
fototrasduzione
fototrasduzione
La fototrasduzione è il processo attraverso il quale i fotorecettori della retina convertono l’assorbimento di onde elettromagnetiche in segnale nervoso. Nella retina si trovano 5 tipi di cellule, tra cui i fotorecettori (coni e bastoncelli); questi contengono il fotopigmento, composto da una parte proteica, chiamata opsina, e da un cromoforo, il retinale, dalla cui isomerizzazione per assorbimento di un fotone parte il processo di trasduzione, con l’attivazione della cascata intracellulare che determina l’insorgenza di un cambiamento del potenziale di membrana. I bastoncelli possiedono un solo tipo di opsina, i coni ne possiedono invece tre. Combinando le risposte dei tre diversi tipi di coni è possibile discriminare lunghezze d’onda diverse e quindi avere una visione a colori. I bastoncelli non danno la visione a colori ma in compenso sono fotodetettori più sensibili dei coni e sono utilizzati nella visione con scarsa visibilità. La fototrasduzione dà luogo a fenomeni di amplificazione che variano in funzione della quantità di luce disponibile, in quanto i fotorecettori modificano la loro sensibilità in relazione alla quantità di luce ambientale (fenomeno dell’adattamento. [➔ percezione del colore; potenziale d’azione; recettore nervoso; visione]
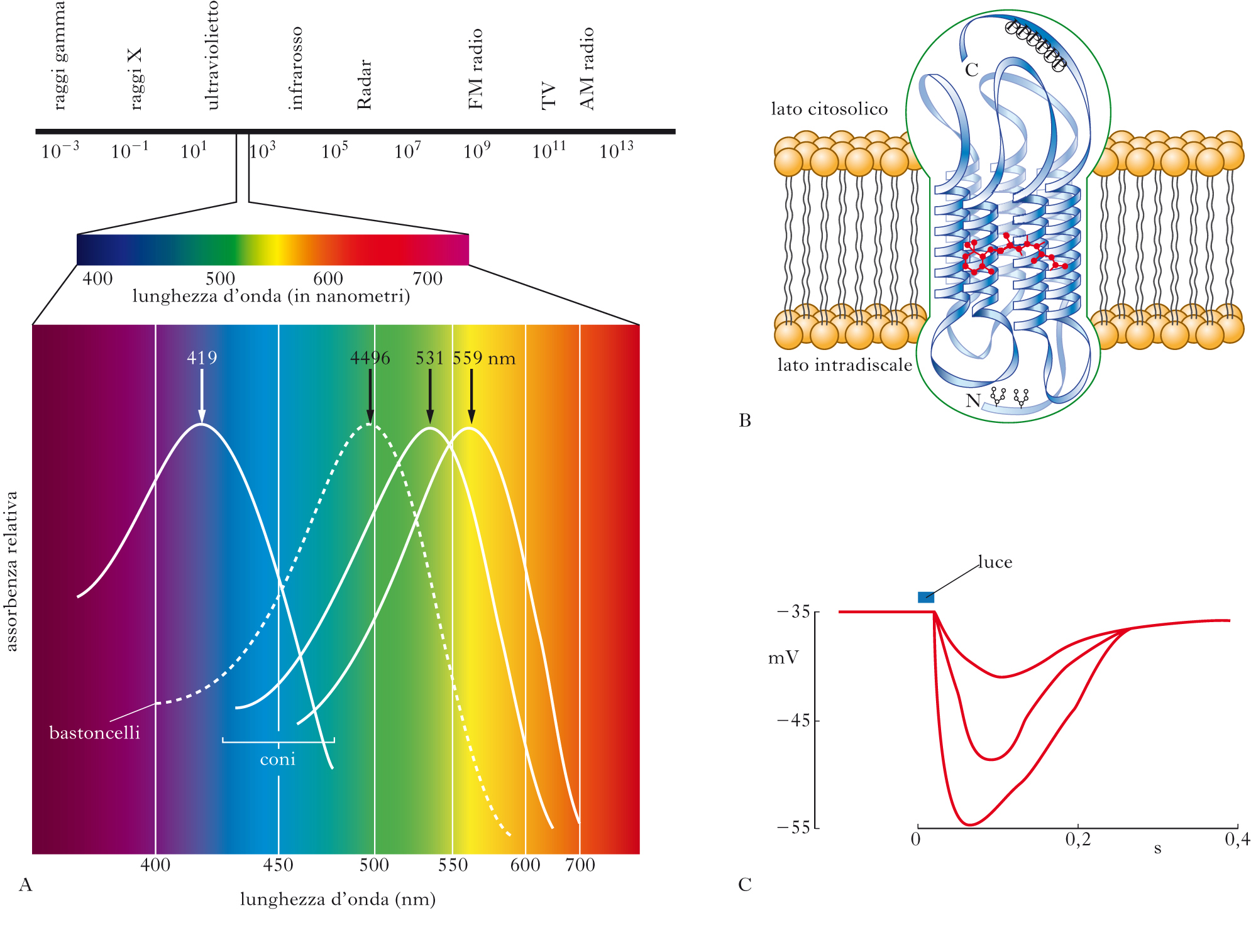
Solo le onde elettromagnetiche di lunghezze d’onda tali da poter essere assorbite dai fotopigmenti dei nostri fotorecettori evocano una sensazione visiva. L’intervallo di lunghezze d’onda delle onde elettromagnetiche che sono per noi visibili va da poco meno di 400 nm (corrispondente al colore violetto) a poco più di 700 nm (corrispondente al colore rosso). Anche se gli esseri umani non percepiscono i raggi ultravioletti o infrarossi come luminosi, altre specie sono sensibili a queste porzioni dello spettro elettromagnetico. Per es., gli insetti sono sensibili alle lunghezze d’onda dell’ultravioletto e alcuni fiori, che a noi appaiono di colore uniforme, agli insetti appaiono invece screziati da venature che puntano alla zona contenente il polline.
Le cellule della retina
Nella retina si trovano 5 tipi di cellule: i fotorecettori (coni e bastoncelli, per un totale di circa 125 milioni di fotorecettori), le cellule bipolari, le cellule orizzontali, le cellule amacrine e le cellule gangliari. Nella via diretta di trasmissione dell’informazione, i fotorecettori sono sinapticamente connessi con le cellule bipolari, le quali a loro volta trasmettono l’informazione alle cellule gangliari. Le cellule orizzontali mediano interazioni di inibizione laterale fra fotorecettori e le cellule amacrine modulano la trasmissione del segnale fra cellule bipolari e cellule gangliari. La luce arriva allo strato dei fotorecettori dopo aver attraversato l’intero spessore della retina, con l’eccezione della fovea. Nella fovea le altre cellule retiniche si ritraggono, lasciando esposti soltanto i fotorecettori. Gli assoni delle cellule gangliari convergono in un punto della retina detto papilla e da qui escono dall’occhio, formando il nervo ottico. Le regioni dei fotorecettori che contengono il fotopigmento, ossia le molecole che hanno la funzione di assorbire la luce, si trovano nel segmento esterno, cilindrico nei bastoncelli e appuntito nei coni. Il segmento esterno contiene ripiegamenti della membrana plasmatica che aumentano la superficie utile per l’assorbimento della luce, e che nei bastoncelli diventano veri e propri dischi, separati tra loro dalla membrana plasmatica e densamente impilati. Le molecole di fotopigmenti si trovano sulla membrana di questi ripiegamenti/dischi e sono in numero molto elevato (fino a 109 molecole di fotopigmento in un singolo bastoncello).
I fotopigmenti
Il fotopigmento è composto da una parte proteica, chiamata opsina, una molecola che si ripiega ripetutamente formando una struttura tridimensionale con 7 domini transmembrana, e da un cromoforo, il retinale, che è un derivato della vitamina A. Il retinale è posto all’interno di una tasca formata dalla struttura proteica dell’opsina ed è legato a essa attraverso un legame covalente. È il retinale che assorbe la luce. Il legame della tasca di opsina con il retinale modifica lo spettro di assorbimento di quest’ultimo, spostandolo verso lunghezze d’onda più lunghe e determinando così lo spettro del visibile. I bastoncelli possiedono un solo tipo di opsina, e quindi un solo fotopigmento, la rodopsina, che assorbe massimamente la luce per lunghezze d’onda intorno a 500 nm. I coni possiedono invece tre tipi di opsina, per cui esistono tre popolazioni di coni, una per cui il fotopigmento ha assorbimento massimo nel rosso (più propriamente nel giallo-arancio), una che assorbe massimamente nel verde e una nel blu. Combinando le risposte dei tre diversi tipi di coni è possibile discriminare lunghezze d’onda diverse e quindi avere una visione a colori. I bastoncelli, con il loro unico fotopigmento, non possono dare invece alcun contributo alla visione a colori. La mancanza o la mutazione dei geni di uno o più dei fotopigmenti dei coni causa specifici deficit nella visione dei colori. La mancanza di due o tutti e tre i fotopigmenti dei coni causa una mancanza totale della visione a colori. La mancanza di un solo fotopigmento causa deficit meno severi: questo è il caso, per es., dei soggetti dicromati, deuteranopi o protanopi (noti come daltonici), che sono incapaci di percepire una differenza fra rosso, arancio, giallo e verde, colori così vividamente diversi per un osservatore normale. Nei protanopi, nei quali manca il fotopigmento che assorbe nel rosso, il colore rosso appare grigio scuro, quasi nero.
Le tappe della fototrasduzione
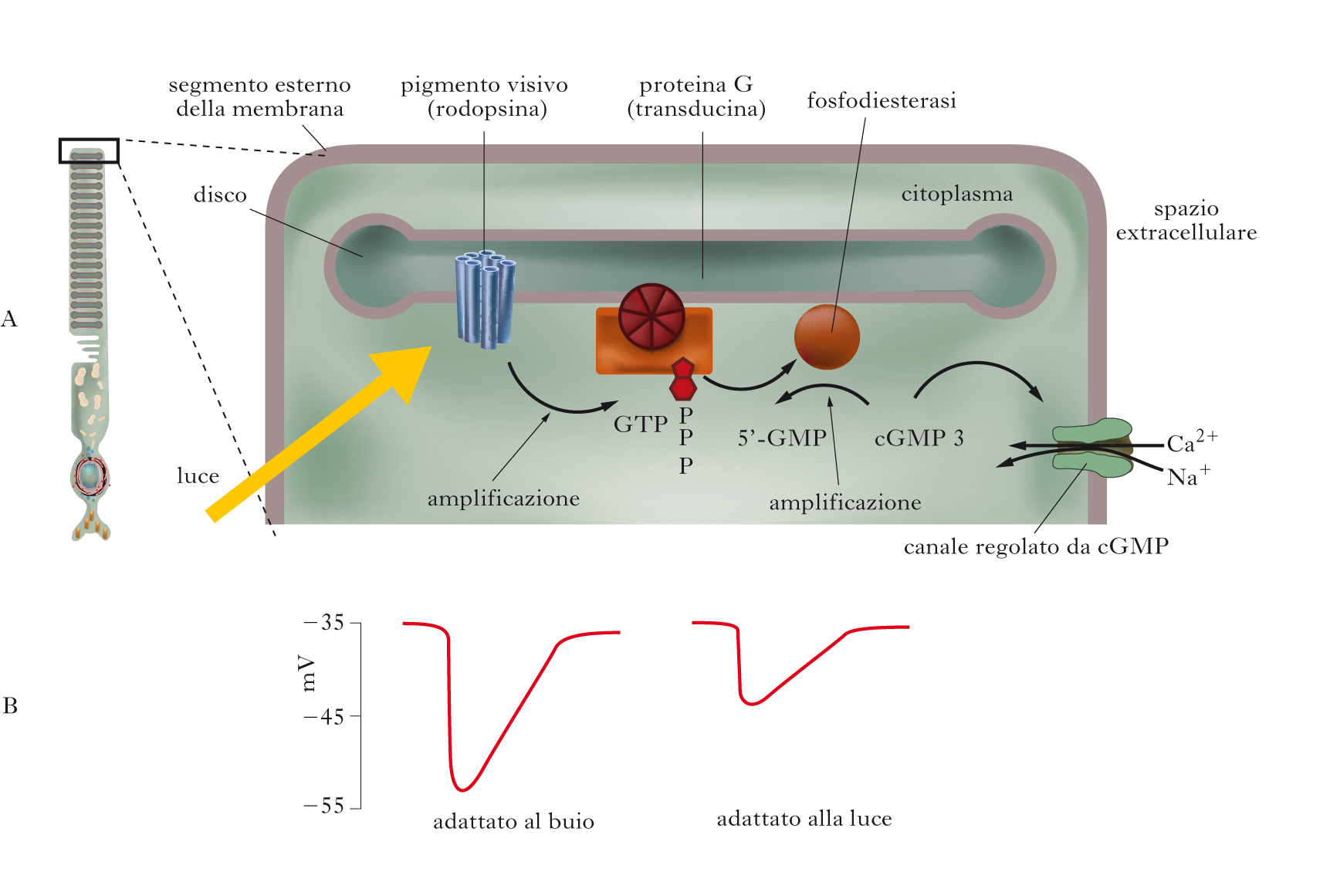
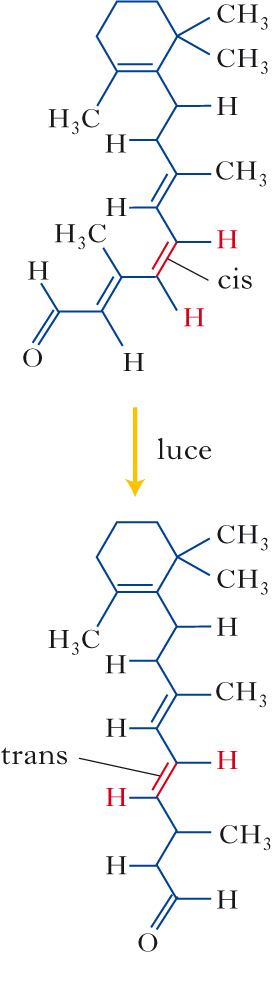
Il primo passo della f. consiste nell’assorbimento di un fotone da parte del retinale, che determina la sua isomerizzazione dalla configurazione 11-cis a quella tutta trans. Questa isomerizzazione innesca cambiamenti conformazionali nell’opsina, che transisce alla forma metarodopsina II (MR II), causando l’attivazione della cascata intracellulare della f., che determina l’insorgenza di un cambiamento del potenziale di membrana, ossia di un segnale nervoso. Tale segnale, chiamato potenziale di recettore, è di tipo analogico e consiste in una iperpolarizzazione, con conseguente riduzione del rilascio di neurotrasmettitore. Per le cellule bipolari, quindi, l’arrivo della luce sui fotorecettori con cui sono connesse sinapticamente si traduce in una riduzione del neurotrasmettitore. Tale riduzione determina a sua volta depolarizzazione di una classe di cellule bipolari, le cellule bipolari centro ON – che segnalano principalmente aumenti locali della intensità luminosa –, e iperpolarizzazione di un’altra classe di cellule bipolari, le cellule centro OFF – che segnalano principalmente riduzioni locali di luminosità. I segnali delle cellule bipolari centro ON e centro OFF sono raccolti da cellule gangliari, rispettivamente centro ON e centro OFF; le cellule gangliari codificano l’informazione in potenziali d’azione che, viaggiando lungo il loro assone, raggiungono le strutture nervose successive lungo le vie visive, per essere ulteriormente elaborate in modo da determinare la percezione cosciente. Come accade per tutti i segnali analogici, l’ampiezza del potenziale di recettore aumenta all’aumentare dell’intensità dello stimolo luminoso, fino a raggiungere un massimo di iperpolarizzazione oltre il quale ulteriori aumenti dell’intensità dello stimolo non provocano altri incrementi (saturazione). La minima intensità luminosa che provoca un potenziale di recettore viene invece chiamata soglia.
Il ruolo della metarodopsina. In che modo la MR II determina il potenziale di recettore? In condizioni di buio, i fotorecettori presentano sulla membrana plasmatica numerosi canali ionici aperti e permeabili a Na+ e anche a Ca2+; di conseguenza, si verifica un continuo flusso di ioni positivi. Questa corrente ionica si chiama corrente di buio. A causa di questa elevata permeabilità ai cationi, il potenziale di membrana dei fotorecettori al buio sarà più depolarizzato di quello presentato dalla maggior parte delle cellule nervose a riposo e si aggira intorno ai −35, −40 mV. La MR II attiva una proteina appartenente alla classe delle proteine G, la transducina, che a sua volta attiva un enzima, la fosfodiesterasi (PDE), che riduce la concentrazione di un secondo messaggero, il guanosinmonofosfato ciclico (GMPc). I canali di membrana permeabili a Na+ e a Ca2+ sono tenuti aperti dal legame con molecole di GMPc, per cui, quando la concentrazione di GMPc si riduce, essi si chiudono, determinando la iperpolarizzazione dei fotorecettori. La risposta alla luce si ‘spegne’ con il contributo di molti fattori, fra cui quello di una proteina specifica, l’arrestina, che inibisce il legame tra MR II e transducina, e nuovo GMPc viene prodotto, causando la riapertura dei canali permeabili a Na+ e Ca2+ e la fine del potenziale di recettore. L’opsina si separa poi dal retinale, che migra fuori dai fotorecettori, nell’epitelio pigmentato, per l’inizio del processo di riconversione alla forma 11-cis che rende possibile la nuova sintesi delle molecole di fotopigmenti nei fotorecettori attraverso la ricombinazione del retinale 11-cis con l’opsina.
Processi di adattamento al buio e alla luce. Il processo di f. presenta due punti di amplificazione: ogni molecola di MR II attivata può attivare fino a circa 500 molecole di transducina e ogni molecola di PDE può eliminare fino a 1000 molecole di GMPc: di conseguenza, un singolo fotone può determinare la eliminazione di 500.000 molecole di GMPc, con conseguente chiusura di centinaia di canali di membrana. L’amplificazione non è costante, ma varia in funzione della quantità di luce disponibile. Infatti i fotorecettori modificano la loro sensibilità in funzione della quantità di luce ambientale: se la quantità di luce ambientale aumenta, la sensibilità dei fotorecettori diminuisce rapidamente (adattamento alla luce); se invece la quantità di luce ambientale diminuisce, la sensibilità dei fotorecettori aumenta progressivamente, raggiungendo il massimo dopo circa 25÷30 minuti (adattamento al buio). Il processo di adattamento alla luce è spiegabile con una riduzione del guadagno della f., legata principalmente alle variazioni della concentrazione di Ca2+ nei fotorecettori in seguito alla chiusura dei canali di membrana sensibili al GMPc causata dalla luce; il processo di adattamento al buio è spiegabile con un aumento del guadagno della f. e con la progressiva nuova sintesi del fotopigmento, fenomeno che richiede tempi lunghi (dell’ordine dei 6 minuti nei coni e dei 25÷30 minuti nei bastoncelli), il che rende ragione della più lunga durata del processo di adattamento al buio rispetto a quello alla luce. Il processo di adattamento nei fotorecettori serve a conciliare la possibilità di rispondere a piccole quantità di luce, che richiede un elevato guadagno del processo di f., con la necessità di evitare la saturazione già a bassi livelli di luminosità, conseguenza di un guadagno elevato fisso. La soluzione è un guadagno variabile in funzione della quantità di luce presente.
Soglia e saturazione dei fotorecettori
Quale è la minima quantità di luce che un fotorecettore deve assorbire per rispondere allo stimolo, nella situazione di massima sensibilità? Per i bastoncelli, è sufficiente che venga assorbito un singolo fotone perché si generi un segnale misurabile. Il bastoncello è quindi un fotodetettore estremamente sensibile. Questo avviene però solo se il bastoncello è adattato al buio. I coni non sono sensibili come i bastoncelli: al massimo della loro sensibilità hanno bisogno di almeno un centinaio di fotoni per iniziare a rispondere alla luce. La loro soglia è quindi circa 100 volte più alta di quella dei bastoncelli, ossia (essendo la sensibilità inversa alla soglia) hanno una sensibilità 100 volte minore. Ci si può rendere conto di questo fatto osservando il cielo stellato: alcune stelle di debole intensità sono visibili solo se non le fissiamo direttamente ma se invece fissiamo un punto distante qualche grado di angolo visivo. Nel primo caso, l’immagine della stella cade sulla fovea, dove sono presenti soltanto i coni, la cui bassa sensibilità non consente di vederla, mentre nel secondo caso cade in una zona della retina ricca di bastoncelli che, con la loro alta sensibilità, consentono la visione dell’oggetto. I coni saturano per quantità di luce molto più alte di quelle per cui saturano i bastoncelli. Quindi, se la luce ambientale è scarsa (visione scotopica) si utilizza la grande sensibilità dei bastoncelli per vedere, mentre, se la luce ambientale è elevata (visione fotopica), i bastoncelli risultano saturati e si utilizzano i coni. La presenza di due tipi di fotorecettori con soglie e punti di saturazione diversi, entrambi in grado di adattare e quindi di lavorare su un ampio intervallo di quantità di luce, permette al sistema visivo di operare in un range di variazioni della quantità di luce disponibile pari a 1011. Ciò significa che tra la minima quantità di luce sufficiente ancora a intravedere i contorni degli oggetti e la massima quantità oltre la quale si ha abbagliamento c’è un rapporto di 100 miliardi. I coni e i bastoncelli sono come due pellicole fotografiche, una poco sensibile e l’altra molto, in grado di sostituirsi l’una con l’altra in funzione della quantità di luce, regolando il guadagno della fototraduzione. Nicoletta Berardi