
Immagini del linguaggio
Immagini del linguaggio
Secondo le teorie del 19° secolo, il linguaggio era localizzato in due piccole aree della corteccia nell'emisfero cosiddetto a dominanza linguistica. Studi recenti di visualizzazione hanno invece mostrato che molte regioni corticali, e perfino nuclei sottocorticali, contribuiscono alla elaborazione delle parole. È emerso che le parole con un contenuto significativo e quelle astratte con funzione grammaticale coinvolgono i due emisferi in misura diversa. Inoltre, si è trovato che i verbi attivano la corteccia motoria, mentre i sostantivi inducono attività in varie aree visive della corteccia occipitale e temporale interna. II diverso coinvolgimento di aree corticali, al di fuori delle aree classiche del linguaggio, si può spiegare sulla base dell'idea che la corteccia sia una enorme memoria associativa in cui i neuroni rafforzano le loro connessioni quando sono attivi contemporaneamente. L'attività simultanea porta probabilmente alla formazione di insiemi di cellule, distribuiti su ampie aree della corteccia, che potrebbero costituire la controparte organica delle parole e di altri elementi della elaborazione cognitiva.
Insiemi di cellule: gli elementi fondamentali della cognizione
Verso la fine degli anni Quaranta, il neuropsicologo Donald Hebb propose che elementi cognitivi, come figure o parole gestaltiche, siano rappresentati a livello corticale da gruppi di neuroni fortemente connessi. Il modello di Hebb fa tre ipotesi fondamentali che, se al tempo della loro formulazione erano molto teoriche, in seguito sono state tutte e tre confermate da dati sperimentali. La prima ipotesi è che i neuroni corticali rafforzino le loro connessioni quando risultano con frequenza attivi contemporaneamente. Questo principio di apprendimento associativo, che è a volte chiamato regola di Hebb, sembra essere valido per la maggioranza dei neuroni corticali (Gustafsson et al., 1987; Ahissar et al., 1992).
La seconda ipotesi è che la corteccia sia un'enorme memoria associativa in cui il rafforzamento delle sinapsi abbia luogo non solo tra neuroni vicini, ma anche tra neuroni in aree corticali distanti. Questa seconda ipotesi trae sostegno dagli studi neuroanatomici che evidenziano percorsi cortico-corticali tra molte aree della corteccia (Braitenberg e Schiiz, 1991; Deacon, 1992). Secondo la terza ipotesi, la contemporanea e frequente attivazione di un gruppo di neuroni che dà luogo al rafforzamento sinaptico ha conseguenze funzionali importanti. l neuroni fortemente connessi probabilmente agiscono insieme, come una unità funzionale. Se vengono attivati solo alcuni dei neuroni, si attiverà l'intero gruppo, a causa delle forti connessioni tra i membri del gruppo stesso. Se l'insieme è attivo, i suoi neuroni sono attivi simultaneamente, o mostrano per esempio schemi di funzionamento sincronizzati in modo preciso, quando l'attività neuronale si diffonde nell'insieme. Le registrazioni multiple di neuroni negli animali hanno fornito prove di un' attività neuronale specifica rispetto allo stimolo e simultanea (Fuster, 1994; Singer e Gray, 1995) o temporizzata in modo preciso (Abeles, 1991), anche tra neuroni appartenenti ad aree corticali distanti. Per esempio, si sono trovati neuroni, in diverse aree visive in entrambi gli emisferi, che oscillano quasi in sincronia quando vengono mostrate barre che si muovono in una particolare direzione, mentre non si è trovata traccia di attività coerente quando la direzione del movimento veniva leggermente cambiata (Engel et al., 1991a; 1991b). Questo conferma l'idea che reti di cellule corticali ampiamente distribuite, specifiche rispetto agli stimoli, possano comportarsi come unità funzionali. Gli insiemi hebbiani di cellule si possono definire come unità funzionali composte da molti neuroni che si formano in una rete associativa, la corteccia, come risultato di una frequente attività neuronale simultanea che causa un rafforzamento sinaptico.
Negli ultimi anni, l'idea hebbiana di insiemi distribuiti con topografie corticali definite è stata incorporata nelle teorie neuronali a grande scala del linguaggio e di altre funzioni cognitive (Braitenberg, 1978; Damasio, 1989; Abeles, 1991; Braitenberg e Schiiz, 1991; Elbert et al., 1994; Fuster, 1994; Mesulam, 1994; Wickens et al., 1994). Attualmente, sembra esservi un ampio consenso sul fatto che neuroni in aree corticali distanti possano cooperare comportandosi come unità funzionali. Lo schema hebbiano, però, non postula soltanto l'esistenza di reti neuronali su grande scala, ma fornisce anche criteri precisi per la formazione di questi insiemi di cellule e, quindi, dà una spiegazione per l'assetto topo grafico di questi insiemi.
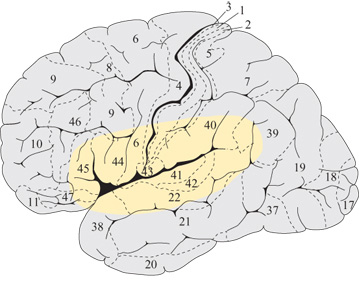
Riguardo alla loro formazione, Hebb (1949) traccia lo scenario seguente: se un certo oggetto viene percepito visivamente di frequente, si attiverà ripetutamente e contemporaneamente un insieme di neuroni nelle aree corti cali visive. Si formerà dunque un insieme di cellule che rappresenta la forma di quell'oggetto. Questo insieme risulta distribuito nelle regioni corticali, nelle quali la stimolazione visiva ha indotto un'attività neuronale simultanea e cioè nella corteccia visiva primaria e nelle aree visive superiori nei lobi occipitali, per esempio le aree di Brodmann 17, 18, 19 e 20 mostrate in figura (fig. 1). Se vi è un'attività neuronale correlata provocata da input che giungono attraverso altre modalità sensoriali, o se essa risulta associata a un output motorio, la distribuzione corticale degli insiemi di neuroni che vengono attivati sarà diversa. Per esempio, se un certo comportamento motorio risulta associato a una stimolazione sensoriale, si possono formare insiemi di cellule che comprendono neuroni sia nelle aree motorie sia in quelle sensoriali. Più in generale, la localizzazione corticale di una certa rappresentazione è una funzione del luogo della corteccia in cui si è presentata un'attività simultanea quando è stata acquisita o appresa quella rappresentazione.
Insiemi di cellule che rappresentano le forme di parola costituiti da neuroni della corteccia frontale e temporale
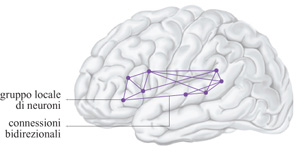
Poiché nel seguito ci occuperemo del linguaggio, è importante domandarsi dove si presenti, nella corteccia, un'attività neuronale correlata durante un'attività verbale nei primi stadi ontogenetici, nei quali ha luogo l'apprendimento del linguaggio (Pulvermiiller e Schumann, 1994). La pronuncia ripetuta delle sillabe da parte del bambino durante la cosiddetta fase della lallazione è controllata dall'attività neuronale nella corteccia motoria inferiore, premotoria e prefrontale. Inoltre, le articolazioni prodotte dal bambino stimolano il sistema uditivo in cui vengono attivati neuroni della corteccia uditiva primaria e di quelle superiori (Fry, 1966). Quindi, quando viene prodotta una sillaba, si avrà un'attività neuronaIe simultanea (o quasi simultanea) nelle regioni motorie e uditive primarie e superiori. Le aree primarie non mostrano forti connessioni cortico-corticali dirette. D'altra parte, i dati neuroanatomici sulle scimmie suggeriscono che queste aree siano connesse indirettamente attraverso aree anteriori a quella motoria e adiacenti a quella uditiva primaria (Pandya e Yeterian, 1985; Deacon, 1992). L'attività neuronale corre lata porta alla formazione di insiemi di cellule distribuiti su queste aree vicine alla scissura di Silvio, le cosiddette aree perisilviane (Braitenberg, 1980; Braitenberg e Pulvermilller, 1992; Braitenberg e Schiiz, 1992; Pulvermilller, 1992). La figura (fig. 2) mostra uno schema di un insieme perisilviano di questo tipo, che potrebbe rappresentare una sillaba prodotta di frequente o una forma verbale acquisita in una fase successiva dell'apprendimento del linguaggio.
Lo schema hebbiano implica che ogni Gestalt (struttura) o forma verbale è associata a un insieme corticale specifico, dato che la percezione di queste entità attiva popolazioni di neuroni diverse, anche se con possibilità di sovrapposizioni. Se un linguaggio non viene appreso attraverso le modalità vocali e uditive, ma attraverso quelle visive e manuali (linguaggio basato sui gesti), la localizzazione corticale degli insiemi di neuroni, che rappresentano gli elementi significativi, si modifica. Poiché i gesti vengono effettuati con le mani e percepiti con gli occhi, essi risultano connessi a un'attività neuronale in zone lontane dalla scissura di Silvio (aree motorie superiori e aree visive occipitali). Dunque, i gesti significativi che compongono il linguaggio saranno rappresentati nelle aree motorie, visive e associative, lontano dalla scissura di Silvio (Pulvermiiller, 1992).
Nell'ipotizzare che gli insiemi di cellule siano distribuiti nelle aree perisilviane, la prospettiva hebbiana è in evidente contrasto con i vecchi modelli di localizzazione, secondo i quali le rappresentazioni motorie e acustiche delle parole vengono immagazzinate in modo separato, rispettivamente nelle aree di Broca e di Wernicke (Wernicke, 1874; Lichtheim, 1885, Geschwind, 1970). La regione di Broca corrisponderebbe alle aree di Brodmann 44 e 45, mentre la migliore collocazione di quella di Wernicke (v. figura l) è forse la parte posteriore del lobo temporale superiore, nell'area 22 (Bogen e Bogen, 1976). La visione hebbiana prevede invece che la rappresentazione motoria e quella acustica di una parola non siano separate, ma fortemente connesse, in modo da formare un'unica unità funzionale distribuita. Affinché questa unità funzioni correttamente, sia le sue parti motorie sia quelle acustiche devono essere integre. Ciò è importante ai fini della spiegazione dell'afasia e in particolare del fatto che questo disturbo organico del linguaggio influenza di solito tutte le modalità attraverso le quali viene trasmesso il linguaggio stesso. In quasi tutti i casi noti, lesioni abbastanza grandi nelle aree del linguaggio posteriori o anteriori influenzano sia la produzione del parlato sia la sua percezione. Mentre i modelli localizzazionisti hanno grandi difficoltà nello spiegare questo fatto (Lichtheim, 1885), un modello basato su insiemi di neuroni può spiegare la multimodalità della maggior parte delle afasie. Ciò è stato dimostrato con simulazioni numeriche di deficit afasici, basate su modelli di insiemi di neuroni (Pulvermiiller e Preissl, 1991).
Essenzialmente, le lesioni nelle aree anteriori dellinguaggio non distruggono soltanto alcuni neuroni dell'insieme coinvolto, ma anche le fibre attraverso le quali l'insieme controlla il suo output motorio. Ciò provoca molteplici menomazioni con deficit più pronunciati nella produzione del parlato. Al contrario, le lesioni nelle aree posteriori del linguaggio distruggono alcuni neuroni dell'insieme e le connessioni uditive che risultano in input, spiegando così i deficit più pronunciati che si verificano nella comprensione. Quindi, l'ipotesi che la base della elaborazione dellinguaggio sia costituita da unità funzionali distribuite sulle aree perisilviane è in grado di spiegare importanti osservazioni cliniche. Tale ipotesi è inoltre corroborata da recenti ricerche di visualizzazione (imaging), che indicano la presenza di una attivazione simultanea di entrambe le aree del linguaggio quando viene percepita una parola (Zatorre et al., 1992; Mazoyer et al., 1993; Fiez et al., 1996).
Si noti che nello schema hebbiano la localizzazione dei meccanismi del linguaggio non viene spiegata esclusivamente sulla base dell'apprendimento associativo. l luoghi della corteccia in cui si ha un'attività simultanea durante prestazioni motorie e durante stimolazioni sensoriali dipendono dal cablaggio delle connessioni corti cali afferenti ed efferenti, che sono determinate geneticamente. l fattori genetici risultano importanti per la formazione dei fasci di fibre cortico-corticali, necessarie per una associazione a distanza tra neuroni di aree diverse che vengono attivati insieme. Inoltre, un approccio associazionista avrebbe difficoltà a spiegare perché, nella maggior parte dei destrimani, sia necessario l'emisfero sinistro, ma non il destro, per la elaborazione del linguaggio.
La 'dominanza per il linguaggio' dell'emisfero sinistro risulta evidente dallo studio degli effetti di lesioni in adulti e in bambini (Woods, 1983) e dagli esperimenti psicofisiologici nei bambini, che dimostrano come sia possibile registrare un'attività elettrocorticale specifica rispetto al linguaggio, maggiore nell' emisfero sinistro rispetto al destro (Molfese e Betz, 1988; Dehaene-Lambertz e Dehaene, 1994). Il corrispettivo neuroanatomico della lateralizzazione dellinguaggio si è visto nella dimensione delle aree perisilviane (Steinmetz et al., 1990; Galaburda et al., 1991) e nella dimensione (Hayes e Lewis, 1993), nell'ordinamento e nell'arborizzazione dendritica (Scheibel et al., 1985; Jacobs et al., 1993; Jacobs e Scheibel, 1993) delle cellule piramidali delle aree del linguaggio. Riguardo alle differenze in dimensione di particolari aree, appaiono molto importanti i processi epigenetici. È ben noto che differenze nella dimensione e nell'arborizzazione dendritica delle cellule possono dipendere dalla stimolazione sensoriale (Diamond, 1990) e, coerentemente, si è avanzata l'ipotesi (Previc, 1991) che la lateralizzazione del linguaggio sia dovuta a fattori ambientali, come la stimolazione uditiva unilaterale prenatale (v. il saggio di Il Hustler, M.E. Gillespie e M. Gazzaniga, in questo volume). Una stimolazione di questo tipo potrebbe essere alla base delle asimmetrie morfologiche sopra menzionate. Vi sono, d'altra parte, anche argomenti a favore di un contributo di fattori genetici alla lateralizzazione del linguaggio (Annett, 1979); attualmente, quindi, sembra prudente non eliminare la possibilità di un ruolo di tali fattori. Un substrato anatomico è necessario per la realizzazione dello schema hebbiano e non vi è dubbio che questo substrato sia determinato da fattori genetici. Ciononostante, partendo dal fatto che il cervello ha i suoi percorsi di input e di output preprogrammati, le sue proiezioni cortico-corticali definite e la sua preferenza dell'emisfero sinistro per il linguaggio, probabilmente determinata geneticamente, l'approccio hebbiano fornisce ipotesi molto specifiche sulla distribuzione corticale delle unità di elaborazione connesse al linguaggio. Rimane aperto il problema di capire quali proprietà anatomiche determinino la dominanza per illinguaggio e quali principi funzionali siano alla base della lateralizzazione. Alcune ipotesi al riguardo sono state avanzate da R. Miller (1987).
Significato della lateralizzazione del linguaggio
Una delle ipotesi che deriva dall'approccio hebbiano riguarda la realizzazione corticale della lateralizzazione dellinguaggio. Secondo i localizzazionisti, l'elaborazione dellinguaggio ha luogo in un solo emisfero. Lo schema hebbiano suggerisce invece una diversa visione.
Sebbene i fattori genetici o ambientali, o entrambi, portino a un'attivazione maggiore della corteccia perisilviana sinistra, in relazione alla produzione o alla percezione del linguaggio, l'articolazione di una parola è probabilmente controllata da un'attività delle regioni motorie in entrambi gli emisferi e la percezione acustica della parola produce necessariamente un' attivazione bilaterale delle aree uditive. Dato che in entrambi gli emisferi vi sono neuroni che vengono attivati contemporaneamente nella produzione o nella percezione di una parola, l'insieme di neuroni che rappresenta quella parola dovrebbe essere distribuito su entrambe le cortecce perisilviane (Mohr et al., 1994). Tuttavia, poiché sembra che i neuroni dell'emisfero sinistro rispondano con maggiore probabilità a stimoli linguistici e che controllino la precisione delle articolazioni, gli insiemi di neuroni che rappresentano una parola sarebbero lateralizzati a sinistra, nel senso che essi comprendono molti neuroni nell'emisfero sinistro e pochi nel destro. In questo modo, un insieme lateralizzato di cellule non si restringe, probabilmente, a un solo emisfero; infatti, potrebbe avvenire che un'alta percentuale dei neuroni dell'insieme sia localizzata nell'emisfero 'dominante' e una percentuale minore nell'emisfero 'non dominante', nel caso tipico di un destrimano. Per fare un esempio arbitrario, nel caso di un destrimano, un insieme di cellule che rappresenta una parola potrebbe avere il 70% dei suoi neuroni nell' emisfero sinistro e il 30% nel destro.
Rappresentazione di parole concrete e astratte da parte di insiemi di cellule con gradi diversi di lateralità
L'apprendimento associativo potrebbe non solo essere rilevante per la rappresentazione corticale delle parole, ma giocare un ruolo importante nell'acquisizione del significato delle stesse. Se si sta acquisendo il significato della parola "pesce", di solito la persona riceve stimoli con diverse modalità legati a questo significato. La persona può per esempio vedere, annusare, assaggiare, toccare o prendere un pesce mentre sente la parola "pesce". Queste percezioni e azioni multimodali sono legate ad attività neuronali in diverse aree corticali. Quindi, neuroni in diverse aree sensoriali e motorie si attivano contemporaneamente a quelli dell'insieme perisilviano che rappresenta la parola. Secondo la logica hebbiana, i neuroni che sono connessi tra loro e si attivano spesso contemporaneamente rinforzano le loro connessioni e si strutturano in un insieme di ordine superiore che rappresenta la parola dal punto di vista fonetico, insieme al suo significato. Dopo la formazione dell'insieme, il segnale fonetico è sufficiente ad attivare l'intero insieme, compresa la rappresentazione semantica, e, viceversa, l'insieme può essere attivato dal solo input semantico. In questo modo, la frequente attivazione contemporanea e la correlazione della parola e degli stimoli legati al significato sono necessarie solo nella fase del processo di acquisizione. In seguito, le forti connessioni di ordine superiore all'interno dell'insieme garantiscono l'attivazione dell'intero insieme quando ne viene attivata una sola parte e, quindi, garantiscono un grado elevato di correlazione tra le attività di tutte le parti dell'insieme e perciò la stabilità dell'insieme stesso.

Quando la forma fonetica della parola acquista significato, si possono innescare processi corticali molto diversi, a seconda del tipo di informazione che viene depositata nella rete associativa. La logica associazionista hebbiana suggerisce che parole che corrispondono a tipi diversi di vocaboli abbiano rappresentazioni corticali radicalmente diverse. Discuteremo in seguito di alcune di queste differenze. È probabile che i neuroni attivati da stimoli connessi al significato di parole con un contenuto concreto (sostantivi, aggettivi e verbi) siano ospitati in entrambi gli emisferi. Per esempio, la percezione visiva di un pesce attiverà probabilmente un uguale numero di neuroni nei due emisferi, perché è ugualmente probabile che il pesce sia percepito dai campi visivi destro e sinistro. Sebbene gli insiemi che rappresentano le forme fonetiche delle parole siano fortemente lateralizzati a sinistra, gli insiemi che rappresentano il contenuto delle parole saranno molto meno lateralizzati. Seguendo l'esempio sopra menzionato, un insieme dedicato alla forma delle parole potrebbe avere il 70% dei suoi neuroni nell'emisfero sinistro e solo il 30% nel destro. Dopo che il significato della parola è stato appreso e che altri neuroni in entrambi gli emisferi si sono accoppiati a quell'insieme, il suo grado di lateralità si ridurrà. In questo caso, per esempio, solo il 60% dei neuroni sarà ospitato ne II 'emisfero sinistro e il 40% nel destro. Nella figura (fig. 3) vengono schematizzati insiemi con diverso grado di lateralizzazione.
Non tutte le parole hanno significati concreti e facilmente raffigurabili. Le differenze di significato più estreme si trovano tra le parole con un contenuto concreto come "pesce" e le parole che svolgono principalmente una funzione grammaticale, come "esso" o "il". Tra queste parole cosiddette funzionali (o a classe chiusa) si annoverano i pronomi, i verbi ausiliari, le congiunzioni e gli articoli. Non sarebbe corretto affermare che queste parole non abbiano alcun significato. Il loro significato, piuttosto, appare molto più complesso rispetto a quello dei sostantivi concreti (Wittgenstein, 1967). L'uso di parole funzionali si può apprendere solo in contesti discorsivi, in cui queste si possono presentare in associazione con una grande varietà di altre parole e di stimoli non linguistici. Questo vuoI dire che, rispetto alla rappresentazione interna cerebrale di tali parole, non vi è un'attività fortemente correlata di un insieme perisilviano che rappresenti la parola, né di neuroni situati in altre aree corticali. Si noti che per la maggior parte delle parole contenuto, almeno per quelle con un significato concreto e facilmente raffigurabile, vi sono motivi per supporre una forte correlazione tra la presentazione delle parole e la presentazione di stimoli multimodali e, quindi, tra l'attività nelle aree perisilviane e quella all'esterno di esse. Al contrario, per le parole funzionali questa correlazione è probabilmente molto bassa. Non vi sono quindi motivi per supporre che la topografia corticale dell'insieme si modifichi durante l'acquisizione di queste parole. Una parola funzionale grammaticale come "esso" o "il" potrebbe quindi essere rappresentata a livello corticale in modo simile alla forma fonetica delle parole. L'insieme corrispondente sarà probabilmente ristretto alle aree perisilviane e risulterà fortemente lateralizzato nell'emisfero sinistro. Le parole con contenuto concreto e quelle con pura funzione grammaticale presentano grandi differenze nel tipo di significato. Le parole di contenuto astratto, come "libertà" o "giustizia", si collocano a un livello intermedio. Non è possibile descrivere visivamente il loro significato, ma si possono raffigurare visivamente scene associate al loro significato, per esempio una statua o i piatti di una bilancia. Probabilmente questa associazione tra la presentazione di parole astratte e quella di stimoli non linguistici di questo tipo legati al significato è rara. Tuttavia, la correlazione tra la percezione della forma fonetica di queste parole e la percezione di oggetti non è forse così bassa come nel caso delle parole con funzione grammaticale, che vengono usualmente percepite e usate in associazione a ogni sorta di stimoli possibili. In questo modo, la correlazione tra gli stimoli non linguistici e le forme fonetiche delle parole appare abbastanza alta per le parole con contenuto concreto, molto più bassa per quelle di contenuto astratto e ancora più bassa per le parole funzionali. Questo fatto suggerisce che, mentre le parole con contenuto concreto e quelle funzionali corrisponderebbero, rispettivamente, a insiemi debolmente e fortemente lateralizzati, il grado di lateralizzazione degli insiemi che rappresentano parole di contenuto astratto si può ipotizzare intermedio tra gli altri due. La stessa osservazione suggerisce l'ipotesi che questi insiemi differiscano anche rispetto alla percentuale di neuroni situati al di fuori delle aree perisilviane. Le rappresentazioni di parole di contenuto concreto avrebbero una percentuale alta di neuroni al di fuori delle aree perisilviane del linguaggio, mentre questa percentuale sarebbe più bassa per gli insiemi di cellule corrispondenti alle parole di contenuto più astratto e le parole funzionali avrebbero solo pochissimi, o nessuno, dei neuroni corrispondenti localizzati al di fuori delle aree perisilviane.
Diversità di percezione tra parole
Le parole contenuto vengono usate per riferirsi a odori, sapori, sensazioni somatiche, percezioni visive o azioni motorie. Durante l'apprendimento del linguaggio, le forme di parola vengono spesso prodotte quando il bambino percepisce gli stimoli o compie le azioni cui le parole stesse si riferiscono. Se la corteccia è una memoria associativa, i canali di elaborazione attraverso i quali le informazioni legate a un significato vengono trasmesse e le modalità di trasmissione devono essere importanti ai fmi della formazione degli insiemi corticali di cellule. Questa idea associazionista ha ispirato recentemente modelli di elaborazione della parola nel cervello che postulano rappresentazioni corticali distinte per classi di parole che possano essere differenziate sulla base di criteri semantici (Warrington e McCarthy, 1987).

Vi sono parole che stimolano forti associazioni visive, mentre altre tendono a richiamare alcune attività del proprio corpo. Chiameremo queste due categorie parole visive e motorie, rispettivamente. Esempi tipici di parole visive sono i sostantivi concreti con significato facilmente raffigurabile (il libro ). Le parole motorie più tipiche si trovano probabilmente tra i verbi di azione (afferrare). Si noti, tuttavia, che in inglese la maggior parte dei sostantivi può essere usata come verbo e viceversa. Per esempio, la parola inglese catch (afferrare) può significare collection (raccolta), e book (libro) può a volte essere un sostituto di reserve (prenotare). Ciononostante, le parole catch e book possono stimolare associazioni motorie o visive quando vengono presentate in un contesto neutro e può essere verosimile che quando se ne impara il significato siano coinvolti principalmente gesti motori o stimoli visivi. Sebbene associazioni occasionali stimolo-risposta non siano certamente sufficienti per la completa acquisizione del significato della parola (Gleitrnan e Wanner, 1982) - si potrebbe dire che non permettono di distinguere tra la stella del mattino e la stella della sera (Frege, 1980) - , tali associazioni potrebbero ugualmente avere implicazioni importanti per il funzionamento interno del cervello. Processi neuronali diversi possono aver luogo durante l'apprendimento del significato delle parole, sulla base dell' attività nei sistemi visivi e motori. Le popolazioni neuronali della corteccia visiva e di quella del linguaggio vengono probabilmente attivate insieme quando vengono acquisiti i significati di parole visive, così come insiemi di neuroni nelle aree motorie e del linguaggio possono attivarsi contemporaneamente durante l'apprendimento del significato di parole che si riferiscono a movimenti del proprio corpo. Secondo l'approccio hebbiano, gli insiemi che rappresentano parole motorie sono quindi distribuiti nelle aree perisilviane e in aree supplementari motorie e prefrontali, mentre le parole visive sono rappresentate da insiemi comprendenti neuroni delle aree perisilviane e visive nei lobi occipitali, parietali e temporali inferiori. La figura (fig. 4) mostra uno schema di questi tipi di insiemi.
Si può ragionare in modo analogo per parole che si riferiscono di solito a stimoli percepiti attraverso altre modalità. Per questi casi si possono ipotizzare ulteriori categorie di parole: parole di odore, sapore, dolore, tatto e suono. l membri di queste classi sono rappresentati da insiemi con topografie corticali specifiche. Per esempio, mentre un insieme che rappresenta una parola di dolore o tatto può avere una frazione rilevante dei suoi neuroni nelle aree somatosensoriali, le parole di suono possono comprendere nei loro insiemi rappresentativi un elevatissimo numero di neuroni appartenenti alle aree uditive. Sottolineiamo di nuovo che i neuroni che rispondono a stimoli che si presentano con diverse modalità e i neuroni che controllano movimenti e azioni motorie sono localizzati in entrambi gli emisferi. Dunque, gli insiemi che rappresentano queste parole dovrebbero essere distribuiti su entrambi gli emisferi ed essere meno lateralizzati rispetto agli insiemi che rappresentano parole funzionali.
La definizione delle parole motorie è particolarmente delicata, perché non tutte le associazioni legate al movimento coinvolgono modalità motorie. È importante distinguere tra i movimenti che possono essere compiuti dal proprio corpo e quelli che di solito vengono percepiti visivamente. Le parole "volare" o "aeroplano", per esempio, possono essere ascoltate frequentemente da un bambino mentre percepisce certi stimoli visivi in movimento. Per quanto non si possa negare una relazione tra lo stimolo visivo e la modalità motoria, visto che la percezione dello stimolo visivo è accompagnata di solito da movimenti oculari legati all'attività neuronale frontale, questa attività neuronale è probabilmente non molto specifica rispetto allo stimolo (movimenti oculari di questo genere si compiono nel guardare diversi oggetti). Quindi, la correlazione tra i pattern visivi di input e l'occorrenza delle parole "volare" o "aeroplano" può essere maggiore e queste parole potrebbero corrispondere a insiemi di cellule che comprendono un numero elevato di neuroni nelle aree visive che rispondono a specifici contorni in movimento. Dovremmo dunque classificare queste parole non come parole motorie, ma come parole visive di un certo tipo (parole visive di movimento). Le parole che si riferiscono di solito a movimenti del proprio corpo, cioè quelle motorie vere e proprie, potrebbero anche comprendere, negli insiemi che le rappresentano, dei rivelatori di movimento, poiché molti movimenti compiuti dal corpo vengono percepiti visivamente mentre vengono compiuti e si possono usare parole motorie anche per riferirsi ad azioni compiute da altri. Da questo si capisce che lo schema della figura 4 fornisce un quadro troppo rozzo degli insiemi di cellule che rappresentano le parole motorie: tali insiemi dovrebbero anche includere neuroni nelle aree visive che elaborano principalmente le informazioni sul movimento, molti dei quali sono probabilmente localizzati nel giro temporale posteriore medio (Zeki et al., 1991). Considerazioni analoghe possono essere fatte per gli stimoli somatosensoriali provocati dai movimenti del corpo e da ciò si può suggerire che agli insiemi che rappresentano le parole di azione si debbano aggiungere neuroni delle aree parietali.
Sulla base delle aree corticali che risultano operanti durante attività motorie legate al significato, si possono delineare altre distinzioni tra le classi di parole; è possibile distinguere diversi tipi di parole motorie sulla base dei particolari muscoli che risultano più importanti per effettuare il movimento (masticare, scrivere, dare un calcio), della complessità del movimento (bussare, scrivere), del numero di muscoli coinvolti (annuire, abbracciare). Questi fattori possono 'spostare' i neuroni dei lobi frontali, aggiunti all'insieme perisilviano, in direzione inferiore-posteriore (rappresentazione della bocca-mano-piede) o anteriore-posteriore (movimenti semplici-complessi), o possono 'ampliare' o 'ridurre' la loro distribuzione corticale (molti o pochi muscoli coinvolti nel movimento).
Anche per le parole visive si potrebbero desiderare analoghe distinzioni più sottili. Alcune parole visive si riferiscono a oggetti statici (casa), altre a oggetti in movimento (treno); alcune si riferiscono a colori od oggetti colorati (iguana), altre a oggetti non colorati (pinguino) e, inoltre, alcuni stimoli visivi sono molto semplici (una linea) mentre altri sono più complessi (un quadrato, un cubo, una casa, una città, una metropoli). Questo fatto suggerisce che diversi gruppi di neuroni si aggiungano all'insieme quando si vengano imparate nuove associazioni tra parole e stimoli diversi. L'insieme che corrisponde a una parola che si riferisce abitualmente a colori od oggetti colorati può comprendere neuroni che rispondono in modo molto elevato al colore; inoltre, come abbiamo già discusso, anche neuroni sensibili a stimoli visivi in movimento possono essere compresi negli insiemi che rappresentano parole che si riferiscono a questi stimoli. Recentemente sono stati scoperti flussi di elaborazione nei lobi temporali che riguardano principalmente le informazioni di colore e movimento provenienti dagli ingressi visivi (Zeki et al., 1991) e si è trovato che alcune aree si specializzano nella elaborazione di facce o di sequenze di lettere (McCarthy et al., 1993; Allison et al., 1994; Puce et al., 1995). Sebbene si discuta ancora sulle prove dell'esistenza di flussi di elaborazione di questo tipo, è possibile che il cablaggio delle aree visive determini il fatto che le cellule in grado di rilevare il movimento siano più frequenti in un'area, per esempio nel lobo temporale medio, e che, invece, i neuroni nella corteccia visiva primaria e secondaria (che rispondono al colore) proiettino segnali preferenzialmente verso altre aree, per esempio il lobo temporale inferiore. Questo suggerisce, per esempio, che le parole che si riferiscono al colore o a oggetti colorati siano rappresentate da insiemi comprendenti anche neuroni nelle aree del colore (per esempio, nel giro temporale inferiore) e che le parole che si riferiscono ai movimenti percepiti visivamente corrispondano a insiemi comprendenti anche neuroni nelle aree del movimento visivo (nella corteccia temporale media).
Si noti che i tipi di parole definiti in questo modo non hanno necessariamente una categoria lessicale congruente: la maggior parte dei verbi, ma non tutti, sono parole motorie e vi possono essere parole motorie in altre categorie lessicali. Inoltre, dalle considerazioni teoriche non è sempre chiaro a quale categoria si debba assegnare una particolare parola. La maggior parte delle parole di contenuto concreto presenta probabilmente una correlazione elevata con stimoli di più modalità e il loro presentarsi può richiamare al soggetto alcuni stimoli multimodali. Inoltre, queste associazioni variano da soggetto a soggetto. Sarebbe quindi desiderabile disporre di dati sperimentali che consentano una classificazione delle parole in categorie di modalità e sarebbe ancor più opportuno valutare quantitativamente le associazioni motorie, visive e di altro tipo.
Previsioni sperimentali
La logica hebbiana suggerisce che parole che appaiono a prima vista molto simili abbiano in realtà equivalenti neurobiologici completamente diversi. Mentre le parole funzionali possono essere rappresentate da insiemi fortemente lateralizzati nella corteccia perisilviana, le parole di contenuto concreto corrispondono probabilmente a insiemi meno lateralizzati che comprendono neuroni di altre aree corticali. Per esempio, parole che ci ricordano principalmente attività motorie possono essere strutturate in insiemi che comprendono anche neuroni della corteccia motoria e parole visive, che stimolano la visualizzazione di oggetti o scene, possono corrispondere a insiemi che comprendono anche neuroni situati nella corteccia visiva. In questo modo la topografia dell'insieme è funzione della categoria semantica. Ci si può quindi attendere che: l) le parole funzionali comportino delle tracce di attività corticale lateralizzata a sinistra nelle aree perisilviane; 2) le parole contenuto comportino delle tracce di attività corticale meno lateralizzata nelle aree perisilviane e al di fuori di esse; 3) le parole motorie provochino anche un'attività nelle aree motorie dei lobi frontali (questa attivià frontale può essere connessa ad associazioni con movimenti del corpo. Come abbiamo discusso sopra, vi può essere un'attivtà supplementare nelle aree temporali e parietali medie connessa alla percezione visiva dei movimenti, sia quelli compiuti da noi stessi sia quelli compiuti da altri, e alla autostimolazione somatosensoriale durante l'effettuazione dei movimenti corporei); 4) le parole visive causino anche un'attività nelle aree visive nei lobi occipitale e temporale inferiore. Anche se si possono ricavare altre previsioni dalla discussione sulla lateralizzazione del linguaggio, nei prossimi paragrafi ci concentreremo sulle quattro ipotesi precedenti.
Alcune considerazioni metodologiche
In neurofisiologia si dispone di numerose tecniche di visualizzazione per indagare i processi cognitivi superiori. Si può visualizzare l'attività di grandi insiemi di neuroni utilizzando tecniche di registrazione elettro fisiologiche, come l'elettroencefalografia (EEG) e la magnetoencefalografia (MEG). Queste tecniche forniscono informazioni esatte circa la dinamica temporale dei processi elettro fisiologici di attivazione e deattivazione, che hanno luogo sulla scala dei millisecondi. Esse consentono anche la localizzazione delle sorgenti, sebbene questa sia di solito molto meno precisa in confronto alle tecniche di visualizzazione del metabolismo del cervello. Le tecniche di visualizzazione metabolica ad alta risoluzione spaziale, come la tomo grafia a emissione di positroni (PET, Positron Emission Tomography) e la visualizzazione a risonanza magnetica funzionale (tMRI, functional Magnetic Resonance Imaging), sono estremamente utili per localizzare le strutture del cervello che diventano più attive e, quindi, aumentano il loro ritmo metabolico durante compiti cognitivi. Comunque, i metodi metabolici forniscono solo un quadro rozzo della dinamica temporale dei processi cerebrali ed è quindi importante utilizzare sia le tecniche di visualizzazione metabolica sia quelle elettro fisiologiche quando si indaga sui processi cerebrali legati alle funzioni cognitive. Negli ultimi anni sono stati effettuati molti studi di visualizzazione sulla elaborazione delle parole, alcuni dei quali sono rilevanti al fine di valutare il modello hebbiano.
Quando si guarda ai risultati degli studi di visualizzazione, comunque, non si dovrebbe ignorare il fatto che molte informazioni importanti circa il dove, il perché e la forma nella quale hanno luogo i processi cognitivi si sono ottenute da studi che non usano l'attività cerebrale, ma il comportamento individuale come grandezza di riferimento. Gli studi sui pazienti neurologici con lesioni focali possono rispondere al quesito sulle strutture cerebrali che sono necessarie per particolari funzioni cognitive (Jackson, 1878). Gli studi su individui sani nei quali l'informazione sullo stimolo raggiunge solo un emisfero (per esempio, utilizzando la tecnica della presentazione tachistoscopica lateralizzata di stimoli visivi) possono fornire elementi importanti di comprensione del ruolo degli emisferi nella elaborazione del linguaggio. Inoltre, gli studi comportamentali, in cui si misurano esattamente i tempi e l'accuratezza delle risposte, hanno mostrato chiaramente che varie proprietà degli stimoli influenzano l'elaborazione dell'informazione nel cervello e molti di questi risultati ottenuti da studi comportamentali possono essere confermati da esperimenti psicofisiologici.
Le tecniche di visualizzazione con buona risoluzione spaziale si usano solo da pochi anni e, quindi, non sono stati ancora effettuati molti studi metodologici sulla influenza delle proprietà degli stimoli per mezzo di queste tecniche. Quando si valutano gli studi di visualizzazione della elaborazione della parola, è essenziale conoscere, sulla base di lavori comportamentali o di precedenti lavori psicofisiologici, le proprietà degli stimoli delle quali è nota una influenza sui processi cerebrali.
Le parole possono variare sotto diversi punti di vista. Tra gli altri fattori, la lunghezza, la frequenza con cui le parole vengono usate nel linguaggio comune, il grado in cui una parola stimola emotivamente i partecipanti all'esperimento e altri fattori psicologici possono fortemente influenzare le risposte psicologiche (Pulvermiiller, 1999). Inoltre, la ripetizione delle parole e il loro contesto possono avere una forte influenza sull'attività cerebrale provocata dalle parole stesse. Quando si indaga su processi cerebrali che distinguono tra classi di parole appare quindi necessario cercare di escludere, per quanto possibile, questi effetti, così come quelli legati alle ripetizioni delle parole, al priming (preattivazione) e al contesto sintattico e semantico della frase. Queste proprietà degli stimoli verbali possono confondere i risultati di qualsiasi studio di visualizzazione che intende evidenziare differenze nell'attività cerebrale provocata da due gruppi di parole. Solo se si escludono queste fonti di confusione si possono trarre conclusioni circa le differenze tra categorie lessicali o semantiche.
Diversi schemi di attività elettrocorticale negli emisferi suscitati da parole funzionali e parole contenuto
Sono stati effettuati molti studi basati su EEG alla ricerca di una diversa attività cerebrale coinvolta nella elaborazione di parole funzionali e contenuto. I primi esperimenti di S.M. Garnsey (1985) hanno messo in luce differenze sottili tra classi di parole nei potenziali evocati, attraverso l'analisi di componenti principali. H.I Neville, D.L. Mills e D.S. Lawson (1992) hanno presentato parole funzionali e parole contenuto nel contesto di frasi a soggetti ai quali era richiesto di indicare se le frasi avessero senso o meno. Le parole delle due classi non erano state bilanciate rispetto alla lunghezza e alla frequenza. Questi autori hanno riferito una componente lateralizzata a sinistra stimolata da parole funzionali, con un massimo a 280 ms dopo la comparsa dello stimolo, mentre le parole contenuto stimolavano un picco più simmetrico tra gli emisferi a 350 ms. Un risultato simile è stato ottenuto da A. C. Nobre e G. McCarthy (1994), i quali hanno usato stimoli bilanciati rispetto alla lunghezza ma non rispetto alla frequenza. Gli autori presentavano le parole una a una e i soggetti studiavano la sequenza cercando di rilevare parole di una particolare classe semantica. Di nuovo, un picco negativo lateralizzato a sinistra seguiva la presentazione di parole funzionali (con latenza di 288 ms), mentre le parole contenuto portavano a un incremento di negatività (con latenza di 364 ms) molto più simmetrico tra gli emisferi.
A. Gevins, B. Cutillo e M.E. Smith (1995) hanno usato un paradigma a due stimoli 'con suggerimento' in cui i soggetti dovevano indicare se tali stimoli erano simili in base a criteri fonetici, sintattici o semantici. È stata osservata una positività lateralizzata (con latenza di 445 ms) stimolata da parole funzionali, più pronunciata nelle regioni frontali sinistre, mentre le parole contenuto non stimolavano una risposta ritardata lateralizzata. Comunque, gli autori non riportano le lunghezze degli stimoli o le frequenze e non è quindi possibile escludere interferenze.
In un esperimento effettuato dal nostro gruppo (Pulvermiiller et al., 1995a) abbiamo presentato parole funzionali e parole contenuto bilanciate rispetto alla lunghezza e alla frequenza e abbiamo chiesto ai partecipanti allo studio di prendere decisioni lessicali veloci. Un'onda negativa con un massimo intorno a 160 ms dopo la comparsa dello stimolo ha mostrato un'interazione significativa tra gli emisferi e i fattori classe di parola. Il picco era ugualmente visibile su entrambi gli emisferi dopo la presentazione di parole contenuto, mentre era pronunciato sull'emisfero sinistro e ridotto sul destro quando venivano percepite parole funzionali. I potenziali evocati medi ottenuti tra 150 e 300 ms dopo la comparsa dello stimolo hanno rivelato la stessa interazione tra emisfero e classe di parola (differenza tra destra e sinistra forte per le parole funzionali, minima o assente per le parole contenuto).
È importante sottolineare alcune delle differenze tra questi studi. Per esempio, i compiti che i partecipanti dovevano svolgere erano molto diversi (decisioni lessicali, giudizi su frasi, ecc.). Malgrado queste diversità, tutti questi esperimenti hanno messo in luce differenze delle risposte elettrocorticali tra i principali tipi di vocaboli. I risultati degli studi di H.I. Neville e collaboratori (1992) e di A.C. Nobre e G. McCarthy (1994) sono stati molto simili. In entrambi i casi sono state trovate una componente precoce lateralizzata a sinistra in relazione alle parole funzionali e una componente simmetrica tra gli emisferi a seguito delle parole contenuto dopo un ritardo più lungo. Nei risultati di A. Gevins e collaboratori (1995) le parole funzionali portavano a una componente lateralizzata a sinistra che compariva molto più tardi rispetto a entrambi gli studi precedenti e, di nuovo, non vi era nessuna componente lateralizzata di questo tipo per le parole contenuto. F. Pulvermiiller e collaboratori (1995a) non hanno trovato nessuna differenza tra le classi di parole in termini di latenza dei potenziali evocati, ma questo studio ha confermato l'esistenza di una componente lateralizzata a sinistra stimolata da parole funzionali e di una componente simmetrica tra gli emisferi stimolata dalle parole contenuto.
La verifica di questi studi rispetto alle possibili fonti di confusione mostra che le parole venivano presentate nel contesto di una frase solo nell'esperimento di Neville, mentre gli effetti di contesto giocano probabilmente un ruolo minore negli altri studi. Il bilanciamento degli stimoli rispetto alla lunghezza delle parole è stato effettuato negli esperimenti di Nobre e McCarthy e di Pulvermiiller e collaboratori. Solo quest'ultimo studio ha usato parole funzionali e contenuto bilanciate rispetto alla frequenza. Tale questione è di particolare importanza per le differenze elettrocorticali tra le parole funzionali e contenuto, poiché i dati riferiti da J.W. King e M. Kutas (1995) indicano che le differenze riportate da Neville e collaboratori e da Nobre e McCarthy potrebbero derivare dalle frequenze delle parole. In effetti, dopo aver bilanciato gli stimoli rispetto alla frequenza, le differenze tra le classi di parole delle latenze dei potenziali evocati sono scomparse.
Ciononostante, le differenze nella lateralizzazione dei potenziali evocati tra le parole funzionali e quelle contenuto erano ancora presenti con stimoli bilanciati rispetto alla frequenza (Pulvermiiller et al., 1995a). Quindi sono le differenze nella lateralizzazione, piuttosto che le differenze nella latenza, che sembrano caratteristiche delle principali classi di parole.
Le differenze elettrocorticali tra le parole funzionali e le parole contenuto si possono usare per trarre conclusioni sulle rappresentazioni corticali di questi tipi di vocaboli. La componente lateralizzata stimolata dalle parole funzionali può essere connessa all'eccitazione di insiemi lateralizzati di cellule corti cali. Invece, i potenziali quasi simmetrici tra gli emisferi che seguono la presentazione di parole contenuto potrebbero indicare che i gruppi di neuroni attivati da queste parole sono meno lateralizzati. L'esistenza di insiemi lateralizzati di cellule che corrispondono alle parole funzionali e di insiemi più simmetrici che rappresentano le parole contenuto è stata ipotizzata sulla base dell' approccio he bbiano (v. figura 3).
Anche se si considerano questi dati come dimostrazione di una diversa distribuzione corticale di insiemi di cellule che rappresentano parole funzionali e parole contenuto, rimane ancora da chiarire quale proprietà dei tipi di parole si correli a queste differenze. Le parole funzionali e contenuto differiscono rispetto alla loro struttura fonetica, alle categorie sintattiche cui appartengono e, come abbiamo discusso, alloro significato. Sulla base dei dati attuali, non è possibile decidere con certezza quale di queste variabili sia rilevante ai fini della differenza psicofisiologica osservata. Secondo il modello hebbiano, le proprietà semantiche delle parole funzionali e delle parole contenuto determinano la distribuzione corticale dei loro insiemi di cellule. A conferma di questo punto di vista, si è trovato che anche i sostantivi con significato concreto e astratto stimolano diverse risposte elettro corti cali tra gli emisferi. Per i sostantivi astratti, si è trovata una differenza tra gli emisferi in un compito di decisione lessicale, mentre i sostantivi concreti stimolano risposte elettrocorticali simili su entrambi gli emisferi (Kounios e Holcomb, 1994). Potrebbe essere quindi la differenza semantica tra i sostantivi concreti rispetto a quelli astratti, e tra le parole contenuto rispetto a quelle funzionali, a essere connessa alla lateralità differenziale dei potenziali evocati. I risultati ottenuti finora sono coerenti con l'ipotesi di insiemi di cellule fortemente lateralizzati per le parole funzionali, insiemi debolmente lateralizzati per le parole di contenuto concreto e un modesto grado di lateralizzazione per gli insiemi che rappresentano parole di contenuto astratto (Pulvermiiller, 1999). Questi risultati suggeriscono che alcuni aspetti del significato delle parole hanno la loro controparte fisiologica nel grado di lateralizzazione delle risposte elettrocorticali e degli insiemi corticali.
Se la distribuzione corticale delle rappresentazioni delle parole è davvero determinata dalle vie corticali attraverso le quali viene trasmessa l'informazione legata al significato, le differenze nella localizzazione corticale dovrebbero non solo distinguere le rappresentazioni delle parole funzionali e delle parole contenuto ma, in più, le rappresentazioni delle parole che differiscono in termini di associazioni motorie e visive, come sostantivi e verbi, o nomi di animali e di utensili. Il modello hebbiano, e probabilmente ogni schema associazionista, suggerisce che è sempre una differenza semantica tra classi di parole che determina differenze nelle rappresentazioni corticali.
È della massima importanza, comunque, il fatto che sulla base di un modello associazionista hebbiano le differenze semantiche tra categorie di parole si possano usare per generare previsioni sulle aree corti cali coinvolte nella elaborazione di parole appartenenti a tali categorie. Come abbiamo discusso in precedenza, ci si può attendere che le parole che stimolano forti associazioni visive siano rappresentate ed elaborate nella corteccia perisilviana e in aree supplementari della corteccia visiva, le aree occipitale e inferotemporale, mentre le parole con forti associazioni motorie possono essere rappresentate in corrispondenza con le aree perisilviane e con aree supplementari motorie nel lobo frontale ed eventualmente con regioni parietali e temporali connesse alla percezione visiva e somatosensoriale dei movimenti corporei. I sostantivi concreti che si riferiscono ad animali o a grandi oggetti manufatti sono tipiche parole visive, i verbi che si riferiscono ad azioni effettuate da esseri umani sono tipiche parole motorie e i nomi di utensili sono un esempio di categoria che evoca forti associazioni sia visive sia motorie.
Dati ottenuti da tecnica PET sulla generazione dei verbi
Dopo gli studi fecondi di S. Petersen e collaboratori (1989) e R. Wise e collaboratori (1991), sono stati compiuti spesso studi di visualizzazione per mettere in luce indizi sulle differenze tra l'elaborazione dei sostantivi e dei verbi (v. anche il saggio di A.L. Roskies e S. Petersen, Visualizzare il funzionamento del cervello umano). Questi autori hanno usato la PET, utilizzata anche in molti studi più recenti, per misurare l'attività del cervello mentre i partecipanti all'esperimento leggevano sostantivi presentati visivamente (compito di lettura), oppure cercavano di generare verbi 'associati' ai sostantivi (compito di generazione dei verbi). Se, per esempio, viene presentata la parola "automobile", ci si può aspettare la generazione dei verbi "guidare" o "gareggiare". Ai fini della valutazione dei risultati, di solito si sottraggono le mappe dell' attività cerebrale derivanti dal compito di lettura da quelle derivanti dal compito di generazione dei verbi. Se vi sono differenze significative nel metabolismo cerebrale, si attribuiscono questi cambiamenti ai processi cognitivi necessari per la generazione dei verbi e non per la lettura dei sostantivi.
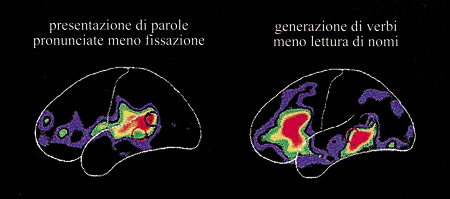
Sebbene non tutti gli studi concordino sulla localizzazione dell'incremento di attività durante la generazione dei verbi, sembra che si possa osservare un incremento di flusso sanguigno sia nelle aree prefrontali sia temporali. È stato riferito un incremento di attività nel lobo frontale sinistro, nell'area di Broca e nella regione anteriore e superiore rispetto a essa; nel lobo temporale sinistro, nell'area di Wernicke (area posteriore 22) e nel giro temporale medio si è osservato un incremento di flusso sanguigno (Petersen et al., 1989; Wise et al., 1991; Fiez et al., 1996). Si è dunque trovato, durante la generazione dei verbi, un incremento di attività nelle aree perisilviane del linguaggio e nelle aree premotorie, prefrontali e temporali medie. La figura (fig. 5) mostra i risultati di uno studio che mette in luce un'attivazione sia prefrontale sia temporale media durante la generazione di verbi connessa alla lettura di sostantivi.
Quando si interpretano questi risultati per trarre delle conclusioni sui processi cognitivi, si devono tener presenti alcune considerazioni. Come l'esempio precedente mostra chiaramente ("automobile" che porta a "guidare" o "gareggiare"), le parole generate non sono necessariamente verbi, in particolare se l'esperimento viene effettuato in inglese in cui molti verbi possono essere usati anche come sostantivi e viceversa. Non appare quindi opportuno chiamare questo un compito di generazione di verbi, quanto di generazione di parole motorie. Anche questo, però, potrebbe non essere giusto, poiché i soggetti potrebbero essere stati istruiti a descrivere cosa possono fare i sostantivi o per che cosa possono essere utilizzati (Fiez et al., 1996), consentendo così la generazione sia di parole motorie sia di parole visive connesse ai movimenti percepiti. Inoltre, sembra difficile interpretare questi risultati in termini psicologici, poiché da un confronto tra i compiti di generazione di parole e di lettura si vedono diverse differenze a livello cognitivo.
La generazione di parole motorie non necessita solo di processi semantici, ma richiede anche, per esempio, un processo di ricerca e porta probabilmente a un livello di attenzione maggiore rispetto alla lettura. Inoltre, nella maggior parte dei casi non viene fornita nessuna informazione sulle proprietà dello stimolo e della risposta che possano consentire una valutazione delle possibili fonti di confusione connesse alle proprietà delle parole.
Sulla base dei soli risultati PET non è quindi possibile attribuire i cambiamenti di flusso sanguigno alla elaborazione dei verbi. Ciononostante, assumendo che nella maggior parte dei casi vengano prodotte parole motorie, questi risultati appaiono coerenti con il seguente quadro: durante la generazione delle parole motorie, rispetto a quanto accade per il compito di lettura, viene attivato un insieme ulteriore di cellule, che comprende non solo neuroni nelle aree perisilviane ma, in più, neuroni nelle aree prefrontale, premotoria e temporale media. Questo è quello che ci si aspetterebbe sulla base dello schema associazionista discusso; tuttavia, da un punto di vista metodo logicamente più rigoroso, piuttosto che confrontare i sostantivi elaborati nel primo compito con i verbi elaborati nel secondo, appare necessario confrontare l'attività cerebrale durante l'elaborazione di parole motorie e visive nello stesso compito.
In recenti studi basati sulla PET (Martin et al., 1995), ai soggetti dell'esperimento sono stati presentati disegni acromatici di oggetti, da cui dovevano essere generati i nomi di azioni e parole di colore associati agli oggetti. Un confronto diretto dei pattern di attività, prodotti durante la generazione di queste categorie di parole, ha mostrato un incremento di ricambio metabolico nel lobo temporale ventrale quando venivano generate parole di colore. La generazione di parole di azione portava invece a una maggiore attività in aree temporali superiori sul giro temporale medio e in aree frontali inferiori, ma non nelle aree motorie. Questo però, come sottolineano gli autori, potrebbe riflettere il fatto che molte delle parole effettivamente generate dai soggetti sottoposti all'esperimento non si riferivano a movimenti che i soggetti potrebbero essi stessi compiere, ma piuttosto a movimenti di oggetti percepibili visivamente. Tra gli esempi di risposte elencati dagli autori vi sono i verbi "volare", "vedere", "dormire", a partire dai quali sono plausibili alcune associazioni visive, ma per i quali appare inappropriata una classificazione come parole motorie. Se sono stati usati molti verbi senza una associazione motoria, questo potrebbe essere il motivo per cui la generazione di queste parole ha attivato le aree visive e non le aree motorie connesse ai movimenti delle mani o dei piedi. Ciò fornisce un'ulteriore dimostrazione del fatto che è molto importante valutare con attenzione le proprietà psicologiche del materiale linguistico prima o durante gli studi psicofisiologici. È d'altra parte importante notare che parte del giro temporale medio sinistro era attiva durante la generazione dei verbi (Martin et al., 1995) e che si era già trovato che la stessa area, o un'area adiacente, risultava attiva durante la generazione dei verbi a partire da sostantivi presentati visivamente.
Risposte elettrocorticali diverse a sostantivi e verbi nelle aree motorie e visive
Le differenze tra le parole motorie e visive sono state studiate anche usando i potenziali evocati calcolati a partire dalle registrazioni di EEG. Questi studi hanno confrontato le risposte elettro corti cali ai sostantivi e ai verbi. Mentre i primi studi hanno riportato potenziali generalmente più positivi in connessione ai verbi in confronto ai sostantivi, lavori più recenti, che hanno utilizzato più elettrodi e tecniche di analisi più sofisticate, suggeriscono differenze tra le classi di parole in termini di topografie corti cali dei potenziali evocati. In uno studio che indagava i potenziali evocati da diverse classi di parole, S. Dehaene (1995) ha presentato numeri, sostantivi (nomi di animali e di persone) e verbi bilanciati rispetto alla lunghezza delle parole. Mentre i potenziali evocati dalle parole erano in generale maggiori nell'emisfero sinistro, si sono scoperte differenze tra le classi di parole, su entrambi gli emisferi, circa 300 ms dopo la comparsa dello stimolo. I verbi hanno stimolato una componente positiva lateralizzata a sinistra, massima su siti corticali frontali inferiori, che non si è trovata per i sostantivi. Sia i sostantivi che si riferiscono ad animali sia i verbi hanno prodotto negatività temporali sinistre quasi identiche. Questi risultati sono coerenti con l'ipotesi di un'attività supplementare frontale sinistra durante l'elaborazione dei verbi, ma non indicano nessuna differenza di elaborazione tra sostantivi e verbi in siti corticali posteriori. Dal punto di vista metodo logico si deve comunque notare che: non è stato effettuato nessun bilanciamento rispetto alla frequenza delle parole o ai valori di stimolo e valenza per i sostantivi e i verbi; i verbi, per un terzo, erano anche sostantivi comuni; gli stimoli venivano ripetuti nell'esperimento. Inoltre, non si è ottenuta nessuna informazione circa i processi cognitivi indotti dagli stimoli.
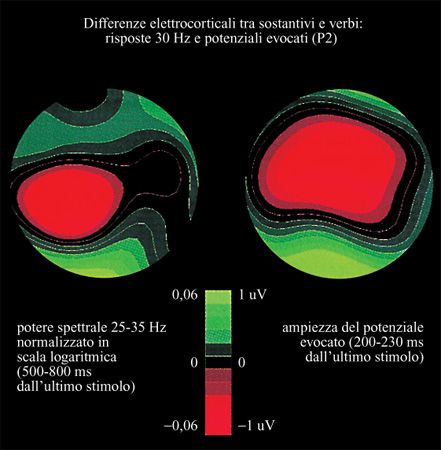
H. Preissl e collaboratori (Preissl et al., 1995; Pulvermiiller, 1999), presentando sostantivi e verbi bilanciati rispetto alla frequenza delle parole, alla lunghezza, ai valori di stimolo e di valenza in un compito di decisione lessicale, hanno trovato differenze corticali già a circa 200 ms dalla comparsa dello stimolo visivo. Nel confrontare i potenziali medi evocati dai sostantivi e dai verbi (tra 200 e 230 ms), si sono trovate differenze significative solo nella corteccia frontale. Dopo aver sottoposto i dati all'analisi di densità di corrente della sorgente (CSD, Current Source Density) per massimizzare il contributo al segnale dei generatori locali, si sono trovati segni elettro corti cali più marcati in relazione ai verbi sulle aree motorie bilaterali, mentre si è rilevata, per i sostantivi, una maggiore attività sulle aree visive nei lobi occipitali. È della massima importanza il fatto che si sono valutati con attenzione gli stimoli rispetto alle associazioni motorie e visive. Le classificazioni operate dai partecipanti all'esperimento hanno confermato differenze nelle associazioni coscienti dei movimenti corporei e delle scene visive provocate dalle parole che fungevano da stimolo. l verbi stimolavano associazioni motorie significativamente maggiori rispetto ai sostantivi e questi ultimi stimolavano associazioni visive maggiori rispetto ai verbi. Le differenze elettro corti cali osservate nelle aree visive e motorie si accordano con queste differenze nelle associazioni motorie e visive coscienti. Questi dati indicano che le parole motorie attivano un generatore supplementare vicino alle aree motorie, mentre le parole visive eccitano neuroni supplementari nelle aree visive primarie, o vicino a esse, nei lobi occipitali. Nell'immagine a sinistra nella figura 8 sono mostrate le differenze nelle topografie dei potenziali evocati dai sostantivi e dai verbi.
Si potrebbe argomentare che, per quanto sia improbabile in questo caso un'influenza dei fattori di confusione che abbiamo menzionato, non è chiaro se queste differenze elettrocorticali tra classi di parole siano connesse ad associazioni semantiche provocate dagli stimoli o piuttosto al fatto che gli stimoli appartenevano a diverse categorie lessicali (sostantivi e verbi). Si deve però notare che l'ipotesi che sia cruciale una differenza semantica può spiegare in parte le differenze trovate nelle topografie delle risposte cerebrali. Una tale spiegazione è possibile sulla base di principi associazionisti. D'altra parte, non vi è nessuna ragione a priori per cui membri di diverse categorie lessicali dovrebbero essere ospitati in diverse aree corti cali. Comunque, per dare ulteriore conferma dell'idea che sono le proprietà semantiche delle parole, e non le loro categorie lessicali, a essere essenziali ai fini dei pattern di attivazione corticale, è opportuno studiare stimoli provenienti dalla stessa categoria lessicale (sostantivi), che nondimeno evocano o principalmente associazioni visive (per esempio, nomi di animali) o associazioni supplementari legate a movimenti corporei (per esempio, nomi di utensili).
Studi con tecnica PET di nomi di utensili e di animali
Recentemente, A.R. Damasio e collaboratori (1996) hanno esaminato le differenze nell'attività cerebrale durante l'operazione di denominazione di animali e utensili (v. anche il saggio di A.R. Damasio, Emozione, processo decisionale ed etica). In uno studio PET che analizzava i cambiamenti nell'attività dei lobi temporali, questi autori hanno trovato una maggiore attività nella parte mediana del giro temporale inferiore sinistro durante l'assegnazione di nomi di animali, mentre hanno trovato un aumento di attività in aree corticali più arretrate nel giro temporale inferiore e medio durante l'assegnazione di nomi di utensili. Questi risultati suggeriscono che vi siano diverse popolazioni neuronali e aree corticali nei lobi temporali che contribuiscono alla elaborazione delle parole visive, in confronto a parole con associazioni motorie supplementari.
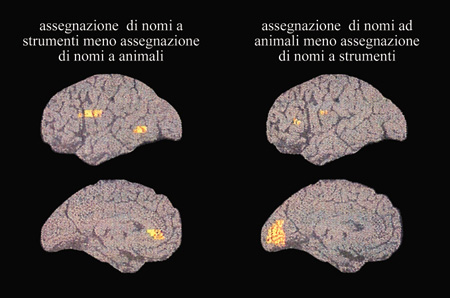
Le differenze nell'attivazione cerebrale durante l'assegnazione di nomi di animali e di utensili sono state studiate anche in una indagine PET svolta da A. Martin e collaboratori (1996). In questo caso, i soggetti dovevano assegnare in silenzio nomi di oggetti riprodotti con un disegno o con il solo profilo per eliminare le differenze nei dettagli interni dei disegni. I nomi degli oggetti rappresentati erano stati bilanciati rispetto alla frequenza delle parole. Un confronto statistico diretto tra i pattern di attivazione suscitati dall' assegnazione di nomi di animali e quelli di utensili ha messo in luce il seguente quadro (fig. 6): l'assegnazione di nomi di animali ha portato a un flusso sanguigno relativamente incrementato nelle aree visive primarie e superiori nella scissura calcarina nell' emisfero sinistro e a piccoli nuclei di attività nel lobo prefrontale; viceversa, l'assegnazione di nomi di utensili si è accompagnata a un aumento di attività nelle aree premotorie sinistre, connesse alla raffigurazione dei movimenti della mano, e a un incremento di attività nel giro temporale medio. Questi dati forniscono una prova ulteriore del fatto che aree al di fuori della corteccia perisilviana contribuiscono all'assegnazione di nomi agli animali e agli utensili. Coerentemente con gli studi precedenti, che usavano un compito di generazione di verbi, l'assegnazione di nomi di utensili ha attivato un sito corticale nel giro temporale medio sinistro. Contrariamente ai risultati dello studio di Damasio, l'incremento di attività durante l'assegnazione di nomi di animali ha riguardato le aree visive occipitali invece dei siti temporali inferiori. È della massima importanza, comunque, il fatto che l'assegnazione dei nomi agli utensili abbia portato a un nucleo supplementare di attività vicino all'area responsabile dei movimenti della mano. Questo fatto è coerente con l'ipotesi che l'elaborazione di parole con associazioni motorie attivi aree motorie coinvolte nella programmazione di tali movimenti.
Sebbene lo studio di Martin abbia diversi vantaggi metodologici rispetto ad altre indagini PET - per esempio, il bilanciamento degli stimoli e delle risposte, il calcolo delle differenze significative tra le condizioni critiche invece del solo calcolo per le differenze tra la condizione critica e quella di controllo - bisogna tener presente che si tratta di uno studio di assegnazione di nomi e che le differenze tra le condizioni in cui l'assegnazione avviene possono essere connesse a diversi processi cognitivi. Ulteriori ricerche sono necessarie per stabilire con certezza se queste differenze siano legate all'attivazione di rappresentazioni semantiche degli oggetti, o all'attivazione delle parole o a quella delle forme di parola, oppure, infine, di insiemi di cellule che includono rappresentazioni sia semantiche sia di forma di parola. Sulla base di questi obiettivi metodologici, si deve dire che nella maggior parte degli studi PET non è chiaro se e in quale misura la complessità, la frequenza, i valori di eccitazione e valenza e la ripetizione degli stimoli o delle risposte abbiano influenzato i risultati. Inoltre, quando si confronta l'assegnazione di nomi alla raffigurazione di animali e di utensili, si deve considerare che le immagini degli animali sono di solito piuttosto complesse e possono comprendere diversi colori o ombreggiature, mentre gli utensili possono essere disegnati con poche linee e mancano di solito di evidente colorazione o ombreggiatura. Se gli stimoli visivi non sono stati bilanciati rispetto alla complessità visiva, le differenze fisiche tra gli stimoli possono spiegare le diverse attivazioni delle vie visive specializzate per l'elaborazione di particolari aspetti degli stimoli stessi.
Alcune possibili influenze di fattori di confusione si possono illustrare sulla base dello studio di H. Damasio e collaboratori (1996) che confronta l'assegnazione di nomi alle rappresentazioni di utensili, animali e facce. La maggiore attivazione nei poli temporali di entrambi gli emisferi è stata osservata durante l'assegnazione dei nomi di persone famose. Non è chiaro a quale variabile psicologica si correli questo incremento di attività. È comunque evidente da indagini psicofisiologiche che le facce sono tra gli stimoli più eccitanti (Lang et al., 1990) e che le parole che si riferiscono a tali stimoli possono avere valori di eccitazione relativamente alti. È stata avanzata l'ipotesi che le parole altamente eccitanti (cioè le parole che evocano forti associazioni emotive) siano rappresentate da insiemi di cellule che comprendono neuroni supplementari nell'amigdala e in strutture sottocorticali (Schumann, 1990; Pulvermiiller e Schumann, 1994). Tutto ciò fornisce una possibile spiegazione all'attività incrementata nei poli temporali trovata da Damasio e collaboratori durante l'assegnazione di nomi alle facce. Quando venivano richiamati i nomi delle persone, si attivavano insiemi di cellule comprendenti un gran numero di neuroni dell'amigdala e, per questo motivo, si è trovato un incremento di flusso sanguigno nell' amigdala e nelle aree corticali adiacenti, cioè nei poli temporali. In questo modo, diversi valori di eccitazione delle parole possono spiegare un diverso coinvolgimento dell'amigdala e dei poli temporali durante l'assegnazione di nomi alle immagini di persone famose. Si noti che è meno probabile che un incremento di attività nei poli temporali e nell'amigdala sia connesso alle proprietà dello stimolo, poiché le facce erano presentate anche nella condizione di riferimento.
Diversità tra parole motorie e visive
Gli studi discussi conducono ai seguenti risultati, rilevanti ai fini dell'idea che le parole motorie e visive abbiano una rappresentazione e una elaborazione corticale diversa. In particolare:
1) gli studi PET sulla generazione di verbi hanno mostrato un incremento di attività nelle aree perisilviane dellinguaggio e in aree prefrontali e temporali adiacenti nell' emisfero sinistro. L'incremento di attività perisilviana si può spiegare ipotizzando che nel compito di generazione venga attivata una rappresentazione supplementare della forma di parola (relativamente al riferimento, di solito la lettura di un sostantivo). L'attivazione di aree corticali supplementari al di fuori della regione perisilviana può essere indizio di processi psicologici accoppiati alla elaborazione della forma di parola: mentre l'incremento di attività prefrontale dorsale rispetto all'area di Broca può essere connesso ai movimenti corporei cui le parole si riferiscono, l'incremento di attività nel giro temporale medio può essere connesso alla raffigurazione visiva del movimento.
2) Gli studi EEG indicano che i sostantivi e i verbi attivano diversi generatori corticali in entrambi gli emisferi. l segni più marcati di attività elettrocorticale successiva ai verbi sono stati registrati nelle regioni anteriori e centrali, mentre i sostantivi portano a segni di un'attività più pronunciata nelle aree visive occipitali. Queste differenze sembrano essere connesse all'attività neuronale all'interno o nelle vicinanze delle aree motorie o visive primarie che sottendono, rispettivamente, le associazioni motorie e visive.
3) Gli studi PET sull'assegnazione di nomi ad animali e utensili forniscono una prova ulteriore delle differenze di elaborazione tra le parole motorie e quelle visive. L'assegnazione di nomi agli utensili con sostantivi che provocano associazioni motorie attiva anche le aree premotorie e siti supplementari nel giro temporale medio; inoltre l'assegnazione di nomi agli animali tramite l'uso di sostantivi visivi può portare a un incremento di attività nelle aree temporali inferiori e in quelle occipitali vicine all'area visiva pnmana.
Sebbene questi studi siano soggetti in diversa misura a problemi metodologici, sulla loro base si può tracciare un quadro coerente. Sia gli studi EEG sia quelli PET indicano un contributo delle aree occipitali vicine alla corteccia visiva primaria all'elaborazione delle parole visive. Tale elaborazione appare così connessa all'attività neuronale nella corteccia visiva.
D'altra parte, le parole che stimolano associazioni motorie, come certi verbi di azione e i nomi di utensili, tendono ad attivare aree nel lobo frontale, anteriormente alla corteccia motoria, come si è visto dagli studi sia EEG sia PET. Inoltre, gli studi PET hanno messo in luce un'area supplementare di attivazione nel giro temporale medio che può essere connessa con la raffigurazione visiva dei movimenti indotta dalle parole motorie; è possibile che questo sito sia localizzato in modo tale che la sua attività sia difficile da registrare in EEG, o che sia necessaria una risoluzione spaziale ancor maggiore, e quindi un maggior numero di elettrodi per registrarla.
Questi risultati sono coerenti con il postulato hebbiano di diverse distribuzioni corticali di insiemi estesi di cellule connessi alla elaborazione cognitiva. Oltre ai neuroni delle aree del linguaggio perisilviane di Broca e di Wernicke, gli insiemi che rappresentano le parole motorie possono includere neuroni appartenenti alle aree motorie, premotorie e prefrontali e alle aree temporali medie. Finora non vi sono prove che l'elaborazione delle parole motorie attivi anche le regioni parietali legate all'elaborazione della auto stimolazione somatosensoriale durante i movimenti. Le parole visive potrebbero essere organizzate nella corteccia in insiemi distribuiti nelle aree perisilviane e nelle aree supplementari visive superiori o anche primarie nei lobi occipitali e temporali. Le ipotesi formulate prima ricevono dunque una conferma dai dati che abbiamo discusso. D'altra parte, le considerazioni teoriche suggeriscono che le differenze fisiologiche tra le classi di parole dovrebbero presentarsi in entrambi gli emisferi, ipotesi che, al momento, viene confermata solo da pochi studi.
Risposte coerenti ad alta frequenza generate da insiemi di cellule
Chiarire quale sia il luogo in cui, nel cervello, hanno luogo i processi cognitivi è solo uno degli argomenti interessanti da lffrontare nelle neuroscienze cognitive. Di pari importanza forse il problema della forma in cui gli elementi fondanentali delle operazioni cognitive (parole dotate di significato o figure gestaltiche) vengono rappresentati nel cervello del modo in cui queste rappresentazioni vengono attivate durante l'elaborazione dell'informazione. Lo schema hebJiano fornisce alcune proposte per rispondere a entrambi :questi problemi. Gli elementi della rappresentazione sono popolazioni fortemente connesse, ma distribuite, di migliaia di cellule nervose. Se uno di questi insiemi viene attivato, si accende e successivamente viene mantenuta l'attività neuronale nell'insieme, con un'eccitazione coerente e ripetuta di molti dei suoi neuroni.
Queste idee hanno ricevuto una parziale conferma empirica dallo studio dell'attività cerebrale coerente ad alta frequenza nei gatti, nei primati non umani e negli esseri umani. Si osserva di solito un'attività ritmica coerente e specifica rispetto allo stimolo in neuroni corticali distanti nell'intervallo di alte frequenze sopra 20 Hz. Le risposte ad alta frequenza a stimoli sensoriali possono quindi considerarsi come indicatori dell'attivazione di insiemi corticali di cellule.
Anche ammettendo che le risposte ad alta frequenza registrate nelle risposte di neurone singolo, nei potenziali locali e nella registrazione non invasiva dell'attività elettrocorticale e biomagnetica possano essere legate all'attivazione di insiemi di cellule, ci si può ancora chiedere se le risposte ad alta frequenza e l'attivazione degli insiemi siano indicatori di processi cognitivi superiori. Lo schema hebbiano suggerisce che un insieme di cellule diventa attivo ogni qual volta viene riconosciuto un elemento di elaborazione cognitiva (una Gestalt visiva o una parola significativa) e l'insieme dovrebbe rimanere attivo mentre l'elemento cognitivo viene elaborato. Le risposte ad alta frequenza dovrebbero essere quindi più intense quando vengono elaborati questi elementi rispetto a uno stato in cui non ha luogo nessuna elaborazione. Quando C. Tallon e collaboratori (1995) studiarono le risposte ad alta frequenza a figure di tipo gestaltico e a stimoli fisicamente simili che non vengono percepiti come Gestalt coerenti, trovarono un'attività più intensa intorno a 30 Hz dopo la presentazione delle Gestalt coerenti. Questo risultato è coerente con la visione secondo cui un insieme di cellule si attiva quando viene percepita una figura gestaltica, mentre la percezione di uno stimolo visivo privo di significato porta a un'attività desincronizzata ad alta frequenza nella corteccia.
Ritmi cerebrali evocati da parole e pseudoparole
Mentre si ipotizza che le parole dotate di significato siano rappresentate da insiemi corticali, è improbabile che sequenze di lettere insolite, anche se obbediscono alle regole fonetiche e ortografiche del linguaggio, posseggano una rappresentazione corticale, visto che tali pseudoparole molto probabilmente non sono mai state percepite o prodotte e quindi non si può essere strutturato nessun insieme di cellule che le rappresenti. Dunque, data la concezione di insieme hebbiano di cellule che abbiamo proposto, è fondamentale chiedersi se vi siano risposte corti cali diverse ad alta frequenza, connesse all'elaborazione delle parole significative e di pseudoparole.
In un compito di decisione lessicale, sono state presentate parole con alta frequenza e pseudoparole equilibrate con le parole rispetto alla frequenza, registrando le risposte EEG (Lutzenberger et al., 1994a; Pulvermiiller et al., 1995c). I segnali EEG sono stati prima sottoposti a un filtraggio in modo da ottenere dati indipendenti dal riferimento e da massimizzare il contributo dei generatori locali al segnale (Perrin et al., 1989). Si noti che l'uso di dati EEG grezzi renderebbe impossibile determinare se l'attività spettrale sia stata generata all'elettrodo critico o a quello di riferimento e, inoltre, l'attività spettrale all'elettrodo di riferimento entrerebbe nei dati come rumore. Le risposte spettrali evocate sono state poi calcolate per ogni sito di registrazione, in diverse bande di frequenza.
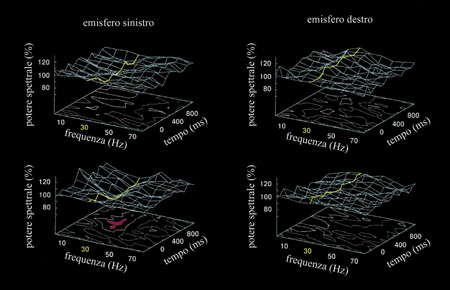
La presentazione delle parole e delle pseudoparole era immediatamente seguita da una marcata riduzione della potenza spettrale nel ritmo α (desincronizzazione α). La desincronizzazione α si osserva in EEG in diverse condizioni in cui il carico di elaborazione cognitiva viene aumentato (Pfurtscheller e Klimesch, 1991; Michel et al., 1994; Pulvermiiller et al., 1995b). Una simile riduzione generale della potenza è risultata visibile anche nel ritmo β. Non si sono osservati cambiamenti o differenze tra le classi di stimoli in questi ritmi bassi, né nell'intervallo di alta frequenza in cui è noto che l'attività muscolare produce gli effetti più marcati (Cacioppo et al., 1990). Il solo ritmo in cui sono stati riportati dati affidabili relativi a differenze specifiche rispetto allo stimolo nelle risposte spettrali è l'intervallo 25÷35 Hz (ritmo γ inferiore). Solo dopo la presentazione di pseudoparole si è avuta una marcata riduzione della potenza spettrale intorno a 30 Hz sull'emisfero sinistro, parallelamente alla riduzione di potenza in ritmo a. Dopo la presentazione di parole comuni, l'attività spettrale intorno a 30 Hz registrata dall' emisfero sinistro è stata significativamente più forte in confronto alla risposta alle pseudoparole (Lutzenberger et al., 1994). Una possibilità per spiegare questa differenza è ipotizzare un generatore supplementare ad alta frequenza che si attiva quando vengono elaborate le parole. Tale generatore potrebbe essere situato nelle aree del linguaggio dell'emisfero sinistro, poiché solo nei siti di registrazione vicini a queste aree si sono ottenute differenze nella risposta EEG ad alta frequenza tra parole e pseudoparole. Sono anche state registrate, utilizzando la magnetoencefalografia (MEG), le risposte spettrali alle parole e alle pseudoparole (Pulvermiiller et al., 1996a). In questo caso si è usato un compito di memorizzazione invece di uno di decisione lessicale e le parole inglesi e le pseudoparole venivano presentate acusticamente a persone destrimane e di madrelingua inglese, mentre negli esperimenti EEG appena discussi le parole venivano presentate visivamente a persone di madrelingua tedesca in un compito di decisione lessicale. L'uso della MEG ha due vantaggi metodologici rispetto alla registrazione EEG: in primo luogo, si possono ottenere dati indipendenti dal valore di riferimento senza usare procedure di analisi complesse. Inoltre, le variazioni di corrente ad alta frequenza sono fortemente attenuate dal cranio e dal cuoio capelluto, mentre variazioni veloci del campo magnetico non sono soggette ad attenuazione. I risultati degli esperimenti MEG sono stati simili a quelli degli esperimenti EEG. In particolare, si è potuta di nuovo osservare una differenza nell' attività spettrale nell'intervallo intorno a 30 Hz. Si sono presentate differenze supplementari in un ritmo di frequenza comprendente parte del ritmo β. Di nuovo, le risposte spettrali ad alta frequenza alle parole sono risultate più forti rispetto alle pseudoparole. Questa differenza si è mostrata significativa nelle registrazioni dall' emisfero sinistro. La figura (fig. 7) mostra l'analisi delle registrazioni da uno dei soggetti.
Vi sono molti processi cognitivi superiori diversi che possono essere indotti dalla presentazione di parole e pseudoparole. Nel caso di una parola, si possono analizzare la sua forma fonetica, le sue proprietà sintattiche e il suo significato. Se viene presentata una pseudoparola, probabilmente si dovrà stabilire innanzitutto che non vi è una rappresentazione corticale disponibile per questo stimolo. Questa è una possibilità per mettere in relazione le risposte differenziali ad alta frequenza con l'elaborazione lessicale delle parole e con il mancato funzionamento del processo di accesso lessicale dopo la presentazione di una pseudoparola. All'interno dello schema hebbiano, si può ipotizzare che la percezione di una parola attivi l'insieme di cellule che rappresenta la forma della parola insieme a tutte le altre sue caratteristiche sintattiche e semantiche, mentre la presentazione di una pseudoparola non riesce ad attivare un insieme specifico, o attiva diversi insiemi in modo asincrono.
Tutti gli insiemi di cellule che rappresentano parole possono avere neuroni localizzati nelle aree perisilviane di sinistra, mentre solo alcuni di essi possono includere neuroni supplementari, per esempio, nelle aree motorie o visive. Questo potrebbe essere il motivo per cui si sono osservate differenze riproducibili nelle risposte spettrali solo nelle aree perisilviane di sinistra del linguaggio. Non necessariamente una dinamica della risposta ad alta frequenza, che distingua tra parole e elementi privi di significato, si presenta intorno a 30 Hz. Uno studio recente ha confrontato le risposte MEG provocate da parole presentate o nella forma uditiva o in quella visiva, con le risposte MEG evocate da parole ascoltate al contrario o da sequenze di falsi caratteri (Eulitz et al., 1996). l soggetti sottoposti all'esperimento dovevano inoltre rilevare stimoli in cui una lettera, o un falso carattere, non era completa (stimolo visivo), oppure una parte del suono di una parola era rimpiazzata da toni (stimoli acustici). In questo compito, in cui in generale i soggetti erano indotti a elaborare lettere e suoni di parole, si è presentata in modo coerente un'attività ad alta frequenza diversa per le parole e per il materiale non linguistico, nell'intervallo di frequenza 60÷65 Hz. Di nuovo, le parole hanno prodotto risposte corticali ad alta frequenza più marcate nell' emisfero sinistro rispetto agli stimoli privi di senso. Questo fatto indica che l'intervallo di frequenza in cui si sviluppa la dinamica della risposta spettrale può variare in funzione del compito cognitivo.
Differenti topografie delle risposte corticali ad alta frequenza indotte da sostantivi e verbi
Dall'approccio hebbiano non consegue solo il fatto che le parole hanno insiemi di cellule mentre le pseudoparole mancano di tali rappresentazioni corticali; ne segue anche il postulato secondo cui gli insiemi di cellule che rappresentano diverse classi di parole hanno diverse distribuzioni corticali. In un compito di decisione lessicale sono stati presentati sostantivi che producono forti associazioni visive e verbi che evocano forti associazioni motorie, registrando le risposte elettrocorticali con EEG. Gli stimoli sono stati equilibrati rispetto alla frequenza delle parole, la lunghezza e i valori di eccitazione e di valenza. I dati EEG sono stati sottoposti ad analisi CSD e a un'analisi di potenza spettrale. A partire da 500 ms dopo la presentazione dello stimolo, si è ottenuta un'interazione significativa tra la categoria della parola e il sito di registrazione, nell'intervallo di frequenza 25÷35 Hz (Pulvermiiller et al., 1996b). Le differenze tra le classi di parole erano più marcate nei siti centrali e posteriori. In accordo con l'ipotesi fatta, le risposte ai sostantivi a 30 Hz erano più forti nelle aree visive, mentre le risposte ai verbi a 30 Hz erano più forti in vicinanza delle aree motorie C3 e C4. Il diagramma a destra nella figura (fig. 8) mostra la topografia corticale delle differenze della potenza a 30 Hz stimolata da queste parole visive e motorie. Non si è registrata alcuna differenza significativa tra le classi di stimoli in nessuna delle bande di frequenza analizzate.
Se si confrontano le topografie delle differenze tra sostantivi e verbi nelle risposte ad alta frequenza con i potenziali evocati, emergono un evidente accordo dei dati ma anche alcune differenze. La mappa delle differenze ottenuta dalle risposte spettrali indica una distribuzione più lateralizzata. Comunque, un confronto statistico dei dati provenienti da entrambi gli emisferi non ha confermato questa impressione. Questi dati sembrano essere compatibili con l'ipotesi che generatori corticali simili sottendano le differenze tra sostantivo e verbo nei potenziali evocati e nelle risposte ad alta frequenza.
I risultati di questo esperimento forniscono un'ulteriore conferma del fatto che le parole visive e motorie hanno diverse controparti corticali nel cervello umano normale e che aree al di fuori delle classiche regioni del linguaggio contribuiscono alla loro elaborazione. I dati forniscono anche una conferma ulteriore dello schema hebbiano, in particolare dell'ipotesi che insiemi con diverse topografie corticali rimangano attivi per un certo tempo e generino risposte ad alta frequenza dopo che siano stati percepiti diversi tipi di elementi significativi.
Se si rappresenta un insieme di cellule come una rete di neuroni che genera un'attività corticale ritmica veloce dopo essere stata stimolata, i dati riportati in questo saggio forniscono una dimostrazione che gli insiemi corti cali si attivano quando vengono elaborate parole significative, ma non si accendono quando vengono presentate pseudoparole e inoltre che la presentazione di parole motorie e visive attiva insiemi corticali con topografie diverse.
Conclusioni
Gli studi di visualizzazione sono stati utilizzati allo scopo di valutare l'ipotesi che le parole siano rappresentate a livello corticale da insiemi distribuiti di cellule con topografie definite, che variano con le proprietà semantiche delle parole. Studi di visualizzazione elettro fisiologici e metabolici forniscono una dimostrazione del fatto che nell' elaborazione delle parole non giocano un ruolo solo le aree del linguaggio dell'emisfero sinistro, ma anche aree corticali supplementari al di fuori delle aree perisilviane di sinistra. Il confronto delle risposte EEG alle parole funzionali e alle parole contenuto indica che l'emisfero destro è coinvolto nell' elaborazione delle parole di contenuto concreto, mentre un'attività prevalentemente lateralizzata a sinistra all'interno o nelle vicinanze delle aree perisilviane sembra essere connessa all'elaborazione di parole funzionali molto astratte. Sia gli studi PET sia quelli EEG hanno messo in luce che, quando vengono elaborate parole con forti associazioni visive o motorie, sono attivate aree corti cali dedicate alla programmazione motoria o alla percezione visiva.
l dati che abbiamo passato in rassegna sostengono un modello hebbiano della rappresentazione delle parole. La diversa lateralizzazione dell'elaborazione corticale delle parole di contenuto concreto e di quelle funzionali astratte si può spiegare attraverso insiemi biemisferici di cellule con diversi gradi di lateralizzazione (v. figura 3). Il diverso coinvolgimento delle aree motorie e visive nell'elaborazione delle parole motorie e visive può indicare il fatto che le rappresentazioni delle parole si estendono in queste aree extraperisilviane (v. figura 4).
Dobbiamo ancora sottolineare che alcune recenti teorie neuronali a grande scala, e le loro applicazioni ai meccanismi del linguaggio (Braitenberg, 1980; Damasio, 1989; Mesulam, 1990; Braitenberg e Pulvermiiller, 1992; Mesulam, 1994), concordano sull'ipotesi che insiemi di cellule con diverse topografie siano la controparte neuronale delle parole e che le proprietà percettive degli stimoli legati a un significato determinino la distribuzione corticale di questi insiemi. Vi sono, però, proposte divergenti, anche se tutte nella tradizione hebbiana. Per esempio, nella maggior parte di queste proposte non viene ipotizzato (o per lo meno non viene postulato esplicitamente) un contributo dell' emisfero destro alla rappresentazione e all'elaborazione dellinguaggio, mentre questa è un'ipotesi necessaria secondo la prospettiva radicalmente hebbiana che abbiamo delineato. Contributi biemisferici alla elaborazione della parola non sono solo suggeriti dalle ricerche sul cervello diviso (Zaidel, 1976) e da esperimenti comportamentali in individui sani (Banich e Karol, 1992; Mohr et al., 1994), ma sono anche coerenti con gli studi PET recenti (Zatorre et al., 1992; Mazoyer et al., 1993; Fiez et al., 1995) e con le registrazioni elettro fisiologiche in profondità (Creutzfeldt et al., 1989). Inoltre, la differente lateralizzazione delle risposte elettrocorticali a parole di diverso tipo e le differenze di elaborazione tra classi di parole registrate dall'emisfero destro sembrano fornire un sostegno ulteriore a un approccio hebbiano radicale. Attualmente, però, è prematuro rigettare una qualsiasi teoria neuronale a grande scala del linguaggio, perché ovviamente è necessario uno studio più approfondito per decidere argomenti controversi relativi al contributo dell' emisfero destro all'elaborazione del linguaggio e, in particolare, a processi specifici di classi di parole.
Lo schema hebbiano ha diverse conseguenze riguardo alla localizzazione delle rappresentazioni delle parole nel cervello. Questo schema comprende d'altra parte anche alcune idee circa il modo in cui l'elaborazione è effettivamente realizzata nel cervello. In accordo con queste idee, le risposte spettrali a frequenze abbastanza alte (superiori a 20 Hz) hanno messo in luce diversità di elaborazione tra parole e pseudoparole equilibrate rispetto alle parole. Questo fatto si può spiegare ipotizzando che gli insiemi di cellule che generano un'attività ritmica ad alta frequenza si attivino quando vengono elaborate delle parole, mentre nessun insieme si attiva completamente quando vengono percepite delle pseudoparole. Si sono potute osservare differenze nelle risposte spettrali ad alta frequenza nelle aree motorie e visive quando venivano elaborate parole con associazioni motorie o visive. Ciò è, di nuovo, coerente con la visione secondo la quale le topografie degli insiemi che rappresentano le parole motorie e visive sono diverse.
Sembra in definitiva che il linguaggio non sia ristretto alle aree del linguaggio dell'emisfero sinistro. Le parole si possono organizzare in insiemi di cellule con un'ampia distribuzione corticale e le proprietà semantiche delle parole sembrano determinare le topografie corticali di queste reti. L'attività persistente degli insiemi può essere legata alla dinamica diffusa delle risposte ad alta frequenza che possono essere registrate con EEG e MEG. Sebbene questi studi dell'elaborazione del linguaggio nel cervello possano contenere indizi circa l'organizzazione corticale delle parole, essi sono certamente solo un primo passo verso una neuroscienza cognitiva del linguaggio.
Ringraziamenti
Questo saggio si basa su un lavoro sperimentale e teorico svolto in collaborazione con i colleghi Niels Birmauer, Valentino Braitenberg, Wemer Lutzenberger, Bettina Mohr e Hubert Preissl. Vorrei ringraziarli tutti per la collaborazione. Questo lavoro è stato possibile grazie a una borsa Heisenberg e a un finanziamento della Deutsche F orschungs gemeinschaft.
Bibliografia citata
ABELES, M. (1991) Corticonics: neural circuits of the cerebral cortex. Cambridge-New York, Cambridge University Press.
AHISSAR, E., V AADIA, E., AHISSAR, M., BERGMAN, H., ARIELI, A., ABELES, M. (1992) Dependence of cortical plasticity on correlated activity of single neurons and on behavioral context. Science, 257, 1412-1415.
ALLISON, T., McCARTHY, G., NOBRE, A.C., PUCE, A., BELGER, A. (1994) Human extrastriate visual cortex and the perception of faces, words, numbers and colors. Cereb. Cortex, 4, 544-554.
ANNETT, M. (1979) Family handedness in three generations predicted by the right shift theory. Ann. Rum. Genet., 42, 479-491.
BANICH, M.T., KAROL, D.L. (1992) The sum of the parts does not equal the whole: evidence from bihemispheric processing. J. Exp. Psychol. Rum. Percept. Perform., 18, 763-784.
BOGEN, J.E., BOGEN, G.M. (1976) Wemicke's region. Where is it? Ann. N. Y. Acad. Sci., 280, 834-843.
BRAITENBERG, V. (1978) Cell assemblies in the cerebral cortex. In Theoretical approaches to complex systems, a c. di Heim R., Palm G., Berlino, Springer, pp. 171-188.
BRAITENBERG, V. (1980) Alcune considerazioni sui meccanismi cerebrali del linguaggio. In L'accostamento interdisdplinare allo studio del linguaggio, a c. di Braga G., Braitenberg V., Cipolli C., Coseriu E., Crespi-Reghizzi S., Mehler J., Titone R., Milano, Franco Angeli, pp. 96-108.
BRAITENBERG, V., PULVERMÜLLER, F. (1992) Model of a neurological theory of speech. Naturwissenschaften, 79, 103-117.
BRAITENBERG, V., SCHUZ, A. (1991) Anatomy of the cortex. Statistics and geometry. Berlino, New York, Springer Verlag.
BRAITENBERG, V., SCHUZ, A. (1992) Basic features of cortical connectivity and some considerations on language. In Language origin: a multidisciplinary approach, a c. di Wind J., Chiarelli B., Bichakjian B.H., Nocentini A, Jonker A, Dordrecht, Boston, Kluwer, pp. 89-102.
BRODMANN, K (1909) Vergleichende Lokalisationslehre der Grofthirnrinde, Lipsia, Barth.
CACIOPPO, J.T., TASSINARY, L.G., FRIDLUND, A.J. (1990) The skeletomotor system. In Principles of psychophysiology. Physical, sodal, and inferential elements, a c. di Cacioppo 1T., Tassinary L.G., Cambrigde-New York, Cambridge University Press, pp. 325-384.
CARAMAZZA, A., BERNDT, R. (1985) A multi-component deficit view of agrammatic Broca's aphasia. In Agrammatism, a c. di Kean M.-L., Orlando, Academic Press, pp. 27-64.
CREUTZFELDT, O., OJEMANN, G., LETTICH, E. (1989) Neuronal activity in the human lateral temporal lobe. I. Responses to speech. Exp. Brain Res., 77, 451-475.
DAMASIO, A.R. (1989) Time-Iocked multiregional retroactivation: a systems-Ievel proposal for the neural substrates of recall and recognition. Cognition, 33, 25-62.
DAMASIO, H., GRABOWSKI, T.J., TRANEL, D., HICHWA, R.D., DAMASIO, A.R. (1996) A neural basis for lexical retrieval. Nature, 380, 499-505.
DEACON, T.W. (1992) Cortical connections of the inferior arcuate sulcus cortex in the macaque brain. Brain Res., 573, 8-26.
DEHAENE, S. (1995) Electrophysiological evidence for categoryspecific word processing in the normal human brain. Neuroreport, 6, 2153-2157.
DEHAENE-LAMBERTZ, G., DEHAENE, S. (1994) Speed and cerebral correlates of syllable discrimination in infants. Nature, 370, 292-295.
DIAMOND, M.C. (1990) Morphological cortical changes as a consequence of leaming and experience. In Neurobiology of higher cognitive Junction, a c. di Scheibel AB., Wechsler AF., New York, Guilford Press, pp. 1-12. ELBERT, T., RAY, W.J., KOWALIK, Z.J., SKINNER, J.E., GRAF, K.E., BIRBAUMER, N. (1994) Chaos and physiology: deterministic chaos in excitable cell assemblies. Physiol. Rev., 74, 1-47.
ENGEL, A.K, KONIG, P., KREITER, A.K, SINGER, W. (1991a) Interhemispheric synchronization of oscillatory neuronal responses in cat visual cortex. Science, 252, 1177-1179.
ENGEL, A.K, KREITER, A.K, KONIG, P., SINGER, W. (1991b) Synchronization of oscillatory neuronal responses between striate and extrastriate visual cortical areas of the cat. Proc. Natl. Acad. Sci. USA, 88, 6048-6052.
EULITZ, C., MAESS, B., PANTEV, C., FRIEDERICI, A.D., FEIGE, B., EIBERT, T. (1996) Oscillatory neuromagnetic activity induced by language and non-Ianguage stimuli. Cogn. Brain Res., 4, 121-132.
FIEZ, J.A., RAICHLE, M.E., BALOTA, D.A, TALLAL, P., PETERSEN, S.E. (1996) PET activation of posterior temporal regions during auditory word presentation and verb generation. Cereb. Cortex, 6, 1-10.
FIEZ, J.A., RAICHLE, M.E., MIEZIN, F.M., PETERSEN, S.E., TALLAL, P., KATZ, W.F. (1995) PET studies of auditory and phonological processing effects of stimulus characteristics and task demands. J. Cognit. Neurosci., 7, 357-375.
FREGE, G. (1980) Uber Sinn und Bedeutung. In Funktion, Begriff, Bedeutung, a c. di Patzig G., Gottinga, Huber, pp. 25-50.
FRY, D.B. (1966) The development ofthe phonological system in the normal and deaf child. In The genesis of language. Proceedings of a conference on 'Ianguage development in children', a c. di Smith F., Miller G.A, Cambridge, Mass., MIT Press, pp. 187-206.
FUSTER, J.M. (1994) Memory in the cerebral cortex. An empirical approach to neural networks in the human and nonhuman primate, Cambridge, Mass., MIT Press.
GALABURDA, A.M., ROSEN, G.D., SHERMAN, G.F. (1991) Cerebrocortical asymmetry. In Cerebral cortex, a c. di Peters A, Jones E.G., voI. 9, New York, Plenum Press, pp. 263-277.
GARNSEY, S.M. (1985) Function words and content words: reaction time and evoked potential measures of word recognition, Rochester, N.Y., University of Rochester.
GESCHWIND, N. (1970) The organization of language and the brain. Science, 170, 940-944.
GEVINS, A., CUTILLO, B., SMITH, M.E. (1995) Regional modulation of high resolution evoked potentials during verbal and non-verbal matching tasks. Electroencephalogr. Clin. Neurophysiol., 94, 129-147.
GLEITMAN, L.R., WANNER, E. (1982) Language acquisition: the state of the state of the art. In Language acquisition. The state of the art, a c. di Wanner E., Gleitman L.R., Cambridge-New York, Cambridge University Press, pp. 3-48.
GUSTAFSSON, B., WIGSTROM, H., ABRAHAM, W.C., HUANG, Y.Y. (1987) Long-term potentiation in the hippocampus using depolarizing current pulses as the conditioning stimulus to single volley synaptic potentials. J. Neurosci., 7, 774-780.
HAYES, T.L., LEWIS, D. (1993) Hemispheric differences in layer III pyramidal neurons of the anterior language area. Arch. Neurol., 50, 501-505.
HEBB, D.O. (1949) The organization of behavior. A neuropsychological theory, New Y ork, Wiley.
JACKSON, J.H. (1878) On affections of speech from disease of the brain (1). Brain, 1, 304-330.
JACOBS, B., BATAL, H.A., LYNCH, B., OJEMANN, G., OJEMANN, L.M., SCHEIBEL, A.B. (1993) Quantitative dendritic and spine analyses of speech cortices: a case study. Brain Lang., 44, 239-253.
JACOBS, B., SCHEIBEL, A.B. (1993) A quantitative dendritic analysis of Wemicke's area in humans. I. Lifespan changes. J. Comp. Neurol., 327, 83-96.
KING, J.W., KUTAS, M. (1995) Proceedings of the 2nd Annual Meeting oJthe Cognitive Neuroscience Society, 68 (Abstract).
KOLK, H.H.J., GRUNSVEN, J.F. VAN, KEYSER, A. (1985) On parallelism between production and comprehension in agrammatism. In Agrammatism, a c. di Kean M.-L., New York, Academic Press, pp. 165-206.
KOUNIOS, J., HOLCOMB, P.J. (1994) Concreteness effects in semantic processing: ERP evidence supporting dual-coding theory. J. Exp. Psychol. Learn. Mem. Cogn., 20, 804-823.
LANG, P.J., BRADLEY, M.M., CUTHBERT, B.N. (1990) Emotion, attention and the startle reflex. Psychol. Rev., 97, 377-395.
LICHTHEIM, L. (1885) On aphasia. Brain, 7, 433-484.
LUTZENBERGER, W., PULVERMÜLLER, F., BIRBAUMER, N. (1994) Words and pseudowords elicit distinct patterns of 30-Hz EEG responses in humans. Neurosci. Lett., 176, 115-118.
McCARTHY, G., ALLISON, T., SPENCER, D.D. (1993) Localization of the face area in human sensorimotor cortex by intracranial recording of somatosensory evoked potential. J. Neurosurg., 79, 874-884.
MARTIN, A., HAXBY, J.V., LALONDE, F.M., WIGGS, C.L., DNGERLEIDER, L.G. (1995) Discrete cortical regions associated withknowledge of color and knowledge of action. Science, 270, 102-105.
MARTIN, A., WIGGS, C. L. , DNGERLEIDER, L.G., HAXBY, J.V. (1996) Neural correlates of category-specific knowledge. Nature, 379, 649-652.
MAZOYER, B.M., TZOURIO, N., FRAK, V., SYROTA, A., MURAYAMA, N., LEVRIER, O., SALAMON, G., DEHAENE, S., COHEN, L., MEHLER, J. (1993) The cortical representation of speech. J. Cognit. Neurosci., 5, 467-479.
MESULAM, M.M. (1990) Large-scale neurocognitive networks and distributed processing for attention, language and memory. Ann. Neurol., 28, 597-613.
MESULAM, M.M. (1994) Neurocognitive networks and selectively distributed processing. Rev. Neurol., 150, 564-569.
MICHEL, C.M., KAUFMAN, L., WILLIAMSON, S.J. (1994) Duration of EEG and MEG α suppression. J. Cognit. Neurosci., 6, 139-150.
MILLER, R. (1987) Representation of brieftemporal patterns, Hebbian synapses, and the left-hemisphere dominance for phoneme recognition. Psychobiology, 15, 241-247.
MOHR, B., PULVERMÜLLER, F., ZAIDEL, E. (1994) Lexical decision after left, right and bilateral presentation of function words, content words and non-words: evidence for interhemispheric interaction. Neuropsychologia, 32, 105-124.
MOLFESE, D.L., BETZ, J.C. (1988) Electrophysiological indices of the early development of lateralization for language and cognition, and their implications for predicting later development. In Brain lateralization in children. Developmental implications, a c. di Molfese D.L., Segalowitz S.J., New York, Guilford Press, pp. 171-190.
NEVILLE, H.J., MILLS, D.L., LAWSON, D.S. (1992) Fractionating language: different neural subsystems with different sensitive periods. Cereb. Cortex, 2, 244-258.
NOBRE, A.C., McCARTHY, G. (1994) Language-related EPRs: scalp distributions and modulation by word type and semantic priming. J. Cognit. Neurosci., 6, 233-255.
PANDYA, D.N., YETERIAN, E.H. (1985) Architecture and counections of cortical association areas. In Cerebral cortex, a c. di Peters A., Jones E.G., voI. 4, New Y ork, Plenum Press, pp. 3-61.
PERRIN, F., PERNIER, J., BERTRAND, O., ECHALLIER, J.F. (1989) Spherical splines for scalp potential and current density mapping. Electroencephalogr. Clin. Neurophysiol., 72, 184-187.
PETERSEN, S., FOX, P., POSNER, M., MINTUN, M., RAICHLE, M. (1989) Positron emission tomography studies of the processing of single words. J. Cognit. Neurosci., 1, 153-170.
PFURTSCHELLER, G., KLIMESCH, W. (1991) Event-related desynchronization during motor behavior and visual inforrnation processing. Electroencephalogr. Clin. Neurophysiol., Suppl. 42, 58-65.
PREISSL, H., PULVERMÜLLER, F., LUTZENBERGER, W., BIRBAUMER, N. (1995) Evoked potentials distingnish between nouns and verbs. Neurosci. Lett., 197, 81-83.
PREVIC, F.H. (1991) A generaI theory concerning the prenatal origins of cerebral lateralization in humans. Psychol. Rev., 98, 299-334.
PRICE,C.J., WISE, R.J., WATSON, J.D., PATTERSON, K., HOWARD, D., FRACKOWIAK, R.S. (1994) Brain activity during reading. The effects of exposure duration and task. Brain, 117, 1255-1269.
PUCE, A., ALLISON, T., GORE, J.C., McCARTHY, G. (1995) Facesensitive regions in human extrastriate cortex studied by functional MRI. J. Neurophysiol., 74, 1192-1199.
PULVERMÜLLER, F. (1992) Constituents of a neurological theory of language. Concepts Neurosci., 3, 157-200.
PULVERMÜLLER, F. (1999) Words in the brain's language. Behavioral and Brain Sciences (in corso di stampa).
PULVERMÜLLER, F., EULITZ, C., PANTEV, C., MOHR, B., FEIGE, B., LUTZENBERGER, W., ELBERT, T., BIRBAUMER, N. (1996a) Highfrequency cortical responses reflect lexical processing: an MEG study. Electroencephalogr. Clin. Neurophysiol., 98, 76-85.
PULVERMÜLLER, F., LUTZENBERGER, W., BIRBAUMER, N. (1995a) Electrocortical distinction of vocabulary types. Electroenceph. Clin. Neurophysiol., 94, 357-370.
PULVERMÜLLER, F., LUTZENBERGER, W., PREISSL, H., BIRBAUMER, N. (1995b) Motor programming in both hemispheres: an EEG study ofthe human brain. Neurosci. Lett., 190, 5-8.
PULVERMÜLLER, F., PREISSL, H. (1991) A celI assembly model of language. Network, 2, 455-468.
PULVERMÜLLER, F., PREISSL, H., LUTZENBERGER, W., BIRBAUMER, N. (1995c) Spectral responses in the gamma-bando Neuroreport, 6, 2057-2064.
PULVERMÜLLER, F., PREISSL, H., LUTZENBERGER, W., BIRBAUMER, N. (1996b) Brain rhythms of language: nouns versus verbs. Eur. J. Neurosci., 8, 937-941.
PULVERMÜLLER, F., SCHUMANN, J.H. (1994) Neurobiologica1 mechanisms of language acquisition. Language Learning, 44, 681-734.
SCHEIBEL, A.B., PAUL, L.A., FRIED, I., FORSYTHE, A.B., TOMIYASU, D., WECHSLER, A., KAO, A., SLOTNICK, J. (1985) Dendritic organization of the anterior speech area. Exp. Neurol., 87, 109-117.
SCHUMANN, J. (1990) The role of the amygdala as a mediator of affect and cognition in second language acquisition. In Georgetown University round table on language and linguistics 1990, a c. di Alatis J., Washington, D.C., Georgetown University Press, pp. 169-176.
SINGER, W., GRAY, C.M. (1995) Visual feature integration and the temporal correlation hypothesis. Annu. Rev. Neurosci., 18, 555-586.
STEINMETZ, H., RADEMACHER, J., JÄNCKE, L., HUANG, Y., THRON, A., ZILLES, K. (1990) Total surface of tempoparietal intrasylvian cortex: diverging left-right asymmetries. Brain Lang., 39, 357-372.
TALLON, C., BERTRAND, O., BOUCHET, P., PERNIER, J. (1995) Gamma-range activity evoked by coherent visual stimuli in humans. Eur. J. Neurosci., 7, 1285-1291.
WARRINGTON, E.K., McCARTHY, R.A. (1987) Categories of knowledge. Further fractionations and an attempted integration. Brain, 110, 1273-1296.
WERNICKE, C. (1874) Der aphasische Symptomencomplex. Eine psychologische Studie auf anatomischer Basis. Breslavia, Cohn.
WICKENS, J., HYLAND, B., ANSON, G. (1994) Cortical celI assemblies: a possible mechanism for motor programs. J. Motor Behav., 26, 66-82.
WISE, R., CHOLLET, F., HADAR, D., FISTON, K., HOFFNER, E., FRACKOWIAK, R. (1991) Distribution of cortical neural networks involved in word comprehension and word retrieval. Brain, 114, 1803-1817.
WITTGENSTEIN, L. (1967) Philosophische Untersuchungen. Francoforte, Suhrkamp.
WOODS, B.T. (1983) Is the left hemisphere specialized for language at birth? Trends Neurosci., 6, 115-117.
ZAIDEL, E. (1976) Auditory vocabulary ofthe right hemisphere following brain bisectionor hemidecortication. Cortex, 12, 191-211.
ZATORRE, R.J., EVANS, A.C., MEYER, E., GJEDDE, A. (1992) Lateralization of phonetic and pitch discrimination in speech processing. Science, 256, 846-849.
ZEKI, S., W ATSON, J.D., LUECK, C.J., FRISTON, K.J., KENNARD, c., FRACKOWIAK, R.S. (1991) A direct demonstration offunctional specialization in human visual cortex. J. Neurosci. 11, 641-649.