
Febbre
Febbre
di Massimo Aloisi
Febbre
sommario: 1. Introduzione. 2. La termoregolazione fisiologica: a) caratteristiche generali; b) acquisizione filogenetica; c) cibernetica della termoregolazione; d) caratteristiche risultanti della omeotermia. 3. Le ipertermie fisiologiche, patologiche e sperimentali. 4. La termoregolazione nella febbre. 5. Patogenesi della febbre: a) introduzione; b) i pirogeni esogeni; c) i pirogeni endogeni. 6. Significato della febbre. 7. Andamento e forme cliniche della febbre. 8. L'organismo del febbricitante. 9. Sostanze che possono interferire con la parte centrale della termoregolazione: gli antipiretici. □ Bibliografia.
1. Introduzione
La febbre è uno stato patologico transitorio (cioè un processo morboso) cui vanno incontro gli animali e l'uomo, durante il quale la temperatura di riferimento della termoregolazione si sposta a un livello maggiore della norma. Ne consegue che la febbre come tale è un fenomeno strettamente inerente agli animali termoregolati (omeotermi o endotermi), il che non esclude che variazioni della temperatura corporea locali o generali possano intervenire in particolari condizioni patologiche anche negli eterotermi (o poichilotermi o ectotermi).
La ragione per cui un uomo può febbricitare, consistendo tale fenomeno nella comparsa, spesso nel corso di altri eventi patologici, di stimoli capaci di modificare il meccanismo fisiologico della termoregolazione, non può essere compresa se non sulla base delle nozioni concernenti questo aspetto della fisiologia. E siccome la termoregolazione e un acquisizione estremamente importante dello sviluppo evolutivo, anche la capacità a febbricitare va probabilmente vista come un ulteriore perfezionamento (sia pure con i suoi lati negativi) del processo evolutivo onde si è stabilizzata l'omeotermia, come una termoregolazione di eccezione per una situazione di eccezione.
Non esiste una dottrina compiuta della febbre, fenomeno clinico naturalmente noto sin dai tempi più lontani, studiato e classificato per le sue forme appariscenti e per gli altri segni che lo accompagnano, sicché la sua descrizione temporale è sempre stata di notevole ausilio diagnostico e prognostico nella medicina pratica, assai prima che si sapesse alcunché di termoregolazione. Se non esiste ancora una dottrina compiuta della febbre lo si deve in primo luogo al fatto che essa è un fenomeno assai complesso, mentre sul piano molecolare è largamente carente l'analisi della stessa fisiologia della termoregolazione; in secondo luogo tale incompiutezza dipende dal fatto che lo spostamento della omeotermia quale si verifica proprio nella febbre si rivela un meccanismo assai intricato e strettamente dipendente da altri circuiti fisiopatologici.
2. La termoregolazione fisiologica
a) Caratteristiche generali
Un omeotermo riesce a mantenere la temperatura interna del proprio corpo intorno a un valore caratteristico della specie annullando quanto più è possibile le differenze rispetto a quel valore, quali possono intercorrere o per effetto di maggior produzione calorica o per la varia incidenza della temperatura ambientale; ma ciò che è ancor più notevole (e difficile a ottenersi) è che un tale organismo riesce a mantenere una temperatura superiore (spesso anche molto superiore) a quella del mondo esterno (v. Thauer, 1967).
I mezzi con i quali si realizza questa regolazione (la cui importanza è talmente grande da non poterne apprezzare ancora tutte le implicazioni) sono fondamentalmente i seguenti: la produzione calorica dell'organismo, sempre più efficientemente isolata nell'evoluzione delle specie da qualunque scambio con l'esterno, si compie attraverso i processi metabolici esotermici cellulari, e in minima misura anche attraverso gli attriti nell'apparato locomotore; la dispersione calorica si compie in vario modo a seconda degli animali (forniti o meno di pelliccia o di piumaggio, di ghiandole sudoripare, ecc.), mediante vasodilatazione e vasocostrizione in regioni del sistema vascolare in contatto con l'esterno (cute, mucose delle vie respiratorie e della bocca), mediante secrezione di sudore destinato all'evaporazione (e quindi alla sottrazione di calore), o attraverso evaporazione sulle mucose per ventilazione superficiale (polipnea termica: cane e altri animali) delle prime vie respiratorie (v. Du Bois, 1937). Questi sono i mezzi dinamici della temoregolazione; gli animali e l'uomo hanno inoltre dei mezzi statici che sono adattamenti che realizzano un determinato isolamento dall'ambiente esterno, in modo che gli aspetti dinamici, e cioè più veri, della termoregolazione non si attuino in situazioni estreme; tali mezzi statici sono le pellicce, i piumaggi, i vestiti; l'uomo vi aggiunge anche, laddove è possibile, una tecnologica stabilizzazione termica di una parte del suo ambiente esterno più immediato (condizionamento dei luoghi di vita e di lavoro).
L'equazione generale del bilancio calorico di un organismo può essere così formalizzata: M = M0 + Ma = R + C + D + E + ΔC, ove M = tasso metabolico; M0 = metabolismo basale; Ma = metabolismo dovuto all'attività; R,C,D,E, = perdite di calore, rispettivamente per irradiazione, per convezione, per conduzione e per evaporazione; ΔC = modificazione nel contenuto termico corporeo (v. Hammel, 1968).
b) Acquisizione filogenetica
Il raggiungimento della omeotermia da parte dei Vertebrati superiori è un evento straordinariamente importante nell'evoluzione delle specie; ed esso è strettamente legato al passaggio dall'ambiente marino e acquatico a quello terrestre, allo sviluppo di un sistema di isolamento, alla riduzione generale della mole corporea, a un notevole perfezionarsi delle funzioni nervose e allo stesso sviluppo del cervello. Occorre tuttavia tener conto che il processo è autoremunerante, nel senso che l'omeotermia è stata condizione dello sviluppo e del consolidarsi di meccanismi complessi di autocontrollo che perfezionano, tra l'altro, la stessa omeotermia.
Ma un intervento regolatore della temperatura corporea, o quanto meno una reattività organismica contro un passivo subire le variazioni più estreme della temperatura ambientale, non si verifica d'improvviso negli omeotermi. La reattività animale è molto più vasta e mentre include embrionali o meno elaborati accorgimenti per la salvaguardia delle condizioni fisiologiche della conservazione individuale, per ciò stesso affonda le sue radici nella reattività generale organismica nei confronti di un cimento ostile (stress), non necessariamente di natura termica. Infatti alcune delle reazioni che troviamo nelle forme superiori come adatte alla lotta contro gli insulti termici, in animali poichilotermi (o ectotermi) terrestri si trovano come reazioni generali di emergenza di fronte al pericolo, quali il tremore (brivido), l'accelerazione cardiaca e la vasomotilità, la secrezione mucosa o cutanea (sudorazione) e lo svuotamento cloacale.
Del resto residui di queste forme di reattività si conservano anche negli omeotermi più perfezionati e del tutto al di fuori del processo termoregolativo.
Nelle specie ectotermiche, e anche questo si conserva in qualche misura negli omeotermi, spesso la dialettica con l'ambiente non è solo nel senso della difesa, ma anche nell'altro senso, cioè quello dello sfruttamento, nei limiti del possibile, delle caratteristiche termiche dell'ambiente per favorire, per esempio in un ambiente mediamente caldo, processi metabolici e quindi fisiologici altrimenti impossibili (per es., nei Rettili, i quali sono anche suscettibili di modulazioni vasomotorie cutanee capaci di favorire in un senso o in un altro lo scambio termico con l'ambiente). Di qui la primordiale difesa della temperatura corporea dalle eccessive variazioni ambientali passivamente imposte (ectotermi) o da un eccessivo e difficile lavoro termoregolativo (endotermi), che consiste nella reazione comportamentistica: l'animale, come l'uomo, cerca l'ambiente termico più adatto, meno cimentante, cambia atteggiamento corporeo, cambia l'entità e la qualità della nutrizione, ecc.
Ma evidentemente tutto questo non rende l'organismo svincolato dalla legge di Arrhenius e di Van't Hoff, cioè non libera le operazioni biochimiche e quindi fisiologiche dalla loro dipendenza dalle caratteristiche termiche dell'ambiente, con tutte le sue variazioni stagionali, imponenti, specie nel settore terrestre.
Come si è accennato, il passaggio alla vita terrestre ha permesso il realizzarsi della omeotermia, non solo nel senso della stabilità della temperatura, che è un evidente fattore di dominio sulle funzioni organismiche, ma anche nel senso della stabilità a un livello termico anche molto superiore alla media dell'ambiente esterno, il che permette una particolare intensità di reazioni, uno sviluppo e un'efficienza del sistema nervoso nemmeno paragonabili alla situazione degli ectotermi. A ciò ha naturalmente contribuito la maggiore disponibilità di ossigeno nell'ambiente terrestre. Si delinea così una macchina cibernetica a combustione il cui prodotto energetico non è immediatamente calore (bensì energia chimica, elettrica, meccanica, ecc.), ma nella misura e lungo il tempo in cui inevitabilmente produce anche calore, ne riutilizza una gran parte per mantenere il milieu intérieur idoneo per quella stessa intensa attività. Che tale aspetto termico del mezzo interno si mantenga realmente costante per tutta la vita o si realizzi a due livelli diversi, uno più elevato e l'altro più basso a seconda delle stagioni, come avviene nell'ibernazione di alcuni endotermi, è qualità secondaria ai fini del presente contesto: semmai l'ibernazione degli omeotermi è un ulteriore adattamento dell'organismo a condizioni di ambiente difficili da fronteggiare e nel caso in cui il combustibile della macchina (nutrimento) subisca inevitabili flessioni periodiche (invernali).
Del resto, come vedremo, variazioni ritmiche, circadiane, estrali e mestruali e d'altra natura sussistono anche nella più perfezionata omeotermia, così come si verificano e sussistono differenze topografiche della temperatura corporea in uno stesso organismo, in parte dovute a circostanze metaboliche particolari, in parte dovute al meccanismo stesso della termoregolazione in quanto l'organismo non è mai veramente ‛isolato' dal mondo esterno e qualcosa di esso deve pur subire le inerenti variazioni termiche, senza di che non esisterebbe neppure una termoregolazione. (È noto, per esempio, che l'optimum termico fisiologico per i testicoli dei Mammiferi nella collocazione scrotale è inferiore alla temperatura media del corpo. Una temperatura più elevata danneggia gravemente, fino alla completa atrofia, questi organi).
L'omeotermia è stata dunque un'acquisizione lunga e complessa e oggi appare come un continuo sforzo non per sfuggire alla legge di Arrhenius e di Van't Hoff, bensì per utilizzarla in un regime tendenzialmente costante. Ciò è costato e costa un alto prezzo, specie per il livello di temperatura interna a cui si opera la regolazione: si pensi che in media esistono circa 20 °C di differenza fra temperatura corporea interna e temperatura esterna (negli Uccelli anche 23-24 °C) e per questo occorre produrre (e disperdere) in media 7-10 volte più calore di quanto non avverrebbe se il livello della termoregolazione fosse più equilibrato con la temperatura media ambientale; l'omeotermo ha quindi, a parità di peso corporeo, un bisogno di nutrimento che è 7-10 volte maggiore di quello del poichilotermo. In condizioni di perfrigerazione tale differenza teorica cresce naturalmente almeno di un fattore due e ciò spiega per alcuni animali la necessità dell'ibernazione. Né questo sforzo è scevro di pericoli; e infatti nello sforzo di mantenere l'omeotermia anche in condizioni di insulto termico estremo, l'uomo può entrare in un vero stato patologico: un aumento da 30 a 40 °C di temperatura esterna con alta umidità atmosferica (che riduce o annulla l'effetto utile della sudorazione) conduce il cuore ad aumentare di 4-5 volte il volume-minuto, mentre le resistenze periferiche diminuiscono dello stesso ordine di grandezza, così che in queste condizioni e nella stazione eretta è facile un collasso generale (v. Thauer, 1965 e 1967).
c) Cibernetica della termoregolazione
Anche seguendo le informazioni più recenti non è possibile oggi fornire uno schema completo e del tutto realistico del complesso di operatori (organi, tessuti, terminali nervosi e centri nervosi) e delle loro vicendevoli connessioni che costituiscono il sistema di controllo termoregolativo. Certamente esso è un sistema a multipli circuiti a retroazione (fedback loops), i quali sono interposti tra i due poli principali della funzione termoregolativa, la termoproduzione da una parte e la termodispersione dall'altra. Così concepito, questo schema (comunque un fisiologo moderno lo voglia costruire o completare con elementi ancora ipotetici) rappresenta la base fisiologica generale della termoregolazione, che in un prossimo futuro si arricchirà degli elementi molecolari del sistema. Ma passando a un'analisi concreta si deve constatare che è difficile ancora oggi valutare nella realtà le molte modulazioni del sistema stesso, le quali variano da animale ad animale, a seconda della mole corporea, delle caratteristiche cutanee, di quelle della respirazione, delle variabili comportamentistiche, dello stato di sonno o di veglia, di riposo o di attività dell'organismo, ecc.
Le strutture che uno schema aggiornato (v. Hammel, 1968) può collegare insieme per l'operatività termoregolativa sono, come si è detto, molteplici. La parte relativa al sistema di controllo è costituita da una zona dell'ipotalamo anteriore includente la regione preottica e un'altra zona più caudale alle quali giungono afferenze da termocettori o sensori di temperatura, sia relativi alla temperatura corporea interna - e quindi anche alla temperatura del flusso sanguigno ipotalamico - sia relativi alla temperatura cutanea; altre afferenze sussidiarie sono quelle talamiche e talamocortico-ipotalamiche (oltre alle afferenze dalla sostanza reticolare) connesse anche con vie sensoriali d'altra natura, comprese quelle dei propriocettori posizionali del corpo. La parte relativa al sistema controllato è costituita dalle ghiandole a secrezione interna (la tiroide in particolare), dai muscoli, dal tessuto adiposo primario e dal fegato per ciò che riguarda il meccanismo di termoproduzione, la vascolarizzazione della cute (e delle mucose in animali diversi dall'uomo), la secrezione sudoripara (e mucosa) per ciò che riguarda la termodispersione. Negli animali anche i muscoli lisci piloerettori hanno notevole importanza termoregolativa contro il freddo.
I centri ipotalamici possono essere suddivisi, per ciò che riguarda la termoregolazione, in due aggruppamenti differenti, di cui uno, in posizione anteriore, attua o favorisce la termodispersione, e l'altro, situato posteriormente, attua o favorisce la termoconservazione e, se necessario, la termoproduzione. Naturalmente questi due aggruppamenti debbono essere intimamente collegati per realizzare una temperatura di riferimento.
Con un meccanismo che è quasi del tutto ignoto alla fisiologia, anche perché sono ancora da individuare le possibili basi molecolari della sensibilità termica (è verosimile che essa sia legata a reazioni chimiche endocellulari, o di membrana, particolarmente termodipendenti) e della sua trasmissibilità, specie a livello dei neuroni centrali, i nuclei ipotalamici e della regione preottica funzionano sulla base: a) della differenza fra la temperatura interna locale rispetto a una temperatura di riferimento (che è caratteristica della specie); b) di informazioni, sempre riguardanti la temperatura interna specie in alcuni animali, che provengono da termocettori situati nel canale vertebrale; infine c) di informazioni provenienti dai termocettori cutanei, i quali rendono conto della temperatura cutanea e quindi delle variazioni di quella ambientale.
L'esistenza di termocettori endovertebrali e perispinali è stata dimostrata per la prima volta nel cane dal gruppo di R. Thauer, con esperimenti con i quali nell'animale si manteneva una temperatura più elevata del normale nella testa e a livello cutaneo in modo da evitare risposte termogenetiche ipotalamiche dirette o mediate attraverso la sensibilità cutanea, e si raffreddava invece il canale vertebrale: l'animale rispondeva con brivido. Questi termocettori endovertebrali sembrano essere importanti anche in rapporto al drenaggio calorico proveniente, lungo i vasi sanguiferi, dalle zone termogenetiche viciniori quali quelle del tessuto adiposo primario (grasso bruno) paravertebrale, paraaortico, interscapolare ecc. (v. sotto). (Del resto, siccome alcuni esperimenti - v. Trabucchi, 1940 - indicano che gli animali cui siano state interrotte le vie nervose sotto l'ipotalamo e per questo siano diventati poichilotermi possono col tempo riacquistare buona parte della capacità termoregolativa, è stato ipotizzato che i centri ipotalamici siano un sistema unificatore e coordinatore di meccanismi molteplici dotati singolarmente di una certa autonomia; v. anche Rondoni, 1954). L'importanza di questi termocettori può variare a seconda degli animali (sono stati studiati anche nel gatto, nella cavia e nel piccione) in dipendenza della loro diversa sensibilità.
I termocettori cutanei sono, come è noto, di due tipi, alcuni per il caldo, altri per il freddo; in realtà l'unico stimolo positivo è l'energia calorica, e l'eccitamento dei sensori avviene o per la quantità assoluta di calore di fatto presente al momento o per il gradiente termico che si determina tra l'esterno e l'interno della cute in corrispondenza della zona sensibile quando l'ambiente non si trova allo zero fisiologico, cioè alla stessa temperatura della cute.
Alla distinzione fisiologica di sensori per il freddo e di sensori per il caldo non corrispondono terminazioni nervose definite e tanto meno morfologicamente distinte. Si sa solo che i termocettori sono più addensati nelle zone pelose della cute (e delle parti più esterne di alcune mucose, a seconda degli animali).
Il sistema termogenetico è evidentemente costituito da tutti gli organi e tessuti del corpo, ma quelli che più degli altri entrano nella parte ‛controllata' ai fini della termoregolazione sono in primo luogo costituiti dalle principali fucine termogenetiche, i muscoli, il tessuto adiposo primario e il fegato; ma l'importanza di quest'ultimo appare più che altro generica in quanto grosso viscere metabolico. I muscoli, invece, si prestano molto bene a variazioni metaboliche estemporanee per gli adattamenti immediati, in buona parte anche sotto il dominio della volontà. E infatti, dato che i muscoli possono intensificare grandemente e acutamente il loro metabolismo solo entrando in attività specifica, ne consegue che la reazione termogenetica è segnalata, dal fenomeno del brivido, che è un susseguirsi di contrazioni cloniche e tonico-cloniche di molti muscoli contemporaneamente.
Il tessuto adiposo primario di Hammar o ‛grasso bruno' è costituito da cellule a potenziale metabolico molto elevato, ricchissime per questo di mitocondri e capaci di produrre quantità di calore relativamente elevate, specie se, come è stato anche supposto, in condizioni di particolare stimolazione l'operatività mitocondriale venga almeno in parte disaccoppiata dalla fosforilazione. L'attività termogenetica del grasso bruno è massima negli animali neonati (e in questo caso va compreso anche l'uomo) e negli animali ibernanti (onde il vecchio nome di ‛ghiandola interscapolare ibernante'), in particolare al loro risveglio o durante la perfrigerazione (lotta contro il freddo).
Ma un controllo dei sistemi termoproduttori può avvenire solo per mezzo degli apparati di correlazione, quello nervoso e quello endocrino: quello nervoso è più idoneo per adattamenti rapidi relativamente fasici o acuti, quali si ottengono col brivido (e con i fenomeni vasomotori cui accenneremo), mentre quello endocrino è più lento ma più duraturo ed entra in azione negli adattamenti cronici, e in modo particolare nell'acclimatazione. Tuttavia anche alcuni ormoni possono intervenire in modo acuto, come nel caso degli ormoni medullo-surrenalici (v. sotto).
Gli ormoni più impegnati nel sistema controllato sono quelli tiroidei e quelli surrenalici, capaci di incidere profondamente sul metabolismo organismico; più precisamente si tratta del sistema ipofiso-tiroideo e di quello ipofiso-surrenale (v. Collins e Weiner, 1968).
Abbiamo parlato di ormoni tiroidei al plurale per tener conto di esperimenti, per altro ancora assai discussi e da alcuni addirittura negati, secondo i quali la tiroide produrrebbe anche un ormone inibente il metabolismo e quindi ipotermizzante, chiamato dal Mansfeld (v. i suoi tre contributi del 1940) ‛termotirina'; questi esperimenti erano stati preceduti da quelli del fisiologo italiano A. Montuori (v., 1904) sulla esistenza di fattori umorali come espressione della risposta al surriscaldamento e alla perfrigerazione e, in quanto passivamente trasmissibili col siero di sangue, inducenti analoghe risposte anche in animali normali (sieri di animali surriscaldati o perfrigerati producevano rispettivamente un abbassamento o un innalzamento della temperatura nei riceventi). Ma più recenti ricerche di Myers e Sharpe (v., 1968) dimostrerebbero fatti analoghi con perfusati di ipotalamo di animali raffreddati o riscaldati.
Della surrenale va presa in particolare considerazione, per risposte rapide e fasiche, la midollare, la quale secerne delle catecolammine, responsabili sia dell'effetto vasomotorio, sia dell'effetto metabolico (mobilitazione del glucosio dal glicogeno, attivazione della fosforilasi, ecc.); la noradrenalina è essenziale nella stimolazione termogenetica del grasso bruno (v. Smith e Horwitz, 1969).
Il sistema termodispersivo (agente sia nel senso di facilitare la termodispersione, sia, all'opposto, nel senso di ridurla e quindi di effettuare una termoconservazione) è costituito, per ciò che riguarda solo gli elementi dinamici (v. sopra), dal gioco vasomotorio, dall'accentuata evaporazione di acqua dalle superfici cutanee e mucose (animali), dalla secrezione delle ghiandole sudoripare e dalla polipnea, in alcuni animali. In questi ultimi anche dall'erezione dei peli e del piumaggio (mobilitazione degli elementi statici).
Il gioco vasomotorio, condizionato dal sistema nervoso e da quello endocrino, riesce a variare notevolmente l'isolamento dell'interno del corpo dall'ambiente esterno (isolamento già basalmente costituito dagli elementi statici); attraverso la vasodilatazione si facilita la dispersione del calore, mentre la vasocostrizione facilita la conservazione del calore interno del corpo, raffreddando la cute.
Le ghiandole sudoripare (le eccrine, non le apocrine) secernono un liquido a concentrazione salina molto bassa e di facile evaporazione; questa sottrae calore in ragione di 585 calorie per ogni litro evaporato; la sudorazione in ambiente molto caldo può spingersi sino a diversi litri. Ma la sudorazione comincia quando la temperatura esterna arriva a 32 °C; prima di questo livello è sufficiente la perspiratio insensibilis, cioè l'evaporazione attraverso gli strati della cute (20-30 g per ora). La secrezione delle ghiandole sudoripare è sotto il controllo neurocolinergico e può intervenire anche per stimolazione dei termocettori cutanei a temperatura interna normale; il fenomeno è però molto variabile individualmente per l'intervento di molti fattori aggiuntivi. All'opposto, si ha sudorazione anche in condizioni di temperatura ambientale normale, quando sia divenuta superiore alla norma la temperatura interna, come può avvenire nella febbre.
Non basta, naturalmente, aver individuato le sorgenti del calore e i meccanismi della sua controllata eliminazione all'esterno per comprendere la termoregolazione: occorre anche conoscere i mezzi con i quali il calore prodotto viene distribuito in modo che si crei un mezzo interno relativamente uniforme, e comunque che il sangue che arriva ai termocettori controllori sia termicamente un campione valido per la funzione regolativa. La distribuzione calorica dagli organi metabolicamente attivi (muscoli, grasso bruno) avviene per scambio transvascolare contro corrente vena arteria (conduzione transmurale) ed è favorita in certi distretti da particolari disposizioni vascolari in parallelo e in serie e da corticircuiti arterovenosi (v. Hammel, 1968; v. Smith e Horwitz, 1969).
Non tutte le parti del corpo però sono esattamente alla stessa temperatura interna, anche per i suddetti dispositivi che forniscono alcuni flussi calorici rispetto ad altri.
Infine si deve ricordare che a questi meccanismi puramente fisiologici si aggiungono quelli comportamentistici dell'organismo (specie terrestre) come insieme coordinato e capace - o con la ricerca volta a volta di un ambiente ecologico più idoneo, o assumendo atteggiamenti abituali coerenti (maggiore o minore superficie corporea esposta) - di favorire il processo termoregolativo.
L'inserimento di tutti questi settori operativi nel campo della termoregolazione in uno schema cibernetico non è facile e va ricordato che la relativa importanza dei vari settori della termoregolazione è molto varia a seconda delle specie animali.
Si è molto discusso sul valore dei termocettori cutanei nel processo termoregolativo; è apparso ai più, a un certo momento, che essi rappresentassero il mezzo per una regolazione fine e tempestiva evitando di utilizzare ordinariamente, come stimolo, una reale variazione di temperatura del sangue circolante e, come termocettori, quelli situati nel sistema nervoso centrale. Questi sarebbero intervenuti in una situazione di cimento più avanzata, quando l'influenza ambientale avesse realmente potuto modificare sensibilmente la temperatura interna, e la loro risposta sarebbe stata conseguentemente meno fine e calma, anzi parossistica e grossolana: qualcosa di simile alla differenza di reattività, nel controllo del respiro, tra chemocettori per la concentrazione del CO2 e chemocettori per la concentrazione di O2 nel sangue.
In realtà si è potuto constatare che la sensibilità dei sensori ipotalamici è molto elevata e bastano variazioni relativamente modeste (0,5 °C) nella temperatura ipotalamica per produrre una risposta termoregolativa cospicua. Ma le informazioni provenienti dai sensori cutanei manterrebbero, anche in questo caso, una notevole importanza nel complesso sistema di circuiti a retroazione (feedback loops), in quanto capaci di regolare la soglia di eccitabilità dei centri ipotalamici, sì da catalizzarne la reazione termoregolativa (v. Hardy, 1967; v. Hammel, 1968). Per l'uomo in particolare si considera che la stimolazione termica della cute sia prevalentemente collegata alla termogenesi chimica (metabolismo, tono muscolare, brivido), mentre la stimolazione termica dei centri ipotalamici sia prevalentemente collegata alla termodispersione o termoconservazione fisica (reazioni vasomotorie, sudorazione).
L'interazione fra informazione proveniente dai sensori della cute e quella realizzantesi a livello dei termocettori ipotalamici sarebbe, così intesa, un esempio di ‛regolazione della regolazione', attraverso espansioni sensitive periferiche che aggiustano tutta la reattività del sistema alle condizioni ambientali.
d) Caratteristiche risultanti della omeotermia
In un uomo la temperatura interna oscilla lievemente intorno ai 37 ‛C, subendo variazioni circadiane di 0,5-1 °C; in taluni soggetti - specie se giovani - esse possono anche raggiungere 1,4 °C. Tali variazioni sono legate al periodismo generale di attività dell'organismo e possono mutare col mutare di questo, con un'operazione di adattamento. Nella donna si sovrappone un periodismo più lungo in corrispondenza del ciclo mestruale, con una maggiorazione di 0,5 °C durante il periodo. Durante la gravidanza questa lieve ipertermia fisiologica accompagna quasi tutta la gestazione. Nel sonno la termoregolazione appare meno efficiente, ma la principale causa dell'abbassamento della temperatura e del bisogno di più efficiente isolamento dall'ambiente esterno è da cercarsi nell'assenza di attività.
Tali variazioni normali della temperatura possono trovare spiegazione ammettendo che il valore di riferimento inserito nel circuito di feedback negativo non sia molto nettamente definito, oppure che tale valore sia a sua volta suscettibile di spostamento a seconda delle condizioni ambientali e dei bisogni (e la interazione dei termocettori cutanei con i termocettori ipotalamici potrebbe esserne una prova). Un'altra possibilità è quella che pur essendo netto il valore di riferimento, esso sia facilmente soverchiabile dagli eventi periferici. Ciò è molto simile alla prima ipotesi, ma nel complesso queste considerazioni saranno da tener presenti in una discussione sulla genesi della febbre (v. Snell e Atkins, 1968).
Nel bambino piccolo, però, la temperatura corporea e intrinsecamente più indisciplinata, un buon ritmo circadiano instaurandosi successivamente con una maggiore regolarità nella sequenza dell'attività e del riposo; la termoregolazione infantile è anche più soggetta all'influenza di stimoli emozionali (ormonici); la presenza di un potente sistema termogenetico ormonodipendente quale il grasso bruno può in parte spiegare le improvvise salite della temperatura corporea dell'infante, così come la facilità alle elevazioni febbrili acute.
Nel vecchio invece la omeotermia, tendente a porsi a un livello inferiore, e la reattività a condizioni estreme di temperatura sono meno efficaci e più psicologicamente fastidiose.
3. Le ipertermie fisiologiche, patologiche e sperimentali
Un aumento fasico fisiologico del livello termico si ha, naturalmente, con una stimolazione termogenetica quale si verifica nell'esercizio muscolare.
Un livello termico modestamente più elevato della norma è riscontrabile nei soggetti ipertiroidei; notevoli ipertermie sono state invece osservate in pazienti con lesioni encefaliche che abbiano investito la zona dell'ipotalamo. Le ragioni di queste ipertermie sono evidenti.
Sperimentalmente si produce una ipertermia, spesso mortale, surriscaldando dall'esterno l'animale; un metodo più efficace è quello di porre l'animale in un campo elettromagnetico a onde corte (per es. 10-12 m). Con tale mezzo si possono anche adattare i ratti più rèsistenti a sopportare una temperatura interna di 44 °C (v. Remelli, 1955). Il punto critico per mantenere questa resistenza è l'inevitabile castrazione termica che interviene e che influisce negativamente, forse attraverso modificazioni ipofisarie. Del resto, animali castrati non si possono adattare.
Si conoscono molte sostanze ipertermizzanti; si potrebbe suggerire di dividerle in gruppi a seconda del settore del sistema omeotermico nel quale si inseriscono, deformandolo. Ma in realtà spesso il loro meccanismo d'azione è composito e complesso: per esempio la 1,2,3,4-tetraidro-2-naftilammina si ritiene che agisca centralmente provocando però una vasocostrizione periferica e divenendo quindi ipertermizzante per un effetto termoconservatore. Tuttavia questa ipertermia è inibita dal curaro (v. Bovet e Virno, 1952), onde è opinabile anche (o prevalentemente) un meccanismo termogenetico muscolare eventualmente sempre mediato da un'azione centrale: infatti la sostanza non interferisce direttamente con i processi ossidativi endocellulari (v. Buffa e altri, 1960).
L'adrenalina e la noradrenalina, mentre agiscono sui vasi periferici determinando va socostrizione e quindi ipertermia termoconservativa, intervengono altresì nel metabolismo dei glucidi attivandolo, e quindi in senso termogenetico; la noradrenalina attiva nel grasso bruno l'utilizzazione dei trigliceridi e quindi anche degli acidi grassi (v. Smith e Horwitz, 1969).
Nell'uomo, non nel coniglio, un particolare steroide C-19, l'etiocolanolone, che è un prodotto di degradazione del testosterone, produce ipertermia (v. sotto).
Le sostanze convulsivanti come la stricnina e la veratrina sono naturalmente ipertermizzanti; altre agiscono inibendo la sudorazione (atropina). Si può invece diminuire la perspiratio insensibilis attraverso un aumento della pressione osmotica (dosi forti di sostanze osmoticamente attive, azione di forti diuretici).
Una particolare posizione hanno le sostanze che intervengono sull'integrità del processo ossidativo endocellulare (mitocondriale) disaccoppiando le ossidazioni dalle fosforilazioni (dinitrofenolo, pentadorofenolo, dinitrocresolo; v. Buffa e altri, 1960 e 1963), onde il valore energetico dei processi ossidativi è, al limite, misurabile tutto in termini di termogenesi. Il fluoroacetato determina invece ipotermia (ed è comprensibile per la sua azione inibitrice sul metabolismo del piruvato e sul funzionamento del ciclo di Krebs); esso non influisce sulla ipertermia da tetraidronaftilammina se essa è già in atto, la impedisce però se viene somministrato prima.
Altre sostanze ipertermizzanti sono le sospensioni colloidali di zolfo, di carbonio, di metalli; le proteine eterogenee e i prodotti batterici: ma in questi ultimi casi il meccanismo può essere molto complesso e si avvicina o si identifica con quello della vera febbre di cui diremo tra poco.
Va infine notato che risultati peculiari e in parte contraddittori si sono ottenuti con l'iniezione di sostanze direttamente nell'ipotalamo o nei ventricoli cerebrali: mentre gli anestetici introdotti nei ventricoli producono da principio ipotermia (e ciò avviene anche con la normale somministrazione per via generale), in seguito producono intensa vasocostrizione, brivido e ipertermia; la 5-idrossitriptammina (serotonina) insieme a creatinina provoca forte ipertermia (2-3 °C), mentre nelle stesse condizioni l'adrenalina determina vasodilatazione e caduta termica. Si suppone che queste due sostanze fisiologiche abbiano importanza nella effettualità molecolare della termoregolazione.
4. La termoregolazione nella febbre
Già quasi un secolo fa i fondamentali esperimenti di C. von Liebermeister (v., 1875) avevano dimostrato che i soggetti febbricitanti termoregolano: cioè che non si tratta, nella febbre, dello scompenso o della rottura di un equilibrio fisiologico, bensì di uno spostamento di questo equilibrio. I soggetti febbricitanti, cioè, tendono a mantenere o a ripristinare la loro temperatura febbrile quando vengano cimentati con un aggravamento o con una sottrazione termica: in altre parole, reagiscono a questi interventi come i soggetti normali, mettendo in opera gli stessi meccanismi (sudorazione, brivido, ecc.).
Successivi esperimenti (v. Giuffrè e Nalli, 1903; v. Grimby, 1962; v. Cooper e altri, 1964) hanno sempre confermato questo dato, anche quando adottavano metodi di rilevamento delle reazioni termoregolative assai più fini ed elaborati. I soggetti febbrili mostrano di rispondere a differenziazioni termiche esterne in modo del tutto simile ai soggetti normali, a meno che non si intervenga con l'esperimento nel periodo di insorgenza della febbre, cioè quando il paziente è impegnato a raggiungere il suo nuovo riferimento termoregolativo, poiché in questo periodo la risposta è quantitativamente maggiore del normale se il senso della stimolazione è quello di generare ancora ipertermia o minore del normale se il senso dello stimolo è quello di generare vasodilatazione e quindi dispersione di calore (v. Snell e Atkins, 1968).
Sembra dunque che la prima modificazione del sistema termoregolativo nella febbre sia quella dello spostamento in alto del livello di riferimento, per cui la risposta organismica è subito correlata a questo nuovo livello piuttosto che alla effettuale temperatura interna del corpo; solo quando la febbre si è stabilizzata (febbre continua) la risposta del febbricitante ritorna simmetrica a quella del soggetto normale. Vi sono infatti prove che dimostrano che è proprio lo stato iniziale dei centri termoregolatori a decidere l'instaurarsi di un episodio febbrile, nel senso che un'artificiale elevazione termica locale dell'ipotalamo annulla o cancella l'azione di una sostanza febbrigena (pirogena), cioè non si determina uno stato febbrile generale (v. Snell e Atkins, 1968).
Pertanto si può concludere che: 1) in condizioni di omeotermia normale per la specie, il valore di riferimento nel sistema di feedback coincide con la temperatura effettiva dell'organismo, le cui modeste oscillazioni fisiologiche sono da interpretare secondo quanto detto nel cap. 2, È d; 2) nell'ipotermico (soggetto ipotiroideo mixedematoso, perfrigerato, sottoposto a sostanze e medicamenti ipotermizzanti) il valore di riferimento rimane normale, e solo la temperatura attuale è patologicamente ridotta; nell'ipertermico (soggetto ipertiroideo, portatore di feocromocitoma, esposto a eccessivo calore ambientale, a strapazzo fisico, sottoposto a sostanze e medicamenti ipertermizzanti) il valore di riferimento rimane normale e solo la temperatura attuale è più elevata di esso; 3) nel febbricitante invece è il valore di riferimento che viene elevato e la condizione è allora da definirsi febbrile indipendentemente dalla temperatura attuale (del momento dato). Con ciò si introduce un criterio obiettivo per distinguere le ipertermie dalle febbri: una ipertermia da dinitrofenolo, per esempio, non è in alcun modo omologabile alla febbre poiché tutta la sua storia naturale si svolge a livello dei tessuti, delle cellule. Tuttavia non è possibile escludere che le sostanze ipo- o ipertermizzanti ad azione periferica a una certa dose non lesionino gli stessi centri nervosi ipotalamici, cioè agiscano direttamente o indirettamente da sostanze febbrigene (pirogene).
5. Patogenesi della febbre
a) Introduzione
Tutto quanto si è detto finora indirizza a una precisa definizione della febbre, anzi rappresenta l'adozione di un concetto patogenetico che stabilisce che l'origine della febbre è centrale.
Ma la storia delle molte indagini e opinioni su questo argomento è tutta vivacemente combattuta tra opposte tendenze che anche in Italia hanno avuto assertori validi e acuti.
A una teoria centrale della febbre, discendente dagli esperimenti di Liebermeister a più riprese confermati e perfezionati (v. sopra, cap. 4), si è opposta una teoria ‛periferica' della febbre secondo la quale l'origine della ipertermia è fondamentalmente dovuta a una necessità termogenetica direttamente connessa con la presenza nel febbricitante di sostanze incongrue prodotte da microrganismi o liberatesi nel corso di danneggiamenti tessutali e cellulari e di degradazioni biochimiche): grazie all'azione di tali sostanze si esaltano i normali processi metabolici di trasformazione e di smaltimento, risultandone in definitiva un'extraproduzione calorica spesso non accompagnata o non sufficientemente accompagnata da un'adeguata contemporanea termodispersione.
La sollecitazione a vedere le cose in questo modo veniva più dalla clinica che dalla sperimentazione, e infatti il primo assertore di questa tesi fu in Italia il Murri (1873-1874). È comprensibile che un clinico, abituato a vedere insorgere la febbre in modo precipuo nelle malattie infettive e specie in quelle in cui vi sono danneggiamenti vari di tessuti e liberazione di prodotti incongrui da parte dei parassiti o la moltiplicazione dei parassiti stessi (vedi coincidenza, scoperta dal Golgi, dell'insorgenza del processo febbrile nella malaria e poussée parassitemica del plasmodio), fosse portato a stabilire un rapporto genetico diretto tra presenza di materiali abnormi e livello metabolico idrolitico-ossidativo. A questo potevasi aggiungere la quota metabolica dovuta all'accensione flogistica e alla fagocitosi: erano note alla clinica le crisi reiterate di piressia alternate a defervescenze non solo in malarici, ma anche in soggetti con suppurazioni migranti o portatori di ascessi con possibilità di svuotamento periodico all'esterno (per es., ascessi polmonari).
L'ipotesi veniva convalidata anche dall'osservazione di quei casi nei quali la febbre, non dovuta a infezione, era tuttavia sempre legata alla introduzione di sostanze incongrue o alla loro liberazione da denaturazione fisica e chimica dei costituenti organismici: febbre dopo l'introduzione di siero eterogeneo (malattia da siero), modesto rialzo febbrile dalla vecchia autoemoterapia, rialzi febbrili da causticazioni, da ustioni di superfici non eccessivamente piccole del corpo (ma talora anche molto limitate), febbre nelle necrosi ischemiche anche non infettate, quale quella dell'infarto miocardico, ecc. Nel contempo non venivano invece messe a fuoco alcune palesi incongruenze come la non corrispondenza, in molti casi e nei due sensi, tra entità della febbre ed estensione plausibile del danno, da un lato, e cessazione di uno stato febbrile ancor prima che il danno tessutale fosse riparato, dall'altro.
Un contributo dottrinale e sperimentale a questa veduta è stato portato in Italia da G. Vernoni e da diversi suoi collaboratori (v. Vernoni, 1916, 1917, 1940, 1948).
Innanzi tutto, rifacendosi anche a esperimenti eseguiti dallo stesso Murri, si è negata gran parte del valore ai dati ricavati da animali a diencefalo escluso, che sono incapaci di febbricitare; e ciò sia perché quell'intervento è in grado di alterare comunque il metabolismo generale indipendentemente dall'origine prima della febbre, sia perché si era nel frattempo visto che animali mesencefalici potevano dopo un certo tempo - se stimolati - ancora febbricitare. Ciò accade anche nell'uomo (v. Thauer, 1939). Ma questo argomento era d'altro canto sopravvalutato poiché si è poi visto che quegli animali riacquistavano la termoregolazione, cioè in essi entravano in funzione altri centri inferiori (v. Trabucchi, 1940), onde la ‛centralità della febbre' si poteva sempre riproporre come spostamento della temperatura di riferimento da parte di quei centri.
In realtà anche nella scuola di Vernoni (v., 1940 e 1948) non veniva escluso uno spostamento di un ‛meccanismo' di termoregolazione, solo che si metteva l'accento su un meccanismo regolatore periferico e in particolare legato al metabolismo muscolare, poiché questo tessuto rappresenta il 40% del corpo.
Un esaltato metabolismo muscolare, in particolare glicolitico e ossidativo, simile a quello connesso con i fenomeni della contrazione, ma non accompagnato da contrazione, finisce per produrre un quantitativo di calore di difficile drenaggio esterno, poiché la vascolarizzazione di riposo del tessuto muscolare è molto meno estesa e attiva che nella contrazione fisiologica (v. Krogh, 1922); di qui il ristagno calorico e l'elevazione della temperatura, ossia la febbre.
A sostegno di questa veduta si portava la riconosciuta compromissione fisiopatologica e persino morfologica delle fibre muscolari scheletriche nel corso delle febbri alte e prolungate (v. Caldera, 1942), la capacità ipotermizzante del massaggio muscolare in animali (e uomini, ma con minore evidenza) febbricitanti (v. Spinelli, 1934), ma soprattutto la constatazione di un aumento di attività metabolica ossidativa (sia pure entro i grossi limiti e i metodi allora a disposizione) di muscoli come di altre cellule (fegato) di animali resi febbricitanti (v. Vernoni, 1916 e 1917; v. Spinelli, 1932). Naturalmente si trattava di tessuti o cellule sopravviventi studiati in vitro a temperatura eguale rispetto ai controlli, poiché gli esperimenti in vivo sono suscettibili di critica nel senso che l'aumento dei processi metabolici dell'organismo febbricitante può essere semplice conseguenza della febbre, secondo la legge di Van't Hoff.
Un altro punto dottrinale che è sempre stato di ostacolo, per i sostenitori della ‛teoria periferica', ad accettare l'ipotesi neurogena di uno spostamento in alto del valore di riferimento per l'omeostasi della temperatura corporea, è la considerazione che nei confronti della molteplicità (che via via andava crescendo) dei mezzi chimici e biochimici atti a produrre febbre, non se ne trovava alcuno che producesse ipotermia; si opinava invece che un centro nervoso può tanto essere esaltato quanto depresso o che - se i centri sono due, uno per la termoproduzione e l'altro per la termodispersione - si potrebbero sempre trovare sostanze agenti ora sull'uno ora sull'altro. A parte il fatto che esistono sostanze ipotermizzanti (antipiretici, barbiturici, ecc.), e che la stessa tetraidro-β-naftilammina che è ipertermizzante nel ratto tenuto a neutralità termica è invece ipotermizzante se l'animale è inizialmente tenuto a temperatura più bassa della normalità (v. Giaja e Dimitrijevic, 1933), è tuttavia vero che nel rapporto infettivo-flogistico, cioè nelle condizioni patologiche naturali, la deviazione quasi universale della termoregolazione è nel senso della febbre, cioè dell'ipertermia febbrile, e non l'inverso (vedremo però che uno stesso pirogeno può produrre febbre in una specie animale e ipotermia in un'altra). Si vedrà che questa esigenza ‛logica' di un meccanismo patogenetico unitario, nonostante la diversità della etiologia febbrile, sarà soddisfatta dalle più moderne ricerche sui pirogeni. D'altra parte la scoperta dei pirogeni agenti, specialmente quelli esterni, in quantità minimali, ha messo fuori causa in modo definitivo ogni semplicistica attribuzione dell'elevazione termica febbrile ai processi biochimici legati allo smaltimento di materiali incongrui.
Tentativi positivi di riprodurre un processo febbrile mediante i prodotti dell'infiammazione (per es. il pus) indussero a ritenere già oltre 100 anni fa che, comunque, la febbre risulta quasi sempre legata al binomio infezione- infiammazione. Tra i primi ricercatori che si sono propriamente orientati alla ricerca di sostanze pirogene specifiche, cioè agenti centralmente in senso febbrile e appartenenti ai prodotti batterici, è stato il patologo italiano E. Centanni. Nel 1893-1896 egli comunicò di avere isolato da vari germi una sostanza ancora tossica ma fortemente pirogena, probabilmente non proteica perché termoresistente, che chiamò ‛pirotossina'. Successivamente la purificò del residuo potere tossico e confermò non trattarsi di una proteina, bensì, ipoteticamente, di polisaccaridi (ma siamo già al 1937).
Nel frattempo si fanno chiari certi dati, dapprima sporadici e poi sempre più frequenti e concordi, di febbri elevatissime prodotte da medicamenti disparati che non avevano alcuna relazione chimica tra loro, iniettati parenteralmente. Si scoprì che l'effetto febbrigeno era dovuto non al medicamento (tanto è vero che variava da preparazione a preparazione dello stesso), ma al comune solvente di esso, cioè alla soluzione fisiologica, all'acqua con cui questa era preparata ecc. (v. Hort e Penfold, 1911). Invece un'acqua accuratamente bi- e tridistillata non produceva più alcuna febbre e risultò evidente che l'effetto febbrigeno dipendeva da minime contaminazioni batteriche dell'acqua distillata prima usata. Tali contaminazioni lasciavano nell'acqua sostanze responsabili dell'elevazione febbrile (talora anche molto elevata), denominate dalla Seibert (1923) ‛pirogeni'.
b) I pirogeni esogeni
Si è così aperta una strada molto fruttuosa almeno per l'interpretazione della febbre insorgente nel corso di malattie infettive. Si è però visto ben presto che anche all'interno di questo settore l'interpretazione non poteva essere univoca (v. Snell e Atkins, 1965 e 1968).
I primi e più potenti pirogeni furono preparati da batteri gram-negativi presenti nell'aria come nell'acqua, non patogeni (v. Seibert, 1925; v. Seibert e Mendel, 1923; v. Bourn e Seibert, 1925; v. Co Tui e Schrift, 1939 e 1942).
Nel 1934 H.M. Banks tentò di specificare i microrganismi più responsabili che individuò negli Pseudomonas; Co Tui, Benaglia e altri collaboratori, invece, diressero le loro indagini su vari altri gram-negativi (compresa la Eberthella typhosa; v. anche De Merre e Probey, 1945). Le sostanze pirogene isolate da questi batteri furono riconosciute come lipopolisaccaridi e pertanto sufficientemente termostabili (v. i dati già rilevati dal Centanni); il loro peso molecolare è piuttosto elevato (circa 0,5-20•106) e molto verosimilmente fanno parte della parete cellulare dalla quale si liberano con difficoltà, per cui rientrano nel tipo di sostanze tossiche batteriche a tipo endotossine alle quali sono state infatti omologate. Talora i due termini sono usati come sinonimi e per questo quando occorrerà useremo anche noi il termine di endotossina per indicare il pirogeno esogeno, al fine di evitare confusione con altre notazioni semantiche. Del resto i pirogeni non si limitano all'azione febbrigena, anche se molto purificati, ma provocano anche modificazioni della reattività del sistema reticolo-istiocitario e in generale della sensibilità vascolo-tessutale, onde sono i mediatori della reazione di Sanarelli-Schwartzmann; caratteristica quasi costante è di possedere anche un'attività antileucocitaria su cui dovremo tornare. Infine, in dosi elevate agiscono sul sistema circolatorio generale provocando shock e morte. Invece le dosi capaci di dare febbre (per es. quelle reperibili nell'acqua anche semplicemente distillata) sono piccolissime: per il coniglio la dose minima di endotossina per avere un rialzo febbrile è di 0,0001-0,001 μg/kg. Anche per l'uomo la minima dose è di quest'ordine di grandezza. Da ciò è facilmente comprensibile come la liberazione di soluzioni acquose dalle endotossine pirogene costituisse all'inizio un problema difficile, tenuto conto. anche della loro termostabilità.
Il fatto che i microrganismi gram-positivi non posseggano endotossine né producano esotossine direttamente pirogene è stato naturalmente il limite maggiore della valutazione del meccanismo pirogeno delle endotossine come stimolazione diretta dei centri ipotalamici, in quanto si hanno in realtà febbri nelle infezioni da gram-positivi, come da vari altri agenti infettanti, virus compresi. Questa considerazione ha riportato l'attenzione su tutte le altre febbri non legate a fatti infettivi, riaccendendo così l'interesse a una spiegazione unitaria della febbre.
In questa direzione si sono dapprima messe in luce alcune caratteristiche del processo non immediatamente spiegabili con l'ipotesi che l'endotossina agisca direttamente sui centri ipotalamici: 1) esiste in generale un periodo di latenza che può essere anche di oltre 30 min (nell'uomo anche 45 min) tra l'iniezione endovenosa di una dose opportuna di endotossina e l'insorgenza della febbre; 2) tale latenza si riduce molto se si inietta nell'animale in esperimento siero di sangue di altro consimile animale pretrattato da breve tempo con endotossina; 3) animali resi leucopenici non rispondono all'endotossina, ma rispondono al siero di animali normali pretrattati con l'endotossina (v. Atkins e Snell, 1965; v. Snell e Atkins, 1968); 4) l'andamento della reazione febbrile dopo la somministrazione di endotossina è spesso caratteristicamente bifasico, i due massimi essendo distanziati di 2 o 3 ore; 5) tale bifasicità non si riscontra usando i sieri di animali pretrattati con endotossina; 6) con ripetute dosi di una endotossina si ha una refrattarietà o ‛tolleranza' verso di essa, la quale, però, risulta aspecifica, cioè valida anche per altre endotossine; tale tolleranza viene rimossa dal bloccaggio del sistema reticolo-istiocitario (v. Grosse-Brockhoff, 1969).
La latenza osservata nel caso dell'endotossina appare essenziale nell'analisi del meccanismo reale sottostante, perché essa si verifica anche facendo pervenire direttamente l'endotossina nell'ipotalamo, onde era difficile ipotizzare un'azione diretta sui centri termoregolatori. Ciò che invece veniva ad aprire una nuova strada da percorrere nello studio della patogenesi della febbre era l'osservazione che, anche in questo caso di introduzione locale dell'endotossina, l'insorgenza della febbre era preceduta da un accumulo flogistico, cioè leucocitario, intorno alla zona di lesione prodotta dall'endotossina.
Si è arrivati così a concludere che con molta probabilità l'effettiva azione pirogena doveva essere almeno in questo caso esercitata da sostanze derivate dai leucociti polimorfonucleati dell'animale in esperimento (e anche da monociti e da altre cellule in diversi altri casi, come vedremo) dette pertanto ‛pirogeni endogeni'. Veniva quindi a rendersi possibile un'interpretazione unitaria della reazione febbrile anche quando non dovuta a endotossine, ma a gram-positivi, a virus, a particelle colloidali e fagocitabili, a lesioni ematiche e tessutali di varia natura, in particolare quelle da sensibilizzazione batterica e non batterica. Va subito detto che in tutti questi casi il periodo di latenza è più lungo che con la endotossina.
c) I pirogeni endogeni
La constatazione che anche le endotossine, o pirogeni esogeni, agiscono mediante la liberazione in seno all'organismo, specialmente dai leucociti polimorfonucleati e dai monociti, di un pirogeno endogeno direttamente responsabile della stimolazione dei centri termoregolatori, non solo ha reso ragione delle incongruità della precedente sperimentazione, ma ha pure fornito la base unitaria del meccanismo patogenetico del processo febbrile quale era richiesta anche dai fautori delle teorie ‛periferiche': la molteplicità degli stimoli primari ed esogeni si generalizza in un danno cellulare (e per le febbri più comuni ed elevate si specifica nel danno leucocitario) e per ciò stesso si traduce in uno stimolo secondario uniforme (o abbastanza uniforme) che è il diretto responsabile dell'elevazione termica. Da ciò si deduce che: 1) la latenza nell'azione della endotossina, che si riduce (10-20 min) usando invece pirogeno endogeno già liberato in un animale pretrattato con endotossina, è dunque spiegabile con la necessità di una reazione primaria fuori dal sistema termoregolativo; 2) la mancata o diminuita risposta degli animali leucopenici diviene egualmente comprensibile; 3) la bifasicità della risposta febbrile da endotossina (che manca invece con pirogeno endogeno) dipende dal fatto che, dopo il primo attacco antileucocitario dell'endotossina (a cui segue il primo massimo della temperatura e uno stato di leucopenia), si ha una risposta organismica con leucocitosi e quindi, rimanendo ancora endotossina disponibile, un nuovo, talora più elevato, massimo di temperatura; 4) una lenta iniezione endoarteriosa nella carotide di pirogeno endogeno produce una risposta più pronta e più elevata a parità di dose di una iniezione endovenosa, poiché con l'iniezione endocarotidea si raggiungono rapidamente e con maggiori concentrazioni di pirogeni endogeni i centri ipotalamici; invece con l'endotossina non si riscontrano differenze di questo genere, poiché essa non è lo stimolo diretto (v. King e Wood, 1958); 5) la quantità di pirogeno endogeno circolante è proporzionale all'elevazione termica febbrile ottenuta (o ottenibile e misurabile per trasporto passivo) (v. Atkins e Wood jr., 1955); 6) la tolleranza acquisibile dagli animali per ripetute somministrazioni di endotossina o di virus (v. Atkins e Huang, 1958) non si verifica usando direttamente pirogeni endogeni; anzi animali resi tolleranti all'endotossina e pertanto non più responsivi possono ancora febbricitare sotto lo stimolo pirogeno endogeno (il pirogeno endogeno non è antigene per lo stesso animale); 7) molti altri agenti patogeni o situazioni patologiche che provocano o si accompagnano a febbre possono liberare pirogeni endogeni, anche con meccanismi diversi, dai leucociti, dai monociti (per es. in seguito ad azione aggressinica, leucocidinica, ecc.), o dopo espletamento della fagocitosi, o per la stimolazione stessa della fagocitosi; forse è concepibile, e alcuni esperimenti lo confermano (v. sotto), che anche da altre cellule o da tessuti altrimenti lesionati (reazioni immunoallergiche, processi necrobiotici) si liberino pirogeni endogeni; in tutti questi casi la latenza è, come si è detto, ancora maggiore che con le endotossine; 8) mentre la febbre da endotossina è accompagnata da altre azioni tossiche proprie di questo genere di sostanze, quella da pirogeni endogeni è del tutto priva di tali ulteriori effetti: la stessa leucopenia iniziale non si verifica naturalmente o - perlomeno - non nel modo molto marcato caratteristico dell'endotossina.
All'individuazione dei pirogeni endogeni hanno servito più linee di ricerca convergenti: cioè non solo quella derivante dall'osservazione delle condizioni di realizzazione del processo febbrile evocato dalle endotossine, ma anche quella - di origine molto antica - derivata dallo studio dell'azione febbrigena degli essudati e dalla ricerca in essi, appunto, di sostanze responsabili di questa reazione generale organismica che accompagna così spesso le manifestazioni infiammatorie per poco che siano sufficientemente estese. Alle vecchie ricerche già ricordate di Weber e di Billroth se ne sono aggiunte molte altre, culminanti nei lavori del Menkin (v., 1944 e 1945) intesi a trovare i mediatori molecolari di tutti gli eventi della flogosi e delle reazioni generali che l'accompagnano (leucocitosi, febbre). Questo autore trovò infatti nella frazione euglobulinica degli essudati flogistici acuti (ottenuti ‛sterilmente' con sostanze piogene quali la trementina) una frazione ad azione pirogena, da lui denominata ‛piressina' (pyrexin), dissociabile da altre frazioni consimili ad azione necrotizzante. La piressina non era dializzabile, era termostabile, si trovava anche nel siero degli animali portatori di un vasto processo infiammatorio e poteva anche comparire nelle orine. È possibile, ma non certo, che nella frazione isolata dal Menkin, pur non ancora chimicamente pura e tanto meno definibile, fosse contenuto vero pirogeno endogeno leucocitario. Il Menkin inseriva i suoi risultati in un quadro anche troppo coordinato, nel senso che secondo le sue ricerche dallo stesso essudato si potevano ricavare, diversamente frazionando, i principali responsabili molecolari degli eventi della flogosi: la necrosi iniziale con vasodilatazione (necrosina); la diapedesi leucocitaria (leucotassina); la leucocitosi (fattore promovente la leucocitosi, FPL); la febbre (piressina). D'altronde, dopo la constatazione della facilità di avere pirogeni esogeni in ogni preparazione, risulta assai difficile, a meno di non fornire prove molto accurate, concludere che un determinato estratto pirogeno è solo pirogeno endogeno, anzi, proprio nel caso della piressina di Menkin fu in seguito documentata la possibilità dell'esistenza in essa di un pirogeno esogeno a tipo endotossina (v. Bennett e Beeson, 1953). Era stato infatti il Beeson, nel 1948, a dimostrare per primo un effetto pirogeno da trasferimento sicuramente sterile di leucociti (del sangue o di essudati sterili). Che questo risultato fosse propriamente quello di un vero e diverso pirogeno endogeno rispetto a quello di Menkin è chiarito dalla sua azione pirogena più rapida (e anche di più breve durata), dall'essere termolabile e dalla sua incapacità a determinare uno stato di tolleranza.
Dunque il pirogeno endogeno è sicuramente derivabile (anche per incubazione in vitro con endotossina) dai granulociti neutrofili per azione di endotossina o di altri agenti. I leucociti normali, non attivati o non incubati, producono solo quantità assai piccole di pirogeno endogeno; dopo attivazione la quantità di pirogeno endogeno rilasciato è abbondante e l'effetto pirogeno è proporzionale alla quantità di leucociti; esperienze in vivo dimostrano che esiste una quantità critica di leucociti per il determinarsi dell'effetto pirogeno.
Al pirogeno endogeno si attribuisce la natura di una lipoproteina ma di un peso molecolare sensibilmente minore di quello dell'endotossina: 1-2•104. La caratterizzazione chimica è ancora da fare.
I pirogeni endogeni liberabili da altre cellule diverse dai granulociti e dai tessuti non sembrano essere essenzialmente differenti da quelli leucocitari, solo che è più difficile estrarne in quantità sufficiente; i tessuti finora esaminati sono quello muscolare, il fegato, il rene, il cuore, il polmone e la milza; non sempre le curve termiche sono pronte e nette come quelle del pirogeno endogeno leucocitario, specie per il fegato e il rene. La quantità di tessuto necessaria è varia, potendo talora bastare 2,5 g di tessuto fresco, quindi in ogni caso con una cospicua differenza rispetto all'erogabilità leucocitaria (naturalmente si è provveduto non solo a evitare la contaminazione endotossinica, ma anche a escludere che l'effetto fosse dovuto ai leucociti contenuti nell'interno dei tessuti esaminati).
Si pone qui la questione se determinati stimoli e lesioni tessutali possano, come nel caso dei leucociti, ‛attivare' le cellule dei tessuti e liberarne quantità maggiori di pirogeni endogeni. Risultati positivi sono stati ottenuti con macrofagi nel caso di una loro indotta ipersensibilità, per es. alla tubercolina o ad antigeni batterici, seguita da preincubazione in vitro con l'antigene: il pirogeno endogeno ottenibile cresce sensibilmente (v. Hall e Atkins, 1959). Allo stesso modo è pensabile avvenga per diversi tessuti, per es. linfogangli (v. Atkins e altri, 1967).
Per i granulociti circolanti lo studio delle tecniche di attivazione per ottenere maggiori quantità di pirogeno si è molto approfondito, anche se ancora i risultati non sono del tutto chiari (v. Bornstein e altri, 1963).
Cooper (v., 1963) nell'ipotesi che i leucociti possano svolgere un'azione pirogena non solo per liberazione di un loro pirogeno endogeno, ma anche per azione sui Batteri con liberazione dai loro lipopolisaccaridi di un composto lipidico pirogeno, propone la sigla LBP - leucocyte-bacterialpyrogen incubate - per definire questo sistema.
Si è detto che senza alcun pretrattamento i leucociti possono liberare solo quantità molto piccole di pirogeni endogeni, meno del 5o% della quantità ottenibile, per es., per azione su di essi dell'endotossina. Ma Kaiser e Wood jr. (v., 1962) dimostrarono che anche leucociti di essudati sterili e non prodotti da endotossina potevano essere forti donatori di pirogeno endogeno dopo incubazione in soluzione fisiologica a 37 °C. Evidentemente il fatto stesso di provenire da un essudato realizzava la liberazione di pirogeni endogeni. Ciò fu confermato dalla dimostrazione che, incubando nella parte liquida di un essudato leucociti normali, questi acquistano la capacità di liberare notevoli quantità di pirogeni endogeni. All'opposto il siero o il plasma normali hanno un potere inibente sulla liberazione di pirogeni endogeni da parte di granulociti di essudato; tale inibizione sembra sia legata alle concentrazioni del Ca2+, e particolarmente a quelle del K+ extracellulare. Anche l'endotossina è incapace di attivare i leucociti se nel mezzo abbonda il K+, a meno che non si aggiunga plasma normale: quest'ultimo effetto sembra sia legato anche alla presenza di Mg2+. Tutto ciò ha fatto pensare che abbiano parte nel fenomeno di attivazione i meccanismi di trasporto attivo cationico e anionico attraverso le membrane leucocitarie.
Queste constatazioni, anche se finora abbastanza disperse e talora non univoche, permettono tuttavia di affermare che il meccanismo di liberazione del pirogeno endogeno dai granulociti, cioè quello del fattore più frequente e forse più attivo della febbre, è collegato con altri meccanismi che hanno luogo nel corso delle infezioni e che nell'insieme rappresentano un gioco reattivo complesso con aspetti regressivi e progressivi strettamente intrecciati: la flogosi, la leucocitosi, l'aggressività batterica e virale o dei complessi antigene-anticorpo sui leucociti, in particolare sulle loro membrane (per es. l'azione delle leucocidine e di alcune lisine), la fagocitosi.
Può essere solo suggestivo (ma appare di notevole significato) che, per esempio, sia nell'azione delle leucocidine, sia nella fagocitosi, sia nell'attivazione e liberazione del pirogeno endogeno si debba tener conto di fatti comuni e molto peculiari: 1) il primum movens è sempre una modificazione della membrana esterna influenzata dai cationi del mezzo; 2) in ogni caso consegue un cambiamento metabolico dei leucociti associato a una degranulazione con ‛secrezione' all'esterno dei prodotti dei granuli (sembra anzi che pirogeno endogeno possa essere estratto appunto dalla frazione granulare dei leucociti; v. Herion e altri, 1966); si ha comunque attivazione del consumo di O2 dei leucociti, attivazione che è cianuro-insensibile; 3) in ogni caso si ha inibizione da agenti che bloccano i gruppi − SH ecc. Con ciò non si vuole in alcun modo andare oltre la persuasione che tutti questi fenomeni siano tra loro collegati proprio in quanto hanno un comune substrato reattivo, la cellula leucocitaria; infatti, anche nell'azione delle leucocidine (v. Woodin, 1968) si distingue una fase specifica da una aspecifica, ed è proprio questa che troviamo comune ad altre fenomenologie che hanno come attore il granulocito neutrofilo. A questo punto appare evidente la centralità del polimorfonucleato nella più immediata reattività organismica al parassitismo. Recentissime osservazioni sulla produzione di pirogeno endogeno da parte di leucociti umani dimostrano l'interconnessione fra fagocitosi e produzione di pirogeno endogeno (v. Nordlund e altri, 1970).
Il pirogeno endogeno è prodotto abbastanza durevolmente dai leucociti ematici attivati dagli essudati e ciò sembra implichi una durevole produzione di proteina, cioè un impegno DNA-RNA (v. anche esperimenti di bloccaggio con puromicina e altri inibitori; v. Atkins e altri, 1967; v. Nordlund e altri, 1970). È anche possibile che, almeno come reattività immediata, l'attivazione operata dallo stimolo consista in una modificazione enzimatica di un precursore prima inattivo.
Per ultimo ricordiamo che la già menzionata (v. cap. 3) ipertermia dell'uomo da etiocolanolone è verosimilmente una vera febbre, cioè un'ipertermia mediata da emissione di pirogeno endogeno; siccome altri steroidi hanno o possono avere un effetto simile, si parla anche di ‛febbre da steroidi', la cui azione sui leucociti differisce da quella dell'endotossina per non produrre leucopenia (non hanno azione leucocidica), per non determinare tolleranza e per essere strettamente specifica per l'uomo.
6. Significato della febbre
L'analisi moderna sul determinismo della febbre ha contemporaneamente contribuito anche a una definizione più esatta del termine e alla sua distinzione da quello di ipertermia: nelle ipertermie non febbrili, cioè non causate da pirogeni endogeni, manca la variazione del valore termico di riferimento, cioè il sistema cibernetico risulta invariato, anche se sovraccaricato (v. sopra, cap. 4).
Come i pirogeni endogeni riescano a variare il valore di riferimento non è noto, come non è noto, del resto, il modo in cui i sensori ipotalamici ed extraipotalamici riescano a tradurre in impulsi nervosi le variazioni termiche ambientali (cioè - come si è detto - non è noto il meccanismo molecolare della termoregolazione). Vi sono alcune prove di una mediazione da parte della 5-OH-triptammina (v. Klastersky e Kass, 1970). Forse il compito del futuro chiarimento sarà facilitato dalla constatazione dell'esistenza di centri distinti per la termoproduzione e per la termodispersione. Nell'instaurarsi della febbre il primo fenomeno è di termoconservazione (vasocostrizione) e di termoproduzione immediata (brivido muscolare), mentre un tono più elevato di termoproduzione generale si instaura secondariamente; nei bambini con grasso bruno ancora presente e funzionante si ha anche il contributo termoproduttore da parte di questo sistema, cui si attribuiscono le accensioni termiche molto vivaci proprie di questi soggetti.
Invece nelle ipertermie ‛chimiche' quali quelle da dinitrofenolo o da tirossina l'azione primaria è, come si è già accennato (v. cap. 4), solo ‛periferica', sui processi ossidativi, senza variazione, attraverso la mediazione dei pirogeni endogeni, dello status del sistema cibernetico (naturalmente, alcuni veleni possono essere pirogeni nella misura in cui lesionano, poniamo, i leucociti o altre cellule o tessuti; se agiscono direttamente sull'ipotalamo determinano ipertermie ma anche ipotermie a seconda della qualità loro o dello stato dei centri).
Mentre negli essudati delle flogosi umane si sono riscontrati molti pirogeni, nel sangue circolante la loro ricerca è stata negativa, forse per effetto della notevole diluizione. Inoltre nell'uomo si riscontrano febbri in diverse circostanze non flogistiche (e oltre ai casi di necrosi tessutale): nel feocromocitoma, nei linfomi, in altri tumori, in discrasie ematiche, in malattie del collageno ecc. In molti di questi casi i leucociti umani appaiono carichi di pirogeno che liberano facilmente se posti in contatto con la parte liquida di essudato sterile di coniglio. Ciò pare si verifichi anche nelle forme allergiche e nell'infarto cardiaco. In ogni modo, per molte delle suddette malattie non si possono escludere rapporti con i fenomeni immunitari e autoimmunitari e le lesioni cellulari conseguenti; a questo proposito va ricordata la febbre che accompagna il lupus eritematoso e l'aggressione immunologica antileucocitaria che si verifica in questa sindrome (fenomeno LE).
Si è molto discusso sul significato evolutivo-difensivo del fenomeno febbrile, specie di quello che consegue alle infezioni (che è di gran lunga il più importante). Non sappiamo se i pirogeni endogeni che stanno alla base del determinismo febbrile, oltre a determinare questa conseguenza, abbiano altri uffici importanti nella reazione antinfettiva, analogamente all'interferon (per fare un esempio) nelle malattie virali. In mancanza di questa possibilità è da discutere se la febbre in quanto tale possa essere di utilità nella difesa antinfettiva: la leucocitosi che è inerente alla febbre, almeno come risposta secondaria alla leucopenia iniziale da pirogeni esogeni, può avere un significato più vasto e complesso; in molti casi la febbre è accompagnata da un aumento dell'indice opsonico del siero, il quale appare seguire molto da vicino le variazioni febbrili (v. Vernoni e Pirone-Naconetschnii, 1938). Tuttavia, questo parallelismo temporalmente molto stretto, come è stato osservato nelle febbri malariche, suggerisce un meccanismo diverso da una semplice stimolazione della plasmopoiesi. Comunque, il generale aumento del livello termico con cui si svolgono, nella febbre, le funzioni cellulari e organismiche può esser considerato in linea con l'attivazione dei processi difensivi (entro certi limiti di elevazione termica) sia pure come effetto secondario, ma interconnesso. La vecchia persuasione medica che la vivacità della febbre nelle forme acute sia un indice di pronta ed efficace reazione organismica (cfr. i bambini sani con gli anziani o i debilitati) è probabilmente basata su un'esperienza certa; ma siccome alla luce dei nuovi fatti febbre e disponibilità leucocitaria sono tra loro collegate, una parte dei fenomeni su cui poggia tale persuasione può essere costituita da situazioni diverse da quelle realizzate dalla febbre in quanto aumento della temperatura.
È difficile invece pensare che l'elevazione termica del febbricitante possa essere particolarmente dannosa per i microrganismi nelle malattie infettive; ma in almeno due casi questo apparentemente si verifica, tanto che nel passato si è giustificata - in deficienza di altro - una ‛piretoterapia' (malarioterapia): per la sifilide tardiva e per la gonorrea cronica (ma è ancora da dimostrare che si trattasse di un effetto antimicrobico della temperatura febbrile).
7. Andamento e forme cliniche della febbre
Una febbre può iniziare repentinamente (insorgenza o incremento acuti), e allora è accompagnata da intenso brivido (per es., malaria); altrimenti, se l'incremento avviene gradualmente (per es., tifo addominale) il brivido iniziale può mancare; naturalmente il febbricitante è pronto a rabbrividire in qualunque fase della sua febbre se viene posto in ambiente più freddo della sua nuova neutralità termica. Tale brivido febbrile è dato infatti dalla sensazione di freddo dovuta: a) allo spostamento del sistema cibernetico termoregolatore operato dai pirogeni e b) (secondariamente) alla stimolazione dei termocettori cutanei per raffreddamento della cute dovuto alla vasocostrizione (v. Grosse-Brockhoff, 1969).
Una volta raggiunto il valore più elevato (acme o fastigio), la temperatura può rimanervi per un tempo più o meno lungo (periodo di stato) o invece cadere di nuovo attuandosi una defervescenza, che può essere anche solo temporanea (v. sotto). Quando la defervescenza è rapida, cioè avviene per crisi (per es., polmonite), essa è accompagnata da intensa sudorazione; quando invece anche la defervescenza o decremento febbrile è graduale, cioè avviene per usi (per es., tifo addominale), il fenomeno della sudorazione profusa può mancare.
In generale, durante il periodo di stato si osservano variazioni circadiane (±0,5-1° C) come nei soggetti normali, accentuandosi il livello termico febbrile nelle ore pomeridiane. Quando il periodo di stato dura abbastanza a lungo (per es., diversi giorni) e le variazioni giornaliere sono solo nell'ambito prima riportato, la febbre è detta continua (per es., tifo addominale); quando le variazioni circadiane o a intervalli diversi e maggiori sono più ampie (1-3 °C), ma non raggiungono mai l'apiressia completa, la febbre dicesi remittente (per es., broncopolmonite, leishmaniosi viscerale); se le defervescenze si ripetono e raggiungono l'apiressia, i periodi della quale si alternano così ai periodi di fervescenza, la forma dicesi intermittente (per es., malaria); nelle forme non complicate della malaria la febbre insorge con regolarità a giorni alterni (terzana) o ogni tre giorni (quartana) e ogni volta essa corrisponde alla liberazione in circolo di una generazione di parassiti (cioè la febbre segue il ciclo di sviluppo del parassita, il quale, a sua volta, è temporalmente diverso a seconda che si tratti di Plasmodium vivax o di Plasmodium falciparum o di Plasmodium malariae, come fu stabilito dal Golgi). Febbri ricorrenti e ondulanti sono quelle in cui a periodi (di giorni) di febbre continua o subcontinua seguono periodi di apiressia o di notevole remittenza (per es., borreliosi, brucellosi). Naturalmente molte volte si hanno decorsi febbrili anomali e irregolari per circostanze accessorie del caso in questione. Il medico utilizza la forma della febbre come elemento semeiologico per la diagnosi, ma è evidente che ciò aveva maggior valore nel passato quando mancavano mezzi di accertamento più diretti e sicuri.
Si descrive anche una ‛febbre algida': caratteristica di questa condizione è uno stato di ipotermia periferica (35-33 °C), mentre la temperatura profonda del corpo è o può essere anche molto elevata, e ciò si verifica, per esempio, nel colera (forma algida).
8. L'organismo del febbricitante
Le manifestazioni che costantemente si rilevano nel febbricitante sono la tachicardia e la tachipnea (polipnea). Di regola, ma non sempre, la tachicardia è proporzionale all'entità della febbre: nelle malattie infettive e tossinfettive gravi, la tachicardia può non essere adeguata (per es., tifo addominale, casi gravi di brucellosi).
La tachicardia, la gittata pulsatoria e quindi il volume minuto aumentano proporzionalmente all'aumento del metabolismo basale (consumo di O2), aumento che è in media del 10% per ogni grado di temperatura sopra la norma; ciò appare in accordo con l'espressione generale della legge di Van't Hoff. Ma bisogna tener presente che il miocardio, specie dei cardiopazienti e degli anziani (come del resto anche il rene), può soffrire delle elevate temperature e subire danni non indifferenti (v. Frankel e altri, 1963). Si spiega in tal modo come in questi soggetti una febbre elevata possa esser causa dell'instaurarsi di una fibrillazione atriale. Sofferenze si hanno anche nel fegato (v. Brauer e altri, 1963).
La pressione del sangue dapprima aumenta come effetto della vasocostrizione, ma arrivati al fastigio spesso diminuisce, sembra per una cospicua vasodilatazione renale (v. Pickering, 19704).
Anche i muscoli scheletrici sono certamente impegnati nella serie di eventi che accompagnano il processo febbrile: sono note l'ipotrofia e la debolezza muscolare dopo le febbri prolungate e persino le lesioni degenerative nelle febbri particolarmente elevate (v. Caldera, 1942).
Il sangue periferico del febbricitante presenta generalmente una leucocitosi; tuttavia talune febbri, come quella del tifo addominale, della brucellosi e della malaria, sono invece accompagnate da leucopenia (granulocitopenia), verosimilmente attribuibile a eccessiva azione endotossinica antileucocitaria o ad altro meccanismo depressore del midollo osseo.
Nel febbricitante è alterato non solo quantitativamente, ma anche qualitativamente il ricambio materiale; vi è soprattutto un accentuato catabolismo proteico, con bilancio negativo dell'azoto, che in parte è dovuto alla disappetenza (vi sono alterazioni della secrezione gastrica e minorata capacità digestiva) e comunque alla riduzione della dieta, ma in parte è propriamente inerente allo stato febbrile. Si ha un'orina carica di sostanze azotate, talune anche relativamente abnormi come la creatina (nell'adulto non compare normalmente nelle orine), e anche piccole quantità di albumine (vedi compromissione renale, rigonfiamento torbido dei tubuli ecc.). Aumentano nell'orina anche alcuni componenti minerali, tra i quali appare importante lo zolfo ridotto, derivante dal catabolismo delle proteine (amminoacidi solforati); diminuiscono invece i cloruri, i quali vengono ritenuti dal febbricitante (ma fanno eccezione le febbri malariche); questo fatto, insieme a una tendenziale acidosi e alle lesioni in sede muscolare, può giustificare l'insorgenza di crampi muscolari nelle febbri elevate.
All'inizio della febbre si ha anche un richiamo di acqua nei tessuti con modesta conseguente ispissatio sanguinis; questi dati, e altri più analitici, parlano per una alterata funzione delle membrane capillari e cellulari, forse mediata da modificazioni. dell'equilibrio ormonico (tiroide, corticosurreni).
9. Sostanze che possono interferire con la parte centrale della termoregolazione: gli antipiretici
Come si è già accennato (v. sopra, cap. 3) molti esperimenti intesi a far pervenire nella zona centrale della termoregolazione, o in prossimità di questa, sostanze a presumibile azione neurotropa hanno dimostrato la possibilità di interferire nel processo termoregolativo e quindi sulla febbre. I risultati di queste ricerche hanno consentito l'acquisizione di dati di notevole interesse teorico e pratico.
Anzitutto occorre tener presente che, naturalmente, solo in casi eccezionali è possibile prevedere il tipo di effetto ottenibile; prevedibili sono i casi come quello delle sostanze usate in anestesia, dalle quali è da aspettarsi, almeno come effetto immediato, un abbassamento del livello termico (quale si verifica, con queste stesse sostanze, somministrandole per via generale), abbassamento che può raggiungere anche 2,5 °C e spesso è accompagnato da vasodilatazione periferica; prevedibile è anche un rapido ritorno alla norma della temperatura (entro poche ore) e che esso sia accompagnato da brivido, fenomeno questo che indica la presenza anche di un meccanismo centrale. Meno prevedibile il fatto che talora l'animale da esperimento (per es., gatto) arrivi poi a superare il livello termico normale e divenga ipertermico.
Tali esperimenti localistici hanno inoltre consentito di constatare una diversa risposta a seconda delle zone della regione responsabile, cioè della regione preottico-ipotalamica. Analogamente agli anestetici, quali i cloralici o i barbiturici, agiscono gli oppiacei, quale la morfina. Non fa meraviglia, quindi, non solo che molti degli ipnotici abbiano anche effetto antifebbrile (antipiretico), ma nemmeno che alcuni degli antipiretici abbiano un più generale effetto analgesico e ipnotico.
Lo studio teorico e l'applicazione pratica degli antipiretici costituiscono argomenti di notevole importanza sia per la farmacologia sia per la clinica medica. Il largo impiego che di tali sostanze si fa nella pratica medica è giustificato dal fatto che, indipendentemente dalla valutazione che si voglia dare del significato biologico-fisiologico della febbre (v. sopra, cap. 6), una febbre elevata e prolungata finisce per risultare comunque dannosa: oltre alle sofferenze miocardiche e renali a cui abbiamo già accennato, va ricordata la compromissione meningea e cerebrale (delirio febbrile), onde la pratica di derivare calore dalla testa con borse di ghiaccio nei febbricitanti in pericolo. Ma in generale nelle febbri elevate e continue periodi di remittenza possono essere molto utili anche per lo stato generale del paziente, per la possibilità di meglio alimentarlo e per provvedere più agevolmente alle norme igieniche. D'altra parte, non bisogna nemmeno dimenticare che alcuni degli antipiretici possono a loro volta essere, a seconda della dose e del periodo di somministrazione, dannosi.
I medicamenti antipiretici sono numerosi: ma, a differenza di molte altre sostanze ipotermizzanti, i veri antipiretici abbassano soltanto la temperatura febbrile; essi cioè non agiscono in modo generale e generico sulla termoregolazione in quanto compromettono tante altre espressioni del controllo nervoso, bensì neutralizzano lo spostamento febbrile della temperatura di riferimento.
Nello schema qui riprodotto sono elencate le condizioni mediche capaci di interferire col processo febbrile; le più importanti di tutte, cioè quelle specifiche solo per la temperatura febbrile e ad azione centrale sono incluse nel riquadro tratteggiato, il quale comprende, appunto, i veri antipiretici.
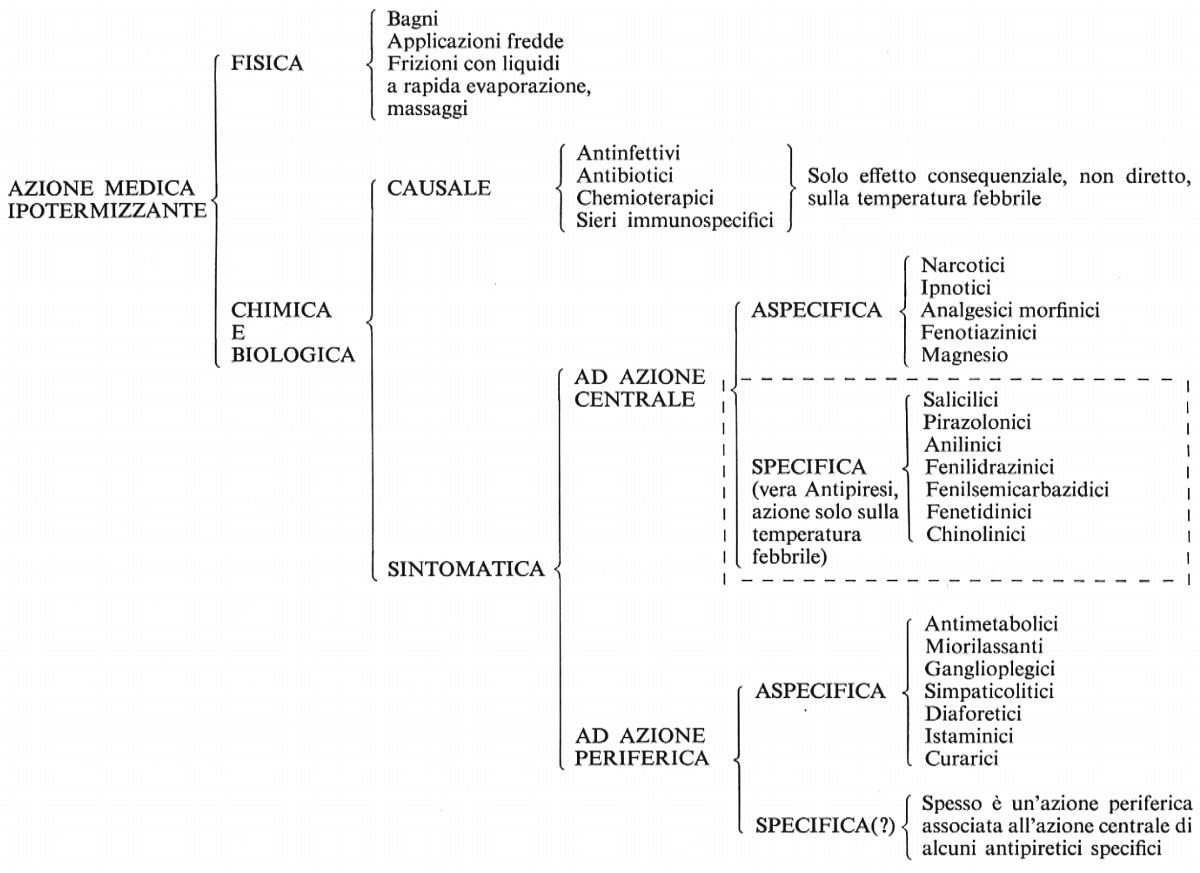
I salicilici (derivati organici dell'acido salicilico di cui il più noto è l'acido acetilsalicilico o Aspirina) corrispondono alla struttura:
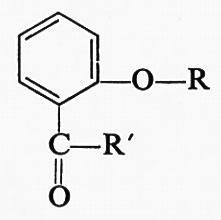
essi hanno, come è noto, anche azione analgesica e antireumatica antiflogistica, ma non sono privi di tossicità locale (stomaco) e generale (sistema nervoso, cardiocircolatorio e respiratorio, coagulazione del sangue). La loro azione è naturalmente centrale, ma si scarica sulla fase termodispersiva della termoregolazione, per vasodilatazione periferica, sudorazione e mobilizzazione di acqua dai tessuti.
I pirazolonici (derivati pirazolici) corrispondono alle strutture di base:
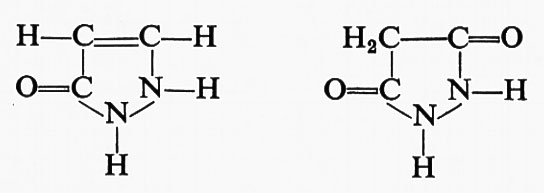
di cui esempi sono l'antipirina (2,3-dimetil-1-fenil-5-pirazolone) e la butazolidina o fenilbutazone (1,2-difenil-4-n-butil-3,5-pirazolone); il noto Piramidone o amminopirina è una dimetilamminoantipirina, ossia 2,3-dimetil-1-fenil-4-dimetilammino-5-pirazolone. Tali composti accentuano l'azione antalgica rispetto alla antipiretica e i derivati butildionici anche quella antireumatica; come antipiretici agiscono sia per accentuata termodispersione, cioè analogamente ai salicilici, sia, sembra, anche per diminuita termogenesi, probabilmente attraverso un meccanismo ormonico. Effetti secondari da evitare sono quelli sul sistema circolatorio e i fenomeni di ipersensibilità (i composti possono legarsi alle proteine corporee).
Gli anilinici, i fenilidrazinici, i fenilsemicarbazidici e i fenetidinici ‛-amminofenilici) corrispondono rispettivamente alle seguenti strutture di base nell'ordine:
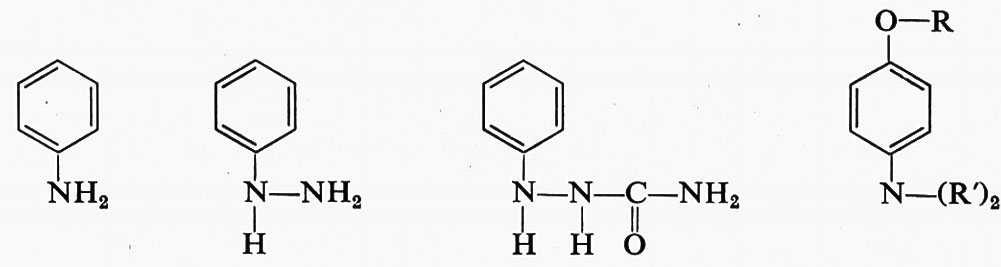
di cui esempi possono essere l'acetanilide o fenilacetammide [o-(acetilmetilammino)-fenolo], o antifebbrina; l'acido fenil-idrazon-levulinico, o antitermina; l'acetil-fenetidina o p-etossiacetanilide o fenacetina. Tali composti agiscono tutti in modo assai simile ai salicilici, e cioè prevalentemente sulla fase termodispersiva; controindicazioni sono i fatti emolitici e metaemoglobinizzanti, la possibilità di idiosincrasie.
I chinolinici rispondono alla formula generale:
di cui è noto il derivato fenolico del corrispondente acido, cioè l'acido fenil-chinolincarbonico (atofan). L'azione antipiretica di questi farmaci può non avere un vero punto di attacco centrale, ma in definitiva interferisce col metabolismo intermedio (v. l'azione sulla mobilizzazione ed escrezione dell'acido urico).
Nel complesso, appare chiaro come lo studio dell'azione antipiretica abbia una base e un'origine essenzialmente empiriche, e tali caratteristiche rimangono per tutto lo sviluppo ulteriore, anche se esso è, come si vede, molto differenziato. È assai probabile che lo stato delle cose si modificherà moltissimo quando possederemo nozioni sui determinanti molecolari della sensibilità dei centri della termoregolazione; potremo allora conoscere meglio i pirogeni endogeni e conseguentemente disporre dei corrispondenti antimetaboliti.
bibliografia
Atkins, E., Bodel, P., Francis, L., Release of an endogenous pyrogen in vitro from rabbit mononuclear cells, in ‟Journal of experimental medicine", 1967, CXXVI, pp. 357-384.
Atkins, E., Huang, W. C., Studies on the pathogenesis of fever with influenzal viruses. I-III, in ‟Journal of experimental medicine", 1958, CVII, pp. 383-435.
Atkins, E., Snell, E. S., Fever, in The inflammatory process (a cura di B. W. Zweifach, L. Grant e R. T. McClusky), London 1965, pp. 495-534.
Atkins, E., Wood, W. B., Jr., Studies on the pathogenesis of fever. II. Identification of an endogenous pyrogen in the blood stream following the injection of typhoid vaccine, in ‟Journal of experimental medicine", 1955, CII, pp. 499-516.
Beeson, P. B., Temperature-elevating effect of a substance obtained from polymorphonuclear leucocytes, in ‟Journal of clinical investigation", 1948, XXVII, p. 524.
Bennett, I. L. Jr., Beeson, P. B., Studies on the pathogenesis of fever. I-II, in ‟Journal of experimental medicine", 1953, XCVIII, pp. 477-508.
Bornstein, D. L., Brendenberg, C., Wood, W. B., Studies on the pathogenesis of fever. XI. Quantitative features of the febrile response to leucocytic pyrogen, in ‟Journal of experimental medicine", 1963, CXVII, pp. 349-364.
Bourn, J. M., Seibert, F. B., The cause of many febrile reactions following intravenous injections. II. The bacteriology of twelve distilled waters, in ‟American journal of physiology", 1925, LXXI, pp. 652-659.
Bovet, D., Virno, M., Proprietà analettiche ed ipertermizzanti della tetraido-β-naftil-etilammina (EtTHN), in ‟Rendiconti dell'Istituto Superiore di Sanità", 1952, XV, pp. 870-904.
Brauer, R. W., Balam, R. W., Bond, H. E., Carroll, H. W., Grisham, J. W., Pessotti, R. L., Reversible and irreversible changes in liver at temperatuers approaching critical upper level, in ‟Federation proceedings", 1963, XXII, pp. 724-728.
Buffa, P., Azzone, G. F., Carofoli, E., Muscatello, U., Modificazioni biochimiche mitocondriali e tessutali in relazione ad alcune ipo- ed ipertermie sperimentali del ratto, in ‟Lo sperimentale", 1960, CX, pp. 79-107.
Buffa, P., Carofoli, E., Muscatello, U., Mithocondrial biochemical lesion and pyrogenic effect of pentachlorophenol, in ‟Biochemical pharmacoloby", 1963, XII, pp. 769-778.
Caldera, M., Le alterazioni strutturali dei muscoli striati in alcune malattie generali, in ‟Archivio De Vecchi", 1942, IV, pp. 266-288.
Centanni, E., Studio sulla febbre infettiva. I. Il veleno della febbre nei Batteri, in ‟Riforma medica", 1893, IX, pp. 361-364.
Centanni, E., Untersuchungen über das Infectionsfieber: das Fiebergift der Bacterien, in ‟Deutsche Medizinische Wochenschrift", 1894, XX, pp. 148-149; 176-178.
Chatonnet, J., Nervous control of metabolism, in ‟Federation proceedings", 1963, XXII, pp. 729-731.
Collins, K. J., Weiner, J. S., Endocrinological aspects of exposure to high environmental temperatures, in ‟Physiological reviews", 1968, XLVIII, pp. 785-839.
Cooper, K. E., Mechanisms of action of pyrogens, in ‟Federation proceedings", 1963, XXII, pp. 721-723.
Cooper, K. E., Cranston, W. I., Snell, E. S., Temperature regulation during fever in man, in ‟Clinical science", 1964, XXVII, pp. 345-356.
Co Tui, Schrift, M. H., Production of pyrogen in sera by bacteria, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1939, XLII, pp. 549-552.
Co Tui, Schrift, M. H., A tentative test for pyrogen in infusion fluids, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1942, IL, pp. 320-323.
De Merre, L. J., Probey, T. F., Les pyrogènes, New York 1945.
Du Bois, E. F., The mechanism of heat loss and temperature regulation, Stanford 1937.
Frankel, H. M., Ellis, J. P., Cain, S. M., Development of tissue hypoxia during progressive hyperthermia in dogs, in ‟American journal of physiology", 1963, CCV, pp. 733-737.
Giaja, J., Dimitrijevic, I., Étude de la thermorégulation dans la fièvre, in ‟Archives intérnationales de pharmaco-dynamie et de thérapie", 1933, XLV, pp. 342-360.
Giuffrè, L., Nalli, V., Il brivido di freddo nelle febbri, in Atti del I congresso medico siciliano, Palermo 1903, pp. 3-7.
Grimby, G., Exercise in man during pyrogen-induced fever, in ‟Scandinavian journal of clinical laboratory investigation", 1962, XIV, suppl. 63, pp. 1-112.
Grosse-Brockhoff, F., Temperaturregulation, in Pathologische Physiologie, Berlin 1969, pp. 697-715.
Hall, C. H. Jr., Atkins, E., Studies on tuberculin fever. I. The mechanism of fever in tuberculin hypersensitivy, in ‟Journal of experimental medicine", 1959, CIX, pp. 339-359.
Hammel, H. T., Regulation of internal body temperature, in ‟Annual review of physiology", 1968, XXX, pp. 641-710.
Hardy, J. D., Central and peripheral factors in physiological temperature regulation, in Les concepts de Claude Bernard sur le milieu intérieur (a cura di B. Halpern), Paris 1967, pp. 247-283.
Hemingway, A., Shivering, in ‟Physiological reviews", 1963, XLIII, pp. 397-422.
Herion, J. C., Spitznagel, J. K., Walker, R. I., Zeya, H. H., Pyrogenicity of granulocyte lysosomes, in ‟American journal of physiology", 1966, CCXI, pp. 693-698.
Hort, E. C., Penfold, W. J., The dangers of saline injections, in ‟British medical journal", 1911, II, pp. 1589-1608.
Kaiser, H. K., Wood, W. B. Jr., Studies on the pathogenesis of fever IX-X, in ‟Journal of experimental medicine", 1962, CXV, pp. 27-47.
King, M. K., Wood, W. B. Jr., Studies on the pathogenesis of fever. IV. The site of action of leucocytic and circulating endogenous pyrogen, in ‟Journal of experimental medicine", 1958, CVII, pp. 291-303.
Klastersky, J., Kass, E. H., Is suppression of fever or hypothermia useful in experimental and clinical infectious-diseases?, in ‟Journal of infectious diseases", 1970, CXXI, pp. 81-86.
Krogh, A., The anatomy and physiology of capillaries, London 1922.
Liebermeister, C. von, Handbuch der Pathologie und Therapie des Fiebers, Leipzig 1875.
Mansfeld, G., Humorale Übertragbarkeit der chemischen Wärmeregulation, in ‟Archiv für experimentelle Pathologie und Pharmakologie", 1940, CXCVI, pp. 573-589.
Mansfeld, G., Weitere Untersuchungen über humorale Übertragbarkeit der chemischen Wärmeregulation, in ‟Archiv für experimentelle Pathologie und Pharmakologie", 1940, CXCVI, pp. 590-597.
Mansfeld, G., Das wirksame Prinzip der oxydationshemmenden Schilddrüsentätigkeit, in ‟Archiv für experimentelle Pathologie und Pharmakologie", 1940, CXCVI, pp. 598-608.
Mansfeld, G., Mészáros, E., Über das Nichtvorhandensein einer zentrenlosen Wärmeregulation, in ‟Archiv für experimentelle Pathologie und Pharmakologie", 1940, CXCVI, pp. 609-622.
Menkin, V., Chemical basis of fever, in ‟Science", 1944, C, pp. 337-338.
Menkin, V., Chemical basis of fever with inflammation, in ‟Archives of pathology", 1945, XXXIX, pp. 28-36.
Montuori, A., Ricerche biomediche, Napoli 1904.
Murri, A., Del potere regolatore della temperatura animale, in ‟Lo sperimentale", 1873, XXV, pp. 43-66; 117-144; 229-257.
Myers, R. D., Sharpe, L. G., Temperature in the monkey. Transmitter factors released from the brain during termoregulation, in ‟Science", 1968, CLXI, pp. 572-573.
Nordlund, J. J., Root, R. K., Wolff, S. M., Studies on the origin of human leukocytic pyrogen, in ‟Journal of experimental medicine", 1970, CXXXI, pp. 727-743.
Pickering, G. W., Fever, in General pathology (a cura di H. W. Florey), London 19704, pp. 394-412.
Remelli, L., Adattamento termoregolativo dei ratti alle forti ipertermie e suoi rapporti con la funzione testicolo-ipofisaria, in ‟La ricerca scientifica", 1955, XXV, pp. 2877-2886.
Rondoni, P., Alterazioni della regolazione termica. La febbre, in Patologia generale (a cura di F. Büchner), Torino 1954, pp. 665-679.
Seibert, F. B., The cause of many febrile reactions following intravenous injections, in ‟American journal of physiology", 1925, LXXI, pp. 621-651.
Seibert, F. B., Mendel, L. B., Protein fevers, in ‟American journal of physiology", 1923, LXVII, pp. 105-123.
Smith, R. E., Horwitz, B. A., Brown fat and thermogenesis, in ‟Physiological reviews", 1969, IL, pp. 330-425.
Snell, E. S., Atkins, E., The presence of endogenous pyrogen in normal rabbit tissues, in ‟Journal of experimental medicine", 1965, CXXI, pp. 1019-1038.
Snell, E. S., Atkins, E., The mechanism of fever, in The biological basis of medicine (a cura di E. E. Bittar e N. Bittar), vol. II, London 1968, pp. 397-419.
Spinelli, A., Contributo allo studio delle ossidazioni organiche nella febbre. Le ossidoriduzioni (deidrogenazioni) e i fermenti ossidasici dei tessuti nei processi febbrili, in ‟Minerva medica", 1932, XXIII, pp. 201-207.
Spinelli, A., Osservazioni intorno alla influenza del lavoro muscolare e del massaggio sull'andamento della ipertermia sperimentale del coniglio, in ‟Archivio di fisiologia", 1934, XXXIII, pp. 519-533.
Thauer, R., Der Mechanismus der Wärmeregulation, in ‟Die Ergebnisse der Physiologie", 1939, XLI, pp. 607-805.
Thauer, R., Circulatory adjustment to climatic requirements, in Handbook of physiology, vol. III, Circulation (a cura di W. F. Hamilton e P. Dow), Baltimore 1965, pp. 1921-1966.
Thauer, R., Homoiothermie als Fortschritt und Schicksal des Menschen, in Jahrbuch der Max Plank-Gesellschaft zur Förderung der Wissenschaften, München 1967, pp. 39-75.
Trabucchi, E., Fisiologia e farmacologia della termoregolazione, in ‟Società Italiana per il Progresso delle Scienze", 1940, XXVIII, pp. 85-117.
Vernoni, G., La respirazione sopravitale dei tessuti nella febbre, in ‟Archivio per le scienze mediche", 1916, XL, pp. 308-348.
Vernoni, G., Il comportamento delle ossidasi (indofenolossidasi) dei tessuti nella febbre, in ‟Bollettino di scienze mediche", 1917, LXXXVIII, p. 1.
Vernoni, G., Sulla patogenesi della febbre, in ‟Lo sperimentale", 1940, XCIV, pp. 3-35.
Vernoni, G., Nuovi fatti e nuove idee sulla febbre, in ‟Recenti progressi in medicina", 1948, V, pp. 289-320.
Vernoni, G., Trattato di patologia generale, vol. I, Firenze 1954.
Vernoni, G., Pirone-Naconetschnii, A., Piroterapia ed indice opsonico del sangue, in ‟Minerva medica", 1938, II, pp. 386-389.
Woodin, A. M., The basis of leucocidin action, in The biological basis of medicine (a cura di E. E. Bittar e N. Bittar), vol. II, London 1968, pp. 372-396.