
I ribosomi e la traduzione del codice genetico
I ribosomi e la traduzione del codice genetico
I ribosomi, particelle costituite da RNA e proteine, sono deputati alla sintesi proteica. Note le strutture primarie e secondarie degli RNA ribosomali (rRNA), sono stati ottenuti dati relativi alloro ripiegamento (folding) tridimensionale ed è stata stabilita l'esatta posizione dei siti di interazione tra le proteine che compongono la subunità piccola del ribosoma e l'rRNA 16S. Numerose prove sperimentali dimostrano che, oltre a possedere un ruolo strutturale e di assemblaggio, gli rRNA partecipano attivamente al compimento della funzione del ribosoma, come risulta anche dall'individuazione di siti di interazione tra l'rRNA e alcuni ligandi. Studi di biochimica e di fisica hanno inoltre evidenziato come il modello classico, ipotizzato per spiegare il meccanismo della traduzione, sia da considerare superato. È molto probabile che in realtà questo sia basato sulla formazione di ibridi molecolari, risultanti dal legame tra l'RNA di trasporto (tRNA) e il ribosoma, che permettono al tRNA di spostarsi indipendentemente rispetto alle due subunità ribosomali.
Introduzione
I ribosomi sono grandi particelle ribonucleoproteiche, cioè formate da RNA e proteine, responsabili della traduzione del codice genetico: un RNA messaggero (mRNA), contenente la sequenza nucleotidica codificante l'informazione relativa a una o più proteine, si lega al ribosoma e ogni codone, costituito da una tripletta di nucleotidi, viene riconosciuto dalla tripletta dell' anticodone appartenente a un corrispondente RNA di trasporto (tRNA). La catena polipeptidica della proteina si assembla mediante la formazione di legami peptidici tra gli amminoacidi che sono legati a tRNA adiacenti nel ribosoma; questa reazione è catalizzata da una regione del ribosoma che possiede attività enzimatica, denominata peptidiltransferasi. L'mRNA e i tRNA si spostano sul ribosoma mediante un processo chiamato traslocazione. Fattori proteici differenti sono necessari in vivo per lo svolgimento delle fasi di inizio, allungamento e terminazione della traduzione. Alcuni di questi fattori sfruttano l'energia derivante dall'idrolisi del GTP. Di notevole importanza sono state numerose osservazioni che hanno dimostrato che i tre passaggi chiave della traduzione (riconoscimento del codone, azione della peptidiltransferasi e traslocazione) sono da ascrivere all'azione del ribosoma stesso; ne consegue che la comprensione dei meccanismi molecolari alla base della traduzione non può prescindere dalla conoscenza della struttura del ribosoma e del modo in cui tale struttura è articolata per esplicare queste tre funzioni. L'architettura molecolare dei ribosomi è vasta e complessa tanto quanto la sequenza dei passaggi molecolari che ne regola l'assemblaggio. Nel corso degli ultimi decenni sono stati fatti grandi progressi nella conoscenza della struttura e della funzione di queste particelle, e questo articolo si prefigge il compito di riassumere i punti fondamentali dell'attuale stato delle conoscenze. Poiché la stragrande maggioranza dei dati sin qui raccolti, relativi alla struttura e alla funzione ribosomale, è stata ottenuta grazie all'utilizzo di ribosomi di Escherichia coli, la discussione riguarderà questi ultimi, tranne nei casi in cui sarà chiaramente specificato.
La prima metà di questo articolo è dedicata alla struttura del ribosoma iniziando, nelle prime due sezioni, dalla discussione delle caratteristiche dei suoi componenti molecolari: le proteine ribosomali (r-proteine) e l'RNA ribosomale (rRNA); una volta esposte le caratteristiche di ripiegamento delle macromolecole che compongono il ribosoma, ne sarà discussa la struttura tridimensionale. Nella terza sezione sono state riassunte le interazioni tra RNA e proteine all'interno del ribosoma. L'architettura ribosomale, così come la conosciamo oggi, è presentata nella quarta sezione che include la morfologia generale delle due subunità del ribosoma di E. coli, la posizione di specifiche proteine e quella di regioni dell'RNA all'interno della struttura stessa. La quinta sezione mette in relazione i dati sulla morfologia, sulla struttura secondaria dell'rRNA e sull'assemblaggio tra r-proteine e rRNA per chiarire come, partendo da approcci differenti, si sia arrivati a un modello della struttura della subunità ribosomale 30S. Nella seconda metà dell'articolo è presentato il processo della traduzione alla luce delle più recenti conoscenze sulla struttura del ribosoma, con particolare riferimento ai tre passaggi che risultano cruciali per la meccanica di tale processo: la selezione accurata dell'amminoacil-tRNA corretto, la traslocazione del tRNA nel ribosoma e il processo catalitico che porta alla formazione del legame peptidico. Come punto di partenza per un maggiore approfondimento degli argomenti che verranno qui trattati, si consiglia il lettore di consultare i numerosi testi classici relativi alla struttura del ribosoma e alla sua funzione (Nomura et al., 1974; Chambliss et al., 1980; Hardesty e Kramer, 1985; Hill et al., 1990).
Proteine ribosomali
Identificazione, purificazione e stechiometria
La complessità strutturale del ribosoma si rese evidente fin dai primi studi effettuati sulle r-proteine. Nel 1964 IP. Waller, mediante elettroforesi su gel d'amido delle proteine della subunità 30S e 50S di E. coli, rilevò la presenza di dozzine di bande proteiche. L'ipotesi che queste corrispondessero ad altrettante singole differenti specie proteiche fu accolta con una certa dose di scetticismo a causa dell'elevata complessità strutturale, senza precedenti fino a quel momento, che ciò avrebbe comportato. l successivi studi di caratterizzazione biochimica e bio fisica, eseguiti sulle proteine purificate, stabilirono infine con certezza che le bande osservate rappresentavano singole specie proteiche differenti per dimensioni, estremità amminoterminali e sequenza amminoacidica (Traut et al., 1967). Attualmente si ritiene che il ribosoma di E. coli contenga circa 52 proteine diverse di cui 21 localizzate nella subunità 30S e circa 31 nella 50S (Wittmann, 1982). l ribosomi eucariotici più grandi e più complessi contengono più di 70 proteine differenti, molte delle quali mostrano una netta omologia con alcune proteine ribosomali batteriche.
La purificazione delle r-proteine è stata solitamente effettuata grazie alla cromato grafia a scambio cationico in urea 6M, seguita da gel-filtrazione (Wittmann, 1974). Poiché le r-proteine sono state isolate indipendentemente in vari laboratori, sono state adottate nomenclature differenti che hanno ulteriormente aumentato la già elevata complessità di questi studi. A un certo punto, la tecnica dell'elettroforesi su gel è stata adottata consensualmente da tutti (Kaltschmidt e Wittmann, 1970) e il suo impiego ha portato alla distinzione delle r-proteine in proteine della subunità piccola (30S), chiamate SI, S2, S3 .... S21 (S dall'inglese small, piccolo) e in proteine della subunità grande (50S), chiamate Ll, L2 .... L36 (L da large, grande). Questo metodo di identificazione delle proteine di E. coli sembrava essere inequivocabile ma in seguito è stato dimostrato che la proteina L8 è, in realtà, un aggregato composto dalle proteine L7/L12 e L10 (Petersson et al., 1976), che la L26 è identica alla S20 (Wittmann et al., 1980) e che le proteine L7 e Ll2 sono identiche se si esclude l'acetilazione amminoterminale (Nterminale) di L7 (Terhorst et al., 1973).
Dopo la purificazione e la caratterizzazione delle proteine, si pose un altro importante quesito: la possibilità che i ribosomi non avessero la stessa composizione proteica ma che, al contrario, esistessero classi differenti di ribosomi dotati di strutture differenti ed eventualmente con funzioni differenti. La stechiometria delle r-proteine è difficile da determinare a causa dell'elevata facilità con cui queste si dissociano dalle particelle durante i processi di purificazione del ribosoma, in particolare se in presenza di elevate concentrazioni di sale. Oggi si è generalmente concordi nel ritenere che tutte le rproteine e le molecole di RNA sono presenti in un rapporto stechiometrico di 1:1, tranne nel caso della proteina L7/L12 di cui sono presenti 4 copie per ogni ribosoma (Hardy, 1975; Subramanian, 1975). Ciò implica che tutti i ribosomi funzionali hanno strutture equivalenti e che, probabilmente, non esistono 'ribosomi specializzati' in E. coli. Studi di ricostituzione in vitro (Held et al., 1973) avvalorano questa ipotesi: le proteine che legano l'RNA raggiungono la saturazione di legame quando il loro rapporto stechiometrico, rispetto alI'RNA stesso, è di 1: 1 (Zimmermann, 1980). Inoltre, esperimenti di sottrazione di singole proteine, durante il processo di ricostituzione, hanno dimostrato che la mancanza di una singola proteina ha effetto sull'assemblaggio o sull'attività di tutti i ribosomi piuttosto che di una specifica frazione di essi (Nomura et al., 1969; Held et al., 1973).
Caratteristiche delle proteine ribosomali
È stata determinata la sequenza di tutte le proteine ribosomali di E. coli, o per sequenziamento diretto delle proteine stesse, o per deduzione dalla sequenza dei geni corrispondenti, o grazie a entrambi i metodi.
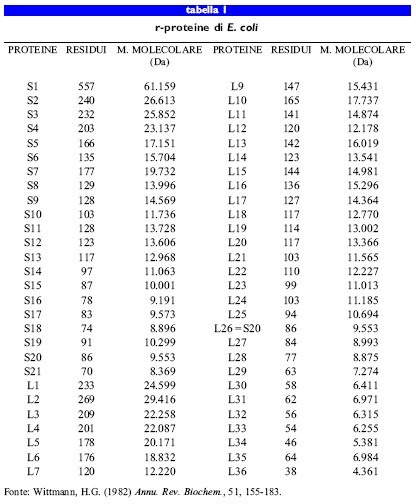
La tabella (tab. I) riporta un confronto tra le masse molecolari delle r-proteine, il cui valore medio è di 14,8 kDa. Oltre alle ridotte dimensioni, queste proteine sono generalmente alquanto basi che, a causa della presenza di un 20% di residui di lisina e di arginina, come del resto ci si aspetta da proteine coinvolte nella formazione di complessi stabili con l'RNA. Fanno eccezione la r-proteina SI, la cui massa molecolare è di 61 kDa, e la L7/Ll2, con massa molecolare di 12 kDa, che sono piuttosto acide e hanno un ruolo rilevante nel processo della traduzione.
Una volta ottenute tutte le sequenze delle 52 r-proteine, la complessità della struttura del ribosoma è diventata ancora più evidente: ricerche effettuate con l'ausilio di elaboratori hanno mostrato che le omologie di sequenza tra le diverse r-proteine sono sorprendentemente basse (Wittmann e Wittmann-Liebold, 1974; Wittmann et al., 1980). Sebbene sia difficile escludere la possibilità di omologie non rilevabili delle sequenze proteiche, questo risultato sembra suggerire un'origine evolutivamente indipendente per ciascuna delle r-proteine o, al massimo, una divergenza evolutiva molto precoce a partire da un progenitore comune. Questo punto di vista è in accordo con l'ipotesi che il ribosoma si sarebbe originato da una struttura composta principalmente, se non esclusivamente, da RNA (Noller e W oese, 1981; Noller 1993; v. oltre). È facile a questo punto supporre il processo graduale con il quale, nel corso dell'evoluzione, si sarebbero aggiunte le singole r-proteine migliorando progressivamente l'efficacia dei meccanismi di assemblaggio e traduzione. Se questo rappresenta il modo di evolvere del ribosoma, non ci dobbiamo necessariamente aspettare di trovare a priori un'omologia di sequenza tra le differenti r-proteine.
Per quanto riguarda le forme delle r-proteine, alcuni studi hanno messo in evidenza strutture prevalentemente allungate, altri forme più compatte e globulari (Liljas, 1982). Questo è un problema di difficile soluzione poiché la struttura che conta è quella della r-proteina nella sua forma funzionalmente attiva, cioè quella che la r-proteina ha quando è inserita nel complesso ribosomale. Alla luce di questo si capisce bene come perfrno le strutture ottenute per analisi ai raggi X dei cristalli proteici (v. oltre) possano far pensare che la proteina, una volta assemblata nel ribosoma, possa avere una conformazione diversa. Non essendo possibile ottenere immagini ad alta risoluzione delle strutture cristalline dei ribosomi stessi, l'unica tecnica che permette di avvicinarsi al problema in modo più diretto è lo studio di scattering neutronico. V. Ramakrishnan e M. Capel, con altri ricercatori, hanno per primi determinato i raggi di girazione di numerose proteine della subunità ribosomale 30S (Ramakrishnan et al., 1984). Anche se l'interpretazione dei dati ottenuti è notevolmente limitata dall'incertezza che circonda le misurazioni effettuate, i risultati ottenuti per molti raggi di girazione sembrano far propendere per una forma globulare delle r-proteine anche se, per le proteine SI, S4 e S8, i dati suggeriscono una struttura più allungata. Una migliore comprensione della struttura delle r-proteine sta ora emergendo dagli studi di cristallo grafia a raggi X.
Struttura tridimensionale delle proteine ribosomali purificate
Come sarà esposto più avanti, le numerose difficoltà di natura tecnica, dovute alle notevoli dimensioni e alla complessità strutturale del ribosoma, hanno frenato sensibilmente l'analisi cristallo grafica della struttura del ribosoma intatto. Inoltre, non pochi sono stati i problemi incontrati nel tentativo di ottenere cristalli di singole r-proteine, sebbene quelle dei batteri termofili siano risultate particolarmente adatte alla cristallizzazione. Finora sono state determinate le strutture cristallografiche di 6 proteine ribosomali: S5 (Ramakrishnan e White, 1992), S6 (Lindhal et al., 1994), L6 (Golden et al., 1993), L7/L12 (Leijonmarck et al., 1980; 1988), L9 (Hoffman et al., 1994), L30 (Wilson et al., 1986; Leijonmarck et al., 1988). Inoltre, la struttura della proteina S 17 è stata ottenuta mediante spettroscopia NMR (Golden et al., 1993). Sebbene la maggior parte delle proteine mostri una forma compatta e globulare, due di esse (L7/L12 e L9) presentano una struttura decisamente allungata, costituita da due domini essenzialmente globulari, tenuti in connessione da un sottile filamento. l domini globulari sono costituiti principalmente da semplici strutture α e β che somigliano ai domini proteici coinvolti nel legame con l'RNA, appartenenti a proteine con funzione analoga (Hoffman et al., 1994). In particolare si ritiene che queste proteine interagiscano con l'RNA mediante la superficie di un foglietto β, che spesso presenta gruppi laterali aromatici in grado di prendere contatti con le basi dell'RNA (Oubridge et al., 1994; Valgard et al., 1994).
RNA ribosomale
L'RNA costituisce più del 60% dei ribosomi batterici: il fatto che, diversamente da altre polimerasi cellulari, i ribosomi contengano RNA come maggior componente strutturale è, probabilmente, un importante indizio relativo alla loro evoluzione e alloro meccanismo d'azione. Attualmente numerose prove rafforzano la tesi secondo la quale l'RNA avrebbe, nel processo di traduzione, un ruolo funzionale anziché di semplice supporto strutturale (Noller, 1991). Tre specie di rRNA fanno parte integrante, come componenti strutturali, dei ribosomi procariotici: la subunità 30S contiene l'rRNA l6S, mentre il 5S e il 23S fanno parte della subunità 50S. Sono note le rispettive strutture primarie e secondarie delle tre molecole e sono stati identificati molti siti di interazione con le r-proteine e con alcuni ligandi funzionali; non è stato scoperto molto, invece, circa i ripiegamenti che formano la struttura terziaria.
Strutture primarie
L'rRNA 5S, il più piccolo dei tre, è stata la prima molecola di RNA a essere sequenziata attraverso l'utilizzo di radioisotopi (Brownlee et al., 1967): è composta da 120 nucleotidi e non è soggetta a modificazioni postrascrizionali. Per ottenere la sequenza completa dell'rRNA l6S, è stata necessaria una ricerca decennale che ha utilizzato il sequenziamento del DNA del corrispondente gene clonato (Brosius et al., 1978) e il sequenziamento diretto dell'RNA (Carbon et al., 1979). Tale filamento è composto da 1542 nucleotidi, dieci dei quali (corrispondenti alle posizioni 527, 966, 967, 1207, 1402, 1407, 1498, 1516, 1517 e 1518) sono metilati e uno è costituito da una pseudouridina (Bakin et al., 1994). Analisi della conservazione, durante la filogenesi, della sequenza dell'rRNA l6S hanno fatto luce sul probabile ruolo biologico delle diverse regioni della molecola (Woese et al., 1983; Gutell et al., 1985); la presenza di modificazioni postrascrizionali in regioni la cui sequenza si è conservata indica che queste hanno un importante ruolo funzionale. Comunque, la presenza di differenze caratteristiche tra le tre maggiori linee evolutive dovrebbe implicare un loro coinvolgimento in un meccanismo di 'regolazione fine' di caratteristiche strutturali cruciali (Noller e W oese, 1981): in alcuni casi, la presenza o meno di gruppi metilici determina la resistenza ad alcuni antibiotici.
L'rRNA 23S di E. coli è costituito da 2904 nucleotidi (Brosius et al., 1980) e, come l'rRNA l6S, contiene numerosi nucleotidi soggetti a modificazioni postrascrizionali, come la presenza di pseudouridine e di metilazioni; molte delle posizioni modificate, ma non tutte, sono state localizzate nella sequenza (Bakin e Ofengand, 1993). L'omologia di sequenza tra i tre rRNA è scarsa o completamente assente, né è stata rilevata alcuna omologia, tranne quella casuale, tra regioni differenti di uno stesso rRNA: ciò avvalora l'ipotesi di un' origine evolutiva indipendente per i tre diversi rRNA (Noller, 1993). L'rRNA non presenta una simmetria strutturale rilevante, quale la ripetizione di motivi strutturali primari o secondari e, come vedremo in seguito, questo fatto sembra estendersi anche a livello dell'organizzazione strutturale di ordine superiore.
Strutture secondarie
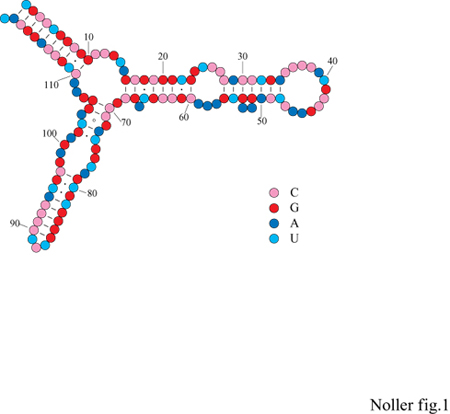
Nei dieci anni che seguirono la pubblicazione della struttura primaria dell'rRNA 5S, furono proposte, per questa molecola, dozzine di modelli di possibili strutture secondarie spesso molto diversi tra di loro, malgrado la disponibilità di dati biochimici e fisici rilevanti, nonché la conoscenza delle regole per il calcolo dell'energia libera che permettono di valutare la stabilità dell'elica di RNA. Si arrivò alla soluzione del problema quando si comprese che, come nel caso dei tRNA, anche gli rRNA 5S, provenienti da specie diverse, hanno probabilmente strutture secondarie piuttosto simili, pur con sostanziali differenze a livello della sequenza. G.E. Fox e C.R. Woese (1975) sono stati i primi a effettuare analisi comparative delle sequenze per dedurre la struttura secondaria dell'rRNA 5S (fig. 1). l suoi quattro principali elementi a elica sono tipiche strutture degli rRNA più grandi (16S e 23S), e comprendono estroflessioni di una o più coppie di basi ed eliche composte, unite da anse interne, tutte strutture che non erano mai state osservate nella struttura secondaria dei tRNA.
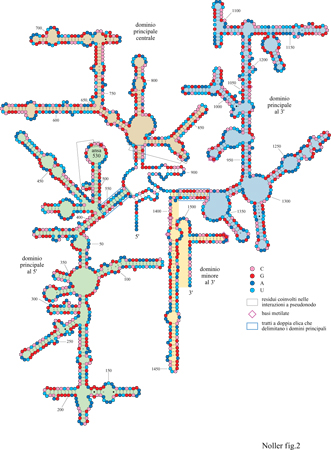
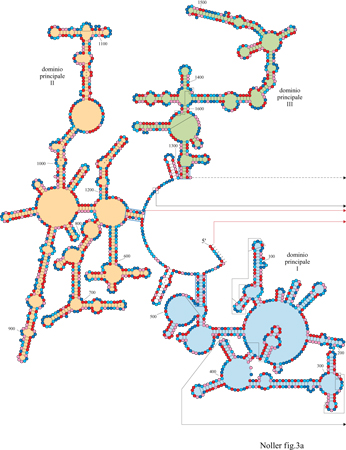
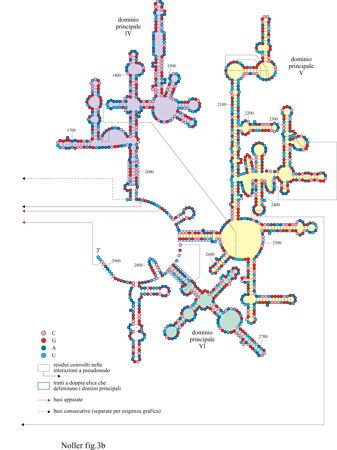
Nelle figure (figg. 2, 3a e 3b) sono mostrate le strutture secondarie degli rRNA l6S e 23S (Noller et al., 1981; Noller e Woese, 1981; Woese etal., 1983; Gutell e Woese, 1990; Gutell, 1994; Gutell et al., 1994).
L'esistenza di ogni struttura a elica è avvalorata da differenti approcci sperimentali, ma le strutture nel loro insieme sono state ricavate da analisi comparative delle sequenze. Significativamente, ogni singola struttura a elica degli rRNA l6S e 23S che compare nelle prime versioni pubblicate, risultante dall'analisi comparativa delle sequenze (Noller et al., 1981; Noller e Woese, 1981) è considerata ancora valida poiché, oltre a essere suffragata da numerose prove sperimentali, è stata confermata da più di 4000 nuove sequenze complete o quasi di rRNA l6S e da più di un centinaio di sequenze di rRNA 23S; ci sono quindi scarse possibilità che si possano individuare ulteriori appaiamenti tra le basi.
La struttura secondaria dell'rRNA l6S è suddivisa in quattro domini strutturali, tre principali e uno minore, che si formano da tre appaiamenti a lunga distanza (v. figura 2). Il dominio principale al 5' (approssimativamente dal residuo 26 al 557) è definito dall'elica 27-37/547-556, cioè formata dai residui 27-37 e da quelli 547-556; il dominio principale centrale (approssimativamente dal 564 a19l2) dall'elica 567569/881-883. Dei due domini a13', il principale (approssimativamente dal 926 al13 91) e il minore (approssimativamente dal 1392 al 1542) sono definiti dall'elica 923-933/1384-1393. Per comprendere l'architettura ribosomale è importante valutare fino a che punto questi domini, definiti finora solo in termini di struttura secondaria, corrispondono ai reali domini strutturali: come sarà descritto più avanti, in occasione dei primi studi sui siti di legame delle r-proteine, è stato possibile isolare alcuni frammenti di rRNA l6S corrispondenti precisamente ai tre domini principali. La capacità dimostrata da alcune r-proteine di rimanere legate a questi frammenti di RNA o, perfino, di legarvisi nuovamente dopo esserne state rimosse (Yuki e Brimacombe, 1975; Zimmermann et al., 1975; Muller et al., 1979; Ehresmann et al., 1981), avvalora l'ipotesi che questi domini corrispondano a entità strutturali realmente esistenti. L'assemblaggio dei singoli domini di RNA con proteine specifiche fornisce un'ulteriore conferma dell'esistenza dei domini strutturali dell'rRNA (Weitzmann et al., 1993; Samaha et al., 1995); comunque, ciò non implica necessariamente che non ci possano essere interazioni strutturali significative tra i domini stessi.
All'interno di ogni dominio la struttura presenta una serie di eliche semplici e composte, separate da numerose anse con più rami laterali, anse interne ed estroflessioni. Contrariamente alle attese, non esistono eliche di rRNA lunghe e regolari, ma la struttura si presenta costituita dall'unione di numerose eliche di piccole dimensioni le cui giunzioni tendono a formare parecchie irregolarità strutturali. Le ragioni di un'architettura così irregolare possono essere molteplici: la presenza di numerose eliche piccole permette la formazione di una struttura tridimensionale più complessa che potrebbe avvicinarsi a quella di una proteina globulare (Noller e Woese, 1981). In secondo luogo, la presenza di eliche lunghe ed estremamente stabili potrebbe intrappolare la molecola di RNA in uno stato conformazionale con un livello di energia libera locale così basso da ostacolare il processo che porta all'assemblaggio della corretta struttura finale. Infine, una struttura di questo tipo permetterebbe l'accesso al solco maggiore dell'elica dell'rRNA da parte delle r-proteine (Weeks e Crothers, 1993; v. oltre).
La figura (v. figura 3a e 3b) mostra la struttura secondaria dell 'rRNA 23 S di E. coli (Noller et al., 1981; Gutell et al., 1993; 1994): come nell'rRNA l6S, appaiamenti di basi a lunga distanza dividono la catena in domini strutturali facilmente identificabili. Se ne individuano sei principali: il dominio l (16-25/515-524), il dominio II (579-584/12561261), il dominio III (1295-1298/1642-1645), il dominio IV (1656-1663/1997-2004), il dominio V (2043-2054/ 2615-2625) e il dominio VI (2630-2637/2781-2788). Questi domini partono da un'ansa centrale creata dall'appaiamento delle estremità 5' e 3' della molecola. Grazie all'analisi comparativa delle sequenze, sono state identificate numerose interazioni a pseudonodo (Gutell et al., 1985; Leffers et al., 1987), di cui almeno due nell'rRNA l6S coinvolgono, nella formazione di legami, i residui 505-507 e 524-526 nel dominio al 5' e i residui 570-571 e 865-867 nel dominio centrale. Il primo dei due pseudonodi, quello formato dai residui 505-507 e 524-526, è coinvolto nella formazione di una delle strutture più conservate dell'rRNA: l'ansa 530, la cui esistenza è stata confermata direttamente anche da studi di genetica (Powers e Noller, 1991). Mutazioni nei residui appartenenti allo pseudonodo dell'ansa 530 causano perturbazioni funzionali dei ribosomi mutati, indicando che questa struttura svolge un ruolo centrale nell'accuratezza del processo di traduzione. Nell'rRNA 23S sono stati individuati 6 pseudonodi, di cui uno coinvolge l'appaiamento delle basi in posizione 413-416 nel dominio l con quelle in posizione 2407-2410 del dominio V, trattandosi, quindi, di una interazione tra due anse molto distanti che fornisce un solido vincolo al ripiegamento dell'rRNA 23S. Anche l'analisi comparativa delle sequenze ha permesso di individuare interazioni tra singole basi a livello della struttura terziaria, ma con tutta probabilità è stata finora identificata solo una frazione di questo tipo di interazioni. Tuttavia, il fatto che si verifichino appaiamenti non canonici tra le basi, come A-G, A-C, C-C e U-U, insieme alle già menzionate interazioni a pseudonodo, indica che l'rRNA ha una struttura di ordine superiore molto complessa, con caratteristiche strutturali ancora da identificare (Gutell e Woese, 1990).
Evidenze sperimentali di strutture di ordine superiore
Un'analisi comparativa fornisce prove convincenti circa l'esistenza in vivo di elementi a elica e anche di singoli appaiamenti tra le basi. Tuttavia, i ribosomi potrebbero essere intesi come strutture dinamiche in cui, durante l'assemblaggio della particella o la sintesi proteica, si verifica l'interconversione tra conformazioni differenti di rRNA. Perciò, gli approcci sperimentali che forniscono informazioni circa la conformazione dell'rRNA non solo servono a confermare la struttura secondaria, ma possono servire a indicare se tutti gli elementi a elica, individuati con l'analisi comparativa, coesistono allo stesso tempo. Attualmente i risultati mostrano che tutte le eliche individuate per confronto filo genetico sono realmente presenti nello stesso momento nel ribosoma e, finora, non si hanno prove convincenti di eliche reciprocamente incompatibili, se si eccettua la competizione esistente tra l'elica presente all'estremità 5' dell'rRNA l6S e un'elica presente nell'RNA precursore che hanno un filamento in comune (Dammel e Noller, 1993). Questa seconda elica si forma presumibilmente solo durante la maturazione della subunità ribosomale 30S.
Lo studio della conformazione dell'rRNA è stato affrontato con una vasta gamma di tecniche sperimentali che comprendono: a) l'uso di sonde chimiche ed enzimatiche, comprese quelle che provocano la formazione di legami crociati, specifiche per il singolo e il doppio filamento; b) l'analisi tramite elettroforesi bidimensionale su gel, in cui frammenti di RNA ottenuti per digestione parziale con nucleasi, associati per complementarità durante la corsa elettroforetica nella prima dimensione, si separano nelle condizioni denaturanti durante la corsa nella seconda dimensione; c) l'uso di oligonucleotidi come sonde molecolari per individuare regioni di rRNA non appaiate tra loro; d) la risonanza magnetica nucleare (NMR).
l nucleotidi appaiati non dovrebbero mostrare re attività quando vengono saggiati con sonde chimiche, specifiche per il filamento singolo. Non è comunque sempre vero il contrario, poiché i nucleotidi che, dall'analisi della struttura secondaria, risultano non appaiati possono resistere all'attacco da parte delle sonde chimiche a causa delle interazioni dovute alla struttura terziaria, a interazioni con proteine, o con ioni come il magnesio, o essere coinvolti in interazioni del tipo sovrapposizione delle basi (stacking), come nel caso del dietilpirocarbonato. Con le attuali metodologie è possibile saggiare rapidamente e sistematicamente con sonde chimiche, virtualmente, tutte le basi degli rRNA 16S e 23S. Secondo un altro approccio, i ribosomi e l'rRNA vengono saggiati con una serie di sonde chimiche, specifiche per il singolo filamento, che modificano tutte e quattro le basi quando sono impegnate in appaiamenti Watson-Crick (Moazed et al., 1986). I siti modificati chimicamente vengono quindi individuati tramite la tecnica primer extension (estensione di un oligonucleotide usato come innesco della reazione di polimerizzazione da parte della trascrittasi inversa) in cui l'oligonucleotide si ferma, momentaneamente o definitivamente, in corrispondenza delle basi modificate. I risultati così ottenuti sull'rRNA 16S nudo (cioè privo di proteine) sono quasi totalmente in accordo con la struttura descritta in figura (v. figura 2), e forniscono una prova ulteriore sull'affidabilità dell'analisi comparativa (Moazed et al., 1986). In altri casi, le discrepanze tra i dati ottenuti utilizzando le sonde e la struttura secondaria dedotta sono state risolte impiegando le sonde suIl'rRNA 16S quando è ancora assemblato alla subunità 30S. Perciò, alcuni aspetti della conformazione dell'RNA, ritenuti determinanti da un punto di vista biologico (individuati solo grazie all'impiego del metodo comparativo), sembrano dipendere dalle interazioni dell'rRNA 16S con le r-proteine della subunità 30S.
Infine, alcuni dati ottenuti con l'impiego di sonde chimiche, sembrano indicare l'esistenza di interazioni di tipo terziario e quaternario, e non possono essere spiegati solo sulla base della struttura secondaria. In alcuni casi, i risultati suggeriscono la presenza di interazioni di ordine superiore predette dall'analisi comparativa delle sequenze.
P.E. Moore e collaboratori (Szewczak et al., 1993), per mezzo di NMR ad alta risoluzione, hanno risolto la struttura tridimensionale dell'ansa ex-sarcina dell'rRNA 23S mostrando che questa regione, composta da un'ansa di 15 nucleotidi molto conservati, è coinvolta in una struttura molto compatta, come indicato dalla bassa re attività delle sue basi nei confronti di sonde chimiche (Moazed et al., 1988). Questa regione è caratterizzata da appaiamenti non canonici tra le basi e da legami tra basi e gruppi fosfato, e rappresenta una delle complessità strutturali dell'rRNA ancora da chiarire.
Interazioni tra RNA e proteine
La comprensione dei processi alla base del riconoscimento tra RNA e proteine è oggi in una fase di rapido sviluppo. Sebbene numerosi meccanismi che regolano le interazioni DNA-proteine siano noti, è presumibile che il riconoscimento RNA-proteine risulterà diverso per molti aspetti fondamentali. Il DNA infatti, a meno di rare eccezioni, ha una semplice struttura a doppia elica, e il riconoscimento di un particolare sito da parte di una proteina dipende, quindi, esclusivamente da un riconoscimento di sequenza o, in un numero ridotto di casi, da variazioni minime che la sequenza può produrre sulla geometria dell'elica. Al contrario, la struttura dell 'RNA si presenta estremamente complessa: non sono presenti solo tratti a elica, ma anche estroflessioni, anse interne, a forcina, a più rami laterali, pseudonodi e appaiamenti non canonici che, attraverso complesse interazioni terziarie, hanno la capacità di formare una struttura globulare e compatta, come può essere quella di un tRNA (Kim et al., 1974). Inoltre, le eliche di RNA presentano quasi sempre una struttura di tipo A o A' in cui il solco maggiore tra le due eliche, risultando stretto e profondo, impedisce l'ingresso di una struttura proteica ad ex-elica, modo con il quale sembra che le sequenze di DNA siano riconosciute dalle proteine. Tuttavia, studi recenti indicano che alcune irregolarità strutturali dell'RNA, quali le estroflessioni e le anse interne, possono facilitare l'accesso alle basi nel solco maggiore delle eliche (Weeks e Crothers, 1993). È rilevante il fatto che queste irregolarità sono distribuite lungo tutta la struttura dell'rRNA e che, quindi, parte della loro funzione può consistere nel permettere il riconoscimento del solco maggiore da parte delle r-proteine.
I ribosomi costituiscono una fonte importante di sistemi modello per lo studio delle interazioni specifiche tra RNA e proteine. Tali studi non solo risultano determinanti per la comprensione dei meccanismi che regolano l'assemblaggio del ribosoma ma contribuiscono a chiarire il problema generale del riconoscimento tra RNA e proteine. Sono stati localizzati numerosi siti di legame delle proteine sull'rRNA ed è utile riassumere qui brevemente le indicazioni più salienti di tali ricerche.
Lo studio del meccanismo di legame delle r-proteine all'rRNA ha avuto inizio con la scoperta del metodo di ricostituzione del ribosoma in vitro (Traub e Nomura, 1969). Il fatto che si ottenessero particelle del tutto funzionali ha fatto supporre che tutte le interazioni molecolari osservate in questo sistema fossero da ritenersi autentiche. Studi compiuti sull'assemblaggio del ribosoma utilizzando singole r-proteine purificate rivelarono che solo una frazione delle proteine della subunità 30S (le proteine che si legano per prime) aveva, in assenza delle altre, la capacità di legarsi indipendentemente all'rRNA 16S. I criteri che indicano la specificità del legame (Zimmermann, 1974) sono:
a) una data proteina dovrebbe legarsi a una sola specie di RNA (per esempio, all'rRNA l6S ma non al 23S);
b) in presenza di un eccesso di proteina il rapporto molare tra proteina ed RNA dovrebbe raggiungere la saturazione in corrispondenza di un rapporto stechiometrico (per esempio, 1:1);
c) l'interazione di una proteina con l'RNA non dovrebbe impedire il legame di un'altra;
d) una proteina già assemblata all'RNA dovrebbe essere incorporata nella particella ribosomale matura senza che l'aggiunta delle proteine rimanenti comporti la sua dissociazione dall'RNA. Seguendo questi criteri si è arrivati alla conclusione che le proteine S4, S7, S8, S15, S17 e S20 si legano direttamente e indipendentemente all'rRNA l6S; Ll, L2, L3, L4, (Ll2k LlO, Lll, Ll5, L20, L23 e L24 all'rRNA 23S e L5, Ll8 e L25 all'rRNA 5S. La localizzazione dei siti di legame di queste proteine sull'RNA è stata effettuata con numerose tecniche differenti:
a) il metodo classico bind and chew (lega e mastica) che si basa su una parziale digestione con un enzima nucleolitico dei complessi proteina-RNA, seguita da una purificazione delle ribonucleoparticelle (RNP) contenenti frammenti dell'RNA (Zimmermann, 1980); la capacità della proteina di legarsi nuovamente al frammento di RNA purificato ha fornito, in molti casi, la conferma che parte del suo sito di legame è contenuto nel frammento di RNA isolato. Questo metodo, pur costituendo un approccio generalmente valido per studi preliminari, si è in seguito mostrato non privo di difetti. In alcuni casi, infatti, si isolavano frammenti di RNA non digerito anche in assenza di proteine legate, suggerendo che fosse la struttura assunta dall'RNA stesso, più che il legame con la proteina, a stabilizzare il frammento protetto (Garrett et al., 1977). Al contrario, determinati siti di legame dell 'RNA possono essere intrinsecamente instabili e non superare il trattamento con nucleasi, e quindi non possono essere identificati con questo metodo.
b) La protezione, dall'attacco chimico o enzimatico, da parte di proteine legate all'RNA (jootprinting): questa tecnica è stata utilizzata soprattutto per lo studio dell'rRNA l6S (Stern et al., 1989; Powers e Noller 1995). Un'ambiguità intrinseca a questo metodo consiste nel fatto che i cambiamenti conformazionali indotti nell'RNA dalla proteina possono essere essi stessi causa di protezione.
c) Gli esperimenti di damage-selection (danno e selezione) con cui è possibile identificare i siti dell'RNA la cui modificazione chimica interferisce con il legame della proteina (Peattie et al., 1981).
d) La formazione di legami crociati, indotta da agenti chimici o da raggi ultravioletti, tra proteine e RNA indica i siti dell 'RNA a contatto con le proteine, anche se in alcuni casi si può verificare la formazione di legami crociati anche con siti non direttamente coinvolti nel legame.
e) L'utilizzo della tecnica di microscopia elettronica che permette di osservare i complessi RNA-proteine.
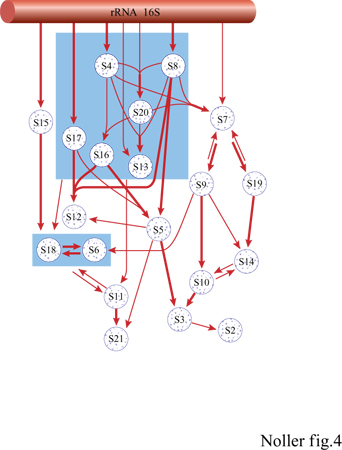
f) L'impiego della mutagenesi sito-specifica, effettuata su geni di rRNA precedentemente clonati, permette di individuare le sequenze e i motivi strutturali necessari per il riconoscimento della proteina. L'approccio mutagenico presenta però, come difetto intrinseco, la possibilità che l'RNA mutato possa ripiegarsi in maniera non corretta in modo da provocare un mancato legame della proteina, dovuto indirettamente agli effetti prodotti dalla mutazione sulla struttura dell'RNA stesso. Le regioni dell'rRNA l6S che costituiscono i siti di legame delle proteine, identificate grazie alla tecnica bind and chew corrispondono, sostanzialmente, ai domini e subdomini della struttura dell 'RNA. Questa osservazione conferma l'esistenza di questi domini, che erano stati ipotizzati sulla base di analisi comparative delle sequenze, come reali entità fisiche. La loro capacità di legarsi nuovamente alle proteine indica, in molti casi, un certo grado di autonomia strutturale all'interno del ribosoma, come dimostrato da esperimenti di ricostituzione in vitro di domini strutturali indipendenti, partendo da interi domini di rRNA l6S (Weitzmann et al., 1993; Samaha et al., 1995). La strategia del jootprinting è stata invece utilizzata per individuare i siti di interazione delle proteine della subunità 30S sull'rRNA l6S (Stern et al., 1989; Powers e Noller 1995): viene effettuata facendo assemblare in vitro, attraverso passaggi successivi, le proteine all'RNA basandosi sulla mappa di assemblaggio specifica per la 30S (fig. 4). A ogni passaggio, l'RNA appartenente al complesso è stato ibridato con una serie di sonde chimiche, specifiche per ciascuna delle quattro basi, individuando poi quelle protette dal legame con le singole proteine. È necessario ricordare che alcune protezioni che si ottengono con l'aggiunta delle proteine potrebbero, in realtà, essere determinate da interazioni indirette, causate da cambiamenti conformazionali dell'RNA indotti dall'assemblaggio delle singole proteine. Le basi che risultano protette, quindi, potrebbero in realtà essere lontane dal sito di legame della proteina. Finora comunque, c'è una stretta corrispondenza tra i risultati ottenuti con questo metodo e quelli ottenuti in esperimenti di legame crociato (come sarà discusso più avanti). Le tecniche che utilizzano la formazione di legami crociati indotti da agenti chimici per individuare, nel ribosoma, i punti di contatto tra proteine e RNA, e tra differenti regioni dell 'rRNA, sono state sfruttate da molti gruppi di ricerca, tra cui quello diretto da R. Brimacombe (Zwieb e Brimacombe, 1979; Wower e Brimacombe 1983; Atmadja et al., 1986; Kyriatsoulis et al., 1986; Stiege et al., 1986; Glotz et al., 1987; Osswald et al., 1987). Spesso usando due differenti approcci, cross-linking e jootprinting, si è comunque giunti alla formulazione di modelli tra loro globalmente congruenti.
Architettura dei ribosomi e delle loro subunità
Morlologia generale delle subunità ribosomali
Le informazioni strutturali che concernono i singoli componenti molecolari, in modo particolare gli rRNA, forniscono dati significativi sull' organizzazione strutturale complessiva del ribosoma di E. coli. Allo stesso tempo, studi di carattere fisico hanno fornito informazioni sulla morfologia delle subunità ribosomali e sulla localizzazione interna di elementi strutturali specifici. Le nostre attuali conoscenze sulla forma dei ribosomi derivano soprattutto da studi di microscopia elettronica e di scattering neutronico.
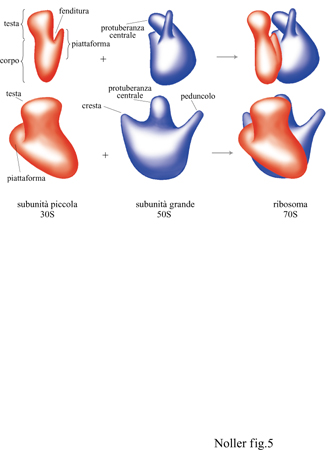
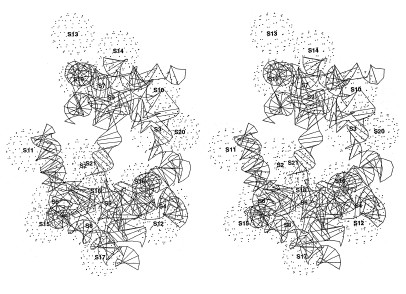
Molti gruppi di ricercatori hanno studiato la struttura delle subunità ribosomali utilizzando la microscopia elettronica (Boublik et al., 1985; Vasiliev et al., 1985; Oakes et al., 1990; Stöffler-Meilecke e Stöffler, 1990; Frank et al., 1991). Sebbene i dettagli dei modelli proposti dai vari gruppi differiscano per l'interpretazione dei dati e per la metodologia applicata, la loro rappresentazione coincide largamente con quella dei modelli delle subunità 30S e 50S descritti da J.A. Lake nel 1980 (fig. 5). Inoltre, utilizzando tecniche di ricostruzione d'immagine in combinazione con la microscopia elettronica, si sono ottenute strutture medie computerizzate del ribosoma e delle sue subunità. Una di queste ricostruzioni per il ribosoma 70S di E. coli, ottenuta da J. Frank e collaboratori (Frank et al., 1991; Penczeck et al., 1992), è mostrata nella figura (fig. 6).
La subunità 30S (v. figura 5) è costituita da un 'corpo' che comprende circa i due terzi dell'intera massa della particella e da una 'testa' che occupa il rimanente terzo della massa. Una sporgenza tipo 'piattaforma' si estende dal corpo della particella, vicino alla giunzione tra il corpo e la testa; con alcuni metodi di identificazione è anche visibile una 'fenditura' tra la testa e la piattaforma.
Alcuni studiosi hanno evidenziato una seconda struttura simile a una piattaforma sulla subunità 30S, posta sul lato opposto rispetto alla piattaforma principale (Stöffler-Meilecke e Stöffler, 1990; Frank et al., 1991). La lunghezza della struttura è di circa 240 ÷ 250 Å. La subunità 50S ha una forma più emisferica o globulare (v. figura 5) e contiene tre strutture che si proiettano all'esterno in maniera più evidente: il 'peduncolo' L7/Ll2 sulla destra; una 'protuberanza centrale' nel mezzo; la 'cresta' LI sulla sinistra. I ricercatori sono generalmente concordi per quanto riguarda la forma della subunità 50S, la cui larghezza massima è stimata intorno a 240 ÷ 250 Å. L'orientamento relativo delle subunità 30S e 50S nel ribosoma 70S è stato anch'esso oggetto di dibattito (Prince et al., 1982), anche se vi è un consenso generale nel ritenere valido l'arrangiamento proposto inizialmente da J.A. Lake (1982). In questo modello (v. figura 5), il lato della piattaforma della subunità 30S è situato di fronte al lato concavo della subunità 50S, con la testa della subunità 30S posta tra la cresta LI e la protuberanza centrale. Questa ipotesi di orientamento è confermata da esperimenti di marcatura semplice o doppia dei ribosomi 70S, mediante anticorpi.
Le tecniche utilizzate per lo studio della forma dei ribosomi e delle loro subunità in soluzione sono la cristallo grafia a raggi X e lo scattering neutronico (Moore, 1980; Liljas, 1982). Sebbene sia molto più difficile interpretare i risultati degli esperimenti di scattering in soluzione, rispetto a quelli di microscopia elettronica, i primi sono comunque molto importanti poiché misurano le proprietà di particelle completamente idratate in condizioni che si avvicinano di più a quelle fisiologiche. È molto positivo, quindi, che il modello per la subunità 50S, ricavato da dati di scattering neutronico (Stuhrmann et al., 1977) sia molto simile a quello ottenuto per mezzo della microscopia elettronica. Informazioni relative alla distribuzione spaziale dell'RNA e delle proteine possono essere ottenute misurando i raggi di girazione dei corrispondenti aggregati in esperimenti che utilizzano la tecnica del contrasto di solvente (contrast-matching). Attualmente c'è un accordo generale nel ritenere che le distribuzioni delle proteine e dell'RNA, nella subunità 50S, abbiano centri di massa all'incirca coincidenti, ma che i componenti proteici presentino un raggio di girazione (90-100 Å) maggiore di quello dell'RNA (55-60 Å) (Crichton et al., 1977). Anche nella subunità 30S, la distribuzione di proteine e RNA risulta all'incirca concentrica, con una maggiore concentrazione proteica in superficie; i raggi di girazione sono pari a circa 78 Å per le proteine e a 61 Å per l'RNA (Damaschun et al., 1979).
l dettagli strutturali delle macromolecole vengono risolti tradizionalmente per mezzo della cristallo grafia a raggi X, ma sfortunatamente i ribosomi, in particolare quelli di E. coli, sono risultati difficili da cristallizzare. Tuttavia, negli ultimi anni si è registrato un progresso significativo nella cristallizzazione di subunità ribosomali ricavate da batteri alofili (Glotz et al., 1987; von Bohlen et al., 1991) e termofili (Mussig et al., 1989; Trakhanov et al., 1989; Yusupov et al., 1991). Recentemente A. Yonath e collaboratori (von Bohlen et al., 1991) hanno ottenuto cristalli delle subunità 50S da Haloarcula marismortui che diffrangono a valori minori di 3 Å. Per trarre informazioni da questi cristalli sarà probabilmente necessario disporre di cristalli isomorfi contenenti gruppi di atomi pesanti in posizioni specifiche delle subunità: e ciò è un obiettivo difficile da raggiungere. Tuttavia, riuscire ad avere una risoluzione della struttura molecolare della subunità ribosomale 50S vicina a quella atomica giustificherà, in larga misura, i molti anni spesi nel lavoro sperimentale.
Localizzazione delle proteine e dell'RNA nei ribosomi
Durante gli anni Ottanta, molto si è appreso sulla distribuzione spaziale delle proteine ribosomali nel ribosoma e sono stati assai determinanti, a tal riguardo, i risultati ottenuti con l'immunomicroscopia elettronica, (lEM, Immuno-Electron Microscopy) e con gli studi di diffrazione neutronica (Capel et al., 1987; Oakes et al., 1990; Stöffler-Meilecke e Stöffler, 1990). Posizionare l'RNA nel ribosoma rappresenta un problema di tipo diverso. A parte la localizzazione dell'rRNA 5S di dimensioni relativamente piccole, quella degli altri rRNA potrebbe, a prima vista, essere considerata un problema banale in quanto la loro massa coincide probabilmente, all'incirca, con quella dell'intera subunità. Tuttavia, l'informazione veramente interessante riguarda la localizzazione nel ribosoma di regioni o siti specifici dell'rRNA perché, in effetti, questo equivale a determinarne alcuni aspetti della struttura terziaria. Tra gli approcci sperimentali importanti per affrontare questo tipo di problema menzioniamo: la lEM (Politz e Glitz, 1977), la mappatura dei siti di legame delle proteine mediante diffrazione neutronica, l'induzione di legami crociati tra RNA-proteina o tra RNA-RNA e, infine, la costruzione di modelli sulla base dei dati riportati sopra e dei vincoli dettati dalle regole stereochimiche.
Immunomicroscopia elettronica
Il principio della lEM si basa sulla possibilità di visualizzare singole molecole anticorpali mediante microscopia elettronica: anticorpi specifici per singole proteine ribosomali o per apteni vengono fatti legare ai ribosomi o alle subunità e successivamente visualizzati (Oakes et al., 1990; Stöffler-Meilecke e Stöffler, 1990; Olah et al., 1993). La posizione occupata dalla proteina all'interno della struttura del ribosoma viene, quindi, dedotta dalla posizione del sito di attacco dell'anticorpo, sulla base di caratteristiche strutturali riconoscibili sulla superficie del ribosoma stesso. La lEM pur essendo in linea di principio un approccio molto diretto, negli anni passati è stata al centro di numerose controversie, per le difficoltà imputabili alla metodica degli esperimenti e all'interpretazione dei dati ottenuti. Fortunatamente, questa situazione è stata in buona parte risolta: con questo metodo è stato possibile determinare la posizione, sulle rispettive subunità, di parecchie r-proteine e spesso essa è in accordo con quella determinata mediante approcci differenti.
Sono stati utilizzati metodi basati sulla microscopia elettronica anche nella determinazione della posizione di alcune strutture dell'rRNA nel ribosoma. In particolare, l'uso di anticorpi diretti contro nucleotidi metilati, ha permesso di localizzare le dimetiladenosine 1518 e 1519, appartenenti all'rRNA 16S sulla 'piattaforma' (Politz e Glitz, 1977) e la 7mG, in posizione 527 sul 'corpo' (Trempe et al., 1982). La marcatura con apteni ha permesso di localizzare l'estremità 5' e 3' dell'rRNA 16S e l'estremità 3' degli rRNA 5S e 23S (Noller e Lake, 1984). Un'altra tecnica impiega oligonucleotidi sintetici di DNA che formano ibridi molecolari con regioni accessibili dell'rRNA 16S; la loro posizione può essere determinata utilizzando un anticorpo contro un aptene attaccato all'oligonucleotide (Lasater et al., 1990) o sfruttando il legame che si forma tra numerose molecole di avi dina e un polimero di biotina attaccato all' oligonucleotide (Oakes et al., 1986). Entrambi questi metodi di indagine, basati sulla microscopia elettronica, hanno permesso di individuare le posizioni di varie importanti strutture dell'rRNA 16S nella subunità 30S.
Diffrazione neutronica
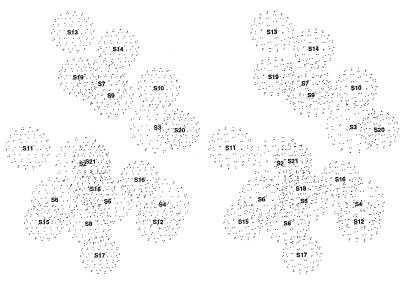
Questa tecnica ha permesso di ottenere una mappa tridimensionale della posizione occupata dalle 21 proteine presenti nella subunità 30S (Capel et al., 1987). Coppie di proteine vengono marcate selettivamente con deuterio e, successivamente, fatte riassociare con gli altri componenti strutturali (non marcati) in una subunità: ciò permette di misurare le distanze tra i centri di massa delle due proteine marcate. Effettuando un gran numero di misurazioni delle distanze relative a numerose coppie di proteine, è possibile determinare, mediante triangolazioni, le posizioni spaziali delle proteine l'una rispetto all'altra. La mappa tridimensionale così ottenuta (fig. 7) è in accordo con i risultati raggiunti con la IEM e permette di correlare i dati ottenuti per mezzo della diffrazione neutronica con la struttura della particella 30S ottenuta mediante microscopia elettronica (v. figura 5). Sebbene nella maggioranza dei casi ci sia corrispondenza sostanziale tra le posizioni della proteina determinate con la IEM e la diffrazione neutronica, sono comunque nate controversie non solo dovute alla discrepanza tra dati provenienti da differenti laboratori, ma anche tra dati ottenuti mediante le due diverse tecniche. In particolare, i dati contrastanti si riferiscono alle proteine S16, Sl9 e S20. Alla proteina Sl9 sono state attribuite, con la tecnica IEM, due posizioni differenti nella testa della subunità 30S (Oakes et al., 1990; Stöffler-Meilecke e Stöffler, 1990) e una terza posizione con la diffrazione neutronica (Capel et al., 1987). Inoltre, risultati preliminari per le proteine Sl6 e S20, ottenuti con la IEM, davano luogo ad ambiguità, ma studi recenti (Schwedler et al., 1993) collocano queste proteine, all'interno della subunità 30S, in una posizione molto più in basso rispetto a quella identificata nella mappa di diffrazione neutronica. Per giungere a un modello chiaro e completo si dovrà comunque ricorrere all'utilizzo di tecniche nuove, dotate di un maggiore potere di risoluzione.
Induzione di legami crociati (cross-linking)
Questa tecnica ha permesso di identificare numerose molecole adiacenti nel ribosoma, fornendo informazioni sulle vicinanze tra proteina e proteina (Traut et al., 1980), tra proteina e RNA o tra RNA e RNA (Brimacombe et al., 1983; 1990). Metodi chimici e foto chimici, utilizzati da gruppi di ricerca differenti, hanno portato a risultati brillanti. Tra i reagenti bifunzionali impiegati, il 2-imminotiolano (Traut et al., 1980) costituisce uno degli esempi più utili ed eleganti da citare. Questa molecola ciclica infatti, reagisce con i gruppi amminici e così facendo rende disponibile un gruppo solfidrilico; l'ossidazione successiva porta alla formazione di ponti disolfuro, tra solfidrili adiacenti, determinando la formazione di legami crociati di natura covalente. Le proteine così legate, vengono poi identificate mediante elettroforesi bidimensionale diagonale su gel, in cui esse migrano come una coppia fuori dalla diagonale.
Un modello molecolare per la subunità 305
Numerosi sono i gruppi di ricerca che hanno ricavato un modello di ripiegamento dell'rRNA 16S all'interno della subunità ribosomale 30S (Brimacombe et al., 1988; Stern et al., 1988; Hubbard e Hearst, 1991; Malhotra e Harvey, 1994). Uno di questi modelli (Stern et al., 1988) è basato sulla posizione delle r-proteine della subunità 30S, determinata mediante diffrazione neutronica. Partendo dalla struttura secondaria dell'rRNA 16S, derivata per confronto filogenetico delle sequenze, i singoli elementi a elica, sono stati agganciati alle proteine corrispondenti, mantenendo sempre distanze stereo chimicamente compatibili tra le eliche. l dati provenienti dagli esperimenti dijootprinting e di cross-linking hanno permesso di verificare quali tratti di rRNA 16S dovessero essere individuati, tramite l'uso di sonde, e con quale tipo di sonde. In figura (fig. 8) è mostrata una versione più recente di questo modello (H.F. Noller, T. Powers, G. Heilek, S. Mian e B. Weiser, articolo non pubblicato) che include una serie supplementare di dati, ottenuti grazie all'uso di sonde costituite da radicali idrossilici per evidenziare le interazioni tra RNA e proteine (Powers e Noller, 1995). È evidente l'elevato grado di sovrapposizione tra questo modello e quello ottenuto mediante la microscopia elettronica (v. figura 5): si possono facilmente identificare gli elementi della 'testa', del 'corpo' e della 'piattaforma'. Un'apparente differenza consiste nella minore massa della parte inferiore del corpo nel modello molecolare, rispetto a quello ricavato con la microscopia elettronica; ciò potrebbe essere dovuto al fatto che solo il 75% della struttura dell'rRNA 16S è stato finora incluso nel modello molecolare e, presumibilmente, il rimanente 25% occupa la parte inferiore della subunità. Una importante predizione di questo modello è che i tre principali domini della struttura secondaria dell'rRNA 16S si ripiegano in tre grandi domini strutturali tridimensionali autonomi. In tal modo, il dominio al 3' definisce la 'testa', quello al 5' il 'corpo' e il dominio centrale le regioni della 'piattaforma' e della 'fenditura'. Recentemente, il dominio al 3' è stato assemblato con le proteine corrispondenti; è stata così prodotta una particella ribonucleoproteica globulare ben definita (Samaha et al., 1995) che fornisce una evidenza diretta dell'autonomia strutturale di questo dominio. In questo modello, inoltre, tutte le basi protette dal tRNA sull'rRNA 16S (Moazed e Noller, 1986; 1991a) sono disposte attorno alla regione della 'fenditura', sulla 'testa' e sulla 'piattaforma', in accordo con la localizzazione del tRNA sulla subunità 30S, ottenuta mediante microscopia elettronica (Gornicki et al., 1984; Wagenknecht et al., 1988). Però, almeno un gruppo di basi protette dal tRNA, nella regione dell'ansa 530, sono localizzate lontano dalla fenditura; la possibilità che la protezione di queste basi fosse causata da un meccanismo allosterico è stata oggetto di numerose discussioni (Brimacombe, 1992; Powers e Noller, 1994).
Modelli di questo tipo sono utili per numerosi aspetti. Oltre a essere un valido approccio (e, fino a ora, forse l'unico) per dedurre la struttura del ribosoma, anche se a bassa risoluzione, l'elaborazione di modelli permette di identificare velocemente il tipo di nuove informazioni che sono necessarie per verificare, confermare o estendere le attuali conoscenze strutturali. Inoltre, essi stimolano e aiutano a focalizzare la discussione sulla struttura e sulla funzione del ribosoma, aspetti sempre più chiaramente connessi ai meccanismi molecolari che stanno alla base della sintesi proteica. Infine, l'interpretazione dei dati cristallo grafici ai raggi X, quando e se saranno disponibili, sarà agevolata da questi tentativi di elaborazione di modelli.
Il meccanismo della traduzione
Al fine di comprendere i meccanismi della traduzione, considerata la sua complessità, è utile chiedersi quali siano i problemi principali che il ribosoma ha dovuto affrontare nella sua evoluzione. Se consideriamo il ribosoma come un enzima, il suo substrato principale è il tRNA. Il primo problema è come il ribosoma riesca a selezionare il corretto tRNA substrato; il secondo è quello della catalisi della formazione del legame peptidico, l'unica reazione per la quale, finora, è stato dimostrato il ruolo catalitico da parte del ribosoma stesso; il terzo, legato all'aspetto più spettacolare, è quello relativo al processo di traslocazione, durante il quale il tRNA compie esatti e, probabilmente, ampi movimenti durante il suo transito sul ribosoma. Si potrebbe quasi dire che molti degli altri aspetti della traduzione - inizio, terminazione, ruolo dei fattori, movimento dell'mRNA, e via di seguito - siano accessori di queste tre attività principali del ribosoma.
Poiché il ribosoma è una particella ribonucleoproteica, è di fondamentale importanza comprendere quali siano i contributi relativi dell'rRNA e delle proteine ribosomali ai tre processi fondamentali sopra citati. Nel 1980 C.R. W oese ha evidenziato l'importanza dello studio del ribosoma dal punto di vista evolutivo, poiché la comprensione della sua evoluzione equivale alla comprensione del suo funzionamento. Se la vita ha avuto origine da un sistema basato sull'RNA (v. il saggio di N. Maizel e A. Weiner, Il mondo a RNA), come è oggi sostenuto dalla gran parte della comunità scientifica, il ribosoma si è certamente evoluto in quel mondo a RNA da un qualcosa di preesistente la cui conoscenza potrebbe aiutarci a comprendere il funzionamento del ribosoma attuale. Non essendo in possesso di tale informazione, possiamo solo ragionevolmente ipotizzare che la grande quantità di RNA contenuta nel ribosoma rispecchi la sua evoluzione a partire da una struttura composta interamente da RNA e che il meccanismo d'azione del ribosoma coinvolga, almeno in parte, l'RNA. Considerando che i ribosomi sono particelle così antiche e complesse, è difficile immaginare come si siano potuti evolvere i primi ribosomi, dal momento che la loro funzione dipendeva da una struttura che doveva assomigliare, anche se alla lontana, a quella attuale. Questo paradosso può essere superato ipotizzando che il ribosoma si sia evoluto dall'RNA, per fusione di molecole di RNA molto più piccole di quelle dell'rRNA odierno, ognuna delle quali potrebbe aver svolto, nel mondo a RNA, una funzione distinta, forse anche non correlata con la sintesi proteica (Noller, 1993). Nella stessa maniera, si può ipotizzare che l'anticodone e il braccio accettore del tRNA si siano anch'essi evoluti, in modo indipendente, da molecole di RNA distinte tra loro. Le interazioni specifiche tra le due estremità opposte del tRNA e le due differenti subunità del ribosoma suggeriscono una coevoluzione dell'anticodone e del braccio accettore del tRNA, rispettivamente con gli rRNA l6S e 23S (Noller, 1993). Anche gli studi sul riconoscimento del tRNA da parte della amminoaciltRNA sintetasi hanno dimostrato che le due estremità del tRNA potrebbero essersi evo Iute separatamente (Schimmel e Henderson, 1994). L'importanza di questo tipo di speculazioni risiede nella loro capacità di semplificare e chiarire la nostra visione di questo problema complesso, in quanto i processi evolutivi sono semplici e procedono per singoli eventi.
Sono riportate delle prove biochimiche, genetiche e derivanti dall'analisi comparativa delle sequenze, a sostegno del diretto coinvolgimento dell'rRNA nel meccanismo di traduzione (Noller, 1991), ed è stata messa in luce la straordinaria conservazione di alcuni elementi nella sequenza dell'rRNA, analoga a quella degli amminoacidi presenti nel sito attivo degli enzimi (Woese et al., 1975; Gutell et al., 1985). Questo è confermato dal fatto che la conservazione di tali elementi nella sequenza dell'RNA si verifica in regioni esattamente equivalenti della struttura secondaria di tutti gli rRNA noti (attualmente migliaia). Sfortunatamente non abbiamo ancora dati sufficienti per poter stabilire se anche le proteine ribosomali presentino una conservazione delle sequenze paragonabile a quella dell'rRNA.
Esperimenti di marcatura per affrnità hanno consentito di localizzare le parti più importanti del tRNA, l' anticodone e l'estremità 3' amminoacetilata, in prossimità di regioni universalmente conservate dell'rRNA (Prince et al., 1982; Barta et al., 1984; Steiner et al., 1988; Wower et al., 1989). Esperimenti di jootprinting chimico hanno inoltre dimostrato che i tRNA proteggono basi conservate negli rRNA l6S e 23S, caratteristiche dello stato legato del tRNA, come sarà descritto più avanti. Anche i fattori di traduzione e gli antibiotici interagiscono con particolari zone dell'rRNA. Considerati nel loro insieme, tutti questi dati biochimici attribuiscono all'rRNA un ruolo cruciale, in accordo con i risultati ottenuti da studi genetici in cui sono state identificate mutazioni dell'rRNA che causano la perdita di funzione del ribosoma (Triman, 1995).
La dimostrazione di un'attività funzionale dell'rRNA privo di proteine, potrebbe fornire la prova più diretta del suo coinvolgimento nella funzionalità del ribosoma. Fino a ora, la realizzazione di questo tipo di esperimenti ha incontrato notevoli difficoltà (4 miliardi di anni di evoluzione potrebbero aver reso l'rRNA dipendente dalle r-proteine), anche se recentemente sono emersi dati incoraggianti. lnnanzitutto, è stato scoperto che diciassette differenti proteine ribosomali possono essere eliminate geneticamente, senza che si origini un fenotipo letale (Dabbs, 1986). In secondo luogo, la straordinaria resistenza della peptidiltransferasi ai procedimenti di estrazione proteica, suggerisce che questa attività catalitica possa essere propria dell'rRNA (Noller et al., 1992). Infine, è stato dimostrato un appaiamento tra basi del tRNA e dell'rRNA 23S nel centro della peptidiltransferasi (Samaha et al., 1995). Questi dati, unitamente ad altri che verranno discussi in seguito, costituiscono prove convincenti a sostegno del paradigma dell'rRNA. Ciò, tuttavia, non significa che la funzione del ribosoma è interamente svolta dall'rRNA, né che possiamo disconoscere il ruolo funzionale delle r-proteine.
l progressi fatti nella comprensione della struttura dell'rRNA, come è stato riassunto precedentemente, sono di particolare importanza poiché la struttura stessa ci permette di comprenderne il meccanismo. In definitiva, la struttura dà origine al meccanismo e, in linea di principio, la relazione tra struttura e funzione può essere studiata in entrambe le direzioni. Un primo esempio è stato fornito dall'analisi della struttura primaria, in cui la conservazione totale di particolari sequenze ha permesso di identificare potenziali siti funzionali (Woese et al., 1975). La maggior parte di queste sequenze conservate è stata poi associata a funzioni ribosomali specifiche (Noller, 1991). Successivamente, la determinazione della struttura secondaria ha permesso di porre vicine tra loro alcune sequenze conservate (per esempio, le regioni 1400 e 1500 conservate universalmente, e quelle dell'ansa centrale del dominio V dell'rRNA 23S) e ciò implica che questi differenti elementi di sequenza si combinano tra loro per formare un sito funzionale.
Selezione del tRNA da parte del ribosoma
Interazioni tRNA-ribosoma, distinte da quelle tRNA-mRNA
Sebbene la specificità dell'amminoacil-tRNA sia determinata dal suo anticodone, la stabilità dell'interazione codone-anticodone è insufficiente a spiegare sia la stabilità dell'interazione tRNA-ribosoma sia l'accuratezza del processo della traduzione osservata in vivo (Grosjean et al., 1978). Evidentemente l'apparato di traduzione, in qualche modo, potenzia il legame del corretto tRNA al ribosoma. Anche se questo processo non è del tutto chiaro, esistono prove a sostegno del coinvolgimento, in questa fase, di un qualche tipo di meccanismo cinetico di 'correzione di bozze' (Hopfield, 1974; Ninio, 1975).
Interazioni nel sito P tra tRNA e rRNA
Il tRNA interagisce con il ribosoma stesso a prescindere dall'interazione codone-anticodone. Ciò è dimostrato chiaramente per il tRNA legato al sito ribosomale P peptidilico. A elevate concentrazioni di ioni magnesio, il tRNA è in grado di legarsi ai ribosomi indipendentemente dall'mRNA (Nirenberg e Leder, 1964). Esperimenti in cui si utilizzano sonde chimiche (Moazed e Noller, 1986; 1989a; 1991) hanno dimostrato che il tRNA interagisce, direttamente o indirettamente, con specifiche basi conservate sia sull'rRNA l6S che 23S del ribosoma (v. figure 2, 3a e 3b). A elevate concentrazioni di ioni magnesio, le basi dell'rRNA protette dal legame mRNA - indipendente del tRNA, sono identiche a quelle protette, in condizioni fisiologiche, dal legame mRNA - dipendente al sito P. Questi risultati suggeriscono che il legame del tRNA, nelle due differenti condizioni, è essenzialmente simile e che l'rRNA contribuisce alla formazione di tale legame.
Nell'rRNA l6S, le protezioni del sito P sono distribuite in diverse regioni della struttura. Come discusso più avanti, le basi maggiormente protette del sito P sono raggruppate, nel modello tridimensionale, intorno alla fenditura della subunità 30S. Nell'rRNA 23S, invece, le basi protette sono localizzate principalmente nel dominio V, e molte di esse si trovano nell'ansa centrale universalmente conservata, che mediante studi di marcatura per affinità è stata localizzata nella regione della peptidiltransferasi. Cinque delle basi protette, A245l, G2505, U2506, U2584 e U2585, corrispondono esattamente alle basi che vengono marcate per affrnità dal Phe-tRNA, modificato dal benzofenone. Le basi A245l e G2505 sono entrambe protette dal cloramfenicolo e dalla carbomicina, due inibitori della peptidiltransferasi (Moazed e Noller, 1987b), a ulteriore conferma che le basi protette dal peptidil-tRNA nell'rRNA 23S sono effettivamente all'interno o in stretta vicinanza del sito della peptidiltransferasi.
Il tRNA interagisce con l'rRNA attraverso le sue estremità
Tutte le protezioni delle basi nell'RNA l6S, dovute al legame del tRNA ai siti A e P nella subunità 30S o nei ribosomi 70S, possono essere simulate legando un frammento di 15 nucleotidi corrispondente al braccio dell' anticodone (Moazed e Noller, 1986; 1991a); ciò indica che tutti gli effetti osservati sono dovuti a interazioni con questa limitata regione del tRNA. In modo analogo, praticamente tutte le protezioni nell'rRNA 23S (dei siti A, P ed E) sono abolite dalla delezione del CA 3' -terminale del tRNA (Moazed e Noller, 1989a), suggerendo che la sequenza conservata CCA 3'-terminale è responsabile delle protezioni nell'rRNA della subunità grande. Queste osservazioni sono state confermate nel caso del legame al sito P, dove oligonucleotidi contenenti l'estremità 3'-terminale CCA (CACCA-[N-Acetil-Phe], UACCA-[N-Acetil-Leu] o CAACCA-[f-Met]) possono essere legati in presenza di sparsomicina ed etanolo. Questi oligonucleotidi acetilati possiedono le stesse capacità di protezione dell'rRNA 23S di un intero tRNA (Moazed e Noller, 1991b), dimostrando che, almeno per quanto riguarda il tRNA legato al sito P, l'estremità CCA è necessaria e sufficiente per la protezione dell'rRNA 23S. Si può, quindi, concludere che le interazioni osservate tra tRNA e rRNA coinvolgono solo le estremità funzionali del tRNA, cioè il braccio dell'anticodone e il terminale accettore.
Interazioni tra tRNA e rRNA 165 nel sito A
La stabilità di legame del tRNA al sito A dipende fortemente dall'mRNA e anche in questo caso vi è, nell'rRNA l6S e nel 23S, una serie specifica di basi protette. Benché non esistano prove definitive, è possibile ipotizzare che le protezioni nel sito A siano dovute al tRNA piuttosto che all'mRNA, considerando, per esempio, che il legame del solo mRNA è insufficiente per la protezione di queste basi e che le protezioni nel sito A, nell'rRNA 23S, non sembrano dipendere dall'mRNA. In ogni caso, l'osservazione che il legame del tRNA al sito A, mediato dall'mRNA, causa la protezione (o, in alcuni casi, la accresce) di una serie di basi nell'rRNA, indica l'esistenza di interazioni tra le due specie di RNA. Nell'rRNA l6S le basi protette sono localizzate in tre specifiche regioni della struttura dell'rRNA (v. figura 2). l residui 1405, 1408, 1492, 1493 e 1494 sono localizzati nel 'sito di decodificazione' che è, a sua volta, adiacente alla C1400, con cui l'anticodone del tRNA del sito P forma legami crociati (Prince et al., 1982). Poiché i codoni del sito A e P sono adiacenti sull'mRNA, i due siti devono esserlo anche nel ribosoma.
Numerose prove sperimentali avvalorano l'ipotesi secondo la quale l'interazione codone-anticodone avviene nella fenditura della subunità 30S. lnnanzitutto la totale conservazione della struttura primaria delle regioni 1400 e 1500 è in accordo con l'importanza biologica di un tale sito funzionale. In secondo luogo molti antibiotici, che inducono errori della traduzione, proteggono basi di questa stessa regione: neomicina, kanamicina, gentamicina, paromomicina, igromicina, neamina e apramicina, tutte proteggono le basi A1408, G149l e G1494, che sono localizzate direttamente nella zona di jootprinting del tRNA sul sito A, o molto vicine a essa (Moazed e Noller, 1987a; Woodcock et al., 1991). Alcuni autori hanno ipotizzato che questi antibiotici agiscano stabilizzando l'interazione aspecifica del tRNA al sito A, probabilmente di storcendo la conformazione dell'rRNA l6S in questa regione. È stato infatti trovato che mutazioni in questa regione fanno insorgere resistenza a tali antibiotici (Li et al., 1982; Spangler e Blackburn, 1985; De Stasio et al., 1989). In terzo luogo, la mutazione della base corrispondente alla A1409 dell'rRNA l6S mitocondriale di lievito, conferisce al ribosoma un'attività di soppressore dello 'sfasamento di lettura del codice', che rappresenta un altro tipo di errore di codificazione (WeissBrummer e Huttenhofer, 1989). In quarto luogo, la colicina E3, che inattiva il ribosoma impedendo il legame del tRNA mediato dall'mRNA, opera un taglio nell'rRNA l6S, tra le basi A1493 e Gl494, adiacenti alle due basi maggiormente protette nel sito A (Bowman et al., 1971; Senior e Holland, 1971).
Interazioni del tRNA con l'rRNA 235 nel sito A
Nell'rRNA 23S, le basi protette dall'interazione del tRNA con il sito A sono localizzate nel dominio V, grossomodo nella stessa regione dove si trovano le protezioni del sito P. Infatti due basi, la A2439 e la A245l, vengono protette in entrambi i tipi di interazione. La protezione di queste basi dipende, però, dall'acilazione del tRNA in quanto non si osserva alcuna protezione nei siti A o P, in seguito alla rimozione del gruppo acile (Moazed e Noller, 1989a). Questi dati suggeriscono che il gruppo acile del tRNA possa interagire con queste due basi da entrambi i siti, cosa molto probabile dato che i due siti devono essere in stretta vicinanza nella regione della peptidiltransferasi per consentire la formazione del legame peptidico. La delezione della A 3' terminale impedisce, durante l'interazione del tRNA con il sito A, la protezione della G2553, mentre la delezione della penultima C impedisce quella della C2254.
EF- Tu protegge l'estremità amminoacilica del tRNA dalla peptidiltransferasi
Sia il fattore di allungamento Tu (EF-Tu, Elongation Factor Tu) che il fattore di allungamento G (EF-G, Elongation Factor G) proteggono le basi universalmente conservate dell'ansa α-sarcina nel dominio VI dell'rRNA 23S (Moazed et al., 1988), in accordo con l'osservazione che essi competono tra loro per il legame al ribosoma. Inoltre, EF -G protegge alcune basi nella regione intorno alla base 1067 del dominio 11, dove sono stati localizzati sia i siti di interazione con le proteine ribosomali L 11, L l O e L 7/Ll2, sia il sito d'azione del tiostreptone, uno specifico inibitore dell'EF-G (Cundliffe, 1981).
Quando il complesso ternario EF-Tu·tRNA·GTP si lega al ribosoma, si osserva l'usuale protezione nell'rRNA l6S a livello del sito A, ma non quella nel sito A dell'rRNA 23S, fino a che il complesso non viene rilasciato (Moazed et al., 1988; Moazed e Noller, 1989a). Invece, si osservano le protezioni a livello dell'ansa α-sarcina dovute all'interazione tra l'EF-Tu e l'rRNA 23S (Moazed et al., 1988; Moazed e Noller, 1989a). Questi risultati sperimentali riflettono, molto probabilmente, un meccanismo di 'correzione di bozze' che si verifica durante la traduzione: fino a che il fattore EF-Tu rimane legato al ribosoma, l'estremità 3' dell'amminoacil-tRNA viene esclusa dal sito A del 23S, impedendo la sua partecipazione alla formazione del legame peptidico; un tRNA non corretto può essere così rilasciato prima di venir utilizzato dalla peptidiltransferasi.
La misteriosa ansa 530
Una seconda regione dell'RNA l6S coinvolta nel legame del tRNA al sito A è l'ansa 530, oggetto di affascinanti osservazioni. Dopo la regione 1400/1500, l'ansa 530 è l'elemento più conservato dell'rRNA l6S. La base G530, una delle più re attive del ribosoma, e la base U53l a essa adiacente, vengono protette dall'interazione del tRNA con il sito A (Moazed e Noller, 1986; 1991a). In accordo, però, a numerose prove sperimentali l'ansa 530 risulta lontana dal sito di decodificazione (più di 75 Å), rendendo scarsamente probabile che questi effetti siano causati dalla diretta interazione del tRNA con le basi dell'ansa 530.
L'importanza della base G530 per la funzionalità del ribosoma è stata verificata direttamente con esperimenti di mutagenesi sito-specifica: la sua mutazione in una qualsiasi altra base genera un fenotipo letale dominante (Powers e Noller, 1990). L'rRNA l6S mutato è in grado di assemblarsi nella subunità 30S, e di formare la particella 70S, ma i ribosomi mutanti non sono in grado di legare il tRNA nel sito A, secondo il processo EF-Tu dipendente.
Queste osservazioni si possono conciliare con i dati ottenuti da studi cinetici della traduzione, in accordo al seguente modello della selezione del tRNA. Al suo ingresso, il complesso ternario interagisce con il ribosoma in almeno due posizioni: una prima interazione avviene tra l'ansa dell'anticodone e il codone disponibile nel sito A a livello della fenditura della subunità 30S; la seconda, invece, avviene tra EF-Tu e l'ansa α-sarcina dell'rRNA 23S.lnoltre EF-Tu lega l'estremità amminoacilica del tRNA, impedendone l'ingresso nel sito A del centro della peptidiltransferasi e, di conseguenza, blocca l'incorporazione del nuovo amminoacido nella catena proteica, fino alla sua dissociazione dal tRNA. Questa è mediata dall'idrolisi del GTP e risulta nella formazione del complesso EF-Tu·GDP che possiede una bassa affrnità per il tRNA e il ribosoma. Il complesso EF-Tu·GDP viene rilasciato prima del tRNA, cosicché l' amminoacido può essere incorporato nella catena polipeptidica, ma se il complesso ternario EF-Tu·tRNA·GTP viene rilasciato prima dell'idrolisi del GTP, l'amminoacido avrà perso l'opportunità di essere incorporato. Perciò, la modulazione dell'idrolisi del GTP può determinare se uno specifico amminoaciltRNA viene utilizzato o meno dalla peptidiltransferasi. Una possibile ipotesi è che una corretta interazione codone-anticodone induca una modificazione conformazionale nella subunità 30S; ciò determinerebbe una modificazione della re attività chimica delle basi G530 e U531. Questo potrebbe corrispondere a un segnale, probabilmente trasmesso attraverso l'EF-Tu, che provoca un'aumentata idrolisi del GTP. In questo modo, un tRNA corretto, ma non uno sbagliato, potrebbe stimolare il rilascio dell'EF-Tu e l'esposizione del gruppo amminoacilico che consentirebbe la sua incorporazione nella catena polipeptidica nascente.
Interazioni tRNA-ribosoma durante i processi di allungamento e di traslocazione
Il modello classico
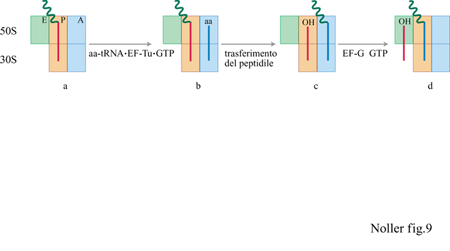
Per tre decenni, il processo di allungamento/traslocazione della sintesi proteica si è basato concettualmente sul modello proposto originariamente da J.D. Watson (1964), il quale aveva ipotizzato che il ribosoma dovesse possedere almeno due siti di legame per il tRNA (generalmente indicati come siti A, o amminoacilico, e P, o peptidilico), in accordo con i dati in suo possesso riguardo al meccanismo della sintesi proteica. Più recentemente è stata dimostrata l'esistenza di un terzo sito, il sito E (dall'inglese exit) o di uscita (Rheinberger et al., 1981). Il modello classico per la traduzione, con l'aggiunta del sito E, è mostrato nella figura (fig. 9).
Iniziando da una situazione in cui un peptidil-tRNA occupa il sito P, un amminoacil-tRNA viene trasferito nel sito A vuoto sotto forma di un complesso temario con EF - Tu e GTP. Il GTP viene idrolizzato, e il complesso binario EF-Tu·GDP rilasciato mentre l'amminoacil-tRNA rimane nel sito A. La peptidiltransferasi, successivamente, catalizza spontaneamente la formazione del legame peptidico, con conseguente trasferimento della catena polipeptidica nascente sul tRNA del sito A, mentre resta un tRNA deacilato nel sito P. A questo punto, EF-G catalizza il movimento dei due tRNA rispettivamente dal sito A al P e da P a E. Sono state formulate varie ipotesi per poter correlare i dati sperimentali con il modello classico; per esempio, l'occupazione del sito P viene normalmente dedotta dalla capacità del peptidil-tRNA di reagire con la puromicina, formando un complesso peptidil-puromicina, caratterizzato dalla presenza di un legame peptidico. Questo fatto corrisponde sostanzialmente alla definizione di sito P data da Watson, poiché è difficile immaginare come un complesso tra puromicina e peptidil-tRNA si possa formare in un altro sito. Comunque, non si può escludere la presenza, in qualche altro sito, di peptidiltRNA non re attivi verso la puromicina, poiché ci potrebbero essere molti motivi per una perdita di re attività. Alcuni ricercatori, in base al modello originale dei due siti, hanno preferito concludere che, se il tRNA viene legato al ribosoma in un sito diverso da P, questo deve essere il sito A.
Nel modello originale di Watson era implicita l'assenza di riferimenti a una dinamica del ribosoma, il che ha dato origine a una sua concezione statica. In effetti, esperimenti sulla dinamica ribosomale sono difficili da realizzare. Una delle prove sperimentali più note a favore di una dinamica del ribosoma, costituita dalle differenti costanti di sedimentazione dei ribosomi 70S prima e dopo la traslocazione (Spirin, 1969), è stata oggetto di una diversa interpretazione: il picco 60S osservato è stato considerato un artefatto dell'equilibrio di dissociazione delle subunità, reso più evidente dall'elevata pressione a cui sono sottoposti i campioni durante la centrifugazione (NolI et al., 1973).
Stati ibridi
Utilizzando sonde chimiche per identificare le interazioni tRNA-rRNA è possibile saggiare, in modo più diretto, i siti, o stati, di legame del tRNA, in modo da valutare la condizione del tRNA nel ribosoma, indipendentemente da molte delle assunzioni richieste dagli approcci precedenti. Il footprinting dei siti A, P ed E può essere considerato un indicatore della presenza, nel ribosoma, di tRNA in diversi stati funzionali. La definizione di uno stato è quindi cruciale: un tRNA nel sito P indica quello stato in cui il tRNA è puromicina-reattivo (nessuna conclusione diretta viene tratta dalla non reattività); il sito A è il luogo in cui un tRNA viene portato dal complesso temario EF-Tu·tRNA·GTP, in seguito all'idrolisi del GTP e al rilascio del complesso binario EF-Tu·GDP; il legame al sito E viene definito come strettamente specifico per i tRNA deacilati con un'estremità CCA 3' terminale intatta.
La verifica sperimentale del modello classico ha fornito risultati inattesi (Moazed e Noller, 1989b). Il primo e il più semplice degli esperimenti è stato effettuato per esaminare lo stato dell'N-Acetil-Phe-tRNA legato al ribosoma, prima e dopo la reazione con la puromicina: prima dell'aggiunta di questa, le basi protette nell 'rRNA l6S e 23 S corrispondono a quelle attese dall'interazione nel sito P. Dopo la reazione con la puromicina, il modello classico prevede che il tRNA deacilato rimanga legato al sito P, ossia nella medesima posizione che occupava prima della reazione della peptidiltransferasi. l dati sperimentali mostrano che le basi protette nell'rRNA l6S rimangono inalterate. Perciò, riguardo alla subunità piccola, il tRNA rimane nel sito P, come previsto dal modello classico. Al contrario, ilfootprinting nell'rRNA 23S risulta completamente alterato: le protezioni del sito P nell'rRNA 23S sono completamente abolite, mentre compare ilfootprinting tipico del sito E. È ovvio che la perdita di footprinting del sito P nell'rRNA 23S non può essere attribuita semplicemente alla mancanza di contatto tra la regione dell'N-Acetil-Phe e l'rRNA 23S, poiché la maggior parte delle protezioni del sito P sono indipendenti dalla presenza del gruppo acile. Questi esperimenti dimostrano che, durante la formazione del legame peptidico, l'estremità 3' del tRNA cambia spontaneamente il suo stato di legame con il ribosoma. In altre parole, il tRNA si trova simultaneamente nel sito p del 16S e nel sito E del 23S: questo stato ibrido di legame del tRNA è detto stato P/E.
In una versione 'più fisiologica' di questo esperimento, la puromicina è stata sostituita da un amminoacil-tRNA. Il Phe-tRNA è stato legato al ribosoma come complesso ternario EF-Tu·Phe-tRNA·GTP. In seguito al rilascio dell'EFTu·GDP e alla formazione del legame peptidico, ilfootprinting dell'rRNA 16S corrispondeva ai siti A e P, mentre quello dell'rRNA 23S ai siti P ed E. La spiegazione più semplice di questi dati è che i due tRNA si trovino in uno stato ibrido: il tRNA deacilato è nello stato ibrido P/E, come dimostrato negli esperimenti con la puromicina, mentre l'NAcetil·Phe·Phe·peptidil-tRNA si trova nello stato A rispetto al 16S e nello stato P rispetto all'rRNA 23S, il cosiddetto stato A/P (Moazed e Noller, 1989b). Se a questo complesso vengono poi aggiunti il fattore d'allungamento EF-G e il GTP, ilfootprinting viene ulteriormente alterato, ma solo per quanto riguarda l'rRNA 16S: il footprinting del sito A viene perso mentre persiste quello del sito P, indicando che il peptidil-tRNA viene convertito, dal fattore d'allungamento EF -G, dallo stato AlP allo stato P/P (equivalente al classico legame al sito P), mentre il tRNA deacilato passa dallo stato P/E a quello E. In altre parole, il legame nello stato ibrido dei due tRNA viene convertito in un legame in stato non ibrido dall'azione dell'EF-G. Quindi, l'evento catalizzato da EF -G coinvolge principalmente un movimento del tRNA rispetto alla subunità piccola, mentre il movimento dell' estremità CCA del tRNA rispetto alla subunità grande sembra avvenire spontaneamente in seguito alla formazione del legame peptidico. In questo modo la traslocazione deve avvenire in almeno due fasi distinte: una coinvolge uno spostamento rispetto alla subunità 50S, l'altra rispetto alla subunità 30S.
Lo studio diretto del legame del peptidil-tRNA al sito A costituisce il terzo tipo di test cui il modello classico è stato sottoposto. Quando il peptidil-tRNA viene legato al ribosoma in presenza di un eccesso di tRNA deacilato, esso si trova in uno stato non reattivo alla puromicina (De Groot et al., 1971; Watanabe, 1972). Questo, in un contesto classico, sta a indicare che il peptidil-tRNA deve essere legato al sito A, mentre il tRNA deacilato si trova nel sito P. È importante notare che questa conclusione era basata più sulla mancanza di re attività verso la puromicina, che su concrete evidenze della sua presenza nel sito A. L'impiego di sonde chimiche su un complesso in cui l'N-Acetil-Phe-tRNA era legato in presenza di un eccesso di tRNA deacilato ha, però, evidenziato una situazione diversa: nell'rRNA 16S sono stati osservati footprinting di A e P, mentre nell 'rRNA 23 Svengono evidenziati quelli di P ed E. Questi risultati dimostrano che, in queste condizioni, l'N-Acetil-Phe-tRNA è legato in uno stato A/P, mentre il tRNA deacilato è legato in uno stato P/E (Moazed e Noller, 1989b), imitando esattamente lo stato in cui si trova il tRNA sul ribosoma immediatamente dopo la formazione del legame peptidico, come nel precedente esperimento.
Implicazioni su stati ibridi e meccanismo di traduzione
Dal modello degli stati ibridi emergono alcune importanti implicazioni sul meccanismo di traduzione:
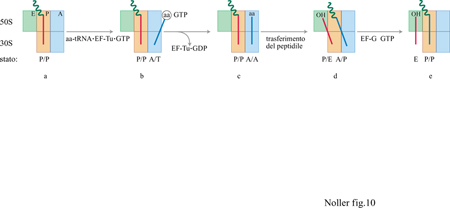
1) si possono distinguere almeno sei differenti stati di legame del tRNA al ribosoma (P/P, AlT, AIA, P/E, AlP, ed E), piuttosto che due o tre (il sito T indica il legame del tRNA con EF-Tu) (fig. 10).
2) La traslocazione avviene in due passaggi, il primo dei quali coinvolge lo spostamento dell'estremità amminoacilica del tRNA rispetto alla subunità grande del ribosoma e avviene spontaneamente dopo la formazione del legame peptidico.
3) Il peptide nascente rimane nel sito P della subunità grande durante le diverse fasi dell' allungamento mentre il tRNA SI muove.
4) La transizione tra stato ibrido e stato non ibrido sembra essere una proprietà del ribosoma poiché, quando due tRNA sono legati, si trovano entrambi o nello stato ibrido o in quello non ibrido.
5) L'esistenza degli stati ibridi suggerisce un ruolo per il sito E. Durante il trasferimento peptidico, l'estremità CCA del tRNA deacilato è spostata dal sito P del 23 S quando la catena polipeptidica viene trasferita all'amminoacil-tRNA. Ciò implica che, in assenza del sito E, non ci sarebbe alcun sito di legame per l'estremità 3' del tRNA deacilato. Questo sito di legame viene fornito dal sito E, e ciò potrebbe contribuire alla stabilità termodinamica del complesso dello stato ibrido, favorendo la progressione della prima, spontanea, fase della traslocazione. In quest'ottica è facile comprendere perché solo la subunità grande del ribosoma ha bisogno di un sito E, visto che il suo principale scopo è quello di fornire la componente 50S allo stato ibrido PIE.
6) L'osservazione che la prima fase della traslocazione sembra coinvolgere principalmente la subunità 50S, mentre la seconda la 30S, suggerisce l'ipotesi che la traslocazione possa coinvolgere un movimento relativo delle due subunità ribosomali. Se quest'ipotesi venisse confermata fornirebbe una spiegazione razionale del fatto che in tutti gli organismi i ribosomi si compongono di due subunità. Quando si verificano ampi movimenti molecolari, il cardine del meccanismo è spesso costituito da movimenti relativi di subunità, piuttosto che da cambiamenti conformazionali all'interno di una stessa macromolecola.
Prove fisiche del meccanismo degli stati ibridi
O.W. Odom e B. Hardesty hanno fornito la prova fisica di un secondo evento di traslocazione. Mediante l'utilizzo di sonde fluorescenti, legate a determinate posizioni del tRNA e del ribosoma, hanno osservato cambiamenti nel rendimento quantico e nell' anisotropia della fluorescenza in seguito alla formazione del legame peptidico (Odom e Hardesty, 1987), dimostrando così che una parte del tRNA si muove rispetto alle proteine ribosomali S21 e L Il. Su questa base, hanno ipotizzato che la catena peptidica rimanga in una posizione costante rispetto alla struttura del ribosoma mentre il tRNA si sposta durante la reazione della peptidiltransferasi. Più recentemente gli stessi autori hanno utilizzato il trasferimento di energia non radiativa per misurare l'ampiezza del movimento del tRNA rispetto al ribosoma, in seguito alla reazione della peptidiltransferasi (Odom et al., 1990). Una sonda fluorescente, legata all'estremità 5' del tRNA, si muove di almeno 20 Å verso la proteina Ll, mentre una sonda, legata alla 4-tiouridina in posizione 8, si allontana dalla proteina S21 di 10 Å. Al contrario, la catena polipeptidica nascente si muove solamente di pochi Å. D'altro canto, studi precedenti in cui si seguiva la posizione dell'mRNA nel ribosoma, utilizzando la protezione alla nucleasi, avevano dimostrato che l'mRNA (e, di conseguenza, l'anticodone dei tRNA) si sposta esattamente di un codone rispetto alla struttura del ribosoma, ma solo in risposta a EF-G e GTP. Questi esperimenti forniscono prove molto convincenti a sostegno del modello degli stati ibridi (v. figura 10).
Peptidiltransferasi
La peptidiltransferasi è una parte del ribosoma
I legami peptidici presenti nelle proteine vengono formati nel ribosoma da una attività enzimatica chiamata peptidiltransferasi. I primi studi hanno dimostrato che essa è parte integrante del ribosoma stesso (Maden et al., 1968). R.E. Momo e K.A. Marker hanno messo a punto la cosiddetta 'reazione del frammento' ifragment reaction), che consiste in un semplice saggio della peptidiltransferasi in cui il tRNA del sito P viene rimpiazzato da un oligonucleotide del tipo CAACCA-(f-Met), e il tRNA del sito A dalla puromicina (Momo e Marcker, 1967). La puromicina contiene un analogo dell'amminoaciladenosina simile all'estremità 3' di un aminoacil-tRNA, capace di donare il suo gruppo amminico al carbonile del substrato peptidico per formare un prodotto contenente un legame peptidico, in questo caso f-Met-puromicina. La reazione richiede componenti molto semplici: oltre ai siti A e P e alcole, necessita solo della subunità ribosomale 50S, di ioni magnesio e potassio. Non richiede, invece, mRNA, subunità 30S, fattori di traduzione, GTP, ATP o tRNA interi.
T. Staehelin e collaboratori (1969) hanno dimostrato che non è necessaria neanche l'intera particella 50S. Separando le subunità in un gradiente di densità di cloruro di cesio, il nucleo della particella, privo della maggior parte delle proteine ribosomali, mostra ancora livelli discreti di attività peptidiltransferasica. La rimozione successiva di altre proteine dal nucleo causa, infine, una perdita di attività che può, comunque, essere recuperata con l'aggiunta delle proteine purificate. In questo modo è stato dimostrato che solo la proteina Ll6 è responsabile del recupero di attività dei nuclei privati delle proteine (Moore et al., 1975). Inoltre, circa la metà delle 30 proteine della subunità grande sono assenti completamente nella particella attiva ricostituita. La possibilità che la proteina Ll6 avesse un ruolo catalitico è stata smentita quando è stato dimostrato il suo ruolo nel mantenimento del corretto ripiegamento tridimensionale della particella (Teraoka e Nierhaus, 1978). Inoltre nessuna proteina isolata, o miscela di proteine ribosomali prive di RNA, è in grado di catalizzare la reazione di peptidiltransferasi. Mediante riassemblaggio di RNA in vitro, con varie combinazioni delle proteine della subunità 50S, K.H. Nierhaus e collaboratori hanno mostrato che le proteine essenziali appartengono a un gruppo limitato, comprendente L2, L3, L4, Ll5 e Ll6 (Hampl et al., 1981).
Marcatura per affinità
Diversi gruppi di ricerca hanno messo a punto metodi di formazione legami crociati per identificare le componenti molecolari del sito della peptidiltransferasi. Considerando che l'estremità amminoacilica (accettore) del tRNA deve interagire con il centro di peptidiltransferasi, sono stati sintetizzati dei reagenti per la marcatura per affinità che contenevano gruppi chimici re attivi attaccati all' amminoacido o al peptide legati al tRNA. Questi reagenti, dopo il legame dei tRNA modificati al ribosoma, reagiscono con le componenti macromolecolari del ribosoma in prossimità del sito catalitico. In molti casi sono stati fatti tentativi per dimostrare che i tRNA legati si trovano in uno stato fisiologicamente rilevante, dimostrando per esempio che essi possiedono ancora la capacità di partecipare alla reazione della peptidiltransferasi.
Da questi esperimenti emergono molti dettagli importanti. Tra le proteine maggiormente soggette a legame crociato ci sono la L2, la L 15, la L 16 e la L27. L'appartenenza di questo gruppo di proteine alla regione circostante il sito catalitico di peptidiltransferasi è convalidata anche da esperimenti di legame crociato proteina-proteina e da esperimenti di immunomicroscopia elettronica che hanno permesso di localizzare queste proteine una vicina all'altra nella subunità ribosomale grande, in una posizione che corrisponde al centro della peptidiltransferasi (Cooperman et al., 1990). Almeno una di queste proteine, L27, non è presente nella particella attiva privata di alcune proteine (Moore et al., 1975; Hampl et al., 1981): ciò limita ulteriormente la lista dei candidati a L2, Ll5 e L16; tuttavia nessuna tra queste proteine mostra attività peptidiltransferasica in assenza di rRNA.
Coinvolgimento dell'rRNA 235 nel funzionamento della peptidiltransferasi
Utilizzando i metodi di marcatura per affinità del tRNA descritti sopra, è stato dimostrato che l'rRNA 23S viene legato in modo crociato con discreta efficienza (Bispink e Matthaei, 1973; Girshovich et al., 1974; Breitmeyer e Noller, 1976). In effetti, la possibilità che l'rRNA potesse svolgere un ruolo funzionale nel processo di traduzione era stata avanzata molti anni prima della scoperta dei ribozimi. La connessione tra peptidiltransferasi e RNA divenne più evidente quando A. Barta e collaboratori (1984) identificarono, nell'rRNA 23S, il sito di legame crociato fotochimico del Phe-tRNA modificato con il benzofenone. Questo legame crociato era particolarmente interessante: veniva ottenuto con una resa eccezionalmente alta (più del 50%) e il tRNA legato in modo crociato manteneva la capacità di prendere parte alla reazione della peptidiltransferasi (Barta e Kuechler, 1983). I siti di legame crociato furono localizzati nell'ansa centrale del dominio V dell'rRNA 23S universalmente conservata, in corrispondenza delle posizioni 2584 e 2585, alle quali si lega il tRNA nello stato AlP, in accordo con il modello degli stati ibridi, e delle posizioni 2451 e 2452 occupate dal tRNA nello stato P/P (Steiner et al., 1988). Il significato della localizzazione di questi legami crociati è stato subito chiaro: mutazioni che conferiscono resistenza al cloramfenicolo, un inibitore della peptidiltransferasi, sono state localizzate in cinque diverse posizioni della stessa ansa. Successivamente, molte altre mutazioni in grado di conferire resistenza a questo tipo di antibiotici sono state localizzate qui, mentre, dopo anni di ricerca, nessuna è stata trovata al di fuori dell'ansa centrale (Vester e Garrett, 1988). Due di questi inibitori, il cloramfenicolo e la carbomicina, hanno inoltre mostrato di proteggere basi specifiche in questa stessa regione dell'rRNA 23S (Moazed e Noller, 1987b).
Recentemente, esperimenti di jootprinting con il tRNA hanno fornito ulteriori prove del ruolo svolto da questa regione del dominio V nell' attività peptidiltransferasica. Quando il tRNA è legato al ribosoma in modo tale che i siti A e P dell'rRNA 23S siano occupati, è possibile osservare protezioni differenti da quelle ottenute mediante rivelazione chimica (Moazed e Noller, 1989a; 1991b). Queste 'impronte' sono caratteristiche dei tre siti differenti e possono venir utilizzate come prova diretta della localizzazione del tRNA nel ribosoma. Le basi protette dal tRNA sono quasi esclusivamente localizzate nel dominio V, più precisamente nell'ansa centrale o nelle regioni che si estendono a partire da essa.
Le prove sperimentali, riportate in precedenza, mostrano che alcune caratteristiche strutturali conservate dell'rRNA 23S sono localizzate nel sito in cui ha luogo l'evento catalitico della peptidiltransferasi. Ma quali sono le prove a sostegno del coinvolgimento diretto dell'rRNA in tale reazione o, addirittura, del suo ruolo catalitico? Gli approcci possibili per la soluzione di tale problema possono essere di due tipi: la rimozione sistematica dal ribosoma delle proteine ribosomali senza alterazione della struttura tridimensionale (o della funzione) dell'rRNA, o la dimostrazione diretta di un appaiamento tra l'estremità CCA del tRNA e l'rRNA 23S.
In studi preliminari (Noller et al., 1992), è stato dimostrato che il ribosoma o anche la sola subunità 50S di E. coli sono in grado di catalizzare la 'reazione del frammento' in presenza di dodecilsolfato di sodio (SDS), un energico detergente ionico comunemente utilizzato per rimuovere le proteine dai complessi nucleoproteici. L'attività resiste anche a digestione con proteinasi K (un enzima che degrada le proteine), effettuata in presenza o meno di SDS. Solo il trattamento con un forte agente chelante, quale l'EDTA, o l'estrazione con fenolo, sono in grado di inibire l'attività della peptidiltransferasi. L'effetto dell'EDTA è in accordo con la ben nota importanza degli ioni metallici nella stabilizzazione delle strutture d'ordine superiore dell'RNA e con la richiesta di ioni Mg²+ nelle reazioni catalizzate da RNA. La perdita di attività dovuta all'estrazione con fenolo, potrebbe essere spiegata con l'eventualità che la struttura d'ordine superiore dell 'rRNA di E. coli, un batterio mesofilico, diventi instabile in assenza di proteine ribosomali.
In considerazione della maggiore stabilità dell 'RNA di organismi termofili, sono stati analizzati i ribosomi provenienti da un eubatterio termofilo, il Thermus aquaticus. L'attività peptidiltransferasica dei ribosomi di Thermus non solo è maggiormente resistente ai trattamenti con SDS e proteinasi K, ma mostra, in seguito a tali trattamenti, perfino un lieve incremento dell'attività (Noller et al., 1992); inoltre, il trattamento con fenolo riduce di poco tale attività. La procedura utilizzata per la preparazione dell'rRNA comprendeva una predigestione con proteinasi K in presenza di SDS, seguita da un'agitazione continua per 45 minuti con un eguale volume di fenolo, a cui seguiva una seconda estrazione (15 minuti) e precipitazione con etanolo. Nonostante questo tipo di trattamento, una quantità significativa di proteine resiste all'estrazione. In esperimenti differenti, condotti con varie preparazioni ribosomali di Thermus, i livelli di proteine residue oscillano tra il 5 e il 20% del contenuto iniziale nella subunità in esame. In altre parole, il materiale analizzato contiene tra il 2 e l' 8% di proteine e tra il 98 e il 92% di RNA. Parte della variabilità del contenuto proteico è da mettere in relazione con quantità variabili di proteine non ribosomali che aderiscono ai ribosomi durante il loro isolamento.
Un punto importante è determinare se la reazione operata da queste particelle private di proteine è realmente catalizzata dalla peptidiltransferasi. Esperimenti condotti con antibiotici hanno dimostrato che è proprio così: il cloramfenicolo e la carbomicina, due inibitori specifici della peptidiltransferasi, inibiscono la catalisi da parte delle particelle private di proteine, tanto quanto inibiscono quella operata dai ribosomi intatti; invece gli antibiotici di cui non si conosce alcun effetto sulla peptidiltransferasi, non alterano in alcun modo la reazione (Noller et al., 1992). In contrasto al mancato effetto di digestione con proteinasi K, l'attività peptidiltransferasica delle particelle private di proteine è completamente inibita da un breve trattamento con ribonucleasi T l.
Esperimenti di elettroforesi bidimensionale su gel hanno mostrato che la componente proteica delle particelle sottoposte a processi d'estrazione comprende un numero di proteine variabile da due fino a otto (Mankin e Noller, 1994). La migrazione corretta di queste proteine, durante l'elettroforesi su gel, suggerisce la loro inaccessibilità alle proteasi. Questa singolare resistenza alla digestione e all'estrazione può essere spiegata con esperimenti di centrifugazione su gradiente di saccarosio: il materiale estratto sedimenta con due picchi, uno più pesante che sedimenta in corrispondenza delle subunità 50S non trattate, l'altro più leggero mostra un coefficiente di sedimentazione pari a 35S. Il picco di 50S mostra un contenuto proteico pari al 10-20% del normale e una piena attività nel saggio della peptidiltransferasi; il picco di 35S è virtualmente privo di proteine e inattivo. Questi dati si possono interpretare in due modi: si può ipotizzare che una o più proteine ribosomali siano coinvolte nella funzione catalitica, cosicché la loro rimozione determinerebbe una perdita di attività; oppure, si può supporre che una o più proteine siano necessarie per il corretto ripiegamento tridimensionale che determina la funzionalità dell'rRNA 23S, e che la loro rimozione si traduca in un mancato ripiegamento della struttura e, quindi, in una perdita di attività. Quest'ultimo modello renderebbe conto della forte variazione del coefficiente di sedimentazione osservata in seguito alla perdita delle poche proteine residue ed è avvalorato da esperimenti di microscopia elettronica i quali hanno dimostrato che le particelle estratte si compongono di una miscela di particelle sferoidali 50S-simili e di RNA non ripiegato (Mankin e Noller, 1994). La singolare resistenza di queste proteine alla digestione proteolitica può essere infine spiegata ipotizzando che i sei domini dell'rRNA 23S formino una gabbia attorno a esse; in accordo a questo modello, le proteine agirebbero da scheletro, all'interno della particella 50S, mediando l'interazione tra i domini dell'rRNA 23S e contribuendo, perciò, al ripiegamento su larga scala della particella nella sua forma compatta e fisiologicamente attiva.
Una coppia di basi appaiate tra tRNA e rRNA 235
Il fatto che la sequenza CCA all'estremità 3', universalmente conservata, sia assolutamente necessaria per un'interazione produttiva del peptidil-tRNA con il sito P del ribosoma, suggerisce che questi nucleotidi possano interagire con alcune sequenze, analogamente conservate, nell'rRNA 23S. Un dato importante a questo proposito è emerso dai dati dijootprinting chimico: due guanine conservate, la G2252 e la G2253, sono protette dall'attacco del chetossale, un agente antivirale, dal legame dell'intero peptidil-tRNA o del frammento contenente la sequenza CCA al sito P della subunità 50S (Moazed e Noller, 1989a; 1991b). Particolarmente rilevante è l'osservazione che la delezione della C75, ma non della A76, del tRNA inibisce la protezione dei due residui di G. Questi risultati, nel loro insieme, suggeriscono un possibile appaiamento tra le due C conservate nel tRNA e le due G dell'rRNA 23S. Questa ipotesi è stata verificata direttamente con esperimenti di genetica in vitro (Samaha et al., 1995): sono state introdotte mutazioni nelle posizioni 2252 o 2253 e si è poi saggiata, analizzando il caratteristico jootprinting del sito P nell'rRNA 23S, la capacità dell'estremità CCA di vari tRNA mutanti di interagire con i ribosomi mutati. I risultati ottenuti indicano chiaramente che il corretto legame nel sito P richiede l'appaiamento di tipo Watson-Crick tra la C74 del tRNA e la G2252 dell'rRNA 23S il quale, nell'interpretazione più semplice, avviene nel sito P. La struttura di questa insolita ansa ricca in G e del suo complesso con un oligonucleotide CCA è stata ottenuta mediante spettroscopia NMR (E.V. Puglisi e J.D. Puglisi, risultati non pubblicati). Probabilmente la comprensione delle sue proprietà strutturali contribuirà a chiarire questa cruciale interazione RNA-RNA.
L'importanza di questa interazione per la reazione della peptidiltransferasi è stata messa in evidenza con una strategia differente (Samaha et al., 1995). In questo caso è stato necessario disporre di una popolazione omogenea di ribosomi mutanti, escludendo la possibilità di una attività peptidiltransferasica di fondo dei ribosomi selvatici. Un possibile approccio prevede la ricostituzione in vitro della subunità 50S, utilizzando rRNA 23S mutato che può essere ottenuto facilmente mediante trascrizione in vitro con l'enzima T7-polimerasi. A questo punto, però, si è dovuto risolvere il problema dell'incapacità di un rRNA 23S prodotto con T7 di ricostituire subunità 50S attive.
Si è trovato che subunità attive possono essere ottenute mediante una ricostituzione chimerica facendo uso di un trascritto di T7 contenente la sequenza di circa 2300 nucleotidi al 5' dell'rRNA 23S, e un frammento naturale dell'rRNA 23S, ottenuto mediante taglio con l'RNasi H, contenente la sequenza di circa 600 nucleotidi dell'estremità 3'. Con questo approccio è stato possibile introdurre mutazioni nella regione del 5' e analizzarle, in seguito alla ricostituzione chimerica, mediante la 'reazione del frammento' di Momo, modificata in modo da renderla molto più sensibile. Questi esperimenti hanno confermato i risultati ottenuti negli esperimenti di legame: i substrati contenenti un sito P in cui la posizione 74 può formare un appaiamento di Watson-Crick con la base 2252 dell'rRNA 23S, sono decisamente più efficaci nella formazione del legame peptidico.
Questi dati sperimentali portano a due importanti conclusioni: innanzitutto esiste un'interazione del tipo WatsonCrick tra la C74 del peptidil-tRNA e la G2252 dell'rRNA 23S nel sito P della subunità ribosomale 50S; in secondo luogo, è ora chiaro che l'rRNA 23S svolge un ruolo funzionale nella sintesi proteica, specialmente a livello della formazione del legame peptidico. Rimane ancora da chiarire come la C75 e la A76 del tRNA interagiscano con la subunità 50S, poiché i metodi di indagine finora utilizzati non sono stati in grado di mettere in evidenza un appaiamento del tipo Watson-Crick tra la C75 e la G2251 o la G2253, né tra la A76 e la U2585, una base che viene protetta dalla stessa A76 durante l'interazione del tRNA con il sito P (R. Green e R. Samaha, risultati non pubblicati; Moazed e Noller, 1989a).
Conclusioni
I dati ottenuti da numerosi laboratori sulle interazioni tra le due estremità del tRNA e gli rRNA delle due subunità fanno emergere un interessante quadro sulla natura della struttura dei siti funzionali del ribosoma, almeno per il sito P. In entrambi i casi, molteplici piccoli differenti elementi di struttura dell'RNA sembrano combinarsi per formare un sito funzionale. Questi elementi funzionali sono molto conservati dal punto di vista filogenetico, sia per quanto riguarda la struttura primaria che quella secondaria. Ancora più interessante, comunque, è il fatto che questi elementi funzionali siano molto dispersi all'interno dell'rRNA, separati spesso l'uno dall' altro da centinaia di nucleotidi, nonostante che i motivi strutturali del tRNA con cui reagiscono siano alquanto compatti. Come spiegare questi risultati? Una possibilità è che questi elementi dell'RNA siano residui dell'evoluzione molecolare, che potrebbero essersi evoluti a partire da strutture di RNA indipendenti, possibilmente funzionali, presenti nel mondo a RNA le quali, in seguito, si sono combinate per formare i predecessori dei grandi rRNA (Noller, 1993). Per esempio, dal modello elaborato per l'rRNA 16S, si evince che differenti elementi strutturali del sito P della subunità 30S convergano, come il pollice e le altre dita di una mano, per afferrare il braccio dell' anticodone del tRNA. Un' altra possibile ragione della dispersione di questi elementi funzionali nell'ampia struttura dell'rRNA è che essi (o almeno alcuni di essi) debbano muoversi uno rispetto all'altro durante la sintesi proteica. Questa visione del ribosoma come macchina a RNA è molto interessante sebbene, considerando le difficoltà incontrate finora in questi studi sul ribosoma, potrebbe rivelarsi estremamente difficile realizzarne una verifica sperimentale.
Bibliografia citata
ATMADJA, J., STEIGE, W., ZOBAWA, M., GREUER, B., OSSWALD, M., BRIMACOMBE, R. (1986) The tertiary folding of Escherichia coli 16S RNA, as studied by in situ intra-RNA crosslinking of 30S ribosomal subunits with bis-(2-chloroethyl) methylamine. Nucleic Acids Res., l, 659-673.
BAKIN, A., KOWALAK, J.A., MCCLOSKEY, J.A., OFENGAND, J. (1994) The single pseudoridine residue in Escherichia coli 16S rRNA is located at position 516. Nucleic Acids Res., 22, 3681-3684.
BAKIN, A., OFENGAND, J. (1993) Four newly located pseudouridylate residues in Escherichia coli 23S ribosomal RNA are all at the peptidyl transferase center: analysis by the application of a new sequencing technique. Biochemistry, 32, 9754-9762.
BARTA, A., KUECHLER, E. (1983) Part ofthe 23s RNA located in the 13s RNA fragment is a constituent of the ribosomal peptidyl transferase centeI. FEBS Lett., 163,319-323.
BARTA, A., STEINER, G., BROSIUS, J., NOLLER, H.F., KUECHLER, E. (1984) Identification of a site on 23S ribosomal RNA located at the peptidyl transferase centeI. Proc. Nat. Acad. Sci. USA, 81,3607-3611.
BISPINK, L., MATTHAEI, H. (1973) Photoaffinity labeling of 23S rRNA in E. coli ribosomes with poly(U)-coded ethyl 2-diazomalonyl-Phe-tRNA. FEBS Lett., 37, 291-294.
BOUBLIK, M., OOSTERGETEL, G.T., WALL, lS., HAINFELD, J.F., RADERMACHER, M., W AGENKNECHT, T., VERSCHOOR, A., FRANK, J. (1985) Structure of ribosomes and their components by advanced techniques of electron microscopy and computer image analysis. In Structure, Junction and genetics of ribosomes, a c. di Hardesty B., Kramer G., New York, Springer Verlag, pp. 68-86.
BOWMAN, C.M., DAHLBERG, J.E., IKEMURA, T., KONISKY, J., NoMURA, M. (1971) Specific inactivation of 16S ribosomal RNA induced by colicin E3 in vivo. Proc. Natl. Acad. Sci. USA, 68, 964-968.
BREITMEYER, J.B., NOLLER, H.F. (1976) Affinity labeling of specific regions of 23S RNA by reaction of N-bromoacetyl-phenylalanyl-transfer RNA with Escherichia coli ribosomes. J. Mol. Biol., 101, 297-306.
BRIMACOMBE, R. (1992) Structure-function correlations (and discrepancies) in the 16S ribosomal RNA from Escherichia coli. Biochimie, 74, 319-326.
BRIMACOMBE, R., ATMADJA, J., STIEGE, W., SCHULER, D. (1988) A detailed model of the three-dimensional structure of Escherichia coli 16S ribosomal RNA in situ in the 30S subunit. J. Mol. Biol., 199, 115-136.
BRIMACOMBE, R., GREUER, B., MITCHELL, P., OSSWALD, M., RÌNKE-APPEL, J. SCHULER, D., STADE, K. (1990) Three-dimensional structure and function of Escherichia coli 16S and 23S rRNA as studi ed by cross-linking techniques. In The ribosome: structure, Junction and evolution, a c. di Hill W.E., Dahlberg A., Garrett R.A., Moore P.B., Schlessinger, D., Warner J.R., Washington, D.C., AmeI. Soc. for Microbiol., pp. 93-106.
BRIMACOMBE, R., MALY, P., ZWEIB, C. (1983) The structure of ribosomal protein. Prog. Nucleic Acid Res. Mol. Biol., 28, 148.
BROSIUS, J., DULL, T.J., NOLLER, H.F. (1980) Complete nucleotide sequence of a 23S ribosomal RNA gene from Escherichia coli. Proc. Natl. Acad. Sci. USA, 77, 201-204.
BROSIUS, J., PALMER, M.L., KENNEDY, P.I., NOLLER, H.F. (1978) Complete nucleotide sequence of a 16S ribosomal RNA gene from Escherichia coli. Proc. Natl. Acad. Sci. USA, 75, 4801-4805.
BROWNLEE, G.G., SANGER, F., BARRELL, B.G. (1967) Nucleotide sequence of 5S ribosomal RNA from Escherichia coli. Nature, 215, 735-736.
CAPEL, M.S., et al. (1987) A complete mapping ofthe proteins in the small ribosomal subunit of Escherichia coli. Science, 238, 1403-1406.
CARBON, P., EHRESMANN, C., EHRESMANN, B., EBEL, J.P. (1979) The complete nucleotide sequence of the ribosomal 16S RNA from Escherichia coli. Experimental details and cistron heterogeneities. Eur. J. Biochem., 100, 399-410.
CHAMBLISS, G., CRAVEN, G.R., DAVIES, J., DAVIS, K., KAHAN, L., NORMURA, M. a c. di (1980) Ribosomes: structure, Junction and genetics. Baltimora, University Park Press.
COOPERMAN, B.S., WEITZMANN, C.I., FERNANDEZ, C.L. (1990) Antibiotic probes of Escherichia coli ribosomal peptydiltransferase. In The ribosome: structure, Junction and evolution, a c. di Hill W.E., Dahlberg A., Garrett R.A., Moore P.B., Schlessinger D., Wamer J.R., Washington, D.C., AmeI. Soc. for Microbiol., pp. 491-501.
CRICHTON, R.R., ENGELMAN, D.M., HASS, J., KOCH, M.H.J., MOORE, P.B., PARFAIT, R., STUHRMANN, H.B. (1977) Contrast variation study of specifically deuterated Escherichia coli ribosomal subunits. ProC. Natl. Acad. Sci. USA, 74, 5547-5550.
CUNDLIFFE, E. (1981) Antibiotic inhibitors of ribosome function. In The Molecular Basis of Antibiotic Action., a c. di Gale, E.F., Cundliffe, E., Reynolds, P.E., Richmond, M.H., Waring, M.H., New York, Wiley, pp. 402-547.
DABBS, E.R. (1986) Mutant studies on the prokaryotic ribosome. In Structure, Function and Genetics of Ribosomes, a c. di Hardesty B., Kramer G., New York, Springer-Verlag, pp. 743-748.
DAMASCHUN, G., MULLER, U., BIELKA, H. (1979) Scattering studies of ribosomes and ribosomal components. Methods Enzymol., 59, 706-750.
DAMMEL, C.S., NOLLER, H.F. (1993) A cold sensitive mutation in 16S rRNA provides evidence for helical switching in ribosome assembly. Genes and Development, 7, 660-670.
DE GROOT, N., PANET, A., LAPIDOT, Y. (1971) The blinding of purified Phe-tRNA and peptidyl-tRNA to Escherichia coli. Eur. J. Biochem., 23, 523-527.
DE STASIO, E.A., MOAZED, D., NOLLER, H.F., DAHLBERG, A.E. (1989) Mutations in 16S ribosomal RNA disrupt antibioticRNA interactions. EMBO J., 8, 1213-1216.
EHRESMANN, C., STIEGLER, P., CARBON, P., VNGEWICKELL, E., GARRETT, R.A. (1981) The topography of the 5' end of 16S RNA in the presence and absence of ribosomal proteins S4 and S20. Eur. J. Biochem., 103, 439-446. Fox, G.E., WOESE, C.R. (1975) 5S RNA secondary structure. Nature, 256, 505-507.
FRANK, J., PENCZEK, P., GRASSUCCI, R., SRIVASTAVA, S. (1991) Three-dimensional reconstruction of the 70S Escherichia coli ribosome in ice: the distribution of ribosomal RNA. J. CelI Biol., 115, 597-605.
GARRETT, R.A., VNGEWICKELL, E., NEWBERRY, V., HUNTER, J., WAGNER, R. (1977) An RNA core in 30S ribosomal subunit of Escherichia coli and its structural and functional significance. CelI Biol. Int. Rep., 1, 487-502.
GIRSHOVICH, A.S., BOCHKAREVA, E.S., KRAMAROV, V.M., OVCHINNIKOV, Y.A. (1974) E. coli 30S and 50S ribosomal subparticle components in the localization region of the tRNA acceptor terminus. FEBS Lett., 45, 213-217.
GLOTZ, C., MUSSIG, J., GEWITZ, H.S., MAKOWSKI, I., ARAD, T., YONATH, A., WITTMANN, H.G. (1987) Three dimensional crystals of ribosomes and their subunits from eu- and archaebacteria. Biochem. Internat., 15, 953-960.
GOLDEN, B.L., RAMAKRISHNAN, V., WHITE, S.W. (1993) Ribosomal protein L6: Structural evidence of gene duplication from a primitive RNA-binding protein. EMBO J., 12, 4901-4908.
GORNICKI, P., NURSE, K., HELLMANN, W., BOUBLIK, M., OFENGAND, J. (1984) High resolution localization of the tRNA anticodon interaction site on the Escherichia coli 30S ribosomal subunit. J. Biol. Chem., 259, 10.493-10.498.
GROSJEAN, H.I., DE HENAU, S., CROTHERS, D.M. (1978) Physical basis for ambiguity in genetic coding interactions. Proc. Natl. Acad. Sci. USA, 75, 610-614.
GUTELL, R.R. (1994) Collection of small subuuit (16S and 16Slike) ribosomal RNA structures. Nucleic Acids Res., 22, 3502-3507.
GUTELL, R.R., GRAY, M.W., SCHNARE, M.N. (1993) A compilation oflarge subunit (23S and 23S-like) ribosomal RNA structures. Nucleic Acids Res., 21, 3055-3074.
GUTELL, R.R., LARSEN, N., WOESE, C.R. (1994) Lessons from an evolving rRNA: 16S and 23S rRNA structures from a comparative perspective. Microbiol. Revs., 58, 10-26.
GUTELL, R.R., WEISER, B., WOESE, C.R., NOLLER, H.F. (1985) Comparative anatomy of 16S-like ribosomal RNA. Prog. Nucleic Acid Res. Mo!. Biol., 32, 155-216.
GUTELL, R.R., WOESE, C.R. (1990) Higher order structural elements in ribosomal RNAs: pseudo-knots and the use of noncanonical pairs. Proc. Natl. Acad. Sci. USA, 87, 663-667.
HAMPL, H., SCHULZE, H., NIERHAUS, K.H. (1981) Ribosomal components from Escherichia coli 50S subunits involved in the reconstitution of peptidyl transferase activity. J. Biol. Chem., 256, 2284-2288.
HARDESTY, B., KRAMER, G. a c. di (1985) Structure, Junction and genetics of ribosomes. New York, Springer-Verlag.
HARDY, S.I.S. (1975) The steichiometry ofthe ribosomal proteins of Escherichia coli. Mol. Gen. Genet., 140, 253-274.
HELD, W.A., MIZUSHIMA, S., NOMURA, M. (1973) Reconstitution of Escherichia coli 50S ribosomal subuuits from purified molecular components. J. Biol. Chem., 248, 5720-5730.
HILL, W.E., DAHLBERG, A., GARRETT, R.A., MOORE, P.B., SCHLESSINGER, D., WARNER, J.R. a c. di (1990) The ribosome: structure, function and evolution. Washington, D.C., AmeI. Soc. for Microbiol.
HOFFMAN, D.W., QUERY, C.C., GOLDEN, B.L., WHITE, S.W., KEENE, J.D. (1994) RNA-binding domain ofthe A protein component of the VI small nuclear ribonucleoprotein analyzed by NMR spectroscopy is structurally similar to ribosomal proteins. Proc. Natl. Acad. Sci. USA, 88, 2495-2499.
HOPFIELD, U. (1974) Kinetic proofreading-new mechanism for reducing errors in biosynthetic processes requiring high specifity. Proc. Natl. Acad. Sci. USA, 71, 4135-4139.
HUBBARD, J.M., HEAST, J.E., (1991) Computer modeling 16S ribosomal RNA. J. Mol. Biol., 221, 889-907.
KALTSCHMIDT, E., WITTMANN, H.G. (1970) Number ofproteins in small and large ribosomal subuuits of Escherichia coli as determined by two-dimensional electrophoresis. Proc. Natl. Acad. Sci. USA, 67, 1276-1282.
KIM, S.H., SUDDATH, F.L., QUlGLEY, G.J., Mc PHERSON, A., SUSSMAN, J.L., WANG, A., SEEMAN, N.C., RICH, A. (1974) Threedimensional tertiary structure of yeast phenylalanine transfer RNA. Science, 185, 435-440.
KYRIATSOULIS, A., MALY, P., GREUER, B., BRIMACOMBE, R., STOFFLER, G., FRANK, R., BLOCKER, H. (1986) RNA-protein crosslinking in Escherichia coli ribosomal subunits, localization of sites on 16S RNA wich are cross-linked to proteins S 17 and S21 by treatment with 2-iminothiolane. Nucleic Acids Res., 14, 1171-1186.
LAKE, J.A. (1980) Ribosome structure and functional sites. In Ribosomes: structure, Junction and genetics, a c. di Chambliss G., Craven G.R., Davies J., Davis K., Kahan L., Nomura M., Baltimora, Vniversity Park Press, pp. 207-236.
LAKE, J.A. (1982) Ribosomal subunit orientations determined in the monomeric ribosome by single- and by double-Iabeling immune electron microscopy. J. Mol. Biol., 161, 89-106.
LASATER, L.S., MONTESANO-RODITIS, L., CANN, P.A., GLITZ, D.G. (1990) Localization of an oligodeoxynucleotide complementing 16S ribosomal RNA residues 520-531 on the small subunit of Escherichia coli ribosomes, electron microscopy of ribosomecDNA-antibody complexes. Nucleic Acids Res., 18, 477-485.
LEFFERS, H., KIEMS, J., OSTERGAARD, L., LARSEN, N., GARRETT, R.A. (1987) Evolutionary relationships amongst archaebacteria. A comparative study of 23S ribosomal RNAs ofa sulphurdependent extreme thermophile, an extreme halophile and a thermophilic methanogen. J. Mol. Biol., 195, 43-61.
LEIJONMARCK, M., APPELT, K., BADGER, J., LILJAS, A., WILSON, K.S., WHITE, S.W. (1988) Structural comparison ofthe prokaryotic ribosomal proteins L 7 /L12 and L30. Proteins Structure, Function and Genetics, 3, 243-251.
LEIJONMARCK, M., ERICKSSON, S., LILJAS, A. (1980) Crystal structure of a ribosomal component at 2.6 Å resolution. Nature, 286, 824-826.
LI, M., TZAGOLOFF, A., VNDERBRINK-LYON, K., MARTIN, N.C. (1982) Identification of the paromomycin-resistance mutation in the 15S rRNA gene of yeast mitochondria. J. Biol. Chem., 257, 5921-5928.
LILJAS, A. (1982) Structural studies ofribosomes. Prog. Biophys. Mol. Biol., 40, 161-228.
LINDHAL, M., et al. (1994) Crystal structure ofthe ribosomal protein S6 from Thermus thermophilus. EMBO J., 13, 1249-1254.
MADEN, B.E.H., TRAUT, R.R., MUNRO, R.E. (1968) Ribosomecatalyzed peptidyl transfer, the polyphenyalanine system. J Mol. Biol., 35, 333-345.
MALHOTRA, A., HARVEY S.C. (1994) A quantitative model of the Escherichia coli 16S RNA in the 30S ribosomal subunit. J. Mol. Biol., 240, 308-340.
MANKIN, A.S., NOLLER, H.F. (1994) Comunicazione personale.
MOAZED, D., NOLLER, H.F. (1986) Transfer RNA shields specific nucleotides in 16S ribosomal RNA from attack by chemical probes. CelI, 47, 985-94.
MOAZED, D., NOLLER, H.F. (1987a) Interaction of antibiotics with functional sites in 16S ribosomal RNA. Nature, 327, 389-394.
MOAZED, D., NOLLER, H.F. (1987b) Chloramphenicol, erythromycin, carbomycin and vemamycin B protect overlapping sites in the peptidyl transferase region of 23S ribosomal RNA. Biochimie, 69, 879-884.
MOAZED, D., NOLLER, H.F. (1989a) Interaction oftRNA with 23S rRNA in the ribosomal A, P and E sites. CelI, 57, 585-597.
MOAZED, D., NOLLER, H.F. (1989b) Intermediate states in the movement of transfer RNA in the ribosome. Nature, 342, 142-148.
MOAZED, D., NOLLER, H.F. (1991a) Binding oftRNA to the ribosomal A and P sites protects two distinct sets of nucleotides in 16S rRNA. J. Mol. Biol., 211, 135-145.
MOAZED, D., NOLLER, H.F. (1991b) Sites of interaction ofthe Cca end of peptidyl-tRNA with 23S rRNA. Proc. Natl. Acad. Sci. USA, 88, 3725-3728.
MOAZED, D., ROBERTSON, J.M., NOLLER, H.F. (1988) Interaction of elongation factors EF-G and EF- Tu with a conserved loop in 23S RNA. Nature, 334, 362-364.
MOAZED, D., STERN, S., NOLLER, H.F. (1986) Rapid chemical probing of conformation in 16S ribosomal RNA and 30S ribosomal subunits using primer extension. J. Mol. Biol., 187, 399-416.
MONRO, R.E., MARCKER, K.A. (1967) Ribosome-catalyzed reaction of puromycin with a formylmethionine-containing oligonucleotide. J. Mol. Biol., 25, 347-350.
MOORE, P.B. (1980) Scattering studies of the three-dimensional organization of the E. coli ribosome. In Ribosomes: structure, function and genetics, a c. di Chambliss G., Craven G.R., Davies J., Davis K., Kahan L., Nomura M., Baltimora, University Park Press, pp. 111-134.
MOORE, V.G., ATCHISON, R.E., THOMAS, G., MORAN, M., NOLLER, H.F. (1975) Identification of a ribosomal protein essential for peptidyl transferase activity. Proc. Natl. Acad. Sci. USA, 72, 844-848.
MULLER, R., GARRETT, R.A., NOLLER, H.F. (1979) The structure of the RNA binding site ofribosomal proteins S8 and S 15. J. Biol. Chem., 254, 3873-3878.
MUSSIG, J., MAKOWSKI, I., VON BOHLEN. K., HANsEN, H., BARTELS, K.S., WITTMANN, H.G., YONATH, A. (1989) Crystals of wild type, mutate d, derivatized and complexed 50S ribosomal subunits from Bacillus stearothermophilus suitable for X-ray analysis. J. Mol. Biol., 205, 619-621.
NINIO, J. (1975) Kinetic amplification of enzyme discrimination. Biochimie, 57, 587-595.
NIRENBERG, M., LEDER, P. (1964) RNA codewords and protein synthesis-effect of trinucleotide upon binding of SRNA to ribosomes. Science, 145, 1399-1407.
NOLL, M., HAPKE, B., SCHREIER, M.H., NOLL, H. (1973) Structural dynamics of bacterial ribosomes 2. Characterization of vacant couples and their relation to complexed ribosomes. J. Mol. Biol., 75, 281-294.
NOLLER, H.F. (1991) Ribosomal RNA and translation. Ann. Rev. Biochem., 191-227.
NOLLER, H.F. (1993) On the origin ofthe ribosome: co-evolution of subdomains of tRNA and rRNA. In The RNA world, a c. di Gesteland R.F., Atkins lR., Cold Spring Harbor, Cold Spring Harbor Laboratory Press, pp. 137-156.
NOLLER, H.F., et. a!. (1981) Secondary structure model for 23S ribosomal RNA. Nucleic Acids Res., 9, 6167-6189.
NOLLER, H.F., HOFFARTH, v., ZIMNIAK, L. (1992) Unusual resistance of peptidyl transferase to protein extraction procedurees. Science, 256, 1416-1419.
NOLLER, H.F., LAKE, J.A. (1984) Ribosome structure and function, localization of rRNA. In Membrane structure and function, VoI. 6, a c. di Bittar E.E., New York, John Wiley and Sons, Inc., pp. 217-297.
NOLLER, H.F., WOESE, C.R. (1981) Secondary structure of 16S ribosomal RNA. Science, 212, 403-411.
NOMURA, M., MIZUSHIMA, S., OZAKI, M., TRAUB, P., LOWRY, C.V. (1969) Structure and function of ribosomes and their molecular components. Cold Spring Harbor Symp. Quant. Biol., 34, 49-61.
NOMURA, M., TISSIERES, A., LENGYEL, P., a c. di (1974) Ribosomes. Cold Spring Harbor, Cold Spring Harbor Laboratory Press.
OAKES, M.I., CLARK, M.W., HENDERsoN, E., LAKE, J.A. (1986) DNA hybridization electron microscopy, ribosomal RNA nucleotides 1392-1407 are exposed in the cleft of the small subunit. Proc. Natl. Acad. Sci. USA, 83, 275-279.
OAKES, M.I., SCHEINMAN, A., ATHA, T., SHANKWEILER, G., LAKE, lA. (1990) Ribosome structure: three-dimensionallocations of rRNA and proteins. In The ribosome: structure, function and evolution, a C. di Hill W.E., Dahlberg A., Garrett R.A., Moore P.B., Schlessinger D., Wamer J.R., Washington, D.C., AmeI. Soc. for Microbiol., pp. 180-193.
ODOM, O.W., HARDESTY, B. (1987) An apparent conformational change in transfer RNAPhe that is associated with the peptidyl transferase reaction. Biochimie, 69, 925-938.
ODOM, O.W., PICKING, W.D., HARDESTY, B. (1990) Movement of transfer-RNA but not the nascent peptide during peptide-bond formation in ribosomes. Biochem., 29, 10.734-10.744.
OLAH, T.V., PERRAULT, A.R., COOPERMAN, B.S., MONTESANO-RODITIS, L., Mc WILLIAMS, R., GLITZ, D.G. (1993) Incorporation of dinitrophenyl derivatives of proteins S6, S13, S16, and SI8 into the 30S subunit of Escherichia coli ribosomes by total reconstitution. J. Biol. Chem., 68, 18.696-18.700.
OSSWALD, M., GREUER, B., BRIMACOMBE, R., STOFFLER, G., BAUMERT, H., FASOLD, H. (1987) RNA-protein cross-linking in Escherichia coli 30S ribosomal subunits; determination of sites on 16S RNA that are cross-linked to proteins S3, S4, S5, S7, S8, S9, SII, S13, SI9 and SI by treatment with metyl p-azidophenyl acetimade. Nucleic Acids Res., 15, 3221-3240.
OUBRIDGE, C., HO, N., EVANS, P.R., TEO, C.H., NAGAI, K. (1994) Crystal structure at 1.92 Å resolution of the RNA-binding domain of the UlA spliceosomal protein complexed with an RNA hairpin. Nature, 372, 432-438.
PEATTIE, D.A., DOUTHWAITE, S., GARRETT, R.A., NOLLER, H.F. (1981) A 'bulged' double helix in a RNA-protein contact site. Proc. Natl. Acad. Sci. USA, 78, 7331-7335.
PENCZEK, P., RADERMACHER, M., FRANK, J. (1992) Three dimensional recostrunction of single particles embedded in ice. UItramicroscopy, 40, 33-53.
PETERSSON, I., HARDY, S.J.S., LILJAS, A. (1976) The ribosomal protein L8 is a complex of L7/Ll2 and LlO. FEBS Lett., 64, 135-138.
POLITZ, S.M., GLITZ, D.G. (1977) Ribosome structure: localization of N6, N6-dimethyladenosine by electron microscopy of a ribosome-antibody complex. Proc. Natl. Acad. Sci. USA, 74, 1468-1472.
POWERS, T., NOLLER, H.F. (1990) Dominant lethal mutations in a conserved loop in 16S rRNA. Proc. Natl. Acad. Sci. USA, 87, 1042-1046.
POWERS, T., NOLLER, H.F. (1991) A functional pseudoknot in 16S ribosomal RNA. EMBO J., 10, 2203-2214.
POWERS, T., NOLLER, H.F. (1993) Evidence for functional interaction between elongation factor Tu and 16S ribosomal RNA. Proc. Natl. Acad. Sci. USA, 90, 1364-1368.
POWERS, T., NOLLER, H.F. (1994) The 530 loop of 16S rRNA, a signal to EF-Tu? Trends Genet., 10, 27-31.
POWERS, T., NOLLER, H.F. (1995) Hydroxyl radical footprinting of ribosomal proteins on 16S rRNA. RNA, l, 194-209.
PRINCE, J.B., TAYLOR, B.H., THURLOW, D.L., OFENGAND, J., ZIMMERMANN, R.A. (1982) Covalent crosslinking oftRNA Val to 16S RNA at the ribosomal P site, identification of crosslinked residues. Proc. Natl. Acad. Sci. USA, 79, 5450-5454.
RAMAKRISHNAN, V., CAPEL, M., K.RELDGAARD, M., ENGELMAN, D.M., MOORE, P.B. (1984) Positions of proteins S14, SI8 and S20 in the 30S ribosomal subunit of Escherichia coli. J. Mol. Biol., 174, 265-284.
RAMAKRISHNAN, V., WHITE, S.W. (1992) The structure of ribosomal protein S5 reveals sites of interaction with 16S rRNA. Nature, 358, 768-77l.
RHEINBERGER, H.-J., STERNBACH, H., NIERHAUS, K. (1981) Three tRNA binding sites on Escherichia coli ribosomes. Proc. Natl. Acad. Sci. USA, 78, 5310-5314.
SAMAHA, R.R., GREEN, R., NOLLER, H.F. (1995) A base pair between tRNA and 23S rRNA in the peptidyl transferase centre of the ribosome. Nature, 377, 309-314.
SCHIMMEL, P., HENDERSON, B. (1994) Possible role of aminoacylRNA complexes in noncoded peptide synthesis and origin of coded synthesis. Proc. Natl. Acad. Sci. USA, 91, 11.283-11.286.
SCHWEDLER, G., ALBRECHT-EHRLICH, R., RAK, K.H. (1993) Immunoelectron microscopic localization of ribosomal proteins BS8, BS9, BS20, BL3 and BL21 on the surface of 30S and 50S subunits from Bacillus stearothermophilus. Eur. J. Biochem., 217, 361-369.
SENIOR, B.W., HOLLAND, I.B. (1971) Effect ofcolicin E3 upon the 30S ribosomal subunit of Escherichia coli. Proc. Natl. Acad. Sci. USA, 68, 959-963.
SPANGLER, E.A., BLACKBURN E.H. (1985) The nucleotide sequence of 17S ribosomal RNA gene of Tetrahymena thermophila and the identification of point mutations resulting in resistance to the antibiotics paromomycin and hygromycin. J. Biol. Chem., 260, 6334-6340.
SPIRIN, A.S. (1969) A model ofthe functioning ribosome: locking and unlocking of the ribosome subparticles. Cold Spring Harbor Symp. Quant. Biol., 34, 197-207.
STAEHELIN, T., MAGLOTT, D., MONRO, R.E. (1969) On the cataIytic site of peptidyl transfer, a part of the 50s ribosome structure. Cold Spring Harbor Symp. Quant. Biol., 34, 39-48.
STEINER, G., KUECHLER, E., BARTA, A. (1988) Photoaffinity labelling at the peptidyl transferase center reveals two different positions for the A- and P-sites in domain V of 23S rRNA. EMBO J., 7, 3949-3955.
STERN, S., POWERS, T., CHANGCHIEN, L.M., NOLLER, H.F. (1989) RNA-protein interactions in 30S ribosomal subunits, folding and function of 16S rRNA. Science, 244, 783-790.
STERN, S., WEISER, B., NOLLER, H.F. (1988) Model for threedimensional folding of 16S ribosomal RNA. J. Mol. Biol., 204, 447-48l.
STIEGE, W., ATMADJA, J., ZOBAWA, M., BRIMACOMBE, R. (1986) Investigation of the tertiary folding of Escherichia coli ribosomal RNA by intra-RNA cross-linking in vivo. J. Mol. Biol., 191, 135-138.
STOFFLER-MEILECKE, M., STOFFLER, G. (1990) Topography ofthe ribosomal proteins from Escherichia coli within the intact subunits as determined by immunoelectron microscopy and protein-protein crosslinking. In The ribosome: structure, function and evolution, a c. di Hill W.E., Dahlberg A., Garrett R.A., Moore P.B., Schlessinger D., Wamer lR., Washington, D.C., AmeI. Soc. for Microbiol., pp. 123-133.
STUHRMANN, H.B., KOCH, M.H.I., PARFAIT, R., HAAS, J., IBEL, K., CRICHTON, R.R. (1977) Shape of the 50S subunit of Escherichia coli ribosomes. Proc. Natl. Acad. Sci. USA, 74, 2316-2320.
SUBRAMANIAN, A.R. (1975) Copies of proteins L7 and Ll2 and heterogeneity of the large subunit of Escherichia coli ribosome. J. Mol. Biol., 95, 1-8.
SZEWCZAK, A.A., MOORE, P.B., CHANG, Y.L., WOOL, I.G. (1993) The conformation of the sarcin/ricin loop from 28S ribosomal RNA. Proc. Natl. Acad. Sci. USA, 90, 9581-9585.
TERAOKA, H., NIERHAUS, K.H. (1978) Protein Ll6 induces a conformational change when incorporated into an Ll6-deficient core derived from Escherichia coli ribosomes. FEBS Lett., 88, 223-226.
TERHORST, C.P., MOLLER, W., LAURSEN, R., WITTMAN-LIEBOLD, B. (1973) The primary structure of an acidic protein from 50S ribosomes of Escherichia coli wich is involved in GTP hydroIysis dependent on elongation factors G and T. Eur. J Biochem., 34, 138-152.
TRAKHANOV, S., YUSUPOV, M., SHIROKOV, V., GARBER, M., MITSCHLER, A., RUFF, M., THIERRY, J.C., MORAS, D. (1989) Preliminary X-ray investigation of 70S ribosome crystals from Thermus thermophilus. J. Mol. Biol., 209, 327-328.
TRAUB, P., NOMURA, M. (1969) Structure and function of Escherichia coli ribosomes. VI. Mechanism of assembly of 30S ribosomes studi ed in vitro. J. Mol. Biol., 40, 391-413.
TRAUT, R.R., LAMBERT, J.M., BOILEAU, G., KENNY, J.W. (1980) Protein topography of Escherichia coli ribosomal subunits are inferred from protein crosslinking. In Ribosomes: structure, function and genetics, a c. di Chambliss G., Craven G.R., Davies J., Davis K., Kahan L., Nomura M., Baltimora, University Park Press, pp. 89-110.
TRAUT, R.R., MOORE, P.B., DELIUS, H., NOLLER, H., TISSIERES, A. (1967) Ribosomal proteins of E. coli I. Demonstration of different primary structures. Proc. Natl. Acad. Sci. USA, 57, 1294-1301.
TREMPE, M.R., OHGI, K., GLITZ, D.G. (1982) Ribosome structure. Localization of 7-methylguanosine in the small subunits of Escherichia coli and chloroplast ribosomes by immunoelectron microscopy. J. Biol. Chem., 257, 9822-9829.
TRIMAN, K.L. (1995) Adv. Genet., 33.
VALGÀRD, K., MURRAY, J.B., STOCKLEY, P.G., STONEHOUSE, N.I., LILJAS, L. (1994) Crystal structure of an RNA bacteriophage coat protein-operator complex. Nature, 371, 623-626.
VASILIEV, V.D., SERDYUK, I.N., GUDKOV, A.T., SPIRIN, A.S. (1985) Self-organization of ribosomal RNA. In Structure, function and genetics ofribosomes., a c. di Hardesty B., Kramer G., New York, Springer Verlag, pp. 128-142.
VESTER, B., GARRETT, R.A. (1988) The importance of highly conserved nucleotides in the binding region of chloroamphenicol at the peptidyl transferase centre of E. coli 23S rRNA. EMBO J., 7, 3577-3587.
VON BOHLEN, K., MAKOWSKI, I., HANSEN, H.A., BARTELS, H., BERKOVITCH-YELLIN, Z., ZAYTZEV BASHAN, A., MEYER, S., PAULKE, C., FRANCESCHI, F., YONATH, A. (1991) Characterization and preliminary attempts for derivatization of crystals of large ribosomal subunits from Haloarcula marismortui diffracting to 3 Å resolution. J. Mol. Biol., 222, 11-15.
WAGENKNECHT, T., FRANK, J., BOUBLIK, M., NURSE, K., OFENGAND, J. (1988) Direct localization of the tRNA-anticodon interaction site on the Escherichia coli 30S ribosomal subunit by electron microscopy and computerized image averaging. J. Mol. Biol., 203, 753-760.
WALLER, J.P. (1964) Fractionation of the ribosomal protein from Escherichia coli. J. Mol. Biol., 10, 319-336.
WATANABE, S. (1972) Interaction of siomycin with acceptor site of Escherichia coli ribosomes. J. Mol. Biol., 67, 443-457.
WATSON, J.D. (1964) Synthesis of proteins upon ribosomes. Bull. Soc. Chim. Biol., 46, 1399-1425.
WEEKS, K.M., CROTHERS, D.M. (1993) Major groove accessibility of RNA. Science, 261, 1574-1577.
WEISS-BRUMMER, B., HUTTENHOFER, A. (1989) The paromycin resistance mutation (par r-454) in the 16S rRNA gene of the yeast Saccharomyces cerevisiae is involve in ribosomal frameshifting. Mol. Gen. Genet., 217, 362-369.
WEITZMANN, C.J., CUNNINGHAM, P.R., NURSE, K., OFENGAND, J. (1993) Chemical evidence for domain assembly ofthe Escherichia coli 30S ribosome. Faseb J., 7, 177-180.
WILSON, K.S., APPELT K., BADGER J., TANAKA I., WHITE S.W. (1986) Crystal structure of a prokaryotic ribosomal protein. Proc. Natl. Acad. Sci. USA, 83, 7251-7255.
WITTMANN, H.G. (1974) Purification and identification of Escherichia coli ribosomal proteins. In Ribosomes, a c. di Nomura M., Tissieres A., Lengye1 P., Cold Spring Harbor, Cold Spring Harbor Laboratory Press, pp. 93-114.
WITTMANN, H.G. (1982) Components of bacterial ribosomes. Annu. Rev. Biochem., 51, 155-183.
WITTMANN, H.G., LITTLECHILD, J.A., WITTMANN-LIEBOLD, B. (1980) Structure of ribosomal proteins. In Ribosomes: structure, function and genetics, a c. di Chambliss G., Craven G.R., Davies J., Davis K., Kahan L., Nomura M., Baltimora, University Park Press, pp. 51-88.
WITTMANN, H.G., WITTMANN-LIEBOLD, B. (1974) Chemical structure of bacterial ribosomal proteins. In Ribosomes, a c. di Nomura M., Tissieres A., Lengyel P., Cold Spring Harbor, Cold Spring Harbor Laboratory Press, pp. 115-140.
WOESE, C.R. (1980) Just-so stories and Rube Goldberg machines: speculations on the origin of the protein synthetic machinery. In Ribosomes: structure, function and genetics, a c. di Chambliss G., Craven G.R., Davies l, Davis K., Kahan L., Nomura M., Baltimora, University Park Press, pp. 357-373.
WOESE, C.R., FOX, G.E., ZABLEN, L., UCHIDA, T., BONEN, L., PECHMAN, K., LEWIS, B., STAHL, D. (1975) Conservation of primary structure in 16S ribosomal RNA. Nature, 254, 83-86.
WOESE, C.R., GUTELL, R., GUPTA, R., NOLLER, H.F. (1983) Detailed analysis of the higher-order structure of 16S-like ribosomal ribonucleic acids. Microbiol. Rev., 47, 621-669.
WOODCOCK, J., MOAzED, D., CANNON, M., DAVIES, J., NOLLER, H.F. (1991) Interaction of antibiotics with A- and P-site-specific bases in 16S ribosomal RNA. EMBO J., 10, 3099-3103.
WOWER, I., BRIMACOMBE, R. (1983) The localization of multiple sites on 16S RNA which are cross-linked to proteins S7 and S8 in Escherichia coli 30S ribosomal subunits by treatment with 2-iminothiolane. Nucleic Acids Res., 11, 1419-1437.
WOWER, J., HIXSON, S.S., ZIMMERMANN, R.A. (1989) Labeling the peptidyl transferase center of the Escherichia coli ribosome with photoreactive tRNAPhe derivatives containing azidoadenosine at the 3' end of acceptor arm: a model of the tRNAribosome complex. Proc. Natl. Acad. Sci. USA, 36, 5232-5236.
YUKI, A., BRIMACOMBE, R. (1975) Nucleotide sequences of Escherichia coli 16S RNA associated with ribosomal proteins S7, S9, S10, S14 and S19. Eur. J. Biochem., 56, 23-24.
YUSUPOV, M.M., GARBER, M.B., VASILIEV, V.D., SPIRIN, A.S. (1991) Thermus thermophilus ribosomes for crystallographic studies. Biochimie, 73, 887-89.
ZIMMERMANN, R.A. (1974) RNA-protein interactions in the ribosome. In Ribosomes, a c. di Nomura M., Tissieres A., Lengyel P., Cold Spring Harbor, Cold Spring Harbor Laboratory Press, pp. 225-270.
ZIMMERMANN, R.A. (1980) Interactions among protein and RNA components ofthe ribosome. In Ribosomes: structure,function and genetics, a c. di Chambliss G., Craven G.R., Davies J., Davis K., Kahan L., Nomura M., Baltimora, University Park Press, pp. 135-170.
ZIMMERMANN, R.A., MACKIE, G.A., MUTO, A., GARRETT, R.A., UNGEWICKELL, E., EHRESMANN, C., STIEGLER, P., EBEL, J.P., FELLNER, P. (1975) Location and characteristics of ribosomal protein binding sites in the 16S RNA of Escherichia coli. Nucleic Acids Res., 2, 279-302.
ZWIEB, C., BRIMACOMBE, R. (1979) RNA-protein cross-linking in Escherichia coli 30S ribosomal subunits: precise localization of the nucleotide in 16S RNA which is coupled to protein S7 by ultraviolet irradiation. Nucleic Acids Res., 6, 1775-1790.
Bibliografia generale
CHAMBLISS, G., CRAVEN, G.R., DAVIES, J., DAVIS, K., KAHAN, L., NOMURA, M. a c. di, Ribosomes: structure, function and genetics, Baltimora, University Park Press, 1980.
HARDESTY, B., KRAMER, G. a c. di, Structure, function and genetics of ribosomes. New York, Springer Verlag, 1985.
HILL, W.E., DAHLBERG, A., GARRETT, R.A., MOORE, P.B., SCHLESSINGER, D., WARNER, J.R. a c. di, The ribosome: structure, function and evolution, Washington, D.C., AmeI. Soc. for Microbiol., 1990.
NOMURA, M., TISSIERES, A., LENGYEL, P. a c. di, Ribosomes, Cold Spring Harbor, Cold Spring Harbor Laboratory Press, 1974.