
Invecchiamento
Invecchiamento
Con il termine invecchiamento si indica il deteriorarsi progressivo, dovuto al tempo, della capacità di un organismo di adattarsi alle modificazioni ambientali nelle quali vive: questo implica maggiore suscettibilità per stati patologici e maggior rischio di morte. Per alcuni ricercatori l'i. è dovuto alla condizione per la quale fattori che agiscono sullo stato di buona salute correlato all'età (longevità) sono sopraffatti dalla progressiva e fisiologica usura interna dovuta al trascorrere del tempo (senescenza). Questo equilibrio tra meccanismi di senescenza e longevità è alla base della grande variabilità del processo di invecchiamento. Si deve aggiungere che l'i. e la velocità con cui un individuo invecchia sono in parte controllati geneticamente, pertanto la durata della vita è differente sia tra specie diverse sia tra individui della stessa specie. In uno schema a più livelli di complessità, le ipotesi sui possibili meccanismi coinvolti nell'i. biologico possono essere classificate prendendo in considerazione aspetti evoluzionistici, fisiologici, cellulari e molecolari.
Evoluzione e invecchiamento
Da un punto di vista evolutivo, l'i. è stato considerato come un progressivo indebolimento della forza della selezione naturale con l'avanzare dell'età. Infatti, la selezione naturale opera in modo da favorire la selezione di quei geni che sono coinvolti nella sopravvivenza e nella riproduzione della specie; pertanto, ne consegue che l'i. è un epifenomeno o un prodotto passivo, 'di scarto', dell'evoluzione. Per interpretare l'i. in termini evolutivi sono stati proposti diversi meccanismi, ognuno dei quali prende in considerazione un aspetto particolare del fenomeno. Secondo la teoria dell'usura, l'i. è dovuto al progressivo deterioramento cui ogni organismo è sottoposto a causa, per es., dell'accumulo di prodotti tossici del metabolismo e dei danni da radiazioni a carico di proteine e DNA. La teoria delle mutazioni nocive sostiene che gli organismi che invecchiano risentono delle mutazioni sfavorevoli che si sono andate accumulando di generazione in generazione attraverso la linea delle cellule germinali. Secondo la teoria del pleiotropismo antagonista, la selezione naturale favorisce i geni che hanno effetti positivi durante gli stadi precoci della vita di un organismo, ma sono anche responsabili di effetti pericolosi, a stadi più avanzati del ciclo vitale di un individuo. Un gene è definito pleiotropico quando la sua espressione può esercitare la sua influenza su più caratteri e, di conseguenza, con l'espressione pleiotropismo antagonista si intende che alcuni caratteri influenzati da un gene pleiotropico hanno effetti opposti sull'efficienza evolutiva. La teoria dell'usura e riparazione parte dal presupposto che meccanismi genetici sono programmati al fine di contrastare i fenomeni di usura imputabili all'interazione dell'individuo con l'ambiente che lo circonda. Poiché questi meccanismi comportano dei costi in termini di metabolismo, che si riduce con l'età, anche l'efficienza evolutiva dell'individuo è ridotta. Dal confronto tra queste teorie emerge il dato comune che il modo di invecchiare può essere influenzato dall'evoluzione, ma questo può avvenire solo dopo che il processo di i. è stato avviato da meccanismi che sono responsabili tanto del deterioramento delle strutture biologiche quanto delle modificazioni fisiologiche legate alla crescita e alla maturazione dell'individuo.
Invecchiamento degli organismi
Molti studi sostengono che nei mammiferi esiste una stretta correlazione tra durata della vita e peso corporeo. Se invece del peso corporeo si prende in considerazione il peso del cervello, i dati mostrano una più significativa correlazione. In relazione al peso corporeo, la specie umana è caratterizzata dall'avere il cervello più pesante e più complesso dal punto di vista strutturale di ogni altro mammifero, e questo è associato a una vita più lunga. Dalla correlazione tra dimensioni del cervello e longevità si possono estrapolare importanti implicazioni fisiologiche. Più alto è il rapporto tra peso del cervello e peso corporeo, più fini sono i meccanismi di regolazione fisiologica cui presiede il cervello e maggiori sono le possibilità di vivere a lungo. Questa spiegazione è ragionevolmente sostenuta dal fatto che il sistema nervoso gioca un ruolo essenziale nella regolazione di adattamenti fisiologici di importanza vitale, particolarmente in risposta a sollecitazioni ambientali. Mantenere le condizioni fisiologiche a un livello ottimale e ridurre l'ampiezza delle fluttuazioni che si possono verificare con il passare del tempo diminuisce la probabilità che si verifichino modificazioni irreversibili in tempi brevi e, di conseguenza, si riducono anche la velocità con cui si invecchia e l'incidenza di morte.
Invecchiamento cellulare
La consapevolezza di un i. a livello cellulare si è venuta affermando intorno agli anni Sessanta del 20° sec., quando L. Hayflick dimostrò che fibroblasti coltivati in vitro si dividono un numero finito di volte. I fibroblasti umani possono raddoppiarsi in media circa 50 volte, cioè possono andare incontro a 50 cicli di divisione cellulare, poi smettono di proliferare e si fermano allo stadio della divisione cellulare prima della sintesi di DNA (fase G0). Questo limite di duplicazioni cellulari (limite di Hayflick) varia a seconda dell'età del donatore e della specie considerata. Dai risultati degli studi in vitro sul limite di Hayflick, sembra essere probabile che anche nell'organismo le divisioni si fermino dopo un certo limite, ed è probabile che i meccanismi che portano all'arresto della proliferazione cellulare possano essere ritenuti responsabili anche del declino delle difese immunitarie nell'invecchiamento.
Biologia molecolare dell'invecchiamento
Sebbene già all'inizio del 20° sec. si fosse osservato che animali caratterizzati da tasso metabolico più elevato muoiono prima rispetto ad altri, non era ancora chiaro il legame tra elevato metabolismo e invecchiamento. Con lo scopo di interpretare le molteplici alterazioni cellulari tipiche dell'i., D. Harman propose, verso la metà degli anni Cinquanta, la teoria dei radicali liberi, che può costituire una base scientifica attendibile per spiegare il rapporto inverso tra tasso metabolico e longevità. L'ipotesi di Harman parte dal presupposto che i radicali dell'ossigeno sono generati all'interno delle cellule nel corso di processi fisiologici, come la respirazione cellulare, e che questi sono responsabili di danni alle macromolecole biologiche. I radicali liberi possono essere definiti come molecole o atomi con uno o più elettroni spaiati. In tutte le reazioni chimiche, affinché il composto che si forma dall'incontro di due reagenti sia stabile nel tempo, è necessario che gli elettroni del prodotto di reazione siano tutti appaiati, in pratica non abbiano alcuna (o scarsa) possibilità di essere trasferiti ad altre molecole.
Per es., a seguito di radiazioni provenienti dall'ambiente (la luce solare) un elettrone può essere allontanato da uno o più atomi di una determinata molecola e costituire un radicale libero, cioè una specie chimica con forte tendenza ad attrarre elettroni o molecole al fine di appaiare di nuovo i suoi elettroni e raggiungere uno stato ottimale di stabilità. In questo processo di acquisizione di elettroni (che dura pochissime frazioni di secondo), i radicali liberi attaccano e modificano non solo quelle molecole che si trovano nel loro raggio di azione, ma innescano una vera e propria catena di reazioni in cui gli atomi e le molecole coinvolti si sottraggono elettroni a vicenda, per cui, per effetto della propagazione dell'azione dei radicali liberi, il danno finale può avvenire a carico di macromolecole molto lontane dal distretto cellulare in cui il primo spostamento di elettroni è avvenuto. I radicali liberi, la cui maggioranza è costituita da specie reattive dell'ossigeno (ROS, Reactive Oxygen Species), vengono continuamente prodotti dagli organismi viventi. In risposta alla produzione di radicali liberi, gli organismi viventi hanno sviluppato, nel corso dell'evoluzione, non solo sistemi antiossidanti di difesa, ma anche sistemi di riparazione in grado di prevenire l'accumulo di molecole danneggiate dalle ossidazioni. Le difese cellulari sono integrate da molecole con funzione antiossidante come superossidodismutasi, catalasi, glutatione perossidasi, vitamina E e il glutatione stesso. Il meccanismo di azione di queste molecole è quello di rallentare la propagazione delle reazioni innescate dai radicali liberi; in particolare, quelle con funzioni enzimatiche interrompono le reazioni dei radicali liberi degradando le molecole che hanno subito il danno. È da precisare che i radicali liberi possono giocare ruoli importanti in molti processi fisiologici e quindi la loro produzione non può essere completamente soppressa. L'ossido nitrico (NO), per es., è un radicale libero che agisce da neurotrasmettitore, è usato cioè come molecola per trasmettere informazioni tra due cellule nervose. I mitocondri pos-sono essere ragionevolmente ritenuti il bersaglio preferito e più fre-quentemente colpito dai radicali liberi e quindi il danno a carico di questi organelli del metabolismo energetico cellulare rappresenta un'alterazione molto precoce dei meccanismi di invecchiamento. Il danno da radicali liberi a carico del DNA mitocondriale può avere conseguenze molto serie specialmente in cellule che hanno la stessa età dell'organismo al quale appartengono (per es., i neuroni). Calcolando i tempi di ricambio di molecole marcate con isotopi radioattivi, è stato dimostrato che la durata media della vita di un mitocondrio neuronale è di circa 28 giorni; nei neuroni di una persona di circa 80 anni, quindi, i mitocondri si sono duplicati circa 1000 volte. A differenza del DNA nucleare del neurone (che non si duplica perché questo tipo di cellula non si divide dopo la differenziazione), il DNA mitocondriale si duplica ogni volta che un nuovo organello viene generato. Inoltre, in ogni mitocondrio sono presenti 2-10 copie di DNA, e pertanto esistono molte possibilità che vengano commessi errori nello stesso processo di duplicazione e che queste alterazioni vengano trasmesse alle generazioni successive di mitocondri. Una singola cellula del corpo umano utilizza circa 1012 molecole di ossigeno al giorno, generando ogni ora circa 30 miliardi di molecole di perossido di idrogeno (H2O2), molecola molto instabile, fonte di radicali liberi. I tipi di danno da ossidazione possono consistere in ripiegamenti della molecola di DNA o in alterazioni delle caratteristiche fisico-chimiche (basicità) di certi siti, o in vere e proprie rotture della doppia elica del DNA. La severità di questi danni è molto maggiore per il DNA mitocondriale rispetto a quello nucleare: il livello di danno può essere circa 10 volte più alto di quello osservato a carico del DNA nucleare anche perché sono pressoché assenti i meccanismi di riparazione del DNA mitocondriale, e questa molecola non è protetta da proteine (gli istoni), presenti invece nel DNA nucleare. Il ruolo centrale dello 'stato di buona salute' ed efficienza funzionale dei mitocondri nei processi di i. è sostenuto da molti ricercatori, e si basa sulla funzione più importante (ma non unica) che viene assolta da questi organelli: la respirazione cellulare con produzione di energia. Il meccanismo di fosforilazione ossidativa con produzione di ATP, che costituisce la fonte primaria di energia di parecchi organi e tessuti, subisce un forte declino non soltanto con l'i., ma anche in numerosi altri stati patologici.
Centinaia di geni nucleari mitocondriali sono coinvolti nella sintesi di molecole della fosforilazione ossidativa. La maggiore vulnerabilità del DNA mitocondriale rispetto al DNA nucleare tende a far ipotizzare che il declino della fosforilazione ossidativa sia il risultato dell'accumulo di danni da ossidazione al DNA mitocondriale. Dal momento che il genoma mitocondriale è solamente di origine materna, ci si potrebbe aspettare di trovare una stretta corrispondenza, in termini di longevità, tra madre e figli. In realtà non è così, perché ogni cellula può presentarsi con un insieme misto di DNA mitocondriale mutato e normale (eteroplasmia) e, ogni volta che una cellula appartenente alla linea somatica o germinale si divide, sia il DNA mitocondriale normale sia quello mutato sono trasferiti a caso nelle cellule figlie, dando così origine a una marcata diversificazione fra una cellula e l'altra e a un mosaico di cellule caratterizzate da diversa efficienza funzionale nelle loro popolazioni mitocondriali. Questo concetto del mosaico sembra rendere ragione delle alterazioni cellulari a macchia di leopardo osservate nel cervello di soggetti anziani sani e in quello di pazienti affetti da malattia di Alzheimer. Un altro tentati-vo di spiegare l'induzione, la promozione e la progressione dell'i. e di alcune patologie a esso correlate è l'ipotesi delle mutazioni somatiche: i. come conseguenza dell'accumulo di mutazioni del DNA delle cellule somatiche. Queste mutazioni sono in grado di modificare l'espressione genica e alterare le funzioni di alcune importanti proteine, come, per es., i fattori di trascrizione che, legandosi al DNA, ne regolano l'attività genica. Un'altra teoria sostiene che responsabili dell'i. siano da ritenersi i cambiamenti correlati all'età nell'attività dei geni. Questi cambiamenti sono selettivi sia per quanto riguarda il sito genetico sia il tipo di cellula e, se colpiscono geni regolatori del ciclo cellulare, i loro effetti possono essere di grande portata. La duplicazione del DNA è un processo piuttosto complesso, ma si basa su principi molto semplici: in una prima fase, un enzima (la DNA elicasi) controlla la distensione (dallo stato arrotolato) della doppia elica della molecola di DNA, e succes-sivamente un altro enzima (la DNA polimerasi) catalizza la sintesi dell'elica duplicata secondo una ben definita sequenza di reazioni. Nel caso del genoma umano, una copia esatta di 6 miliardi di coppie di basi deve essere portata a termine in 6-12 ore. È stato calcolato che, nel corso della vita di un uomo, il tasso di errore nel copiare il DNA è dello 0,01%, cioè, ogni 10.000 copie di basi una è sbagliata. Ci sono sistemi deputati alla conservazione della precisione del processo di replica del DNA come, per es., il sistema di controllo della lettura che verifica il provvisorio accoppiamento tra l'elica stampo e la nuova elica duplicata che è stata sintetizzata; altri sistemi sono deputati a rilevare modificazioni chimiche come la metilazione di alcune basi.
Questi sistemi riducono significativamente il tasso di errore nella duplicazione del DNA fino al punto che si stima un errore dell'ordine di una base errata ogni 100 milioni. Altre stime mostrano che solo una coppia di nucleotidi su 1000 viene cambiata a caso ogni 200.000 anni. A fronte di questa relativa stabilità della molecola di DNA, le ricom-binazioni genetiche sono responsabili di modificazioni di grande portata del genoma che avvengono con sorprendente frequenza: per es., il genoma può espandersi o contrarsi a causa di duplicazioni o delezioni e le sue parti possono essere trasposte da una regione all'altra per creare nuove combinazioni. Sebbene siano stati chiaramente documentati difetti del genoma e attività di sistemi di riparazione durante l'i. (soprattutto a carico delle cellule del soma) sembra che non ci siano difetti del genoma abbastanza severi da essere ritenuti diretti responsabili del progressivo i. delle cellule. Un'altra caratteristica alterazione degli organismi vecchi è il deterioramento delle proteine dopo la loro sintesi e l'accumulo progressivo di quelle alterate. Questo è dovuto al fatto che, in molte varietà di organismi, la sintesi proteica è meno efficiente con l'avanzare dell'età e questo comporta che anche il ricambio delle proteine usurate risulti rallentato.
Strategie anti-invecchiamento
È ampiamente documentato che adottare una dieta adeguata, ossia 'mangiare meno', fa vivere più a lungo. Il meccanismo (o i meccanismi) alla base di questi benefici effetti sembra essere costituito dalla forte riduzione dello stress ossidativo. Una strategia di risposta allo stress ossidativo è anche quella di aumentare i meccanismi di difesa interni dell'organismo e sembra che il miglior meccanismo di attivazione di questa risposta sia lo stesso stress. Un evento stressogeno controllato può indurre un aumento della sopravvivenza e un ridotto danno a carico dei tessuti dell'organismo in seguito a uno stress successivo, anche se molto più severo. Il termine con il quale è stato definito il concetto è ormesi. Si pensa che, nell'uomo, il ben documentato benefico effetto esercitato da un moderato (e adeguato al singolo soggetto) esercizio fisico sullo stato di salute e sulla longevità possa rappresentare un fenomeno di ormesi. Sia il metabolismo sia lo stress ossidativo aumentano durante e immediatamente dopo un singolo episodio di esercizio fisico. Ciò si può spiegare considerando che la produzione di radicali superossido aumenta e tale fenomeno è dovuto al fatto che con l'esercizio fisico il consumo di ossigeno è circa 10 volte più alto. La risposta dell'organismo all'esercizio fisico implica non solo un rapido adattamento cellulare all'attacco dei radicali liberi, e anche l'attivazione a lungo termine del meccanismo di trasmissione delle informazioni tra cellula e cellula innescato proprio dai radicali liberi. Questi ultimi, infatti, possono attivare diversi fattori di trascrizione: proteine che si legano alla regione promotrice di specifici geni e ne attivano la trascrizione.
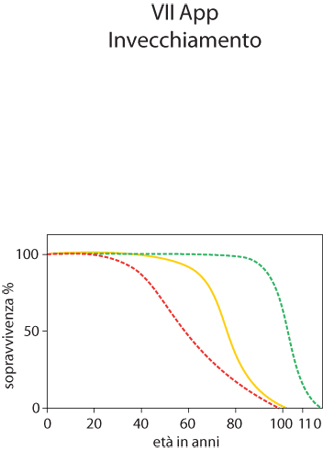
Di fatto questo porta a far scattare immediatamente il processo di sintesi di molecole antiossidanti e altre proteine di difesa contro i radicali liberi. Studi sull'uomo hanno dimostrato che in soggetti adulti che praticano una moderata attività fisica la mortalità è ridotta anche del 44%. È dimostrato che una regolare attività fisica porti in sostanza a una diminuzione del peso corporeo che rappresenta l'anello di congiunzione tra dieta ipocalorica ed esercizio fisico. Ambedue gli interventi sono in grado di modificare significativamente lo stile di vita individuale e questo porta a ottenere benefici che possono migliorare stato di salute e longevità. Nel 1825 l'attuario inglese B. Gompertz formulò un'equazione matematica che descrive la crescita del tasso di mortalità e che ha dato origine all'elaborazione grafica nota come curva di Gompertz. In questa curva (v. figura) riferita agli esseri umani viene preso in considerazione il tasso di sopravvivenza invece del tasso di mortalità. Una combinazione di rischi di natura genetica e ambientale può essere causa di una più rapida diminuzione della percentuale di sopravvissuti: in questo caso la curva (linea gialla intera) si sposta verso sinistra (linea rossa tratteggiata). In condizioni di vita migliori, la durata media della vita aumenta e la curva si sposta verso destra e diventa più 'rettangolare' (linea verde tratteggiata). Molti gerontologi sostengono che esiste la concreta possibilità di una 'rettangolarizzazione' della curva di Gompertz, cioè una modificazione della decelerazione lineare dell'invecchiamento. Ottenere questo risultato implica che ciascun individuo possa vivere in buona salute fino a raggiungere la massima durata di vita (110-120 anni) e muoia senza complicazioni imputabili a stati patologici oppure a malattie invalidanti negli ultimi anni della sua vita.
Considerazioni finali
Allo stato delle conoscenze raggiunte si può affermare che, in quelle popolazioni di organismi che sono caratterizzate da ripetuti periodi riproduttivi nell'arco della vita, con invecchiamento si intende una serie di alterazioni di funzioni e di strutture che si sovrappongono, lentamente e in modo insidioso, dopo il periodo della maturità nel corso della loro vita. La somma totale di queste alterazioni a carattere degenerativo può portare a un progressivo aumento dell'incidenza della mortalità e a un calo nel tasso di riproduzione. Sebbene l'i. sia sicuramente da considerare un processo multifattoriale, la produzione di specie reattive dell'ossigeno (i radicali liberi) e una risposta proporzionata allo stress ossidativo rappresentano i fattori chiave che determinano la longevità. Quello che resta ancora da chiarire è se i radicali liberi agiscano semplicemente a caso come composti pericolosi o se si debba considerare il loro ruolo fisiologico di regolatori di specifici processi biologici direttamente collegati alla capacità di rispondere allo stress e all'invecchiamento.
bibliografia
D. Harman, Aging: a theory based on free radical and radiation chemistry, in Journal of gerontology, 1957, 11, 3, pp. 298-300.
L. Hayflick, The limited in vitro lifetime of human diploid cell strains, in Experimental cell research, 1965, 37, 3, pp. 614-36.
A.W. Linnane, S. Marzuky, S. Ozawa et al., Mitochondrial DNA mutations as an important contributor to aging and degenerative diseases, in Lancet, 1989, 1, pp. 642-45.
D.C. Wallace, Mitochondrial genetics: a paradigm for aging and degenerative diseases?, in Science, 1992, 256, pp. 628-32.
B.N. Ames, M.K. Shigenaga, T.M. Hagen, Mitochondrial decay in aging, in Biochimica et biophysica acta, 1995, 1271, pp. 165-70.