
L'elaborazione dell'informazione per il movimento
L'elaborazione dell'informazione per il movimento
University of Minnesota, Medical School Minneapolis, Minnesota, USA
L'elaborazione dell'informazione per il movimento
Questo saggio ha per oggetto l'elaborazione dell'informazione legata alla direzione del movimento nello spazio, dal punto di vista del comportamento dei neuroni e delle reti neurali artificiali. Verranno analizzati alcuni meccanismi neuronali di codifica nella corteccia motoria dei primati, con riferimento sia a neuroni singoli sia a popolazioni neuronali. Le singole cellule codificano la direzione del movimento secondo una curva che pone in relazione la frequenza di scarica della cellula e la direzione. Le informazioni che provengono da queste cellule selettive rispetto alla direzione permettono di determinare il vettore di popolazione, la cui direzione fornisce una previsione univoca, accurata e consistente della direzione del movimento. La direzione, variabile nel tempo, del vettore di popolazione costituisce uno strumento valido per decifrare i meccanismi neuronali di elaborazione delle informazioni direzionali da parte della corteccia motoria in vari compiti, come i movimenti legati ai tempi di reazione, i compiti motori ritardati appresi e memorizzati e le operazioni cognitive direzionali.
Movimenti di raggiungimento
Il movimento che ci permette di raggiungere un oggetto appare generalmente armonioso e non sembra richiedere alcuno sforzo. Questo atto, apparentemente semplice e naturale, implica però la contrazione e il rilassamento coordinati nel tempo di molti muscoli che agiscono su diverse articolazioni del braccio, ognuna delle quali possiede un certo numero di gradi di libertà di movimento, e che, nell'insieme, costituiscono un sistema enormemente complesso dal punto di vista computazionale. I calcoli che entrerebbero in gioco, se si volesse risolvere in modo diretto il problema che lega l'attivazione muscolare alla realizzazione del movimento, sarebbero tanto complessi da risultare proibitivi. Ciò è dovuto principalmente all'aspetto combinatorio del problema, nel senso che lo stesso obiettivo si può raggiungere con un numero quasi infinito di combinazioni di attivazione dei muscoli e di movimenti delle articolazioni: si può infatti raggiungere un oggetto attraverso molte traiettorie di movimento, che corrispondono a molte combinazioni diverse di muscoli e articolazioni. Fortunatamente, i movimenti vengono di solito effettuati in modo stereotipato, e con molti vincoli, cosicché il numero di gradi di libertà si riduce in maniera significativa. Per esempio, un movimento di raggiungimento coinvolge movimenti ben coordinati e strettamente connessi delle articolazioni della spalla e del gomito che, di fatto, riducono il numero di gradi di libertà. Sembra quindi naturale che il cervello controlli gli arti, nel loro movimento, come un sistema vincolato e connesso, e non come un insieme di unità indipendenti. Considerato da questo punto di vista, un movimento di raggiungimento può essere considerato come un vettore nello spazio, che va dal punto in cui ha origine il movimento fino al punto di arrivo, e che si può descrivere quindi compiutamente attraverso la sua direzione e il suo modulo.
In questo saggio verranno discussi, in particolare, alcuni aspetti dell'elaborazione dell'informazione riguardante la direzione dei movimenti di raggiungimento, e i meccanismi neuronali coinvolti.
Come già menzionato, i movimenti di raggiungimento coinvolgono le articolazioni del gomito e della spalla e sono una conseguenza dell'applicazione di coppie di forze generate in ogni articolazione dalla contrazione dei muscoli. Quali muscoli vengano attivati, e quale sia l'andamento nel tempo dell'intensità della loro contrazione, dipenderà dalla traiettoria del movimento nello spazio, dalla velocità del movimento, e dall'entità e dalla direzione di eventuali carichi esterni, fissi o variabili. Le relazioni tra gli andamenti temporali di queste coppie e la traiettoria della mano sono complicate, anche per traiettorie in due dimensioni. Questo è vero sia che si voglia dedurre la traiettoria, date le coppie (cinematica integrale), sia che si vogliano dedurre le coppie per ogni articolazione, data la traiettoria (cinematica inversa).
La causa della complessità dello schema di attivazione dei muscoli durante i movimenti di raggiungimento è da addebitarsi a molti e diversi fattori. In primo luogo, questi movimenti derivano dalla contrazione concomitante di muscoli differenti. Si deve quindi prendere in considerazione una popolazione di muscoli, anziché una coppia agonista-antagonista, come si fa in genere per i movimenti relativi ad articolazioni con un solo grado di libertà, per esempio quella del ginocchio. In secondo luogo, spesso è difficile misurare il contributo esatto di ciascun muscolo alla coppia generata in ogni articolazione durante il movimento, poiché la direzione lungo la quale un muscolo esercita la sua azione meccanica può variare durante il movimento stesso. Infine, per un'analisi completa dello schema di attivazione muscolare sotteso a questi movimenti si deve tenere conto delle differenze temporali relative tra le contrazioni dei muscoli implicati nel movimento e della loro evoluzione nel tempo, poiché queste differenze possono influenzare la traiettoria del movimento. Il problema è complicato ulteriormente da considerazioni metodologiche. Per esempio, l'attivazione di un muscolo si stabilisce di solito attraverso l'elettromiogramma (EMG), cioè attraverso la registrazione dei fenomeni elettrici che accompagnano l'attività muscolare. Non c'è tuttavia un accordo generale sulla relazione tra l'attività elettromiografica e la forza esercitata dal muscolo. Questo è un problema particolarmente difficile nel caso dei muscoli pennati, le cui fibre non si sviluppano lungo l'asse che connette i punti di ancoraggio del muscolo; in questo caso, per stabilire in modo approssimato la relazione tra l'attività elettromiografica e la forza sviluppata lungo quell' asse, è necessario prendere in considerazione l'organizzazione geometrica delle fibre.
Malgrado queste difficoltà, alcuni principi che regolano l'attivazione dei muscoli durante i movimenti di raggiungimento risultano evidenti: per esempio, in un'analisi elettromio grafica dei movimenti compiuti da scimmie in otto diverse direzioni in un piano si è trovato il coinvolgimento di almeno dieci muscoli, nell' articolazione della spalla e del cingolo scapolare coinvolti (Georgopoulos et al., 1984). L'intensità dell'attivazione elettromiografica era correlata in maniera significativa tra muscoli diversi, suggerendo l'esistenza di sinergie tra i muscoli sottese ai movimenti. Inoltre, queste sinergie variavano in modo regolare con la direzione del movimento. Sebbene si possano applicare considerazioni simili anche ai movimenti liberi di raggiungimento nello spazio tridimensionale, manca al momento una descrizione adeguata delle caratteristiche di attivazione simultanea dei muscoli in questo caso generale.
Teoria dell'informazione
Generalità
Formalmente, l'informazione è definita come una riduzione di incertezza. Secondo le parole di C.E. Shannon, "se il numero di messaggi nell'insieme è finito, si può considerare questo stesso numero, o qualunque funzione monotona di esso, come una misura dell'informazione prodotta quando si sceglie un messaggio dall'insieme, nel caso in cui tutte le scelte siano equiprobabili". Si noti che l'incertezza non dipende dall'evento che si è verificato, ma dall'intero insieme degli eventi che si sarebbero potuti verificare.
La scelta più naturale come misura dell'informazione è una funzione logaritmica di N, invece di N stesso, come è stato messo in evidenza da R.V.L. Hartley nel 1928. Questa funzione risulta più pratica da usare in quanto N potrebbe crescere molto rapidamente al crescere della complessità del sistema. In un sistema di due relè, per esempio, vi sono quattro possibili stati corrispondenti alle condizioni di apertura e chiusura dei contatti dei due relé; l'aggiunta di un ulteriore relè raddoppia il numero di stati possibili. La quantità di incertezza eliminata dal verificarsi di un evento si può considerare come il numero minimo medio di domande "vero o falso" necessarie per ridurre l'incertezza a zero. Questo concetto è intuitivamente più vicino al nostro modo di pensare e ha portato, in modo naturale, all'uso della base 2 per la misura logaritmica dell'informazione. J.W. Tuckey propose la parola bit per indicare questa unità logaritmica; bit è la contrazione delle due parole binary digit (cifra binaria). La quantità di informazione H si può quindi esprimere come
H=10g2N [l]
in cui N è il numero delle alternative equiprobabili. Poiché, in questo caso, la probabilità che si verifichi l'i-esimo evento è
formula [2]

si ha che
formula [3]

Nella realtà, spesso gli eventi non accadono con uguali probabilità. L'equazione [3] indicherebbe quindi che gli eventi con bassa probabilità forniscono più informazione di quelli ad alta probabilità.
Di solito, siamo interessati all'informazione media fornita dal verificarsi di un certo numero di eventi; questa si può calcolare come:
formula [4]

in cui l'informazione per evento è pesata dalla probabilità che quell' evento si verifichi, e si somma su tutti gli eventi i dell'insieme N. La massima quantità possibile di informazione media si ottiene per un insieme di N eventi se questi sono equiprobabili. Sia le variazioni della probabilità degli eventi rispetto al caso equiprobabile, sia l'imposizione eventuale di ulteriori vincoli, per esempio di correlazioni tra gli eventi, hanno l'effetto di ridurre l'informazione.
Da un punto di vista soggettivo, due aspetti fondamentali dell'informazione emergono da quanto detto sopra. In primo luogo, la quantità d'informazione non dipende dalle cause e dagli effetti di un evento; in secondo luogo, l'informazione è una grandezza quantificabile e astratta, il cui valore non dipende dalla natura degli eventi, ma solo dalla probabilità che essi si verifichino; è quindi indipendente dalla metrica o dalle unità usate per la misura di specifici eventi. Questo fatto è importante perché consente, nel contesto della teoria dell'informazione, l'analisi e il confronto di insiemi di eventi di tipo molto diverso. Nell'ambito di questo saggio ciò verrà utilizzato per confrontare aspetti neuronali e comportamentali dei movimenti di raggiungimento.
Applicazioni al comportamento
La prima applicazione della teoria dell'informazione a studi psicofisici si è avuta nel campo della percezione, e si basava sulla analisi di Shannon del canale discreto, pubblicata nel 1948. In breve, se si trasmettono dei segnali di input A, B, C e D attraverso un canale discreto, i cui possibili output sono gli stessi A, B, C e D, si ha una trasmissione senza errori se i messaggi di output corrispondono esattamente a quelli forniti in input al canale; per esempio, se ad A corrisponde A, a B corrisponde B, e così via. In presenza di errori di trasmissione, invece, si possono avere degli errori di identificazione, come nel caso in cui si ricevono come output A o B avendo in input solo A. Le frequenze relative di comparsa degli output corrispondenti a un certo ingresso si possono tabulare costruendo una matrice delle probabilità congiunte input-output, dalla quale è possibile calcolare le incertezze in input e in output e l'informazione trasmessa. Negli studi psicofisici sulla percezione, un insieme di stimoli presentati a un soggetto diventa una sorgente di informazione; il soggetto viene considerato come un canale rumoroso attraverso il quale deve passare l'informazione; la risposta del soggetto rappresenta l'informazione in output. In questo modo, se l'informazione in input dalla sorgente è di r bit, e il soggetto risponde correttamente a ogni stimolo, anche l'informazione trasmessa è di r bit; se il soggetto ignora la sorgente e risponde a caso, l'informazione trasmessa è zero; il caso reale può variare tra questi estremi. Si è trovato, in molti esperimenti condotti su soggetti umani in cui si richiedeva un giudizio assoluto su un insieme di stimoli (sapori, tonalità sonore, intensità luminosa) che, all'aumentare dei livelli in cui lo stimolo è reso discreto, si verifica una saturazione, e l'informazione contenuta nelle risposte cessa di crescere. Il valore massimo dell'informazione trasmessa definisce la capacità del canale per quel compito. Il risultato interessante, e inatteso, è che per diverse modalità sensoriali unidimensionali la capacità del canale rispetto al giudizio assoluto è risultata di circa 2,5 bit per stimolo, che corrisponde a 7 ± 2 alternative (Miller, 1956).
Informazione trasmessa dalla direzione dei movimenti planari del braccio
La teoria dell'informazione può essere applicata in modo simile al sistema motorio; si sceglie un certo parametro relativo al movimento del braccio, come l'ampiezza, la direzione o il punto fmale, e si effettua un esperimento in cui si chiede a un soggetto di compiere dei movimenti secondo un determinato insieme di valori di quel parametro, per esempio di una certa ampiezza, in una certa direzione o verso un certo punto finale. L'insieme dei valori di input del parametro e l'output fornito dal soggetto defmiscono una matrice inputoutput dalla quale si può calcolare l'informazione trasmessa. B. Sakitt e collaboratori (1983) hanno misurato l'informazione trasmessa assumendo come parametro il punto finale dei movimenti del braccio, effettuati in uno spazio bidimensionale e in assenza di retro azione visiva; la capacità del canale per questa variabile è risultata di 3,1 bit.
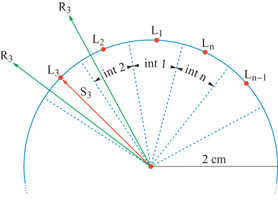
L'informazione trasmessa usando come parametro la direzione dei movimenti di raggiungimento è stata studiata da A.P. Georgopoulos e J.T. Massey (1988). In questi studi, le scimmie (Georgopoulos et al., 1982) o i soggetti umani (Georgopoulos e Massey, 1988) muovevano una leva dal centro di una superficie di lavoro planare verso un bersaglio rappresentato da un punto luminoso su di un cerchio. In questo caso, la direzione del bersaglio rappresenta la direzione ideale: se i movimenti del soggetto fossero delle linee rette dal centro al bersaglio, le sue prestazioni sarebbero perfette, e si potrebbe dire che il movimento ha trasmesso la massima informazione possibile. Viceversa, la direzione dei movimenti dei soggetti varia, in generale, da una prova all'altra, anche per lo stesso bersaglio e, quindi, l'informazione trasmessa è raramente massima: quanto più varia la direzione, tanto minore è l'informazione trasmessa. Ai soggetti veniva richiesto di muovere nella direzione del bersaglio (fig. 1), senza imporre restrizioni sull'ampiezza o sul punto terminale del movimento: in tal modo, veniva realizzato un compito puramente direzionale (Georgopoulos e Massey, 1988).
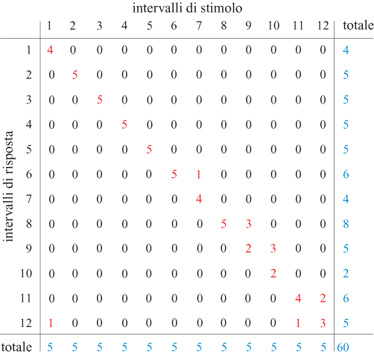
La variabile sperimentale sotto controllo era la posizione della luce bersaglio sul cerchio, che definisce la direzione dello stimolo. Il numero di direzioni dello stimolo veniva variato sistematicamente in un dato insieme di prove, da 4 a 100, corrispondenti a un'incertezza dello stimolo tra 2 e 6,64 bit, rispettivamente. Le direzioni dello stimolo, definite dalla posizione delle luci sul cerchio, venivano distribuite in modo equidistante sull'intero intervallo continuo di direzioni di 360°, con un inizio casuale per ogni insieme di prove. In un dato insieme di esperimenti, ogni direzione dello stimolo veniva presentata cinque volte in modo casuale. Il calcolo dell'informazione trasmessa dalla direzione del movimento richiede la costruzione di una matrice di prestazioni in cui vengono tabulate le direzioni ideali e quelle effettive, e da cui si può calcolare l'informazione trasmessa (Georgopoulos e Massey, 1988). A questo scopo, si è costruita una matrice di prestazioni stimolo-risposta per ogni soggetto e per ogni insieme di prove (fig. 2). La matrice è quadrata; l'ordine della matrice, cioè il numero delle righe e delle colonne, è uguale al numero di direzioni dello stimolo, e gli intervalli sono centrati sulla direzione dello stimolo, come illustrato nella figura 1. Gli intervalli della direzione ottenuta in risposta sono uguali a quelli delle direzioni degli stimoli. L'informazione trasmessa Tviene calcolata nel modo seguente:
T =H(S) +H(R) -H(S,R) [5]
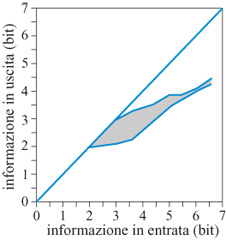
in cui H(S) è l'informazione nell'insieme degli stimoli, H(R) è l'informazione nell'insieme delle risposte, e H(S,R) è l'informazione congiunta nella matrice stimolo-risposta (Georgopoulos e Massey, 1988). Per quanto riguarda i dati mostrati nella figura 2, il numero di direzioni degli stimoli era 12, corrispondente a un'informazione di input di 3,59 bit; l'informazione trasmessa era di 3,11 bit. l risultati su tutti i soggetti sono mostrati nella figura seguente (fig. 3). L'area ombreggiata indica la variazione per soggetti diversi; si può vedere come l'informazione trasmessa cresca con l'informazione in input, ma a un tasso minore del massimo possibile, indicato dalla diagonale. Al livello più alto d'informazione in input che è stato provato negli esperimenti, 6,64 bit, la prestazione migliore è stata di 4,5 bit.
Approccio di Fitts all'informazione motoria
P.M. Fitts (1954) ha applicato un metodo basato sulla teoria dell'informazione alle relazioni che legano accuratezza dei movimenti e distanza. Egli ha innanzitutto definito l'indice di difficoltà del compito (in bit) come
formula [6]

o, in modo equivalente,
formula [7]

in cui W è la larghezza del bersaglio, e A è l'ampiezza del movimento. Una semplice interpretazione di questo indice è che l'informazione maggiore è portata dai movimenti di maggiore accuratezza, cioè con W minore, e di maggiore ampiezza, cioè con maggiore A; questi sono i movimenti più difficili da compiere. Fitts ha anche dedotto una relazione tra la durata del movimento, cioè il tempo del movimento, MT, e l'indice di difficoltà del compito:
formula [8]

in cui k e b sono coefficienti di regressione. L'ultima espressione nell'equazione [8], nota come legge di Fitts, si è dimostrata valida in un'ampia gamma di condizioni sperimentali. Fitts, infine, ha anche definito un indice di prestazione, lp, come il tasso di informazione trasmessa da un singolo movimento:
formula [9]

in cui lp è misurata in bit/s. I limiti superiori dell'informazione trasmessa da soggetti umani variano al variare del compito (Fitts, 1954).
Velocità di elaborazione dell'informazione nei compiti di raggiungimento
I movimenti di raggiungimento sono stati studiati nel contesto di vari compiti sperimentali. Rispetto al movimento stesso, si può operare una distinzione tra i compiti nei quali viene consentito un raggiungimento libero in tre dimensioni, e quelli in cui il movimento di raggiungimento è confinato in una o due dimensioni. Poiché il fatto di vincolare il movimento produce, probabilmente, un effetto sull'elaborazione dell' informazione, si discuteranno questi tipi di compiti in maniera distinta.
Movimenti di raggiungimento non vincolati in tre dimensioni. - In un compito di raggiungimento libero in tre dimensioni (Schwartz et al., 1988) le scimmie compivano movimenti di ampiezza A=12,5 cm verso bersagli visivi di larghezza W=I,6 cm. In queste condizioni, dall'equazione [7] si calcola la = 3,97 bit; considerando che il tempo medio di movimento era di 431 ms, dall'equazione [9] si ottiene che lp = 9,2 bit/s. Si noti che, in un precedente esperimento di raggiungimento in tre dimensioni condotto nel 1975 da V.E. Mountcastle e collaboratori (dati non pubblicati), è stato ottenuto lo stesso valore di lp: le scimmie dovevano raggiungere un bersaglio luminoso posto di fronte; prima dovevano premere una leva, e quando la luminosità del bersaglio si affievoliva dovevano rilasciarla e colpire il bersaglio. Quando diminuiva la luminosità, il bersaglio poteva essere fermo o muoversi a 12 o 21 o/s. In questo caso, il valore di lp era di 9,2 bit/s per il bersaglio fermo, e di 8,3 e 8,4 bit/s rispettivamente per il bersaglio in movimento. Questi valori di 8÷9 bit/s sono quindi ben consolidati per i compiti di raggiungimento in tre dimensioni.
In un altro studio sperimentale, J.N. Sanes (1985) ha osservato dei soggetti umani normali e altri affetti da morbo di Parkinson, durante movimenti ripetuti tra due bersagli. Nel gruppo parkinsoniano è stata trovata una riduzione consistente della velocità di elaborazione dell'informazione. In un caso, venivano effettuati movimenti di 32 cm di ampiezza tra bersagli larghi 0,5 cm, e ld aveva un valore di 7 bit. Il tempo di movimento medio in queste condizioni era di 764 e 1118 ms, rispettivamente per il gruppo normale e per quello parkinsoniano, con conseguenti valori di lp pari a 9,2 bit/s nel gruppo normale, e a 6,3 bit/s nel gruppo parkinsoniano. Il valore di lp ottenuto dai soggetti normali uguaglia quello delle scimmie sottoposte a compiti di raggiungimento in tre dimensioni, mentre quello dei soggetti affetti da morbo di Parkinson è inferiore di quasi 3 bit/s. Questo attesta in modo formale la menomazione principale del morbo di Parkinson, cioè la riduzione della velocità di elaborazione motoria spazio-temporale.
Movimenti di raggiungimento vincolati in due dimensioni. - In un compito di raggiungimento bidimensionale (Massey et al., 1986) i soggetti umani normali e le scimmie muovevano, in diverse direzioni, una leva articolata verso bersagli visivi posti su di una superficie di lavoro piana. All'inizio, i soggetti tenevano l'oggetto al centro del piano, e poi lo muovevano verso bersagli sistemati radialmente, in un compito di misurazione dei tempi di reazione. L'lp medio è risultato di 7,25 bit/s, molto simile nelle scimmie e nei soggetti umani. Questo valore è inferiore a quello ottenuto nel caso dei movimenti non vincolati (9,2 bit/s) e tale riduzione riflette, probabilmente, il vincolo imposto ai movimenti in due dimensioni.
Movimenti del braccio in una dimensione. - Alcune scimmie sono state addestrate a muovere un oggetto con attrito molto basso lungo una dimensione, in risposta a un bersaglio visivo. Alle scimmie venivano mostrate due file di 128 LED (Light Emitting Diodes, diodi a emissione di luce); le due file erano sistemate una sotto l'altra, e la distanza tra due LED adiacenti in una fila era di 2,54 mm. La fila superiore indicava il bersaglio del movimento, quella inferiore la posizione dell'oggetto. La scimmia doveva muovere l'oggetto in modo che il LED inferiore (di retroazione) fosse allineato al LED superiore (bersaglio), in un compito di misurazione dei tempi di reazione. Venivano effettuati movimenti di tre ampiezze (2,5 cm, 6,25 cm, 10 cm). Per i movimenti più ampi, che erano anche i più veloci, si è ottenuto un tempo medio di movimento di 770 ms; considerando che per questi movimenti A= 10 cm e W= 0,67 cm, ld =4,89 bit e lp =6,99 bit/s. Questo valore è leggermente più basso di quello (7,25 bit/s) precedentemente calcolato per i movimenti in due dimensioni. I risultati sin qui discussi indicano che la velocità di elaborazione dell'informazione motoria cresce con il numero di gradi di libertà disponibili per il movimento. Queste velocità risultano essere pari a 6,99 bit/s per i movimenti in una dimensione e, rispettivamente, a 7,25 bit/s e a 9,2 bit/s, per i movimenti in due e in tre dimensioni. Questo fatto è alquanto paradossale, poiché ci si aspetterebbe che la velocità di elaborazione diminuisca quando viene effettuato un movimento in un numero maggiore di dimensioni. Una spiegazione plausibile è che i movimenti non vincolati (in tre dimensioni) siano i più naturali per il sistema motorio, e la limitazione della loro dimensione con dispositivi artificiali, per esempio con l'utilizzo di vari tipi di leve, aumenti la richiesta informazionale, includendo quella relativa all'uso di tali oggetti, e causi velocità di elaborazione dell'informazione più basse.
Velocità di elaborazione dell'informazione in una sequenza di movimenti
Nei compiti descritti nel paragrafo precedente, venivano eseguiti movimenti singoli. Descriveremo ora un altro esperimento in cui, al contrario, vengono effettuati movimenti bidimensionali di raggiungimento, in rapida successione, in risposta a uno spostamento del bersaglio (Massey et al., 1986). In questo esperimento si richiedeva ai soggetti di indicare un bersaglio visivo più rapidamente possibile. Poco tempo dopo la comparsa del primo bersaglio (50÷400 ms) esso si spostava, e il soggetto doveva muovere verso il bersaglio nuovo. Si è trovato che la velocità di elaborazione dell'informazione aumentava in maniera significativa (fino a un fattore 2) durante il secondo movimento. La velocità più alta (14,4 bit/s) si otteneva quando il bersaglio si spostava 150 ms dopo la sua prima comparsa, quasi il doppio rispetto alla velocità di 7,25 bit/s osservata per il movimento verso un bersaglio fisso. Questi risultati mostrano che è possibile incrementare la velocità di elaborazione dell'informazione del sistema motorio, aumentare cioè la velocità del movimento, senza degradare la sua accuratezza. In un certo modo questo caratterizza le prestazioni motorie di destrezza. Per esempio, un buon giocatore di tennis è spesso sia veloce sia accurato, particolarmente se si considera che l'accuratezza richiesta in queste circostanze copre diversi aspetti del movimento, per esempio l'inclinazione della mano, la forza nel colpire la palla, la scelta precisa dei tempi. Occorre considerare in che modo si ottiene un incremento nell'elaborazione dell'informazione. Noi suggeriamo che un fattore chiave responsabile del miglioramento delle prestazioni nelle abilità motorie che implicano delle sequenze di movimenti possa essere un'elaborazione motoria parallela. Sembra che, in certe condizioni, la generazione di un movimento, mentre un altro si sta evolvendo o viene generato, preattiva il sistema percettivo-motorio, cosicché esso, invece di esserne vincolato, elabora l'informazione in modo più efficiente e produce movimenti più veloci, e altrettanto accurati, di quelli generati in isolamento. F. Soechting e F. Lacquaniti hanno descritto un fenomeno correlato a questo incremento di efficienza nell' elaborazione dell'informazione evidenziando che, nel rispondere a un cambiamento del bersaglio, si verifica una riduzione dei gradi di libertà del movimento; questo si ottiene imponendo dei vincoli alle variabili cinematiche, per esempio decelerazioni angolari all'articolazione della spalla e del gomito, e generando degli schemi di attività più stereotipati nei muscoli che agiscono su queste articolazioni.
Aspetti neuronali dell'elaborazione dell'informazione motoria
Generalità
Vi sono diverse aree cerebrali coinvolte nell'innesco e nel controllo dei movimenti di raggiungimento. Lo studio del ruolo delle varie aree in questa funzione è divenuto possibile grazie allo sviluppo di una tecnica che ha consentito la registrazione dell'attività delle cellule singole nel cervello di animali attivi, durante i movimenti di raggiungimento. Questa tecnica è indispensabile per lo studio dei meccanismi neuronali che sono alla base degli aspetti motori del comportamento. Tipicamente, si addestrano le scimmie a svolgere compiti motori diversi; successivamente attraverso la dura madre vengono inseriti microelettrodi nell'area cerebrale in questione per registrare in modo extracellulare i potenziali d'azione isolati delle singole cellule. Questo tipo di esperimento comportamentale e neurofisiologico fornisce uno strumento diretto e dotato di buona risoluzione, attraverso il quale si possono studiare i meccanismi cerebrali che sono alla base delle prestazioni motorie. Un risultato importante di questi studi è che diverse aree cerebrali sono coinvolte nei movimenti di raggiungimento, tra cui sia aree della corteccia cerebrale sia varie strutture sottocorticali.
R. Porter e M.M. Lewis hanno studiato i cambiamenti di attività delle cellule corticali motorie che si verificavano nelle scimmie mentre esse raggiungevano e manipolavano una maniglia posta di fronte a loro. Si è trovato che le singole cellule cambiavano la loro attività durante lo svolgimento del compito, e che la latenza dell'attivazione di cellule diverse si spostava verso tempi successivi via via che parti sempre più distali dell'arto venivano coinvolte nell'atto motorio. L'attivazione sequenziale delle popolazioni corti cali motorie nel raggiungimento e nella prensione è stata studiata in maggior dettaglio da J.T. Murphye collaboratori (1985), registrando l'attività di singole cellule nell'area della corteccia motoria pertinente agli arti anteriori, in un compito in cui le scimmie puntavano a bersagli posti di fronte a loro. La relazione funzionale tra il sito di registrazione e l'articolazione del braccio è stata determinata attraverso microstimolazioni intracorticali. In questo modo è stato possibile classificare le diverse cellule in base al tipo di movimenti a cui esse erano principalmente correlate (movimenti della spalla, del gomito, della mano o delle dita). Si è trovato che nel compito di puntamento le cellule erano in genere attivate in sequenza, da quelle prossimali a quelle distali, in un modo che riflette il coinvolgimento sequenziale delle parti via via più distali del braccio.
Murphy e collaboratori hanno anche studiato le possibili relazioni tra l'attività cellulare a livello della corteccia motoria, il movimento articolare e l'attività elettromiografica nei muscoli dell'arto anteriore durante il raggiungimento, ottenendo tre risultati principali. In primo luogo, non si è osservata nessuna relazione semplice tra l'attività di singole cellule e l'attività elettromiografica, anche quando il muscolo da cui veniva registrata tale attività era attivato da microstimolazioni intracorticali. In secondo luogo, le cellule connesse al movimento delle articolazioni della spalla o del gomito si sono comportate in modo simile nel compito in questione, sebbene i movimenti prodotti in queste arti colazioni potessero essere molto diversi. In ultimo, l'emissione di impulsi da parte delle cellule connesse alla spalla sembrava variare in modo sistematico con la traiettoria del movimento. Questi risultati indicano che le relazioni tra l'attività delle singole cellule della corteccia motoria e le componenti del movimento di raggiungimento (rotazione dell'articolazione, attività elettromiografica) sono complesse.
Codifica della direzione del movimento da parte di cellule singole

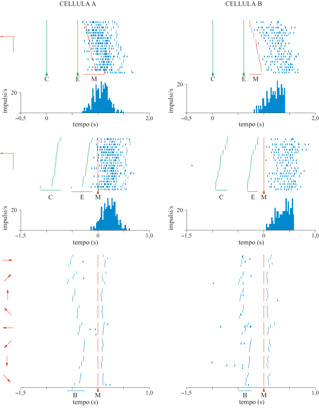
Un movimento di raggiungimento è un vettore nello spazio che può essere descritto completamente dalla sua direzione e dalla sua ampiezza, relativamente al punto di origine del movimento. Georgopoulos e collaboratori hanno iniziato nei primi anni Ottanta lo studio della codifica neuronaIe di questi aspetti spaziali, cioè la direzione e l'ampiezza dei movimenti di raggiungimento. Nel 1982 è stato pubblicato il primo studio sistematico delle relazioni tra l'attività delle cellule corticali motorie e la direzione del raggiungimento (Georgopoulos et al., 1982). Questo studio fu integrato, undici anni dopo, da un'analisi esaustiva delle relazioni tra l'attività cellulare e l'ampiezza del movimento, effettuata da T. Ebner e collaboratori. Il problema di come varia l'attività di una cellula rispetto alla direzione del movimento della mano nello spazio è stato esaminato addestrando delle scimmie a muovere la mano verso bersagli visivi su di un piano o nello spazio tridimensionale (fig. 4), e registrando l'emissione di impulsi dei singoli neuroni in specifiche aree del cervello durante la prestazione motoria.
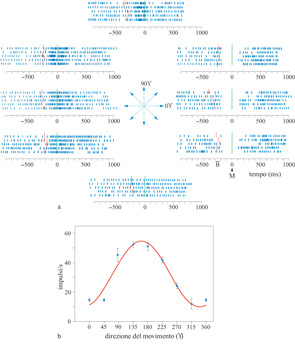
L'emissione di impulsi registrata viene rappresentata graficamente sia in forma di raster, sia riportando la frequenza di scarica in funzione della direzione del movimento (fig. 5). Queste analisi hanno rivelato che l'attività cellulare varia in maniera regolare con la direzione del movimento. In particolare, la scarica è maggiore quando il movimento si svolge in una certa direzione, cioè quella preferenziale della cellula, e diminuisce progressivamente per movimenti compiuti in direzioni via via più lontane da quella preferenziale.
Questi cambiamenti dell'attività cellulare sono correlati alla direzione, non al punto terminale del movimento di raggiungimento (Georgopoulos et al., 1984). Da un punto di vista quantitativo, l'attività della cellula è una funzione lineare del coseno dell'angolo tra la direzione preferita della cellula e la direzione di un certo movimento
di(Mk) = bi + ai cos θCiMk [10]

in cui di (Mk) rappresenta la scarica dell'i-esima cellula, quando il movimento è nella direzione Mk, bi e ai sono coefficienti di regressione, e B rappresenta l'angolo formato tra la direzione preferita della cellula, Ci, e la direzione del movimento Mk. L'equazione [10] è valida sia per movimenti di raggiungimento bidimensionali effettuati su un piano (Georgopoulos et al., 1982) sia per movimenti tridimensionali liberi (fig. 6) (Schwartz et al., 1988). Le direzioni preferite sono differenti per cellule diverse, e sono distribuite su tutte le direzioni nelle tre dimensioni (Schwartz et al., 1988). Esse rimangono molto simili per movimenti di diversa ampiezza, ma tendono a cambiare a seconda della postura del braccio. Coppie di cellule con direzioni preferite simili tendono a mostrare interazioni sinaptiche eccitatorie, mentre coppie di cellule con direzioni preferite opposte tendono a mostrare interazioni sinaptiche inibitorie (Georgoponlos et al., 1993).
Codifica della direzione del movimento da parte di popolazioni neuronali
Il controllo neuronale della direzione del movimento nello spazio si può interpretare in termini di cellule sensibili alla direzione che, come popolazione neuronale, forniscono un segnale direzionale unico e non ambiguo. Data una particolare direzione di movimento, questa popolazione si può considerare formalmente come un insieme di 'vettori cellulari', nel modo seguente: una cellula è rappresentata come un vettore che giace lungo la direzione preferenziale della cellula stessa; la lunghezza del vettore associato alla cellula, per un dato movimento del braccio, rappresenta il cambiamento della attività della cellula durante quel particolare movimento; il vettore risultante dalla somma di questi vettori fornisce il risultato univoco della codifica di insieme, ed è chiamato vettore neuronale di popolazione (Georgopoulos et al.,1983a; 1986; 1988). Questaoperazione si può esprimere nel modo seguente:
formula [11]

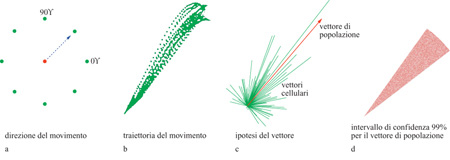

in cui Pk è il vettore di popolazione per la direzione di movimento k, Wik è il peso relativo all'i-esima cellula per la direzione di movimento k, e α è la direzione preferenziale della i-esima cellula. Data la direzione preferita, α è un vettore di lunghezza unitaria nello spazio e il peso Wik determina essenzialmente la lunghezza del vettore Pk, ed è solitamente una funzione della variazione di attività della cellula rispetto a un livello di riferimento (Georgopoulos et al., 1988). Si è trovato (Georgopoulos et al., 1983a; 1986; 1988) che il vettore di popolazione punta nella direzione del movimento (fig. 9) e della forza dinamica isometrica (Georgopoulos et al., 1992). Nelle figure seguenti è rappresentata la codifica neuronale nella direzione di raggiungimento in due dimensioni e in tre dimensioni (figg. 10, 11). La figura seguente (fig. 12) illustra la variabilità direzionale intorno al vettore di popolazione.
Vi sono tre aspetti notevoli dell'analisi in termini di vettore di popolazione: cioè la sua semplicità, la sua robustezza e la sua risposta spaziale. Rispetto alla semplicità, il calcolo del vettore di popolazione è una procedura semplice: si basa sulla selettività direzionale delle singole cellule, che risulta evidente; implica l'attribuzione di un peso ai contributi vettoriali delle singole cellule sulla base dei cambiamenti delle loro attività, il che è ragionevole; si fonda sulla somma vettoriale di questi contributi, che è in pratica la procedura più semplice per ottenere un risultato univoco. In effetti, un aspetto importante dell' analisi basata sul vettore di popolazione è che essa si fonda sulla selettività direzionale, quale risulta definita operativamente dalla procedura sopra descritta; non viene fatta, né si richiede, nessuna ipotesi particolare sull' origine di questa selettività.

Per quanto riguarda la robustezza, il vettore di popolazione è un codice distribuito, e come tale non dipende in modo esclusivo da nessuna cellula in particolare. La sua robustezza è evidenziata dal fatto che può fornire un buon segnale direzionale con un piccolo numero di cellule. Per esempio, mentre il metodo originale (Georgopoulos et al., 1988) richiedeva circa 100 cellule, l'applicazione di procedure nuove e ottimizzate, cioè il metodo della stima lineare (Salinas e Abbott, 1994), ha ridotto il numero di cellule necessarie a meno di 10 (fig.13). Il vettore di popolazione fornisce ancora una stima non di storta della direzione del movimento quando cambia la postura del braccio, anche nel caso in cui le direzioni preferite delle singole cellule possono cambiare al variare del punto di origine del movimento. Infine, il vettore di popolazione è una misura spaziale, isomorfa alla direzione del movimento nello spazio. In effetti, l'analisi basata sul vettore di popolazione trasforma insiemi di sequenze di impulsi puramente temporali, cioè le scariche delle cellule neuronali eccitate, in un segnale direzionale.
L'approccio basato sul vettore di popolazione si è dimostrato utile non solo nello studio della corteccia motoria, ma anche in quello di altre aree cerebrali, tra le quali il cervelletto, la corteccia premotoria, l'area 5, l'area 7 e la corteccia superiore e inferotemporale (Georgopoulos et al., 1993).
Informazione direzionale trasmessa dal vettore neuronale di popolazione
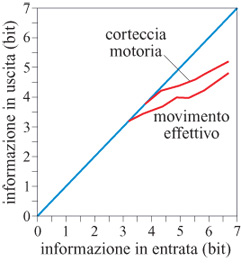
La tecnica usata per calcolare l'informazione trasmessa dalla direzione del movimento si può anche applicare per valutare l'informazione trasmessa dalla direzione del vettore di popolazione (Georgopoulos e Massey, 1988). Poiché il vettore di popolazione è la somma vettoriale dei contributi pesati delle singole cellule, e poiché i pesi possono variare da prova a prova a causa della variabilità tra le prove sperimentali delle scariche neuronali, la direzione del vettore di popolazione può cambiare anch'essa in modo sensibile da prova a prova. Questa variazione si può trattare esattamente nello stesso modo della direzione del movimento e si può calcolare l'informazione trasmessa. L'informazione trasmessa dalla direzione del movimento e dalla direzione del vettore di popolazione è stata calcolata effettivamente (Georgopoulos e Massey, 1988); si è trovato che, in primo luogo, l'informazione trasmessa da entrambe queste misure cresce con l'informazione di input, ma più lentamente del massimo possibile, e tende a un valore di saturazione (fig. 14). Tale perdita di informazione è probabilmente dovuta al rumore generato durante lo stadio iniziale (percettivo) di elaborazione, e quello successivo (percettivo-motorio). In secondo luogo, l'informazione trasmessa dal vettore di popolazione è risultata sistematicamente più alta di circa 0,5 bit rispetto a quella trasmessa dal movimento; questo significa che c'è un'ulteriore perdita di informazione tra la corteccia motoria e il movimento. Questa perdita, però, è diversa dalla precedente, poiché non cresce all'aumentare dell'informazione sullo stimolo ma rimane costante a circa 0,5 bit per tutti i livelli di informazione in input. Essa potrebbe verificarsi durante l'elaborazione in altre strutture motorie o a livello della realizzazione biomeccanica del movimento.
Codifica della direzione della forza isometrica
Recentemente sono state studiate le relazioni tra l'attività delle cellule nella corteccia motoria e la direzione della forza isometrica in due dimensioni (Georgopoulos et al., 1992). Descriveremo ora un esperimento che ha consentito la separazione tra le componenti dinamica e statica della forza esercitata. Scimmie opportunamente addestrate applicavano forze di tipo impulsivo su di una maniglia isometrica, in presenza di una forza costante di deviazione, cosicché la forza risultante, cioè la somma vettoriale della forza esercitata dalla scimmia e della forza di deviazione, risultava in una determinata direzione che veniva visualizzata. Si richiedeva che la forza risultante sviluppata nel tempo rimanesse nella direzione specificata e crescesse in intensità in modo da superare una soglia di intensità richiesta. Il caso più interessante è quello in cui le direzioni della forza risultante e di quella di deviazione sono diverse, per esempio ortogonali. Affinché il compito sia svolto con successo in queste condizioni, la forza dell'animale deve cambiare continuamente in direzione e in intensità, cosicché, in un momento qualsiasi durante lo sviluppo della forza, la somma vettoriale di questa forza e di quella di deviazione sia nella direzione specificata visivamente. Questo tipo di esperimento separava in modo efficace il vettore della forza dell'animale, la cui direzione variava continuamente durante la prova, dal vettore della forza risultante, la cui direzione rimaneva invariata. Sono state utilizzate otto direzioni della forza risultante e otto direzioni della forza di deviazione. Le registrazioni dell'attività neuronale nella corteccia motoria hanno rivelato che l'attività delle singole cellule era direzionalmente selettiva in assenza della forza di deviazione, e che questa selettività permaneva senza variazioni quando veniva prodotta la stessa forza risultante, in presenza di diverse direzioni della forza di deviazione.
Questi risultati hanno dimostrato che l'attività delle cellule non è in relazione con la direzione della forza esercitata dall'animale. Poiché la forza risultante è equivalente alla componente dinamica della forza esercitata dall' animale, dopo aver sottratto una componente vettoriale statica (uguale e opposta alla forza di deviazione) questi risultati suggeriscono che la corteccia motoria fornisce il segnale di forza dinamica durante lo sviluppo della forza; altre strutture, forse sottocorticali, potrebbero fornire il segnale statico di compensazione. Quest'ultimo può essere fornito da sistemi neuronali antigravità, dato che la maggior parte dei carichi statici incontrati sono di natura gravitazionale. Secondo questa visione generale, la forza esercitata dal soggetto consiste in una componente dinamica e in una statica, ognuna delle quali è controllata da sistemi neuronali diversi; questi segnali convergerebbero nel midollo spinale e fornirebbero un segnale integrato dinamico agli insiemi di motoneuroni. Questi risultati hanno stabilito che la codifica dell'informazione direzionale si applica all'output motorio in generale, anche in assenza di movimento dell'articolazione. Inoltre, la direzione specificata dalla corteccia motoria non è quella della forza totale esercitata dall' animale, ma quella della componente dinamica della forza, cioè la componente della forza che rimane dopo aver sottratto una forza statica, costante.
Elaborazione dell'informazione direzionale in vari compiti
L'analisi basata sul vettore neuronale di popolazione è stata applicata con successo per la soluzione di vari problemi concernenti l'elaborazione dell'informazione nel tempo.
Movimenti continui di tracciamento
Il vettore neuronale di popolazione predice la traiettoria inmovimenti continui di tracciamento, come è stato osservato da A.B. Schwartz. In questo esperimento alcune scimmie erano addestrate a tracciare in modo regolare con il dito indice delle sinusoidi visualizzate su uno schermo, da un estremo all'altro. La direzione dei vettori di popolazione, calcolati in istanti successivi lungo la traiettoria, cambiava nel corso di tutto il movimento sinusoidale, in stretta corrispondenza con la direzione del percorso seguito dal dito nella sua variazione regolare. Inoltre, unendo le estremità dei successivi vettori di popolazione, si è ottenuta una 'immagine' della traiettoria sinusoidale del movimento. Poiché la lunghezza di questi vettori di popolazione, variabili nel tempo, era calcolata per un intervallo temporale costante, questo risultato suggerisce che la lunghezza del vettore di popolazione porta informazione circa la velocità istantanea, cioè per intervallo temporale, del movimento.
Tempo di reazione

Il vettore neuronale di popolazione predice la direzione del movimento di raggiungimento durante il tempo di reazione; questo rappresenta il caso più semplice di previsione nel tempo della direzione di un movimento imminente. Dato che i cambiamenti di attività cellulare nella corteccia motoria precedono l'inizio del movimento, in media, di circa 160÷180 ms (Georgopoulos et al., 1982), è importante aver scoperto che il vettore di popolazione predice la direzione del movimento imminente durante il periodo in cui il movimento viene programmato (Georgopoulos et al., 1984; 1988). La figura (fig. 15) mostra un esempio di questo fatto. Inoltre, gli aspetti essenziali del movimento in preparazione nello spazio tridimensionale risultano descritti in maniera accurata da una serie temporale di vettori di popolazione connessi come descritto sopra.
Elaborazione dell'informazione direzionale durante un ritardo 'istruito'

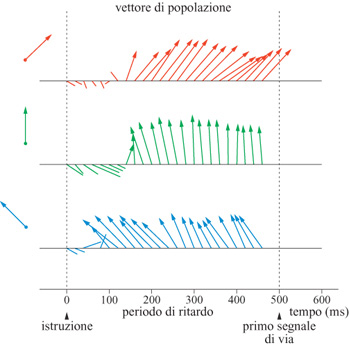
Le cellule della corteccia motoria sono state studiate tradizionalmente nell'ambito dell'esecuzione di un atto motorio. J. Tanji e E.V. Evarts, nel 1976, hanno utilizzato un compito più complesso in cui venivano presentati due stimoli: dapprima uno stimolo visivo 'di istruzione' (l'accensione di un punto luminoso in una di due posizioni periferiche) che indicava alla scimmia la direzione (spingi o tira) del movimento successivo, poi una perturbazione somestesica alla mano che impugnava la leva per innescare il movimento. Si è trovato che 103 neuroni del sistema piramidale su 122 (84%), e 132 su 137 (96,4%) dei neuroni non appartenenti al sistema piramidale hanno cambiato la loro attività entro 0,5 s seguenti lo stimolo di istruzione e prima della somministrazione della perturbazione di innesco. Non è stato osservato nessun cambiamento nell'attività elettromio grafica dei muscoli durante il periodo di attesa. Questi risultati hanno mostrato che gli schemi di attività delle cellule corticali motorie dipendono dal modello comportamentale usato e che possono verificarsi cambiamenti nell'attività delle cellule corticali motorie durante il periodo di istruzione, cioè in assenza di un'uscita motoria immediata. Cambiamenti nell'attività delle cellule corticali motorie durante un periodo di attesa precedente un movimento sono stati osservati, anche in diversi studi successivi. I cambiamenti nell'attività neuronale, connessi alla direzione di movimenti bidimensionali, in compiti con o senza ritardo 'istruito', sono stati esaminati in uno studio (Georgopoulos et al., 1989a) in cui si presentava uno stimolo visivo come bersaglio del movimento, ma le scimmie erano addestrate a trattenere il movimento fino all'abbassamento di una luce. Lo scopo dello studio era di determinare se cambiamenti nell'attività delle cellule corticali motorie, durante il periodo di ritardo, forniscono informazione per quanto riguarda la direzione del movimento seguente nello spazio bidimensionale. Durante il periodo di ritardo, venivano effettivamente osservati cambiamenti nell' attività delle cellule (fig. 17), ed essi erano spesso, ma non sempre, congruenti con i cambiamenti osservati nei compiti senza ritardo. Inoltre, c'era un sottoinsieme di cellule che non mostrava cambiamenti durante il periodo di ritardo, sebbene lo facesse nel compito senza ritardo; questo significa che i cambiamenti 'anticipatori' riguardano solo certe cellule. Infine, un risultato rilevante di questo studio è stato il fatto che il vettore neuronale di popolazione, calcolato durante il periodo di ritardo, prediceva bene la direzione del movimento da eseguirsi successivamente (fig.18).
Il fatto che si possa visualizzare un comando motorio in assenza di un movimento immediato, offre nuove opportunità per lo studio dei correlati neuronali dei processi nascosti spazio-motori, come le trasformazioni direzionali, o per la visualizzazione delle rappresentazioni neuronali delle traiettorie spaziali con cambiamenti di direzione (v. oltre).
Elaborazione dell'informazione direzionale durante il mantenimento in memoria

Sono stati compiuti vari studi finalizzati all'identificazione dei segnali neuronali connessi al mantenimento in memoria di informazioni concernenti la direzione di un movimento imminente (Smyrnis et al., 1992). A questo scopo, inizialmente le scimmie sono state addestrate a muovere una maniglia su di un piano in direzioni specificate da uno stimolo visivo su di esso: la maniglia era posizionata in corrispondenza di una luce al centro del piano e, successivamente, veniva mossa nella direzione indicata da una luce periferica (segnale di indizio). Il segnale per il movimento (segnale di via) veniva dato spegnendo la luce centrale. Nel compito senza ritardo la luce periferica veniva accesa nello stesso istante in cui quella centrale veniva spenta. Nel compito con ritardo memorizzato la luce periferica rimaneva accesa per 300 ms (periodo di indizio) e la luce centrale veniva spenta 450÷750 ms dopo (periodo di ritardo). Infine, nel compito con ritardo non memorizzato la luce periferica rimaneva sempre accesa, mentre quella centrale si spegneva 750÷1050 ms dopo l'accensione della luce periferica. Le registrazioni compiute nell'area della corteccia motoria corrispondente al braccio hanno messo in evidenza cambiamenti nell'attività delle singole cellule in tutti i compiti. Il vettore di popolazione veniva calcolato ogni 20 ms, dopo l'accensione della luce periferica. Sono stati studiati due aspetti dell'informazione associata al vettore di popolazione: uno riguarda la sua direzione, che si può interpretare come l'informazione portata dal segnale di popolazione; l'altro aspetto riguarda la lunghezza del vettore di popolazione, che si può considerare come l'intensità del segnale direzionale. La direzione del vettore di popolazione durante il compito con ritardo memorizzato (fig.19) era vicina alla direzione del bersaglio, che era la stessa del movimento seguente.
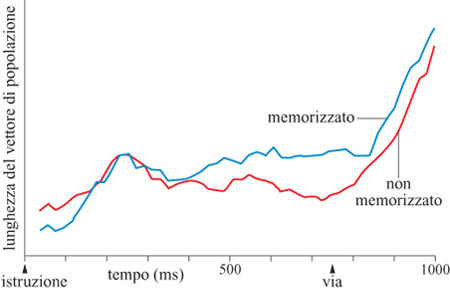
Per quanto riguarda la lunghezza del vettore di popolazione (fig. 20), è interessante il fatto che, rispetto alla parte non memorizzata del ritardo, essa risultava simile nel periodo di indizio, ma maggiore durante la parte memorizzata del ritardo. In questa evoluzione temporale sono state distinte tre fasi. Nella prima, il vettore di popolazione ha mostrato un aumento iniziale di lunghezza, con inizio a circa 100 ms dopo la comparsa dell'indizio, e con un massimo a 250 ms. Interpretiamo questo picco iniziale come l'espressione di un processo di codifica. Questo aumento è risultato molto simile nei compiti con ritardo memorizzato e non memorizzato. Seguiva una seconda fase, che differiva nei compiti con ritardo memorizzato e non, in quanto vi era un segnale persistente e forte durante il periodo di ritardo memorizzato, assente in quello non memorizzato. Questo sembra riflettere il processo di mantenimento in memoria. Infine, dopo il segnale di 'via', la lunghezza del vettore di popolazione cresceva in modo simile in tutti i compiti, riflettendo il processo di generazione del movimento.
L'informazione direzionale, portata dal vettore di popolazione nel compito con ritardo memorizzato, ha identificato l'informazione memorizzata in modo diretto. Inoltre, quest'analisi ha gettato luce sull'evoluzione temporale della codifica e del mantenimento in memoria dell'informazione direzionale. Un aspetto interessante di questi risultati è che l'aumento del segnale, durante il periodo di ritardo memorizzato, è stato osservato in assenza del bersaglio, sebbene le aspettative fossero per un segnale più forte in presenza, invece che in assenza, dello stimolo visivo. Questo fatto rafforza l'interpretazione di questo aumento come un segnale di memoria, anziché come un segnale sensoriale, e suggerisce l'ipotesi più generale che la corteccia motoria possa essere particolarmente coinvolta quando viene fornita solo una parte dell'informazione visiva riguardante un movimento imminente.
Traiettorie complesse memorizzate
Gli studi che abbiamo riassunto sopra riguardavano l'attività corticale motoria durante un compito che richiedeva la memorizzazione della direzione di un movimento rettilineo. Una questione diversa è quella dei meccanismi neuronali sottesi alle traiettorie memorizzate di movimenti complessi. Questo problema è stato analizzato in uno studio recente (Ashe et al., 1993), in cui alcune scimmie erano addestrate a effettuare a memoria un movimento ad angolo retto del braccio, verso l'alto e a sinistra, dopo un periodo di attesa. Le scimmie tenevano una manopola bidimensionale al di sopra di una luce localizzata al centro della superficie planare di lavoro. Quando questa luce si spegneva, agli animali era richiesto di tenere ferma la leva per almeno 600 ms e, quindi, di muovere la manopola in alto e a sinistra per ricevere un succo come ricompensa. Non vi erano segnali esterni indicanti il tempo di via o la traiettoria del movimento. Dopo 20 prove della traiettoria di movimento memorizzata, sono state effettuate 40 prove di movimenti innescati in modo visivo, in direzioni radiali. È stata registrata l'attività di 137 cellule della corteccia motoria durante lo svolgimento del compito. Una buona percentuale (62,8%) cambiava attività durante il periodo di attesa. L'attività di altre cellule non mutava fino a che non era terminato il tempo minimo di attesa di 600 ms e, a volte, l'attività cellulare cambiava quasi esattamente 600 ms dopo lo spegnimento della luce centrale. L'osservazione più interessante, comunque, è consistita nel fatto che un piccolo numero di cellule cambiava l'attività esclusivamente durante l'esecuzione del movimento memorizzato (fig. 21); queste cellule erano completamente inattive durante lo svolgimento di movimenti simili nel compito di controllo guidato visivamente.
Questi risultati suggeriscono che l'esecuzione di una traiettoria di movimento a memoria può coinvolgere un gruppo specifico di cellule, oltre a quelle attivate sia durante i movimenti guidati visivamente sia in quelli memorizzati. Questa ipotesi è in accordo con i risultati di alcuni studi di modellistica che verranno discussi più avanti (Lukashin et al., 1994).
Elaborazione dell'informazione direzionale durante una rotazione mentale
Il nostro gruppo di ricerca ha condotto vari esperimenti basati su compiti che richiedevano la trasformazione della direzione di un movimento programmato. In questi studi, sono state dapprima effettuate alcune osservazioni di natura psicologica su soggetti umani. Le scimmie sono state addestrate a svolgere lo stesso compito ed è stata registrata l'attività delle singole cellule neuronali di questi animali durante lo svolgimento del compito. Infine, si è cercato di mettere in relazione i risultati neuronali con quelli degli studi sui soggetti umani, e di interpretare i secondi sulla base dei primi: lo scopo era di spingersi il più avanti possibile nel mettere in relazione la neurofisiologia e la psicologia cognitiva.
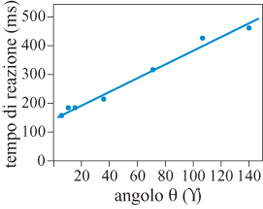
Il compito richiedeva ai soggetti di muovere una maniglia di un certo angolo rispetto a una direzione di riferimento, definita da uno stimolo visivo su un piano. Poiché la direzione di riferimento cambiava da prova a prova, il compito richiedeva che, in un dato esperimento, la direzione del movimento fosse specificata secondo quella direzione di riferimento. Negli studi di tipo psicologico (Georgopoulos e Massey, 1987), i soggetti umani effettuavano blocchi di 20 prove, in cui l'angolo suddetto e il suo verso (orario o antiorario) erano fissati, ma la direzione di riferimento variava; si sono usati sette angoli (5°÷140°). Il risultato principale è stato che il tempo necessario per iniziare il movimento (il tempo di reazione) cresceva in modo lineare con l'angolo (fig. 22).
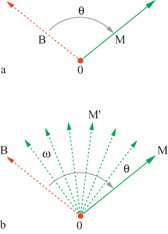
L'ipotesi più semplice per spiegare questi risultati è che i so ggetti arrivano alla determinazione della direzione giusta del movimento spostando la loro intenzione motoria dalla direzione di riferimento a quella del movimento, passando per gli angoli intermedi (fig. 23).
Questa idea è molto simile all'ipotesi della rotazione mentale avanzata da R.N. Shepard e collaboratori per spiegare l'aumento monotono del tempo di reazione con l'angolo di orientazione, quando si richiedeva di giudicare se un'immagine visiva ruotata fosse normale o speculare: in entrambi i casi si postula una rotazione mentale.
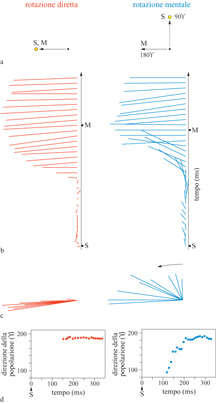
Negli studi neurofisiologici (Georgopoulos et al., 1989b; Lurito et al., 1991), si sono addestrate due macachi a muovere la maniglia di 90° in senso antiorario, rispetto alla direzione di riferimento; queste prove erano intervallate da altre in cui gli animali muovevano la manopola in direzione del bersaglio. Quando si calcolava il vettore neuronale di popolazione, variabile nel tempo, durante il tempo di reazione, si trovava che esso ruotava dallo stimolo (di riferimento) alla direzione di movimento in senso antiorario nel caso in cui l'animale doveva allontanarsi dallo stimolo, oppure ruotava in direzione dello stimolo, quando l'animale doveva muoversi verso di esso, come illustrato in figura (fig. 24).
È notevole il fatto stesso che il vettore di popolazione ruotasse, e anche che questa rotazione si verificasse verso l'angolo minore, in senso antiorario. Questi risultati hanno mostrato in modo chiaro che il processo cognitivo attivo in questo compito coinvolge realmente la rotazione di un segnale analogico. Il fatto che si abbia una vera rotazione è stato documentato ulteriormente mostrando che si verificava una crescita transiente verso la metà del tempo di reazione, nel reclutamento di cellule con direzioni preferite a metà strada tra lo stimolo e le direzioni di movimento (fig. 25): questo ha indicato che la rotazione del vettore di popolazione non era il risultato dell'attivazione variabile nel tempo di due soli gruppi di cellule, una con le direzioni preferite centrate sullo stimolo, e l'altra in direzione del movimento.
Tale processo di rotazione, quindi, passando attraverso l'insieme delle cellule selettive alla direzione, ha fornito per la prima volta una visualizzazione diretta di un processo cognitivo dinamico. Da questo punto di vista, è interessante notare come il vettore di popolazione sia, essenzialmente, una grandezza che può assumere valori continui di direzione. Riassumendo, i risultati di questi studi forniscono i correlati neuronali di una rappresentazione cognitiva dinamica. Il contributo fondamentale di questo lavoro sta nell'identificazione neuronale, e nella visualizzazione, della rappresentazione dinamica, variabile nel tempo, della direzione nella corteccia motoria, quando viene richiesta, e ottenuta, una trasformazione di questa direzione.
È interessante notare come la velocità di rotazione media e l'intervallo di velocità, osservati per diverse direzioni di riferimento, erano molto simili a quelli osservati negli studi condotti su soggetti umam.
Velocità di elaborazione dell'informazione motoria nella rotazione mentale. - L'ipotesi di una rotazione mentale nei compiti descritti sopra è stata confermata dai risultati di un'analisi dei dati, basata sull'ipotesi che il processo presunto di rotazione mentale coinvolga, in effetti, qualche tipo di movimento di un vettore di movimento immaginario. Assumendo questo modello di movimento interno, abbiamo analizzato le relazioni tra l'ampiezza e l'accuratezza applicando il metodo di Fitts ai movimenti reali. In particolare, eravamo interessati a scoprire se la legge di Fitts, valida per i movimenti reali, lo fosse anche nel nostro caso. Questa legge, come riportato precedentemente nell'equazione [8], collega il tempo del movimento, MT, a una misura informazionale, Id.
Quando l'accuratezza del movimento non è vincolata a priori, si può usare una misura della dispersione dei punti terminali del movimento come stima di W nell' equazione [7]. In questi studi Id è stato definito assumendo che A abbia come valore l'angolo effettivo ottenuto, e che W=2so, dove So è la deviazione standard circolare (Georgopoulos e Massey, 1987). In questo caso, si chiedeva ai soggetti di compiere un movimento di un certo angolo in senso orario a partire dalla direzione di uno stimolo. Id veniva calcolato dai dati mediati su 9 soggetti (Georgopoulos e Massey, 1987); si è trovato che cresceva come una funzione lineare dell'angolo prefissato, nel modo seguente:
Id = 0,46 + 0, 015 θ (r² = 0,98) [12]
in cui e è l'angolo prefissato (v. sopra) e r² è il coefficiente di espressione (r²= l se i punti sperimentali sono allineati esattamente sulla retta di regressione). Infine, l'aumento del tempo di rotazione (R T) era una funzione lineare di h secondo l'equazione seguente:
RTθ - RTo = 86 + 154 Id (r² = 0,99). [13]
In accordo con la legge di Fitts, quindi, abbiamo trovato che l'aumento di RT, considerato come un tempo di movimento mentale, era una funzione lineare della difficoltà del compito, calcolata dall'angolo ottenuto e dalla sua variabilità; tale legge è valida per il movimento rotatorio ipotizzato per l'immaginario vettore di movimento, e sia i movimenti reali sia quelli immaginari potrebbero essere governati da relazioni simili tra l'ampiezza e l'accuratezza. In effetti, quest'idea ha stimolato la serie seguente di esperimenti, che sfruttavano i risultati più recenti riguardanti la codifica neuronale della direzione di un movimento effettivo, per estenderli al caso di un movimento mentale, in particolare una rotazione mentale. Il valore medio dell'indice di prestazione (Ip) per questo compito è stato di 4,21 bit/s, che è quasi la metà della velocità di elaborazione dell'informazione per i movimenti rettilinei (Ip =7,25 bit/s). Questo è dovuto, probabilmente, al peso cognitivo di tale compito, insieme al fatto che la rotazione implica di lavorare sulla direzione, e ciò costituisce un processo lento.
Riguardo ai meccanismi neuronali sottesi a tale rotazione mentale da parte di un vettore immaginario di movimento, effettuato il seguente calcolo, basato sul fatto che, durante il tempo di reazione, il vettore di popolazione neuronale ruotava in senso antiorario dalla direzione dello stimolo, verso la direzione del movimento (v. sopra). Erano disponibili otto direzioni per lo stimolo, e quindi otto serie temporali di vettori di popolazione ruotanti durante RT (Lurito et al., 1991). Da questi dati, il valore di Ip è risultato di 4,785 bit/s. Questa è la stima della velocità di elaborazione dell'informazione direzionale da parte dell'insieme neuronale durante la rotazione mentale. Il valore di Ip per le stesse scimmie è stato calcolato nel modo seguente. L'angolo di rotazione richiesto era di 90°, con una tolleranza di ±30°, e quindi la larghezza del bersaglio angolare era di 60°; il valore medio per RT era 413,7 ms. Da questi dati, risulta Ip = 3,832 bit/s. Occorre sottolineare, in primo luogo, che questo valore comportamentale di Ip per le scimmie è vicino al valore di 4,21 bit/s calcolato per i dati sui soggetti umani di cui sopra. In secondo luogo, il valore neuronale per Ip di 4,785 bit/s era di circa l bit più alto di quello comportamentale di 3,832 bit/s, calcolato nelle stesse condizioni e negli stessi animali. Questo fatto supporta ulteriormente l'analisi, e suggerisce che ci sia un po' di perdita di informazione dall' elaborazione corticale motoria al comportamento, come ipotizzato precedentemente (Georgopoulos e Massey, 1988).
Elaborazione dell'informazione direzionale durante l'esplorazione della memoria in compiti di richiamo del contesto

Negli studi descritti, sono stati identificati i correlati neuronali di un processo di rotazione mentale, come la rotazione graduale del vettore di popolazione neuronale a partire da uno stimolo, verso la direzione di un movimento, attraverso direzioni con un angolo specifico. Questa rotazione esemplificava la regola spaziale operante nel compito di rotazione mentale, che richiedeva la produzione di un movimento di un certo angolo rispetto alla direzione di uno stimolo. In un altro studio (Pellizzer et al., 1995) si è cercato invece di determinare i correlati neuronali di un processo cognitivo la cui regola era basata non su un vincolo spaziale, ma sulla posizione seriale degli stimoli in una sequenza: data una sequenza arbitraria di stimoli su un cerchio, uno dei quali sia stato identificato come stimolo di test, la risposta motoria doveva essere verso lo stimolo che seguiva lo stimolo di test nella sequenza. Questo compito è una versione visuomotoria del compito di esplorazione della memoria 'richiamo del contesto' sviluppato da S. Sternberg. Per determinare il meccanismo neuronale in questo compito è stata registrata l'attività di cellule singole nella corteccia motoria di una scimmia addestrata a effettuare un compito di richiamo del contesto e uno di controllo, di ritardo istruito (fig. 26). Sono stati inoltre analizzati di nuovo i dati dello studio sulla rotazione mentale (Lurito et al., 1991), per confrontarli con quelli ottenuti con questo nuovo compito. Nel compito di controllo, veniva presentato uno stimolo giallo in una tra otto direzioni, e questo rimaneva per 400 ms, dopo di che diventava blu.
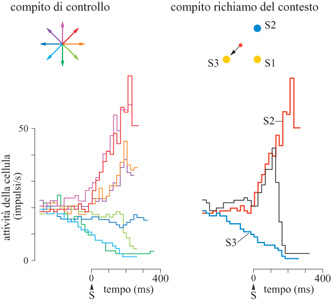
Questo forniva il segnale di via affrnché la scimmia esercitasse brevemente una forza, tale che un indicatore di forza di retro azione superasse una certa soglia. Nel compito di richiamo del contesto (v. figura 26), venivano presentati in successione, ogni 400 ms, tre stimoli gialli, cioè quelli della lista, in posizioni diverse sul cerchio, i quali rimanevano sullo schermo. Dopo ulteriori 400 ms uno di questi stimoli, eccetto l'ultimo, diventava blu. Questo identificava lo stimolo di test, e forniva il segnale di via: ora, la scimmia doveva muovere il cursore nella direzione dello stimolo che seguiva lo stimolo di test nella sequenza. Abbiamo trovato che, durante il tempo di risposta, gli schemi di attività neuronale nella corteccia motoria ricordavano inizialmente quelli associati alla direzione del secondo stimolo. Quando lo stimolo di test era il primo della sequenza, l'attività della cellula continuava a riflettere la direzione del secondo stimolo che, in questo caso, costituiva la risposta motoria corretta. Quando però lo stimolo di test era il secondo della sequenza, l'attività neuronale commutava improvvisamente, in modo da riflettere lo schema di attività associato con la direzione del terzo stimolo, che rappresentava adesso la risposta motoria appropriata. Questo cambiamento era davvero improvviso, si verificava circa 100÷150 ms dopo il segnale di via, e risultava evidente sia nell' attività di singole cellule (fig. 27) sia nel vettore di popolazione, variabile nel tempo, che cambiava direzione entro 50÷60 ms.
Questi risultati identificano i correlati neuronali di un processo di commutazione che è diverso dalla rotazione mentale (Lurito et al., 1991) descritta sopra. Una dimostrazione ulteriore della natura fondamentalmente diversa dei due processi neuronali è stata fornita da un'analisi della selettività direzionale di cellule che cambiavano attività durante il tempo di risposta. L'idea è che, in un processo di rotazione, l'insieme delle cellule che cambiano attività durante il tempo di risposta dovrebbe includere cellule la cui direzione preferita sia in una direzione intermedia tra lo stimolo e la direzione di risposta, come è stato osservato effettivamente (Lurito et al., 1991). D'altra parte, un'operazione di commutazione, come quella ipotizzata per il compito di richiamo del contesto, non dovrebbe coinvolgere l'attivazione di cellule in direzioni intermedie tra lo stimolo di test (S2) e la risposta motoria (S3). Anche questo è stato osservato (Pellizzer et al., 1995). Sembra che il tempo necessario per derivare la direzione motoria nel compito di rotazione mentale rifletta una trasformazione, mentre il tempo necessario per il compito di richiamo del contesto rifletta un processo di selezione. Infine, occorre sottolineare che questi studi hanno gettato luce sui meccanismi neuronali che sono alla base di questi processi in una particolare area cerebrale, la corteccia motoria, ma è ovvio che altre aree cerebrali sono verosimilmente coinvolte; sono necessari altri esperimenti per identificarle e chiarire i relativi contributi di ciascuna allo svolgimento del compito.
Aspetti temporali dell'elaborazione dell'informazione direzionale
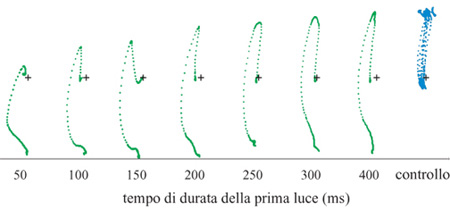
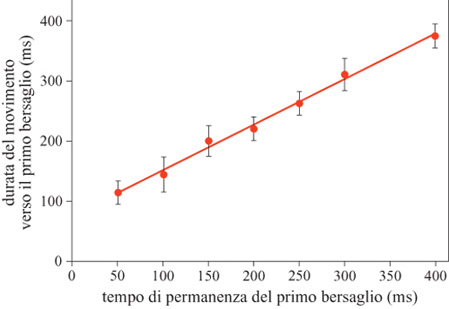
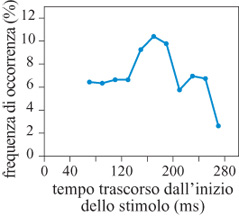
Gli studi fin qui illustrati trattano la codifica neuronale dell'informazione direzionale e la sua elaborazione in diversi compiti. Essi si basano su compiti coinvolgenti la produzione di movimenti di raggiungimento verso bersagli visivi stazionari. Occorre però chiedersi come si influenzi il movimento, se i suoi bersagli cambiano in modo non predicibile durante il tempo di reazione, o quello di movimento, e se il processo che genera il movimento desiderato è del tipo 'tutto o niente' o è modificabile. Questo costituisce un quesito importante: spesso in natura l'oggetto che suscita interesse da parte della scimmia, verso il quale cioè essa dirige la mano, cambia posizione; quindi è interessante sapere se il sistema motorio può seguire il bersaglio e cambiare il movimento a mezza strada, o se esiste un ritardo apprezzabile tra schemi motori commutabili. Lo studio di tali problemi è stato condotto attraverso il seguente esperimento (Georgopoulos et al., 1981). Sono state usate due luci periferiche in posizioni opposte (per esempio alle ore 6 e alle ore 12). La scimmia reggeva la maniglia nel centro per un periodo di tempo variabile, dopo il quale uno dei due bersagli periferici veniva acceso; rimaneva acceso per 50, 100, 150, 200, 250, 300 o 400 ms dopodiché veniva spento, e si accendeva l'altro bersaglio. L'animale otteneva una ricompensa dopo esser stato sulla seconda luce per 0,5 s. Esperimenti in cui la prima luce rimaneva accesa, senza cambiare posizione, erano in1Tammezzati ad altri in cui la posizione del bersaglio cambiava, cosicché l'animale non poteva predire se il bersaglio avrebbe cambiato posizione in una data prova; inoltre l'animale non poteva predire per quanto tempo il primo bersaglio sarebbe rimasto acceso, nelle prove in cui esso cambiava posizione. Abbiamo trovato sistematicamente che il braccio si muoveva inizialmente verso il primo bersaglio, quindi cambiava direzione e si muoveva verso il secondo (fig. 28). Quantitativamente, la durata del movimento del braccio verso il primo bersaglio era una funzione lineare del tempo per il quale il primo bersaglio rimaneva acceso (fig. 29), indicando uno stretto accoppiamento temporale tra lo stimolo visivo e il movimento del braccio controllato da esso. Un problema diverso riguarda le caratteristiche spaziali della traiettoria del primo movimento. Nella maggior parte dei casi studiati, le due luci bersaglio erano collocate lungo la stessa linea, cosicché il primo e il secondo movimento avevano direzioni opposte. Quando, però, le due luci erano sistemate in modo tale che i movimenti corrispondenti avessero direzioni opposte, è stata osservata in alcuni esperimenti una modificazione della direzione della traiettoria del primo movimento, per cui questa deviava verso la direzione del secondo bersaglio, tanto più quanto più breve era l'intervallo tra gli stimoli. Questi risultati indicano che il comando motorio per la direzione del movimento generato, dopo la comparsa del primo bersaglio, può essere modificato dal secondo bersaglio, quando i bersagli cambiano rapidamente. Infine, è stato interessante esaminare le caratteristiche di velocità delle traiettorie del movimento verso il secondo bersaglio, dopo il loro scambio. In effetti, questi movimenti erano molto più veloci di quelli compiuti verso il primo bersaglio; la velocità di picco dei movimenti verso il secondo bersaglio, approssimativamente, era 3 volte superiore a quello dei movimenti verso il primo bersaglio. Questo si verificava anche quando gli intervalli tra gli stimoli erano di 50 ms: i movimenti verso la prima luce erano piccoli e, quindi, l'ampiezza del secondo movimento era molto simile a quella del movimento fatto direttamente verso il secondo bersaglio negli esperimenti di controllo. Questi risultati si possono spiegare con il tipo di strategia che le scimmie, e i soggetti umani, usavano per risolvere i problemi posti da questo compito di spostamento del bersaglio. Questa strategia è stata messa in luce da un'analisi delle forze esercitate sulla leva durante lo svolgimento del compito (Massey et al., 1986). In breve, i soggetti producevano una grande forza frenante in risposta al secondo bersaglio, che fermava il movimento della mano verso il primo bersaglio, e si muovevano poi verso il secondo bersaglio. Tale forza era eccessiva per i movimenti brevi verso il primo bersaglio, ma era efficace in tutti i casi, e non richiedeva correzioni individuali per esperimenti particolari. Dunque, una strategia semplice e meccanicamente efficace, ma energeticamente inefficiente, sta alla base della coordinazione efficiente tra l'occhio e la mano in questo compito.
Meccanismi neuronali dell'accoppiamento visuomotorio

Quando la posizione degli stimoli visivi cambia in rapida successione, gli schemi di attività cellulare nell'area della corteccia motoria corrispondente al braccio seguono questi cambiamenti con notevole fedeltà temporale (Georgopoulos et al., 1983b), testimoniando l'efficace coinvolgimento di questa struttura durante la coordinazione tra l'occhio e la mano. In maniera specifica, lo schema di attività inizialmente corrispondeva a quello associato al movimento verso il primo bersaglio; questo schema durava per un tempo proporzionale al tempo per cui il primo bersaglio rimaneva acceso, e diveniva poi simile allo schema di attività associato al movimento verso il secondo bersaglio (fig. 30). Le relazioni temporali tra la durata della presentazione del primo bersaglio, la durata del cambiamento dell'attività della cellula associata alla presentazione del primo bersaglio, e la durata del movimento della mano verso il bersaglio sono lineari (Georgopoulos et al., 1983b). Questi risultati indicano che quando viene generata una sequenza di movimenti della mano verso bersagli che cambiano posizione, l'attività corticale motoria segue bene i cambiamenti nell'informazione visiva, e genera i comandi motori appropriati senza dispersione temporale. Questo sta alla base, a quanto pare, della coordinazione efficiente tra l'occhio e la mano, osservata a livello comportamentale.
Elaborazione dell'informazione direzionale attraverso il midollo spinale
Sistemi interneuronali spinali
In generale, i segnali neuronali dalla corteccia motoria non attivano direttamente i muscoli, ma influenzano l'attività dei motoneuroni attraverso il midollo spinale. A volte, gli assoni corticospinali terminano all'interno dei nuclei motori, come avviene nei motoneuroni che innervano i muscoli distali degli arti dei primati adulti, ma nella grande maggioranza dei casi l'influenza sugli insiemi di motoneuroni viene esercitata attraverso l'interposizione di interneuroni, presenti sia a livello segmentale sia a livello cervicale superiore (C3-C4). Un'analisi dettagliata del modo in cui i sistemi spinali di intemeuroni possano influenzare l'uscita spinale motoria è stata condotta di recente nel midollo spinale della rana da E. Bizzi e collaboratori (v. il saggio di E. Bizzi e F. Mussa-lvaldi, Apprendimento e controllo degli atti motori). In questi studi è stata utilizzata la tecnica della microstimolazione per determinare gli effetti delle popolazioni di interneuroni sulla direzione e sull'entità della forza sviluppata dall'arto della rana, ottenendo tre risultati principali. In primo luogo, la microstimolazione di una particolare zona interneuronale nella spina dorsale ha provocato lo sviluppo di una forza da parte dell'arto, di direzione ed entità diverse a seconda della posizione dell'arto nello spazio; l'insieme di queste forze defmisce un campo di forze. In assenza di microstimolazione, vi erano delle forze passive dovute alla resistenza allo stiramento dei tessuti dell'arto; durante la microstimolazione, venivano sviluppate delle forze attive che, aggiunte a quelle passive, davano come risultante la forza esercitata dall'arto. Nell'esperimento venivano misurate la forza passiva e quella totale, e la forza passiva veniva sottratta da quella totale per ottenere il campo di forza attiva. È stato evidenziato che la microstimolazione in una data area produceva un campo di forza attiva in cui i vettori di forza convergevano in un punto particolare dello spazio, nel quale la forza sviluppata era nulla; tale punto è stato chiamato il punto di equilibrio di quel campo. In molti casi, il campo di forza prediceva il movimento dell'arto, quando a questo era consentito di muoversi; questi campi si possono quindi considerare come sotto stanti a delle primitive di movimento. In secondo luogo, attraverso una microstimolazione dettagliata e sistematica di ampie aree del midollo spinale, sono stati trovati solo 3÷4 gruppi di punti di equilibrio, ciascuno dei quali era rappresentato in posizioni diverse nel midollo spinale. Infine, Mussa-lvaldi e collaboratori hanno trovato che la microstimolazione concomitante in due punti qualsiasi produceva un nuovo campo di forza che era il risultato di una somma lineare punto a punto dei campi di forze prodotti in ognuno dei due punti di stimolazione; venivano dunque creati nuovi punti di equilibrio.
Questi risultati suggeriscono i possibili meccanismi attraverso i quali i comandi sovraspinali possono essere tradotti in azioni motorie: in primo luogo, poiché le microstimolazioni attiveranno un numero apprezzabile di neuroni, questi meccanismi intrinseci spinali motori coinvolgono popolazioni di neuroni e la somma di influenze da insiemi neuronali potenzialmente separati. Secondariamente, è probabile che questi insiemi siano destinatari di influenze sovraspinali. Per esempio, una caratteristica comune degli assoni del sistema piramidale è la loro apprezzabile divergenza nel midollo spinale con collaterali verso vari segmenti spinali. Questa divergenza abiliterebbe dei comandi motori centrali a indirizzarsi in modo concomitante verso diverse popolazioni spinali associate a campi di forza differenti, cioè con primitive di movimento differenti. lnfine, questa organizzazione potrebbe fornire il contesto per la traduzione di un comando corticale motorio, da come esso viene compreso a livello della singola cellula e della singola popolazione neuronale all'attivazione motoneuronale (Georgopoulos, 1996).
Modelli di reti neuronali delle operazioni direzionali
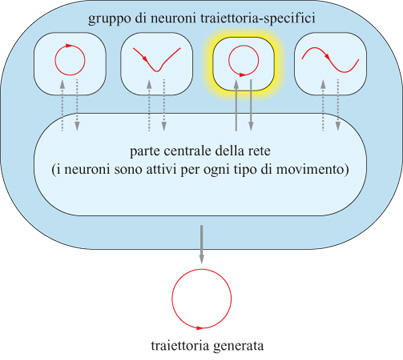
La corteccia motoria si può considerare essenzialmente come una rete di neuroni. Gli elementi della rete sono selettivi rispetto alle direzioni, e una delle possibili modalità di funzionamento della rete è il calcolo del vettore neuronaIe di popolazione. L'ampia curva di selettività direzionale della singola cellula sembra essere un effetto generale del funzionamento a livello di popolazione, considerato che A.V. Lukashin ha trovato che questa proprietà appariva nello strato nascosto di una rete a tre strati addestrata a calcolare il vettore di popolazione. Inoltre, un importante problema riguarda le interazioni tra le cellule, poiché è noto che vi sono molte interconnessioni locali. Il nostro scopo è stato quello di identificare queste interazioni tra le cellule della corteccia motoria, selettive rispetto alla direzione, e scoprire le regole che determinano la loro presenza; di studiare una rete neurale artificiale composta di elementi selettivi rispetto alla direzione, con interconnessioni estese; di confrontare i risultati ottenuti per la rete reale, cioè la corteccia motoria, e per quella artificiale. l risultati ottenuti sono i seguenti (Georgopoulos et al., 1993). In primo luogo, le interazioni tra le coppie di cellule della corteccia motoria sono in relazione con la selettività rispetto alla direzione delle cellule di ogni coppia: le interazioni sono risultate frequenti più del doppio quando le cellule erano selettive, rispetto al caso in cui non lo erano e, per le cellule selettive rispetto alla direzione, questa andava dall'essere fortemente positiva (cioè eccitatoria) a fortemente negativa (cioè inibitoria), via via che l'angolo tra le direzioni preferite delle cellule in una coppia variava tra 0° (cioè stessa direzione preferita) e 180° (cioè direzioni preferite opposte). In secondo luogo, la stessa tendenza si è osservata tra gli elementi selettivi rispetto alla direzione, in una rete dinamica artificiale, fortemente interconnessa, durante un periodo stabile di calcolo del vettore neuronale di popolazione. In terzo luogo, si è trovato che affinché la rete sia stabile, il peso delle interazioni sinaptiche doveva essere basso: nel caso migliore (cioè il più stabile) il peso sinaptico medio tende a 2/N, dove N indica il numero di elementi della rete. Questo è coerente con il fatto che le cellule corticali in un'area sono estensivamente, ma debolmente, interconnesse. I risultati ottenuti confermano la corrispondenza tra la rete neuronale corticale motoria e quella artificiale, e aprono la possibilità di usare questa rete nel dominio del tempo, per esplorare i meccanismi del funzionamento cognitivo descritti in precedenza. A tal fine, recentemente, sono stati creati dei modelli utilizzando una rete neurale artificiale estesamente interconnessa, composta di neuroni selettivi rispetto alla direzione, che produce come output il vettore neuronale di popolazione (Lukashin et al., 1994). Questa rete ha ben riprodotto molti dei risultati sperimentali, e ha condotto a una nuova ipotesi sul modo in cui potrebbero essere immagazzinate le traiettorie memorizzate di movimenti complessi nelle connessioni sinaptiche di reti neuronali che si sovrappongono (Lukashin et al., 1994). È stata ipotizzata l'esistenza di una rete di tipo generale coinvolta in tutti i tipi di movimenti, memorizzati o meno, che però non porti informazione circa le traiettorie memorizzate di forme specifiche (per esempio, cerchi, ellissi, scarabocchi, ecc.), e che, se attivata da sola, produrrebbe traiettorie rettilinee. Attualmente, si suppone che vi siano anche delle reti altamente specializzate per una particolare traiettoria (per esempio, un cerchio in senso orario) che sono interconnesse con la rete di tipo generale: quando bisogna effettuare una traiettoria specifica, la rete specifica appropriata si fonde con quella di tipo generale e, come singola rete, produce la traiettoria desiderata (fig. 31).
È notevole il fatto che, affrnché la traiettoria desiderata sia efficacemente immagazzinata e riprodotta, è sufficiente che la rete specializzata sia grande meno del 5% della rete di tipo generale (Lukashin et al., 1994). Vale la pena notare che cellule molto specifiche di questo tipo si sono osservate, in piccole proporzioni, nelle registrazioni neurofisiologiche durante la realizzazione di traiettorie memorizzate (Ashe et al., 1993).
L'ipotesi dell' esistenza di reti molto specializzate solleva il problema del grado di specializzazione, e del modo in cui tali reti vengano create inizialmente. Su questo punto possiamo solo avanzare delle ipotesi. Riguardo alla rete di tipo generale, è ragionevole assumere che essa sia presente alla nascita, poiché si ipotizza che essa sottenda tutti i movimenti. Per le reti specializzate vi sono diverse possibilità. Una è che un certo numero di reti di piccole dimensioni, specifiche per alcune forme fondamentali (per esempio, linee rette, curve e alcune loro combinazioni - primitive motorie di forma), siano presenti alla nascita. L'apprendimento di altri atti motori complessi consisterebbe allora nella calibrazione dei pesi delle connessioni tra la rete di tipo generale e le reti specializzate. Quest'ipotesi implica che tutte le primitive specializzate vengano usate regolarmente, sebbene in modo non così frequente come la rete di tipo generale. Un'altra possibilità è che le reti specifiche innate codifichino forme più complicate, e siano molte. Il meccanismo dell'apprendimento motorio sarebbe quindi simile a quello descritto sopra, ma in questo caso sarebbe usata solo una piccola frazione delle reti specializzate. Questo vuoI dire che molte delle complesse reti specializzate potrebbero non essere mai usate. Questa situazione sarebbe simile a quella incontrata nel sistema immunitario, in cui c'è una grande potenzialità per la costruzione di molti anticorpi dei quali, però, solo alcuni vengono realmente costruiti, a seconda dell' esposizione da parte dell' organismo ad antigeni specifici. In entrambi i casi vi è una selezione: quella di una traiettoria specializzata o quella di un anticorpo, entrambe a partire da un grande insieme disponibile. Infine, un'ipotesi intermedia consisterebbe nel cominciare con primitive motorie di forma, mentre le traiettorie più complesse risultanti dalla combinazione di queste primitive con la rete generica diventerebbero esse stesse molto specializzate, e si comporterebbero come tali nella formazione di altre traiettorie in nuove associazioni.
Ringraziamenti
Questo lavoro è stato finanziato dall 'United Public Health Service, dal Department ofVeterans e dall'American Legion Chair in Brain Sciences.
Bibliografia citata
ASHE, J., TAIRA, M., SMYRNIS, N., PELLIZZER, G., GEORGAKOPOULOS, T., LURITO, J.T., GEORGOPOULOS, A.P. (1993) Motor cortical activity preceding a memorized movement trajectory with an orthogonal bend. Exp. Brain Res., 95, 118-130.
CAMINITI, R., JOHNSON, P.B., GALLI, C., FERRAINA, S., BURNOD, Y., URBANO, A. (1991) Making arm movements within different parts of space: the premotor and motor cortical representation of a coordinate system for reaching at visual targets. J. Neurosci., Il, 1182-1197.
FITTS, P.M. (1954) The information capacity of the human motor system in controlling the amplitude of movement. J. Exp. Psychol., 47, 381-391.
GEORGOPOULOS, A.P. (1996) On the translation of directional motor cortical commands to activation of muscles via spinal intemeuronal systems. Cogn. Brain Res., 3, 151-155.
GEORGOPOULOS, A.P., ASHE, J., SMYRNIS, N., TAIRA, M. (1992) Motor cortex and the coding offorce. Science, 256,1692-1695.
GEORGOPOULOS, A.P., CAMINITI, R., KALASKA, J.F., MASSEY, J.T. (1983a) Spatial coding of movement. A hypothesis concerning the coding of movement direction by motor cortical populations. Exp. Brain Res. Suppl., 7, 327-336.
GEORGOPOULOS, A.P., CRUTCHER, M.D., SCHWARTZ, A.B. (1989a) Cognitive spatial motor processes. 3. Motor cortical prediction of movement direction during an instructed delay period. Exp. Brain Res., 75, 183-194.
GEORGOPOULOS, A.P., KALASKA, J.F., CAMINITI, R , MASSEY, J.T. (1982) On the relations between the direction of two dimensional arm movements and celI discharge in primate motor cortex. J. Neurosci., 2, 1527-1537.
GEORGOPOULOS, A.P., KALASKA, J.F., CAMINITI, R., MASSEY, J.T. (1983b) Interruption of motor cortical discharge subserving aimed arm movements. Exp. Brain Res., 49, 327-340.
GEORGOPOULOS, A.P., KALASKA, J.F., CRUTCHER, M.D., CAMINITI, R., MASSEY, J.T. (1984) The representation of movement direction in the motor cortex: single celI and population studies. In Dynamic aspects of neocortical Junction, a c. di Edelman G.M., Cowan W.M., Gall W.E., New York, Wiley, pp. 501-524.
GEORGOPOULOS, A.P., KALASKA J.F., MASSEY, J.T. (1981) Spatial trajectories and reaction times of aimed movements: effects of practice, uncertainty and change in target location. J. NeurophysioI., 46, 725-743.
GEORGOPOULOS, A.P., KETTNER, R.E., SCHWARTZ, AB. (1988) Primate motor cortex and free arm movements to visual targets in three-dimensional space. II. Coding of the direction of movement by a neuronal population. J. Neurosci., 8, 2928-2937.
GEORGOPOULOS, A.P., LURITO, J.T., PETRIDES, M., SCHWARTZ, A.B., MASSEY, J.T. (1989b) Mental rotation of the neuronal population vector. Science, 243, 234-236.
GEORGOPOULOS, A.P., MASSEY, J.T. (1987) Cognitive spatial-motor processes. 1. The making of movements at various angles from a stimulus direction. Exp. Brain Res., 65, 361-370.
GEORGOPOULOS, A.P., MASSEY, J.T. (1988) Cognitive spatial-motor processes. 2. Information transmitted by the direction of two-dimensional arm movements and by neuronal populations in primate motor cortex and area 5. Exp. Brain Res., 69, 315-326.
GEORGOPOULOS, A.P., SCHWARTZ, AB., KETTNER, R.E. (1986) Neuronal population coding of movement direction. Science, 233, 1416-1419.
GEORGOPOULOS, A.P., TAIRA, M., LUKASHIN, A.V. (1993) Cognitive neurophysiology ofthe motor cortex. Science, 260, 47-52.
LUKASHIN, A.V., WILCOX, G.L., GEORGOPOULOS, A P. (1994) Overlapping neural networks for multiple motor engrams. Proc. NatI. Acad. Sci. USA, 91, 8651-8654.
LURITO, J.L., GEORGAKOPOULOS, T., GEORGOPOULOS, A.P. (1991) Cognitive spatial-motor processes. 7. The making of movements at an angle from a stimulus direction: studies of motor cortical activity at the single celI and population levels. Exp. Brain Res., 87, 562-580.
MASSEY, J.T., SCHWARTZ, A.B., GEORGOPOULOS, A.P. (1986) On information processing and performing a movement sequence. Exp. Brain Res. Suppl., 15, 242-251.
PELLIZZER, G., SARGENT, P., GEORGOPOULOS, A.P. (1995) Motor cortical activity in a context-recall task. Science, 269, 702-705.
SALINAS, E., ABBOTT, L.F. (1994) Vector reconstruction from firing rates. J. Comput. Neurosci., 1, 89-107.
SANES, J.N. (1985) Information processing deficits in Parkinson's disease during movement. Neuropsychologia, 23, 381-392.
SCHWARTZ, A.B., KETTNER, R.E., GEORGOPOULOS, A.P. (1988) Primate motor cortex and free arm movements to visual targets in three-dimensional space. I. Relations between single celI discharge and direction of movement. J. Neurosci., 8, 291-327.
SMYRNIS, N., TAIRA, M., ASHE, J., GEORGOPOULOS, A.P. (1992) Motor cortical activity in a memorized delay task. Exp. Brain Res., 92, 139-151.
Bibliografia generale
BEAR, M.F., CONNORS, B.W., PARADISO, M.A. Neuroscience. Exploring the brain. Baltimora, Williams & Wilkins, 1996.
GAZZANIGA, M.S., a c. di, The cognitive neurosciences. Cambridge, Mass., MIT Press, 1995.
KANDEL, E.R., SCHWARTZ, J.H., JESSEL, T.M., a c. di, Principles of neural science., 3a ed., Norwalk, Appleton & Lange, 1991.
SHANNON, C.E., WEAVER, W. The mathematical theory of communication. Urbana, University of Illinois, 1949, p.3.
SHEPARD, R.N., COOPER, L.A. Mental images and their transformations. Cambridge, Mass., MIT Press, 1982.