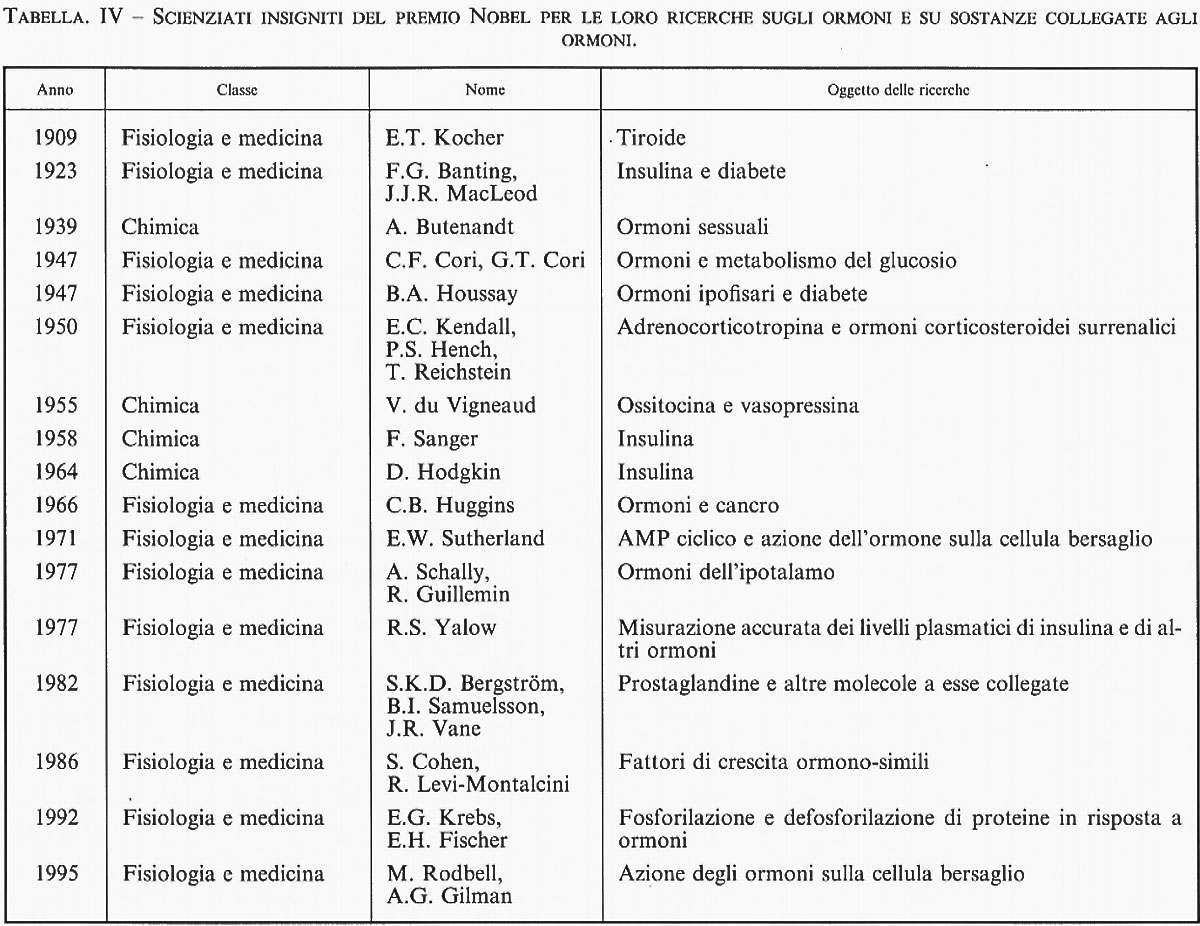
Contemporaneamente, i fisiologi procedevano alla rimozione sistematica di singoli organi in animali da esperimento per apprenderne le funzioni. Gli ormoni prodotti da organi che svolgono molte altre funzioni - come il cervello, il cuore, il fegato e il rene - potevano essere difficilmente identificati in questo modo; le anomalie dovute alla perdita della funzione ormonale venivano sommerse dagli effetti della perdita di altre funzioni. Tali ormoni sono stati scoperti più tardi e con maggior fatica. Viceversa, le ghiandole endocrine propriamente dette - quali la tiroide, le paratiroidi, l'ipofisi, le ghiandole surrenali - organi in cui le funzioni ormonali sono dominanti, furono più facili da individuare con questo metodo. Allo stesso modo è stata determinata la natura endocrina di ghiandole che svolgono anche altre funzioni, non endocrine, quali i testicoli, le ovaie e il pancreas.
Seguendo questa impostazione classica, un organo veniva rimosso chirurgicamente e se ne osservavano le conseguenze fisiologiche sull'organismo. Estratti delle ghiandole espiantate venivano quindi iniettati nell'animale da cui erano state rimosse per osservare quali anomalie venivano compensate. Il materiale attivo presente negli estratti ghiandolari veniva poi purificato e concentrato, finché la sostanza responsabile dell'attività biologica era ottenuta in forma pura e la sua struttura determinata. Attualmente, per essere certi dell'identità del materiale attivo, se ne prepara una copia sintetica che si confronta in ogni modo possibile con la forma naturale, originale dell'ormone.
Ulteriori studi, oggi di routine per tutti gli ormoni, mirano a: 1) identificare il gene o i geni che codificano per quell'ormone (nel caso di ormoni peptidici), ovvero il gene o i geni che codificano per gli enzimi che sintetizzano quell'ormone (nel caso di ormoni che non sono peptidi); 2) identificare il recettore specifico dell'ormone oggetto di studio; 3) identificare i geni che codificano per il recettore in questione; 4) definire il secondo messaggero o altre importanti vie intracellulari attivate dal complesso ormone-recettore.
Il metodo sperimentale tradizionale della rimozione chirurgica di ghiandole da una cavia è stato notevolmente ampliato in diversi modi creativi che lo hanno reso più specifico e flessibile. Un meccanismo consiste nel neutralizzare l'ormone in vivo o in provetta, introducendo anticorpi specifici; l'anticorpo o impedisce all'ormone di interagire col proprio recettore, o ne accelera il trasporto ai siti di degradazione. Sono anche stati creati antagonisti dell'ormone che gli assomigliano sotto diversi aspetti, ma che, quando si legano al recettore, lo bloccano anziché attivarlo. Negli ultimi anni è stato introdotto un metodo particolarmente potente: i ricercatori creano razze di topi in cui il gene relativo all'ormone, o quello relativo al recettore, viene eliminato selettivamente (topi knock-out). All'estremo opposto c'è il topo ‛transgenico', in cui si introducono copie extra del gene relativo a un dato ormone, o recettore; da ciò segue uno stato di iperfunzionamento.
Fondamentale, per tutte queste indagini, è il ‛dosaggio', un metodo in grado di misurare specificamente, accuratamente e con grande sensibilità la concentrazione dell'ormone. I dosaggi basati sulle proprietà biologiche dell'ormone, o biodosaggi (bioassays), sono di importanza cruciale, ma spesso vengono sostituiti da altri dosaggi specialmente perché sono più precisi, sensibili e specifici, ma anche solo per convenienza. Per piccole molecole, come gli ormoni tiroidei o gli ormoni steroidei, i dosaggi chimici sono in genere affidabili, mentre per gli ormoni peptidici e per altri ormoni la tecnica d'elezione è l'immunodosaggio (immunoassay). Oggi si usano spesso anticorpi capaci di riconoscere l'ormone, sviluppati in vivo ma impiegati in provetta, perché possiedono un'altissima sensibilità e un'altissima specificità; essi possono riconoscere minime quantità di ormone e sono in grado di farlo in soluzioni molto complesse. Il dosaggio ignora tutte le sostanze, eccetto quelle poche specie molecolari ormono-simili che sono riconosciute dall'anticorpo.
Dato che i dosaggi chimici e gli immunodosaggi misurano proprietà o regioni della molecola di ormone che non coincidono necessariamente con le sue parti biologicamente rilevanti, ogni nuovo dosaggio deve, di norma, essere convalidato dall'accertamento delle proprietà biologiche; da un punto di vista concettuale la tecnica più importante per misurare un ormone è il biodosaggio, che registra l'attività biologica specifica dell'ormone interagente col recettore in funzione della concentrazione dell'ormone. In origine i biodosaggi venivano eseguiti su cavie normali; in seguito si è passati ad animali con livelli ormonali previamente ridotti quanto più possibile, in modo da poter riconoscere più facilmente l'ormone aggiunto dall'esterno. I biodosaggi in cui si usano animali vivi sono costosi, imprecisi e meno sensibili - rispetto ad altri tipi di biodosaggi - in condizioni di basse concentrazioni ormonali. Gli organi che contengono le cellule bersaglio possono essere perfusi in situ o rimossi per essere perfusi in un termostato. Oggigiorno le cellule bersaglio per biodosaggi più diffusamente utilizzate sono quelle appena isolate (colture primarie) o quelle derivanti da linee cellulari immortali cresciute in provetta e che conservano la propria reattività all'ormone. Questi dosaggi possono offrire alti livelli di specificità e sensibilità.
È opportuno ricordare che gli animali hanno sempre svolto un ruolo indispensabile nella ricerca sugli ormoni (gli esperimenti sui cani, per esempio, hanno portato alla scoperta e all'isolamento dell'insulina per il trattamento del diabete) e continuano a essere importanti in questo campo così come in tutti i settori della ricerca bio-medica.
BIBLIOGRAFIA
Addison, T., On the constitutional and local effects of diseases of the suprarenal glands, London 1855.
Basedow, V. von, Exophthalmus durch Hypertrophie des Zellegewebes in der Augenhöhle, in ‟Wochenschrift für die gesamte Heilkunde", 1840, VI, pp. 197-204.
Becker, K. L. (a cura di), Principles and practice of endocrinology and metabolism, Philadelphia, Pa., 19952.
Berson, S. A., Yalow, R. S., Radioimmunoassay of peptide hormones in plasma, in ‟New England journal of medicine", 1967, CCLXXVII, pp. 640-647.
Bliss, M., The discovery of insulin, Chicago 1982.
Burgus, R., Guilleman, R. e altri, Structure moléculaire du facteur hypothalamique hypophysiotrope TRF d'origine ovine: mise en évidence par spectrométrie de masse de la séquence PCA-His-Pro-NH2, in ‟Comptes rendus hebdomadaires des séances de l'Académie des Sciences", 1969, CCLXIX, pp. 226-228.
Collip, J. B., The original method as used for the isolation of insulin in semi-pure form for the treatment of the first clinical case, in ‟Journal of biological chemistry", 1923, LV, pp. 50-51.
Cushing, H., The basophil adenomas of the pituitary body and their clinical manifestations (pituitary basophilism), in ‟Bulletin of the Johns Hopkins Hospital", 1932, L, pp. 137-195.
DeGroot, L. J. e altri (a cura di), Endocrinology, 3 voll., Philadelphia, Pa., 19943.
Felig, P., Baxter, J. D., Frohman, L. A. (a cura di), Endocrinology and metabolism, New York 19953 (tr. it.: Endocrinologia e metabolismo, Milano 19912).
Fitzgerald, M. J. T., Fitzgerald, M. (a cura di), Human embryology, Philadelphia, Pa., 1994.
Fye, B., The history of medicine: an annotated list of key reference works, in ‟Annals of internal medicine", 1993, CXVIII, pp. 59-62.
Graves, R. J., Clinical lecture, in ‟London medical and surgical journal", 1835, VII, pp. 599 ss..
Guyton, A. C., Hall, J. E. (a cura di), Textbook of medical physiology, Philadelphia, Pa., 19969.
Kadowaki, T., Bevins, C. L. e altri, Two mutant alleles of the insulin receptor gene in a patient with extreme insulin resistance, in ‟Science", 1988, CCXL, pp. 787-790.
Kendall, E. C., Mason, H. L., McKenzie, B. F., Myers, C. S., Koelsche, G. A., Isolation in crystalline form of the hormone essential to life from the suprarenal cortex: its chemical nature and physiologic properties, in ‟Proceedings of the Mayo Clinic", 1934, IX, pp. 245-250.
Laron, Z., Pertzelan, A., Mannheimer, S., Genetic pituitary dwarfism with high serum concentration of growth hormone. A new inborn error of metabolism?, in ‟Israel journal of medical sciences", 1966, II, pp. 152-155.
McCann, S. M. (a cura di), Endocrinology, people and ideas, Baltimore, Md., 1988.
Medvei, V. C., A history of endocrinology, Lancaster 1982.
Meites, J., Donovan, B. T., McCann, S. M. (a cura di), Pioneers in neuroendocrinology, 2 voll., New York 1975-1978.
Mendeloff, A. J., Smith, D. E. (a cura di), Acromegaly, diabetes, hypermetabolism, proteinuria and heart failure, in ‟The American journal of medicine", 1956, XX, pp. 133-144.
Mering, J. von, Minkowski, O., e altri, Diabetes mellitus nach Pankreasexstirpation, in ‟Archiv für die experimentelle Pathologie und Pharmakologie", 1890, XXVI, pp. 371-387.
Pierce, J. G. (a cura di), Protein and peptides hormones, New York 1982.
Roth, J., Taylor, S. I., Information transfer, cell regulation, and disease mechanism: insight from studies of cell surface receptors, in ‟The Harvey lectures", 1983, LXXVII, pp. 81-127.
Scharrer, E., Scharrer, B., Neuroendocrinology, New York 1963.
Scriver, C. A., Beaudet, A. L., Sly, W. S., Valle, D. (a cura di), The metabolic bases of the inherited diseases, New York 19957.
Stryer, L. (a cura di), Biochemistry, New York 19954 (tr. it.: Biochimica, Bologna 19964).
Thorn, G. W., Firor, W. M., Desoxycorticosterone acetate therapy in Addison's disease: clinical consideration, in ‟Journal of the American Medical Association", 1940, CCXXXI, pp. 76 ss..
Williams, P. L. (a cura di), Gray's anatomy, Edinburgh 199538.
Wilson, J. D., Foster, D. W. (a cura di), Williams textbook of endocrinology, Philadelphia, Pa., 19928 (tr. it.: Trattato di endocrinologia, Padova 19855).
Yalow, R. S. (a cura di), Radioimmunoassay, New York 1983.