
Imprinting
Imprinting
Gli etologi definiscono 'imprinting' lo sviluppo, in un animale giovane, di una preferenza sociale per il suo genitore o per un altro individuo al quale esso sia stato esposto sin dal momento della nascita. Grazie alle brillanti ricerche condotte da Konrad Lorenz (1937), l'imprinting è diventato un argomento centrale nello studio del comportamento animale ed ha generato una ricca messe di studi empirici sui meccanismi fondamentali dell'apprendimento. Gli etologi hanno classificato diverse forme di imprinting, a seconda delle conseguenze funzionali a breve o a lungo termine sul comportamento del soggetto, oppure a seconda dei diversi repertori comportamentali che ne vengono modificati. L'imprinting filiale per esempio, ossia la formazione di un attaccamento e di una preferenza da parte di un animale giovane per un genitore, per un suo surrogato o per i fratelli, ha lo scopo di assicurare al piccolo la vicinanza e la protezione dell'adulto che ne dovrà assicurare la sopravvivenza; questa prima forma di imprinting si distingue dall'imprinting sessuale, che riguarda invece la formazione delle preferenze nella scelta del partner, che verranno espresse nel corso della vita dell'individuo e che orienteranno le scelte riproduttive. In entrambi i casi, gran parte delle ricerche è stata effettuata sugli uccelli.
Mediante l'associazione di osservazioni comportamentali e di metodi di indagine neurofisiologica, è stato possibile individuare alcuni dei processi di elaborazione dell'informazione che presiedono a questo comportamento. Dei due meccanismi percettivi che svolgono un ruolo importante nello sviluppo del comportamento filiale, il primo è essenzialmente una rappresentazione o memoria degli stimoli a cui l'animale viene esposto; la formazione di questa memoria è appunto nota come 'imprinting filiale'. Il processo di apprendimento che porta all'imprinting filiale differisce da quello caratteristico del condizionamento pavloviano, ma alcuni meccanismi di base sono simili a quelli tipici dell'apprendimento associativo. Il secondo meccanismo responsabile dello sviluppo del comportamento filiale era stato già individuato da Lorenz, quando egli sottolineò che l'imprinting non è un fenomeno universalmente valido per tutte le specie di Uccelli. Nella terminologia di Lorenz alcune specie di Uccelli possiedono un 'modello innato di percezione' del 'compagno' (Kumpan) appropriato, mentre in altre specie il modello percettivo si forma in seguito all'esperienza con un particolare oggetto. Solo quest'ultimo caso equivale all'imprinting, mentre l'altro è approssimativamente corrispondente al meccanismo percettivo detto 'predisposizione'. In questo contesto le predisposizioni si riferiscono a preferenze percettive che si sviluppano in giovani animali senza che essi abbiano avuto esperienza dei particolari stimoli implicati. Studi recenti hanno dimostrato che, più in generale, le predisposizioni svolgono un ruolo importante nello sviluppo del comportamento filiale in alcune specie di Uccelli a prole precoce; sarà qui discussa la natura di queste predisposizioni nei pulcini di gallo domestico (Gallus gallus spadiceus).
Imprinting e apprendimento
La formazione delle preferenze filiali è stata studiata soprattutto in specie di Uccelli a prole precoce, come anatre e galli domestici. Questi uccelli sono in grado di muoversi autonomamente poco dopo la schiusa e tendono ad avvicinarsi e a seguire qualsiasi oggetto appariscente a cui siano stati esposti. In natura l''oggetto' è in genere la madre o un consanguineo, ma in loro assenza un'ampia varietà di surrogati è in grado di provocare nel giovane il comportamento filiale: per esempio, quando un pulcino si trova presso un oggetto appropriato, si avvicina e tenta di accovacciarsi accanto a esso e, quando l'oggetto viene rimosso, diventa irrequieto ed emette insistenti pigolii di richiamo. Inizialmente il piccolo si avvicina a molti degli oggetti a cui è esposto ma, dopo aver fatto una certa esperienza con un particolare oggetto, vi rimane fedele ed evita nuovi oggetti. Se gli si concede di scegliere tra lo stimolo familiare e uno stimolo nuovo, l'uccello si avvicina di preferenza a quello familiare. Si definisce allora 'imprinting filiale' il processo attraverso il quale il comportamento sociale del giovane animale diviene limitato a un particolare oggetto o classe di oggetti a cui è esposto.
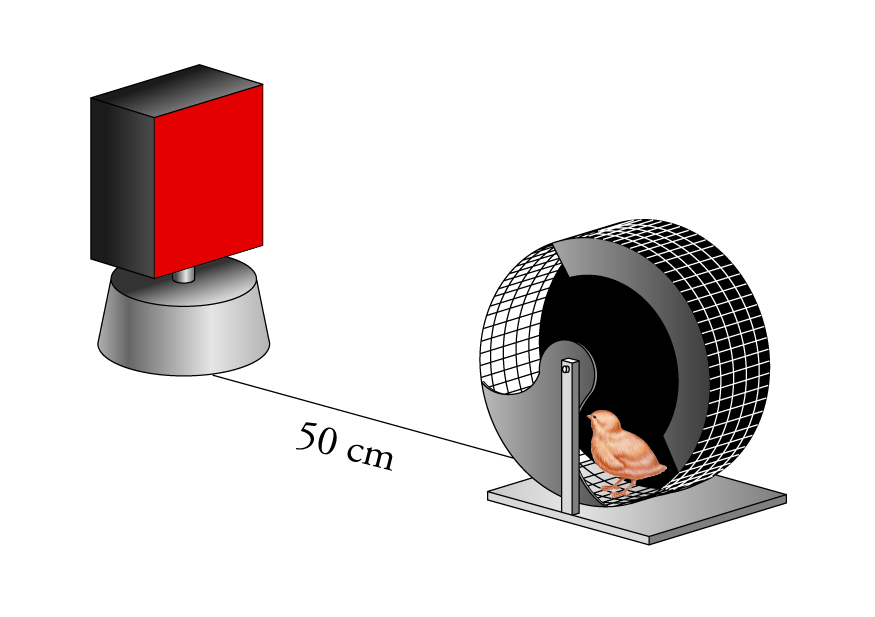
Per studiare l'imprinting in laboratorio, spesso i pulcini o gli anatroccoli vengono fatti nascere e allevati in oscurità ed esposti a un oggetto ben visibile e definito a circa 24 ore dalla nascita. Il pulcino viene esposto a uno stimolo mentre si trova in una ruota mobile costituita da una rete metallica (come quella illustrata nella fig. 2), e può vedere l'oggetto attraverso le maglie della rete. Dopo la sessione di imprinting, gli animali vengono riportati in incubatrice e tenuti in oscurità fino a quando vengono misurate le loro preferenze in un test di scelta, che comprende l'esposizione sequenziale o simultanea allo stimolo presentato nella sessione di imprinting e a uno stimolo nuovo. In genere, quanto più un oggetto è appariscente per l'occhio umano, tanto più sarà efficace come stimolo per l'imprinting. Il movimento dell'oggetto aumenta inoltre la sua attrattiva e lo stesso avviene se, oltre allo stimolo visivo, viene presentato anche uno stimolo uditivo. L'imprinting non è tuttavia un processo istantaneo: per un pulcino può essere necessaria più di un'ora di esposizione affinché manifesti una preferenza significativa per un oggetto artificiale semplice e il periodo di esposizione necessario può variare a seconda dell'oggetto e della procedura sperimentale adottata.
Konrad Lorenz inizialmente suggerì che il processo di acquisizione della conoscenza dell'oggetto verso cui il comportamento sociale veniva diretto non avesse nulla a che vedere con l'apprendimento, soprattutto perché pensava che tale processo avvenisse durante un periodo sensibile e fosse irreversibile. Ricerche successive hanno invece dimostrato che le preferenze sociali indotte dall'imprinting possono essere modificate e che non esiste un periodo sensibile in senso assoluto. Inoltre, questi criteri teorici non fanno differenza tra imprinting e altre forme di apprendimento. Lorenz operava una distinzione tra l'imprinting, che vedeva come l'acquisizione di un 'modello percettivo' della specie, e un processo di apprendimento simile al condizionamento classico. È chiaro attualmente che l'imprinting filiale non implica semplici associazioni stimolo-risposta, ma crea una rappresentazione mentale dello stimolo noto che può essere utilizzata in maniera flessibile. Per esempio, in seguito a imprinting filiale, i pulcini mostrano ritmi di apprendimento discriminativo significativamente più veloci se uno dei due stimoli da distinguere è quello che ha indotto l'imprinting. Ciò avviene nonostante il fatto che nella prova di discriminazione l'avvicinamento allo stimolo nuovo venga premiato con una ricompensa, mentre l'avvicinamento all'oggetto usato per l'imprinting non viene rinforzato.
Gli etologi oggi propendono per un'interpretazione dell'imprinting in termini di apprendimento associativo e hanno descritto gli elementi essenziali per questo tipo di interpretazione. Fino a poco tempo fa erano disponibili dati relativamente scarsi su questo argomento. Alcuni dei meccanismi alla base dei processi di apprendimento dell'imprinting sono stati chiariti quando si è tenuto conto che, in natura, la chioccia possiede elementi che la caratterizzano, quali taglia, forma e colorazione, ed emette un richiamo specifico. Dunque, durante le sessioni di imprinting in laboratorio, alcuni giovani uccelli sono stati esposti a uno stimolo visivo accompagnato da una registrazione del richiamo di una chioccia. In alcuni studi, un gruppo di pulcini veniva esposto a uno stimolo visivo accompagnato dal richiamo materno, mentre un altro gruppo veniva esposto al solo stimolo visivo. In seguito, entrambi i gruppi sono stati sottoposti a un test di preferenza in cui lo stimolo visivo della sessione di imprinting e un nuovo stimolo visivo venivano presentati senza il richiamo materno. I pulcini a cui era stato fatto ascoltare il richiamo materno durante la prova di imprinting mostravano una preferenza significativamente maggiore per lo stimolo di imprinting rispetto a quelli che non avevano ascoltato il richiamo. Così, la presenza di un richiamo sonoro durante la prova di imprinting è in grado di aumentare la preferenza acquisita per lo stimolo visivo.
Un'interpretazione interessante di queste osservazioni è che un meccanismo analogo potrebbe spiegare la facilitazione dell'imprinting e il potenziamento della capacità di apprendimento in soggetti condizionati mediante stimoli complessi, composti cioè dall'associazione di due stimoli diversi (come una luce e un suono). Per esempio, nei ratti il condizionamento avversivo nei confronti di un odore viene incrementato se durante la sessione di condizionamento l'odore è accoppiato a un sapore. È stato suggerito che tale potenziamento potrebbe essere il risultato di un apprendimento durante l'evento: in altre parole, durante la fase di condizionamento, potrebbero formarsi associazioni tra le rappresentazioni mentali dei due elementi che costituiscono lo stimolo composito. Analogamente, durante la sessione di imprinting effettuata con uno stimolo composito uditivo e visivo, potrebbe determinarsi un'associazione tra le rappresentazioni delle componenti uditiva e visiva. La formazione di tale associazione farebbe sì che, durante un successivo test in cui venisse presentata solo la componente visiva, vi sarebbero due potenziali sorgenti della risposta preferenziale: quella acquisita tramite la componente visiva e quella dovuta alla rappresentazione, attivata in maniera associativa, della componente uditiva dello stimolo composito.
Sembrerebbe per alcuni versi che l'imprinting filiale e il condizionamento classico (o pavloviano) procedano in modo differente. Nel condizionamento uno stimolo neutro viene accoppiato a uno stimolo biologicamente rilevante. Lo stimolo neutro è detto 'stimolo condizionato'; nel famoso esperimento sui cani di Ivan Pavlov, lo stimolo condizionato era costituito dal suono di una campana. Lo stimolo biologicamente rilevante è noto come 'stimolo incondizionato'; nel caso dei cani di Pavlov esso corrispondeva alla distribuzione del pasto, che induceva una forte salivazione negli animali. Dopo diversi abbinamenti del suono della campana alla distribuzione del pasto, il suono di per sé stesso era divenuto sufficiente a indurre la salivazione, indicando che gli animali associavano allo stimolo condizionato quello incondizionato.
Nel caso di imprinting con uno stimolo composito, uditivo e visivo, un'interpretazione alternativa a quella dell'apprendimento durante l'evento è che lo stimolo uditivo agisca come una sorta di stimolo incondizionato rispetto allo stimolo visivo inizialmente neutro (lo stimolo condizionato). Dunque l'imprinting corrisponderebbe a una forma di condizionamento pavloviano. È stato dimostrato che gli stimoli capaci di indurre imprinting hanno un certo significato motivazionale per il giovane animale: essi, infatti, possono fungere da stimolo incondizionato in prove di apprendimento strumentale. Tuttavia, nel condizionamento pavloviano la presentazione sequenziale dello stimolo condizionato e dello stimolo incondizionato è di gran lunga più efficace della presentazione simultanea dei due stimoli. Come abbiamo visto, invece, nell'imprinting con uno stimolo composito uditivo e visivo avviene proprio il contrario: la presentazione simultanea dello stimolo composito è più efficace della presentazione sequenziale.
I meccanismi della predisposizione filiale e dell'imprinting
Le osservazioni comportamentali che suggeriscono l'esistenza del meccanismo della predisposizione sono state compiute da Mark H. Johnson e collaboratori (1985). Questi ricercatori hanno sottoposto pulcini di un giorno, allevati in oscurità, a sessioni di imprinting in cui i piccoli venivano esposti a una scatola rossa rotante oppure a un individuo impagliato rotante di sesso femminile della specie Gallus gallus spadiceus. Altri individui, usati come controllo, venivano esposti invece a una luce bianca. Le preferenze dei pulcini venivano misurate in due test successivi, condotti rispettivamente 2 ore (test 1) e 24 ore (test 2) dopo la sessione di imprinting, in cui la gallina e la scatola venivano presentate simultaneamente. Mentre nel test 1 i pulcini mostrarono di preferire l'oggetto a cui erano stati esposti precedentemente, nel test 2 venne osservata una preferenza significativamente maggiore per la gallina impagliata in entrambi i gruppi sperimentali. La preferenza per la gallina impagliata era dunque maggiore nel test condotto dopo 24 ore e questo indipendentemente da quale fosse lo stimolo a cui i pulcini erano stati esposti durante la sessione di imprinting. Anche i pulcini del gruppo di controllo, che erano stati posti nell'apparato sperimentale ma esposti al solo stimolo di una luce bianca, mostrarono un aumento della preferenza per la gallina impagliata se sottoposti al test dopo 24 ore.
Johnson e collaboratori hanno suggerito che le preferenze dei pulcini sottoposti a sessioni di imprinting dipendano da almeno due differenti sistemi. Da una parte, vi è l'effetto dell'esperienza con particolari stimoli, che determina la differenza di comportamento tra i pulcini appartenenti al gruppo di controllo e quelli sottoposti a stimoli specifici: questo è l'imprinting filiale. Dall'altra, si osserva una predisposizione ad accostarsi a stimoli che assomigliano a individui appartenenti alla stessa specie, che si riflette nel diverso comportamento del gruppo di controllo nel test 1 e nel test 2. I risultati di questo esperimento suggeriscono che per far emergere tale predisposizione non è necessario esporre i piccoli a un particolare stimolo; essa, infatti, può manifestarsi in pulcini allevati in oscurità a condizione che ricevano una certa quantità di stimolazione non specifica entro un determinato periodo dello sviluppo. In altri studi è stato osservato che l'esperienza visiva non è necessaria per 'scatenare' o indurre la predisposizione. In un esperimento si è visto che pulcini collocati in una ruota mobile (fig. 2) in totale oscurità, per due periodi di 1 ora, preferiscono la gallina impagliata alla scatola rossa in un test di scelta simultanea effettuato 24 ore dopo il periodo trascorso nella ruota. Al contrario, pulcini tenuti nell'incubatrice oscura dal momento della schiusa a quello del test non sviluppano alcuna preferenza. È dunque sufficiente una minima esperienza associata al trovarsi nella ruota per far emergere la preferenza per la gallina impagliata.
In tali studi sulla predisposizione, il periodo in cui i pulcini si trovavano nella ruota in oscurità iniziava approssimativamente 24 ore dopo la schiusa. Una preferenza significativa per la gallina impagliata fu riscontrata 24 ore, ma non 2 ore, dopo la fine del periodo trascorso nelle ruote mobili. Neil G. Hampton e collaboratori (1995) si sono chiesti se siano effettivamente necessarie 24 ore affinché il meccanismo alla base di questo fenomeno si sviluppi o sia in grado di esercitare il suo influsso sul comportamento dei pulcini. Si è scoperto che la predisposizione si manifesta 10 ore dopo il collocamento nelle ruote mobili, oppure 5 ore dopo che i pulcini sono stati esposti a una sessione con la scatola rossa rotante. Inoltre questi ricercatori hanno dimostrato che non è necessario collocare i pulcini nella ruota per far emergere la predisposizione. Pulcini che venivano tolti dall'incubatrice e manipolati nello stesso tempo e per la stessa durata per cui gli altri pulcini sarebbero normalmente stati collocati nella ruota, mostravano una preferenza significativa per la gallina impagliata rispetto alla scatola rossa 5 ore, ma non 2 ore, dopo la fine di questa esperienza. In maniera analoga Hampton e collaboratori trovarono che l'esposizione di pulcini di 24 ore allevati in oscurità a un richiamo materno di una gallina per 2 ore era sufficiente a indurre lo sviluppo di una predisposizione visiva 24 ore più tardi. Le prove fornite da questi studi suggeriscono che differenti tipi di esperienza, in differenti modalità, sono sufficienti per indurre la predisposizione.
Si è inoltre cercato di capire se la predisposizione funga da filtro o da stampo, limitando le informazioni che il pulcino può memorizzare, o se, viceversa, essa agisca indipendentemente dagli effetti dell'apprendimento durante lo sviluppo. In alcuni pulcini allevati in oscurità è stata indotta la predisposizione, manifestatasi come una preferenza significativa per la gallina impagliata rispetto alla scatola rossa, collocandoli per 2 ore in oscurità all'interno di ruote mobili. In successivi test di preferenza si è osservato che, dopo lo sviluppo della predisposizione, questi pulcini erano ancora in grado di apprendere le caratteristiche di un oggetto appariscente come una scatola rossa. Questi risultati suggeriscono che la predisposizione non agirebbe limitando la quantità di informazione memorizzata, bensì che i due meccanismi interagirebbero a livello comportamentale. In altre parole, l'attivazione della predisposizione orienterà la risposta di approccio del pulcino verso un certo tipo di stimoli e l'animale potrà apprendere le caratteristiche di questi stimoli, ma sarà ancora in grado di apprendere le caratteristiche di altri oggetti.
Ma quali sono le caratteristiche determinanti dello stimolo per l'espressione della predisposizione filiale? Mark H. Johnson e Gabriel Horn (1988) hanno cercato di 'titolare' la complessità dello stimolo visivo proveniente dalla gallina selvatica impagliata rispetto alle sue caratteristiche di 'modello di gallina'. Gli stimoli utilizzati durante i test erano una gallina impagliata intatta e una serie di modelli di gallina sempre più degradati. Questi ultimi spaziavano da un modello in cui le differenti parti (ali, capo, tronco, zampe) venivano riassemblate in maniera innaturale, fino a scatole rotanti su cui era stato incollato il piumaggio di una gallina tagliato in piccoli pezzi. Nel secondo test, condotto quando i pulcini avevano circa 52 ore, il modello intatto risultava quello preferito solo quando l'oggetto degradato non possedeva più alcuna caratteristica distinguibile di una gallina selvatica. Inoltre, Johnson e Horn hanno dimostrato che i pulcini non preferivano un modello intatto di gallina rispetto a un oggetto alternativo che conteneva solo la testa e il collo di un soggetto impagliato; ciò suggerisce che le regioni del capo e del collo contengano stimoli particolarmente rilevanti per indurre la predisposizione.
In successivi esperimenti, Johnson e Horn hanno osservato che i pulcini non preferivano una gallina selvatica impagliata rispetto a un'anatra impagliata o anche a una puzzola impagliata. Dunque la predisposizione non è specie-specifica. Sembrerebbe che stimoli posti sul capo dell''oggetto stimolo' possano avere un ruolo importante nell'espressione della predisposizione, mentre gli occhi non costituirebbero uno 'stimolo segnale' indispensabile. È possibile che la selezione dello stimolo coinvolto nella predisposizione si conformi alla regola della sommazione eterogenea, per cui sue differenti caratteristiche contribuiscono in modo indipendente all'induzione della risposta comportamentale: così, gli occhi costituirebbero uno stimolo importante come altri. Alternativamente, si può immaginare che la risposta dei pulcini rappresenti un esempio di apprendimento configurazionale di uno stimolo composto da una testa, dagli occhi, da un collo e un corpo.
Periodi sensibili e imprinting sessuale
Piuttosto che la locuzione 'periodo critico' utilizzata originariamente da Lorenz, che implica un intervallo di tempo fisso, nella ricerca contemporanea viene utilizzata l'espressione 'periodo sensibile'. I dati sulla durata dei periodi sensibili variano ampiamente anche nell'ambito di una stessa specie; si è chiarito che non esiste un periodo sensibile rigido e nettamente definito per l'imprinting filiale. L'esperienza esterna svolge un ruolo importante nel modificare la sensibilità all'imprinting; la fine di un periodo apparentemente sensibile è dovuta in larga misura all'esperienza stessa di imprinting filiale e non è soltanto una conseguenza di modificazioni interne all'individuo. Più avanti vedremo che, anche nel caso di imprinting sessuale, un periodo ritenuto sensibile può rivelarsi meno determinante di quanto ipotizzato e che le preferenze sessuali possono essere influenzate fino al momento in cui si manifesta il comportamento sessuale.
Lorenz per primo suggerì che una delle conseguenze dell'imprinting sarebbe la formazione di una preferenza sessuale per la specie da cui il giovane uccello è stato allevato. Attualmente si opera la distinzione tra imprinting filiale e imprinting sessuale e sono state elencate alcune delle differenze tra imprinting filiale e sessuale, quali la diversa fase della vita in cui i loro effetti si manifestano (età precoce in un caso, età adulta nell'altro) e il diverso periodo di tempo durante il quale l'esperienza influisce sul comportamento (ore o giorni nel primo caso e settimane nel secondo). Inoltre, prove fornite da Paul P.G. Bateson (1982) e da altri ricercatori suggeriscono che, al contrario di quanto Lorenz sosteneva inizialmente, le preferenze sessuali potrebbero essere rivolte agli individui piuttosto che alla specie. In particolare, Bateson ha dimostrato che la preferenza sessuale finale potrebbe rivolgersi a un individuo differente da quelli con cui l'animale è cresciuto.
Negli studi sull'imprinting sessuale gli Uccelli in genere sono allevati fino a una certa età da genitori della propria specie o da genitori adottivi di una specie differente. Le preferenze sessuali vengono poi analizzate quando gli animali sono sessualmente maturi, talora dopo un certo periodo di isolamento sociale. Le prove dell'imprinting sessuale sono in genere dedotte dalle risposte di preferenza verso un membro di sesso opposto della specie che ha allevato il soggetto analizzato (come la tendenza ad avvicinarsi allo stimolo o le risposte di corteggiamento diretto). Tuttavia Bateson ha dimostrato che le quaglie giapponesi (Coturnix coturnix japonica) possono sviluppare una preferenza sessuale per un individuo con caratteristiche precise, e in particolare per un individuo diverso (ma non troppo) da quelli con cui sono cresciute. Bateson ha suggerito che questa risposta preferenziale sia un meccanismo utile per ottenere un grado di esoincrocio ottimale, tale da evitare gli effetti deleteri sia dell'inincrocio sia dell'esoincrocio estremo.
Per un certo periodo si è pensato che predisposizioni specie-specifiche svolgessero un ruolo nell'imprinting sessuale. Per esempio, maschi di diamante mandarino (Taeniopygia guttata castanotis) allevati in successione da genitori di passero del Giappone (Lonchura striata) e da genitori della propria specie, o da una coppia mista (cioè costituita da un genitore di ciascuna di queste due specie), preferirono apparentemente una femmina della propria specie. Tuttavia, Carel ten Cate scoprì che esisteva una differenza significativa tra i genitori della specie diamante mandarino e quelli di passero del Giappone nel numero di interazioni sociali stabilite con i nidiacei. Il grado di preferenza sessuale era correlato alla quantità di interazioni sociali che i giovani maschi avevano ricevuto dai differenti genitori. Inoltre, fu scoperto che nei maschi di diamante mandarino sottoposti ad adozione incrociata vi era una correlazione tra il numero di fratelli presenti nel nido e la successiva preferenza per femmine della propria specie.
Un altro punto importante, una volta stabilito il periodo sensibile, è se la preferenza sessuale sia irreversibile. Esperimenti recenti effettuati sul diamante mandarino in due diversi laboratori hanno modificato considerevolmente questo quadro e hanno dimostrato che in realtà vi è una notevole plasticità nella formazione delle preferenze sessuali. Quando i maschi di diamante mandarino sono allevati da genitori adottivi di passero del Giappone per i primi 40 giorni di vita, nei test di preferenza condotti quando sono diventati adulti mostrano una forte preferenza verso femmine di quest'ultima specie. Quando questi maschi sono esposti per diversi mesi a una femmina della propria specie, manifestano verso di lei un comportamento di corteggiamento ma, se ne vengono successivamente separati, nei test di scelta tornano a preferire una femmina della specie adottiva. Tuttavia, se i maschi di diamante mandarino non sono sottoposti al test di preferenza prima dell'esposizione alla femmina della propria specie, gran parte di essi finisce per indirizzare in maniera stabile il proprio comportamento di corteggiamento verso le femmine di diamante mandarino. In ogni caso, non è necessaria l'interazione fisica diretta con le femmine, né durante i brevi test di preferenza né durante i periodi di più lunga esposizione, quando gli animali sono sessualmente maturi.
Da queste osservazioni si può concludere che le preferenze sessuali possono essere modificate da esposizioni più tardive a una femmina di un'altra specie; inoltre una breve esposizione a una femmina della specie da cui il soggetto è stato allevato è sufficiente a consolidare o stabilizzare la preferenza sessuale dell'animale. Quando i maschi di diamante mandarino allevati da genitori di passero del Giappone sono esposti a femmine della specie adottiva, in seguito corteggiano le femmine di questa specie; tuttavia, quando sono esposti prima a una femmina di diamante mandarino, gran parte dei maschi muta la preferenza rivolgendosi alla femmina della propria specie. Sulla base di questi e di altri risultati, Hans-Joachim Bischof (1994) ha suggerito che l'imprinting sessuale sia un processo che si svolge in due stadi, o periodi sensibili: una fase di acquisizione, in cui l'animale apprende le caratteristiche dei suoi genitori e dei fratelli, e una fase di consolidamento, in cui la preferenza sessuale si stabilizza o si modifica a seconda della specie a cui appartiene l'individuo al quale il maschio viene esposto.
I termini 'mantenimento', 'facilitazione' e 'induzione' furono introdotti da Gilbert Gottlieb per distinguere alcune delle modalità con cui l'esperienza può influenzare lo sviluppo della 'percezione delle caratteristiche della specie'. È stato dimostrato che tutte e tre le modalità possono verificarsi durante lo sviluppo delle preferenze filiali. Hampton e collaboratori sostengono che durante le fasi di vita oggetto dei loro studi e di altri studi simili, gli effetti dell'esperienza sullo sviluppo della predisposizione possono essere classificati come un caso di induzione; la predisposizione inizialmente non è espressa ed emerge solo in seguito a esperienza non specifica. Certamente la terminologia di Gottlieb è descrittiva e non dice nulla sui meccanismi alla base dei fenomeni; tuttavia essa può costituire un utile modello per classificare gli effetti dell'esperienza sullo sviluppo del comportamento.
Il fatto che l'esposizione a differenti tipi di stimoli possa portare all'emergere di una predisposizione costituisce un esempio di induzione di un meccanismo percettivo tramite esperienza non specifica. Altri studi sull'imprinting filiale suggeriscono che lo stesso principio si può applicare alla facilitazione dello sviluppo dei meccanismi percettivi. Nell'imprinting sessuale del diamante mandarino si sono evidenziate prove del mantenimento (definito 'consolidamento' o 'stabilizzazione') delle preferenze sessuali attraverso esperienze non specifiche. Gli effetti dell'induzione, del mantenimento o della facilitazione provocati da un'esperienza non specifica, e apparentemente non correlata, su meccanismi percettivi specifici, sembrano essere un principio fondamentale dello sviluppo comportamentale.
Una conclusione importante che si può trarre da una rassegna delle ricerche recenti è che, in contrasto con quanto affermato originariamente da Lorenz, l'imprinting filiale e quello sessuale non sono fenomeni irreversibili e rigidamente determinati, vincolati a periodi sensibili o a particolari predisposizioni, anche se questi fattori svolgono certamente, in un modo o nell'altro, un ruolo importante. Vi è invece una considerevole plasticità nel sistema, che permette a differenti tipi di esperienze di influenzare la formazione delle preferenze sociali di un animale in differenti fasi del suo sviluppo.
Bibliografia
Bateson 1982: Bateson, Paul P.G., Preferences for cousins in Japanese quail, "Nature", 295, 1982, pp. 236-237.
Bischof 1994: Bischof, Hans-Joachim, Sexual imprinting as a two-stage process, in: Causal mechanisms of behavioural development, edited by Jerry A. Hogan, Johan F. Bolhuis, Cambridge-New York, Cambridge University Press, 1994, pp. 82-97.
Gottlieb 1976: Gottlieb, Gilbert, The roles of experience in the development of behavior and the nervous system, in: Neural and behavioral specificity, edited by Gilbert Gottlieb, New York-London, Academic Press, 1976, pp. 237-280.
Hampton 1995: Hampton, Neil G. - Bolhuis, Johan J. - Horn, Gabriel, Induction and development of a filial predisposition in the chick, "Behaviour", 132, 1995, pp. 451-477.
Horn 1984: Horn, Gabriel - McCabe, Beverly J., Predispositions and preferences. Effects on imprinting of lesions to the chick brain, "Animal behaviour", 32, 1984, pp. 288-292.
Johnson, Horn 1985: Johnson, Mark H. - Bolhuis, Johan J. - Horn, Gabriel, Interaction between acquired preferences and developing predispositions during imprinting, "Animal behaviour", 33, 1985, pp. 1000-1006.
Johnson 1988: Johnson, Mark H. - Horn, Gabriel, Development of filial preferences in dark-reared chicks, "Animal behaviour", 36, 1988, pp. 675-683.
Lorenz 1937: Lorenz, Konrad, The companion in the bird's world, "Auk", 54, 1937, pp. 245-273.
Ten Cate 1989: ten Cate, Carel, Behavioral development: toward understanding processes, in: Perspectives in ethology, edited by Paul P.G. Bateson, Peter H. Klopfer, New York, Plenum, VIII, 1989, pp. 243-269.