
Evoluzione del genere Homo
Evoluzione del genere Homo
La storia del genere Homo è lunga, complessa e di grande rilevanza per la comprensione dell'attuale variabilità umana. Dall'inizio di questa storia evolutiva, più di due milioni di anni fa, almeno tre specie di Homo sono comparse prima degli ultimi 500 mila anni, e almeno tre o quattro possibili specie (o forse sottospecie) potrebbero essersi evolute nelle ultime centinaia di migliaia di anni. Solo una, Homo sapiens, sopravvive al giorno d'oggi. Tra le prime caratteristiche adattative derivate del genere Homo rispetto ad Australopithecus vi sono un cervello relativamente grande rispetto alle dimensioni del corpo e una serie di adattamenti specializzati per la locomozione bipede. H. sapiens non possiede un cervello più grande delle altre specie recenti del genere Homo, ma differisce da queste principalmente per la fisionomia craniofacciale. Sembra che H. sapiens si sia evoluto approssimativamente 100 ÷ 200 mila anni fa in Africa o forse in Medio Oriente, e che abbia ampiamente sostituito in tutto il Vecchio Mondo i taxa preesistenti, le cui storie evolutive risalirebbero alla migrazione di Homo erectus fuori dall'Africa, durante l'Era Glaciale.
Introduzione
Il genere Homo, vecchio più di due milioni di anni, comprende almeno quattro specie, forse sei. Come genere, Homo si distingue da Australopithecus per il considerevole incremento delle dimensioni del cervello rispetto alla massa corporea, per l'evoluzione di una stazione eretta pienamente sviluppata e di una locomozione bipede specificamente adatta a camminare e correre su grandi distanze, per la tendenza a procurarsi il sostentamento tramite battute di caccia su larga scala, per la grande dispersione geografica, dagli habitat dell'Africa tropicale agli ambienti più diversi del Vecchio e Nuovo Mondo, e naturalmente per le straordinarie innovazioni sociali e tecnologiche. In questo saggio sono riportate le principali testimonianze relative a tre problemi tassonomici e filogenetici fondamentali che riguardano l'evoluzione del genere Homo: il periodo e l'area in cui si è evoluto; l'origine di Homo erectus e la sua migrazione al di fuori dell'Africa attraverso il Vecchio Mondo; l'origine di Homo sapiens e il destino dei taxa più arcaici, come Homo neanderthalensis. Il saggio verterà principalmente su tali questioni sistematiche che sono alla base di altre ipotesi evolutive di più alto livello sul comportamento, l'ecologia e lo sviluppo del genere Homo, ognuna delle quali verrà discussa brevemente.
Controversie
Malgrado gli incredibili progressi resi possibili dalle nuove tecnologie e dalle scoperte degli ultimi decenni, lo studio degli ultimi due milioni di anni della storia evolutiva dell 'uomo rimane una delle branche più controverse della biologia. Almeno quattro fattori contribuiscono alle numerose discussioni sull'evoluzione del genere Homo. Primo fra tutti, e più importante, è che le ipotesi sulla sua evoluzione trattano inevitabilmente di questioni quali razza, identità umana e potenziali riflessi del nostro passato evolutivo sul concetto moderno di umanità. Inoltre, i paleoantropologi sono fortemente ostacolati dal numero relativamente piccolo di reperti fossili, nei quali disponibilità e qualità dei dati sono spesso insufficienti per verificare molte ipotesi di notevole interesse. In più i resti fossili e archeologici provenienti da molte parti del mondo non vengono sufficientemente campionati e classificati. Infatti vi sono esemplari abbastanza completi provenienti dall'Europa e dal Medio Oriente, mentre sono ancora scarsi i fossili datati e i siti archeologici in Asia orientale e Africa orientale e meridionale, e sono praticamente assenti in aree geografiche importanti come l'Asia meridionale e l'Africa occidentale.
Un'ulteriore difficoltà nel verificare le ipotesi sull'evoluzionedel genere Homo consiste nella necessità di integrare le testimonianze fossili non solo con i dati molecolari ma anche con i dati archeologici, nessuno dei quali è in completo accordo con le evidenze paleontologiche umane. Infine, la paleoantropologia, come molte altre branche della paleontologia, risente del dibattito epistemologico sulle interpretazioni dei reperti fossili. Tre questioni principali sono particolarmente controverse. Innanzitutto, non c'è un accordo generale su come definire valide unità tassonomiche all'interno del genere Homo, in particolare a livello di specie, ma anche a quello di sotto specie (Tattersall, 1994; Wolpoff, 1994). Inoltre, vi è una grande controversia sull'applicazione teorica e pratica della sistematica cladistica, che basa la classificazione delle specie esclusivamente sulle loro relazioni evolutive, rispetto a quella fenetica, che invece basa la classificazione sul livello di somiglianza tra le specie, per risolvere le relazioni sistematiche tra taxa recenti (per esempio, popolazioni viventi o razze) che potrebbero non rappresentare cladi, cioè gruppi tassonomici i cui membri discendono tutti da un antenato comune, o linee distinte (Lieberman, 1995). Infine, gli antropologi non concordano sul significato biologico della variabilità morfologica, specialmente per quanto riguarda il contributo relativo di stimoli, genetici e non, alle modificazioni dello scheletro (Lieberman, 1995; Churchill, 1996).
Tassonomia
Homo habilis e Homo rudolfensis

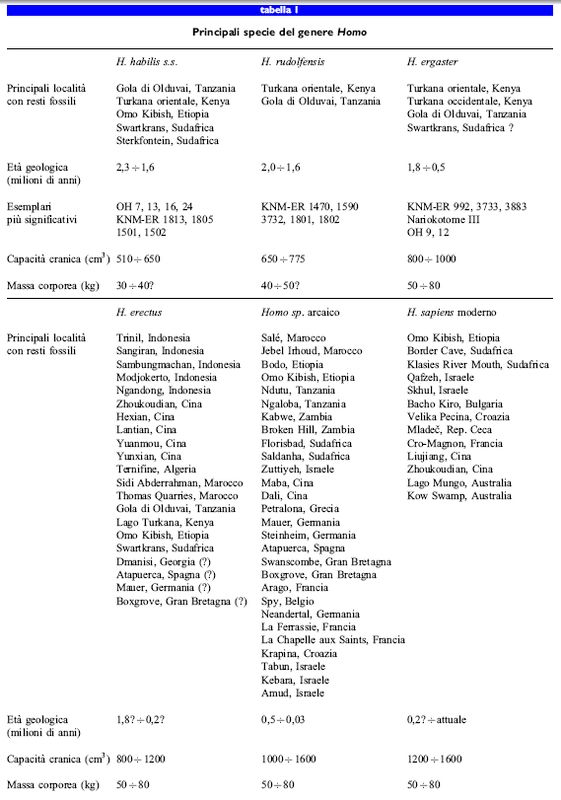
I primi resti fossili attribuiti al genere Homo, provenienti dall' Africa orientale, risalgono ad almeno 2,2 milioni di anni fa, ma potrebbero avere fino a 2,5 milioni di anni (Hill et al., 1992). Alcune specie del tardo Pliocene sono state ritrovate esclusivamente in Africa, ma è importante notare che la loro attribuzione tassonomica è complicata dalla storia del loro recupero e della loro classificazione. Basandosi soprattutto sui resti dell'Ominide 7 di Olduvai, ritrovato nel 1960, L. Leakey, P. Tobias e J. Napier (1964) istituirono la specie H. habilis. Inizialmente, le caratteristiche più importanti che definivano la specie H. habilis consistevano nelle dimensioni del cervello (da 600 a 750 cm³) e dei molari, intermedie tra quelle di H. erectus e quelle di diverse specie di Australopithecus già ben conosciute e studiate. Le successive scoperte di fossili riconducibili a questa categoria intermedia furono quindi classificate come H. habilis, malgrado presentassero un enorme grado di variabilità sia nelle dimensioni sia nella forma (Howell, 1978; Lieberman et al., 1988). Studi recenti hanno portato a una revisione formale di tale raggruppamento che viene suddiviso in almeno due specie, H. habilis sensu stricto (s.s.) e H. rudolfensis (Stringer 1986; Wood, 1991; 1992; Rightmire, 1993; Lieberman et al., 1996). H. habilis s.s., ben rappresentato dai crani KNMER 1813 e OH 24, è caratterizzato da una capacità cranica tra 510 e 650 cm³, da una faccia ortognata separata dal neurocranio da un piccolo ma ben sviluppato toro sopraorbitario, dalla zona superiore della faccia più larga di quella intermedia e del palato, da una cavità nasale a forma triangolare con ossa nasali rovesciate, da un'arcata zigomatica verticale e da piccoli denti sottilmente smaltati delle dimensioni di quelli di H. erectus (v. oltre), con una restrizione del diametro bucco-linguale della corona dei canini e una riduzione del terzo molare - il nostro dente del giudizio - rispetto al secondo molare. Una rappresentazione schematica delle ossa del cranio è riportata nella figura (fig. 2). La maggior parte dei fossili di H. habilis s.s. è stata datata tra 2,3 e 1,6 milioni di anni fa (tab. I). L'altro taxon dei primi Homo, H. rudolfensis, rappresentato dai crani fossili che includono KNM-ER 1470, differisce da H. habilis s.s. per alcune caratteristiche importanti. H. rudolfensis ha una capacità cranica decisamente più grande, tra 650 e 775 cm³, una regione sopraorbitale in cui manca un toro distinto che, al contrario, si fonde col neurocranio senza un solco, una faccia intermedia larga e piatta più ampia di quella superiore, una regione zigomatica inclinata anteriormente, un ampio palato prognato e denti decisamente più grandi e più finemente smaltati, con un'ampia corona dei canini e senza riduzione del terzo molare (Wood, 1992). In altre parole, H. habilis s.s. ha faccia e dentatura più simili a quella degli altri rappresentanti del genere Homo assieme a un cervello di dimensioni più piccole, mentre H. rudolfensis ha un cervello decisamente più grande associato a una faccia e a una dentatura più simili ad Australopithecus. l fossili di H. rudolfensis sono stati datati tra 2,0 e 1,6 milioni di anni fa, ma molti potrebbero risalire fino a 2,5 milioni di anni fa. Poco si sa sulla morfologia dello scheletro postcraniale di H. habilis s.s. e di H. rudolfensis. A causa della scarsità dei reperti scheletrici postcraniali associati ai crani ancora non si conosce la taglia corporea e quindi il rapporto cervello/massa corporea delle due specie. Tuttavia, i resti postcraniali di un individuo pressoché completo, OH 62, presumibilmente H. habilis s.s., assomigliano a quelli degli australopitecini per dimensioni, proporzioni e morfologia (Johanson et al., 1987; Hartwig-Scherer e Martin, 1991). Inoltre, alcuni resti postcraniali isolati come KNMER 813,1472 e l48lA, generalmente attribuiti a H. habilis sensu lato (s.I.), ma che provengono da depositi che contenevano esemplari di H. rudolfensis come KNM-ER 1470, appartengono evidentemente a individui di grossa corporatura che sono, dal punto di vista dello scheletro postcraniale, del tipo Homo sia per la morfologia sia per le proporzioni (McHemy e Corruccini, 1978; Wood, 1992). Se queste attribuzioni fossero corrette, allora H. habilis s.s. e H. rudolfensis dovrebbero aver avuto un rapporto cervello/taglia corporea equivalente.
Homo erectus e Homo ergaster
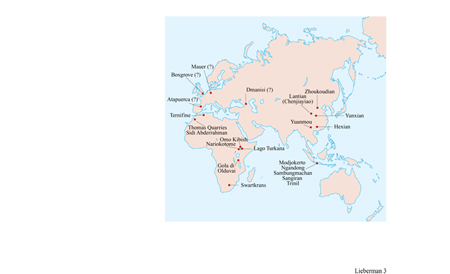
La specie H. erectus, così come è conosciuta oggi, venne definita per la prima volta nel 1894 come Pithecanthropus erectus dal medico olandese E. Dubois, che ne rinvenne i primi esemplari fossili nel sito di Trinil sul fiume Solo a Giava (fig. 3). Altri fossili di rilievo, ora attribuiti alla specie H. erectus ma originariamente classificati come Sinanthropus pekinensis, vennero scoperti tra gli anni Venti e Trenta nella famosa grotta superiore di Zhoukoudian vicino a Pechino, in Cina. Negli anni Trenta, le successive scoperte di fossili simili nelle località di Modjokerto e Sangiran a Giava spinsero i ricercatori a classificare tali taxa tutti insieme in una unica specie (Mayr, 1950; Howell, 1960). Sin dalla seconda guerra mondiale, in Asia l'ipodigma di H. erectus, cioè la lista di fossili assegnati alla specie, è stato sostanzialmente ampliato con la scoperta di altri fossili a Giava, per esempio a Trini!, Ngandong e Sambungmachan, e in Cina, per esempio a Lantian, Yuanmou, Chenjiayiao, Hexian e Yunxian (Wu e Poirier, 1995). Inoltre, fossili generalmente attribuiti a H. erectus sono stati rinvenuti nell'Africa sub-sahariana - come quelli della gola di Olduvai in Tanzania, del Lago Turkana in Kenya, di Orno Kibish in Etiopia e di Swartkrans in Sudafrica - e nell'Africa settentrionale - come quelli di Ternifine in Algeria e di Sidi Abderrahman e Thomas Quarries in Marocco; inoltre fossili probabilmente di H. erectus sono stati rinvenuti in Europa (v. oltre).
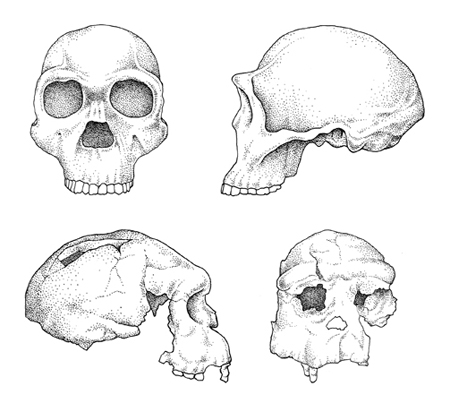
H. erectus è solitamente identificato in base all'aumento della sua capacità cranica, che varia da 800 a 1200 cm³, ma non c'è ancora accordo su come definire la specie con maggiore precisione. l tratti caratteristici, probabilmente esclusivi, del cranio di H. erectus (fig. 4) includono un neurocranio lungo e basso, un massiccio toro sopraorbitario separato dal neurocranio da un solco ben definito, un toro occipitale a forma di angolo acuto all'intersezione della sua porzione nucale e squamo sa, che spesso coincide con una protuberanza, e un prominente toro angolare all'angolo mastoideo. Inoltre, molti crani di H. erectus si distinguono da quelli di H. habilis e H. rudolfensis per alcune caratteristiche derivate che includono un lieve grado di increspatura lungo le suture metopica e sagittale, una spessa volta cranica, una larga cavità nasale con i margini rovesciati e una faccia massiccia ma ortognata, con l'eccezione della porzione alveolare della mascella che è prognata. Dal punto di vista della morfologia dentaria, H. erectus ha molari leggermente più grandi dell 'uomo moderno, con larghi incisivi superiori spesso a forma di spatola. Se si considera lo scheletro postcraniale, H. erectus appare del tutto simile a H. sapiens per taglia corporea, proporzioni e morfologia complessiva. l fossili di H. erectus sono tipicamente molto robusti, con ossa piatte estremamente spesse. Inoltre, il bacino di H. erectus è più stretto di quello dell'uomo moderno, con un caratteristico ispessimento osseo (pilastro iliaco) sopra l'acetabolo.
Il contesto cronologico e geografico di H. erectus rimane controverso per almeno tre motivi. Primo e più importante è che alcuni fossili provenienti dall'Africa sub-sahariana, come KNM-ER 3773, 3883, Nariokotome lll, e forse Swartkrans 847, datati tra 1,8 e 1,2 milioni di anni fa, sono generalmente attribuiti a H. erectus, anche se in realtà sono abbastanza distinti dal punto di vista morfologico (v. oltre) da costituire un taxon a sé stante, H. ergaster (Wood, 1991). Il secondo motivo è che le nuove datazioni radiometriche suggeriscono la possibilità che qualche fossile di H. erectus rinvenuto in Cina e in Indonesia possa essere altrettanto antico se non addirittura più vecchio di tutti gli altri fossili conspecifici dell'Africa. Infatti, sulla base di dati radiometrici su sedimenti vulcanici trovati nel canale uditivo, è stato suggerito che un individuo, l'infante di Modjokerto, rinvenuto in depositi alluvionali vicino a Sangiran, possa risalire a 1,8 milioni di anni fa (Swisher et al., 1994), ma questa data resta controversa e ha bisogno di essere confermata. Inoltre, alcuni frammenti di materiale mandibolare e dentario recuperati nella grotta di Longgupo nella Cina centrale (Huang et al., 1995) sono stati datati, mediante correlazioni paleomagnetiche e faunistiche, a 1,9 milioni di anni fa; sebbene molti ricercatori ritengano che il frammento mandibolare appartenga al genere Homo, l'attribuzione a H. erectus non è sicura. Infine, le date dell' estinzione di H. erectus sono poco definite a causa di seri disaccordi su questioni tassonomiche. Comunemente, i fossili del genere Homo del Pleistocene, che non sono uomini moderni ma che sono più giovani di circa 500 mila anni e hanno una capacità cranica maggiore di 1200 cm³, vengono raggruppati senza ulteriore distinzione nel taxon H. sapiens arcaico, secondo un criterio che si basa più sul grado di somiglianza che sulla nozione di clado. Alcuni ricercatori hanno adottato il punto di vista estremo per cui il taxon H. erectus deve essere inglobato all'interno di H. sapiens (Wolpoff, 1996). Questa tassonomia basata sul grado di somiglianza resta un problema serio e necessita di una risoluzione poiché essa, accorpando assieme taxa differenti, può, in linea di principio, ostacolare gli sforzi per saggiare con rigore ipotesi filo genetiche alternative.
Studi recenti (Wood, 1991 e 1992) hanno evidenziato una possibile distinzione a livello di specie tra il materiale classico di H. erectus, in gran parte proveniente dall'Asia, e alcuni fossili più antichi rinvenuti in Africa. Questi sono feneticamente simili a H. erectus asiatico, ma non presentano molte delle autapomorfie (caratteri derivati esclusivi di un taxon) di H. erectus precedentemente descritte. Paragonati agli esemplari asiatici, i fossili dell' Africa sub-sahariana come KNM-ER 3733, KNM-ER 3883 e Nariokotome III, hanno le ossa della volta cranica più sottili, gli occipitali meno angolati e non presentano una cresta sagittale. Di conseguenza, tali individui sono stati provvisoriamente attribuiti a H. ergaster (Wood, 1991), ma tale taxon non è stato ancora formalmente definito con chiarezza. Il riconoscimento della specie Homo ergaster solleva interessanti questioni sistematiche. Come già menzionato, il frammento mandibolare di Longgupo potrebbe essere di H. ergaster, suggerendo così non solo la possibilità che H. ergaster sia stato il primo ominide a migrare fuori dall' Africa, ma anche l'ipotesi che H. erectus s.s. si sia evoluto esclusivamente in Asia senza aver mai abitato l'Africa sub-sahariana o l'Europa. Infatti, in Europa sono stati trovati solo pochi fossili che potrebbero appartenere a H. erectus - a Dmanisi in Georgia, a Boxgrove in Gran Bretagna, a Atapuerca in Spagna e a Mauer in Germania - ma la maggior parte di questi fossili ha anche alcuni tratti derivati in comune con Homo sp. indet. (specie indeterminata) arcaico. Con l'eccezione della mandibola rinvenuta a Dmanisi, che probabilmente risale a 1,4 milioni di anni fa, non ci sono testimonianze certe che H. erectus s.s. abbia mai occupato l'Europa.
Homo sapiens e Homo sp. indet.
La tassonomia delle specie recenti del genere Homo è forse persino più complessa di quella delle specie più antiche; ciò deriva dalla pratica comune di collocare nella nostra stessa specie, H. sapiens, tutti i fossili del Pleistocene medio e superiore con caratteristiche derivate non condivise da H. erectus s.I. Infatti, molti studiosi impiegano uno schema tassonomico basato chiaramente sul grado di somiglianza, che suddivide ulteriormente questo esemplare secondo criteri temporali e morfologici in due sotto specie distinte: H. sapiens arcaico e H. sapiens moderno. In accordo con tale schema, i fossili che rappresentano gli 'umani arcaici' includono gli individui del Pleistocene medio e superiore rinvenuti in Africa e in Asia, tra cui gli esemplari che potrebbero rappresentare il più recente H. erectus, e quelli rinvenuti in Europa, compreso l'Uomo di Neandertal. Poiché una definizione di H. sapiens basata sul grado di somiglianza porta probabilmente alla costruzione di un taxon polifiletico, se non addirittura parafiletico, che include linee evolutive potenzialmente diverse del tardo Homo, si è nuovamente tentato di definire taxa distinti filogeneticamente nell'ambito di Homo arcaico, tra cui Homo heidelbergensis, Homo neanderthalensis, Homo rhodesiensis e Homo soloensis (Tattersall, 1986). Tuttavia, non si è ancora raggiunto un accordo unanime su quanti di questi taxa esistano o su come debbano essere chiamati: di conseguenza è stata utilizzata la denominazione Homo sp. indet., da qui in poi indicato come Homo sp. arcaico.

In generale, i fossili attribuiti a Homo sp. arcaico sono estremamente variabili sia tra le diverse aree geografiche sia all'interno di una stessa area, e molti di essi provengono da contesti geologici o archeologici di età incerta. Dal punto di vista morfologico, Homo sp. arcaico condivide con H. erectus molti tratti primitivi del cranio, incluse una lunga, bassa e spessa volta cranica che si allarga verso la base, una robusta mandibola che manca del mento, una faccia larga ma verticale ben posizionata di fronte al neurocranio e una massiccia arcata sopraccigliare che separa la parte alta della faccia dalla volta cranica. I fossili di Homo sp. arcaico si distinguono da quelli di H. erectus soprattutto per l'aumentata dimensione del cervello, tra 1000 e 1600 cm³, che rientra nelle dimensioni di quello di H. sapiens moderno. Altre comuni caratteristiche derivate di Homo sp. arcaico (fig. 5) includono una fronte più ampia e quindi una faccia superiore più grande, un delicato osso del timpano praticamente allineato con la porzione della rocca petrosa dell' osso temporale, un tubercolo articolare a forma di barra al margine anteriore della fossa temporale glenoidea, un osso temporale corto con una sutura squamosa regolarmente curvata, ossa parietali relativamente verticali e occipitali più arrotondate (Stringer, 1985; Tattersall, 1986; Rightmire, 1990). Se non tutte, molte di queste differenze nell'architettura della volta cranica di Homo sp. arcaico e di H. erectus sono probabilmente correlate all'espansione del volume dell'endocranio. H. sapiens moderno si distingue da Homo sp. arcaico principalmente per alcune particolari caratteristiche derivate che comprendono una bassa base cranica, che fa sì che la faccia sia posta completamente al di sotto della fossa cranica anteriore riducendo o addirittura eliminando la presenza di una vera e propria arcata sopraccigliare, e una volta cranica arrotondata e sferica, con una regione occipitale tondeggiante (Lieberman, 1995). Queste differenze di architettura del cranio sono associate a una serie di altre caratteristiche derivate che comprendono una fronte alta, un toro sopraorbitario discontinuo e relativamente ridotto, un sostegno cranico più piccolo e un lungo e alto arco parietale più stretto in basso e più largo nella regione superiore (Day e Stringer, 1982; Stringer, 1987; Stringer e Andrews, 1988). H. sapiens è caratterizzato anche dalla presenza di un vero e proprio mento e da denti di dimensione ridotte. Contrariamente a quanto si è soliti credere, molti tra i primi H. sapiens avevano cranio e postcranio molto robusti (Ruff et al., 1993; Lieberman, 1996).
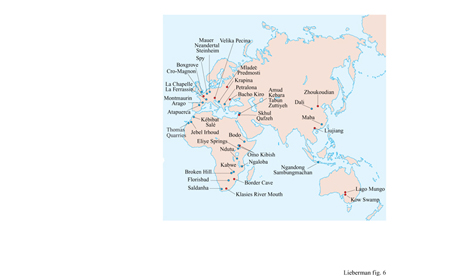
Poiché le definizioni di H. erectus, Homo sp. arcaico e H. sapiens, così come le distinzioni fra di loro, rimangono controverse, l'interpretazione dei reperti fossili è difficilmente collocabile nell' ambito di un inquadramento tassonomico e quindi filogenetico. Dal nostro punto di vista, i resti del Pleistocene medio rinvenuti in Africa, che condividono numerose caratteristiche con H. erectus s.I. ma che sono probabilmente meglio classificati come Homo sp. arcaico, comprendono i fossili rinvenuti a Salé e a Thomas Quarries in Marocco, a Bodo in Etiopia e a Ndutu in Tanzania (fig. 6). Esemplari un po' più recenti, rappresentativi di Homo sp. arcaico africano, includono i resti scheletrici recuperati a Jebel Irhoud in Marocco, a Omo Kibish in Etiopia, a Eliye Springs in Kenya, a Ngaloba in Tanzania, a Kabwe nello Zambia e a Saldanha e Florisbad in Sudafrica. La classificazione degli ominidi fossili asiatici del Pleistocene medio non è meno difficile: in Asia, individui con affmità a H. erectus s.s. persistono fino alla fine del Pleistocene medio in siti quali Ngandong, a Giava, e Hexian, in Cina. Questi fossili sembrano essere contemporanei di altri fossili rinvenuti in Cina e ragionevolmente attribuiti a Homo sp. arcaico, inclusi i crani di Dali e Maba e lo scheletro incompleto di Jinniu Shan.
I fossili di Homo sp. arcaico meglio caratterizzati sono quelli rinvenuti in Europa e Asia occidentale, dove vengono convenzionalmente divisi in: antichi, Pleistocene medio, e recenti, Pleistocene superiore. La mandibola del Pleistocene inferiore (risalente forse a 1,4 milioni di anni fa) rinvenuta a Dmanisi, in Georgia, ha alcune caratteristiche primitive che la collegano sia a H. erectus sia a H. ergaster, e potrebbe rappresentare un membro europeo di una di queste specie. Anche i fossili del Pleistocene medio rinvenuti nei siti di Mauer e Bilzingsleben in Germania e di Vértesszöllös in Ungheria, presentano delle affinità con H. erectus ma mostrano un maggior numero di caratteristiche derivate nella faccia e nella volta cranica, e sono quindi probabilmente meglio definiti come Homo sp. arcaico antico. Altri fossili di Homo sp. arcaico antico sono stati rinvenuti a Swanscombe in Gran Bretagna, a Steinheim in Germania, a Arago e Montmaurin in Francia, a Sima de los Huesos e Atapuerca in Spagna, a Petralona in Grecia e a Zuttiyeh in Israele. Probabilmente anche gli esemplari rinvenuti recentemente a Boxgrove, in Gran Bretagna, e a Bari, in Italia, appartengono a questo gruppo.

L'Uomo di Neandertal è probabilmente il taxon di Homo sp. arcaico più studiato ed è quello per cui ci sono le maggiori controversie (fig. 7). Questo Homo sp. arcaico recente, che abitava l'Europa, il Medio Oriente e l'Asia orientale, condivide le fattezze di Homo sp. arcaico precedentemente descritte, ma possiede anche alcune caratteristiche craniofacciali e forse postcraniali uniche, che lo contraddistinguono come clado a sé stante. Tali caratteristiche includono una faccia media marcatamente prognata, mascellare rigonfio, apertura nasale larga e sporgente, tori sopraccigliari a doppio arco che contengono ampi seni frontali, una larga prominenza iugomastoidea, un tubercolo mastoideo anteriore, un appiattimento lamboideo combinato con un piano occipitale curvo che gli conferisce il tipico aspetto a 'crocchia', protuberanze occipitali a doppio arco e fossa soprainiaca, molari taurodonti e uno spazio retromolare tra l'ultimo molare mandibolare e il margine anteriore del ramus (Stringer, 1985; Condemi, 1991; Wolpoff, 1996). Un carattere derivato probabilmente esclusivo (autapomorfo) dello scheletro postcraniale dell'Uomo di Neandertal è un ramo pubico superiore allungato in senso anteroposteriore (Trinkaus, 1976). È significativo che alcuni fossili del Pleistocene medio del primo Homo sp. arcaico rinvenuti in Europa, come quelli di Atapuerca, presentino molte, ma non tutte, le autapomorfie di H. neanderthalensis (Arsuaga et al., 1993). Fossili che possono ragionevolmente essere attribuiti a questa specie compaiono nel tardo Pleistocene medio, a partire da circa 200 mila anni fa, nei siti di Biaché, Fontéchevade e La Chaise, in Francia. Tuttavia, l'Uomo di Neandertal è meglio conosciuto per i reperti rinvenuti nei depositi dell'ultimo interglaciale e del Pleistocene superiore, che hanno fornito i resti di più di 275 individui in circa 70 siti (Klein, 1989). Alcuni degli esemplari più noti o più completi rinvenuti in Europa provengono da Spy in Belgio, Neandertal in Germania, La Ferrassie, La Chapelle aux Saints, La Quina e Regourdou in Francia, Krapina in Croazia, Monte Circeo e Saccopastore in Italia e Forbe's Quarry a Gibilterra. Resti dell'Uomo di Neandertal sono stati trovati anche in Oriente fino in Uzbekistan, a Teshik-Tash, e in zone meridionali, fino in Iraq, nella grotta di Shanidar. Alcuni resti rinvenuti in Israele, nelle grotte di Tabun, Kebara e Amud, mostrano leggere differenze nella morfologia craniofacciale rispetto a H. neanderthalensis europeo 'classico', ma molto probabilmente appartengono allo stesso clado.
Finora, i fossili più antichi con caratteri derivati di H. sapiens moderno sono stati rinvenuti nell'Africa sub-sahariana e forse in Medio Oriente, sebbene lo status tassonomico di tali fossili sia stato enormemente dibattuto (ChurchilI et al., 1996). Resti craniofacciali e frammenti di scheletro postcraniale con fattezze moderne sono stati trovati in depositi del Pleistocene medio, che risalgono probabilmente a più di 100 mila anni fa, nei siti di Klasies River Mouth e Border Cave in Sudafrica, e di Orno Kibish, in Etiopia (v. figura 6). Altri resti, denti e frammenti di scheletro postcraniale di età confrontabile sono stati rinvenuti in Sudafrica nei siti di Equus Cave, di Die Kelder Cave e di Sea Harvest, e in Tanzania in località Mumba Rock Shelter. Inoltre, fossili che presentano caratteri derivati del moderno H. sapiens e che le tecniche radiometriche fanno risalire a 81 ÷ 101 mila anni fa, sono stati recuperati in grotte a Qafzeh e Skhul in Israele (Bar-Yosef, 1994). È interessante osservare che tali fossili di H. sapiens sono indiscutibilmente antecedenti ai campioni di Neandertal che provengono dalle stesse regioni (a Tabun, Kebara e Amud) e che datano tra gli 80 e i 40 mila anni fa (Grun e Stringer, 1991).
Finora nessun H. sapiens moderno più vecchio di 40 ÷ 60 mila anni è stato mai rinvenuto in Europa o in Asia centrale e orientale. Resti di H. sapiens compaiono tra le testimonianze fossili dell'Europa centromeridionale: forse possono essere datati a più di 43 mila anni fa quelli della grotta di Bacho Kiro in Bulgaria, e tra 34 e 36 mila anni fa quelli di Hahn6fersand in Germania, di Velika Pecina in Croazia e di Mladec nella Repubblica Ceca (Smith et al., 1989). I primi H. sapiens moderni, determinati con certezza e rinvenuti in Europa occidentale, sono quelli di Cro-Magnon in Francia, che risalgono a circa 30 mila anni fa. Una possibile contemporaneità tra i primi uomini moderni e l'Uomo di Neandertal potrebbe essersi verificata nell'Europa centrale e occidentale, come indicano i fossili di H. neanderthalensis più recenti, quelli di Vindija in Croazia, che risalgono a 33 mila anni fa (Karavanic, 1995) e quelli di 30 mila anni fa di Zafarraya, in Spagna (Hublin et al., 1996). In Cina i fossili più antichi che si possono attribuire con certezza al moderno H. sapiens provengono dal sito di Liujiang e dalla grotta superiore di Zhoukoudian e risalgono rispettivamente a circa 60 mila anni i primi, e tra 27 e 33 mila anni fa i secondi (Wu e Poirier, 1995). In Australia, dove non sono mai stati trovati resti di Homo sp. arcaico, i fossili più vecchi di H. sapiens moderno provengono dal Lago Mungo e risalgono a circa 35 mila anni fa. Tali reperti sono estremamente moderni per quanto riguarda la morfologia e più vecchi dei più robusti - ma forse deformati artificialmente - fossili di H. sapiens recuperati a Kow Swamp, Cohuna e Keilor. Le testimonianze archeologiche, tuttavia, suggeriscono che l'occupazione umana del continente australiano possa risalire almeno a 60 mila anni fa (Roberts et al., 1984).
Filogenesi
Homo antico
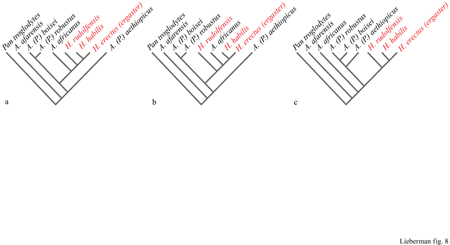
Tre recenti analisi cladistiche principali (W ood, 1991; Lieberman et al., 1996; Strait et al., 1997) forniscono una possibile base per risolvere le relazioni cladistiche, e quindi filogenetiche, tra i primi Homo. Le differenze significative tra i tre cladogrammi (fig. 8), cioè i diagrammi relativi alle analisi cladistiche, mettono in evidenza alcuni problemi tassonomici che devono essere analizzati e revisionati con attenzione. In particolare, le notevoli differenze tra questi e altri cladogrammi fanno pensare che vi sia una bassa probabilità che siano corretti; infatti essi incorporano un'alta incompatibilità tra i caratteri che potrebbe essere il risultato di omoplasie (per esempio, convergenza), ma potrebbe anche dipendere dall'uso di caratteri morfologici poco definiti che hanno scarsa base genetica e che quindi non forniscono prove attendibili da cui dedurre o valutare le relazioni filogenetiche.
Uno dei principali motivi di disaccordo è la relazione delle varie specie di Homo antico tra loro e con gli australopiteci. Sebbene sembri probabile che le specie H. habilis s.s. e H. rudolfensis appartengano entrambe al clado Homo, distinto dal clado degli australopiteci robusti (a volte denominato come genere Paranthropus), molti ricercatori hanno notato che la specie H. rudolfensis possiede alcuni tratti derivati degli australopiteci robusti, in particolare da Australopithecus boisei, suggerendo che essa potrebbe essere stata impropriamente assegnata al genere Homo (Lieberman et al., 1996). Un cladogramma di questo tipo suggerirebbe che, negli ominidi, cervelli di grandi dimensioni si siano evoluti più volte. In alternativa, se H. rudolfensis appartenesse al genere Homo, molte delle somiglianze che questo presenta nella faccia e nei denti con A. boisei e Australopithecus robustus dovrebbero essere esempi di convergenza evolutiva. Una seconda questione di considerevole importanza filogenetica è la relazione evolutiva di H. habilis e H. rudolfensis con la prima specie africana H. erectus (H. ergaster). Alcuni studi (Wood, 1991; Lieberman et al., 1996) sottolineano le sinapomorfie (caratteristiche comuni a diversi cladi collegati tra loro) facciali e dentali tra H. habilis s.s. e H. erectus, mentre D. Strait e collaboratori (1997) considerano la dimensione assoluta del cervello come il carattere derivato significativo che lega H. rudolfensis con H. erectus.
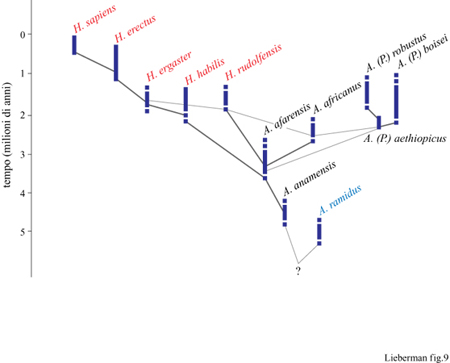
Una possibile filogenesi è basata sulle analisi cladistiche precedentemente discusse (fig. 9). Sebbene molti dettagli necessitino ancora di un chiarimento, alcuni punti meritano un'attenzione particolare. Prima di tutto, è chiaro che le origini del genere Homo sono parte di una radiazione generale degli ominidi avvenuta alla fine del Pliocene, probabilmente in relazione a un cambiamento globale del clima (Vrba, 1993). Tra 2,5 e 1,8 milioni di anni fa in Africa comparvero almeno sei specie di ominidi, tre delle quali sono forse membri del genere Homo. In secondo luogo, pensiamo sia probabile (sebbene difficilmente accertabile) che delle due specie di Homo antico quella con la corporatura più piccola, H. habilis s.s., sia l'antenato del primo H. erectus africano (H. ergaster), apparso approssimativamente 1,8 milioni di anni fa o forse prima. Tuttavia la relazione che proponiamo pone due problemi principali: l'elevata somiglianza dello scheletro postcraniale di H. habilis con quello degli australopiteci e la possibile sovrapposizione temporale di H. habilis con H. ergaster. Inoltre anche altri problemi che riguardano la relazione tra H. ergaster e H. erectus richiedono una soluzione: se per esempio, in accordo con B. Wood (1991), H. ergaster fosse una specie distinta da H. erectus, allora non ci sarebbero prove sufficienti per stabilire quale dei due sia stato il primo ominide a migrare fuori dall'Africa. Infine è anche possibile che la specie H. erectus si sia evoluta da H. ergaster in Asia e debba essere circoscritta ai fossili rinvenuti in questa regione; questo implicherebbe la possibilità che H. erectus sia un ramo evolutivo laterale e non un antenato di H. sapiens.
Homo recente

La filogenesi del genere Homo più recente è, e non ci stupisce, altrettanto controversa e irrisolta quanto la sua tassonomia. Molti ricercatori e in particolare D. Frayer e i suoi collaboratori (1993), che considerano Homo sp. arcaico e H. sapiens come taxa conspecifici basati sul grado di somiglianza, hanno suggerito l'assenza di vere e proprie linee filogenetiche nell'ambito del genere Homo recente, e teorizzano invece una evoluzione multiregionale. In accordo con tale teoria, H. sapiens sarebbe una specie polimorfa estremamente variabile che si è evo Iuta in modo graduale e policentrico in diverse aree geografiche durante il Pleistocene medio e superiore; l'elevato livello di flusso genico e di mobilità delle popolazioni avrebbe, infatti, impedito la speciazione di questi gruppi geograficamente circoscritti di ominidi (Wolpoff, 1996). L'ipotesi multiregionale sostiene pertanto che le relazioni evolutive tra taxa di Homo recente non possano essere riassunte o studiate con metodi cladistici (Wolpoff e Crummett, 1995). Una posizione contrastante, spesso chiamata ipotesi della origine africana recente (RAO, Recent African Origin), è stata sostenuta da ricercatori quali I. Tattersall (1986), C. Stringer e P. Andrews (1988), D. Lieberman (1995) e altri. Secondo tale ipotesi, H. sapiens moderno si sarebbe evoluto monocentricamente come linea evolutiva distinta in un'unica regione del mondo, probabilmente in Africa o in Medio Oriente, e avrebbe successivamente rimpiazzato le popolazioni preesistenti di Homo sp. arcaico in altre zone (fig. 10).
Finché le questioni che riguardano la variabilità in H. erectus s.I., Homo sp. arcaico e i primi H. sapiens non saranno risolte, ricostruire accuratamente la filogenesi dei rappresentanti del genere Homo più recenti sarà praticamente impossibile. Ciononostante, diverse evidenze suggeriscono che il moderno H. sapiens possa essere un clado distinto che si è evoluto in Africa o in Medio Oriente. Innanzitutto, i fossili più antichi che mostrano la maggior parte dei caratteri derivati di H. sapiens anatomicamente moderno (v. sopra), provengono chiaramente dall'Africa e dal Medio Oriente, e non dall'Europa o dall'Asia. In contrapposizione, i primi e più antichi fossili europei (con la possibile eccezione della mandibola di Dmanisi, di 1,4 milioni di anni fa) condividono alcuni tratti derivati solo con i primi rappresentanti di Homo sp. arcaico. Quindi, basandosi sulle testimonianze correnti, sembra probabile che l'antico Homo sp. arcaico, e non H. erectus, abbia colonizzato per primo l'Europa e che tale linea abbia dato origine all'Uomo di Neandertal. Inoltre, in Asia orientale, a Hexian, e in Indonesia, a Ngandong, H. erectus sembra rimanere ancora a lungo dopo la comparsa, più a occidente, di Homo sp. arcaico, dando indicazioni del persistere di entrambe le specie fino al Pleistocene superiore. Tuttavia, altri individui asiatici che presentano caratteri leggermente più moderni, come quelli rinvenuti a Maba e Dali in Cina, potrebbero indicare che in Asia, durante il Pleistocene medio, siano forse vissute due specie di ominidi oppure una specie caratterizzata da elevata variabilità. Verso la fine del Pleistocene medio, dunque, il Vecchio Mondo era apparentemente popolato da almeno tre taxa morfologicamente distinti: H. sapiens in Africa e nel Medio Oriente, Homo sp. arcaico in Europa e in parte dell'Asia e H. erectus nell'Asia orientale. Il grado di variabilità tra questi taxa contemporanei e strettamente correlati è probabilmente troppo grande per poterli incorporare tutti in un'unica specie.
Ulteriori prove a favore del fatto che H. sapiens si sia evoluto recentemente e come linea a sé stante vengono da studi cladistici e fenetici sulla variabilità regionale all'interno di popolazioni recenti e fossili di Homo sp. recente. Sebbene coloro che propongono l'ipotesi multiregionale puntino sull'esistenza di tratti 'razziali' regionali, che si suppone derivino da una continuità evolutiva regionale tra Homo sp. arcaico e le popolazioni moderne, è stato obiettato che tali tratti o siano in realtà caratteristiche disperse globalmente oppure che possano essere caratteri primitivi che non forniscono alcuna informazione sulle relazioni evolutive (Groves, 1989; Lahr, 1994; Lieberman, 1995). Al contrario, le analisi cladistiche e fenetiche sia di fossili sia di popolazioni moderne indicano che queste ultime sono caratterizzate da un grado estremamente basso di variabilità morfologica che è difficile spiegare considerando i lunghi tempi della separazione e dell'evoluzione regionale (Howells, 1989; 1995). Anche studi di genetica hanno dato risultati in favore dell' origine africana recente delle moderne popolazioni umane, fornendo quindi ampio supporto al modello RAO. In particolare, analisi recenti delle variazioni nel DNA nucleare e mitocondriale, sebbene controverse, indicano che la variabilità genetica che si riscontra tra le diverse popolazioni umane è una piccola frazione rispetto alla variabilità genetica presente all'interno delle singole popolazioni, e che gli africani sono molto diversi fra loro dal punto di vista genetico; è dunque assai probabile che essi siano gli ultimi antenati comuni di tutte le popolazioni viventi.
Uno scenario alternativo, noto come diffusione demica o modello africano di ibridazione e sostituzione (african hybridization and replacement model; Smith et al., 1989; Braiier, 1992), suggerisce che H. sapiens sia comparso inizialmente in Africa o in Medio Oriente, ma che poi si sia incrociato con Homo sp. arcaico piuttosto che averne rimpiazzato completamente le popolazioni nel resto del Vecchio Mondo. In altre parole, un certo grado di flusso genico o di mobilità delle popolazioni potrebbe aver caratterizzato le interazioni tra popolazioni durante l'espansione geografica di H. sapiens nel Pleistocene superiore. Tale scenario è estremamente difficile da verificare senza ulteriori accertamenti tassonomici. Tuttavia, se Homo sp. arcaico e il primo H. sapiens moderno erano conspecifici e si incrociavano tra loro, allora ci si dovrebbe aspettare di trovare dei fossili che combinino le caratteristiche peculiari dei tratti precedentemente descritti di entrambi i taxa: fino a oggi, nessun fossile con tali caratteristiche è mai stato rinvenuto o documentato, anche se tale possibilità richiede ulteriori ricerche.
Ricostruzione comportamentale
Postura e locomozione
Nessun elemento di scheletro postcraniale è direttamente associato con i reperti cranici di H. rudolfensis, lasciando senza risposta molte domande sulla taglia corporea, la locomozione e gli adattamenti posturali di questa specie. Tuttavia, alcuni grossi resti di ossa delle gambe, inclusi KNM - ER 813, 1472 e l48lA, contemporanei del cranio KNM-ER 1470, indicano la possibilità che H. rudolfensis possedesse uno scheletro postcraniale derivato del tipo Homo (Lieberman et al., 1996). Qualcosa di più si sa di H. habilis s.s. grazie al rinvenimento di uno scheletro assai frammentario, OH 62, nella gola di Olduvai (Johanson et al., 1987), di alcune ossa della mano associate a un teschio incompleto, OH 7, e di alcuni frammenti postcraniali isolati, incluso un piede quasi completo, OH 8. Tali resti suggeriscono che H. habilis, come Australopithecus afarensis, fosse un bipede eretto abituale, di bassa statura e piccola taglia corporea, di dimensioni analoghe allo scimpanzé (McHenry, 1992), che conservava una serie di adattamenti per la locomozione arboricola come lunghe braccia, gambe corte e ossa delle mani ricurve, per potersi muovere in modo efficiente sospeso ai rami (Susman e Stern, l 982).
l resti di scheletro postcraniale dei primi rappresentanti di H. erectus (H. ergaster) africano, e in particolare lo scheletro quasi completo di Nariokotome (Walker e Leakey, 1993), indicano senza ambiguità che questa specie rappresenta un chiaro mutamento verso una morfologia adatta alla locomozione più moderna (fig. 11). H. erectus è il primo ominide che ha uno scheletro postcraniale in molti aspetti simile a quello di H. sapiens e che non presenta gli adattamenti per muoversi tra gli alberi descritti prima. Questa specie ha un torace più cilindrico, dita dei piedi e delle mani corte e diritte e una colonna vertebrale che si curva anteriormente nella regione toracica e posteriormente sul collo e sopra il bacino. Lo scheletro postcraniale di H. erectus, tuttavia, differisce da quello dell'uomo moderno in quanto presenta un bacino più stretto, un collo del femore più lungo e ossa degli arti più robuste con una ridotta cavità midollare (Ruff et al., 1993; Walker e Leakey, 1993). l crani di Homo più antichi mostrano una serie di adattamenti specifici per la corsa, compresi una accentuazione del canale semicircolare verticale dell' orecchio interno che aiuta a mantenere l'equilibrio in posizione verticale (Spoor et al., 1994), un legamento nucale che stabilizza la testa contro lo spostamento inerziale in avanti causato dalla corsa e un grande naso esterno che permette uno scambio efficiente di calore e umidità durante la respirazione (Franciscus e Trinkaus, 1988).
Dopo la dispersione iniziale degli ominidi fuori dall'Africa, cominciarono a emergere le differenze regionali nella forma del corpo caratteristiche delle popolazioni viventi. Queste variazioni di statura, dimensioni del tronco e proporzioni degli arti si possono facilmente interpretare in termini di meccanismi di termoregolazione che rendono gli individui adatti ai diversi regimi climatici (Ruff, 1994). l resti di scheletri postcraniali sono scarsamente rappresentati nei documenti fossili, fatta eccezione per gli ominidi europei del Pleistocene superiore, e quindi le generalizzazioni si devono ricavare da pochi reperti isolati. Lo scheletro di H. erectus (ergaster) di Nariokotome, di 1,6 milioni di anni fa, così come i fossili del Pleistocene medio di Homo sp. rinvenuti a Kabwe, presentano statura alta, tronco stretto e hanno lunghi segmenti degli arti distali caratteristici delle popolazioni moderne che vivono nelle regioni calde e secche dell'Africa equatoriale (Ruff, 1994). l primi abitanti dell'Europa, d'altra parte, hanno una forma del corpo più corta e più tozza, meno assottigliata, come è tipico delle popolazioni adattate ad ambienti caratterizzati da stagioni fredde. Ciò potrebbe essere vero per gli abitanti di mezzo milione di anni fa di Boxgrove, nella Gran Bretagna meridionale (Trinkaus et al., 1996), ed è sicuramente il caso dell'Uomo di Neandertal. Questi ominidi, che hanno vissuto nelle regioni periglaciali tra 200 e 30 mila anni fa, erano relativamente bassi di statura, ma avevano una corporatura massiccia, con un ampio tronco a forma di botte e arti spessi e muscolosi con segmenti distali corti (Holliday, 1995). Fisionomie simili si riscontrano nelle popolazioni artiche moderne, come gli eschimesi, e, in misura minore, in altre popolazioni viventi europee. L'Uomo di Neandertal della costa più calda del Mediterraneo, nel Vicino Oriente, aveva una fisionomia simile, sebbene con forme meno marcate. Sfortunatamente, l'anatomia postcraniale dell'Homo sp. arcaico dell'Asia è di fatto sconosciuta.
l resti di scheletro postcraniale dei primi umani moderni africani sono scarsi e finora non vi è nulla tra i reperti rinvenuti in siti quali Orno Kibish e Klasies River Mouth, che suggerisca che tali popolazioni fossero sostanzialmente differenti nella forma del corpo da quelle dei moderni africani. l primissimi umani moderni del Vicino Oriente e d'Europa, a differenza dell'Uomo di Neandertal di queste regioni, avevano proporzioni del corpo più simili a quelle delle popolazioni tropicali adattate al caldo e quindi statura maggiore, tronco stretto e lunghi segmenti distali degli arti. T. Holliday (1995) ha suggerito che tali cambiamenti indicano un aumento di flusso genico dalle regioni più calde, forse sotto forma di sostituzione di popolazioni, al momento della comparsa dell 'uomo moderno al di fuori dell'Africa. Durante l'ultimo massimo glaciale (circa 18 ÷ 17 mila anni fa) le popolazioni europee moderne hanno evoluto nuovamente una forma corporea di tipo artico, adattata al freddo. Quindi le somiglianze fra la forma del corpo dell'Uomo di Neandertal e quella degli europei viventi potrebbero riflettere l'evoluzione parallela di un morfotipo artico in risposta a simili eventi climatici stressanti.
Uno scheletro quasi completo di H. neanderthalensis e alcuni resti di individui contemporanei rinvenuti al di fuori dell'Europa indicano che, fino a tempi relativamente recenti, tutti i taxa di Homo tardivo avevano uno scheletro robusto con spesse ossa piatte, grandi superfici articolari e siti di attacco dei muscoli grandi e resistenti. Tuttavia, poiché molte caratteristiche dello scheletro si adattano continuamente alle variazioni del carico meccanico abituale, aggiungendo o riassorbendo osso a seconda del bisogno, molto probabilmente tale robustezza riflette l'adattamento a elevati livelli di stress biomeccanico sullo scheletro, probabile conseguenza di alti livelli di mobilità e di lavoro faticoso (Ruff, 1992). In particolare, le analisi dei resti postcraniali indicano che sia l'Uomo di Neandertal sia i primi uomini moderni avevano gambe strutturalmente adatte ad alti livelli di carico, suggerendo un alto grado di mobilità (Ruff et al., 1993). La robustezza degli arti superiori, tuttavia, differisce tra Homo sp. arcaico e i primi H. sapiens (Churchill, 1994): Homo sp. arcaico possiede arti che tendono a essere più robusti. Tale contrasto suggerisce che si sia verificata una modificazione nella quantità di carico sugli arti superiori, forse in relazione a un cambiamento delle pratiche di sussistenza al tempo della prima comparsa dell 'uomo moderno. Tuttavia, sia i fossili umani arcaici sia quelli moderni sono caratterizzati da un alto livello di asimmetria bilaterale nella robustezza degli arti superiori (Trinkaus et al., 1994), che potrebbe riflettere l'uso di tecniche di caccia simili (per esempio, lance scagliate con un solo braccio) in entrambi questi gruppi. L'Uomo di Neandertal, inoltre, ha un'apertura nasale insolitamente ampia e una voluminosa cavità toracica, che potrebbero indicare una forte necessità respiratoria dovuta a elevati livelli di attività in ambienti freddi e secchi (Franciscus, 1989).
Tecnologia e sussistenza
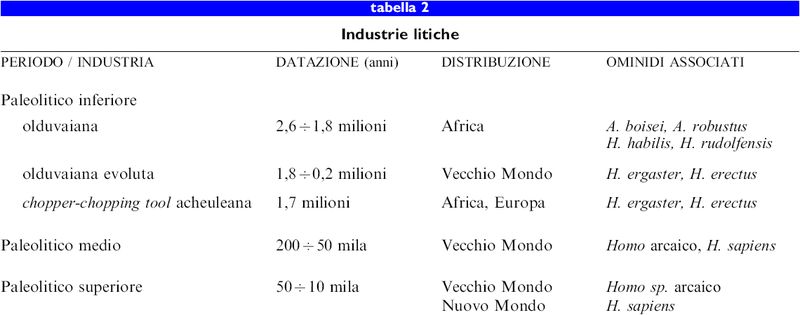
Sebbene sia quasi certo che i primi ominidi, come gli scimpanzé, producessero e usassero strumenti di materiale deperibile come il legno, la più antica testimonianza della produzione di utensili duraturi in pietra viene dai siti archeologici dell'Africa orientale che risalgono approssimativamente a 2,6 milioni di anni fa. Tali strumenti, che vengono classificati come la prima variante dell'industria olduvaiana del Paleolitico inferiore, consistono in semplici ciottoli scheggiati dai quali, per percussione, venivano rimosse una o più schegge per creare un bordo tagliente. l nuclei così ottenuti e le schegge rimosse venivano inizialmente usati per tagliare materiali vegetali e per scarnificare carcasse di animali (Toth, 1987; Blumenschine et al., 1994). Non ci sono al momento prove certe su chi realizzò i primi strumenti o con quale frequenza questi venissero utilizzati; si presume che venissero fabbricati da H. habilis o da H. rudolfensis, ma forse anche dai robusti australopitecini, presenti contemporaneamente in Africa orientale (Susman, 1988). L'industria olduvaiana è stata rinvenuta in numerosi siti dell'Africa orientale e meridionale e risale a più di 1,7 milioni di anni fa, quando comparvero per la prima volta due nuove industrie del Paleolitico inferiore, quasi contemporaneamente all'evoluzione di H. erectus (ergaster). La più famosa di queste, l'industria acheuleana, è caratterizzata dalla produzione di asce a mano simmetriche, in cui numerose schegge venivano staccate da un nucleo per creare un grande strumento bifacciale, e le numerose piccole schegge taglienti rimosse costituivano a loro volta degli strumenti. Siti acheuleani sono stati rinvenuti soltanto in Africa, Europa e Medio Oriente e non sono presenti in Asia orientale. Un'altra industria del Paleolitico inferiore - conosciuta in Africa come industria olduvaiana evoluta, in Asia come industria del chopper-chopping tool e in Europa da varianti regionali quali industria clactoniana e industria tayaziana - è all'incirca contemporanea all'acheuleana, ma non produceva le asce a mano. Industrie non acheuleane del Paleolitico inferiore sono state trovate in gran parte del Vecchio Mondo, compresa l'Asia orientale (tab. 2). I pochi resti archeologici del Paleolitico inferiore associati ai primi Homo non forniscono indicazioni su un grado di complessità cognitiva e tecnologica paragonabile a quello raggiunto successivamente nel Paleolitico medio e superiore. Durante il periodo di Olduvai molto, se non tutto, l'approvvigionamento di carne derivava probabilmente dalla scarnificazione delle carcasse di animali piuttosto che dalla caccia (Blumenschine e Cavallo, 1992) ed è probabile che la dieta di H. rudolfensis e di H. habilis fosse in gran parte vegetariana. Sebbene esistano alcune testimonianze di attività di caccia da parte di H. erectus durante l'Acheuleano e l'Olduvaiano avanzato, cibarsi delle carcasse di animali potrebbe aver costituito ancora una strategia importante per l'acquisizione di proteine animali. La tecnica di caccia durante il Paleolitico inferiore era semplice e gli strumenti di pietra erano probabilmente usati più per macellare le carcasse che per cacciare. Testimonianze sull'uso e il controllo del fuoco da parte di H. erectus potrebbero essere quelle di Zhoukoudian in Cina, 500 mila anni fa, di Terra Amata in Francia, circa 400 mila anni fa, e forse dell'Africa meridionale, 1,6 milioni di anni fa. Le ipotesi sulle capacità linguistiche e cognitive delle specie più antiche di Homo sono molto speculative e difficilmente verificabili.
Tra 200 e 130 mila anni fa in Africa, Asia occidentale ed Europa, le industrie del Paleolitico inferiore aprirono la via alle strumentazioni, in qualche modo più sofisticate, del Paleolitico medio. La natura di tale evoluzione nell'Asia orientale è scarsamente documentata e datata. Denominate industria musteriana in Europa, industria levalloisiana-musteriana nel Vicino Oriente ed età della Pietra media in Africa, queste industrie del Paleolitico medio, sebbene diversificate, presentano un maggior numero di strumenti di diverso tipo e mostrano una tecnologia più avanzata rispetto a quelle precedenti. Tali innovazioni, che compaiono in molti siti a partire da 400 mila anni fa ma che diventano sempre più frequenti nel Paleolitico medio, includono l'uso diretto di un percussore più morbido per rimuovere schegge dai nuclei e l'uso di questi per produrre schegge di forma e dimensione predeterminate. Gli utensili del Paleolitico medio sono associati a Homo sp. arcaico in molte regioni, ma erano prodotti anche dai primi umani moderni in Africa e nel Vicino Oriente.
Molti degli utensili del Paleolitico medio, come coltelli, punte e raschiatoi, servivano probabilmente per procacciarsi e lavorare le risorse animali, come la carne e le pelli (Shea, 1989). Tuttavia, l'abilità nella caccia della gente del Paleolitico medio resta un problema dibattuto. Le rappresentazioni di parti di animali nel Musteriano, le associazioni faunistiche dell'età della Pietra media e le ossa lavorate dall'uomo (ossa con segni di taglio), che presentano ferite causate da carnivori, hanno indotto L. Binford (1984, 1985 e 1989) a supporre che il cibarsi delle carcasse, in particolare di animali di grossa taglia, fosse una componente importante nell' approvvigionamento di carne da parte degli umani del Paleolitico medio. Binford riconosce la possibile esistenza di battute di caccia regolari su piccola e media scala nel Paleolitico medio, ma sostiene che gli umani arcaici o addirittura anche quelli moderni potrebbero non aver avuto la raffinatezza comportamentale e tecnologica sufficiente per cacciare regolarmente parecchi tipi di prede. Siti musteriani in alcune regioni d'Europa mettono in evidenza l'esistenza di un tipo di caccia focalizzata su un solo animale, indicando così la mancanza di battute di caccia cooperative e uccisioni di massa. Ciò potrebbe riflettere l'uso di pratiche di caccia opportunistiche (Freeman, 1973; Binforcl, 1982; Straus e Clark, 1986). Tali argomenti hanno dato luogo a una visione generale dello stile di vita dell'uomo arcaico che appare poco razionale e privo di un comportamento tattico ben pianificato e di attenzioni durature alle tecnologie per la sussistenza (Binford, 1989).
Un comportamento propriamente razionale di ricerca del cibo, quindi, potrebbe essere una caratteristica dell'uomo pienamente moderno, emersa con il moderno H. sapiens, se non più tardi (Binford, 1985). Le differenze nella capacità di procurarsi le risorse e nel tempo dedicato al lavoro per la sussistenza, potrebbero spiegare la differente robustezza degli arti superiori nell'uomo arcaico e in quello moderno. È importante notare, tuttavia, che esistono numerose argomentazioni a sostegno della scarsa differenza tra l'organizzazione per la sussistenza e il comportamento di caccia durante il periodo che va dal Paleolitico medio a quello superiore (Chase, 1986, 1988 e 1989; Farizye David, 1989; Patou, 1989). Gli studi di A. Jelinek e collaboratori (1989) hanno evidenziato alcune testimonianze che si pensa rappresentino uccisioni di massa nel Musteriano francese. Vi sono anche delle prove che, in alcune regioni dell'Italia centrale, una riduzione nella frequenza dell'abitudine di cibarsi delle carcasse e un intensificarsi del comportamento di caccia (Stiner, 1994; Stiner e Kuhn, 1992) si siano verificati durante il Musteriano piuttosto che durante la transizione tra il Paleolitico medio e quello superiore. A tutt'oggi, la natura del comportamento di sussistenza nel Paleolitico medio rimane oscura.
I comportamenti simbolici sembrano essere poco sviluppati nel Paleolitico medio. Forse l'Uomo di Neandertal seppelliva alcuni, ma non la maggior parte, dei morti e non è certo in che misura i suoi contemporanei fuori dall'Europa facessero lo stesso, anzi tale pratica sembra per loro ancora meno frequente. Le forme di espressione artistica non sono molto frequenti e non è affatto noto fino a che punto questi ominidi usassero una forma di linguaggio articolata.
Tra 45 e 30 mila anni fa, le industrie del Paleolitico medio vennero sostituite da quelle del Paleolitico superiore (tarda età della Pietra in Africa) che presentano una varietà maggiore di tipi di utensili, una accresciuta produzione di lame finemente lavorate e un incremento nella lavorazione di prodotti in osso e como. Mentre una delle prime industrie del Paleolitico superiore, l'industria castelperroniana nel sud-ovest della Francia, è associata con l'Uomo di Neandertal, molti dei siti del Paleolitico superiore sono associati esclusivamente con resti di H. sapiens moderno. Altri cambiamenti nel Paleolitico superiore includono la comparsa di un numero crescente di armi sofisticate e diverse per la caccia e di oggetti elaborati che servivano come ornamenti personali e artistici.
Conclusioni
Il genere Homo si è evoluto come parte di un generale evento di radiazione degli ominidi alla fine del Pliocene. Le prime specie di Homo, compresi H. habilis, H. rudolfensis, H. ergaster e H. erectus, sono tipicamente, ma non sempre, caratterizzate da un cervello più grande in rapporto alle dimensioni del corpo, una faccia piccola e una serie di adattamenti per la locomozione bipede. Presumibilmente, alcuni di tali adattamenti permisero a H. erectus (forse H. ergaster) di migrare fuori dall'Africa, attraverso il Vecchio Mondo, e colonizzare gli habitat temperati. Eventi evolutivi successivi diedero luogo a una varietà di taxa con cervello di dimensioni maggiori, attualmente raggruppati sotto il nome di Homo sp. indet. arcaico, del quale la tassonomia e le relazioni evolutive restano oscure e ampiamente dibattute. Testimonianze fossili e testimonianze genetiche indicano che a un certo punto, tra 100 e 200 mila anni fa, in Africa o in Medio Oriente si è evoluto il moderno H. sapiens, che ha sostituito in gran parte i taxa di Homo preesistenti con tempi diversi in luoghi diversi. Questi uomini moderni hanno una configurazione craniofacciale particolare che non è correlata alle dimensioni del cervello, ma che potrebbe invece essere correlata ad alcuni adattamenti dovuti ai cambiamenti nella dieta e nello stile di vita, o alla produzione del linguaggio. Le testimonianze archeologiche delle variazioni comportamentali hanno scarsissime relazioni con i dati paleontologici.
Forse le due domande più interessanti al centro delle teorie sull'evoluzione del genere Homo sono il modo e le ragioni per cui il moderno H. sapiens ha rimpiazzato le popolazioni contemporanee di Homo arcaico come l'Uomo di Neandertal, e quanto tali eventi evolutivi sono in relazione con l'origine delle popolazioni moderne. Attualmente, la maggior parte dei fossili e dei dati genetici indicano che gli umani moderni si sono evoluti abbastanza recentemente e che le piccole differenze genetiche e morfologiche che si riscontrano tra le popolazioni viventi sono il risultato di eventi molto recenti, forse meno di 50 mila anni nella maggior parte del mondo. Tuttavia, per una soluzione più soddisfacente di questo problema è necessario un approccio più approfondito, che risolva i problemi tassonomici e filogenetici legati alla grande diversità morfologica tra le specie del genere Homo. I tentativi di distinguere i differenti taxa del genere Homo esclusivamente sulla base della variabilità morfologica si sono rivelati in gran parte inutili, perché solo una piccola parte dei caratteri conservati nei resti fossili fornisce informazioni accurate sulle relazioni progenitore/discendente in taxa strettamente correlati. È probabile che un approccio maggiormente basato su aspetti dello sviluppo sia in grado di offrire migliori contributi, aiutando a distinguere i caratteri che sono il risultato di stimoli genetici da quelli che non lo sono.
Lo studio della filo genesi delle specie umane rimane alle frontiere della biologia perché le domande che essa pone vanno al di là dell' attuale capacità esplicativa dei nostri dati, pur rivestendo un grande significato. La risoluzione delle relazioni evolutive tra le popolazioni umane viventi è cruciale per la comprensione dei dati sulla variabilità genetica umana che si stanno velocemente accumulando (per esempio, con il Progetto Genoma Umano), così come per la comprensione del modo in cui operano i processi evolutivi in animali culturali altamente dispersi e complessi dal punto di vista comportamentale. l paleontologi che studiano gli ominidi necessitano non solo di fossili più completi dal mondo intero, ma anche di integrare le tradizionali analisi morfologiche e metriche di tali fossili con studi funzionali e sullo sviluppo degli scheletri in taxa viventi. Inoltre, servono tecniche migliori per distinguere con precisione i taxa degli ominidi fossili in quanto molte, se non tutte, le controversie sulla recente evoluzione umana si fondano principalmente su questioni tassonomiche basilari. Lo studio di chi siamo e di come ci siamo evoluti resta una delle grandi sfide della biologia.
Bibliografia citata
ARSUAGA, J.L., MARTINEZ, I., GRACIA, A., CARRETERO, J.M., CARBONELL, E. (1993) Three new human skulls from Sima de los Huesos Middle Pleistocene site in Sierra de Atapuerca, Spain. Nature, 362, 534-537.
BAR-YOSEF, O. (1994) The contribution of Southwest Asia to the study of the origin of modem humans. In Origins of anatomically modern humans, a c. di Nitecki, M.H., Nitecki, D.V., New York, Plenum Press, pp. 23-66.
BINFORD, L.R. (1984) Faunal remains from Klasies River Mouth. New York, Academic Press.
BINFORD, L.R. (1985) Human ancestors: changing views of their behavior. J. Anth. Arch., 4, 292-327.
BINFORD, L.R. (1989) Isolating the transition to cultural adaptations: an organizational approach. In The emergence of modern humans, a c. di Trinkaus, E., Cambridge, Cambridge University Press, pp. 18-41.
BLUMENSCHINE, R.J., CAVALLO, J.A. (1992) Scavenging and human evolution. Sci. Amer., 267, 90-96.
BLUMENSCHINE, R.J., CAVALLO, J.A., CAPALDO, S.D. (1994) Competition for carcasses and early hominid behavioral ecology: a case study and conceptual framework. J. Rum. Evol., 27, 197-213.
BRAUER, G. (1992) A craniological approach to the origin of anatomically modem Romo sapiens in Africa and implications for the appearance of modem humans. In Continuity or re placemento Controversies in Homo sapiens evolution, a c. di Brauer, G., Smith, F.H., Brookfield, Balkema, pp. 83-98.
CHASE, P.G. (1986) The hunters of Combe Grenal: approaches to Middle Paleolithic subsistence in Europe. Brit. Arch. Rep. IS 286.
CHASE, P.G. (1988) Scavenging and hunting in the Middle Palaeolithic: the evidence from Europe. In The upper Pleistocene prehistory of Western Eurasia, a c. di Dibble, H., Monet-White, A, Philadelphia, University of Peunsylvania Press, pp. 225-232.
CHASE, P.G. (1989) How different was Middle Palaeolithic subsistence? In The human revolution: behavioural and biological perspectives on the origins of modern humans, voI. I, a c. di Mellars, P., Stringer, C.B., Edinburgh, Edinburgh University Press, pp. 321-337.
CHURCHILL, S.E. (1994) Ruman upper body evolution in the Eurasian Later Pleistocene, Ph.D. Thesis, University New Mexico.
CHURCHILL, S.E. (1996) Particulate versus integrated evolution of the upper body of Late Pleistocene humans: a test of two models. Am. J. Phys. Anth., 100, 559-583.
CHURCHILL, S.E., PEARSON, O.M., GRINE, F.E., TRINKAUS, E., HOLLIDAY, T.W. (1996) Morphological affinities of the proximal ulna from Klasies River main site: archaic or modem? J. Rum. Evol., 31, 213-237.
CONDEMI, S. (1991) Some considerations conceming Neandertal features and the River main site: archaic or modem? Riv. Antropol., 69, 27-38.
DAY, M.H., STRINGER, C.B. (1982) A reconsideration of the Orno Kibish remains and the erectus-sapiens transition. In L'Homo erectus et la pIace de l'Romme de Tautavel parmi les hominides Fossiles, Paris, Louis-Jean Scientific and Literary Publications, voI. 2, pp.814-846.
FARIZY, C., DAVID, F. (1989) Chasse et alimentation camée au Paléolitique moyen, l'apport des gisements de plein air. In L'Homme de Néandertal, la subsistance, a c. di Otte, M., Liège, Etudes et Recherches Archéologiques de l'Université de Liège, voI. 6, pp. 59-62.
FRANCISCUS, R.G. (1989) Am. J. Phys. Anthrop., 78, 223.
FRANCISCUS, R.G., TRINKAUS, E. (1988) Nasal morphology and the emergence of Romo erectus. Am. J. Phys. Anth., 75, 578-585.
FRAYER, D.W., WOLPOFF, M.H., THORNE, A.G., SMITH, F.H., POPE, G.G. (1993)
Theories of modem human origins: the palaeontological test. Amer. Anthropol., 95, 14-50.
FREEMAN, L.G. (1973) The significance of mammalian faunas from Paleolithic occupations in Cantabrian, Spain. Am. Antiquity, 38, 3-44.
GROVES, C.P. (1989) A regional approach to the problem of the origin of modem humans in Australasia. In The human revolution: behavioural perspectives in the origins of modern humans, a c. di Mellars, P., Stringer, C.B., Edinburgh, Edinburgh University Press, pp. 245-273.
GRUN, R., STRINGER, C.B. (1991) Electron spin resonance data on the evolution of modem humans. Archaeometry, 33, 153-199.
HARTWIG-SCHERER, S., MARTIN, R.D. (1991) Was 'Lucy' more human than her 'child'? Observations on early hominid postcrani al skeletons. J. Hum. Ev., 21, 439-449.
HILL, A., WARD, S., DEINO, A., CURTIS, G., DRAKE, R. (1992) Earliest Homo Nature, 355, 719-722.
HOLLIDAY, T.W. (1995) Body size and proportions in the Late Pleistocene western old world and the origins of modern humans. Ph.D. Thesis, University New Mexico.
HOWELL, F.C. (1960) European and north west african Middle Pleistocene hominids. Curr. Anthro., l, 195-232.
HOWELL, F.C. (1978) Hominidae. In Evolution of african mammaIs, a c. di Maglio, Y.l, Cooke, H.B.S., Cambridge, Mass., Harvard University Press, pp. 154-248.
HOWELLS, W.W. (1989) Skull shapes and the map. Peabody Museum Papers, 79.
HOWELLS, W.W. (1995) Who's who in skulls. Peabody Museum Papers, 82.
HUANG, W., CIOCHON, R., YUMIN, G., LARICK, R., QIREN, F., SCHWARCZ, H., YONGE, C., DEVOS, J., RINK, W. (1995) Early Romo and associated artefacts from Asia. Nature, 378, 275-278.
HUBLIN, J.J., SPOOR, F., BRAUN, M., ZONNEVELD, F., CONDEMI, S. (1996) A late Neandertal associated with Upper Palaeolithic artefacts. Nature, 381, 224-226.
JELINEK, A.J., DEB NATH, A., DIBBLE, H.L. (1989) A preliminary report on evidence related to the interpretation of economic and social activities of Neandertals at the site of La Quina (Charente), France. In L'Homme de Néandertal, la subsistance, voI. 6, a c. di Otte, M., Liège, Etudes et Recherches Archéologiques de l'Université de Liège, pp. 99-106.
JOHANSON, D.C., MASAO, F.T., ECK, G.G., WHITE, T.D., WALTER, R.C., KIMBEL, W.H., ASFAW, B., MANEGA, P., NDESSOKIA, R., SUWA, G. (1987) New partial skeleton of Romo habilis from Olduvai Gorge. Nature, 327, 205-209.
KARAVANIC, I. (1995) Upper Paleolithic occupation levels and late-occurring Neandertal at Vindija Cave (Croatia) in the context of centraI Europe and the Balkans. J. Anthrop. Res., 51, 9-35.
KLEIN, R.G. (1989) The human career: human biological and cultural origins, Chicago, University of Chicago Press.
LAHR, M.M. (1994) The multiregional model of modem human origins: a reassessment of its morphological basis. J. Hum. Evol., 26, 23-56.
LEAKEY, L.S.B., TOBIAS, P.V., NAPIER, J.R. (1964) A new species of the genus Homo from Olduvai Gorge. Nature, 189, 649-650.
LIEBERMAN, D.E. (1995) Testing hypotheses about human evolution from skulls: integrating morphology, function, development, and phylogeny. Curo Anthro., 36, 159-197.
LIEBERMAN, D.E. (1996) How and why recent humans grow thin skulls: experimental data on systemic cortical robusticity. Am. J. Phys. Anthrop., 101, 217-236.
LIEBERMAN, D.E., PILBEAM, D.R., WOOD, B.A. (1988) A probabilistic approach to the problem of sexual dimorphism in Homo habilis: a comparison of KNM-ER 1470 and KNM-ER 1813. J. Hum. Ev., 17, 503-511.
LIEBERMAN, D.E., PILBEAM, D.R., WOOD, B.A. (1996) Homoplasyand early Homo: an analysys ofthe evolutionary relationships of H. habilis sensu stricto and H. rudolfensis. J. Hum. Ev., 30, 97-120.
MAYR, E. (1950) Where are we? Cold Spring Rarbor Symp., Quant. Biol., 25, 109-118.
McHENRY, H.M. (1992) Body size and proportions in early hominids. Am. J. Phys. Anth., 87, 407-431.
McHENRY, H.M., CORRUCCINI, R.S. (1978) The femur in early human evolution. Am. J. Phys. Anth., 49, 473-488.
PATOU, M. (1989) Subsistance et approvisionement au Paleolitique Moyen. In L'Homme de Néandertal, la subsistance, vol. 6, a c. di Otte, M., Liège, Etudes et Recherches Archéologiques de l'Université de Liège, pp. 11-18.
RIGHTMIRE, G.P. (1990) The evolution of Homo erectus: comparative anatomical studies of an extinct human species, Cambridge, Cambridge University Press.
RIGHTMIRE, G.P. (1993) Variation among early Homo crania from Olduvai Gorge and the Koobi Fora region. Am. J. Phys. Anth. 90, 1-33.
ROBERTS, R.G., JONES, R., SMITH, M.A. (1984) Beyond the radiocarbon barrier in Australian prehistory. Antiquity, 68, 611-616.
RUFF, C.B. (1992) Biomechanical analisys of archaeological human material. In Skeletal biology of past peoples: advances in research methods, a c. di Saunders, S.R., Katzenburg, M.A., New York, Wiley-Liss, pp. 41-61.
RUFF, C.B. (1994) Morphological adaptation to climate in modem and fossil hominids. Yrbk. Phys. Anthrop., 37, 65-107.
RUFF, C.B., TRINKAUS, E., WALKER, A., LARSEN, C.S. (1993) Postcranial robusticity in Homo I: temporal trends and mechanical interpretation. Am. J. Phys. Anth., 91, 21-53.
SHEA, II (1989) A functional study of the lithic industries associated with Hominid fossils in the Kebara and Qafzeh caves, Israel. In The human revolution, a c. di Mellars, P., Stringer, C., Edinburgh, Edinburgh University Press, pp. 611-625. SMITH, F.H., FALSETTI, A.B., DONNELLY, S.M. (1989) Modem human origins. Yrbk. Phys. Anthrop., 32, 35-68.
SPOOR, F., WOOD, B., ZONNEVELD, F. (1994) Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion. Nature, 369, 645-648.
STINER, M.C. (1995) Honor among thieves: a zooarchaeological study of Neandertal ecology. Princeton, Princeton University Press.
STINER, M.C., KUHN, S.L. (1992) Subsistence, technology and adaptative variation in Middle Palaeolithic Italy. Am. Anthrop., 94, 306-339.
STRAIT, D.S, GRINE, F.E., MONIZ, M. (1997) A reappraisal of early hominid phylogeny. J. Rum. Ev., 32, 17-82.
STRAUS, L.G., CLARK, G. (1986) La Riera Cave. Anthropological Research Papers 36, University of Arizona.
STRINGER, C.B. (1985) Middle Pleistocene hominid variability and the origin of Late Pleistocene humans. In Ancestors: the hard evidence, a c. di Delson, E., New York, Wiley-Liss, pp. 289-295.
STRINGER, C.B. (1986) The credibility of Homo habilis. In Major topics in primate and human evolution, a C. di Wood, B.A., Martin, L., Andrews, P., Cambridge, Cambridge University Press, pp. 266-294.
STRINGER, C.B. (1987) A numerical cladistic analysis for the genus Homo. J. Rum. Evol., 16, 135-146.
STRINGER, C.B., ANDREWS, P. (1988) Genetic and fossil evidence for the origin of modern humans. Science, 239, 1263-1268.
SUSMAN, R.L. (1988) Hand of Paranthropus robustus from Member I, Swartkrans: fossil evidence for tool behavior. Science, 240, 781-784.
SUSMAN, R.L., STERN, J.T. (1982) Functional morphology of Homo habilis. Science, 217, 931-934.
SWISHER, C.C. CURTIS, G.H., JACOB, T., GETTY, A.G., SUPRIJO WIDIASMORO, A. (1994) Age of the earliest known hominids in Java, Indonesia. Science, 263, 1118-1121.
TATTERSALL, I. (1986) Species recognition in human palaeontology. J. Hum. Evol., 15, 165-175.
TATTERSALL, I. (1994) Morphology and phylogeny. Ev. Anthro., 3, 40-41.
TOTH, N.I. (1987) Behavioral inferences from Early Stone Age artifact assemblages: an experimental model. Hum. Evol., 16, 763-787.
TRINKAUS, E. (1976) The morphology of European and Southwest Asian Neandertal pubis bones. Am. J. Phys. Anthrop., 44, 95-104.
TRINKAUS, E., CHURCHILL, S.E., RUFF, C.B. (1994) Postcranial robusticity in Homo II: humeral bilateral asymmetry and bone plasticity. Am. J. Phys. Anthrop., 93, 1-34.
TRINKAUS, E., STRINGER, C.B., RUFF, C.B., HENNESSY, R.I., ROBERTS, M.B., PARFITT, S.A. (1996) The Boxgrove Tibia. Am. J. Phys. Anthrop. Suppl., 22, 230-231.
VRBA, E. (1993) The pulse that produced us. Nat. Rist., 102, 47-51.
WALKER, A., LEAKEY, R.E.F. (1993 ) The Nariokotome skeleton. Cambridge, Mass., Harvard University Press.
WOLPOFF, M.H. (1994) Time and phylogeny. Ev. Anthro., 3, 38-39.
WOLPOFF, M.H. (1996) Human evolution. New York, McGrawHill.
WOLPOFF, M.H., CRUMMETT, T.L. (1995) Response to Lieberman. Curr. Anthrop., 36, 186-188.
WOOD, B.A. (1991) Koobi Fora research project, vo!. 4, Hominid cranial remains. New York, Oxford University Press.
WOOD, B.A. (1992) Origin and evolution of the genus Homo. Nature, 355, 783-790.
WU, X., POIRIER, F. (1995) Human evolution in China. New York, Oxford University Press.
Bibliografia generale
KLEIN, R.G. The human career: human biological and cultural origins. Chicago, University of Chicago Press, 1989.
RIGHTMIRE, G.P. The evolution of Homo erectus: comparative anatomical studies of an extinct human species. Cambridge, Cambridge University Press, 1990.
WOLPOFF, M.H. Human evolution. New York, McGraw-Hill, 1996.
WOOD, B.A. Koobi Fora research project, vol. 4, Hominid crani al remains. Oxford, Oxford University Press, 1991