
Flusso energetico e produzione primaria
Flusso energetico e produzione primaria
In questo saggio analizzeremo il modo in cui le risorse della biosfera e le trasformazioni energetiche che in essa avvengono sono in grado di influenzare le nostre condizioni di vita. Cominceremo col descrivere i processi che regolano il ciclo biogeochimico dell'acqua e i flussi energetici che si verificano tra la biosfera e l'atmosfera, soffermandoci in particolare sui fattori che controllano i meccanismi di dissipazione dell'energia da parte della vegetazione. Come esempio dell'influenza esercitata dagli organismi vegetali sul clima discuteremo l'impatto della deforestazione delle foreste tropicali umide dell'Amazzonia e delle foreste boreali della Siberia. Successivamente analizzeremo l'importanza della produttività primaria come elemento fondamentale dell'organizzazione trofica del pianeta e descriveremo il ciclo del carbonio a livello planetario, sottolineando il ruolo degli ecosistemi nella regolazione degli equilibri climatici del pianeta.
Introduzione
La vegetazione terrestre, costituita per il 40% da foreste, ricopre circa il 23% della superficie complessiva del nostro pianeta. Se fosse distribuita in un unico strato sottile di superficie foto sintetizzante, la superficie fogliare che forma il mantello verde delle nostre coperture vegetali sarebbe in grado di coprire circa 2,5 volte l'intera superficie del globo.
Quando pensiamo all'importanza della vegetazione in rapporto alla nostra vita o a quella di altri organismi viventi, in genere ci riferiamo alla possibilità di utilizzare la sostanza organica vegetale come materiale di consumo alimentare necessario per lo svolgimento delle funzioni vitali, oppure pensiamo ai prodotti estraibili che possono servire a migliorare le nostre condizioni di vita, come il legno per costruzione, la pasta di cellulosa, gli oli e le fibre vegetali per alcune applicazioni industriali. Raramente ci soffermiamo a pensare all'importante ruolo che gli ecosistemi terrestri hanno nella regolazione delle condizioni della nostra vita, per esempio nell'influenzare le caratteristiche dell'atmosfera, il clima, il ciclo dell'acqua, l'erosione e altri processi che, in modo meno diretto ma estremamente significativo, condizionano e rendono possibile la vita sul nostro pianeta (Valentini et al., 1998a).
Gli ecosistemi terrestri, attraverso i processi di fotosintesi e traspirazione, influenzano i flussi energetici superficiali e modificano la composizione chimica dell'atmosfera. Circa 4,5 miliardi di anni fa, all'origine del nostro pianeta, i fattori fisici e chimici dominavano la distribuzione degli elementi nel suolo e la composizione delle acque e dell'atmosfera. Da quando per la prima volta la vita comparve sulla Terra, circa 3,5 miliardi di anni fa, i processi biologici sono diventati sempre più importanti nel determinare la distribuzione degli elementi e nell'influenzare le caratteristiche chimicofisiche del pianeta.
Con la pubblicazione nel 1979 di Gaia, J.E. Lovelock portò all'attenzione della comunità scientifica il fatto che la composizione chimica dell'atmosfera attuale è estremamente 'anomala', diversa da quella che ci si attenderebbe all'equilibrio termodinamico (Lovelock, 1979). Se non si fosse sviluppata la vita sulla Terra la concentrazione di diossido di carbonio nell'atmosfera sarebbe rimasta molto elevata e la temperatura superficiale del pianeta sarebbe molto differente, probabilmente più alta dell'attuale. La concentrazione di ossigeno sarebbe cresciuta lentamente nel tempo a causa della dissociazione dell'acqua per effetto della radiazione solare, ma non sarebbe arrivata che all'1% della concentrazione attuale. Se oggi scomparisse la vita sulla Terra, si verificherebbero l'arresto dei processi di denitrificazione, che causerebbe un aumento del livello dei nitrati, e una riduzione della produzione di ammoniaca, che è quasi esclusivamente di origine biologica. L'ammoniaca regola il pH delle piogge, mantenendo lo a un valore adeguato, pari a circa 6; in sua assenza, questo valore si ridurrebbe a 3 (un'acidità simile a quella dell'aceto!) e ciò avrebbe un impatto notevole sugli oceani.

Nella tabella (tab. I) sono riportate le abbondanze di alcuni gas neII'attuale atmosfera terrestre, che contiene circa il 20% di ossigeno, insieme alle ipotetiche concentrazioni all'equilibrio. Si può notare come, a causa dell'abbondanza di ossigeno, la composizione attuale dell'atmosfera non potrebbe rimanere costante se non vi fossero flussi biogenici in grado di mantenerla. Vi sono infatti oggi nella nostra atmosfera troppo azoto, metano (più di 29 ordini di grandezza rispetto alla concentrazione di equilibrio), ossido nitroso, ammoniaca e idrogeno. Almeno per altri 40 gas di origine biogenica esiste il problema di spiegare la loro enorme concentrazione atmosferica rispetto alla quantità di ossigeno presente (Levine, 1989). Poiché queste sostanze sono soggette continuamente alla trasformazione da parte dell'ossigeno, la loro abbondanza può essere spiegata soltanto dall'esistenza di flussi costanti e ingenti provenienti dalla biosfera.
La quantità di energia disponibile sulla superficie terrestre è il risultato del bilancio tra l'energia proveniente dal Sole e quella riemessa per irraggiamento. La quantità di energia in ingresso è funzione della riflettività (albe do) della superficie terrestre. Il ghiaccio riflette l'80÷95% della radiazione incidente, una prateria il 30÷40% e una foresta di conifere il 10÷15%. Di conseguenza, le variazioni stagionali nella vegetazione, così come l'impatto delle attività umane, possono provocare sostanziali cambiamenti nella quantità di radiazione assorbita dalla superficie, alterando sensibilmente la temperatura superficiale. È stato dimostrato in molti studi che il bilancio energetico superficiale ha importanti conseguenze sul regime climatico regionale e globale.
Sono numerose, oggi, le simulazioni del clima globale che tengono conto della vegetazione terrestre e delle sue proprietà in termini di flussi energetici superficiali (Sellers et al., 1996). Alcuni esempi dell'importanza degli scambi energetici superficiali sono stati riportati in uno studio di Y. Xue e J. Shukla (1993) con il quale è stato mostrato come la deforestazione e i cambiamenti di uso del suolo nelle regioni del Mediterraneo e del Sahel possano portare a una riduzione delle precipitazioni e a un conseguente aumento dell'aridità e dei processi di desertificazione.
In questo saggio discuteremo il modo in cui gli ecosistemi terrestri, attraverso le trasformazioni dell'energia e i processi di sintesi della sostanza organica, possono esercitare un impatto significativo sul clima del nostro pianeta sia a livello regionale sia a livello globale.
Una definizione operativa di biosfera
La biosfera è la parte del nostro pianeta dove esistono le condizioni indispensabili alla vita animale e vegetale e dove gli organismi viventi, essi stessi parte della biosfera, scambiano materia ed energia. La biosfera comprende l'atmosfera, la superficie terrestre, dove risiedono le piante, la rizosfera, dove dominano le radici e i microrganismi, i corpi idrici liberi quali fiumi, laghi e oceani. Nel caso degli ecosistemi acquatici il ciclo degli elementi e la natura degli organismi coinvolti sono così differenti che si usa suddividere la biosfera in geobiosfera, che include gli ecosistemi terrestri, e idrobiosfera, che racchiude gli ecosistemi acquatici (Walter, 1973).
Le trasformazioni di energia a livello di ecosistema
La circolazione dell'acqua attraverso le piante e gli ecosisterni rappresenta il più grande trasferimento di energia mediante una sostanza chimica nella biosfera. Le piante sono in grado di assimilare acqua grazie all'azione di assorbimento radicale e di trasportarla lungo il fusto fino alle foglie da dove, attraverso gli storni (i minuscoli pori sulla superficie fogliare), viene restituita all'atmosfera sotto forma di vapore acqueo. La quantità di acqua che le piante possono estrarre dal suolo è davvero notevole. Una foglia può traspirare una quantità di acqua pari a circa 200 mI al giorno, un albero circa 200 l e una foresta di un ettaro 10.000 l. Il trasporto idrico e la successiva evaporazione sono processi termodinamici che richiedono ingenti quantità di energia: si consideri che per l'evaporazione di 1 g di acqua sono necessari 2440 J. Gli ecosistemi terrestri possono essere considerati gigantesche e silenziose macchine termodinamiche capaci di trasformare, giorno dopo giorno, rilevanti quantitativi di energia.
L'energia che muove tutto ciò è sempre quella solare, che viene assorbita dalle piante e utilizzata in vari processi, tra cui quello traspirativo e quello fotosintetico. Il modo in cui viene trasformata questa energia è peculiare di ciascun ecosistema terrestre e varia a seconda della sua struttura, della sua composizione specifica e della sua funzionalità.
Poiché la maggior parte dell'energia assorbita dagli ecosistemi viene utilizzata nei processi di traspirazione ed evaporazione, cominceremo con l'esaminare lo schema generale del ciclo dell'acqua su scala globale, e successivamente osserveremo come la dissipazione di energia possa determinare una serie di proprietà funzionali a livello di ecosistema. Descriveremo infine alcuni casi in cui l'azione della vegetazione terrestre influenza il clima su scala regionale.
Il ciclo globale dell'acqua
Attraverso la traspirazione, l'evaporazione e le precipitazioni (oltre che mediante le correnti oceaniche) l'acqua trasferisce la maggior parte dell'energia ricevuta dal Sole dalle regioni tropicali a quelle polari, quasi come una conduttura che trasferisce il calore da una caldaia ai vari locali di un'abitazione.
l movimenti dell'acqua nell'atmosfera determinano la distribuzione delle precipitazioni sulla Terra e la disponibilità annuale di risorse idriche e, di conseguenza, la crescita e lo sviluppo delle piante e, più in generale, le condizioni favorevoli alla vita degli organismi. Se le precipitazioni eccedono l'evaporazione e la traspirazione si può produrre lo scorrimento (o deflusso) superficiale che determina l'erosione e il trasporto di materiale solido verso gli oceani.
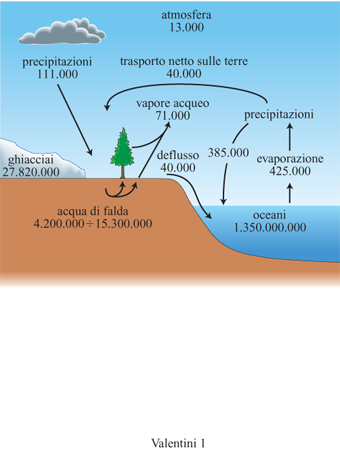
Le quantità di acqua coinvolte nel ciclo globale idrologico (fig. 1) sono tradizionalmente espresse in unità di km³. Gli oceani, come si può facilmente intuire, sono i serbatoi di acqua più grandi del nostro pianeta: ne contengono 1.350.000.000 km³, circa il 97% di tutta quella presente sulla superficie terrestre. l ghiacci polari e i ghiacciai continentali, con 27.820.000 km³, forniscono il secondo contributo al nostro patrimonio di acqua. Il suolo contiene circa 121.800 km³ di acqua, dei quali circa 58.100 km³ si trovano nella zona interessata dalle radici delle piante (Webb et al., 1993). l fabbisogni umani sono soddisfatti da una quantità relativamente ridotta di acqua, contenuta nei laghi e nei fiumi. L'acqua presente nelle falde sotterranee non è facilmente valutabile, ma recenti stime mostrano che essa varia globalmente da 4.200.000 a 15.300.000 km³; a eccezione della piccola parte estratta dall'uomo, in genere quest'acqua non è disponibile per i processi vitali della biosfera.
La quantità di acqua presente nell'atmosfera è relativamente ridotta, circa 13.000 km³, tuttavia grandi quantità di acqua si muovono da e verso l'atmosfera e all'interno di essa come risultato degli scambi energetici superficiali. L'evaporazione rimuove dagli oceani circa 425.000 km³ di acqua ogni anno. Da questo dato si può dedurre che il tempo medio di residenza dell'acqua negli oceani è pari a circa 3200 anni. Soltanto una parte di quest'acqua, circa 385.000 km³/anno, ritorna agli oceani con le precipitazioni; il resto, pari a 40.000 km³/anno, viene trasferito come vapore acqueo sulle terre. Le piante sono in grado di assorbire acqua tramite l'apparato radicale e di restituirla all'atmosfera sotto forma di vapore: questo processo va sotto il nome di traspirazione. L'insieme della traspirazione e dell'evaporazione che inevitabilmente vi si accompagna in presenza di superfici bagnate viene definito evapotraspirazione, termine utilizzato per descrivere il flusso di acqua dalle superfici vegetali all'atmosfera.
Lo spostamento di acqua dovuto all'evapotraspirazione delle terre ammonta a circa 71.000 km³ l'anno, che insieme ai 40.000 km³/anno provenienti dagli oceani determinano una precipitazione sulle terre di 111.000 km³/anno. Tale valore testimonia la grande importanza della vegetazione terrestre nel riciclare l'acqua dell'atmosfera. Il tempo medio di residenza nell'atmosfera dell'acqua proveniente dall'evapotraspirazione è circa l anno, assai più breve di quello dell'acqua negli oceani. Questi dati evidenziano come gli ecosistemi terrestri giochino un ruolo determinante nel ciclo dell'acqua e nelle trasformazioni energetiche a esso associate.
Il bilancio energetico superficiale
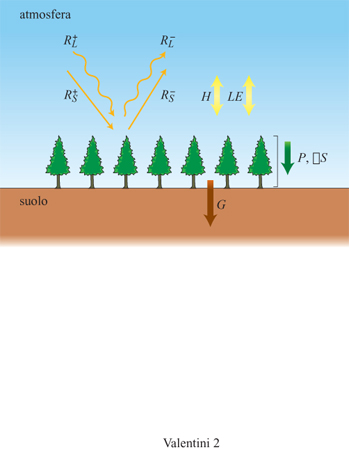
Se consideriamo una superficie ideale relativamente uniforme e omogenea e sufficientemente estesa, separata dall'atmosfera sovrastante mediante una sottile interfaccia e posta immediatamente sopra al suolo, il calcolo del bilancio energetico risulta semplificato in quanto devono essere considerati solo i flussi verticali d'energia. Ci sono essenzialmente quattro tipi di flussi d'energia per una superficie vegetale ideale: il flusso radiativo netto (RN) da e verso la superficie, il flusso di calore sensibile (H) e latente (LE) da e verso l'atmosfera e il flusso di calore (G) da e verso il suolo (fig. 2).
Il flusso radiativo netto (RN) è il risultato del bilancio radiativo nelle bande spettrali del visibile (Rs) e dell'infrarosso (RL). Se separiamo la radiazione incidente verso la copertura e quella uscente dalla superficie possiamo scrivere:
formula. [
l]
Per convenzione, i flussi diretti verso la superficie sono considerati positivi, quelli diretti verso l'atmosfera negativi. La radiazione solare visibile che giunge sulla superficie è costituita in parte dalla radiazione solare diretta, in parte da quella diffusa. La componente che esce dalla superficie è la frazione di radiazione incidente che viene riflessa dalla copertura vegetale:
formula, [2]
dove A è l'albedo della superficie, il cui valore dipende dalle proprietà di questa. Alti valori di albedo si osservano per le coperture nevose, per i suoli nudi privi di vegetazione, ecc., mentre per le coperture vegetali tale grandezza varia tra 0,1 e 0,3, indicando un grado significativo di assorbimento della radiazione visibile. In assenza di nuvole, la radiazione incidente a elevata lunghezza d'onda, appartenente alla regione infrarossa dello spettro (RL dipende principalmente dalla temperatura, dall'umidità e dalla concentrazione dei gas serra nell'atmosfera, e non mostra variazioni rilevanti durante le ore diurne. La presenza di nuvole aumenta significativamente la quantità di radiazione infrarossa che raggiunge la superficie. La radiazione infrarossa uscente dipende dalla temperatura della superficie secondo la legge di Stefan-Boltzmann:
formula, [3]
dove s è l'emissività della superficie (tipicamente 0,90÷0,98 per la vegetazione), σ è la costante di Stefan-Boltzmann (5,67 10-⁸m -2K-¹) e T è la temperatura assoluta.
Poiché la radiazione infrarossa uscente dalla superficie è proporzionale alla quarta potenza della temperatura, essa mostra significative variazioni diurne, con valori massimi nel primo pomeriggio e minimi all'alba. Frequentemente nelle prime ore della notte la temperatura della superficie vegetale è ancora elevata, mentre la radiazione infrarossa proveniente dal cielo è ridotta, essendo presente solo quella di corpo nero delle masse d'aria; ciò fa sì che il flusso radiativo netto diventi negativo.
La radiazione netta può essere calcolata misurandone le singole componenti, oppure mediante appositi strumenti, chiamati radiometri netti, costituiti da due sensori che misurano simultaneamente, con una risposta spettrale piuttosto ampia (0,4÷50 μm), la radiazione incidente e quella uscente, e forniscono automaticamente la differenza tra le due, cioè il flusso radiativo netto RN.
Il flusso energetico di calore sensibile (H) è la quantità di calore che viene trasportata per via convettiva e turbolenta tra la superficie e l'atmosfera, a causa del gradiente di temperatura esistente tra esse:
formula, [4]
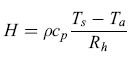
Rh dove ρ e cp sono rispettivamente la densità e il calore specifico a pressione costante dell'aria secca, Tg e Ta le temperature della superficie vegetale e dell'aria circostante in un punto di riferimento al di sopra della copertura. Rh è la resistenza termica (cioè l'inverso del coefficiente di trasporto del calore), dipendente dalla velocità del vento e dalle caratteristiche aerodinamiche della superficie. Il flusso di calore sensibile dipende quindi dalle sole caratteristiche fisiche del sistema, in particolare dalla temperatura superficiale e dal vento. Le caratteristiche delle strutture vegetali (rami, foglie, altezza) giocano anch'esse un ruolo importante nella dissipazione dell'energia. Foglie di dimensioni ridotte, aghiformi o lanceolate favoriscono la dissipazione del calore in quanto aerodinamicamente efficienti. Per questo motivo negli ambienti aridi la maggior parte delle forme di vita vegetali ha sviluppato adattamenti fogliari e strutturali di questo tipo, al fine di aumentare la dissipazione del calore.
In modo analogo il flusso energetico di calore latente di evaporazione (LE), che rappresenta l'energia spesa nei processi evapotraspirativi, si può scrivere:
formula, [5]
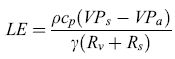
ove γ è la costante psicrometrica, VPs e VP a le pressioni parziali di vapore acque o rispettivamente all'interno delle foglie (più precisamente nelle cavità substomatali) e in un punto nell'atmosfera al di sopra della copertura, Rv la resistenza al trasporto turbolento del vapore acqueo e Rs la resistenza al trasporto attraverso gli storni (Monteith, 1975). È interessante notare che Rv è un parametro di tipo fisico, analogo a Rh, che dipende dalle caratteristiche aerodinamiche della superficie e dalla velocità del vento, mentre Rs è un parametro di natura biologica, correlato all'apertura degli storni, la cui dimensione è regolata dal metabolismo della vegetazione.
Il flusso energetico da e verso il suolo (G) può essere stimato come
formula, [6]
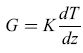
dove K è la conducibilità termica del suolo e dT/dz, il gradiente termico verticale nel suolo. Il flusso di calore nel suolo dipende largamente dalle condizioni di irraggiamento della sua superficie. Nel caso di suolo privo di copertura vegetale il flusso di calore che entra nel suolo può rappresentare anche il 50% di tutto il flusso radiativo netto, mentre se il suolo è protetto da una densa copertura vegetale, come quella di una foresta, tale valore si riduce a circa il 2÷5% del flusso radiativo netto.
Di giorno il suolo assorbe calore e il flusso è quindi positivo, mentre nelle prime ore della notte il suolo tende a cedere calore rapidamente, mostrando un flusso negativo. Una superficie ideale, orizzontalmente omogenea e uniforme, raramente si incontra in pratica. In particolare l'ipotesi di interfaccia sottile tra atmosfera e superficie è puramente ideale, in quanto le coperture vegetali sono costituite da spessi strati fogliari. A tale proposito basti considerare le foreste, i cui alberi possono raggiungere dimensioni di oltre 50 m e quindi inglobare tra suolo e atmosfera sovrastante un ampio volume di spazio, all'interno del quale si svolgono altri processi energetici come l'immagazzinamento del calore e i processi biochimici.
l flussi energetici associati con la presenza di un volume finito (∆SB, ∆ST, ∆SE) possono essere ricondotti ai seguenti tre processi che avvengono all'interno del volume considerato: il riscaldamento della biomassa presente nel volume, il riscaldamento dell'aria e le variazioni della concentrazione di vapore acque o nell'atmosfera interna al volume. L'entità di tali flussi energetici dipende dalla densità, dal calore specifico e dalla temperatura della biomassa presente nel volume.
Un ulteriore termine che contribuisce al bilancio energetico è il flusso di energia biochimica (P) che viene utilizzata all'interno del volume di riferimento. l processi fotosintetici utilizzano la radiazione solare per produrre sostanza organica, associando al flusso di energia la sintesi biologica. Viceversa, i processi respiratori rappresentano un consumo di energia. Tuttavia, nonostante tali flussi energetici siano di estrema importanza per la vita degli organismi biologici e dell'ecosistema nel suo insieme, la quantità di energia che viene utilizzata in questi processi è trascurabile rispetto a quella disponibile per l'ecosistema. Essa infatti ammonta a circa il 2% (eccezionalmente, per colture agrarie ad alta produttività, può arrivare all'8%) di tutta l'energia disponibile.
In sintesi, tenendo presente il principio di conservazione dell'energia, si può scrivere il bilancio riferito a una superficie vegetale (v. figura 2) mediante l'espressione:
RN +H +LE + G + ∆ST + ∆SB + ∆SE + P = 0. [7]
Dalle considerazioni sopra esposte, i termini che riguardano i flussi energetici associati con le reazioni biochimiche sono trascurabili. Se, in prima approssimazione, si considerano non importanti i termini riguardanti l'energia immagazzinata all'interno del volume dell'ecosistema, il bilancio si riduce a quello di una superficie ideale:
RN + H + LE + G = 0, [8]
da cui, ponendo A uguale a RN + G, si ricava:
A = − (H + LE). [9]
In questa relazione A è il flusso di energia effettivamente disponibile a livello di ecosistema, che viene impegnato nei due processi principali: l'evapotraspirazione e il trasporto di calore sensibile.
Meccanismi di dissipazione dell'energia
Esaminando la forma semplificata dell'equazione del bilancio energetico ci possiamo porre il problema di come gli ecosistemi terrestri usino l'energia, di come la dissipino nei vari processi e, quindi, di quali siano le loro proprietà funzionali. Tradizionalmente lo studio del bilancio energetico è stato affrontato per ricavare informazioni di tipo pratico sui consumi idrici di colture agrarie. Infatti, conoscendo l'energia spesa nel processo di evapotraspirazione, si potevano calcolare i fabbisogni idrici delle colture e quindi regolare in modo più efficiente le irrigazioni. In tempi più recenti numerosi studi, estesi anche alla vegetazione naturale e alle foreste, hanno mostrato che il modo con cui gli ecosistemi terrestri ripartiscono l'energia solare assorbita è una proprietà funzionale caratteristica che varia a seconda del tipo di copertura vegetale.
P.G. Jarvis e K.G. McNaughton (1986) furono tra i primi a osservare che le modalità di dissipazione dell'energia sono sotto il controllo biologico e strutturale da parte della vegetazione, introducendo il concetto di accoppiamento atmosferico, espresso mediante il cosiddetto fattore Omega (v. oltre). Un modo per esprimere l'evapotraspirazione di una copertura vegetale in funzione dei vari parametri biologici e ambientali è l'equazione di Penmann-Monteith (Monteith, 1975):
formula, [10]
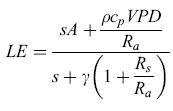
dove A è il flusso di energia disponibile nell'ecosistema, p e cp sono rispettivamente la densità dell'aria secca e il suo calore specifico, VPD è il deficit di pressione parziale di vapore a un'altezza di riferimento sopra la copertura, s è la pendenza della curva della pressione di vapor saturo in funzione della temperatura, γ la costante psicrometrica, Rs e Ra rispettivamente la resistenza stomatica e quella aerodinamica. Quest'ultima, per il principio della similarità del trasporto turbolento di massa ed energia, è la stessa per il trasporto sia del calore sensibile che del vapore acqueo (Ra = Rh = Rv). La resistenza aerodinamica è una funzione della velocità del vento, della stabilità atmosferica e delle proprietà strutturali della vegetazione, quali altezza, area fogliare, irregolarità della copertura. In particolare, per coperture vegetali di bassa statura e relativamente regolari e omogenee (come, per esempio, le colture agrarie o le praterie) Ra assume un valore elevato, indicando una bassa efficienza aerodinamica per il trasporto di energia. Al contrario, la vegetazione di elevata statura e irregolare (come possono essere le foreste) mostra valori di Ra più bassi, che denotano una maggiore efficienza aerodinamica e quindi un più agevole trasporto di energia.
Vi sono due limiti dell'equazione di Penmann-Monteith che rappresentano le due situazioni descritte:
formula [11]
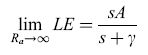
formula. [12]
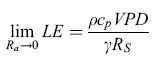
Il primo caso è quello di una copertura vegetale con resistenza aerodinamica molto alta. In queste condizioni l'evapotraspirazione dipende solo dall'energia disponibile, indipendentemente dal controllo biologico effettuato da parte degli storni, e il valor limite di LE rappresenta l'evapotraspirazione di equilibrio, che è il valore massimo in rapporto all'energia disponibile. Tale limite può essere superato solo se vi sono input di energia esterni.
Nel secondo caso l'evapotraspirazione è più direttamente controllata dal deficit di pressione parziale di vapore e dagli storni. In questo caso la vegetazione può controllare più efficacemente l'evapotraspirazione, regolando l'apertura degli storni.
Il primo caso rappresenta una vegetazione bassa e omogenea, come le colture agrarie, la cui evapotraspirazione è maggiormente dipendente dall'energia disponibile. Secondo la terminologia di Jarvis-McNaughton, queste vegetazioni sono 'disaccoppiate' dall'atmosfera, e il loro fattore Omega è prossimo a 1. Al contrario, le coperture forestali sono maggiormente dipendenti dal deficit di pressione di vapore nell'atmosfera e dalle condizioni di apertura stomatica. Esse sono cioè 'accoppiate' all'atmosfera, e il loro fattore Omega è prossimo a 0. Questo sta a dimostrare che nel primo caso l'energia è dissipata prevalentemente sotto forma di evapotraspirazione (quando vi sia sufficiente disponibilità idrica nel suolo), mentre nel secondo caso il controllo biologico esercitato dalle piante riduce la dissipazione dell'energia sotto forma di evapotraspirazione a vantaggio del calore sensibile.
È possibile analizzare la sensibilità del processo di evapotraspirazione al controllo biologico, esprimendo la variazione percentuale dell'evapotraspirazione in rapporto alla variazione della resistenza stomatica. Derivando l'equazione [10] si ottiene:
formula. [13]
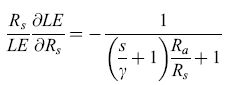
Il segno negativo indica una retro azione negativa, cioè per ogni incremento della resistenza stomatica (chiusura degli storni) vi è una diminuzione dell'evapotraspirazione. Aumentando Rs aumenta la sensibilità dell'evapotraspirazione alla chiusura stomatica. Nella figura (fig. 3) vengono confrontate tre differenti coperture vegetali (grano, foresta di latifoglie decidue, foresta di conifere) usando come valori di Ra rispettivamente 80, 15 e 5 s m-1.
l valori più negativi della sensibilità indicano un maggiore controllo biologico sull'evapotraspirazione. Come atteso, la sensibilità dell'evapotraspirazione a cambiamenti di Rs risulta più grande per le foreste e in particolare per quelle di conifere. È chiaro quindi che sia la struttura della vegetazione (espressa attraverso il parametro di efficienza aerodinamica) che il controllo biologico (espresso dalla resistenza stomatica) determinano il controllo sull'evapotraspirazione e quindi sulla dissipazione di energia.
È interessante notare come molti studi riguardanti i rapporti tra biosfera e atmosfera tendano a indicare una convergenza funzionale di vari ecosistemi terrestri nell'amplificare le loro proprietà di dissipazione dell'energia. Per esempio le foreste, e in generale la vegetazione naturale, che possiedono la più elevata efficienza aerodinamica e sono quindi più accoppiate all'atmosfera, presentano anche i valori di resistenza stomatica più elevati, che accrescono ulteriormente la sensibilità dell'evapotraspirazione al controllo biologico. Il contrario risulta per la vegetazione agraria o per le praterie, dove i valori più bassi di resistenza stomatica amplificano ancora di più il loro disaccoppiamento dall'atmosfera.
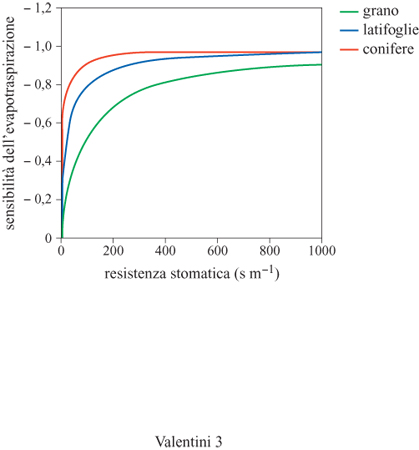
In figura (fig. 4) è riportato l'andamento del rapporto tra l'evapotraspirazione e il suo valore di equilibrio (equazione [11]) in funzione della resistenza stomatica per una serie di ecosistemi terrestri. Si può notare come gli ecosistemi che presentano un forte controllo stomatico, cioè un elevato valore di Rs dissipano energia per evapotraspirazione in quantità molto inferiore alle condizioni di equilibrio. Ciò sta a significare che la maggior parte dell'energia assorbita è dissipata in calore sensibile, cioè attraverso uno scambio fisico con l'atmosfera circostante, con conseguente impatto sul riscaldamento di quest'ultima.
Viceversa, a mano a mano che si passa a ecosistemi con un controllo stomatico meno intenso, cioè in cui gli storni sono più aperti, l'energia dissipata per evapotraspirazione tende al valore di equilibrio, che dipende solo dalla quantità di energia disponibile fornita dal Sole. Quando la maggior parte dell'energia è dissipata sotto forma di evapotraspirazione, la superficie risulta più fredda e si ha un minore impatto sul riscaldamento dell'atmosfera circostante.
Prima di esaminare l'effetto della diversità di dissipazione dell'energia sul clima regionale, è interessante porsi alcune domande: come mai la capacità di dissipazione di energia sotto forma di evapotraspirazione varia così tanto negli ecosistemi terrestri? Perché le foreste controllano l'evapotraspirazione più di altri tipi di vegetazione, per esempio delle praterie? E in particolare, perché tra le foreste quelle di conifere sono più sensibili al controllo biologico rispetto a quelle di latifoglie?
Una possibile spiegazione è che le foreste e la vegetazione naturale di solito colonizzano ambienti più severi, dove le disponibilità idriche sono scarse, per lo più variabili e altamente imprevedibili. In queste condizioni è necessario un forte controllo biologico dell'evapotraspirazione, in modo da controllare le perdite di acqua nei periodi più sfavorevoli e favorire la ricarica delle riserve in quelli più favorevoli. Inoltre, le piante sempreverdi devono necessariamente regolare i flussi idrici mediante controllo stomatico, in quanto si trovano a vegetare in condizioni altamente variabili nel corso dell'anno, mentre la vegetazione decidua ha a disposizione un periodo più breve per i processi vegetativi e quindi non può limitare troppo l'apertura stomatica. Queste differenze si riflettono nelle diverse strategie osservate in natura tra specie consumatrici di acqua (water spenders) e specie conservatrici di acqua (water savers). È interessante notare come le proprietà di dissipazione dell'energia, osservate a livello di specie e di singoli organi fogliari, si riflettano anche a livello di ecosistema nella sua totalità, portando a confermare l'esistenza di proprietà funzionali a questo livello nei rapporti biosfera-atmosfera.
Implicazioni climatiche su scala regionale
Alla luce di quanto abbiamo descritto, quando osserviamo una determinata porzione della superficie terrestre in termini di utilizzazione dell'energia, ci troviamo di fronte a un mosaico di diverse porzioni di territorio che mostrano altrettante modalità di scambio energetico con l'atmosfera circostante. In questo paragrafo analizzeremo il modo in cui la dissipazione dell'energia al livello di superficie terrestre, e quindi a livello di ecosistema, può influenzare il clima regionale ed eventualmente quello globale.
Nel passato, come si è detto, la visione delle interazioni tra biosfera e atmosfera era di tipo unidirezionale, cioè veniva presa in considerazione l'influenza del clima sulla vita degli organismi biologici, ma non veniva contemplata la possibilità che anche questi ultimi potessero giocare a loro volta un ruolo importante sul clima. Recentemente l'influenza delle caratteristiche della superficie terrestre e delle proprietà funzionali degli ecosistemi che la compongono è stata riconosciuta importante per i processi climatici. Oggi sono sempre più diffusi i modelli di circolazione globale che includono le caratteristiche della superficie e gli scambi energetici a livello di ecosistema nella simulazione del clima del pianeta (Sellers et al., 1996). Ulteriori studi hanno mostrato l'esistenza di importanti effetti dei cambiamenti del suolo sul clima regionale (Pielke et al., 1991) e l'esistenza di sistemi in cui l'impatto della vegetazione sul clima regionale è così importante da determinare le condizioni di vita dell'intero ecosistema.
Un caso di retroazione tra vegetazione e clima regionale ampiamente studiato è quello del bacino amazzonico (Schukla et al., 1990), il quale è particolarmente noto per le attività di deforestazione che ogni anno lo investono e per l'impatto che queste hanno sul bilancio del carbonio planetario (v. oltre) e sulla biodiversità. Tuttavia un aspetto poco noto, ma estremamente importante per la vita dell'ecosistema amazzonico, è il danno che la deforestazione sta arrecando al bilancio idrologico della regione. Il sistema amazzonico è un sistema chiuso dal punto di vista idrologico. Circa il 70% delle precipitazioni è costituito dal vapore acqueo evapotraspirato dalla vegetazione. Di conseguenza, se si riduce il manto vegetale si riducono anche le precipitazioni, con conseguente alterazione delle condizioni vitali di molte specie vegetali e animali abituate al clima tropicale umido della foresta. Tale tendenza è stata prevista dai modelli di simulazione del clima quando si sono incorporati in essi anche gli effetti dell'evapotraspirazione delle piante. In seguito tali risultati sono stati confermati da osservazioni sulla diminuzione delle precipitazioni sul bacino amazzonico a partire dagli anni Ottanta.
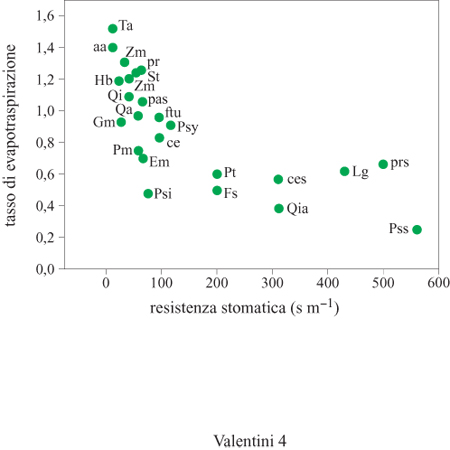
Un altro caso di notevole interesse per le implicazioni ecologiche riguarda la regione siberiana. Le regioni centrali della Siberia sono tra le zone più continentali del nostro pianeta. In queste condizioni la vegetazione può giocare un ruolo determinante sul clima regionale. Tale territorio, che si estende per migliaia di chilometri dagli Urali al fiume Lena, è dominato da un alternarsi di paludi e foreste. Queste ultime, inoltre, sono rinnovate per effetto dei frequenti incendi determinati da cause naturali, come i fulmini originati dalla formazione di nubi convettive. In questo mosaico di differenti ecosistemi le paludi e le foreste presentano comportamenti diversi nella dissipazione di energia. Durante il periodo estivo, che in queste regioni dal clima continentale è caratterizzato da temperature elevate (spesso superiori ai 28°C), le paludi presentano un rapporto tra flusso di calore sensibile e calore latente di evapotraspirazione pari a circa 1; ciò sta a indicare che l'energia disponibile è equamente distribuita tra i due processi. Le foreste invece mostrano un rapporto pari a circa 4, il che vuoI dire che l'energia disponibile è dissipata prevalentemente sotto forma di calore sensibile, mentre l'evapotraspirazione è una componente molto ridotta. Le coperture forestali generano perciò un flusso di calore che riscalda l'atmosfera sovrastante innescando moti convettivi i quali, trasportando ingenti quantità di acqua evaporata dalle paludi, danno origine alla formazione di nubi temporalesche e quindi alle precipitazioni (fig. 5). Mediante l'analisi isotopica dell'acqua piovana è stato mostrato che la maggior parte di essa si forma in seguito alla condensazione del vapore acqueo traspirato dalle piante. Si tratta quindi di un ciclo chiuso.
L'aspetto più interessante del ciclo idrologico in questa regione è comunque l'importanza dei fulmini che si generano in seguito ai fenomeni convettivi. Grazie agli incendi che da questi scaturiscono è possibile mineralizzare la sostanza organica presente nel suolo e permettere la rinnovazione di queste foreste, normalmente assai povere di nutrienti. Questo esempio mostra come i fenomeni fisici e quelli biologici siano intimamente connessi in una complessa rete di relazioni. Ogni perturbazione che modifichi la distribuzione degli ecosistemi nel territorio, intervenendo sull'alternanza di paludi e foreste, può avere un significativo impatto sul clima regionale e, di conseguenza, anche sui fenomeni di successione ecologica che sono alla base dell'evoluzione della vegetazione in questa regione.
Produttività primaria e ciclo del carbonio
Abbiamo osservato in precedenza che negli ecosistemi terrestri la maggior parte dell'energia è utilizzata nei processi di evapotraspirazione e trasporto di calore turbolento (calore sensibile). È attraverso questi flussi energetici che gli ecosistemi terrestri giocano un ruolo fondamentale nelle interazioni tra biosfera e atmosfera, influenzando sia il clima regionale che quello globale.
Sebbene solo una piccola parte dell'energia disponibile (circa il 2%) venga spesa nei processi di sintesi della sostanza organica, tali processi rappresentano un pilastro fondamentale per la vita di tutti gli esseri viventi, in quanto la produzione primaria costituisce il mattone fondamentale per il sostentamento delle catene trofiche dell'ecosistema. Tuttavia, il ruolo del metabolismo del carbonio degli ecosistemi terrestri nel ciclo globale del carbonio e l'impatto sulla concentrazione di diossido di carbonio della nostra atmosfera non sono stati studiati in modo approfondito.
Per descrivere tali processi è necessario richiamare alcune definizioni di produttività primaria. La produzione primaria lorda rappresenta la quantità di sostanza organica che viene sintetizzata nel processo della fotosintesi, la produzione primaria netta è ciò che rimane della produzione lorda al netto dei processi respiratori delle piante, mentre la produzione primaria netta dell'ecosistema è la parte di produzione primaria netta che rimane una volta detratte le perdite dovute alla respirazione degli organismi eterotrofi, tra cui i processi di decomposizione della sostanza organica. Tali definizioni si applicano a qualsiasi livello dell'ecosistema e possono essere considerate su base annua. Tuttavia molti processi che influenzano la produzione primaria si svolgono su scale temporali di decine di anni o secoli. Per esempio gli incendi, le catastrofi naturali come alluvioni o tempeste di vento e fattori biotici come gli attacchi di insetti defogliatori sono in grado di sconvolgere l'ecosistema e influenzare notevolmente la sua produttività primaria. Questi fenomeni sono particolarmente significativi se considerati su scale spaziali estese. Se consideriamo la biosfera nel suo insieme vi troveremo, per esempio, foreste molto produttive, altre sfruttate dall'uomo, altre ancora soggette a disturbi naturali. Per questo motivo è stato introdotto un nuovo concetto, quello di produzione primaria netta della biosfera, che rappresenta l'insieme del bilancio del carbonio biosferico, comprendente tutti i fattori che lo compongono.
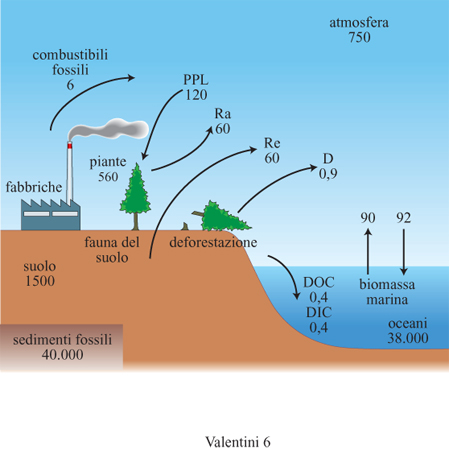
Se si osserva il ciclo del carbonio nelle sue componenti (fig. 6) si può notare come le maggiori riserve di questo elemento si trovino nei sedimenti fossili, dove sono contenute circa 40.000 Gt (1 Gt = 10⁹ t) di carbonio, di cui circa 4000 utilizzabili come combustibili fossili. Gli oceani ne contengono circa 38.000 Gt, pari a circa 51 volte il contenuto dell'atmosfera. Sulle terre emerse il maggiore serbatoio di carbonio è costituito dal suolo, che ne contiene 1500 Gt, mentre solo poco più di un terzo di questo ammontare si trova nella biomassa epigea. Nell'atmosfera vi è il serbatoio più ridotto di carbonio, con circa 750 Gt.
L'aspetto più interessante del ciclo globale del carbonio è rappresentato dai flussi di carbonio tra i vari compartimenti. Tra le terre emerse e gli oceani vi è il trasporto di carbonio organico disciolto nelle acque dei fiumi, pari a circa 0,4 Gt/anno. Un flusso equivalente si registra in termini di carbonio inorganico. Tuttavia i flussi di carbonio più importanti sono quelli che si verificano tra oceani e atmosfera e tra terre emerse e atmosfera. Lo scambio di carbonio tra oceani e atmosfera è pari a circa 92 Gt/anno nella direzione atmosfera → oceani e a circa 90 Gt/anno nella direzione opposta. Gli oceani agiscono dunque come un elemento in grado di assorbire il carbonio atmosferico, grazie soprattutto alla capacità del diossido di carbonio di diffondersi nelle acque superficiali e di depositarsi sotto forma di carbonati nelle profondità oceaniche.
Gli scambi di carbonio tra terre emerse e atmosfera sono caratterizzati da più componenti, che sono anche quelle maggiormente influenzate dalle attività umane. Gli ecosistemi terrestri ogni anno scambiano 60 Gt di carbonio con l'atmosfera. Questo valore rappresenta la produzione primaria netta degli ecosistemi terrestri, risultando dalla differenza tra le 120 Gt di produzione primaria lorda e le 60 Gt liberate dalla respirazione delle piante. La decomposizione della sostanza organica libera circa 60 Gt di carbonio ogni anno, cosicché in linea di principio gli ecosistemi terrestri sarebbero in equilibrio dal punto di vista del bilancio del carbonio: la produzione primaria netta è consumata totalmente dai decompositori e quindi la produzione primaria netta dell'ecosistema è nulla. Vedremo però tra breve che l'asserzione secondo la quale la componente terrestre della biosfera è in equilibrio deve essere rianalizzata sulla base di nuove osservazioni.
Il bilancio del carbonio planetario presenta inoltre due altri fattori inattesi che dipendono largamente dalle attività umane: l'uso dei combustibili fossili e la deforestazione tropicale. L'utilizzo dei combustibili fossili introduce ogni anno nell'atmosfera circa 6 Gt di carbonio, con un andamento in progressiva crescita. Tra tutte le componenti del bilancio del carbonio, quella legata allo sfruttamento dei combustibili fossili è la più documentata ed è in continua espansione. La deforestazione tropicale è responsabile dell'emissione nell'atmosfera di circa 1,6 Gt di carbonio ogni anno. Considerando la ricrescita parziale delle aree sottoposte a deforestazione (si tratta comunque di ecosistemi semplificati, per lo più costituiti da erbe infestanti, molto lontani dalla vegetazione preesistente), che determina un assorbimento di carbonio pari a 0,7 Gt/anno, vi è un'emissione netta di carbonio pari a 0,9 Gt di carbonio ogni anno.
Come è noto, a fronte di questi due ultimi fattori, sconosciuti alla nostra atmosfera prima della rivoluzione industriale, la concentrazione di diossido di carbonio nell'atmosfera è passata dalle 280 ppm dell'era preindustriale alle 360 ppm attuali e si prevede per l'inizio del prossimo secolo un suo raddoppio rispetto ai valori iniziali, cioè un livello di circa 560 ppm. È noto che l'aumento di diossido di carbonio è piuttosto preoccupante per le sue implicazioni nei cambiamenti climatici.
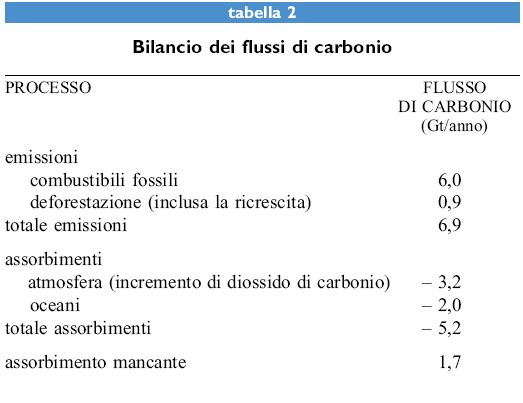
È possibile verificare il bilancio del carbonio dell'atmosfera, conoscendo i flussi in ingresso e in uscita e i cambiamenti di concentrazione annuale. In base ai dati raccolti negli anni Novanta (tab. 2) si è stimato che ogni anno vengono introdotte nell'atmosfera circa 6,9 Gt di carbonio, mentre gli oceani sono in grado di assorbime circa 2 Gt. Tuttavia, sulla base delle misurazioni di concentrazione si è valutato che nell'atmosfera terrestre entrano soltanto 3,2 Gt di carbonio ogni anno. Rimane quindi da capire dove finiscano 1,7 Gt/anno di carbonio che risultano non spiegate dalle osservazioni e dalle stime dei flussi. Bisogna cioè identificare il cosiddetto carbon sink (assorbitore di carbonio) mancante, ovvero quel fattore che sembra contrastare l'aumento di concentrazione di diossido di carbonio nell'atmosfera, rallentando le possibili conseguenze dell'effetto serra sul clima.
Una prima ipotesi formulata per spiegare lo squilibrio osservato è la possibilità che gli oceani in realtà assorbano più carbonio di quanto stimato. Tuttavia, nuove campagne di misura, e soprattutto lo studio della distribuzione del 14C sulla superficie degli oceani, mostrano che valori più elevati di assorbimento non sono possibili nelle condizioni attuali. Infatti lo scambio di diossido di carbonio con gli oceani, pur non essendo limitato dalla diffusione superficiale, in quanto il diossido di carbonio diffonde facilmente in acqua, è ostacolato dalla difficoltà con cui le acque superficiali si mescolano con le acque profonde dove il diossido di carbonio può effettivamente essere immagazzinato. L'altra ipotesi, confermata da recenti studi sulle fluttuazioni stagionali degli isotopi dell'ossigeno e dell'azoto nell'atmosfera, è che gli ecosistemi terrestri non siano in equilibrio dal punto di vista del bilancio del carbonio, come invece previsto nello schema teorico, ma che in realtà essi rappresentino un elemento assorbitore, ovvero che la produttività primaria netta della biosfera sia diversa da zero e positiva.
Le stime di produttività primaria netta degli ecosistemi terrestri sono oggi disponibili su scala globale grazie ai dati raccolti dall'lnternational Biosphere Programme. Tuttavia questi dati, per lo più limitati alla sola componente epigea, non sono in grado di spiegare il carbonio mancante nell'atmosfera, in quanto il bilancio netto di carbonio è rappresentato dalla produzione primaria netta dell'ecosistema che include anche i processi di decomposizione della sostanza organica. Soltanto recentemente, grazie allo sviluppo di nuove metodologie di misura, è divenuto possibile misurare direttamente il flusso netto di carbonio con l'atmosfera e ricavare quindi delle stime sul ruolo degli ecosistemi terrestri nel bilancio del carbonio.
La tecnica di misura eddy eovarianee (Baldocchi et al., 1996; Moncrieff et al., 1997) si basa sulla correlazione tra le fluttuazioni della componente verticale della velocità del vento e della concentrazione di diossido di carbonio nell'atmosfera in prossimità della superficie vegetale. Il flusso netto (F) che attraversa l'unità di superficie nell'unità di tempo è dato dalla media temporale del prodotto della velocità del vento (w) per la concentrazione del gas (c), in questo caso diossido di carbonio:
formula, [14]
dove la barra rappresenta il valore medio nel tempo. Poiché nell'atmosfera il moto dell'aria è turbolento, sia la velocità verticale del vento che la concentrazione possono essere decomposte in una componente media e una fluttuante (w+w', c+e') da cui:
formula. [15]
Inoltre, poiché è lecito assumere che non vi sia un moto medio dell'atmosfera verso l'alto, cioè che w = O, il flusso turbolento si può calcolare dalle sole componenti fluttuanti:
Da un punto di vista operativo è necessario disporre di strumenti adeguati in grado di misurare la componente verticale della velocità del vento e la concentrazione di diossido di carbonio nell'atmosfera sopra la copertura vegetale tanto velocemente (circa 20 volte al secondo) da catturare le fluttuazione rapide che compongono il flusso turbolento. La componente verticale della velocità del vento viene misurata con un particolare anemometro funzionante a impulsi ultrasonori, in grado di decomporre la velocità del vento nelle sue tre componenti spaziali. La concentrazione di diossido di carbonio viene misurata con un analizzatore funzionante sul principio dell'assorbimento della radiazione infrarossa da parte del gas. Infatti il diossido di carbonio, come tutti i gas serra, mostra una forte banda di assorbimento nella regione infrarossa dello spettro. Gli strumenti vengono posti al di sopra della superficie vegetale e forniscono in continuo, con cadenza oraria, il flusso netto di diossido di carbonio tra la superficie vegetale e l'atmosfera. Il vantaggio di questa metodologia è che il valore di flusso netto si riferisce direttamente alla produzione primaria netta dell'ecosistema integrata sull'area di misura, che è dell'ordine di 100 ha.
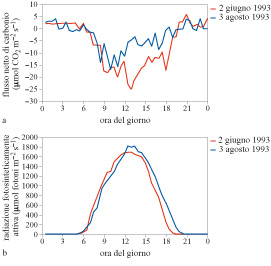
In figura (fig. 7) è presentato un esempio di dati giornalieri per una foresta di faggio dell'Appennino centro-meridionale (Valentini et al., 1996). Si può notare come il flusso netto di carbonio abbia un caratteristico andamento giornaliero in risposta alla luce e quindi ai processi foto sintetici. l valori negativi durante il giorno stanno a indicare un flusso di diossido di carbonio diretto verso la superficie. Nel periodo notturno, invece, il flusso diventa positivo, cioè è diretto dalla superficie verso l'atmosfera, indicando una perdita di diossido di carbonio nei processi respiratori.
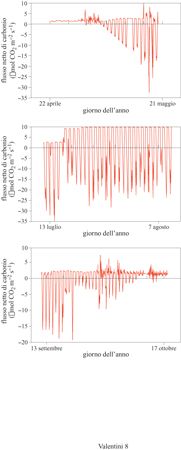
Le osservazioni possono essere ripetute per più giorni, come mostrato nella figura successiva (fig. 8). In tal modo è possibile notare la dinamica dei processi fotosintetici e respiratori della foresta di faggio nel corso della stagione. In inverno la foresta, priva di foglie, è una sorgente di diossido di carbonio, cioè i processi respiratori dominano gli scambi gassosi superficiali. Parallelamente allo sviluppo fogliare aumenta progressivamente l'assorbimento di carbonio per effetto della fotosintesi e il bilancio giornaliero dell'ecosistema si sposta complessivamente a favore dell'assorbimento di carbonio. Durante l'estate il processo è abbastanza stazionario, con l'assorbimento fotosintetico che comunque supera le perdite dovute alla respirazione. In autunno, con la diminuzione della superficie fogliare, le perdite per respirazione diventano più importanti e l'ecosistema nel suo complesso da elemento di assorbimento diventa nuovamente sorgente di diossido di carbonio verso l'atmosfera. Calcolando il bilancio annuale netto dei flussi di diossido di carbonio si può valutare se l'ecosistema agisca da sorgente o da assorbitore di carbonio atmosferico. Tale analisi (Valentini et al., 1996) mostra che la foresta di faggio analizzata funziona, su base annuale, come elemento di assorbimento di carbonio, con una capacità di accumulo pari a circa 432 g m-² di carbonio per anno.
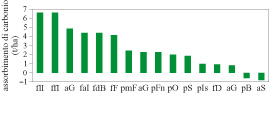
Alla fine degli anni Novanta è stata avviata un'iniziativa internazionale, denominata FLUXNET (Valentini et al., 1998b), a cui afferiscono vari progetti in diversi paesi del mondo, finalizzata allo studio delle capacità di assorbimento o di rilascio del carbonio atmosferico da parte degli ecosistemi terrestri. In figura (fig. 9) è presentata una sintesi di alcuni dati relativi a varie foreste europee. Come si può notare, alcune foreste sono sorgenti di diossido di carbonio (pino silvestre in Belgio e abete rosso in Svezia), mentre altre, soprattutto ne II ' area mediterranea, mostrano un significativo assorbimento di carbonio atmosferico.
L'insieme di questi dati mostra una distribuzione latitudinaIe degli scambi di diossido di carbonio. In particolare si può notare come le foreste delle regioni boreali mostrino valori di scambio netto di carbonio annuale molto ridotti rispetto alle foreste delle zone temperate. Tale particolare comportamento può essere spiegato dall'importanza dei processi respiratori dei vari ecosistemi. Nelle zone boreali la componente respiratoria del bilancio del carbonio è relativamente superiore a quella degli ecosistemi temperati. Ciò è spiegato dal fatto che nelle regioni boreali la quantità di carbonio organico nei suoli è molto maggiore, soprattutto nella componente volatile: nonostante le temperature siano più basse, il flusso respiratorio risulta quindi relativamente superiore rispetto ai processi foto sintetici. Inoltre, la componente respiratoria è influenzata in modo sostanziale dalla disponibilità di risorse idriche che determinano una riduzione della respirazione negli ecosistemi delle zone temperate. Tali osservazioni dimostrano comunque che, nonostante le evidenze sperimentali su scala globale suggeriscano la presenza di un significativo assorbimento di carbonio da parte degli ecosistemi terrestri, la sua localizzazione geografica è ancora oggetto di discussione. In ogni caso, le osservazioni compiute mediante le nuove metodologie di studio dei flussi di carbonio negli ecosistemi confermano l'importanza della componente biologica nel ciclo del carbonio e, più in generale, nella regolazione della composizione chimica dell'atmosfera e dei processi alla base del nostro sistema climatico.
Bibliografia citata
BALDOCCHI, D., V ALENTINI, R., RUNNING, S., OECHEL, W., DAHLMAN, R. (1996) Strategies for rneasuring and modelling carbon dioxide and water vapour fluxes over terrestrial ecosystem. Global Change Biology, 2, 159-168.
JARVIS, P.G., McNAUGHTON, K.G. (1986) Stornatal control of transpiration: scaling up frorn leaf to region. Adv. Ecol. Res., 15, 1-49.
LEVINE, J.S. (1989) Photochemistry of biogenic gases. In Global ecology: towards a science of the biosphere, a c. di Rambler M.B., Margulis L., Fester R., Boston, Academic Press, 51-74.
LOVELOCK, J.E. (1979) Gaia: a new look at life on Earth. Oxford-New York, Oxford University Press.
MONCRIEFF, J., VALENTINI, R., GRECO, S., SEUFERT, G., CICCIOLI, P. (1997) Trace gas exchange over terrestrial ecosystems: methods and perspectives in micrometeorology. J. Exp. Bot., 48, 1133-1142.
MONTEITH, J.L., a c. di (1975) Vegetation and the atmosphere, 1. Londra-New York, Academic Press.
PIELKE, R.A., DALU, G., SNOOK, J.S., LEE, T.J., KiTTEL, T.G.P. (1991) Non linear influence of mesoscale land use on weather and climate. J. Climate, 4, 1053-1069.
SCHUKLA, J., NOBRE, C., SELLERS, P. (1990) Amazon deforestation and climate change. Science, 247, 1322-1325.
SELLERS, P.J. et al. (1996) Modelling the exchanges of energy, water and carbon between continents and the atmosphere. Science, 275, 502-509.
VALENTINI, R., BALDOCCHI, D.D., TENHUNEN, J. (1998a) Ecological controls on land-surface atmospheric interactions. In Integrating hydrology, ecosystem dynamics and biogeochemistry in complex landscapes, a c. di Tenhunen J., Kabat P., Chichester, John Wiley & Sons Ltd.
VALENTINI, R., BALDOCCHI, D., RUNNING, S. (1998) The 1GB P BARC global flux network initiative (FLUXNET): current status and perspectives. Global Change Newsletter, 28,14-16.
VALENTINI, R., DE ANGELIS, P., MATTEUCCI, G., MONACO, R., DORE, S., SCARASCIA MUGNOZZA, G.T. (1996) Seasonal net carbon dioxide exchange of a beech forest with the atrnosphere. Global Change Biology, 2, 199-208.
WALTER, R. (1973) Vegetation of the Earth. New York, Springer Verlag.
WEBB, R.S., ROSENZWEIG, C.E., LEVINE, E.R. (1993) Specifying land surface characteristics in generaI circulation model. Soil profile data set and derived water holding capacities. Global Biogeochem. Cycles, 7, 97-108.
XUE, Y., SHUKLA, J. (1993) The influence of land surface properties on Sahel climate. Part I: desertification. J. Climate, 4, 345-363.