
L'Io cosciente: una visione neuroeuristica della mente
L'Io cosciente: una visione neuroeuristica della mente
I più recenti avanzamenti della ricerca scientifica permettono di affrontare il problema della coscienza in modo interdisciplinare, utilizzando un paradigma che qui chiameremo neuroeuristica. In questo saggio presentiamo, in particolare, alcuni risultati di ricerche specifiche svolte nel campo delle neuroscienze, della psicofisiologia e dell'informatica. Tali ricerche permettono di elaborare un modello di mente relazionale, basato su principi fondamentali controlla bili sperimentalmente. A partire da tale modello vengono illustrati i possibili meccanismi neuronali associati alla cosiddetta sensazione cosciente, altrimenti detta coscienza passiva . Questi meccanismi sono alla base di processi cognitivi, quali l'introspezione e l'intenzionalità, che formano la cosiddetta coscienza attiva.
Introduzione
Attualmente una considerevole attività di ricerca tenta di comprendere la natura della coscienza (Nagel, 1974). Molte discipline sono interessate a questo studio e possono fornire un valido contributo: filosofia, psicologia, neuroscienze, farmacologia, fisica, intelligenza artificiale, ingegneria, informatica e matematica. Questa interdisciplinarità rende il problema più interessante e tuttavia più complicato, perché si deve tener conto del diverso linguaggio e delle conoscenze di base che ciascuna disciplina possiede. Un'opportuna interdisciplinarità appare quindi necessaria (Hameroff et al., 1996). Lo scopo di questo saggio è presentare un nuovo modo di affrontare lo studio della coscienza, basato su un paradigma interdisciplinare, la neuroeuristica. Cominceremo presentando i presupposti concettuali che conducono alla formulazione del nostro paradigma operativo. Lo studio della coscienza si sviluppa in termini di strutture di controllo e strategie di elaborazione; inoltre cerca le loro possibili identificazioni neurobiologiche in determinate aree cerebrali. Verranno presentate alcune prove, ottenute utilizzando la neurofisiologia e la psicologia, per chiarire il ruolo di queste strutture e delle loro funzioni e mostreremo che tali prove potrebbero essere applicabili anche a un programma di ricerca più vasto. Alla luce dei risultati raggiunti, saranno infine discusse direzioni future di ricerca.
Un paradigma dinamico per le neuroscienze
Le neuroscienze sono una delle principali aree della ricerca biomedica, data l'importanza che rivestono per la salute pubblica e il benessere generale dell'individuo. Studi statistici indicano che negli Stati Uniti e nell'Europa occidentale circa il 5% della popolazione totale è affetta da una malattia nervosa. Si stima che la spesa annuale pro capite sia di circa 500.000 dollari, dovuta in parte ai costi indiretti delle conseguenze socioeconomiche delle malattie nervose. Oltre il 90% degli individui affetti da tali malattie soffre di sintomi che debilitano le funzioni cognitive, in modi e forme diverse. l principi fondamentali delle scienze cognitive non possono essere concepiti indipendentemente dalle loro basi neurobiologiche, dalle loro relazioni con la rappresentazione mentale, e dalle teorie logiche e computazionali derivanti dallo studio del comportamento animale e umano (Segundo, 1985). Si è dimostrato, per esempio, che ratti esposti sin dalla nascita a un ambiente ricco di stimoli variabili tendono a sviluppare un ispessimento della corteccia cerebrale maggiore rispetto ai ratti allevati in un ambiente statico e immutabile. Tuttavia, quando la ricerca è ridotta alla mera correlazione tra attività mentale (determinata dalla ricchezza dell'ambiente circostante) e stato del cervello (spessore della corteccia), il metodo scientifico tradizionale delle scienze cognitive tende a produrre dei risultati troppo circoscritti per essere utilizzati in ambito generale.
Nella teoria dell'analisi causale sostenuta da K. Popper e J.C. Eccles l'informazione ascendente (bottom-up), che passa da livelli di complessità inferiori verso livelli di complessità superiori, si distingue da quella discendente (topdown), che procede in senso opposto. Applicando la bottom-up si potrebbe concludere che l'alterazione morfologica della corteccia cerebrale del ratto, precedentemente citata, sia causata da una successione di stimoli sconosciuti che in un animale naif (cioè, sottoposto per la prima volta a quella esperienza) provoca un comportamento esplorativo. Il punto di vista top-down, legato a una visione di tipo antropomorfico sosterrebbe, invece, che l'ispessimento della corteccia cerebrale sia causato da un'esperienza arricchita da eventi sorprendenti e interessanti.
Potrebbe sembrare legittimo sostenere che le caratteristiche proprie del livello superiore della mente, relative all'individuo, riflettano un particolare livello inferiore definito dai costituenti del sistema nervoso centrale. Tuttavia, l'osservazione microscopica del cervello di Albert Einstein, eseguita dopo la sua morte, non ha rivelato le informazioni attese dai sostenitori dell' analisi causale (Diamond et al., 1985). l ricercatori coinvolti in questo studio hanno osservato soltanto un aumento del numero di cellule gliali in alcune regioni dell'emisfero sinistro della corteccia. Pertanto, nel cervello di Einstein gli elementi computazionali, i neuroni, sono risultati di minor importanza rispetto agli elementi trofici, le cellule gliali. Allo stesso modo, gli anatomisti neopositivisti hanno avuto serie difficoltà a spiegare come mai nel cervello di Anatole France sia stata osservata una quantità inferiore di materia solida, giacché la sua massa risultava di 1017 g, di molto inferiore alla massa media (1300 ÷ 1400 g). È interessante osservare che la massa del cervello di questo scrittore è risultata simile a quella dell'Uomo di Giava (Pitecanthropus erectus). All'inizio del 20° secolo questa anomalia venne giustificata secondo la razionalità del paradigma neopositivista, con la seguente argomentazione del paleontologo A. Keith: "Anatole France traeva vantaggio da uno straordinario aumento di circonvoluzioni nel lobo frontale che compensava la mancanza di materia cerebrale".
Le strategie che si potrebbero usare per comprendere il funzionamento del cervello dipendono in maniera determinante dall'impostazione degli studi interdisciplinari relativi al comportamento molecolare, cellulare, individuale e sociale. La strategia di ricerca basata sul percorso dell'informazione del tipo bottom-up, preferita dai neurobiologi, sembra essere potenzialmente adeguata a questo scopo; tuttavia, non è ancora completamente applicabile, poiché è impossibile esaminare simultaneamente tutti gli elementi cellulari del cervello, persino nel caso di specie primitive. La strategia di tipo top-down, con l'assistenza di ipotetiche scatole chiuse, è più semplice da portare a termine, ma risulta insufficiente a spiegare i meccanismi che coordinano le reti locali. Ci è parso quindi necessario oltrepassare il confronto tra le due teorie, bottom-up e top-down, e proporre un nuovo approccio nel quale un risultato non è ridotto a essere semplicemente un risultato positivo o negativo, perché il procedimento stesso che ci consente di raggiungerlo diventa parte del risultato. Questo approccio pone l'accento sulle caratteristiche dinamiche e irriducibili di una nuova metodologia che progredisce per tappe successive allorché caratteristiche essenzialmente nuove e inaspettate si sviluppano a partire da proprietà preesistenti alla tappa precedente. Chiameremo questa metodologia neuroeuristica e cercheremo di dame una defrnizione esauriente.
La neuroeuristica, o neuristica (dal greco, νευpov = nervo e εὐpίσκειν = trovare, scoprire) è un ramo della scienza che ricerca la comprensione del funzionamento del cervello nella sua interezza tramite un processo di esplorazione dei principi fondamentali delle neuroscienze. Questo particolare processo progredisce continuamente grazie al rinnovamento che è messo in atto al conseguimento di ogni risultato parziale.
L'organizzazione del sistema nervoso è strutturata in modo tale da trattare le informazioni in una dimensione temporale: si pensi, per l'uomo, alla memoria e alla proiezione mentale di se stesso nel futuro. Le relazioni dinamiche tra memoria e attività sensomotoria avvengono essenzialmente nel corso del sonno. Durante l'attività onirica, le informazioni possono essere riorganizzate improvvisamente in una dimensione temporale diversa da quella reale, che è associata ai meccanismi classici stabiliti dagli studi effettuati durante lo stato di veglia.
La formulazione delle leggi delle neuroscienze, così come la costruzione di assiomi matematici, nel 19° e 20° secolo è stata basata su osservazioni e conoscenze postulate a priori. Pertanto l'osservazione è risultata spesso riduttiva e sterile, limitandosi esclusivamente alla descrizione. Quando una singola misurazione quantitativa non risulta sufficiente a determinare un asse di congruenza, si può ricordare la posizione una volta espressa da A. Einstein, secondo il quale le parole e il linguaggio, parlato o scritto, contrariamente all'opinione comune non giocavano un ruolo così secondario nella strutturazione dei suoi pensieri: le entità psichiche, elementi costitutivi del suo pensiero, erano immagini simboliche più o meno decifrabili, riproducibili o opportunamente combinabili. Questo processo evolve necessariamente dalla rottura del vincolo temporale, e risulta simile al metodo estetico del riconoscimento delle opere d'arte.
Un approccio interdisciplinare
L'analogia tra i livelli astratti di organizzazione nell' elaboratore e nel cervello va oltre l'osservazione di base che la programmazione all'elaboratore rappresenta un modello prescelto o un' imitazione artificiale dell' attività intellettiva umana. Nonostante i progressi ottenuti nella comprensione delle basi neurobiologiche della percezione, dell'apprendimento e della memoria, mediante la sperimentazione animale, le neuroscienze hanno contribuito poco alla conoscenza dei substrati biologici della creatività. Le scienze cognitive (Pylyshyn, 1986) hanno comunque affrontato il problema principale del cervello: separare la conoscenza dichiarativa (conoscere cosa) dalla conoscenza procedurale (conoscere come). A questo proposito sono innegabili i notevoli benefici che le scienze cognitive hanno ottenuto dallo scambio di informazioni con l'intelligenza artificiale, ma è nostra opinione che l'attività intellettuale non possa essere ridotta soltanto a un calcolo o a una semplice prestazione. Vogliamo qui sottolineare la sinergia tra l'informatica, intesa come scienza della comunicazione e delle procedure di analisi dell'informazione, e le neuroscienze; già in passato i primi scienziati, tra i pionieri delle neuroscienze, hanno valorizzato l'importanza di tale sinergia.
Negli anni tra il 1753 e il 1755 il fisiologo bernese Albrecht von Haller pubblicò un trattato, De partium corporis humani sensibilius et irritabilus. Questo lavoro era basato su numerosi esperimenti di vivisezione e di stimolazione di organi, eseguiti applicando le nuove conoscenze di fisiologia, fisica, chimica e storia naturale. Tramite una tecnica rudimentale di stimolazione elettrica, von Haller classificò le parti del corpo in irritabili, sensibili o elastiche, e osservò che le reazioni allo stimolo variavano nelle diverse aree del cervello. L'importanza storica del lavoro di von Haller non è propriamente dovuta ai risultati ottenuti, ma piuttosto al metodo di ricerca basato sull'integrazione sistematica delle nuove tecnologie del suo tempo. Anche la filosofia ha contribuito in modo interessante, in particolare chiarendo alcuni dei più importanti interrogativi relativi a mente e coscienza. In questo contesto una questione fondamentale è quella sollevata da T. Nagel relativa a "cosa sia X", dove X è un qualsiasi essere vivente diverso dal sé, che può potenzialmente sviluppare coscienza (Nagel, 1974). Secondo J. Searle (Searle, 1992) la scienza non può rispondere a tale domanda, e ha formulato l'utile distinzione tra domande 'facili' e 'difficili' riguardo la coscienza. Le domande facili si riferiscono ai meccanismi cerebrali alla base della coscienza, e sono state in gran parte trattate dai recenti sviluppi in psicologia, neuroscienze e farmacologia. Le scienze, come l'ingegneria, la fisica e la matematica stanno recentemente portando contributi essenziali allo studio teorico relativo alle diverse strategie di ricerca da usare. Studi cerebrali non invasivi di analisi di immagine, studi di lesioni ed esperimenti con elettrodi singoli e multipli, su animali e sull'uomo, stanno producendo risultati di fondamentale importanza anche per l'identificazione delle reti dei moduli neuronali preposti a un ampio spettro di compiti psicologici. l progressi sulla comprensione di queste connessioni cerebrali hanno aumentato l'ottimismo circa la possibilità di costruire un modello globale del cervello come sistema multimodulare. La strategia di ricerca consiste nel partire da simulazioni di neuroni e moduli semplici per poi gradualmente svilupparne la complessità. Tale compito richiede un approccio di tipo interdisciplinare, e il metodo neuroeuristico può essere un approccio in grado di dare risposte alle domande semplici sulla coscIenza.
La mente relazionale
Le strutture relazionali sono state da molto tempo riconosciute parti integranti del cervello e della mente. Duemila anni fa Aristotele sostenne che il pensiero procede secondo le relazioni fondamentali di contiguità, analogia e opposizione. Questa ipotesi venne largamente sviluppata dalle scuole associazioniste di psicologia nel 18° e 19° secolo. Tuttavia, già nel 17° secolo i filosofi empirici J. Locke, G. Berkeley e D. Hume studiarono in modo più approfondito il modo in cui le idee si susseguono. In particolare Hume dichiarò che "la mente non è altro che un accumulo o collezione di percezioni differenti, unificate da certe relazioni" (Hume, 1896). Nel seguito cercheremo di specificare tali relazioni, per poter giustificare la tesi di base del modello di mente relazionale secondo cui "il contenuto cosciente di un'esperienza mentale è determinato dall'evocazione e dalla miscela di memorie remote evocate, per lo più inconsciamente, dagli stimoli contingenti che sono alla base di quell'esperienza" (Taylor, 1991).
Generalmente si suppone che l'evocazione attivi le memorie passate che, a loro volta, possono assumere molteplici forme. A tale proposito si consideri quanto la memoria semantica sia drasticamente coinvolta per le parole. D'altra parte il processo preliminare della visione avviene solo attraverso le aree visive primarie del lobo occipitale. Pertanto, l'analisi nelle vie temporali (il cosa) e parietali (il dove) sarebbe ottenuta successivamente a partire dalla codificazione delle nozioni di forma, struttura, colore e moto che prendono origine nelle varie aree occipitali. Anche le memorie episodiche e autobiografiche (ovvero quelle in cui è presente l'lo) sono estremamente importanti, poiché vengono utilizzate per attribuire un aspetto più personale e sociale al contenuto cosciente. Sono altresì rilevanti gli aspetti affettivi che impartiscono una spinta emotiva a un contenuto cosciente, che altrimenti rimarrebbe puramente cognitivo. Essi originano da memorie immagazzinate nei circuiti del sistema limbico, più specificatamente nell'amigdala e nella corteccia frontale mediale. Il procedimento di miscelamento delle informazioni, necessario per risolvere una possibile ambiguità, richiede che alcune informazioni afferenti siano evidenziate più di altre. In effetti una parte consistente di parole e scene visive contiene elementi ambigui. La parola "collo", per esempio, ha almeno due significati ed è il contesto che di solito determina quale di essi debba essere scelto in una frase. Analogamente la visione del cubo di Necker presenta due possibili percezioni, con il lato inferiore verso l'osservatore oppure lontano da esso. Quale di essi venga scelto dipende dai movimenti e dall'adattamento oculare. Per risolvere l'ambiguità e definire le informazioni per un'elaborazione successiva viene adottato un processo naturale di competizione tra alternative. Il completamento di tale processo può richiedere anche molto tempo, ed eventualmente possono risultare necessari ulteriori dati per precisare il contesto e ridurre l'ambiguità. Si rende così indispensabile un contenitore, ovvero una memoria di lavoro (WM), che conservi una traccia dei precedenti dati, e consenta quindi una scelta. La competizione richiede la presenza di connessioni inibitorie tra i competitori, situate in regioni distanti della corteccia cerebrale oppure tra le aree corticali e sottocorticali. È naturale supporre che la competizione tra più componenti porti alla selezione di un singolo vincitore, 'chi vince piglia tutto', ma questo risulta vero soltanto con un'inibizione globale della rete neurale. Inoltre, se l'inibizione è locale, così come sembra avvenire nei principali centri nervosi, è possibile avere contemporaneamente diverse attività persistenti. Nel caso in cui le inibizioni giungano differite nel tempo, la loro azione in qualsiasi competizione e nella rete neurale potrà essere solamente di tipo sequenziale. Pertanto, le tracce di decadimento dei precedenti vincitori possono favorire, oppure opporsi, all'ingresso degli ultimi dati acquisiti dalla rete neurale durante la battaglia per la supremazia. Il modello di mente relazionale va oltre quello di Hume, estendendo le relazioni da lui descritte alle percezioni e includendo una serie di esperienze passate che interagiscono con quelle presenti. Le esperienze passate, al momento della loro prima apparizione o della loro evocazione, non devono essere obbligatoriamente coscienti. L'idea fondamentale è che la sensazione abbia origine grazie al confronto tra attività cerebrali originate da informazioni provenienti con varie modalità dal mondo esterno. Talvolta, analoghe attività passate vengono immagazzinate nelle cosiddette memorie di analisi preliminare e semantica (PM) e nelle memorie episodiche. Il contenuto mentale di un'esperienza consiste dunque nell'insieme delle relazioni di quell'esperienza con le memorie di altre esperienze precedentemente immagazzinate. In questo modo la sensazione cosciente di un cielo blu, come noi lo vediamo, è determinata da un insieme di memorie riguardanti le nostre esperienze trascorse di cieli blu, per esempio su una collina, sdraiati sulla spiaggia al mare, o su un'amaca con lo sguardo rivolto al cielo. La struttura del modello della mente relazionale presuppone che gli stimoli siano codificati in opportuni livelli superiori di analisi preliminare in relazione alla propria modalità sensoriale. Queste informazioni, trattate precedentemente a livelli periferici o inferiori del sistema nervoso, vengono quindi dirette verso la memoria di lavoro più appropriata, dove saranno in competizione con altre attività e con altre memorie di lavoro caratterizzate da altri codici. Le relazioni così attivate possono essere molto semplici, come per esempio l'analisi visiva di un colore puro, che implica la separazione dei dati afferenti in opportuni intervalli di lunghezze d'onda, oppure complesse, come l'analisi della parola e la relativa codificazione della semantica delle parole stesse nella rete della memoria di lavoro.
Una o più coscienze?
Se le relazioni definite tra le memorie di lavoro sono tali da non generare una competizione assoluta, per esempio tramite l'attività persistente ma decrescente delle precedenti memorie di lavoro vincenti, si evita il bisogno di una rete neurale che si comporti come 'chi vince piglia tutto'. Inoltre, l'attività di queste memorie di lavoro potrebbe aumentare con l'uso di opportune memorie episodiche o di valore. La coscienza sarebbe quindi prodotta dall'attivazione continua di un vincitore per un tempo minimo, che permette a quella particolare memoria di lavoro di vincere la competizione nei confronti delle altre memorie. L'attività della coscienza descritta in questi termini corrisponde a ciò che definiremmo sensazione cosciente o coscienza passiva, che qui indicheremo con Cp. Questo tipo di coscienza è in contrasto con la coscienza attiva, che qui indicheremo con Ca, che richiede un coinvolgimento maggiore delle componenti di introspezione e intenzionalità. In questo modo la coscienza nel suo insieme è definita come una funzione di due componenti distinti, Cp e Ca.
Tale distinzione era già stata fatta da D. Davidson, che aveva diviso l'esperienza cosciente in due parti: una fenomeno logica e l'altra associata alle opinioni ed emozioni (Davidson, 1970). Una suddivisione simile è stata effettuata anche da N. Block (1995) che, pur escludendo dal suo schema l' autocoscienza, attribuisce a Cp e Ca, rispettivamente l'aspetto fenomenologico e quello accessibile della coscienza. Il metodo da noi sviluppato, proponendosi di distinguere le regioni neuronali collegate ai vari aspetti informativi della coscienza, ha portato a una divisione analoga in modo indipendente (Taylor e Alavi, 1995). Il modello della mente relazionale può essere esteso anche alla coscienza attiva in modo tale da evidenziare le relazioni indispensabili che coinvolgono l'introspezione e l'intenzionalità. Alcune di queste relazioni sono eseguite come fenomeno emergente, a partire dal livello inferiore del contenuto cosciente Cp, tramite gli specifici meccanismi di competizione che derivano dalle successive attivazioni semantiche e delle altre memorie di lavoro.
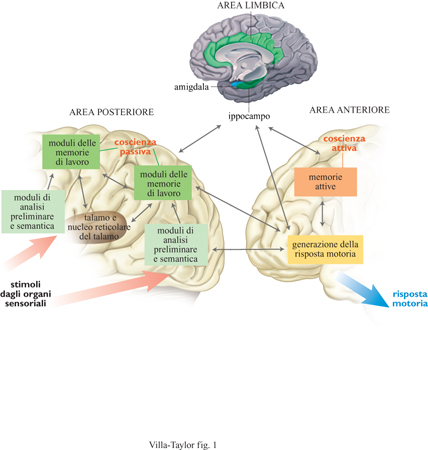
Tentando di indicare una corrispondenza neurobiologica con i concetti definiti precedentemente, appare evidente come gli indici p e a possano anche indicare le aree cerebrali posteriori e anteriori di trattamento dell'informazione. Le regioni di memoria preliminare e semantica PM e di memoria di lavoro WM, relativamente vicine alle informazioni afferenti, sarebbero collocate soprattutto nella regione posteriore. Alla regione anteriore è ragionevole attribuire un ruolo per l'intenzionalità, poiché il lobo frontale è fortemente legato all'azione e al suo eseguirsi. Ulteriori dati in Ca potrebbero venire da quelle strutture del lobo frontale collegate alla memoria individuale e sociale, ai sistemi di predizione, all'informazione volitiva e intenzionale. Ci sono inoltre altri scomparti, detti memorie attive, che possono mantenere le attivazioni per periodi uguali o superiori a 30 s, il cui contenuto è determinato da moduli posteriori. L'estensione di questa finestra temporale aumenta enormemente il numero di relazioni che possono essere attivate da Ca, rispetto a Cp, e consente pertanto una maggiore flessibilità della risposta. Ciononostante il contenuto cosciente di Ca può ancora essere considerato in termini di mente relazionale e verrà discusso in seguito. Nella figura (fig. 1) è descritta la struttura del modello di mente relazionale in funzione del collegamento tra le aree cerebrali coinvolte. Le frecce denotano la direzione del flusso di attivazione, in particolare quando viene innescata la competizione nel sistema TRC, costituito dal talamo, dal nucleo reticolare talamico (NRT) e dalla corrispondente area corticale. Il risultato finale di queste operazioni è una risposta motrice, generata da un modello neuronale dei gangli basali. Il sistema di guida (in particolare quello che proviene dall'ipotalamo e da altre parti del sistema limbico) non è qui discusso, ma è stato incluso per spiegare il legame tra motivazioni e azione.
Modello del filtro attivo dell'Io cosciente
Nell'esperienza quotidiana la coscienza ci appare come un processo unico. l moduli della memoria di lavoro, situati nelle cortecce parieto-temporale, temporale inferiore e frontale, sono tra loro distanti. Come possono dunque combinarsi le loro attività per produrre quella sensazione di unicità che si prova con la coscienza? Si può ipotizzare che tra i moduli distanti della corteccia si possa instaurare un'effettiva competizione globale tramite un'opportuna inibizione di questi moduli ottenuta, per esempio, mediante l'eccitazione dei neuroni inibitori locali da parte di afferenze eccitatorie provenienti da altre aree del cervello. Tuttavia, è dubbio che tale meccanismo possa verificarsi nei vertebrati superiori. È noto come la presenza di un corretto bilanciamento tra eccitazione e inibizione (Hill e Villa, 1997) sia essenziale per la selezione dell'orientamento visivo a livello della corteccia cerebrale. In questo caso l'inibizione si estende lateralmente solo per una distanza limitata, inferiore al diametro di un'ipercolonna corticale che corrisponde al più piccolo elemento funzionale di analisi (Braitenberg e Schiiz, 1991). Si deve inoltre notare che la selezione dell'orientamento visivo, osservata a livello della corteccia visiva primaria (VI), sembra piuttosto dovuta a un'attività neuronale pro attiva e già orientata, proveniente dal corpo genicolato laterale del talamo (LGN) e legata a un'attività riverberante tra corteccia e talamo che affina i campi recettivi. L'attivazione pro attiva a lungo raggio, verso i lobi temporali e parietali, è verosimilmente solo di tipo eccitatorio, come indicano le registrazioni elettrofisiologiche di singole cellule nervose. Un'inibizione laterale a lungo raggio appare dunque poco probabile nella corteccia.
'Chi vince piglia tutto'
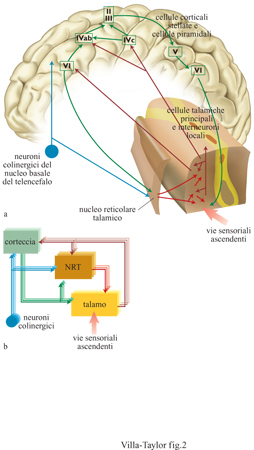
I collegamenti reciproci tra talamo e corteccia cerebrale sono una caratteristica generale di tutte le modalità sensoriali del sistema nervoso centrale, a eccezione dell'olfatto, e la loro funzione è generalmente intesa come integrazione talamo-corticale e retro azione corticotalamica (Steriade e Llinas, 1988). In questo circuito molti moduli del sistema TRC funzionano parallelamente e sono caratterizzati dalla stessa connettività di base che si articola nei tre componenti che seguono: i neuroni del talamo dorsale (per esempio del corpo genicolato mediale per la via uditiva o del corpo genicolato laterale per la via visiva) che ricevono le afferenze sensoriali dalla periferia; le cellule del nucleo reticolare del talamo che è uno dei componenti maggiori del talamo ventrale; l'area corticale che riceve le corrispondenti afferenze talamiche. Le proiezioni talamo-corticali e cortico-talamiche sono in larga misura reciproche. Lo schema di questa organizzazione modulare è illustrato nella figura (fig. 2). Nei nuclei del talamo dorsale (per brevità, ci riferiremo a questi nuclei come al talamo in quanto tale) potrebbe svilupparsi un'inibizione laterale sufficiente a mantenere il processo 'chi vince piglia tutto' (winner take all). Tuttavia, ci sono poche prove neuroanatomiche a sostegno di questa ipotesi, dato che gli intemeuroni inibitori del talamo hanno collegamenti molto circoscritti con le altre cellule talamiche. Nelle ricerche più recenti non hanno trovato conferma altre ipotesi di connessioni laterali tra i diversi nuclei del talamo, che spieghino l'attività corticale riscontrata durante il sonno e caratterizzata da un alto grado di sincronia e coerenza. Alla luce delle attuali conoscenze neuroanatomiche dobbiamo concludere che i nuclei del talamo dorsale non sono capaci di generare intrinsecamente un meccanismo del tipo 'chi vince piglia tutto'.
Il sistema TRC
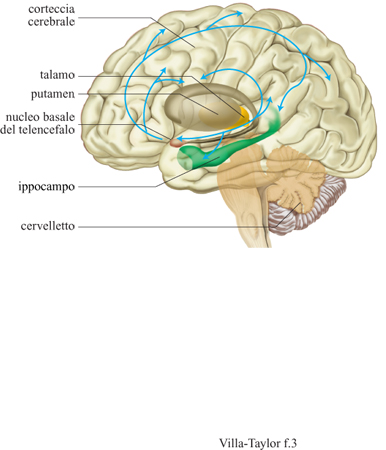
Un fine strato di cellule inibitrici forma il nucleo reticolare del talamo e si estende come un drappo sopra i nuclei del talamo dorsale. Ogni assone che parte dalla corteccia verso il talamo passa attraverso l'NRT e dà origine a fibre collaterali eccitatorie verso le cellule dell'NR T, così come ogni assone che ha origine nel talamo e si proietta verso la corteccia cerebrale. l neuroni dell'NRT proiettano le loro efferenze inibitrici solamente verso il talamo, regolando in questo modo l'attività dei neuroni talamocorticali. L'NR T riceve alcune afferenze anche da diversi nuclei del telencefalo e del mesencefalo noti per il loro ruolo modulatorio in ampie aree del cervello. È necessario, a questo punto, ricordare le fibre colinergiche del telencefalo basale perché anche nell'individuo adulto hanno recettori specifici per il fattore di crescita nervoso (NGF) e innervano l'NRT (fig. 3). Questo nucleo sembra essere una sede di regolazione dell'attività dell'NGF, e inoltre è ipotizzabile che i fattori neurotrofici possano influenzare il trattamento dell'informazione attraverso l'NRT stesso, come confermato da recenti indagini di elettro fisiologia (Villa et al., 1996). La regolazione dei fattori neurotrofici nell'NRT potrebbe essere selettivamente legata ai cambiamenti che si verificano nei terminali assonici colinergici del telencefalo basale, sensibili all'NGF, e alloro ruolo nell'apprendimento e nella memoria. L'NRT occupa una posizione così strategica nel trattamento dell'informazione a livello talamo-corticale da essere coinvolto, secondo diverse ipotesi, nella regolazione dell'attività cerebrale a livello globale. Spesso l'NRT è stato definito come un passaggio obbligato verso la corteccia cerebrale. A.B. Scheibel ha scritto che "la porta del nucleo reticolare diventa un mosaico di cancelletti ognuno dei quali è legato a specifici campi recettivi o a tipi particolari di afferenze", e suggerisce che "forse è qui che risiede il substrato morfofunzionale per l'attenzione selettiva, ed è nella delicatezza e complessità delle sue connessioni che si trova la nostra sorgente del sapere e della consapevolezza del sapere" (Scheibel, 1980).
L'ipotesi che noi proponiamo è che l'NRT, attraverso la sua intrinseca connettività globale, possa operare la selezione tra i vari moduli corticali specializzati, che entrano in competizione tramite l'attività congiunta di diverse regioni talamiche, della corteccia e dell'NRT. La perdita di attività nell'NRT provocherebbe la perdita delle tipiche caratteristiche globali di questa competizione. In un certo senso si potrebbe dire che se l'NRT fosse dissecato, o rimanesse collegato solo localmente, il sistema funzionerebbe in un modo simile a quello suggerito da Scheibel, ossia come tanti cancelletti. Durante lo stato di veglia si suppone che l'NRT funzioni come un cancello globale, in modo tale che il trattamento dell'informazione corticale avvenga con una competizione globale. Discutiamo ora diverse prove a favore di questa tesi.
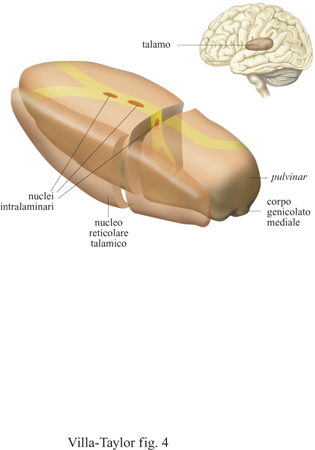
Un primo sostegno alla nostra tesi è di tipo neuroanatomico. Il ruolo chiave dell'NRT può essere descritto utilizzando l'analogia del passaggio da un cancello: se immaginiamo la corteccia come il magazzino dei dati afferenti, sia presenti che passati, allora il talamo comprende un insieme di nuclei, corrispondenti a diverse modalità sensoriali, ognuno dei quali viene identificato da una specifica porta. In questa analogia l'NRT agisce come un cancello, permettendo il flusso dei dati afferenti solamente all'una o all'altra porta (fig. 4). Ci sono altri nuclei talamici che non appartengono alle vie sensoriali principali. Uno di questi nuclei è il pulvinar che ha importanti afferenze ed efferenze corti cali organizzate in modo ben definito. Nella scimmia è stato sperimentalmente dimostrato che le lesioni delle vie visive afferenti, dalla retina verso il pulvinar inferiore, hanno scarso effetto sulle caratteristiche di risposta dei neuroni del pulvinar stesso. Le lesioni nella corteccia visiva primaria eliminano invece la risposta di tutti i neuroni del pulvinar inferiore agli stimoli visivi per un periodo di almeno tre settimane, a partire dal momento della lesione. Si pensa che il pulvinar, e la regione dell'NRT a esso collegata, possano giocare un ruolo importante nei processi di attenzione. Gli studi basati su lesioni selettive del pulvinar nei primati hanno mostrato, infatti, che in tutti i casi in cui veniva riscontrata la corrispondente lesione dell'NRT si osservavano chiari deficit nei tempi di reazione delle risposte a test comportamentali.
A ulteriore sostegno della nostra ipotesi, misure elettroencefalografiche effettuate in pulvinar di scimmie durante esperimenti di attenzione visiva, hanno mostrato un aumento di attività in questa regione talamica, confermato peraltro da recenti studi sull'uomo (La Berge, 1990) tramite tomo grafia a emissione di positroni (PET, Positron Emission Tomography). Non è possibile tuttavia definire con precisione quale sia il grado di coinvolgimento dell'NRT nelle varie fasi di svolgimento degli esperimenti comportamentali di attenzione visiva. Ciononostante, la ben nota connessione eccitatoria tra il pulvinar e il vicino NRT suggerisce fortemente un coinvolgimento concomitante di questo nucleo come parte del processo. Misure dell'attività cerebrale (Llinas et al., 1992) eseguite con tecniche di magnetoencefalografia, con sistemi di rilevamento multipli e durante compiti di attenzione, hanno fornito ulteriori argomenti favorevoli all'idea di un processo globale di controllo. In un compito di attenzione uditiva, durante il quale il soggetto doveva prestare attenzione a particolari suoni puri, è stato osservato un rapido spostamento, dalla parte anteriore verso la parte posteriore del cervello, di un onda di attività cerebrale (le cosiddette oscillazioni a 40 Hz), caratterizzata da picchi di attività talamici che precedevano di circa 3 ms i picchi di attività corticali. Si è inoltre osservato che il fronte di quest'onda di attività talamo-corticale è situato nell'NRT. L'origine di questa attività a 40 Hz potrebbe risiedere nelle proprietà di alcune cellule talamocorti cali caratterizzate da oscillazioni intrinseche del potenziale elettrico della membrana cellulare, a frequenze comprese tra 20 e 40 Hz (Steriade et al., 1990). Tale attività oscillatoria può essere collegata a quella originariamente osservata nella corteccia visiva del gatto, benché la dipendenza di queste oscillazioni dagli stati di veglia non sia ancora chiara.
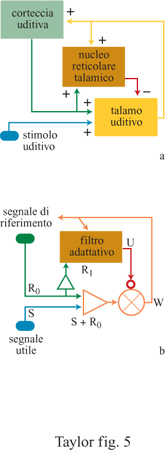
Il ruolo fisiologico cruciale del sistema talamo-reticolocorticale (TRC) nel trattamento dell'informazione sensoriale è illustrato dal seguente esempio per il sistema uditivo. Risultati sperimentali hanno mostrato che l'attività corticofuga (ovvero l'attività esercitata dalla corteccia cerebrale sulle strutture sottocorticali) modula le proprietà di risposta delle reti neurali talamiche, cambiando la loro sensibilità a stimoli acustici che includono gamme di frequenze sonore ben precise. In questo modo è possibile estrarre selettivamente, in funzione dell'attività corticale, un'informazione contenuta nel segnale sensoriale afferente (Villa et al., 1991). Di fatto, esistono strette analogie tra un modulo TRC e la descrizione dei filtri elettronici adattativi usati dagli ingegneri (fig. 5). Infatti, supponiamo che un segnale proveniente dalla periferia contenga il segnale utile S. Il segnale di riferimento Ro è prodotto dall'attività corticale che si propaga al corpo genicolato mediale (il nucleo principale nel talamo appartenente alla via uditiva) e non è correlato in alcun modo con il segnale proveniente dalla periferia. Un altro segnale di riferimento Rl è correlato solo con Ro. Se U è il segnale in uscita dal filtro corrispondente al profilo di attività delle cellule dell'NRT (Villa, 1990), come accade per i filtri elettronici quando i coefficienti del filtro cambiano in modo da minimizzare l'errore (cioè la distanza tra il segnale di uscita e la prestazione attesa), allora il segnale in uscita W, che corrisponde al profilo di attività riscontrato nel talamo uditivo, rappresenta la migliore approssimazione del segnale d'entrata S (Tetko e Villa, 1997).
Controllo globale e onde di attività
Un altro metodo di studio è basato sulla simulazione all'elaboratore. In questi anni sono stati eseguiti diversi lavori preliminari atti a costruire un modello realistico dell'NRT come rete competitiva. Tramite simulazioni di semplici modelli (La Berge et al., 1992), basati sul modo di azione inibitoria dell'NRT, è stato mostrato che la competizione laterale tra neuroni può essere efficace per evidenziare un massimo, ossia un picco di attività, in un insieme locale di informazioni afferenti. Questi modelli hanno sfruttato esplicitamente la presenza della retroazione esercitata dall' attività delle fibre collaterali, provenienti da diverse aree corti cali, al fine di ottenere una relazione competitiva tra le attività corticali locali. Uno studio, indipendente dal precedente (Taylor, 1992), ha invece analizzato la presenza di reciproche azioni inibitorie locali tra cellule dell'NRT, dimostrate negli studi neuroanatomici. In questo modello le onde di attività oscillatoria possono propagarsi nell'intero NRT, a partire da particolari configurazioni iniziali di attività locale. Ciò è dovuto al modo in cui l'inibizione laterale locale è capace di sopprimere l'attività entro un certo raggio. Oltre questo limite l'attività può riorganizzarsi a un ulteriore livello locale ottimale. A sua volta, questa attività svanisce a distanze maggiori, via via propagandosi e poi svanendo fino al raggiungimento di una determinata zona di influenza. Si ha in questo modo un andamento ondulatorio; la lunghezza d'onda di tale oscillazione può essere determinata con una dettagliata analisi della dinamica, e può essere ottenuta tramite opportune simulazioni all'elaboratore. Analoghe reti neurali appaiono anche nell'analisi del modo in cui si formano particolari profili di attività corticale, nei quali coesistono attività oscillatorie eterogenee (Cohen e Murray, 1981), così come nel modellamento di effetti allucinatori (Ermentrout e Cowan, 1978). Sono queste onde di attività che consentono all'attività evocata localmente nell'NRT di controllare l'attività in un'altra sua regione. Esiste inoltre un'importante interrelazione tra l'NRT e i nuclei intralaminari del talamo (v. figura 4). Questi formano un insieme di piccoli nuclei che hanno un ruolo importante nella coscienza visivo-spaziale, nell'attenzione e nella memoria, poiché diffondono un segnale di stimolo generalizzato verso la corteccia. È quindi verosimile che attraverso il reclutamento dei nuclei intralaminari del talamo, l'NRT sia capace di produrre lente oscillazioni corticali sincronizzate in molte regioni.
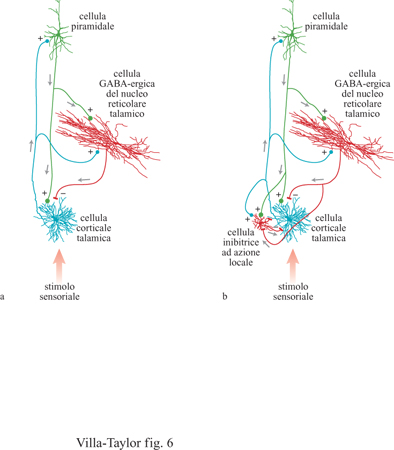
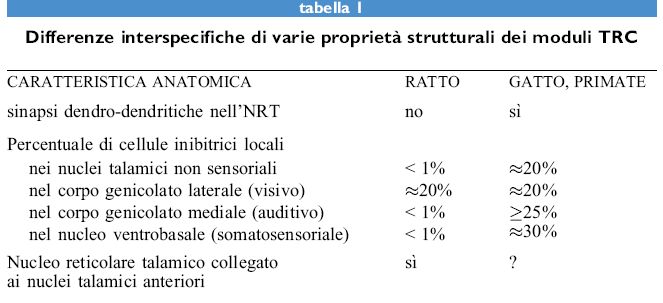
Poiché come abbiamo già notato, l'NRT riceve afferenze sia dal talamo che dalla corteccia ma si proietta, per vie inibitorie, esclusivamente verso il talamo, l'NRT può esercitare un controllo globale sull'attività afferente o sull'attività in elaborazione nella corteccia cerebrale. È importante notare che ci sono differenze nel livello di retro azione esercitato dall'NRT sul talamo (Winer e Larue, 1996) a seconda delle specie animali prese in esame (fig. 6). Alcune simulazioni indicano che, quando un certo tipo di attività di retroazione esercitata dall 'NR T non corrisponde ai parametri ottimali, le proprietà tipiche che permettono all'NRT di fungere da controllo globale vengono perse. Può tuttavia persistere un'attività in aree indipendenti dell'NRT, a condizione che queste aree siano sufficientemente distanti da evitare interferenze di corto raggio (Taylor e Alavi, 1993; 1995). Sono interessanti le conseguenze di questi effetti sul ratto che, come sappiamo dagli studi di neuroanatomia, non possiede sinapsi dendro-dendritiche nell'NRT. Ci si può dunque chiedere se, secondo il modello descritto in queste pagine, il ratto possieda una singola o più coscienze. In effetti, le informazioni analizzate e ritrasmesse dalla corteccia limbica sono collegate agli stati emozionali, così come alle memorie remote. Esistono reciproche connessioni tra i nuclei anteriori talamici e le strutture limbiche, ma questi nuclei talamici non hanno connessioni dirette con l'NRT tranne che nel ratto (tab. I). Una connessione tra le strutture limbiche e l'NRT procede in modo indiretto e unidirezionaIe dal complesso del subicolo verso il talamo mediodorsale, il quale è specificatamente collegato con la parte anteriore dell'NR T. Osserviamo che nel ratto non esistono quelle connessioni di controllo che permetterebbero all'attività presente nei circuiti limbici di riversarsi sull'NRT per poi ricevere la retro azione dal talamo anteriore. Potremmo quindi concludere con l'ipotesi che nel ratto le emozioni sono inestricabilmente legate alla coscienza.
Sensazione cosciente
Interessanti risultati, risalenti a trenta anni fa (Libet et al., 1964), riguardano la natura del controllo globale e forniscono un solido sostegno al modello qui sviluppato. Gli esperimenti di B. Libet si proponevano di determinare la soglia di corrente elettrica necessaria a provocare una sensazione cosciente, quando sulla circonvoluzione cerebrale postcentrale veniva inserito un elettrodo stimolatore del diametro di l mm. La sensazione cosciente nel soggetto risultava molto simile a uno stimolo superficiale localizzato sulla pelle, come un pizzico sul dorso della mano. Lo stimolo era costituito da una serie di brevi impulsi elettrici della durata di alcuni millisecondi. Questi risultati evidenziano due particolarità collegabili con il funzionamento del sistema TRC. La prima osservazione è legata all'esistenza di una soglia di sensazione cosciente. Allorché una stimolazione di durata inferiore a 500 ms superava un certo valore critico di energia elettrica (proporzionale alla frequenza degli impulsi moltiplicata per la durata della stimolazione e per l'intensità della corrente al quadrato) il soggetto riportava una sensazione cosciente di prurito. La seconda osservazione è che, per stimolazioni con durata superiore a 500 ms, quel valore cntIco cambiava: per mantenere la sensazione cosciente della stimolazione occorreva aumentare l'energia elettrica in modo proporzionale alla durata.
La necessità di una quantità minima di energia elettrica per generare la sensazione cosciente per un breve intervallo di tempo è coerente con il precedente modello di controllo dell'NRT che, come già accennato, funziona come un circuito globale di controllo. Dapprima, il circuito selezionerebbe il modulo di attività vincente in funzione della richiesta e dell'energia elettrica. In seguito, considerato il secondo risultato di Libet, si richiederebbe una maggior potenza dello stimolo per mantenere il circuito attivato a controllare il sistema. Poiché il circuito in attività richiede una potenza elettrica maggiore, lo stimolo deve avere una potenza elettrica superiore al valore critico rappresentato dalla soglia di sensazione per mantenere il controllo della coscienza per periodi più lunghi.
La natura dello stimolo necessario al raggiungimento della sensazione cosciente costituisce un altro aspetto importante degli studi iniziati dal gruppo di Libet. La percezione è un fenomeno centrale, cioè derivante dall'attività del cervello, ma generalmente gli stimoli sono variazioni di energia che vengono recepite dai recettori posti alla periferia del sistema nervoso. La sensazione di un pizzico sulla pelle può richiedere soltanto un singolo impulso nervoso a condizione che provenga dalla cellula sensoriale adeguata. Viceversa, nel caso in cui lo stimolo sia applicato direttamente al cervello, come abbiamo visto prima, la sensazione diventa cosciente soltanto tramite una serie di impulsi elettrici di durata corrispondente alla corretta attivazione neuronale. Libet (1982) ha mostrato che un singolo impulso d'attivazione periferica sulla pelle genera dei potenziali tardivi negativi, registrati con macroelettrodi inseriti nella corteccia cerebrale. Supponiamo che tali potenziali corrispondano a quello stimolo afferente che ha rapidamente ottenuto l'accesso all'appropriata memoria di lavoro e che ivi rimanga attivo. La nostra definizione stessa di memoria di lavoro (WM) è legata a questo concetto di mantenimento dell'attivazione e concorda con un numero sempre maggiore di osservazioni sperimentali effettuate in diverse regioni cerebrali delle scimmie. l primi risultati che misero in rilievo l'attività congiunta e prolungata, sia del talamo che della corteccia cerebrale, vennero presentati da E.W. Dempsey e R.S. Morison negli anni Quaranta (Dempsey e Morison, 1942). Le loro ricerche si riferivano a una generica attività di proiezione, caratterizzata da una risposta complessa alla stimolazione sensoriale, e piuttosto sensibile all'anestesia. Questi dati furono completati nei decenni successivi e sono ancor oggi oggetto di specifiche ricerche che sostengono l'esistenza di veri e propri fenomeni di risonanza cortico-talamica basati sulla coattivazione di questi nuclei cerebrali, analogamente al fenomeno di risonanza osservabile in musica. Le simulazioni del modello competitivo 'chi vince piglia tutto' (Taylor e Alavi, 1995) indicano che anche la stimolazione della corteccia, per quanto sia intensa, richiede un tempo minimo (mai nullo) per vincere la competizione, in accordo, quindi, con i risultati sperimentali (Libet, 1982). Inoltre, Libet e i suoi collaboratori hanno dimostrato che l'uso di stimolazioni multiple, sia nel nucleo VPL (Ventrale Posteriore Laterale) del talamo (corrispondente alla suddivisione talamica preposta alla modalità somatosensoriale) sia nella corteccia, non provoca la consapevolezza della stimolazione, poiché ciascuna stimolazione è caratterizzata da un livello energetico inferiore alla soglia necessaria per innescare la sensazione cosciente. Lo stesso risultato è riscontrabile nelle simulazioni all'elaboratore, allorché una stimolazione intensa venga interrotta immediatamente dopo la sua attivazione: la variazione di energia è molto modesta e di conseguenza la possibilità d'innescare un meccanismo competitivo vincente è molto scarsa. Pertanto, essendo necessario attivare ripetutamente con una serie di impulsi i circuiti TRC, per attivare, a loro volta, le corrispondenti memorie di lavoro WM, si suppone che l'attivazione prodotta da un singolo stimolo superficiale sulla pelle non sia da sola sufficiente per accedere direttamente alla memoria di lavoro. Per interpretare questo dato bisogna considerare che l'attivazione periferica non si riduce alla semplice attivazione della via sensoriale confluente al talamo e alla corteccia, ma alla rapida attivazione della WM (in circa 50 ms) che fa verosimilmente intervenire specifici sistemi neuromodulatori mesencefalici. Questi circuiti sono attivati da fibre collaterali delle vie sensoriali ascendenti e si proiettano direttamente (in modo monosinaptico) verso la corteccia cerebrale con proiezioni collaterali verso specifici nuclei del talamo (come per esempio i nuclei intralaminari citati in precedenza). Le aree corticali che ricevono queste proiezioni modulatrici sono molto vaste cosicché i neurotrasmettitori tipici di queste proiezioni provocano degli effetti diffusi su tutta l'attività corticale. Consideriamo la proiezione colinergica eccitatoria (v. figura 3) che ha origine nell'area peduncolopontina tegmentale del mesencefalo e che raggiunge l'NRT (Steriade et al., 1990). Di conseguenza, un'attività collegata a uno stimolo periferico in entrata nel sistema TRC, per esempio il pizzico sulla pelle, può attivare direttamente una parte specifica dell 'NR T attraverso il suo cancelletto locale, coattivato dalla corteccia somatosensoriale, per poi attivare la memoria di lavoro appropriata e innescare la competizione 'chi vince piglia tutto'. La contemporanea attivazione del circuito neuromodulatore provoca la selezione del circuito vincente nell'NRT, e ciò costituisce una prova oggettiva del modello competitivo della coscienza.
Il modello esteso a livello cellulare
Occupiamoci ora di ciò che potrebbe succedere a livello cellulare. l mutamenti dell'eccitabilità cellulare sono generalmente associati alle proprietà della membrana neuronale. Il potenziale di membrana può essere iperpolarizzato o depolarizzato da innumerevoli meccanismi e sarebbe difficile, se non impossibile, voler considerare tutte le possibili reazioni biofisiche. Gli effetti simulati all'elaboratore dell'attivazione delle vie neuromodulatrici sul sistema TRC possono essere semplificati riducendoli a due parametri essenziali. Il primo parametro, l' eccitabilità della cellula, corrisponde alla differenza tra il potenziale di membrana a riposo e il potenziale di soglia. Il secondo parametro, relativo alla cinetica delle reazioni elettriche della membrana cellulare, corrisponde alla costante di tempo dei potenziali postsinaptici eccitatori e inibitori. Questo valore è effettivamente costante in un determinato momento dell'attività cellulare, quando viene utilizzato nel calcolo delle equazioni del potenziale di membrana, ma è variabile nell' arco di minuti o ore. Stimolando il sistema TRC, dopo un certo tempo e se non intervengono altre perturbazioni, la risonanza talamo-corticale permette all'attività di stabilizzarsi in modo riverberante. Definiamo modo di attività quella particolare attività che si è stabilizzata nel sistema TRC quando esso analizza una specifica informazione afferente, corrispondente a particolari serie di impulsi nervosi detti profili d'attività. In uno stesso modulo TRC si possono osservare diversi modi di attività a seconda dei cambiamenti della connettività interna o dell'eccitabilità degli elementi neuronali che compongono il modulo o di entrambi i fenomeni.
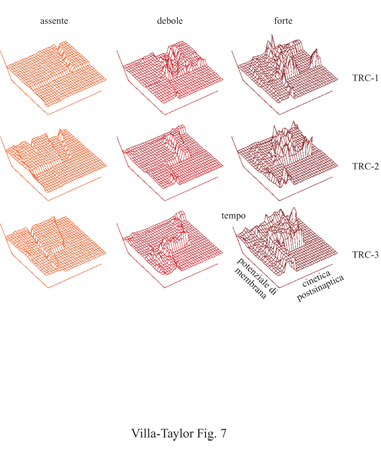
Un insieme composto da differenti profili iniziali d'attività può a sua volta generare uno stesso modo di attività, alla maniera degli attrattori nella teoria dei sistemi dinamici. Infatti, in questo contesto il sistema TRC viene considerato come un sistema dinamico non lineare che opera all'interno di uno spazio definito da parametri di controllo, quali la cinetica postsinaptica e il potenziale di membrana. Queste considerazioni teoriche hanno trovato di recente riscontro nella ricerca sperimentale che, tramite l'analisi delle sequenze dei potenziali d'azione neuronali, con metodi matematici propri dell'analisi dinamica non lineare, ha provato l'esistenza di attrattori di bassa dimensionalità nel talamo del ratto (Celletti e Villa, 1996). Nella figura (fig. 7) è illustrato il risultato di una simulazione di tre diversi sottotipi di un sistema di base TRC (Villa e Tetko, 1995). Nella simulazione più semplice i moduli TRC hanno tra loro una debole interazione e l'informazione è trattata in modo indipendente all'interno di ogni modulo. In questo caso il paesaggio dello spazio delle fasi illustrato nella figura 7 è piuttosto piatto, lasciando intravedere soltanto la presenza di pochi massimi locali. Con l'incremento dell'interazione tra i moduli, l'attività competitiva che si viene a creare, genera un numero maggiore di massimi locali. l profili d'attività che corrispondono a questi massimi possono essere interpretati come un correlato dell'insorgenza della sensazione cosciente nella memoria di lavoro appropriata, associata a uno specifico modulo di analisi preliminare o semantico (Taylor e Alavi, 1993; 1995). Si può quindi ipotizzare che nella rete neurale TRC sia possibile osservare i profili di attività riverberanti solo quando una specifica afferenza neuromodulatrice, per esempio la proiezione colinergica del nucleo basale del telencefalo (un nucleo particolarmente colpito nel morbo di Alzheimer), sia associata a uno specifico profilo di attività sensoriale. In conclusione, gli esperimenti di Libet convalidano il modello competitivo suggerito per spiegare l'insorgere della sensazione cosciente associata alla coscienza passiva Cp e altresì confermano il sistema TRC nel suo ruolo determinante per l'insorgere della sensazione cosciente. Questi esperimenti rafforzano inoltre l'ipotesi che la sensazione cosciente sia un fenomeno che dipenda da attività localizzate in siti ben definiti: i moduli delle memorie di lavoro, che entrano in competizione tra loro per mezzo delle inibizioni laterali esistenti nel nucleo reticolare del talamo. Il tempo necessario affinché una particolare attività afferente nel sistema TRC possa trasformarsi in sensazione cosciente può essere interpretato come la latenza necessaria alla creazione di un sito specifico di memoria di lavoro. La normale attività afferente alla corteccia raggiungerebbe quindi tale sito di memoria di lavoro, che si trova in uno stato già innescato, tramite il sistema attivatore reticolare e la sua vasta rete di connessioni che include l'NRT.
Ritorno alla coscienza
Per sostenere una persistente attività afferente e quindi rendere possibile la manifestazione della coscienza, viene utilizzata la memoria a breve termine o, più precisamente, la memoria di lavoro, così definita da Baddeley e Hitch: "nel nostro pensiero tendiamo a equiparare la memoria a breve termine alla coscienza, ossia i pensieri e l'informazione di cui siamo consapevoli al momento possono essere considerati parte della memoria a breve termine" (Atkinson e Shiffrin, 1971). Ciò richiede che una certa attività deve persistere per almeno alcuni secondi in una lavagnetta mentale (un registro di memoria) prima che sia cancellata, senza possibilità di recupero. Questa identificazione della coscienza con la memoria primaria o a breve termine era stata originalmente introdotta da W. James negli anni Quaranta. In seguito, è stata estesa e modificata per tener conto dei differenti tipi di memoria a breve termine, come le memorie di lavoro e le memorie attive in diverse regioni cerebrali.
Queste considerazioni sulla memoria a breve termine ci inducono a riassumere la nostra argomentazione nei seguenti concetti: l) la teoria relazionale della mente, per la quale le informazioni afferenti precedentemente codificate secondo una semantica propria e le memorie episodiche, a esse collegate, sono confrontate in specifici moduli decisionali; 2) l'lo cosciente che si identifica con l'NRT nel quale la competizione globale è attuata grazie alle sue caratteristiche interne di connettività, e più precisamente tramite il controllo retro attivo al quale è sottoposto; 3) la sensazione cosciente, o consapevolezza, intesa come fenomeno originato dalla competizione localizzata all'interno di un modulo i cui nodi codificano diverse interpretazioni della stessa informazione afferente, a partire dalla codificazione semantica di origine. Per poter formulare un insieme di principi fondamentali che completano il modello originale, occorre definire più in dettaglio il modello di mente relazionale legato alla coscienza passiva, inserendo i punti (2) e (3) sopra descritti. l principi possono essere così formulati: l) per ogni codice legato alle informazioni afferenti (per esempio un codice α o un codice β) esiste una coppia, o un insieme di coppie, di moduli di memoria per l'analisi preliminare, PM, e di memoria di lavoro, WM (indicati rispettivamente PMα, WMα e PMβ, WMβ quando si riferiscono a due codici distinti α e β); 2) per uno specifico modulo WMα esiste una competizione tra le diverse attività neuronali, che rappresentano interpretazioni diverse degli input appena arrivati; 3) si stabilisce una competizione tra le attività delle diverse memorie di lavoro di modo tale che il vincitore, indicato WMwin, guadagna l'accesso alla coscienza; 4) le memorie episodiche (EM) esercitano un controllo di ritorno o retroattivo sulla competizione tra le varie coppie PM e WM; 5) l'aggiornamento delle memorie episodiche viene eseguito dalle informazioni provenienti dalla memoria di lavoro vincente, WMwin; 6) l'attività nella memoria di lavoro vincente è immagazzinata nell'ippocampo e nelle strutture a esso direttamente collegate, come un 'tampone' utilizzabile (eventualmente anche in modo non cosciente) in momenti successivi; 7) l'attività prodotta dalla coppia vincente PMwin e WMwin guida il pensiero e la pianificazione, ma a causa di questo stesso ruolo guida non è possibile richiamare WMwin dalla memoria a lungo termine; 8) le attività presenti negli altri moduli di analisi preliminare (quindi i diversi PMα e PMβ a eccezione di PMwin) non hanno accesso alla coscienza ma possono essere usate per trattamenti automatici o non coscienti dell 'informazione; tale attività perdente pertanto non viene completamente perduta; 9) l'aggiornamento della memoria, sia essa memoria dichiarativa, a livello del complesso dell'ippocampo, o nei moduli semantici PM, oppure memoria non dichiarativa nei circuiti riverberanti formati dalle connessioni tra lobo frontale, gangli basali e talamo frontale, può avvenire in momenti successivi al manifestarsi della sensazione cosciente; 10) le informazioni provengono da WMwin con diverse modalità, con possibili difetti di trasmissione che aumentano con la diminuzione del livello del segnale.
Riscontri sperimentali al decalogo della coscienza passiva
Tentiamo ora di collegare questi dieci principi e chiarire alcune definizioni. lnnanzitutto precisiamo il termine codice che qui si riferisce alle informazioni in uscita dagli analizzatori non coscienti, eventualmente organizzati in vari livelli di complessità, che agiscono esclusivamente sulle informazioni sensoriali primarie. Queste informazioni provengono direttamente dalla periferia e sono legate in maniera specifica a una particolare modalità sensoriale. l livelli più alti di tale gerarchia sono i moduli che abbiamo denominato PMα, allorché ci riferiamo a un codice α. Per una stessa modalità sensoriale, per esempio quella visiva, esistono molti codici diversi quali la struttura, la forma, il moto, la posizione, il colore e altri ancora. Il livello del codice può considerarsi primitivo, come nel caso del colore e del moto, ossia legato direttamente ad alcune caratteristiche fisiche costitutive dell'oggetto. Il livello del codice può anche appartenere a un alfabeto di livello superiore, come suggerito dai geoni di Biederman (Biederman, 1993). l geoni, il cui termine deriva dalla contrazione delle parole inglesi generalized cane, rappresentano archetipi volumetrici la cui percezione è invariante rispetto al punto di osservazione. Questi archetipi possono formarsi a partire dall'analisi preliminare dei rivelatori, creatisi durante processi di apprendimento, a base di funzioni logiche di tipo e, di volta in volta più complicate. Nell'udito il livello più alto della gerarchia è occupato dai moduli di analisi preliminare e semantica, che trattano codici complessi fondati su un alfabeto di fonemi e persino di parole con poche sillabe.
In psicofisiologia esistono ampi riscontri sperimentali a sostegno delle definizioni date e del punto l sopracitato (Baddeley, 1993) sull'attivazione della memoria di lavoro in vari codici: l'immagazzinamento fonologico dei fonemi, la rappresentazione visivo-spaziale per gli attributi visivi di spazio e di forma, e la rappresentazione della superficie corporea per le afferenze somatosensoriali. Inoltre, l'esistenza di specifiche memorie di lavoro corticali localizzate per l'udito e per la visione sono state evidenziate da analisi di neuroimmagini eseguite con la PET (Paulesu et al., 1993). Spesso i moduli coinvolti sono numerosi e si riferiscono sia all'attività di analisi preliminare e semantica, sia a quella delle memorie di lavoro continuamente sollecitate.
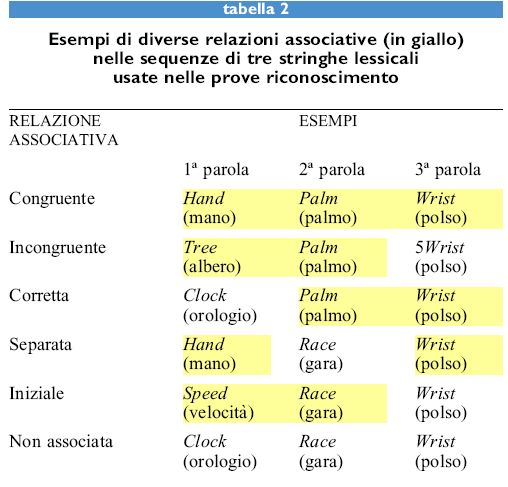
Per generalizzare il modello è necessario considerare esperimenti che studino il modo in cui le prime informazioni afferenti possano modificare la risposta alle afferenze successive. La specifica manipolazione sperimentale di queste prime afferenze, che le rende percepibili o no, permette di scoprire vari aspetti dell'insorgere della sensazione cosciente nella percezione di parole, per esempio nell' analisi di decisioni lessicali trattate in maniera subliminale. In questo tipo di esperimenti eseguiti da A.J. Marcel e collaboratori (Marcel, 1980), un soggetto doveva prendere una decisione sul carattere di parola o di non parola di una stringa lessicale. Una prova consisteva in una serie di tre stringhe lessicali, ove la seconda lettera della stringa era mascherata, in modo da rendere necessario l'uso di diversi livelli coscienti o subliminali per poterne percepire il significato. Alcuni esempi di stringhe formate da 3 parole tipiche in lingua inglese sono presentati nella tabella (tab. 2). Quando il soggetto doveva pronunciarsi sul carattere lessicale della terza parola, si è osservato che il tempo necessario per formulare una risposta era influenzato dalla relazione semantica che la terza parola aveva con le parole precedenti. Inoltre, questo risultato suggerisce che, al momento dell'analisi della seconda parola, tutti i possibili codici semantici erano apparentemente ancora accessibili da parte del soggetto, ma che ciò non avveniva a livello cosciente. Nel caso di una parola polisemica, come per esempio la parola inglese race, che può indicare sia la gara di corsa che l'insieme degli individui di una specie con tratti visibili in comune, soltanto uno dei significati era accessibile a livello cosciente quando questa parola era presentata in seconda posizione. Pertanto, i tempi di risposta da parte del soggetto erano ritardati se l'evocazione cosciente della parola aveva carattere semantico opposto al contesto delle altre stringhe lessicali.
Questo esperimento può essere simulato con un modello composto da due moduli, uno per la codificazione semantica a livello preconscio PM e l'altro, la memoria di lavoro WM, per l'insorgere della sensazione cosciente allorché l'eccitazione dell' area appropriata supera il livello critico di soglia. Solamente nel modulo WM tutte le competizioni possono svilupparsi per inibizione laterale, propagandosi da un nodo attivo a uno che sia semanticamente contraddittorio. È utile ricordare anche l'esistenza di un'eccitazione laterale tra una parola elaborata in maniera cosciente e una parola successiva, collegata alla precedente in modo semantico. Le interazioni laterali sembrano seguire un tipo di funzione detta' sombrero messicano', caratterizzata da un picco centrale e un avvallamento laterale che aumenta il contrasto tra le due parole e ne facilita la percezione. Come osservato da Marcel, è attraverso le interazioni eccitatorie laterali (sia quelle che procedono dal modulo PM al modulo WM, sia quelle laterali tra i nodi del modulo WM) che il tempo di reazione può diminuire, grazie all'analisi subliminaIe delle parole in seconda posizione nella stringa di tre parole. Un ritardo del tempo di reazione nell' analisi di una seconda parola elaborata coscientemente è invece dovuto alle interazioni laterali inibitorie tra i nodi della rete WMwin. È in questo caso che intervengono le costanti di tempo più lunghe della memoria di lavoro; la presenza nella memoria di lavoro vincente dell'attività precedente vincola un'ulteriore attivazione dello stesso modulo WMwin. Secondo questa interpretazione la sensazione cosciente, al livello più elementare, dipende inizialmente da un processo capace di risolvere l'ambiguità della stringa lessicale. Partendo dalle tracce di attivazione delle informazioni trattate inizialmente e ancora presenti nella memoria di lavoro, il processo mentale fornisce gli elementi contestuali favorevoli a risolvere l'ambiguità delle informazioni più recenti.
L'inibizione intramodulare, postulata sopra, può invero essere aumentata, o persino essere interamente sostituita, dagli effetti sottocorticali prodotti dalle inibizioni laterali nell'NRT che, passando da un modulo all'altro, riescono ad agire a distanze sufficienti a spiegare i dati di Libet. Simulazioni che prendono in considerazione queste inibizioni forniscono risultati che si accordano bene con i dati sperimentali elettroencefalografici sulle onde negative di non corrispondenza (mismatch negativity). Queste onde sono state osservate in soggetti sottoposti al riconoscimento di frequenze acustiche devianti in un insieme standardizzato e possono essere interpretate come il risultato della competizione che si sarebbe instaurata tra i nodi attivati per i suoni standard e quelli attivati dalle frequenze devianti. Inoltre, fenomeni di alterazione percettiva, come quelli provocati dalla visione del cubo di Necker, forniscono una prova meno diretta della competizione intramodulare. Un'analisi con due moduli neuronali, ognuno dei quali responsabile della codificazione di ciascuna interpretazione alternativa del cubo di Necker, ha mostrato un buon accordo con i tempi di alternanza riportati dai soggetti (Riani e Masulli, 1990). Anche in questo caso il modello soddisfa il principio di cui al punto 2 in quanto attribuisce ai due moduli in competizione un ruolo specifico all'interno dello schema percettivo visivo-spaziale.
Pensieri intrusivi
l risultati degli esperimenti compiuti dal gruppo di Libet offrono un importante sostegno sperimentale al principio espresso dal punto 3, precedentemente discusso. Ulteriori dati a conferma di questo principio sono stati forniti dai recenti esperimenti che Baddeley e i suoi collaboratori hanno svolto sui pensieri intrusivi (Baddeley, 1993). La depressione è una delle patologie psichiatriche più diffuse nella società contemporanea e i soggetti che ne sono affetti riferiscono spesso che i loro ragionamenti sono costantemente interrotti da pensieri di varia natura. È pertanto spiegabile l'attenzione volta alla comprensione e al controllo dei pensieri intrusivi. Dobbiamo distinguere due tipi di stimoli intrusivi indipendenti dal pensiero volontario: gli stimoli sequenziali e coerenti da un lato e quelli di carattere più frammentato, dall'altro. Chiedendo in modo irregolare a un soggetto seduto in silenzio, in una stanza tranquilla, quali siano i suoi pensieri, risulta che egli ha pensieri intrusivi durante circa 1'80% del tempo totale, costituiti da pensieri sequenziali e coerenti per un tempo pari al 65% e da pensieri frammentari per il rimanente 15%. L'esecuzione di opportuni compiti distraenti (come l'ascolto e la successiva ripetizione di una sequenza di 5 cifre presentate una volta al secondo) riduce in modo consistente i pensieri coerenti (sino a circa il 15%), ma determina solo piccoli cambiamenti al livello del pensiero frammentario. Si è inoltre osservato che una simile riduzione dei pensieri intrusivi risulta più o meno indipendente dalla modalità della distrazione, per esempio anche se il compito di distrazione visiva consiste nel seguire un bersaglio con lo sguardo. Nel caso dell'esperimento con le cinque cifre, i soggetti possono essere raccolti in due gruppi ben distinti: quelli coscienti delle cifre durante la ripetizione e quelli che non lo sono. Tra i soggetti coscienti si osserva che meno del 10% ha pensieri intrusivi. Al contrario, i soggetti capaci di ripetere la sequenza di cifre senza esserne coscienti hanno pensieri indipendenti dalla volontà per circa il 60% del periodo totale. Pertanto, con tale compito distraente, si osserva una riduzione pari al 20% dei pensieri intrusivi rispetto ai soggetti semplicemente posti in un luogo silenzioso. Da questi risultati emerge il ruolo fondamentale che la consapevolezza delle cifre assume nella riduzione (di ben sei volte) della frequenza dei pensieri intrusivi, rispetto a quella osservata senza lo svolgimento di un compito o con la ripetizione automatica.
Considerando due codici distinti α e β questi dati sono in accordo con un modello del processo costituito da due coppie di moduli (PMα/WMα) e (PMβ/WMβ), ciascuna associata a una memoria episodica EM e al sistema TRC per la competizione globale. Quando una valenza affettiva è associata alla memoria EM il modello suggerisce che i pensieri intrusivi vengano prodotti in relazione a spiacevoli memorie passate e a bassi livelli di auto stima. Lo sviluppo di pensieri intrusivi può coinvolgere circuiti fortemente autoeccitatori e dipendere da un eccesso di rumore, come nel caso delle voci esterne dei pazienti schizofrenici. Il quesito cruciale è in che modo i pensieri intrusivi possono trasformarsi in sensazioni coscienti. Quando gli stimoli esterni, trattati nelle memorie semantiche PMα e PMβ, si riducono, i frammenti o le sequenze d'informazioni generati internamente dalla memoria episodica EM possono assumere il controllo dell' attività delle memorie di lavoro (WMα e WMβ). In questo modello, come riportato dal principio espresso nel punto 3, è attesa una riduzione del livello dei pensieri intrusivi ogniqualvolta una delle memorie di lavoro vince la competizione, come avviene nel caso della consapevolezza delle cifre durante il compito di distrazione. Tale riduzione dei pensieri intrusivi non può aver luogo se le cifre sono esaminate automaticamente, poiché, in questo caso, gli stimoli esterni generano un'attività afferente al modulo di analisi preliminare semantico PMα che passa direttamente alla risposta motoria, senza consapevolezza (v. figura 1). Il modello produce risultati in accordo con quelli di Baddeley (1993). Inoltre, nel caso del processo automatico, la riduzione di circa il 20% dei pensieri intrusivi può essere spiegata da un'inibizione generale che parte dalla risposta stessa, in questo caso la ripetizione delle cifre, e procede retro attivamente verso la memoria episodica EM e il sistema a essa collegato. È importante notare che questo risultato costituisce anche una conferma al principio descritto dal punto 8. Si noti, infine, che l'uso di un compito di distrazione visivo-spaziale, come la pressione di una determinata sequenza di tasti, provoca una riduzione dei pensieri intrusivi dall'80% al 30%.
False memorie
Il principio espresso nel punto 4 è fondamentale per il modello di mente relazionale, ma l'analisi dei pensieri intrusivi fornisce solo indirettamente elementi a suo favore. Una più forte e diretta conferma a questo principio proviene dai recenti lavori di H.L. Roediger e K.B. McDermott (Roediger e McDermott, 1995). Questi ricercatori hanno studiato la costruzione di false memorie, con esperimenti in cui i soggetti forniscono risposte sbagliate a proposito di stimoli con i quali in realtà non sono stati posti a confronto. Un tale processo mentale può essere considerato memoria ricostruttiva, cioè una complementazione attiva degli elementi mancanti di un ricordo, in contrasto con il ricordo puro, ossia l'accurata riproduzione del materiale memorizzato. In questi esperimenti (Roediger e McDermott, 1995) i soggetti ascoltavano e ripetevano elenchi di 12 parole, ognuna era collegata a uno stesso termine con il ruolo di esca per lo scopo prefissato. In un esperimento tipo, ai soggetti veniva consegnato un insieme di 6 elenchi di 12 parole, ognuno collegato a parole esca differenti. Dapprima, gli elenchi erano letti ad alta voce e a ogni soggetto veniva richiesta una rievocazione libera delle parole degli elenchi immediatamente dopo la loro presentazione. Dopo alcuni minuti, veniva presentato un elenco di 42 parole, comprendente le 12 appartenenti a una delle liste studiate precedentemente e 30 parole nuove. Il risultato più impressionante è stato che, per il 40% delle volte, nella rievocazione libera veniva richiamata la parola usata come esca, cioè la parola associata alla lista ma che in realtà non era presentata. Se confrontata con il 14% delle volte in cui veniva introdotta una nuova parola a caso, la percentuale di risposta indica che i soggetti non provavano affatto a indovinare a caso. Se interrogati, i soggetti si dichiaravano sicuri che le parole nuove dell'elenco di riconoscimento apparivano almeno per la metà delle volte negli elenchi presentati originariamente.
La domanda fondamentale sollevata da questo esperimento è quale sia il meccanismo che permette al soggetto di ricostruire coscientemente un supposto ricordo episodico, verosimilmente innescato dall'attivazione di associazioni implicite tra le parole dell'elenco e la parola associata con ruolo di esca. Il ricordo della parola nuova risultava maggiore quando questa era proferita immediatamente dopo la prima lettura del test (probabilità di 0,58 contro 0,38 rispettivamente). Questo dato può essere interpretato come un'indicazione che c'è un 20% addizionale di probabilità in cui la parola nuova è ricordata liberamente, in modo implicito, per poi restare nella memoria episodica ed essere usata nel test di riconoscimento come base del ricordo. Durante l'esame della lista di riconoscimento la risposta e l'esperienza cosciente associata risultano quindi modificate dal ricordo della parola nuova e da quello implicito, inesatto, delle esche, mediante l'intervento delle memorie di analisi preliminare semantica o della memoria episodica (Taylor, 1996). In particolare è importante notare che il coinvolgimento della memoria episodica, formatasi per ultima (quando il soggetto ricorda a memoria subito dopo la lettura), interviene nell'insorgere della coscienza, come proposto dal principio di cui al punto 4.
Processi automatici
l dati degli esperimenti precedenti sulle liste di parole (Roediger e McDermott, 1995) confermano anche l'ipotesi dell'esistenza di un aggiornamento delle memorie episodiche in funzione dell'attività della memoria di lavoro vincitrice. Questo principio, espresso nel punto 5, evidenzia il modo in cui le memorie episodiche e di lavoro svolgono un'azione eccitatoria reciproca e sempre attivata. Verranno ora esaminati i principi definiti nei punti 6, 7 e 8, relativi al modo in cui un processo automatico può svolgersi. Per chiarire a quale tipo di processo automatico ci stiamo riferendo, consideriamo, per esempio, quello che succede quando ci poniamo alla guida di un'automobile: normalmente non siamo coscienti di tutto ciò che i nostri sensi ci trasmettono negli istanti immediatamente precedenti al presente. È tuttavia possibile dimostrare che esiste una consapevolezza di queste informazioni e che la loro memoria viene mantenuta per circa 30 s. Sembrerebbe che l'attività della memoria di lavoro vincente non sia salvaguardata nella corrispondente memoria di analisi preliminare. Questa salvaguardia costituisce una fase necessaria per l'immagazzinamento nella memoria a lungo termine, la quale può quindi fornire informazioni accessibili in tempi successivi, ben oltre il decadimento della memoria a breve termine.
Consideriamo il modo in cui le informazioni richiamabili dalla memoria possono essere usate durante il sogno, in assenza di stimoli sensoriali periferici del sistema nervoso. In termini puramente elettro fisiologici, lo stato di veglia e lo stato di sonno paradossale, caratterizzato da rapidi movimenti oculari, rappresentano entrambi uno stato di attivazione molto simile, con depolarizzazione delle cellule talamo-corticali e attivazione del sistema TRC. Ci sono quindi i presupposti per una competizione tra i moduli di memorie di lavoro che consenta soltanto al modulo vincente di raggiungere la coscienza. Durante questo tipo di sonno, tuttavia, le afferenze sensoriali sono profondamente alterate. Diversi studi sugli effetti della privazione del sonno e sul ricordo dei sogni, indicano che il sonno paradossale è associato a una particolare percezione e sensazione cosciente. Al contrario le altre fasi del sonno, caratterizzate nell' elettroencefalogramma da onde lente, sarebbero più vagamente associate a una sensazione di consapevolezza (ovvero di qualcosa che sta accadendo nella mente ma non descrivibile in termini precisi). L'assenza di attivazione sensoriale durante il sonno paradossale conduce a ipotizzare il coinvolgimento dei moduli di analisi preliminare e semantica PM nell'analisi di un'informazione di origine interna. In un modello di formazione dei sogni legati a una esperienza cosciente, devono essere le memorie a lungo termine ad attivare il sistema, provvedendo ad alimentarlo con gli elementi necessari all'aggiornamento della memoria episodica (Bosinelli, 1995). In questo caso tale aggiornamento della memoria si svolge in differita, in accordo al principio espresso dal punto 9.
Un ulteriore insieme di risultati sperimentali a sostegno del nostro modello è stato fornito da Marcel nel suo studio sullo slittamento dell'unità della coscienza (Marcel, 1993). l suoi esperimenti hanno dimostrato come stimoli, molto disturbati o di bassa intensità, possano essere riportati nell' esperienza cosciente in diverse modalità sensoriali. Questi esperimenti forniscono anche solidi argomenti a favore del principio espresso nel punto 10. Ai soggetti era richiesto di indovinare la presenza o meno di una luce fioca. Il livello dell'illuminazione era tale che soltanto nella metà dei tentativi i soggetti riuscivano a indovinare correttamente quando la luce fosse accesa. l test più importanti consistevano in serie di prove in cui la luce veniva accesa per il 50% delle volte in modo casuale; si chiedeva quindi ai soggetti di rispondere simultaneamente in tre modi diversi alla domanda se essi avvertivano o meno la presenza di una luce: strizzando l'occhio destro, spingendo un bottone sotto l'indice destro e dicendo "sì!". Se il compito richiesto era riferire, il più velocemente e accuratamente possibile, la sensazione cosciente di aver avvertito la luce, si osservava una dissociazione simultanea: in una stessa prova si poteva osservare una risposta affermativa con una modalità di risposta, ma non con le altre. Nelle successive serie di prove, cambiando la proporzione di prove con la luce accesa, si osservavano diversi livelli significativi di successo. Abbandonando il presupposto di una sensazione cosciente unitaria, Marcel conclude "se è così, dobbiamo concludere che o l'esperienza fenomenologica della luce viene rappresentata in modo molteplice oppure esistono autocoscienze separate, che hanno accesso alle singole esperienze in modo differenziato" (Marcel, 1993).
Il modello di mente relazionale suggerisce che l'attività risultante nel modulo WMwin sia trasmessa agli altri moduli WM. L'attività nel modulo WMwin, che in quest'ultimo esperimento potrebbe corrispondere allo schema percettivo visivo-spaziale, potrebbe essere messa in una memoria 'tampone' a livello dell'ippocampo, secondo il principio esposto nel punto 6 e contribuire ugualmente a trasmettere le informazioni afferenti al circuito di sintesi fonetica per verbalizzare la risposta. Il modello è compatibile con il fatto che la trasmissione tra moduli WM sia disturbata dal rumore, in accordo con gli esperimenti di Marcel. È interessante notare quanto i risultati sui tempi di reazione rafforzino in modo sostanziale il modello che abbiamo elaborato. In effetti la risposta più rapida è la modalità visiva con la strizzatina dell'occhio (circa 315 ms), seguita dalla risposta con le dita (circa 580 ms) e dalla risposta verbale (850 ms). Questo scaglionamento delle risposte era prevedibile, se si considera che la trasmissione delle informazioni parte dall'area visiva frontale della corteccia, si dirige verso l'area motoria (rappresentante la superficie corporea), e quindi verso i circuiti fonologici (per la sintesi fonetica e la verbalizzazione). Se lo stimolo esterno fosse emesso in un'altra modalità, per esempio mediante il tatto o l'udito, è prevedibile che il tasso di errore e lo scaglionamento temporale dei vari tipi di risposta risulterebbero diversi da quelli osservati usando la luce fioca. Tutto questo in pieno accordo con il principio espresso dal punto 10.
Coscienza attiva
Cerchiamo ora di applicare il modello di mente relazionaIe alla coscienza anteriore o attiva Ca. Per fare questo è necessario considerare le versioni frontali (nel senso che ci riferiamo al lobo frontale del cervello) del processo di evocazione e miscelamento che sostengono Ca e consentire così la formulazione di un insieme di principi analoghi per Ca come è stato fatto per Cp. l siti dell' evocazione della memoria attivati da Ca sono di tre tipi. l siti del primo tipo si trovano nella corteccia posteriore, cioè le coppie di memoria di analisi preliminare e di memoria di lavoro, collegate a Cp.
l siti del secondo tipo sono localizzati nel sistema limbico, come le regioni dell'amigdala e dell'ippocampo, e consistono in memorie affettive, episodiche o biografiche. Infine, i siti del terzo tipo sono localizzati nelle regioni frontali vere e proprie (corteccia, gangli basali e la parte del talamo a essi associato). l siti interessati da Ca sono coinvolti principalmente nell'ordinamento sequenziale delle risposte motorie, nelle memorie attive a esse collegate, e nella pianificazione del comportamento. Potrebbe darsi che, in questo schema il cervelletto debba essere collocato tra i siti del primo tipo. Gli aspetti della pianificazione del comportamento potrebbero essere immagazzinati nel sistema limbico, sebbene anche la corteccia cingo lata frontale sembra essere una sede coinvolta in tali rappresentazioni. Inoltre, possono esistere alcune frammentazioni delle memorie autobiografiche; in questo caso i frammenti sono immagazzinati in diverse regioni del cervello, quali le aree paraippocampali e, alternativamente (o in aggiunta), parti della corteccia mediale frontale (Damasio, 1994).
Organizzazione della risposta motoria
L'identificazione di un luogo di memorizzazione dell'ordinamento sequenziale delle azioni rappresenta un problema ancora irrisolto. Secondo J. Grafman (1989) le azioni vengono memorizzate nel lobo frontale, come parzialmente confermato da recenti lavori sperimentali, anche se quest'ultimo sembra sia ritenuto predisposto più alla conoscenza manageriale che al contenuto specifico di sequenze dettagliate. Inoltre, non c'è alcuna indicazione sufficiente sulla natura delle rappresentazioni della memoria attiva. Tuttavia si sono ottenuti importanti riscontri sperimentali che sostengono l'esistenza di un codice nell'attività combinata (vettore risultante) di popolazioni di cellule nervose nelle diverse aree della corteccia motoria, utilizzato per pianificare, nello spazio tridimensionale, la direzione della risposta motoria. Tale codice fornisce inoltre un meccanismo che consente alle regioni di memoria attiva della corteccia frontale di essere usate come lavagne. Mantenendo questa metafora, l'informazione può essere scritta sulle lavagne dalla stessa attività afferente che provoca l'attivazione ciclica talamocorticale e intracorticale. Mediante le connessioni laterali l'informazione scritta sulla lavagna permette di selezionare rapidamente gli attrattori, ossia i vettori ottimali. Pertanto, una modificazione delle soglie d'attivazione dell'attività afferente, modulata dall' attività dei gangli basali, permette di orientare o cambiare gli attrattori.
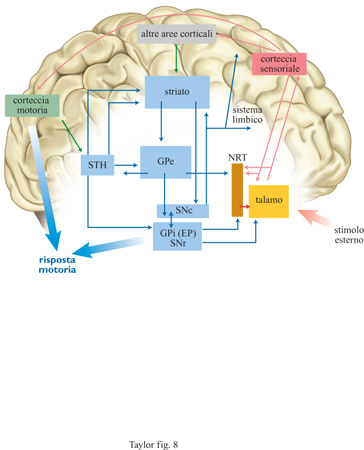
Attualmente, il complesso circuito formato dai gangli basali e dai loro arrangiamenti sinaptici è un importante oggetto di studio (Parent e Hazrati, 1995). Un diagramma delle relazioni dei gangli basali con il sistema TRC è illustrato nella figura (fig. 8). A tale proposito esistono prove sperimentali sia a favore della confluenza dell'informazione verso un unico canale che a favore dell'esistenza di vari canali di informazione che funzionano in parallelo. Tuttavia, come è stato detto da A. Parent e L.N. Hazrati "la possibilità di una diminuzione dell'inibizione esercitata sulle cellule talamiche dal nucleo reticolare del talamo potrebbe costituire un elemento di grande importanza per capire l'organizzazione funzionale dei gangli basali, giacché per iniziare un movimento è essenziale, a livello del talamo motorio, la soppressione dell'inibizione delle cellule talamocorticali, altrimenti mantenute dalle efferenze dei nuclei basali, durante lo stato di riposo, in uno stato di inibizione tonica" (Parent e Hazrati, 1995).
La rete neurale action
Un modello di rete neurale dei gangli basali che agiscono disinibendo i circuiti TRC (con annesse connessioni laterali) è la rete denominata action (Alavi e Taylor, 1996). Questa rete permette di selezionare un attrattore di attività in funzione delle afferenze esterne, mediante la scelta selettiva dei parametri di entrata che definiscono uno spazio multidimensionale di dimensione elevata. Questo procedimento allarga la codifica bidimensionale della direzione della risposta a tutti i circuiti riverberanti del sistema frontale della corteccia motoria. Attualmente è di particolare interesse lo studio di modifiche delle connessioni tra i nodi della rete neurale artificiale tali che l'insieme delle afferenze esterne, legate a un attrattore, possa scrivere sulla lavagna della corteccia frontale le informazioni provenienti dalle memorie di lavoro (in particolare quelle situate nella parte posteriore del cervello). Queste connessioni consentono la scrittura sulle memorie attive del materiale relativo a Cp, delle informazioni dei moduli di memoria EM, memorie autobiografiche o episodiche collegate a questo materiale, e delle altre memorie immagazzinate nell'ippocampo.
Anche il processo di miscelamento indispensabile per l'insorgenza della coscienza attiva deriva da un processo competitivo. È stato già suggerito che tale competizione possa essere simulata dalla rete neurale action, e secondo questo modello i nuclei inibitori dei gangli basali (il nucleo striato e il pallido) hanno un ruolo simile a quello dell'NR T nel sistema TRC. In questo caso la competizione coinvolgerebbe meccanismi di variazione della soglia di attivazione, come descritto a proposito dell'azione disinibitoria esercitata dallo striato sul talamo. Esistono diverse varianti della rete action corrispondenti ai grandi circuiti frontali che coinvolgono la corteccia motoria, premotoria e motoria supplementare (per l'ordinamento sequenziale di gesti e azioni), la corteccia limbica (per l'affettività e le emozioni), la corteccia orbitofrontale (per il comportamento sociale), l' area visiva frontale (per i movimenti oculari) e la corteccia prefrontale dorsolaterale (che interviene nei processi cognitivi e di attenzione). Come confermato dagli studi di neuroanatomia, ognuna di queste regioni corticali è dotata di opportune connessioni per ricevere le afferenze dalle aree posteriori e limbiche. Il richiamo di memorie episodiche deve essere, nel suo insieme, dipendente da memorie passate che diano all'esperienza il colore della coscienza (sia essa Cp o Ca).
Un modello basato sulla complementarità funzionale dell'informazione (cioè la possibilità di completare un'informazione parziale in funzione di un'attività risultante ben specifica) o sulle reti legate agli attrattori (reti attrattive) non costituisce un'ipotesi ottimale perché, alla fine, si genererebbe un solo profilo di attività alla volta. La struttura di memoria più opportuna sembra essere una matrice di memoria associativa basata su una rete neurale artificiale pro attiva, nella quale un dato profilo di attività afferente permette di generare un numero di attività finali corrispondente a quello dei profili di attività precedentemente memorizzati, che siano largamente sovrapponibili con l'attività afferente. D'altra parte l'uso di una rete attrattiva può aiutare a spiegare il tempo necessario per una ricerca in memoria, poiché vari attrattori vengono attivati e rifiutati dopo il confronto con un riferimento attivo che coinvolge i lobi frontali. Alcuni circuiti elettronici basati sulla rete action sono stati realizzati per simulare i circuiti dei lobi frontali e sono stati applicati allo studio degli effetti di deficit della rete quando questa è coinvolta nello svolgimento di compiti temporali (Weinberger et al., 1996). È interessante notare che i risultati, ottenuti studiando le lesioni delle reti artificiali, hanno mostrato molte analogie con la perdita della capacità di analisi dell'informazione che presentano i pazienti affetti da sindrome frontale, schizofrenia e altri disturbi mentali collegati a tali patologie.
Predire per agire meglio
Dopo aver costruito alcuni modelli di possibili meccanismi neuronali della coscienza attiva, esaminiamo, più da vicino, il loro funzionamento. In particolare, è importante fornire alcune nozioni su come si possano ottenere i processi frontali di attenzione, apprendimento e selezione di schemi logici, predizione, pensiero e pianificazione. La rete neurale action suddivide l'attenzione in specifici sottoprocessi di confronto, attivazione o inibizione e di controllo, così da rendere possibile imparare e generare le sequenze, come avviene nell' apprendimento e nella produzione di schemi logici (Alavi e Taylor, 1996). La generazione di sequenze coinvolge anche un'altra fondamentale attività frontale, la predizione, dove l'attività dell'ippocampo svolge un ruolo importante. J.A. Gray (1995) ha suggerito che nel cervello esiste un comparatore che a ogni istante è in grado di paragonare lo stato attuale del mondo percettivo con uno stato predetto. Il risultato di tale operazione di paragone corrisponderebbe al contenuto cosciente e sarebbe trasmesso nel cervello, secondo Gray, grazie alle peculiarità dei circuiti ippocampali.
Il pensiero, in quanto attività frontale, trasforma in azione le rappresentazioni iscritte nell' attività della corteccia prefrontale in modo da renderle confrontabili e più simili alle mete prefissate, immagazzinate nel sistema limbico. lnfine, l'attività di pianificazione utilizza tecniche analoghe per trovare quelle trasformazioni che da uno specifico stato di partenza permettano di arrivare a una specifica meta. Si noti la differenza tra pensiero e pianificazione: il primo usa ogni possibile trasformazione per poter raggiungere la meta, ed è quindi la successione di stati che risulta fondamentale, mentre la seconda enfatizza l'insieme delle trasformazioni e non gli stati stessi. Nel nostro modello queste trasformazioni forniscono un segnale globale di controllo, di tipo affettivo, che il sistema limbico genera in modo sempre maggiore al crescere della differenza tra lo stato desiderato, la meta, e lo stato attuale. Questo controllo globale, indirizzato verso i siti corti cali della coscienza anteriore Ca e posteriore Cp, può influenzare immediatamente il comportamento in atto. Questo segnale di controllo può risultare cruciale per il cambiamento del piano d'azione così come ha un effetto fondamentale nella motivazione e nell'intelligenza. Recenti riscontri sperimentali hanno dimostrato che è effettivamente possibile osservare una particolare attività elettrofisiologica in relazione a scelte predeterminate dal soggetto (Villa et al., 1999).
Interdipendenza tra Ca e Cp
Vogliamo qui sottolineare che la visione neuroeuristica della coscienza non ci porta ad analizzare il dettaglio, ma i principi coinvolti. Con ciò vogliamo affermare che numerose importanti funzioni svolte dai lobi frontali possono essere svolte da strutture neurali artificiali, costruite sulla base delle informazioni raccolte dagli studi di neuroscienze, senza peraltro avere la pretesa di riprodurre esattamente ciò che il cervello compie. Si capisce che stiamo proponendo un programma di lavoro per il futuro, ma torniamo ora al nostro quesito di partenza: come fa il modello ad adeguarsi, sia a livello strutturale che funzionale, a entrambe le coscienze, Cp e Ca? All'inizio abbiamo indicato la necessità di costruire le strutture neurali appropriate e di identificare le caratteristiche di Cp e Ca. È ora giunto il momento di affrontare questa questione fondamentale.
Lo schema tracciato nella figura (v. figura 1) potrebbe lasciar intendere che Cp e Ca siano indipendenti, ma abbiamo visto che ciò non può essere vero, dato che Ca sembrerebbe dipendere dall'attività risultante in Cp. Senza l'ausilio di Ca è stato possibile sviluppare un modello per Cp e per alcune delle sue caratteristiche più importanti, considerando soltanto l'aspetto fenomenologico della coscienza. Abbiamo descritto il modo in cui la manifestazione di particolari aspetti di Cp dipende da appropriate memorie di lavoro WM. In alcune situazioni passive, l'attività proveniente da Cp può essere trasferita a Ca, pertanto la coscienza attiva può essere considerata come uno stato mentale non indipendente. D'altro canto può esserci indipendenza quando Ca si sviluppa attraverso le attività generate al suo interno, come nel pensiero o nella pianificazione. Anche in questo caso, comunque, il contenuto di Ca deve provenire dalle strutture usate da Cp, per esempio le memorie di analisi preliminare e semantica PM, dalle memorie di lavoro WM a esse associate dopo lo specifico procedimento di analisi, e da Cp stessa come parte fondamentale dell'attività globale nella mente. Tuttavia, quando i processi creativi si svolgono a livello subconscio, l'attivazione dei moduli di memoria per le analisi preliminare e semantica può anche non provocare la sensazione cosciente Cp. Più in generale, l'interdipendenza tra Ca e Cp sembra essere di una complessità tale da non poter essere facilmente descritta in termini matematici o logici. Per questo motivo non ci sembra possibile riuscire a elencare principi relativi a Ca analoghi a quelli precedentemente enunciati per la sensazione cosciente Cp. Inoltre, questa interdipendenza ha la conseguenza importante di non ammettere generalizzazioni di questi principi che non tengano conto dei cambiamenti risultanti dalle relazioni reciproche.
Invece di tentare di ride finire i principi della sensazione cosciente e di cercarne per la coscienza attiva, esamineremo solo due questioni principali. In primo luogo si consideri il valore aggiunto di Cp derivato dall'insorgere di Ca. La sensazione cosciente, così come l'attività evocata dalle memorie di analisi preliminare, possono rifornire le memorie attive collegate a Ca. Queste informazioni, trattate soltanto passivamente da Cp, possono adesso essere elaborate attivamente da Ca, come è necessario nella pianificazione, nel pensiero e nelle altre operazioni mentali. La seconda questione riguarda la precisazione della relazione tra Cp e Ca. Possiamo ritenere che i siti di memoria attiva partecipino alla competizione globale innescata dal sistema TRC? Invero sembra che sia così, dato che tutti i nuclei talamici non limbici, compresi quelli coinvolti nei circuiti chiusi con la corteccia frontale, sono collegati all'NRT in modo analogo ai nuclei sensoriali del talamo posteriore. Ciononostante, la corteccia frontale, considerata l'esistenza di onde di attivazione che si propagano dalle regioni anteriori verso quelle posteriori, ha probabilmente un legame, ancora mal definito dagli studi anatomici, con le aree posteriori (Llinas e Ribary, 1992). Soltanto i nuclei limbici sono slegati dall'NRT, nel senso che, pur proiettando si con fibre collaterali verso l'NRT, non ricevono nessuna proiezione di ritorno.
Da quanto esposto sopra, possiamo concludere che esiste un'interrelazione tra la sensazione cosciente e la coscienza attiva che le rende componenti del sistema TRC in senso lato, con i segnali affettivi e le emozioni che si scaricano su di loro, pur senza alcuna influenza reciproca diretta e, quindi, con scarso controllo da parte dei moduli corticali. D'altronde sappiamo che l'esistenza di un preciso accoppiamento tra i siti di immagazzinamento delle memorie attive anteriori e quelli delle memorie di lavoro posteriori, fornisce la base per un'attività combinata anteriore e posteriore che contribuisce all'insorgere della coscienza attiva. La coscienza globale diventa quindi una complessa funzione di Cp in quanto essa dipende sia direttamente da Cp sia da ogni carattere fenomenologico di Ca che nasce da Cp e da ulteriori operazioni legate ai processi di trasformazione dell'informazione che si svolgono intrinsecamente a livello frontale. Questo contenuto addizionale esprime a sua volta un carattere fenomenologico poiché, come detto sopra, nasce nel sistema TRC come parte della competizione globale, con modalità già discusse precedentemente. Il modello di mente relazionale può essere testato con predizioni sperimentali, analizzando le diverse sequenze temporali di attività che dovrebbero esistere a livello del talamo, e le altrettanto precise relazioni temporali per l'innesco dell'attività nelle relative memorie di lavoro da parte di stimoli generati nella periferia sensoriale, nel talamo stesso o a livello della corteccia cerebrale. Anche particolari situazioni neuropatologiche, quali la sindrome di negligenza unilaterale e la cosiddetta visione cieca (blind sight), sono espressioni fenomeno logiche che possono essere testate. A partire dagli anni Novanta la possibilità di usufruire di tecniche analitiche computerizzate e non invasive offre l'opportunità di fare esperimenti sull'uomo sempre più mirati allo studio del corretto funzionamento dei circuiti cerebrali, con l'obiettivo di scoprire in futuro l'esatta natura della coscienza, utilizzando modelli simili a quello proposto in questo saggio.
Conclusioni
Usando una metafora potremmo dire che, grazie alla retroazione e alla sinergia tra congetture e sperimentazioni, la visione neuroeuristica della mente permette di osservare i risultati sperimentali oltrepassando il muro che circonda le ipotesi provvisorie, così come un bambino osserva ciò che accade attorno a sé mentre gioca in un giardino recintato da una siepe, da un cancello o da una grata. Questa metafora serve anche a indicare il carattere, talvolta passionale, che deve avere l'atto della scoperta scientifica, come insegna la massima latina [udendo docere. La ricerca scientifica rigorosamente frnalizzata a uno scopo utilitario e indirizzata in una prospettiva prestabilita, nuoce alla creatività scientifica. Come osservato dal neurobiologo A. Hodgkin a proposito del clima che circonda la ricerca scientifica "i risultati che si ottengono durante una ricerca spesso si conciliano più con la ragione a continuare o cessare il lavoro, che con l'idea che ha dato l'impulso ai primi esperimenti. Nella stesura degli articoli scientifici gli autori sono incoraggiati a essere logici e, anche se volessero ammettere che un esperimento prodotto secondo la logica è stato eseguito per una ragione assolutamente poco giustificabile, se non casuale, essi non sarebbero certo invitati a ingombrare la letteratura scientifica specializzata con reminiscenze personali irrilevanti".
Dall'Era neolitica, allorché le prime pietre di silice furono intagliate dall'uomo, gli utensili hanno preso forme diverse, ma l'interazione che essi hanno avuto con le grandi scoperte è rimasta essenzialmente la stessa. Metodi di ricerca senza precedenti nella storia stanno per essere adottati nelle neuroscienze, grazie alle moderne tecnologie informatiche. Tuttavia, la complessità dei problemi che si pongono oggi al ricercatore è di tale entità che un'analisi, anche se basata su amplissime banche dati, non può essere limitata a un'unica dimensione disciplinare. Inaugurando un nuovo modo di affrontare il problema della coscienza, la visione neuroeuristica della mente cerca di utilizzare un paradigma fondato sulla reciproca sinergia tra la creazione di modelli e gli esperimenti, parallelamente alla strada tracciata dalla sinergia tra informatica e neuroscienze, che va sempre più affermandosi.
Bibliografia citata
ALAVI, P., TAYLOR, J.G. (1996) A basis for long range inhibition across cortex. In Lateral interactions in the cortex: structure and function, a c. di Sirosh J., Miikkulainen R., Choe Y., Hypertext Book ISBN.
ATKINSON, R.C., SHIFFRIN, R.M. (1971) The control ofshort-term memory. Sci. Am., 225, 82-90.
BADDELEY, A. (1993) Working memory and conscious awareness. In Theories of memory, a c. di Conway M.A., Morris P.E., Hillsdale, N.J., Erlbaum Associates, pp. 11-28.
BIEDERMAN, I. (1993) Geon theory as an account of shape recognition in mind and brain. Irish Journal of Psychology, 14, 314-327.
BLOCK, N. (1995) On a confusion about a function of consciousnesso Behaviour and Brain Sciences, 18, 227-287.
BRAITENBERG, V., SCHlÜZ, A. (1991) Anatomy of the cortex: statistics and geometry. Berlino, Springer-Verlag.
CELLETTI, A., VILLA, A.E. (1996) Low dimensional chaotic attractors in the rat brain. BioI. Cybern., 74, 387-394.
COHEN, D.S., MURRAY, J. (1981) A generalized diffusion model for growth and dispersal in a population. Journal of Mathematical Biology, 12, 237-249.
DAMASIO, AR. (1994) Descartes' error: emotion, reason, and the human brain. New York, Avon Books.
DAVIDSON, D. (1970) Mental events. In Experience and Theory, a C. di Foster L., Swanson l, Duckworth. Amberst, University of Massachusetts Press.
DEMPSEY, E.W., MORISON, R.S. (1942) The electrical activity of a thalamo-cortical relay system. Amer. J. PhysioI., 138, 283-289.
DIAMOND, M.C., SCHEIBEL, A.B., MURPHY, G.M. JR, HARVEY, T. (1985) On the brain of a scientist: Albert Einstein. Exp. NeuroI., 88, 198-204.
ERMENTROUT, G.B., COWAN, J.D. (1978) Studies in mathematics. The Math. Assoc. of America, 15, 67-117. GRAFMAN, J. (1989) Plans, actions and mental sets. In Integrating theory and practice in clinical neuropsychology, a c. di Perecman E., Hillsdale, N.J., Erlbaum Associates.
GRAY, J.A. (1995) The contents of consciousness: a neurophysiological conjecture. Behaviour and Brain Sciences, 18, 659-676.
HAMEROFF, S., KASZNIAK, A, SCOTT, A., a c. di (1996) Towards a science of consciousness. Cambridge, Mass., MIT Press.
HILL, S.L., VILLA, A.E.P. (1997) Dynamic transitions in global network activity influenced by the balance of excitation and inhibition, Network in Computation in Neural System, 8, 165-184.
HUME, D. (1896) A treatise on human nature, a c. di Selby-Briggs E.A., Kensington, Clarendon Press.
LABERGE, D. (1990) Thalamic and cortical mechanisms of attention suggested by recent positron emission tomographic experiments. J. Cognitive Neuroscience, 2, 358-373.
LABERGE, D., CARTER, M., BROWN, V. (1992) A network similation ofthalamic circuit operations in selective attention. Neural Comp., 4, 318-333.
LIBET, B. (1982) Brain stimulation in the study of neuronal functions for conscious sensory experience. Rum. Neurobiol., l, 235-242.
LIBET, B., ALBERTS, W.W., WRIGHT, E.W. JR., DELATTRE, D.L., LEVIN, G., FEINSTEIN, B. (1964) Production of threshold levels of conscious sensation by electrical stimulation of human somato-sensory cortex. J. NeurophysioI., 27, 546-578.
LLINAS, R., RiBARY, U. (1992) In Induced rhythms in the brain, a c. di Basar E., Bullock T., Boston, Birkhauser.
MARCEL, A.J. (1980) Consciousness and preconscious recognition of polysemous words. In Attention and Performance VIII, a c. di Nickerson R.S., Hillsdale, N.J., Erlbaum Associates.
MARCEL, A.J. (1993) Slippage in the unity of consciousness. In Experimental and theoretical studies in consciousness, Chichester, New York, Wiley (Ciba Foundation Simposium n. 174).
NAGEL, T. (1974) What is it like to be a Bat? Philosophical Reviews, 83, 435-450.
PARENT, A., HAZRATI, L.N. (1995) Functional anatomy of the basaI ganglia. I. The cortico-basal ganglia-thalamo-cortical loop. Brain Res. Rev., 20, 91-127.
PAULESU, E., FRITH, C.D., FRACKOWIAK, R.S. (1993) The neural correlates ofthe verbal component of working memory. Nature, 362, 342-345.
PYLYSHYN, Z.W. (1986) Computation and cognition: toward a foundation for cognitive science. Cambridge, Mass., MIT Press.
RIANI, M., MASULLI, F. (1990) Modelling perceptual alternation by using ANNs. In Parallel architectures and neural networks: second Italian workshop, Vietri sul Mare, Salerno, 26-28 ApriI 1989, a c. di Caianiello E.R., Singapore, Teaneck, N.J., World Scientific.
ROEDIGER, H.L., McDERMoTT, K.B. (1995) Creating false memories; remembering words not presented in lists. J. Experimental Psychology: Learning Memory and Cognition, 21, 803-814.
SCHEIBEL, A.B. (1980) The reticular formation revisited: specifing function for a non specific system, a c. di Hobson J.A., Brazier B.A, New York, Raven Press.
SEARLE, J. (1992) The rediscovery of mind. (Representation and Mind). Cambridge, Mass., MIT Press.
SEGUNDO, J.P. (1985) Mind and matter, matter and mind? J. Theoret. NeurobioI., 4, 47-58.
STERIADE, M., GLOOR, P., LLINAS, R.R., LOPES DA SILVA, P.H., MESULAM, M.M. (1990) Report of IFCN Commitee on Basic Mechanisms. Basic mechanisms of cerebral rhythmic activities. Electroencephalogr. Clin. NeurophysioI., 76, 481-508.
STERIADE, M., LLINAS, R.R. (1988) The functional states of the thalamus and the associated neuronal interplay. PhysioI. Rev., 68, 649-742.
TAYLOR, J.G. (1991) Can neural networks ever be made to think? Neural Network World, 1,4-11.
TAYLOR, J.G. (1992) Towards a neural network model of mind. Neural Network World, 2, 797-812.
TAYLOR, J.G. (1996) Breakthrough to awareness - a preliminary neural network model of conscious and unconscious perception in word processing. BioI. Cybern., 75, 59-72.
TAYLOR, J.G., ALAVI, F. (1993) Mathematical analysis of a competitive network for attention. In Mathematical approaches to neural networks, a c. di Taylor J.C., Amsterdam-New York, North Holland, pp. 341-382.
TAYLOR, J.G., ALAVI, F.N. (1995) A global competitive neural network. Bio!. Cybern., 72, 233-248.
TETKO, I.V., VILLA, A.E. (1997) Efficient partion of learning datasets for neural network training. Neural Networks, 10, 1361-1374.
VILLA, A.E. (1990) Physiological differentiation within the auditory part ofthe thalamic reticular nucleus of the cat. Brain Res. Rev., 15, 25-40.
VILLA, A.E., BAJO LORENZANA, V.M., VANTINI, G. (1996) Nerve growth factor modulates information processing in the auditory thalamus. Brain Res. Bull., 39, 139-147.
VILLA, A.E., ROUlLLER, E.M., SIMM, G.M., ZURITA, P., DE RIBAUPIERRE, Y., DE RIBAUPIERRE, F. (1991) Corticofugal modulation of information processing in the auditory thalamus of the cat. Exp. Brain Res., 86, 506-517.
VILLA, A.E., TETKO, I.V. (1995) Spatio-temporal patterns of activity controlled by system parameters in a simulated thalamocortical neural network. In Workshop on Supercomputing in brain research: from tomography to neural networks, HLRZ, KFA Julich, Germany, November 21-23, 1994, a c. di Hermann H., Wolf D.E., Poppel E., Singapore, River Edge, N.J.,World Scientific, pp. 379-388.
VILLA, A.E., TETKO, I.V., HYLAND, B., NAJEM, A. (1999) Spatiotemporal activity patterns of rat cortical neurons predict responses in a conditioned task. Proc. Natl. Acad. Sci. USA, 96, 1006-1011.
WEINBERGER, D., ROBBINS, T.W., TAYLOR, J.G., MORRIS, R.G. (1996) Dissociating executive functions of the prefrontal cortex. Philos. Trans. R. Soc. Lond. B Biol. Sci., 351, 1470-1471.
WINER, J.A., LARUE, D.T. (1996) Evolution of GABAergic circuitry in the mammalian mediaI geniculate body. Proc. Natl. Acad. Sci. USA, 93, 3083-3087.
Bibliografia generale
BERGER, R. (1994) Du miroir à l'après-histoire. Diogène, 167, 118-141.
BOSINELLI, M. (1995) Mind and consciousness during sleep. Behav. Brain Res., 69, 195-201.
DENNET, D.C. Quining Qualia. In Consciousness in contemporary science, a c. di Marcel A.J., Bisiach E., Oxford, Clarendon Press and Oxford-New York, Oxford University Press, 1988.
JELSMA, T.N., AGUAYO, A.J. (1994) Trophic factors. Curr. Opin. Neurobiol., 4, 717-725.
LEVI MONTALCINI, R., CALISSANO, P. (1986) Nerve growth factor as a paradigm for other polypeptide growth factors. Trends in Neuroscience, 9, 473-477.
PRIGOGINE, I., STENGERS, I. La nouvelle alliance: métamorphose de la science. Parigi, Gallimard, 1979.
WIDROW, B., STEARNS, S.D. Adaptive signal processing. Englewood Cliffs, Prentice Hall, 1985.