
genetica e malattie genetiche
genetica e malattie genetiche
La scienza che studia l’ereditarietà
I progressi della genetica, nata nell’Ottocento con Gregor Mendel, ci hanno permesso di capire come i geni paterni e materni si distribuiscono nei gameti (uova e spermatozoi) e come dall’incontro dei gameti possano nascere organismi diversi dai genitori. Lo studio delle mutazioni nelle popolazioni umane ha poi permesso di conoscere e capire molte malattie genetiche, fra le quali quelle dovute a mutazioni recessive, dominanti o legate al cromosoma X e quelle dovute a gravi alterazioni del numero e della struttura dei cromosomi
Perché siamo biondi o bruni
Tutti i ragazzi sono interessati a sapere perché sono biondi o bruni, da dove proviene la forma del loro naso, perché il loro amico suona bene la chitarra e loro sono stonati, o come è potuto nascere, da due genitori come tutti gli altri, il ragazzino affetto dalla sindrome di Down che frequenta la loro scuola.
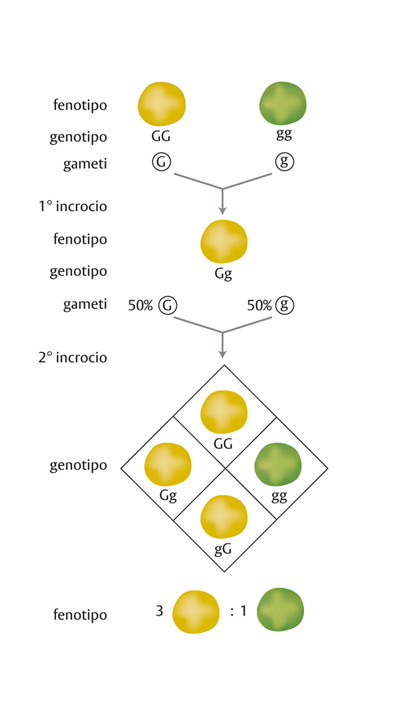
La risposta a queste domande cominciò a venire da un monaco, ottimo matematico, Gregor Mendel (vissuto nel 19° secolo), che coltivava l’orto del suo convento di Brno in Moravia. Studiando i caratteri delle piante di piselli, Mendel descrisse le leggi dell’ereditarietà, base della moderna genetica (fig.). Molti anni dopo Mendel, altri biologi si dedicarono a questi studi e definirono gene quel qualcosa, allora ancora non chiaro, che, passando dai genitori ai figli, ne determina le caratteristiche del corpo e anche qualche loro capacità particolare, come per esempio l’orecchio musicale.
Gli esperimenti di genetica conobbero uno sviluppo meraviglioso grazie soprattutto alle drosofile, i moscerini della frutta, che come materiali di laboratorio hanno tre vantaggi rispetto ai piselli: si riproducono molto più in fretta – ogni due settimane, nutriti di miele e messi dentro bottiglioni tappati con cotone –, hanno caratteri molto più numerosi e i loro cromosomi si vedono al microscopio con grande facilità.
Analizzando le caratteristiche delle drosofile da una generazione all’altra, e mettendole in relazione con i cromosomi visibili all’interno delle cellule con buoni microscopi ottici, è stato dimostrato che, come aveva previsto Mendel, ogni carattere è dovuto a una coppia di geni, derivati dal padre e dalla madre, chiamati alleli. Durante la formazione dei gameti (uova e spermatozoi), il numero dei geni allelici viene ridotto da due a uno in un processo di divisione cellulare chiamato meiosi, in modo che, per ogni carattere, i gameti posseggono solo un allele, quello paterno o quello materno. Dopo l’incontro dell’uovo e dello spermatozoo, si forma la prima cellula del nuovo essere, lo zigote, da cui si svilupperà il nuovo organismo. In questa fase si ricostituisce il numero doppio di cromosomi – tipico degli organismi con due genitori, come la drosofila o l’essere umano – che formeranno 23 coppie di cromosomi omologhi, uno ricevuto dal padre e uno dalla madre. Di queste coppie di cromosomi omologhi, per effetto della meiosi uno solo – a caso, il paterno o il materno – andrà, quasi fosse tirato a sorte, a costituire l’informazione genetica di ogni gamete.
Come possono diverse combinazioni di geni dare caratteri diversi?
Un problema lasciato aperto dagli esperimenti di Mendel era quello delle differenze fra i due alleli. Non si riusciva a capire perché uno determinasse il colore giallo dei semi di pisello e l’altro il colore verde. Usando un termine derivato dalla lingua greca, si definisce come fenotipo ogni carattere visibile corrispondente a un gene, e nel caso specifico il fenotipo è il colore dei semi, giallo o verde.
Mendel aveva osservato che dall’incrocio fra piante a semi gialli e quelle a semi verdi nascevano solo piante di colore giallo, mentre il colore verde ricompariva soltanto se si incrociavano tra loro i piselli figli, ma in una pianta su quattro. La differenza fra i due alleli era così evidente che Mendel e i genetisti dopo di lui definirono dominante l’allele del colore giallo e recessivo quello del colore verde.
L’allele dominante in molti casi è anche quello che permette la funzione normale dell’organismo, ed è indicato con una lettera maiuscola. L’allele recessivo è invece la conseguenza di una mutazione che danneggia la funzione normale, viene chiamato mutato e indicato con una lettera minuscola. Per l’appunto, negli esperimenti di Mendel il colore giallo è la condizione normale di quel tipo di piselli ed è dovuto all’allele dominante G grande che produce una proteina capace di legarsi a un pigmento (una sostanza colorata) giallo. L’allele G, a seguito di una mutazione, diventa recessivo g piccolo, perché produce una proteina diversa dalla precedente, incapace di legarsi al pigmento, per cui vediamo il seme verde com’è la pianta perché la clorofilla non è mascherata dal pigmento giallo.
Prepotenti e sottomessi?
Cosa ha ottenuto Mendel negli incroci dei piselli? Ha incrociato fra loro ripetutamente piselli gialli fino a ottenere sempre e soltanto piselli gialli, cioè piante in cui entrambi gli alleli erano sicuramente solo di tipo G grande. Per indicare i due tipi di geni presenti scriviamo GG e diciamo che questo è il genotipo responsabile del fenotipo colore giallo della pianta. Siccome nella formazione dello zigote partecipano due alleli dominanti uguali, diciamo che questi piselli sono omozigoti per l’allele dominante giallo G grande. Facendo lo stesso con i piselli verdi, in cui è assente la capacità di prendere il pigmento giallo, Mendel ha ottenuto piante tutte a fenotipo verde e a genotipo gg, cioè omozigoti per l’allele recessivo g piccolo. Fino a qui tutto sembra facile: al genotipo GG corrisponde il fenotipo giallo e al genotipo gg corrisponde il fenotipo assenza di giallo, cioè verde.
Ma cosa succede incrociando un pisello giallo GG, omozigote dominante, con uno verde gg, omozigote recessivo? A seguito della meiosi uno solo dei due alleli di ogni genitore va a costituire il nuovo zigote, e quindi in questo incrocio l’allele G grande della pianta gialla e l’allele g piccolo di quella verde formano il nuovo genotipo Gg, definito eterozigote, perché entrano nello zigote due alleli diversi. Ebbene, al genotipo eterozigote Gg, in cui sono rappresentati entrambi gli alleli, corrisponde un solo fenotipo, quello corrispondente all’allele giallo dominante, mentre l’allele g piccolo recessivo verde è sparito dal fenotipo. Allora, esistono alleli ‘prepotenti’ e alleli ‘sottomessi’?
Zelanti e pasticcioni
Solo molti anni dopo, fra il 1950 e il 1970, quando è nata una nuova scienza chiamata biologia molecolare, si è visto che il gene è una parte della molecola del DNA e contiene l’informazione per costruire, utilizzando il codice genetico, una precisa proteina. Ecco quindi spiegato il rapporto diretto fra genotipo e fenotipo: alla parte del DNA (v. DNA e RNA) che forma un dato gene corrisponde una proteina e a ogni proteina una funzione; questa funzione stabilisce il fenotipo che noi possiamo osservare con i nostri occhi.
Anche la genetica è allora diventata molecolare: da questa è nata, all’inizio degli anni Ottanta, l’ingegneria genetica e la tecnologia del DNA ricombinante. Grazie a questi progressi si sono trovate molte risposte alle domande a cui gli strumenti a disposizione dei genetisti classici non potevano dare soluzione. L’allele dominante G grande è un gene capace di formare la proteina funzionante, in grado cioè di legarsi al colore giallo per cui il seme sarà giallo, mentre una mutazione che altera la struttura del gene, dando luogo all’allele recessivo g piccolo, porta l’informazione per una proteina non funzionante cioè incapace di legarsi al colore giallo.
Ora possiamo rispondere alla domanda sull’allele ‘prepotente’ e quello ‘sottomesso’, analizzando cosa accade veramente nei piselli a genotipo eterozigote Gg. Il fenotipo corrispondente al genotipo Gg è giallo perché è sufficiente 50% di proteina funzionante prodotta dall’allele dominante G grande a dare il colore giallo. L’allele dominante non è prepotente, ma semplicemente fa il suo lavoro per entrambi, mentre l’allele recessivo non è sottomesso, ma è soltanto un ‘pasticcione’ che finché c’è l’altro se ne sta a guardare. È quello che succede anche a coppie di esseri umani!
Mutazioni e malattie genetiche
Come può nascere un bambino con una malattia genetica da due genitori sani? Anche la risposta a questa domanda viene dagli studi originali di Mendel sulle piante, che utilizzano gli stessi meccanismi di base della genetica valida per gli animali e per l’uomo.

I geni da cui dipendono funzioni importanti dell’organismo possono mutare, provocando così la mancanza di quelle funzioni e quindi malattie genetiche più o meno gravi, che sono spesso già evidenti alla nascita. Al contrario di un raffreddore o di un’influenza, una malattia genetica ha spesso origine molte generazioni prima, dai nonni dei nonni dei nonni, e si prolunga per molte generazioni che seguono. Una nuova mutazione è un evento raro, che avviene durante la formazione di un gamete, e danneggia sempre solo uno dei due alleli, rendendolo recessivo, mentre l’altro allele, rimasto integro e dominante, continua a fare bene il suo lavoro. Come dicevamo sopra, un eterozigote ha un allele normale e uno mutato. Pertanto, una persona eterozigote per una malattia genetica ha un solo allele recessivo mutato e sta bene perché l’altro allele, normale e dominante, è capace di svolgere da solo tutto il lavoro necessario (fig.).
L’anemia mediterranea
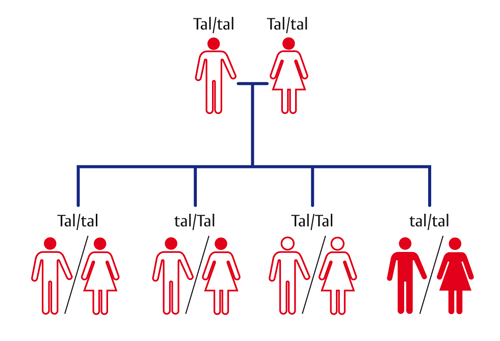
Possiamo ora fare l’esempio di una malattia genetica dell’emoglobina (la proteina dei globuli rossi che trasporta l’ossigeno), che è abbastanza comune in alcune regioni italiane come la Sardegna o nella zona del Po. Si tratta dell’anemia mediterranea o talassemia (v. anemie, leucemie, mononucleosi), il cui nome indica che è frequente nell’area del Mediterraneo (fig.). L’allele mutato tal è recessivo e negli individui eterozigoti, con un allele normale Tal e uno mutato tal, provoca soltanto la formazione di globuli rossi più piccoli (microcitemia), senza manifestazioni patologiche. Un eterozigote Tal/tal produce due tipi di gameti, uno con l’allele normale dominante Tal e l’altro con quello mutato recessivo tal. Se sposa una persona Tal/Tal, le probabilità dicono che i figli saranno per metà normali Tal/Tal e per metà eterozigoti Tal/tal, comunque nessuno sarà malato.
Se una persona eterozigote Tal/tal sposa un altro eterozigote Tal/tal, questa coppia avrà figli con quattro possibili genotipi, due eterozigoti Tal/tal non malati, un omozigote dominante Tal/Tal e un omozigote recessivo tal/tal. In quest’ultimo caso entrambi gli alleli sono mutati e provocano la formazione di molecole di emoglobina non funzionanti che deformano e distruggono i globuli rossi in cui sono contenute, dando luogo ad anemia molto grave. Per questo motivo gli eterozigoti con un allele recessivo vengono chiamati portatori della malattia.
Esistono alleli mutati dominanti?
La situazione di mutazioni recessive descritta sopra si capisce facilmente se si immagina che i due alleli siano due studenti che abitano assieme in un appartamento e si danno una mano per tenere in ordine la casa. Se uno dei due è malato (o semplicemente pigro), l’altro riesce ugualmente a fare i lavori di casa. Se entrambi sono ammalati o fannulloni, la casa si ‘ammala’ presto di disordine.
Ma cosa succede se, come può accadere, l’allele mutato è dominante? Immaginiamo ancora una volta la casa di due studenti, entrambi bravi e operosi. Improvvisamente uno dei due cambia carattere (ha studiato troppo? ha avuto una delusione d’amore?), diventa prepotente e cattivo e non tiene più in ordine la casa. Lo studente rimasto normale non riesce a rimediare e presto la casa diventa invivibile. Questa è esattamente la situazione delle malattie genetiche dovute a un allele mutato dominante.
La corea di Huntington è una malattia genetica di questo tipo. George Huntington era un medico americano che nel 1872 aveva scoperto fra i suoi pazienti una strana malattia del sistema nervoso che iniziava sempre in persone adulte, tra i 35 e i 45 anni, e provocava movimenti inarrestabili alle braccia e alla testa (come una danza, per questo il termine greco còrea) e portava alla morte in 10÷20 anni.
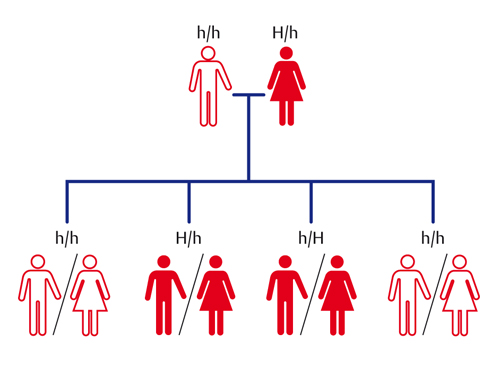
La malattia è rara, solo un malato su 10.000 sani, ed è dovuta a una proteina ‘impazzita’, chiamata huntingtina, prodotta da un unico allele dominante mutato. Quindi colpisce anche individui eterozigoti e si ammalano inesorabilmente metà dei figli di una persona a sua volta ammalata (ma che non lo sapeva a 30 anni, quando ha avuto i bambini) che ha sposato una persona normale, secondo lo schema riportato nella figura (fig. a, fig. b e fig. c).
Malattie genetiche legate al sesso
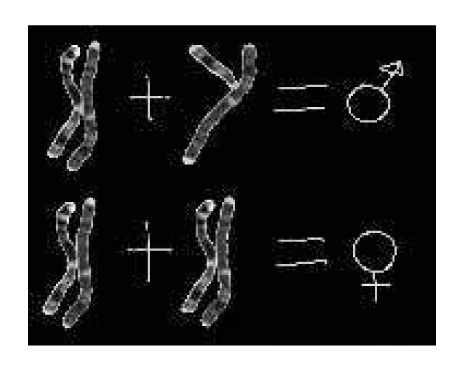
Alcune malattie genetiche associate al cromosoma X colpiscono quasi solo i figli maschi. Questa è la situazione di molte malattie genetiche, fra cui la distrofia muscolare di Duchenne, una grave alterazione progressiva dei muscoli dovuta a un allele mutato recessivo d che produce una proteina, la distrofina, inattiva. In questa malattia una madre eterozigote e sana che sposa un uomo normale ha un figlio maschio ammalato di distrofia su due, mentre le figlie femmine hanno tutte fenotipo sano (fig.).
Questa strana situazione si spiega facilmente se consideriamo che il gene per la distrofina è presente sul cromosoma X. I cromosomi sessuali X e Y sono distribuiti nella disposizione XX nelle femmine e XY nei maschi. Mentre il cromosoma Y contiene soltanto l’informazione genetica per far diventare maschio l’embrione che cresce, il cromosoma X contiene, oltre quelli necessari a regolare il sesso della nascitura, anche molti altri geni per funzioni importanti, ma non riguardanti il sesso, fra cui la distrofina normale, necessaria alla funzione muscolare. Una donna eterozigote XDXd (significa che su un cromosoma X c’è l’allele dominante De sull’altro l’allele recessivo d) portatrice di una mutazione recessiva d a carico di un solo allele, produce 50% di distrofina mutata e 50% di distrofina normale e quindi ha fenotipo sano. La stessa mutazione provoca invece gravi danni al fenotipo dei maschi, che avendo genotipo XdY (associazione cromosomica detta emizigote), senza l’allele normale D, non hanno la distrofina normale e sono colpiti dalla malattia.
Nella tabella 3 sono riportate le meiosi per la formazione dei gameti femminili e maschili e le possibili conseguenti unioni fra uova e spermatozoi. Ne risulta chiaro il motivo per cui una donna sana, ma portatrice, ha 50% di figli maschi ammalati e 100% di figlie femmine sane, ma a loro volta per il 50% eterozigoti portatrici.
I bambini down hanno un cromosoma in più
Le mutazioni non riguardano soltanto la struttura di un singolo gene (mutazioni geniche), ma anche il modo come sono costruiti i cromosomi (mutazioni cromosomiche) o addirittura il loro numero complessivo (mutazioni genomiche).
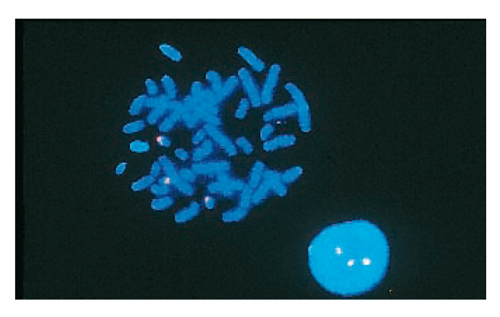
Lo sviluppo degli embrioni con mutazioni cromosomiche e genomiche è quasi sempre impossibile per la gravità del danno genetico e si avrà un’interruzione spontanea di gravidanza (aborto spontaneo). Però alcune fra queste mutazioni sono meno gravi e quindi gli embrioni che ne sono affetti riescono a nascere. Fra questi ci sono i bambini, che hanno una malattia chiamata sindrome di Down. John Langdon Haydon Down è il nome del medico che per primo ha analizzato la malattia. Le persone down hanno tre cromosomi 21 (trisomia 21), perché durante la meiosi per la formazione dell’uovo materno i due cromosomi omologhi 21 non si separano e questi due cromosomi 21 materni, uniti al cromosoma 21 dello spermatozoo paterno, portano a tre il numero di cromosomi 21 e a un numero complessivo di 47 cromosomi invece di 46 (fig.).