
Evoluzione. Selezione naturale
Evoluzione. Selezione naturale
A centocinquanta anni dalla sua nascita l'idea di selezione naturale rimane la chiave di volta della teoria dell'evoluzione, un ruolo che ricoprì nella formulazione originaria che ne diedero Charles Darwin e Alfred Russel Wallace e conservò nella teoria sintetica dell'evoluzione della metà del Novecento. La selezione naturale, o riproduzione differenziale dei genotipi di una popolazione, ha il doppio ruolo di causa e di effetto. Dal punto di vista delle sue manifestazioni, essa è un effetto; la selezione è il risultato dell'interazione tra variazione genetica fenotipicamente espressa e variazione dei fattori ecologici direttamente agenti sugli individui; tali fattori ne modificano la probabilità di sopravvivenza o la fecondità. Praticamente, ogni volta che si passa da una generazione a quella successiva, nella popolazione ha luogo un processo spontaneo di selezione. A selezione avvenuta, la composizione genotipica, quali-quantitativa, della nuova popolazione sarà differente dalla composizione genotipica della popolazione di partenza. Mentre a livello fenomenologico la selezione naturale rappresenta un effetto, a livello teorico della spiegazione dei processi evolutivi essa invece rappresenta una causa di evoluzione.
I processi evolutivi basati sul meccanismo di selezione sono processi deterministici, perché la probabilità di riproduzione dei differenti genotipi non è dovuta al caso: gli individui che riescono a sopravvivere, e tra questi quelli che lasciano il maggior numero di figli capaci di diventare adulti, sono anche nel complesso i più idonei al loro ambiente di vita. La selezione naturale è un meccanismo che in presenza di vincoli ambientali produce evoluzione a patto che gli individui della popolazione possiedano delle differenze che influenzano la loro idoneità riproduttiva e a patto che tali differenze siano ereditabili. In definitiva, c'è evoluzione attraverso selezione solo a carico di caratteri variabili, ereditabili e collegati alla fitness. Nel concetto di selezione, alla dimensione genetico-riproduttiva si trova intimamente intrecciata la dimensione ecologico-economica, perfettamente riflessa nell'espressione ottocentesca di 'lotta per l'esistenza'. Il meccanismo selettivo funziona ed è onnipervasivo; c'è sempre selezione perché la capacità riproduttiva di qualsiasi specie è teoricamente infinita, mentre le risorse ecologiche sono limitate; di conseguenza gli individui, in quanto geneticamente differenti, possiedono ineguale probabilità di sopravvivenza o ineguale successo riproduttivo. I vincitori non sono però estratti a sorte; il successo va a chi in quel contesto riesce a sopravvivere e a riprodursi con più efficacia.
Il meccanismo di selezione è alimentato dal caso; perché possa esservi selezione è necessario che vi sia variabilità genetica e questa è promossa da mutazioni casuali del materiale genetico. L'ordine biologico riscontrabile nei sistemi viventi è in larga parte un prodotto della selezione naturale, un meccanismo che da miliardi di anni, generazione dopo generazione, è responsabile dell'evoluzione adattativa.
Cenni storici
Uno dei pilastri dello studio moderno dei viventi è costituito dall'assunto secondo il quale tutti gli adattamenti di un organismo al proprio ambiente possono essere spiegati ricorrendo a un processo chiamato 'selezione naturale'. Com'è noto, i primi a identificare nella selezione la causa principale del cambiamento evolutivo, e a indicarla come forza guida dell'adattamento e della diversità biologica, furono i due naturalisti inglesi Charles Darwin e Alfred Russell Wallace. È documentato che ciascuno di loro elaborò all'insaputa dell'altro una propria teoria dell'evoluzione per selezione naturale, basandosi su un'enorme quantità di osservazioni personali combinate con la lettura critica di alcuni testi chiave. Cruciali per entrambi furono lo scritto di demografia umana An essay on the principle of population (1798) dell'economista Thomas R. Malthus (1766-1834) e il trattato Principles of geology (1830) di Sir Charles Lyell (1797-1875).
Nonostante alcune differenze di accento su certi punti, la teoria di Darwin e quella di Wallace sono sostanzialmente identiche e la paternità della scoperta della selezione naturale appartiene perciò a entrambi. Darwin, in particolare, formulò un'articolata spiegazione meccanicistica dell'adattamento, connettendo tra loro in maniera altamente originale fatti e idee noti a molti naturalisti suoi contemporanei. Dal 1838 (anno in cui, leggendo Malthus, venne colpito dall'idea ‒ come scrive nella sua autobiografia ‒ che nella "lotta per la vita [...] le variazioni vantaggiose tendessero a essere conservate e quelle sfavorevoli a essere distrutte") al 1858, quando la teoria venne resa pubblica per la prima volta, Darwin lavorò instancabilmente al perfezionamento del proprio modello esplicativo.
La grande novità della concezione darwiniana della selezione consiste nell'idea che essa non si limita a essere un processo eliminativo (questa conclusione, collegata a una visione essenzialistica e statica dei viventi, la si può ritrovare già nei pensatori dell'antica Grecia), ma può consentire anche la manifestazione di proprietà nuove per la popolazione. L'insistenza sugli aspetti innovativi della selezione si accorda perfettamente con la visione popolazionista e dinamica che pervade l'opera On the origin of species (1859). Benché le conclusioni di Wallace collimino con quelle di Darwin (entrambi ritengono infatti che la grande variabilità delle popolazioni congiunta alla severa decimazione operata regolarmente dall'ambiente debba portare a un cambiamento evolutivo), l'approccio wallaceano si distingue da quello darwiniano per il taglio spiccatamente ecologico dell'analisi.
Nel saggio del 1858, Wallace argomenta con chiarezza riguardo alla necessità logica di una selezione intra-popolazionale, non molto diversamente da come farebbe oggi un moderno ecologo di popolazione. Muovendo da osservazioni sulla sostanziale e generale costanza numerica delle popolazioni naturali, Wallace dimostra come questo stato di fatto sia da ricondurre all'azione stabilizzatrice della selezione: escludendo la maggior parte degli individui dalla riproduzione (uno dei fattori ambientali più critici è per Wallace la competizione per le risorse), la selezione consente solo a pochi di sopravvivere e di riprodursi. Anche per Wallace le differenze tra gli individui di una popolazione forniscono la campionatura in cui verranno scelte quelle varianti che solo a posteriori potremo definire meglio adattate a quella certa situazione ambientale. Per Wallace, quindi, come per Darwin, variazione e selezione sono elementi indispensabili alla produzione del cambiamento.
Definizione e descrizione
Data una popolazione di organismi, se essa è caratterizzata da variazione di un carattere, se il carattere è ereditabile ed esistono differenze di fitness a esso collegate, allora è possibile prevedere che la distribuzione del carattere nella progenie sarà differente da quella della generazione parentale. Se sono soddisfatte le tre condizioni (variazione, ereditabilità, diversa idoneità), è estremamente probabile che si avrà una divergenza tra le due distribuzioni di frequenza, quella della generazione parentale e quella della progenie. Tale divergenza è indice di un cambiamento evolutivo che a sua volta dipende da differenze nella capacità riproduttiva (o nella capacità di sopravvivenza) degli individui della generazione parentale. Per questa ragione la selezione naturale è definibile come 'riproduzione differenziale di genotipi'.
Dal punto di vista della moderna teoria dell'evoluzione, nella forma in cui è trattata in genetica di popolazione, la selezione naturale rappresenta, insieme alla deriva genetica, alla mutazione, alla migrazione e alla suddivisione della popolazione in isolati genetici, una delle cause principali dell'evoluzione delle popolazioni di organismi. Tra tutti questi fattori, la deriva genetica (cioè la fluttuazione casuale degli alleli tra generazioni) si manifesta in modo simile alla selezione. In via di principio la loro distinzione è assai semplice, dato che nella deriva genetica non esistono per definizione differenze di fitness tra le varianti del carattere, e che, in secondo luogo, l'effetto statistico della fluttuazione casuale delle frequenze alleliche è direttamente collegato alla dimensione della popolazione: più piccola è la popolazione, maggiore è la probabilità di fluttuazione casuale. Tuttavia, anche in una piccola popolazione assieme alla deriva può agire la selezione naturale; solo che in questo caso diventa difficile per ragioni statistiche discriminare gli effetti separati delle due componenti del cambiamento. Se può esservi evoluzione (intesa come origine e diffusione di alleli, varianti, valori e stati di un carattere), o per deriva genetica o per selezione naturale o per effetto combinato di entrambe, ciò non vuol dire che la selezione naturale produca sempre evoluzione. Infatti, se la popolazione si mantiene in regime di equilibrio, la distribuzione di frequenza di un qualsiasi carattere resta costante da una generazione all'altra. Per definizione, in una popolazione in equilibrio non c'è mai evoluzione. Nonostante la definizione univoca data in precedenza, l'espressione 'selezione naturale' è impiegata con molteplici significati a seconda dello specifico processo a cui viene riferita.
Distinguendo tra processi che sono alternativi nella forma e nel risultato, avremo una selezione sessuale e una selezione non sessuale (che corrisponde alla selezione naturale in senso ristretto). La distinzione tra selezione sessuale ‒ il processo che determina il successo differenziale negli accoppiamenti ‒ e selezione naturale viene fatta perché qualche volta i caratteri favoriti dalla selezione sessuale possono essere svantaggiati oppure contrastati dai componenti della selezione naturale. Tuttavia, oggi molti autori non ritengono valida questa distinzione, per cui la selezione sessuale viene vista soltanto come una componente della selezione naturale. Nella selezione naturale in senso stretto possiamo distinguere due sottoprocessi demografici: uno connesso alla mortalità, l'altro collegato alla fecondità. Secondo un'altra distinzione vi sarebbero nella selezione naturale due processi sequenziali, non alternativi, detti 'selezione fenotipica' e 'risposta evolutiva'. In questo approccio, molto frequente nella genetica quantitativa (che studia caratteri controllati da più geni e a variazione continua come la statura), la selezione fenotipica corrisponderebbe alla modifica della distribuzione di un carattere tra gli individui di coorti diverse ma appartenenti alla stessa generazione. La cosiddetta risposta evolutiva consisterebbe nel cambiamento prodotto dall'interazione tra selezione fenotipica e sistema genetico. Siccome nella selezione fenotipica non viene soddisfatta la condizione di ereditabilità del carattere, a rigore essa non può essere considerata propriamente un tipo di selezione naturale. D'altra parte, è l'ambiente a operare la selezione e l'ambiente (fattori fisico-chimici e fattori biologici) agisce sui fenotipi, non direttamente sui corredi genetici individuali (genotipi). Parlare di selezione fenotipica significa sottolineare che la selezione naturale è sopravvivenza e perpetuazione dei fenotipi, trascurando arbitrariamente che i caratteri debbano essere ereditari. Se la selezione tra fenotipi determina infatti le distribuzioni di frequenza al momento della riproduzione, è però l'ereditarietà dei caratteri a consentire il trasferimento di quella distribuzione di frequenza alla successiva generazione. Anche la selezione fenotipica rappresenta solo una parte del processo di selezione naturale.
Modi e tipi di selezione
Nonostante l'esito della selezione naturale sia sempre uno solo (la riproduzione differenziale dei genotipi), molteplici sono i suoi modi d'azione. I differenti aspetti del processo sono collegabili alla distribuzione di frequenza di un carattere, al numero di fenotipi selettivamente equivalenti, alla micro-variazione spaziale dell'habitat, ai livelli di selezione. In una popolazione i caratteri possono avere variazione continua (caratteri quantitativi come, per es., la statura) o discontinua (cosiddetti 'caratteri polimorfici', per es., il gruppo sanguigno). I modelli di selezione agenti sulle distribuzioni di frequenza dei caratteri sono di tre tipi. Nella selezione 'direzionale' sono favoriti gli individui posti a uno dei lati della curva normale; dopo la selezione ci sarà una nuova media e la varianza potrà essere minore. Nella selezione 'stabilizzante' sono favoriti gli individui posti nella zona media della curva: il valore ottimale del carattere prima della selezione coincide con quello medio della distribuzione dopo la selezione; la media non cambia e la varianza diminuisce.
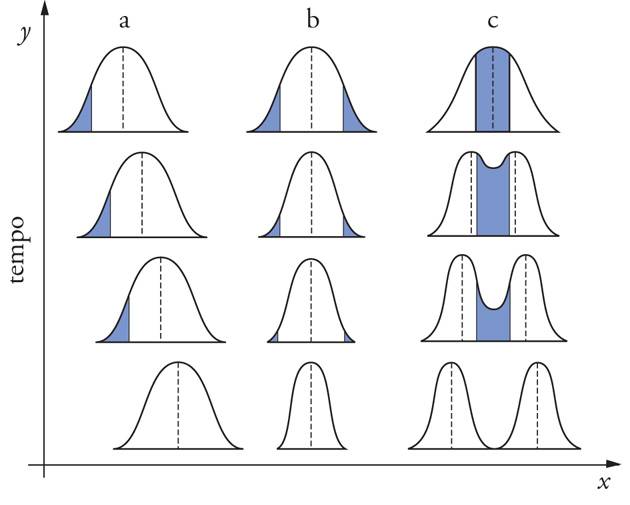
Nella selezione 'disruptiva' (anche detta 'diversificante') vengono favoriti gli individui posti all'estremo della curva e sono sfavoriti quelli che presentano i valori intermedi del carattere; la media può cambiare e la varianza aumenta (fig. 2). Una selezione di questo tipo è quella detta 'dipendente da frequenza' (ovvero 'densità-dipendente'), in cui i fenotipi più comuni sono svantaggiati rispetto a quelli più rari (in una popolazione di Insetti predati da Uccelli, per es., tutti gli individui di aspetto più vicino al tipo medio sono più predati dagli Uccelli insettivori, mentre gli individui di aspetto più lontano dal tipo medio sono più rari e perciò meno esposti alla predazione). Sotto l'azione della selezione dipendente da frequenza, con il passare delle generazioni i tipi inizialmente più rari tenderanno a divenire più frequenti. Non necessariamente, però, a differenze fenotipiche osservabili corrispondono differenze selettive; gli stessi fenotipi, infatti, possono essere selettivamente neutrali, selettivamente avvantaggiati o svantaggiati a seconda dell'ambiente considerato e del gruppo di fenotipi della stessa popolazione con cui vengono confrontati. D'altra parte, fenotipi differenti possono anche equivalersi sotto il profilo adattativo (in una popolazione di piante, per es., una protezione egualmente efficace contro i fitofagi può essere ottenuta in modo diverso dai differenti individui, grazie alla sintesi di differenti tipi di metaboliti secondari).
Le modalità di azione della selezione naturale possono differire da luogo a luogo, a seconda dei distretti componenti l'area di distribuzione geografica della specie. Questa variabilità geografica della selezione, combinata con la migrazione genica, può provocare una variazione delle frequenze geniche e dei valori di un carattere che sarebbero del tutto inavvertibili limitandosi all'esame locale di una sola popolazione. Anche se in via di principio è possibile che la selezione naturale sortisca effetti differenti a seconda che agisca su geni, genotipi, gruppi, popolazioni o specie, mancano prove irrefutabili dell'esistenza di modalità selettive veramente alternative alla selezione che agisce sui fenotipi individuali. In questi ultimi anni molti biologi evoluzionisti stanno nondimeno lavorando a una teoria della selezione agente a più livelli gerarchici. L'ostacolo maggiore, lo stesso incontrato da Darwin che già si era posto il problema, è quello di dimostrare in che modo, attraverso l'aumento di fitness di un'entità sopraindividuale, si riesca a realizzare un adattamento che allo stesso tempo possa essere neutrale o addirittura svantaggioso a livello degli individui. I modelli teorici indicano che ci sono condizioni in cui questo è possibile, ma al momento mancano evidenze sperimentali a sostegno di queste conclusioni. D'altra parte, la natura gerarchica del vivente è un dato di fatto, ma questo non comporta di per sé che a ognuno di questi livelli (individuo uni- o pluricellulare, popolazione, specie) corrisponda automaticamente un'unità di selezione.
Fitness
Il termine fitness (appropriatezza, idoneità) rimanda a uno dei concetti più importanti di tutta la biologia evolutiva: quello di 'idoneità riproduttiva'. Nonostante l'apparente semplicità, la nozione di fitness presenta parecchie difficoltà sia teoriche, sia pratiche. La quantità di figli prodotti non è una misura accettabile dell'idoneità riproduttiva di un membro di una popolazione. È necessario conoscere anche l'idoneità riproduttiva dei suoi figli insieme al numero e alla fitness dei figli degli altri individui della stessa popolazione. Per fortuna questi ostacoli sono aggirabili, perché di solito non si è interessati al successo biologico di un determinato individuo, ma, piuttosto, al successo di parecchi individui portatori di un certo allele genico, un fatto che implica l'esistenza di differenze genetiche tra questi individui e i portatori di stati allelici alternativi. È chiaro, perciò, che la fitness piuttosto che essere proprietà di un individuo va riferita a una categoria di individui: quelli che condividono le stesse caratteristiche genetiche, lo stesso genotipo (per es., hanno lo stesso genotipo gli omozigoti per l'allele A a un dato locus genico). La fitness descrive dunque il contributo medio che un genotipo apporta alla generazione successiva, confrontato con quello di altri genotipi. La fitness così intesa (conosciuta anche come 'fitness darwiniana', 'fitness relativa', 'valore selettivo di un genotipo') è relativa a un ambiente determinato e, poiché l'ambiente agisce sull'individuo nel suo complesso e non sui singoli geni, è opportuno che sia riferita ai genotipi piuttosto che ai singoli geni.
Il concetto di fitness è collegato inoltre con quello di adattatività e di adattamento. L'adattatività di un individuo è il grado in cui tale individuo è capace di vivere e di riprodursi all'interno di un certo insieme di ambienti. Questa capacità è misurabile mediante la fitness assoluta. Un fenotipo con un elevato grado di adattatività può manifestare una fitness relativa non alta (e viceversa). Le differenze di adattatività osservabili tra i diversi individui generano il processo di selezione naturale. Diversamente dall'adattatività, che è una caratteristica osservabile alla scala temporale di una generazione, l'adattamento richiede più generazioni ed è la serie di eventi storici con cui si è realizzata la congruità tra organismi e ambiente. Durante il processo di adattamento cambia la fitness relativa media della popolazione. Una conoscenza completa della selezione osservabile in una popolazione si ottiene con una descrizione quantitativa del processo selettivo. L'approccio matematico ai problemi di selezione può essere visto come l'effettuazione di operazioni di misura, o meglio di stima, del tasso di selezione a carico di un carattere polimorfico o quantitativo. Le variabili da stimare in questo caso sono note come 'coefficienti di selezione' e 'differenziali di selezione'. Una volta conosciute, queste grandezze possono aiutare a comprendere i motivi di ordine biologico per cui alcune varianti di un carattere possiedono una fitness maggiore di altre. Inoltre, riconosciuta l'esistenza di un campo di variazione della fitness e stimate le variabili, si potranno tentare previsioni sulla dinamica evolutiva del carattere sotto selezione, nonché sulle eventuali configurazioni di equilibrio.
Bibliografia
Endler 1986: Endler, John A., Natural selection in the wild, Princeton (N.J.), Princeton University Press, 1986.
Futuyma 1986: Futuyma, Douglas J., Evolutionary biology, 2. ed., Sunderland (Mass.), Sinauer Associates, 1986.
Sober 1984: Sober, Ellis, The nature of selection, Cambridge (Mass.)-London, MIT Press, 1984.
Williams 1992: Williams, George C., Natural selection: domains, levels, and challenges, New York-Oxford, Oxford University Press, 1992.
Tavola I
Selezione sessuale
Il successo riproduttivo di un individuo non dipende soltanto dalla sua adattabilità ai fattori fisico-chimici e a quelli biologici, interspecifici, rappresentati da competitori, predatori e parassiti, ma è influenzato in grande misura anche dai rapporti, intraspecifici, che ogni adulto intrattiene con i potenziali partner dell’altro sesso. C’è selezione sessuale ogni volta che uno dei due sessi influisce sulla fitness dei caratteri espressi nell’altro individuo attraverso la scelta del partner in vista dell’accoppiamento.
In molti animali gli individui dei due sessi sono facilmente distinguibili sulla base di caratteri somatici extragonadici. Tra questi caratteri sessuali secondari, i più vistosi comprendono l’apparato genitale e i caratteri morfologici extragenitali come, per esempio, la criniera del leone, lo sperone del gallo, il piumaggio vistoso dei maschi di molti Uccelli, la presenza di corna nei maschi dei Cervidi e in certi Coleotteri, le dimensioni corporee. Un forte dimorfismo sessuale può anche interessare i caratteri fisiologici e soprattutto quelli etologici, come tipicamente accade nel caso delle molto elaborate e bizzarre pergole di accoppiamento costruite dal maschio degli uccelli giardiniere. Da molto tempo i ricercatori si sono interrogati sull’esistenza nei maschi di eventuali vantaggi collegati al possesso di strutture che tuttavia sembrano ostacolare la loro stessa sopravvivenza. È dimostrato infatti che l’esibizione di livree vistose da parte di molti uccelli
può esporli a una maggiore predazione, sicché qui come in casi analoghi l’esistenza di un forte dimorfismo sessuale pone un problema esplicativo piuttosto complesso.
Perché i maschi sono diversi dalle femmine? C’è un vantaggio nel dimorfismo sessuale, e qual è? Darwin affronta la questione nel quarto capitolo di On the origin of species by means of natural selection (1859) e in tutta la seconda parte di The descent of man, and selection in relation to sex (1871), proponendo due spiegazioni. La prima è che i caratteri
sessuali secondari sarebbero utili ai maschi nella lotta per accedere alle femmine; la seconda è che, siccome le femmine di certi uccelli durante il corteggiamento sembrano essere attratte da alcuni caratteri ornamentali come le code vistose e il piumaggio elaborato dei maschi (esempi canonici ne sono rispettivamente la coda del pavone o degli uccelli vedova e le livree degli uccelli del paradiso), questi caratteri sessuali secondari potrebbero essersi affermati perché attraenti per le femmine. Per questa via Darwin arriva a configurare due modalità di selezione sessuale; quella cosiddetta ‘intrasessuale’, attuata attraverso competizione tra maschi per la femmina con cui accoppiarsi, e quella ‘intersessuale’, attuata dalla femmina quando sceglie tra maschi. Il terzo processo attraverso cui sappiamo che la selezione sessuale
agisce è la ‘competizione spermatica’, un fenomeno che Darwin conosceva ma cui non diede grande peso. Darwin in effetti ricorre a due principî e fornisce una spiegazione che sembra duplice ma che in realtà chiarisce solo una metà (la selezione intrasessuale) del fenomeno, lasciando invece inesplicata la scelta operata dalla femmina (la selezione intersessuale).
Alfred R. Wallace, coautore della teoria della selezione naturale, accettò l’idea di competizione tra maschi per l’accesso alle femmine, ma rifiutò nettamente, giudicandola assurda, l’idea di una femmina capace di scegliere il maschio con cui accoppiarsi e quindi respinse la spiegazione darwiniana dell’evoluzione degli ornamenti dei maschi. L’interesse per i vari aspetti della sessualità in senso lato (la sua origine, determinazione ed evoluzione, il dimorfismo sessuale, i sistemi nuziali, la selezione sessuale, il rapporto numerico tra maschi e femmine di una popolazione, o sex ratio, le cure parentali), è sempre rimasto vivo e la ricerca è ripartita ogni volta dalla posizione di Darwin. La doppia ipotesi darwiniana fu affrontata per la prima volta sul piano teorico da Ronald Fisher, che si propose di superare l’empasse in cui veniva a
trovarsi la teoria della selezione naturale, impossibilitata a spiegare le ragioni del dimorfismo sessuale e segnatamente i tratti appariscenti di certi maschi. D’altra parte, se era fuori discussione l’esistenza di una scelta da parte delle femmine, non era invece altrettanto palese quale potesse essere per una femmina il beneficio derivante da questa sua capacità di scegliere un maschio vistoso con cui accoppiarsi.
Nel suo The genetical theory of natural selection del 1930, Fisher elaborò un modello unitario (conosciuto come runaway hypothesis) dell’evoluzione dei caratteri maschili appariscenti e della preferenza femminile in cui i tratti maschili e quelli femminili sono fatti coevolvere fino a raggiungere un equilibrio. Fisher interpretò correttamente la preferenza delle femmine per gli ornamenti vistosi come un adattamento raggiunto attraverso l’azione della selezione naturale. La sua ipotesi era che di per sé non vi fossero differenze biologiche rilevanti tra maschi appariscenti e maschi non appariscenti; giudicò costoso esibirsi e quindi giudicò svantaggiati in termini di selezione naturale i maschi portatori di ornamenti costosi e che perciò sprecano risorse, ma convenne che le femmine rispondono molto di più al corteggiamento di questi maschi esibitori di segnali costosi piuttosto che al corteggiamento dei maschi poco appariscenti. In sostanza, secondo Fisher, le femmine tendono a considerare più attraenti i maschi riccamente ornati, i quali perciò da questa tendenza ricaveranno un premio riproduttivo. Secondo questo modello, visto che attraggono un maggiore numero di femmine, solo i maschi sono avvantaggiati; le femmine sono svantaggiate quando generano figli appariscenti. D’altronde, però, solo questo tipo di prole attirerà le femmine e quindi la vistosità dei maschi si perpetuerà nella popolazione attraverso le generazioni.
Una diversa spiegazione della presenza di ornamenti vistosi e stravaganti nei maschi di alcuni uccelli venne data negli anni Settanta da Atmotz Zahavi, il quale ipotizzò che caratteristiche come la coda del pavone rappresentino veri e propri handicap per il portatore. Questi handicap, d’altra parte, proprio in quanto tali agirebbero come avvertimenti ‘onesti’ della vitalità del maschio. Osservando la capacità di un maschio di tollerare il proprio handicap, la femmina potrebbe perciò scegliere i maschi migliori con cui accoppiarsi. Ecco allora spiegato come i geni codificanti per questi fenotipi si diffonderebbero nella popolazione. Secondo Zahavi le femmine valuterebbero la qualità dei maschi sfruttando le informazioni rappresentate dall’esibizione di segnali molto costosi quali sono le strutture ornamentali corporee. La principale obiezione al modello di Zahavi riguarda la dinamica temporale della variazione fenotipica (e quindi della variazione genetica sottostante).
L’ipotesi di Zahavi prevede che col tempo le differenze tra maschi debbano ridursi fino a scomparire per controselezione delle varianti con minore fitness; ma questa previsione è contraddetta dai fatti. Una soluzione a questo problema venne avanzata nel 1982 da William Hamilton e Marlene Zuk, che proposero di interpretare caratteri appariscenti ed esibizioni maschili come handicap rilevatori della resistenza, controllata geneticamente, all’infezione da parte di parassiti. Dimostrare la correttezza dell’ipotesi di Hamilton e Zuk è difficile, mentre è facile obiettare che una femmina potrebbe evitare di accoppiarsi con un maschio malato (e quindi senza ornamenti) semplicemente per evitare di contrarre essa stessa i parassiti. Mentre nel modello di Zahavi tutti i maschi investono in eguale misura nell’handicap, ma solo i più vitali possono sostenere il costo di un tale investimento (l’handicap è dipendente dallo stato di benessere dell’animale), nel modello di Hamilton e Zuk solo i maschi migliori sono portatori di handicap (l’handicap è una spia del vigore dell’animale).
A partire dagli anni Settanta, le osservazioni e gli studi sperimentali sull’evoluzione dei caratteri sessuali secondari hanno ricevuto nuovo impulso dalla messa a punto di tecniche e di metodi innovativi che hanno permesso di stimare parametri (per es., il numero di copule, il loro costo metabolico, la quantità di figli generati, ecc.) influenti sulla fitness di un individuo, hanno consentito di accertare la paternità (mediante il test del DNA fingerprint) e di valutare fattori e modelli comportamentali coinvolti nell’evoluzione della sessualità, in particolare quelli relativi alle colorazioni maschili, alla scelta del partner, alle cure parentali. Tra le domande più frequenti c’è quella se gli animali scelgano o meno il partner in base al fatto che esso possiede caratteri appariscenti. In alcuni casi è quello che avviene. Nel 1982, ipotizzando che nella vedova codalunga, la specie africana poliginica Euplectes progne, la selezione sessuale favorisse l’evoluzione di code lunghe, l’etologo Malte Andersson manipolò la lunghezza della coda accorciandola ad alcuni esemplari e tagliandola e poi rincollandola (perciò senza alterarne la lunghezza) ad altri. La vedova codalunga è un uccello dalle dimensioni di un passero caratterizzato da un marcato dimorfismo sessuale: le femmine hanno una coda di circa 7 cm, una lunghezza giudicata pressoché ottimale per il volo, mentre la coda dei maschi può raggiungere i 50 cm. In natura, i maschi difendono un’area in cui si esibiscono in complessi rituali di corteggiamento. Lo sperimentatore dimostrò che la lunghezza della coda non aveva alcun peso nella competizione tra maschi durante la fase di difesa delle arene di corteggiamento, mentre ne aveva nella loro capacità di suscitare l’interesse delle femmine. I maschi con coda accorciata artificialmente attraevano infatti meno femmine di quelli con coda integra o forniti di code tagliate e rincollate. I maschi con le code allungate avevano arene con un maggiore numero di nidi rispetto a quelle degli altri maschi.
Diversamente dal punto di vista tradizionale che giudicava corteggiamento e accoppiamento come processi eminentemente cooperativi, oggi ha sempre più spazio l’ipotesi dell’esistenza di un vero e proprio conflitto di interesse tra maschio e femmina.
Questo in realtà non deve stupire perché, anche se è vero che entrambi i partner sono beneficiati dalla riproduzione, esiste tuttavia una fondamentale asimmetria di ruoli generata dal differente investimento parentale (allocazione di risorse necessarie a nutrire e allevare la prole) che controlla la fitness maschile e quella femminile. La principale differenza tra maschio e femmina risiede nella differente dimensione dei rispettivi gameti (stato di anisogamia): le femmine producono gameti grandi immobili e ricchi di sostanze nutritive, mentre i maschi producono gameti piccoli e solitamente molto mobili (lo spermatozoo comprende una testa contenente DNA e una coda dotata di un flagello propulsore). La situazione è perciò chiaramente distinta: i maschi aumentano la propria fecondità in maniera direttamente proporzionale al numero di accoppiamenti effettuati (vi sarà dunque una selezione sui maschi a favore delle caratteristiche che aumentano
la probabilità di essere scelti per l’accoppiamento), mentre le femmine investono nella prole una maggiore quantità di risorse sia nella produzione dei gameti che nelle cure parentali (e perciò la selezione massimizzerà la loro capacità di scegliere i maschi ‘migliori’ insieme a qualunque attributo in grado di favorire l’allevamento della prole). Per questo motivo i maschi sono per loro natura etologicamente ‘focosi’, pronti al combattimento specialmente nel periodo in cui le femmine sono sessualmente ricettive, mentre la diversa condizione delle femmine le rende particolarmente accorte nella scelta del maschio. In molte specie di Uccelli e Insetti la ritrosia delle femmine viene vinta attraverso un dono nuziale sotto forma di cibo, che viene consumato durante il corteggiamento o durante la copula. Anche lo sforzo di accoppiamento – che, con l’investimento parentale, rappresenta la seconda componente dello sforzo riproduttivo – differisce tra maschio e femmina. L’investimento necessario per procurarsi il partner risulta maggiore nel maschio che nella femmina. Complessivamente la selezione sessuale è più intensa nelle specie con sistema nuziale poligamico, promiscuo (cioè un misto di poliginia – il maschio si accoppia con molte femmine e ciascuna femmina si accoppia con un solo maschio – e poliandria – la femmina si accoppia con molti maschi), che nelle specie monogame, in cui si stabilisce un legame di coppia e spesso entrambi i genitori concorrono alla cura della prole.