
La seconda rivoluzione scientifica: scienze biologiche e medicina. La biologia evoluzionistica e la genetica
La seconda rivoluzione scientifica: scienze biologiche e medicina. La biologia evoluzionistica e la genetica
La biologia evoluzionistica e la genetica
L'eredità di Darwin
Alla fine del XIX sec. la teoria dell'evoluzione per selezione naturale, pubblicata contemporaneamente da Charles Darwin e Alfred R. Wallace in due brevi articoli del 1858, e in forma estesa in On the origin of species da Darwin nel 1859, ebbe un profondo impatto sulla biologia in due direzioni abbastanza diverse. Come primo effetto determinò enorme interesse e attenzione per lo studio della filogenesi animale e vegetale al fine di rintracciare le linee evolutive. Una volta che il concetto base dell'evoluzione stessa venne accettato, divenne possibile, ed enormemente popolare, ripercorrere le storie filogenetiche di numerosi gruppi di organismi. Nel periodo 1860-1910, gli scopi della filogenesi acquisirono un'importanza tale che praticamente ogni disciplina biologica, eccetto forse la fisiologia generale e la biochimica, si inchinò o venne asservita alla teoria evoluzionistica. Un secondo effetto fu la metodologia evidenziata dagli studi di Darwin. In On the origin of species Darwin scrisse di aver proceduto per induzione, mettendo insieme una grande quantità di dati (informazioni) che la teoria della selezione naturale riuscì a sintetizzare. L'idea della selezione naturale fu per Darwin una generalizzazione induttiva che diede un significato a un gran numero di fatti altrimenti sconnessi. Studi storici hanno comunque dimostrato che Darwin procedette molto meno induttivamente di quanto avesse dichiarato o, almeno, di quanto avesse scelto di enfatizzare, tuttavia l'immagine che l'approccio darwiniano divulgò al pubblico scientifico alla fine dell'Ottocento valorizzava soprattutto le grandi teorie generali, al cui interno era possibile riunire un ampio ventaglio di problemi. Darwin, nella sua opera, raccolse le prove a favore della teoria della selezione naturale da diverse aree della ricerca biologica, tra cui la sistematica, la zootecnia, l'agricoltura, la biogeografia, l'anatomia comparata, l'ecologia e l'embriologia. In tal modo ottenne il sostegno necessario alla sua idea generale dell'evoluzione per selezione naturale; dimostrò inoltre la validità di ogni teoria che fosse in grado di includere dati provenienti da aree biologiche così diverse. Fu il trionfo del metodo della concordanza di William Whewell, in cui i dati raccolti indipendentemente da due o più fonti si supportano l'uno con l'altro. Questo retaggio fu talmente importante da condizionare le numerose teorie dell'eredità, dello sviluppo e dell'evoluzione che sorsero verso la fine del secolo.
Negli anni Novanta dell'Ottocento, la teoria darwiniana stava però incontrando numerosi problemi. Dal punto di vista del contenuto, il principale problema era costituito dall'asserzione di Darwin, senza alcuna prova diretta, che le piccole variazioni ereditate si manifestano di quando in quando in una popolazione e sono in effetti la materia prima su cui agisce la selezione (negativamente o positivamente). L'intero meccanismo di evoluzione per selezione naturale si basava infatti su questa affermazione, perché se le piccole differenze individuali non fossero ereditabili, la selezione pro o contro di esse non produrrebbe modificazioni nella generazione successiva e quindi non si avrebbe evoluzione. Di conseguenza, la natura delle variazioni (la loro origine e l'ereditarietà) divenne oggetto di considerevoli preoccupazioni nel periodo postdarwiniano.
Il principale problema metodologico dell'opera di Darwin era costituito dal meccanismo della selezione naturale, all'epoca ancora non passibile di verifica sperimentale. Mentre Darwin stesso fece riferimento a esperimenti di selezione artificiale (condotti da allevatori e agricoltori), nondimeno utilizzò questi esperimenti solo come modello (per analogia) per la selezione naturale; la teoria darwiniana sembrava dunque limitata all'inferenza logica a partire dalle osservazioni.
L'estensione e la profondità dell'influenza di Darwin possono essere apprezzate meglio se si considera l'importanza assunta dalla morfologia nel periodo 1860-1900. Il cosiddetto studio della forma comprendeva un gruppo di discipline oggi considerate indipendenti: l'anatomia comparata, l'embriologia, la paleontologia, la citologia e in qualche misura l'ecologia. Sebbene come branca della biologia la morfologia avesse già una lunga storia, con diversi scopi e metodologie, nel periodo postdarwiniano pose la sua attenzione principalmente alla determinazione delle relazioni filogenetiche tra gruppi di organismi (dalle specie alle famiglie e ai phyla) per mezzo della comparazione delle strutture omologhe nelle forme adulte ed embrionali (per es., strutture che derivano da un progenitore ancestrale comune che però nel tempo potrebbero aver subito modifiche per l'adattamento a differenti funzioni). Determinando le caratteristiche veramente omologhe (per es., le appendici frontali dei vertebrati), sarebbe stato possibile inferire le relazioni evolutive dalle prove anatomiche ed embriologiche disponibili.
Uno dei più noti tentativi di formulare ricostruzioni filogenetiche su larga scala a partire dall'anatomia comparata e dall'embriologia era stato proposto dal morfologo tedesco Ernst Heinrich Haeckel (1834-1919) nella Generelle Morphologie (Morfologia generale) del 1866. Fondata sulla sua idea di ricapitolazione o 'legge biogenetica' (l'affermazione che ogni organismo nel suo sviluppo embrionale attraversa i principali stadi adulti nella storia filogenetica della propria specie), Haeckel azzardò che i morfologi dovessero osservare solamente lo sviluppo di un organismo dall'uovo fecondato fino all'adulto per vedere la sequenza in miniatura del passato evolutivo della specie. Per spiegare come l'embriologia potesse riflettere la filogenesi, Haeckel introdusse il concetto di 'addizione terminale' nel quale le novità evolutive venivano aggiunte al termine dello sviluppo embrionale ancestrale, mentre gli stadi di sviluppo precedenti venivano compressi per far posto alle nuove addizioni. Anche se il modello di Haeckel è stato confutato da lungo tempo (attualmente è noto che gli stadi di sviluppo embrionale degli organismi rappresentano la derivazione degli stadi embrionali e non adulti dei progenitori), diede comunque impulso a un programma di ricerca che coinvolse un'intera generazione di morfologi. Il fatto che le prove a sostegno di molte ricostruzioni filogenetiche fossero circostanziali rese infine l'intera tradizione morfologica soggetta alla critica dei biologi più giovani. Tale tradizione enfatizzava comunque l'integrazione tra le teorie dell'eredità (quello che viene trasmesso dai genitori alla discendenza), lo sviluppo embrionale (in particolare il problema del differenziamento) e l'evoluzione. Per i morfologi della fine del XIX sec. i tre processi erano così intrecciati che ogni teoria capace di spiegarne uno doveva essere in grado di comprendere anche gli altri. Il termine 'eredità' alla fine del secolo comprendeva infatti i processi di trasmissione, sviluppo embrionale ed evoluzione.
L'opposizione al darwinismo
L'opera On the origin of species convinse un gran numero di naturalisti che l'evoluzione stessa ‒ ossia, la discendenza con modificazioni ‒ fosse la spiegazione più probabile per la diversità. Tuttavia, l'ipotesi avanzata da Darwin su come questa avesse avuto luogo non fu assolutamente accettata da tutti. Le critiche sollevate dai naturalisti della fine dell'Ottocento alla teoria darwiniana comprendevano: (a) incertezza sul tipo di variazioni ‒ grandi o piccole ‒ su cui la selezione potesse lavorare; (b) efficacia della selezione naturale nella produzione di specie interamente nuove; (c) scetticismo nella maniera in cui gli adattamenti ‒ e in modo particolare ciò che Darwin e i suoi contemporanei chiamavano 'organi di estrema perfezione' (come l'occhio dei vertebrati) ‒ potessero emergere da adattamenti piccoli e in gran parte casuali; (d) problemi relativi ai dati geologici e paleontologici, come la mancanza di un tempo geologico sufficiente perché l'evoluzione si verifichi mediante il lento ritmo casuale di Darwin e mostri il tasso di variazione, la mancanza di forme di transizione, l'apparente presenza di tendenze di aumento di dimensioni e di complessità dagli strati inferiori ai superiori, l'improvvisa scomparsa di forme di vita da uno strato all'altro (per es., i grandi rettili al confine tra Cretaceo e Terziario). Di conseguenza, una serie di processi evolutivi ‒ tra cui l'eredità neolamarckiana, l'ortogenesi e il mutazionismo ‒ vennero proposti a cavallo tra Ottocento e Novecento quali possibili meccanismi alternativi per l'evoluzione.
In parte prendendo spunto dalla fiducia dello stesso Darwin nel principio dell'ereditarietà degli effetti dell'uso, del disuso e dell'ambiente sulle strutture e le funzioni corporee, un diffuso movimento neolamarckiano crebbe nel periodo 1880-1920, associato ai nomi di Theodor Eimer in Germania, Paul Kammerer in Austria, Yves Delage in Francia, Edward D. Cope e Henry Fairfield Osborn negli Stati Uniti. Questa teoria, chiamata così in onore del naturalista francese Jean-Baptiste Lamarck (1744-1829), sosteneva che le modificazioni del corpo degli organismi adulti, sia anatomiche sia fisiologiche, sia ‒ in alcune versioni ‒ psicologiche, potessero essere trasmesse alle cellule germinali e da queste alla discendenza. Per esempio, gli adulti utilizzando alcuni muscoli o ossa, oppure imbattendosi in nuovi tipi di cibo, acquisiscono nuovi adattamenti individuali che possono diventare parte delle caratteristiche ereditarie della specie. Il neolamarckismo offrì aiuto ai naturalisti per aggirare il problema dell'apparente improbabilità di adattamenti altamente sviluppati nati da variazioni casuali. Poiché le variazioni sarebbero state prodotte come risposta diretta alle reali condizioni ambientali alle quali l'organismo si sarebbe dovuto adattare, il processo non avrebbe dovuto attendere le variazioni casuali per aver luogo. Questo tipo di adattamento diretto avrebbe inoltre ridotto in maniera considerevole il tempo geologico richiesto perché le nuove caratteristiche potessero essere fissate nella popolazione.
Un forte colpo alla teoria neolamarckiana venne assestato dagli studi di August Weismann (1834-1914) con la sua teoria della separazione del germoplasma e del soma, che precludeva qualsiasi influenza delle cellule dell'organismo adulto sui tessuti germinali. Nei primi anni Novanta Weismann effettuò una serie di esperimenti sui topi tagliandone sistematicamente le code per circa 18 generazioni, senza osservare alcuna riduzione della lunghezza media della coda nei discendenti. Per molti, questa prova fu decisiva, ma non pose fine alla controversia sull'eredità dei caratteri acquisiti. Sebbene nel XX sec. i sostenitori del neolamarckismo continuassero ancora a esercitare qualche influenza, specialmente con l'affermarsi in Unione Sovietica delle teorie di Trofim Denisovič Lysenko (1898 - 1976), dagli anni Trenta del Novecento la maggior parte dei biologi occidentali aveva abbandonato la dottrina dell'ereditarietà dei caratteri acquisiti. Il genetista e agronomo russo riteneva che le caratteristiche acquisite potessero essere ereditate attraverso un controllo ambientale, opponendosi in questo modo alla scuola di genetisti che al contrario le considerava innate.
Un'altra alternativa al darwinismo ortodosso fu la teoria dell'ortogenesi, che considerava la direzionalità incorporata nel processo evolutivo. Appoggiata in particolare da alcuni paleontologi tra i quali Eimer e Cope, l'ortogenesi tentava di spiegare le tendenze di lunga durata osservate nei dati fossili. Un esempio particolarmente famoso era l'evoluzione dei palchi nell'alce irlandese (un grosso cervo che era vissuto nelle zone prive di alberi in Europa e nelle isole britanniche del Pleistocene) che, si riteneva, ne avesse determinato l'estinzione. Dal momento che era stata raggiunta un'ampiezza di circa quattro metri poco prima dell'estinzione, si pensava che la grande dimensione dei palchi fosse il risultato di una sorta di 'momento inerziale' dell'evoluzione. Una volta iniziata la corsa all'ingrandimento, originariamente adattativo, la dimensione del palco era cresciuta fino a diventare un impedimento per la mobilità e l'equilibrio dell'animale; di conseguenza, il 'momento' non poté essere fermato e l'alce fu portato all'estinzione. Tendenze simili furono osservate nel numero di spire dei gasteropodi marini, nei canini della tigre dai denti a sciabola e in numerosi altri casi. Studi successivi mostrarono che spiegazioni darwiniane più convenzionali avrebbero potuto render conto di ciò che appariva come una tendenza nei dati fossili (nel caso dell'alce irlandese, per es., la sua estinzione fu molto probabilmente il risultato, durante l'ultima era glaciale, della migrazione verso sud nelle aree forestali dell'Europa centrale e occidentale, dove palchi così grandi erano decisamente svantaggiosi). Poiché il 'momento evolutivo' sembrava essere un concetto metafisico per il quale non esisteva alcun meccanismo noto, l'ortogenesi venne abbandonata dalla maggior parte dei biologi evoluzionisti a partire dai primi decenni del XX secolo.
Uno degli aspetti concettualmente più difficili della teoria della selezione naturale di Darwin era la sua affermazione che la selezione agiva principalmente su piccole differenze individuali all'interno di una popolazione, in opposizione ai cambiamenti su larga scala, noti al tempo come 'sport' o 'mostruosità'. Il problema che numerosi contemporanei ebbero con l'enfasi posta da Darwin sulla selezione di piccole differenze individuali, fu che sebbene gli agricoltori e gli allevatori avessero agito efficacemente sugli organismi grazie alla selezione artificiale di sottili differenze individuali, nondimeno non avevano mai prodotto una nuova specie con tale metodo. Quindi, si sosteneva che la selezione di queste piccole variazioni fosse in grado di produrre solamente varietà; serviva qualcos'altro per poter dare vita a una specie interamente nuova. Tra le alternative più comuni al gradualismo darwiniano vi fu la teoria della mutazione, proposta dal botanico olandese Hugo De Vries (1848-1935) in due corposi volumi del 1901 e del 1903. De Vries si basò principalmente su esperimenti effettuati sull'enagra Oenothera lamarckiana (detta anche rapunzia a grandi fiori), incrociando due ceppi e osservando nella discendenza numerose forme talmente diverse dal tipo parentale che sembravano appartenere a nuove specie. Queste nuove forme non si incrociavano con i loro progenitori, anche se in alcuni casi erano tra loro interfeconde. I risultati ottenuti convinsero De Vries di aver trovato un meccanismo che permetteva la nascita di nuove specie (invece delle varietà prodotte dai selezionatori). Le mutazioni erano cambiamenti massicci nella costituzione ereditaria dell'organismo che attraversavano i confini delle specie.
Anche se la teoria mutazionista ebbe un notevole seguito nei primi anni del secolo, nel 1915 venne scoperto che l'Oenothera è un organismo anomalo per ciò che riguarda il meccanismo riproduttivo. Le 'forme mutate' di De Vries si rivelarono essere non nuove specie bensì il risultato di un comportamento eccezionale dei loro cromosomi nel corso della formazione dei gameti. I discendenti di ogni incrocio tra due tipi rappresentano un gran numero di forme di diverso aspetto che a una prima occhiata sembrano specie nuove. Solamente quando divenne più chiaro il ruolo dei cromosomi nell'ereditarietà, si comprese che l'Oenothera è, dal punto di vista riproduttivo, un caso molto raro.
Teorie dell'eredità e dell'evoluzione alla fine del XIX secolo
Darwin riconobbe che la sua teoria dell'evoluzione aveva bisogno di un meccanismo di eredità delle variazioni permanenti e in Variation of animals and plants under domestication (1868) propose tale meccanismo ‒ la sua 'teoria provvisoria della pangenesi'. L'ipotesi suggeriva che i caratteri individuali fossero controllati da particelle chiamate 'pangeni', specifiche per i differenti tessuti (muscoli, pelle, nervi, e così via) e in essi presenti. I pangeni venivano rilasciati dai tessuti e raccolti nelle gonadi, dove si riversavano nell'uovo o nello sperma. Secondo Darwin la caratteristica importante di questa teoria consisteva nella variazione. Nel momento in cui l'organismo utilizza un certo tessuto (per es., un determinato muscolo), la produzione di pangeni in quel muscolo aumenta e viene raccolta nelle cellule germinali. La teoria della pangenesi fornì a Darwin un meccanismo plausibile, anche se successivamente screditato, per l'origine e la trasmissione ereditaria delle variazioni.
Un approccio alternativo al problema dell'eredità e della sua relazione con l'evoluzione e lo sviluppo venne proposto da Weismann, zoologo di Friburgo. Microscopista di notevole abilità, Weismann osservò in alcuni idroidi marini (Cnidaria) che alla prima segmentazione dello zigote una cellula veniva messa da parte per diventare successivamente il tessuto che nell'adulto genera l'uovo o lo sperma, la linea cellulare germinale, mentre l'altra cellula dà luogo a tutti i restanti tessuti corporei, la linea cellulare somatica. Weismann aveva concluso che in tutti gli organismi pluricellulari avviene una separazione completa dei tessuti germinali e somatici, così che eventuali cambiamenti nelle cellule somatiche non avrebbero alcun effetto sulla linea germinale. L'idea di Weismann della separazione dei tessuti germinali e somatici creò le basi per la sua strenua opposizione alle affermazioni tra cui quelle di Darwin, sull'ereditarietà degli effetti dell'uso e del disuso. Dal punto di vista evoluzionistico, le variazioni nella linea germinale avrebbero procurato i cambiamenti ereditari su cui agisce la selezione naturale; le variazioni nella linea somatica, invece, mentre in qualche modo potrebbero avere effetto sulle strutture e le funzioni dell'adulto, non possono essere ereditate e quindi non possono contribuire alla futura evoluzione della specie.
Come Haeckel, Weismann andò oltre nell'elaborazione di uno schema concettuale estremamente ambizioso, che comprendeva i processi evolutivi, ereditari e di differenziamento embrionale. Per spiegare l'eredità, Weismann ipotizzò che il nucleo cellulare avesse una gerarchia di particelle ereditarie che controllano le caratteristiche generali dell'organismo nonché i suoi tratti specifici. Queste particelle erano trasmesse alla prole attraverso l'uovo e lo sperma. Per spiegare come le cellule si diversificano durante la crescita embrionale, Weismann suggerì che queste particelle venissero distribuite successivamente a cellule differenti, in modo che alla fine ogni cellula risultasse avere solo un tipo di determinante che indirizza una cellula qualsiasi a divenire muscolo, nervo o pelle. Per spiegare l'insorgenza e l'ereditarietà delle variazioni, Weismann suggerì che la competizione per il nutrimento tra i determinanti avrebbe consentito a quelli metabolicamente più attivi di avere il sopravvento sugli altri e quindi di esprimersi, mentre i meno attivi metabolicamente sarebbero divenuti latenti. Queste erano le variazioni su cui avrebbe agito la selezione naturale. Gli studi di Weismann erano più accurati di quelli di Haeckel e in qualche modo obbligarono i biologi seri a prestargli maggiore attenzione. Comunque, come le teorie di Haeckel, le ipotesi di Weismann andarono molto oltre i dati disponibili e non furono quindi dimostrabili.
Sebbene la teoria mutazionistica fosse alla fine confutata come meccanismo di speciazione, alimentò le speranze nella possibilità di studiare sperimentalmente il processo evolutivo. I biologi più giovani, maldisposti verso le vetuste e speculative teorie morfologiche di Haeckel, Weismann e altri della loro generazione, pensavano che l'evoluzione potesse essere studiata con lo stesso rigore dei problemi di fisica e chimica. Per coincidenza, contemporaneamente a Darwin, un meccanismo verificabile empiricamente venne proposto da Gregor Mendel (che lo pubblicò nel 1866), ma rimase ignoto a Darwin e in effetti a tutta la generazione successiva di biologi. La teoria mendeliana sarebbe infine divenuta il fondamento della genetica e della teoria evoluzionistica nel XX secolo.
Gregor Mendel e la nascita della genetica
Tutte le teorie dell'eredità, dello sviluppo e dell'evoluzione avanzate nella seconda metà del XIX sec. postulavano che i caratteri venivano trasmessi alla prole per mezzo di particelle ereditarie. Nello stesso periodo, tra il 1856 e il 1863 Mendel, un monaco agostiniano di Brünn in Austria (ora Brno, Repubblica Ceca), stava conducendo esperimenti di coltivazione con il pisello comune, Pisum sativum. Il lavoro di Mendel era molto diverso da quello dei suoi contemporanei, sia concettualmente sia metodologicamente. Dal punto di vista concettuale, per quanto la teoria di Mendel fosse compatibile con le teorie particellari dell'eredità proposte negli stessi anni, non era stata formulata come una grande teoria unificata dell'eredità, dello sviluppo e dell'evoluzione. Gli interessi di Mendel erano limitati al processo di ibridazione e alla trasmissione di tratti specifici negli incroci tra varietà che presentavano differenze in una singola caratteristica ‒ per esempio, altezza (alto o basso), colore del baccello (giallo o verde). Con una formazione di insegnante di scienze naturali e matematica, Mendel importò nello studio dell'ibridazione una metodologia quantitativa e sperimentale che rifuggiva da speculazioni teoriche di vasta portata. Per quanto Mendel postulasse l'esistenza, nelle cellule dell'uovo e del polline dei suoi ibridi, di una qualche sorta di 'fattori' per ogni tratto dell'adulto, la sua enfasi era nella progettazione di esperimenti che avrebbero dovuto dimostrare come questi fattori venivano trasmessi e ricombinati nella discendenza. Per esempio, incrociando una pianta di una linea pura per il carattere 'alto' con una linea pura per il carattere 'basso', scoprì che tutta la prima generazione (F1) era di carattere 'alto'. Incrociando tra loro le piante F1, invece, trovò che su 1064 piante di seconda generazione F2, 787 erano alte e 277 erano basse. Grazie a una mente matematica, e in particolare alle leggi della probabilità, Mendel riconobbe rapidamente che i rapporti si approssimavano a 3:1. Inoltre, le piante basse erano in media della stessa altezza dei loro progenitori di linea pura ‒ ossia la combinazione con i fattori per l'altezza nell'ibrido F1 non ne modificava l'espressione.
Per spiegare questi e altri risultati simili, Mendel ipotizzò che per ogni tratto o carattere dell'organismo esistessero due fattori, uno ereditato dalla madre e l'altro dal padre. Per ogni tratto possono quindi esserci almeno due forme: per esempio, per quanto riguarda l'altezza le forme alta e bassa, mentre per il rivestimento del seme quella liscia o rugosa. Per ogni tratto studiato, Mendel si accorse che una forma sembrava 'dominare' sull'altra, come nel caso dell'altezza, dove la forma alta era dominante su quella bassa. Il fattore 'alto' nascondeva l'effetto del fattore 'basso' ma, come dimostravano i risultati in F2, quest'ultimo riappariva immutato. Inoltre, Mendel sviluppò una notazione per questo processo, che avrebbe reso possibile verificare la sua ipotesi. Se la forma 'alta' viene simbolizzata con una lettera maiuscola (T) e quella 'bassa' con una lettera minuscola (t), allora i progenitori di linea pura possono essere scritti in simboli come TT e tt. Mendel ipotizzò ulteriormente che nella formazione dei gameti (in questo caso, polline e cellula uovo), i due fattori per ogni tratto si dividessero e si distribuissero in gameti separati (ciò divenne noto come principio di segregazione). Nel caso delle linee pure, da piante alte nascono solo gameti con il fattore T, mentre le piante basse producono gameti con il solo fattore t. La notazione usata da Mendel rese possibile indicare che, sebbene le piante F2 sembrassero uguali alle piante alte di linea pura, erano in realtà ibridi, o eterozigoti (Tt), e quindi potevano dar vita a discendenti sia alti sia bassi. Per esempio, retroincrociando un ibrido F1 con una pianta di linea pura bassa, il risultato F2 dovrebbe essere una divisione a metà tra ibridi alti e linee pure basse. I risultati sperimentali confermarono questa previsione.
Mendel esaminò allora l'andamento dell'eredità di due tratti per volta, per esempio altezza e colore del seme (il giallo, Y, è dominante sul verde, y), riuscendo a scoprire che durante la segregazione ogni tratto si separava in maniera indipendente dall'altro, così che negli ibridi F1 (TtYy) erano possibili quattro classi di polline o cellule uovo: TY, Ty, tY, ty. Quando questi doppi ibridi venivano incrociati, la discendenza risultava divisa in quattro categorie: alto-giallo, alto-verde, basso-giallo, basso-verde, in un rapporto, rispettivamente, di 9:3:3:1. Ciò è esattamente quanto ci si sarebbe aspettati se i fattori dei due caratteri si assortissero indipendentemente nel corso della formazione dei gameti nelle cellule germinali dei genitori.
I lavori di Mendel non suscitarono molta attenzione nei suoi contemporanei, e coloro i quali avrebbero maggiormente potuto apprezzare le sue conclusioni, come Darwin o Weismann, non videro il suo articolo del 1866. Solamente una ventina di riferimenti alla pubblicazione originale di Mendel sono stati rintracciati nella letteratura sulla coltivazione, l'ibridazione e lo studio dell'eredità nel periodo 1866-1900. In quest'ultimo anno tuttavia, diversi ricercatori, in particolare Carl Erich Correns (1864-1933) in Germania e De Vries in Olanda, notarono che i rapporti trovati da Mendel erano molto simili a quelli osservati nei propri esperimenti di ibridazione.
Sia Correns sia De Vries, negli articoli pubblicati separatamente nel 1900, fecero riferimento all'opera di Mendel (un terzo riferimento da parte dell'austriaco Erich Tschermak von Seysenegg non viene più riconosciuto come una riscoperta veramente indipendente), e compresero il significato dei rapporti costanti nella discendenza degli ibridi. Quando in Inghilterra William Bateson (1861-1926) venne a conoscenza del saggio di Mendel e lo lesse per la prima volta in treno, da Cambridge (dove abitava) a Londra, dove avrebbe dovuto tenere una conferenza, fu talmente colpito che inserì una breve descrizione dei risultati di Mendel nel testo preparato per quella sera. Bateson diventò presto il maggiore sostenitore della teoria mendeliana nel mondo anglofono, traducendone l'articolo e scrivendo numerosi saggi per esporre la teoria mendeliana ai biologi e ai selezionatori. Con i colleghi Reginald C. Punnett (1875-1967) ed Edith R. Saunders (1865-1945), Bateson mostrò nuovi casi di eredità mendeliana in diversi animali e piante. La promozione del mendelismo da parte di Bateson fu particolarmente ben accetta tra i selezionatori agricoli in Inghilterra e negli Stati Uniti, dove un'intera generazione di ricercatori cominciò ad applicare (sottoponendola a ulteriori elaborazioni) la teoria alle pratiche selettive.
L'aperto sostegno di Bateson al mendelismo lo portò in diretto conflitto con un'altra scuola di ricercatori britannici, i biometrici. Ispirandosi agli studi di Francis Galton (1822-1911), cugino di primo grado di Darwin, i biometrici cercarono di sviluppare non soltanto metodi per la misurazione delle caratteristiche biologiche, ma anche una serie di strumenti statistici per analizzare tali misure (per es., le curve di distribuzione normale, la media, la mediana e la varianza, nonché i principî di correlazione e regressione per analizzare la distribuzione dei caratteri all'interno di una popolazione). I biometrici si dedicarono in particolare al problema dell'eredità visto in una prospettiva statistica. Essi calcolavano i coefficienti di correlazione per caratteri come la statura di genitori e figli o di fratelli e sorelle, analizzandone dal punto di vista statistico i cambiamenti nelle popolazioni da una generazione all'altra.
Come Darwin, i biometrici erano interessati alle relazioni tra eredità ed evoluzione, enfatizzando l'importanza delle piccole differenze individuali tra membri di una popolazione come materia prima per l'evoluzione. La loro idea del processo evolutivo era quindi gradualista, in aspra opposizione al saltazionismo, come la teoria di De Vries. Agli occhi dei biometrici la riscoperta dell'opera di Mendel apparve dunque solamente un altro esempio di discontinuità. Mentre alcuni biometrici, quale Galton, pensavano che i principî mendeliani fossero compatibili con la teoria biometrica, la maggioranza riteneva che tali principî fossero contraddittori, o potessero essere ignorati perché si riferivano solamente a caratteri insignificanti dal punto di vista evolutivo (come il colore dei semi nei piselli).
Una lunga, e a volte molto aspra, disputa tra i mendeliani guidati da Bateson e i biometrici capeggiati da un allievo di Karl Pearson, Walter F.R. Weldon (1860-1906), determinò una frattura nella comunità dei biologi, specialmente in Gran Bretagna ‒ direttamente dal 1901 al 1906, quando Weldon morì, e successivamente per almeno una generazione ‒ fino agli anni Trenta del Novecento. Anche se in altri paesi, come gli Stati Uniti, il dibattito non raggiunse la stessa intensità, era comunque largamente condivisa l'idea che la teoria mendeliana, per quanto interessante e utile nell'ibridazione e in agricoltura, fosse in grado di apportare poche aggiunte alla comprensione della selezione. L'applicazione della teoria mendeliana al darwinismo fu costretta ad aspettare ancora; nel frattempo, i principî di Mendel venivano via via messi alla prova in un numero sempre maggiore di organismi, compresi quelli umani, e legati alle strutture e alle funzioni delle cellule (il dominio della citologia).
La citologia e la teoria cromosomica dell'eredità
Immediatamente dopo la riscoperta di Mendel, due questioni diventarono rilevanti nel dibattito sulla teoria: fino a che punto i fattori mendeliani fossero da considerare entità reali e materiali contenute nelle cellule germinali e quale fosse il processo fisico della loro trasmissione da una generazione all'altra. Le basi materiali per la teoria mendeliana vennero costituite tra il 1880 e il 1920 con la teoria cromosomica dell'eredità, secondo la quale veniva ipotizzato che i fattori, o geni come furono definiti da Wilhelm Johannsen nel 1909, potessero essere considerati segmenti specifici dei cromosomi, strutture filiformi visibili nel nucleo cellulare. La teoria cromosomica dell'eredità rappresentava la fusione di due tradizioni della biologia tra la fine dell'Ottocento e l'inizio del Novecento: la sperimentazione agrozootecnica, esemplificata dagli studi di Mendel, Bateson, Johannsen (in Svezia) e William E. Castle (negli Stati Uniti), e la citologia, lo studio delle strutture cellulari, in particolare del nucleo cellulare, caratterizzato dai lavori di Edouard van Beneden e Richard Hertwig in Germania, ed Edmund B. Wilson, Nettie M. Stevens e Walter S. Sutton negli Stati Uniti. La fusione di questi due filoni di ricerca venne realizzata nel laboratorio di Thomas H. Morgan alla Columbia University di New York tra il 1910 e il 1925, e poi allargata dagli anni Venti agli anni Cinquanta del Novecento, dagli studi sul mais del gruppo di Rollins A. Emerson alla Cornell University, dal laboratorio di citogenetica di Cyril D. Darlington in Inghilterra, e da molti altri.
Negli ultimi due decenni dell'Ottocento gli indizi di un coinvolgimento dei cromosomi nel processo ereditario erano aumentati notevolmente: (1) ogni specie ha il suo numero caratteristico di cromosomi; (2) ogni cellula degli organismi multicellulari contiene lo stesso numero di cromosomi; (3) i cromosomi si presentano a coppie, uno che sembra provenire dal padre, l'altro dalla madre; (4) prima della divisione cellulare i cromosomi si condensano in strutture non più filiformi, ma a forma di bastoncino, intraprendono la replicazione e vengono distribuiti nel corso di un complesso processo noto come mitosi, in modo che alla fine ogni cellula figlia riceva lo stesso numero e tipo di cromosomi della cellula originaria; (5) durante la formazione dei gameti, le coppie di cromosomi vengono divise in un processo noto come meiosi, e i membri della coppia vengono suddivisi tra gameti separati. Questo dimezzamento, o aploidia, è caratteristico dei gameti di quasi tutte le specie (sia animali sia vegetali) a riproduzione sessuale; la condizione doppia, o diplodia, viene ricreata con la fecondazione.
Le osservazioni più convincenti, che indicavano un ruolo centrale dei cromosomi nell'eredità, scaturirono dallo studio sulla determinazione del sesso. Stevens del Bryn Mawr College (Pennsylvania) e Wilson, della Columbia University (New York) notarono contemporaneamente che tra gli animali il sesso sembra essere determinato da una delle coppie di cromosomi, chiamati appropriatamente cromosomi sessuali. Diversamente dalle altre coppie presenti nel genoma, i due cromosomi sessuali hanno strutture visibilmente diverse l'uno dall'altro. Il più grande fu chiamato cromosoma 'X' e il più piccolo 'Y'. Nella gran parte delle specie (tra cui i mammiferi, la maggioranza degli altri vertebrati e quasi tutti gli artropodi) i maschi sono XY e le femmine XX; negli uccelli, nelle falene e nelle farfalle, i maschi sono XX mentre le femmine XY. In alcune specie di insetti, i maschi sono XO (a indicare che manca il cromosoma Y). Tutte queste osservazioni furono confrontate e riassunte nel 1901 dallo zoologo Thomas H. Montgomery, che si basò sui dati di 42 specie diverse.
Con la riscoperta di Mendel nel 1900, molti ricercatori tra i quali Sutton, uno studente di Wilson, suggerirono un possibile parallelo tra il comportamento dei cromosomi durante la meiosi e la 'segregazione' delle due forme alternative dei caratteri (per es., A e a) che era stata ipotizzata da Mendel. Era chiaro che ogni cromosoma non potesse controllare un solo carattere, poiché ogni organismo possiede di gran lunga più caratteri che cromosomi. La risposta alla domanda se i fattori mendeliani fossero in qualche modo parti dei cromosomi, disposti linearmente lungo le strutture filiformi cominciò ad arrivare nel 1910 con i lavori di Morgan e dei suoi studenti, anche loro della Columbia University. Uno degli studenti di Morgan aveva allevato il moscerino della frutta, Drosophila melanogaster, nota al tempo anche come la mosca dell'aceto, Drosophila ampelophila, facendola crescere al buio per molte generazioni, allo scopo di verificare la teoria dell'ereditarietà dell'uso e del disuso delle parti. Nonostante l'esperimento avesse dato risultati negativi (non fu osservata alcuna perdita o diminuzione né per quanto riguarda la dimensione né la funzione dell'occhio), Morgan intuì la potenzialità del piccolo insetto come organismo sperimentale da laboratorio. Si riproduce velocemente (una nuova generazione ogni 10-12 giorni), ha una prole numerosa, e può essere allevato facilmente, in bottiglie da latte in un semplice mezzo di banana e agar. Tra il 1900 e il 1910, lo stesso Morgan era stato molto critico nei confronti sia della teoria mendeliana sia della teoria darwiniana (e in particolare della selezione naturale), considerate troppo speculative e non verificabili sperimentalmente. In realtà, Morgan aveva aderito entusiasta alla teoria mutazionista di De Vries, come proficua alternativa sia a Mendel sia a Darwin. Tra il 1908 e il 1910, aveva perfino iniziato esperimenti di allevamento con Drosophila per verificare se fosse possibile rintracciare (o produrre, per irradiazione con tavolette di radio) in una specie animale macromutazioni, confrontabili con le osservazioni realizzate da De Vries su Oenothera.
Nel 1910 Morgan trovò nelle colture di Drosophila un maschio con gli occhi bianchi, invece del normale colore rosso mattone; si trattava sicuramente di una variazione discontinua ed evidente, ma ovviamente non era una nuova specie. Tuttavia, Morgan incrociò questa mosca con una femmina con gli occhi rossi, creando una discendenza (F1) solamente con occhi rossi. Facendo accoppiare nuovamente la generazione F1, trovò che nella generazione successiva F2 circa 3/4 delle mosche avevano gli occhi rossi e 1/4 gli occhi bianchi. Era esattamente il rapporto mendeliano che ci si sarebbe aspettati per un carattere completamente dominante. Morgan, però, notò anche qualcosa che si accordava male con il modello mendeliano: tutte le mosche dagli occhi bianchi erano maschi. Che il colore degli occhi fosse in qualche modo legato al sesso era l'ovvia interpretazione, ma Morgan non formulò esplicitamente questa ipotesi. Continuò ad allevare Drosophila, osservando altri caratteri, tra cui diversi colori degli occhi, del corpo e differenti forme delle ali, che avevano diverse varianti distinte. Sebbene alcuni caratteri non sembrassero avere maggiore correlazione con un sesso o con l'altro (vale a dire, sembravano seguire il principio di Mendel dell'assortimento casuale), un certo numero si comportava come il carattere occhio bianco, con le forme recessive che apparivano più frequentemente nei maschi. Morgan chiamò questo fenomeno 'eredità limitata al sesso' (in seguito eredità legata al sesso) e nel 1911 gli sembrò che i fatti avrebbero avuto una semplice spiegazione ipotizzando che i fattori legati al sesso fossero parte del cromosoma X. Le femmine, essendo XX, avrebbero avuto minor probabilità di mostrare il carattere occhio bianco recessivo, perché l'omologo (l'altro X) avrebbe probabilmente avuto il gene dominante normale; i maschi invece, non avendo un secondo X, avrebbero sempre mostrato il carattere recessivo occhio bianco nel caso lo avessero ereditato per via materna.
Nel biennio 1911-1912 Morgan portò nel suo laboratorio due laureandi della Columbia, Alfred H. Sturtevant e Calvin B. Bridges; nel corso del 1912, si unì al laboratorio Hermann Joseph Muller. Il gruppo si espanse rapidamente, includendo altri dottorandi e ricercatori. Con l'aumento dei caratteri presi in esame, Morgan e i suoi studenti osservarono un fenomeno che Bateson aveva già rilevato: mentre alcuni fattori si segregavano secondo le previsioni mendeliane, altri erano 'accoppiati' (termine usato da Bateson) e sembravano ereditati insieme (ma non associati a un sesso piuttosto che all'altro). Bateson aveva anche notato che in molti casi i caratteri 'accoppiati' si separavano in una piccola percentuale dei discendenti. Per diverse ragioni (soprattutto il suo antimaterialismo), Bateson aveva rifiutato l'interpretazione cromosomica dell'accoppiamento e della separazione, sviluppando invece una teoria piuttosto complessa nota come ipotesi della riduplicazione. Morgan pensava invece che l''accoppiamento' fosse forse da ascrivere all'associazione sui cromosomi, ma la 'separazione' appariva come un'anomalia.
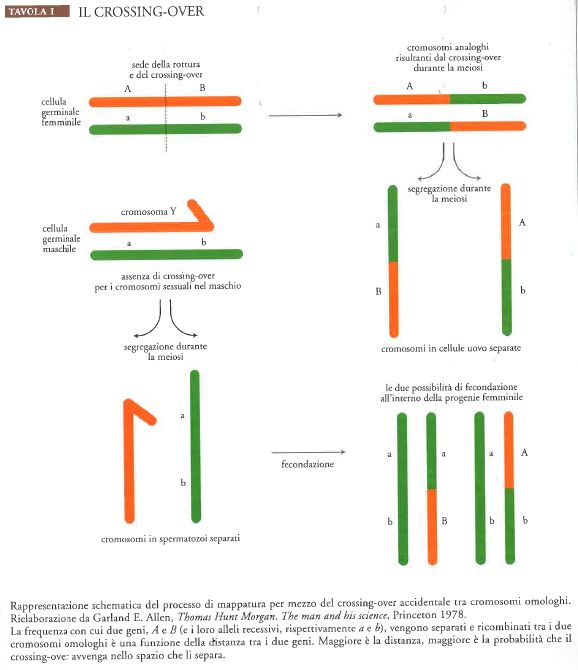
La risposta al problema dell'accoppiamento si presentò abbastanza casualmente. Un giorno Wilson richiamò l'attenzione di Morgan su un curioso articolo del 1904 del citologo belga Frans Alfons Janssens: nei preparati microscopici di cromosomi durante la fase di meiosi, Janssens aveva notato che nel processo chiasmatico, in cui i due membri di una coppia di cromosomi (omologhi) si arrotolano l'uno sull'altro, i cromosomi sembravano rompersi e scambiarsi alcune parti. Morgan dedusse subito che questo processo, detto crossing-over, avrebbe spiegato l'accoppiamento e la separazione di Bateson. In una discussione immediatamente successiva, Morgan e Sturtevant compresero che se veramente i fattori di Mendel erano disposti linearmente lungo i cromosomi, allora la frequenza delle rotture durante i chiasmi sarebbe stata più o meno direttamente proporzionale alla distanza tra i fattori stessi; di conseguenza, la frequenza con cui i fattori accoppiati vengono separati sarebbe potuta risultare utile per costruire mappe per ogni cromosoma. Il giorno successivo, Sturtevant preparò una mappa di molti dei fattori mutanti noti sul cromosoma X di Drosophila (Tav. I).
La mappatura divenne l'obiettivo principale del gruppo della Drosophila per il decennio successivo. La procedura fu applicata anche ad altri organismi quali topi e mais. Sebbene alcuni ricercatori (tra gli altri Bateson e Richard Goldschmidt in Germania) mostrassero un perdurante scetticismo nei confronti della teoria cromosomica dell'eredità, nel 1931 furono ottenute le prove citologiche del crossing-over per il mais (Barbara McClintock e Harriet Creighton) e per Drosophila (Curt Stern), eliminando ogni dubbio. Non sfuggì d'altra parte all'attenzione di alcuni scienziati, tra cui lo stesso Morgan, che le variazioni mendeliane (a partire dal 1915 chiamate 'mutazioni', indicando però mutamenti di grandezza minima) insieme alla ricombinazione dei caratteri per mezzo del crossing-over, creavano una grande variabilità su cui la selezione naturale poteva agire, in misura maggiore di quanto ipotizzato fino ad allora. Era inoltre diventato chiaro che il modello di Mendel non si poteva ritenere una formalizzazione puramente speculativa.
Allo stesso tempo Morgan e il suo laboratorio, insieme al gruppo ‒ in rapida crescita ‒ del mais di Emerson, iniziarono a trovare esempi di interazione tra geni, che indicavano, come gli embriologi avevano previsto da tempo, che i geni non erano semplicemente caratteri in miniatura, bensì erano coinvolti (in un modo che all'epoca non era ancora compreso) nella direzione dello sviluppo. Tra gli esempi di questa interazione c'era la cosiddetta 'epistasi', in cui gli effetti combinati di due geni in diverse posizioni o loci sul cromosoma agivano sulle rispettive funzioni, e l'effetto posizionale, in cui alcuni cambiamenti cromosomici (causati dalla duplicazione di regioni cromosomiche, o dalla rottura di un pezzo del cromosoma durante il chiasma e la sua reinserzione in senso inverso ‒ chiamata 'inversione') possono modificare la posizione di un gene e quindi anche la sua espressione. Nonostante queste scoperte, l'immagine prevalente che si affermò fu quella di geni mendeliani indipendenti, allineati lungo il cromosoma, i quali producono caratteri adulti invarianti (il fenotipo). Questa visione è ancora prevalente in molti manuali e libri di divulgazione, con espressioni del tipo 'Il gene per...' che, anche se fuorvianti, sono divenute parte della genetica moderna.
La biometria e la teoria sintetica dell'evoluzione
Per quanto il movimento biometrico si fosse occupato direttamente dei problemi dell'evoluzione, di per sé la biometria non formulava meccanismi evolutivi nei termini della nuova genetica. La durevole opposizione dei biometrici al mendelismo portò al rifiuto di qualsiasi ruolo per i geni nel processo evolutivo. Il vero contributo dei biometrici alla sintesi evoluzionistica che ebbe inizio negli anni Trenta del Novecento fu il ragionamento in termini statistici e il riconoscimento dell'importanza della matematica come strumento per affrontare l'evoluzione a livello popolazionale. Questo modo di vedere fu condiviso negli anni Venti da diversi studiosi, quali John B.S. Haldane e Ronald A. Fisher in Inghilterra, Sewall Wright negli Stati Uniti, e Sergej Četverikov in Unione Sovietica. Una delle prime applicazioni del pensiero statistico riguardò la questione dei caratteri recessivi, ossia se la loro scomparsa avvenga o meno in modo graduale. Nel 1908, il matematico inglese Godfrey H. Hardy e il fisico tedesco Wilhelm Weinberg dimostrarono matematicamente ‒ in maniera indipendente l'uno dall'altro ‒ che le frequenze geniche rimangono stabili in una popolazione se vengono mantenute alcune condizioni: una popolazione abbastanza grande da evitare errori di campionamento, un accoppiamento casuale, nessuna apprezzabile selezione, niente mutazioni né migrazioni. Noto come 'equilibrio di Hardy-Weinberg', questo semplice enunciato è divenuto la base per la comprensione dell'evoluzione in termini di genetica mendeliana. Sono fattori come l'accoppiamento non casuale, le mutazioni e altre forme di variazione (incluse le inversioni cromosomiche et similia), le migrazioni e la nascita di nuove piccole popolazioni isolate, e la selezione naturale, che producono l'evoluzione. Questa poteva ora essere considerata in termini di cambiamento di frequenze geniche nel tempo, prodotto da uno di questi processi. L'equilibrio di Hardy-Weinberg rappresentò il principale punto di partenza per gli studi di Fisher, Wright e Haldane dalla seconda metà degli anni Dieci fino agli anni Trenta del Novecento.
La prima fase della sintesi evoluzionistica
Fisher, Wright e Haldane, per quanto lavorando indipendentemente, erano tutti e tre interessati in particolare a misurare quantitativamente l'effetto della selezione naturale in determinate condizioni. Essi affrontarono però il problema da diverse angolazioni. Fisher era il più avvezzo all'uso della matematica, avendo studiato astronomia a Cambridge con Sir James Jeans e avendo poi lavorato, successivamente alla laurea, come attuario presso una compagnia di assicurazioni. Non c'è da meravigliarsi del fatto che i suoi studi risultassero i più astratti e teorici, e i meno orientati alle popolazioni reali in Natura. Haldane era, fin dal principio, più indirizzato verso la biologia, avendo studiato matematica, genetica e fisiologia a Oxford e biochimica con Frederick Gowland Hopkins a Cambridge. Wright conseguì invece il suo dottorato a Harvard sotto la guida del genetista Castle, e successivamente lavorò presso lo United States Department of Agriculture (USDA), sviluppando programmi di selezione delle colture; inoltre, da autodidatta, studiò matematica e statistica. Tra i tre studiosi, Fisher era il più interessato nell'applicazione dei suoi modelli matematici al miglioramento della razza umana tramite programmi di eugenetica; addirittura, l'ultimo terzo del suo pionieristico volume, The genetical theory of natural selection (1930), è interamente dedicato all'eugenetica.
Gran parte degli storici considera gli studi di Fisher come il fondamento della sintesi evoluzionistica, dal momento che enuncia i principî della genetica di popolazione e sviluppa metodi statistici per quantificare i modelli. Il titolo del libro di Fisher è significativo perché indica i limiti che si autoimpose. Egli scelse appositamente un ambito ristretto, mettendo in relazione la genetica mendeliana con il processo di selezione naturale, senza prendere in considerazione le altre questioni come le migrazioni, l'isolamento, le interazioni epistatiche e la ricombinazione genetica. Fisher comincia la sua discussione da ciò che denomina il parametro maltusiano di incremento della popolazione, cioè le statistiche vitali degli organismi in relazione al loro ambiente come descrizione del relativo tasso di crescita delle popolazioni. Il parametro maltusiano misura i valori produttivi degli individui di ogni età o in ogni stadio dei loro cicli vitali, e dunque si prevede una variazione a seconda del genotipo; esso fornisce un indice della capacità di sopravvivenza di un genotipo in specifiche condizioni ambientali. A questa definizione, Fisher aggiunse il concetto di varianza genetica, che esprime quantitativamente lo spettro di variazioni che un particolare carattere genetico può mostrare in diverse condizioni ambientali (stagioni differenti, cicli riproduttivi, ecc.). Ogni singola misura in una popolazione sarà dunque l'espressione non solo della variabilità genetica, ma anche delle interazioni tra i geni e delle influenze ambientali. Tuttavia, se le misurazioni sono fatte su un campione abbastanza ampio da comprendere l'intero spettro di ambienti in cui vive la popolazione, è possibile distinguere le variazioni realmente genetiche da quelle indotte dall'ambiente. Questo fu un passo cruciale per Fisher (ciò che lui denominò analisi della varianza), perché gli permetteva di analizzare gli effetti della selezione su variazioni mendeliane reali, e forniva un modello grazie al quale si potevano prevedere quantitativamente gli effetti di numerosi fattori, come la quantità di variabilità nella popolazione o l'intensità della selezione.
Fisher mise poi in relazione la sua analisi della varianza genetica con il processo di selezione naturale. La prima componente di questo modello è la variabilità genetica, che Fisher pensava dovuta quasi totalmente alle mutazioni. Basandosi sugli studi di Muller sulle mutazioni indotte dalle radiazioni, condotti negli anni Venti del Novecento, ipotizzò che la mutazione di un gene in una delle sue forme alleliche avvenisse a un ritmo prevedibile (anche se i ritmi sono bassi, nell'ordine di 1:10.000 o 1:100.000 gameti), consentendo quindi una misura quantitativa della rapidità con cui nuove variazioni entrano in una popolazione. Più grande è la popolazione, maggiore è la variazione: per Fisher le grandi popolazioni erano quindi i luoghi chiave in cui l'evoluzione avveniva con maggior efficacia. Il secondo componente del modello di Fisher era la selezione naturale medesima, i fattori ambientali che determinavano le possibilità del gene o di un gruppo di geni di essere trasmessi, e con quale frequenza, alla generazione successiva.
Il concetto centrale proposto da Fisher in The genetical theory definì il teorema fondamentale della selezione naturale. Argomentando in maniera puramente quantitativa, Fisher metteva in evidenza che, se un cambiamento di 1 mm in un carattere ha un valore selettivo di x, allora un cambiamento di 0,1 mm avrebbe un valore di 0,1x. Il tasso con cui una mutazione incrementa la propria frequenza non dipende dal vantaggio selettivo che essa conferisce al singolo individuo, ma dalla variazione totale ereditabile nella popolazione (senza essere influenzata dall'entità delle singole variazioni). Questo rappresentava un importante preludio al teorema fondamentale stesso, perché in un colpo solo eliminava la distinzione tra variazioni continue e discontinue (rispettivamente, piccole o grandi mutazioni) che Fisher considerava inutile e che tanto aveva ostacolato nel passato l'applicazione della genetica mendeliana ai problemi dell'evoluzione. Il teorema fondamentale continua definendo il concetto di fitness, o grado di vantaggio (o svantaggio) selettivo che un particolare gene possiede in confronto ai suoi alleli, stabilendo che il tasso di incremento di fitness per ogni organismo in qualsiasi momento è uguale alla varianza genetica della fitness in quel momento. Il valore selettivo diventava dunque una caratteristica dei geni, non degli organismi nella loro interezza. The genetical theory riduceva gli organismi a collezioni di geni (quello che è detto pool genico della popolazione), rendendo il processo evolutivo trattabile con le modellizzazioni matematiche e le analisi quantitative che Fisher prediligeva.
I modelli di Fisher erano molto semplificati, ma proprio nella semplicità risiede la loro importanza. Egli separò attentamente i fili più importanti nella trama della variazione popolazionale, mostrando che l'ampiezza della variabilità in una data popolazione era il risultato di tre fattori separati: la reale variazione genetica, gli effetti ambientali e l'errore di campionamento da parte dell'osservatore. In questo processo, Fisher affermava di fare per la teoria evoluzionistica ciò che le leggi dei gas avevano fatto per la fisica: fornire gli strumenti per l'analisi statistica di eventi casuali, così da poter produrre risultati regolari e prevedibili. Come i sistemi fisici, le popolazioni di Fisher erano pensate infinitamente grandi e panmittiche, cioè tali che gli individui delle popolazioni potessero incrociarsi liberamente. Sebbene egli ammettesse che non tutte le popolazioni avrebbero avuto queste caratteristiche, pensò tuttavia che i cambiamenti evolutivi significativi si verificassero soprattutto all'interno di grandi popolazioni.
L'approccio di Fisher fu criticato da Haldane, che pensava fosse eccessivamente idealizzato e semplificato. Marxista molto legato culturalmente alla Dialektik der Natur (Dialettica della Natura) di Friedrich Engels (uno scritto postumo che applica il materialismo dialettico a diversi sistemi naturali), Haldane era particolarmente interessato alle interazioni tra i geni e gli altri parametri nelle popolazioni. Considerando i sistemi di Fisher 'troppo meccanici', si concentrò sui processi interattivi, come la dominanza incompleta, i sistemi multigenici, come l'epistasi, l'incrocio casuale in opposizione all'incrocio selettivo, e l'autofecondazione rispetto all'incrocio. Per mezzo dei calcoli basati sui dati di Biston betularia (una farfalla notturna per la quale la varietà melanica più scura aveva sostituito in meno di un secolo la forma normale bianca nelle aree industriali come Manchester), Haldane aveva concluso che in generale la pressione selettiva dovesse essere maggiore di quanto avesse pensato Fisher, aumentando quindi, in media, la velocità dell'evoluzione. Con una formazione in biochimica e matematica, Haldane, al pari di Fisher, sviluppò modelli basati su supposizioni ‒ riguardanti le popolazioni, le loro dimensioni, la variabilità, le diverse pressioni selettive e l'esistenza di differenti parametri ‒ non direttamente tratti dall'osservazione naturalistica. Sia per Fisher che per Haldane, le popolazioni erano astrazioni da descrivere in termini di frequenze di particolari genotipi.
Gli studi di Sewall Wright erano invece attentamente costruiti in base ai dati raccolti sulle popolazioni reali, per quanto anch'egli sviluppò modelli altamente matematizzati che per molti biologi dell'epoca erano difficili, se non impossibili, da capire. Entrando in scena qualche anno dopo rispetto a Fisher e Haldane, Wright approfittò delle loro precedenti pubblicazioni, riconoscendo in maniera esplicita l'importanza del pensiero di Haldane per il proprio crescente interesse nella genetica di popolazione. Provenendo dal laboratorio di Castle a Harvard, era sicuramente ben preparato per cogliere la relazione tra la genetica mendeliana e la selezione (Castle aveva eseguito una serie di esperimenti selettivi sulle cavie e i ratti per dimostrare che la selezione spingeva realmente i geni mendeliani a cambiare secondo la direzione della pressione selettiva). Il punto di partenza di Wright era una situazione semplice in cui l'equilibrio genetico prevaleva, per poi osservare i fattori che potevano spostare questo equilibrio. I fattori studiati nel dettaglio da Wright comprendevano la mutazione, la selezione, la dimensione delle popolazioni e la migrazione; egli introdusse quindi condizioni che le analisi di Fisher e Haldane avevano in larga misura ignorato.
La deriva genetica
Wright enfatizzò l'importanza della struttura e della dimensione della popolazione per il processo evolutivo. Al contrario di Fisher, Wright non considerava le popolazioni in natura infinitamente grandi ed equamente distribuite sul territorio, ma piuttosto le pensava disperse in piccole sottopopolazioni, o demi, che abitano nicchie locali all'interno dell'area geografica di tutta la popolazione. Queste piccole popolazioni occupano ciò che Wright chiamava un 'picco adattativo' all'interno di questo paesaggio. I picchi adattativi erano più metaforici che reali, ma ognuno di essi rappresentava una composizione genetica unica che aumentava la propria fitness nel microambiente del deme. Data la piccola dimensione delle popolazioni, queste risentivano in misura maggiore degli effetti dei processi casuali, come le migrazioni, le mutazioni, le ricombinazioni, la casualità dell'accoppiamento e i cambiamenti ambientali. Wright introdusse il concetto di 'deriva genetica' per descrivere le modalità in cui le frequenze genetiche si fissano nelle popolazioni per effetto di agenti diversi dalla selezione naturale. Pur senza negare il ruolo della selezione naturale, Wright sostenne che altri fattori hanno una parte importante nel creare le differenze di frequenze geniche tra demi. Allo stesso tempo, le piccole popolazioni di Wright erano in equilibrio precario sui loro picchi adattativi, poiché ogni mutamento ambientale, ogni migrazione o l'accoppiamento casuale avrebbe potuto alterare significativamente le frequenze geniche nello spazio di una generazione. Diversamente dal modello di Fisher, nel quale la selezione spingeva costantemente una grande popolazione verso una fitness crescente, il modello di Wright suggeriva che la maggior parte delle piccole popolazioni alla fine va incontro all'estinzione. Il vantaggio per la popolazione nel suo complesso risiedeva nelle collezioni uniche di geni che ogni deme rappresentava, fornendo dunque un serbatoio di variabilità maggiore di quanto accadesse per le grandi, ma più omogenee, popolazioni di Fisher. Inoltre, i demi potevano rapidamente evolvere da un picco adattativo all'altro, fintanto che riuscivano a sopravvivere alla traversata attraverso la 'valle' di valore adattativo minore. Il modello dei paesaggi adattativi fu introdotto da Wright in un articolo del 1932 e divenne uno strumento molto utile per visualizzare e descrivere la deriva genetica e le altre conseguenze delle dimensioni ridotte delle popolazioni.
Lo schema della deriva genetica di Wright venne però fortemente criticato da darwiniani ortodossi come Ernst Mayr, perché all'epoca sembrava sminuire o addirittura escludere un ruolo importante per la selezione e inoltre riduceva il potere dell'adattamento come principale 'ragion d'essere' del cambiamento evolutivo. Le ipotesi di Wright, per quanto sempre riconosciute, entrarono a far parte a pieno titolo della sintesi evoluzionistica solo negli anni Sessanta.
Studi sulle popolazioni naturali
La prima fase della sintesi evoluzionistica fece dunque convergere la genetica mendeliana e la teoria darwiniana della selezione (insieme al suo assunto del gradualismo) in un insieme di modelli altamente matematizzati e astratti. Mancava quindi l'applicazione e/o la verifica di questi modelli nelle popolazioni naturali. Tali applicazioni vennero realizzate da Theodosius Dobzhansky negli Stati Uniti (in parte collaborando con Wright), Edmund B. Ford in Inghilterra (in collaborazione con Fisher), George G. Simpson negli Stati Uniti (che applicò la genetica di popolazione ai dati fossili) e Mayr, utilizzando l'approccio popolazionale per trattare il problema della specie.
Dobzhansky (1900-1975) nacque a Odessa, Russia, e nei primi anni Venti del Novecento fu allievo di uno dei più importanti naturalisti russi, Sergej Četverikov. Con una formazione da sistematico, Četverikov divenne anche decisamente darwiniano e, quando Muller visitò l'Unione Sovietica nel 1922, gli comunicò l'entusiasmo che circondava la genetica di Drosophila.
Abile negli studi sul campo, Četverikov tentò di applicare i principî della genetica mendeliana alle popolazioni naturali, raccogliendo organismi (soprattutto Drosophila) e portandoli in laboratorio, dove li incrociava con individui di composizione genetica nota per osservarne la variabilità. Da questi studi, egli concluse che tutte le popolazioni dovevano ospitare una gran quantità di variazione nascosta o 'criptica', che avrebbe potuto avere conseguenze importanti sull'evoluzione. Per Četverikov, così come per il suo allievo Dobzhansky, il lavoro sul campo e in laboratorio erano complementari.
Nel 1927 Dobzhansky ricevette un borsa di studio dalla Rockefeller Foundation per recarsi negli Stati Uniti a lavorare nel laboratorio di Morgan alla Columbia University; egli avrebbe dovuto cominciare da zero lo studio dei dettagli della genetica e della citogenetica di Drosophila. In parte per le fruttuose collaborazioni avviate negli Stati Uniti, in parte per i crescenti attacchi alla genetica mendeliana in Unione Sovietica da parte dei neolamarckiani come Trofim Denisovič Lysenko, Dobzhansky rimase negli Stati Uniti per il resto della sua carriera.
Quando Morgan si trasferì dalla Columbia al California Institute of Technology (Caltech) nel 1928, Dobzhansky fu invitato a seguirlo. La vicinanza del deserto a sud-ovest di Pasadena fu lo stimolo che spinse Dobzhansky a intraprendere gli studi sul campo come nei primi anni Venti aveva fatto Četverikov. Dalla California meridionale all'Arizona orientale e al Texas, egli raccolse campioni di popolazioni di Drosophila a diverse altitudini, in habitat e stagioni differenti. Utilizzando le tecniche citologiche originariamente sviluppate da Theophilus S. Painter della University of Texas, ampiamente sfruttate dal gruppo di Morgan, Dobzhansky identificò le diverse configurazioni delle bande cromosomiche (utilizzando i cromosomi giganti delle ghiandole salivari delle larve). In questo modo riuscì a seguire diverse inversioni cromosomiche e altre alterazioni da popolazione a popolazione e da stagione a stagione, variando le condizioni che considerava correlate ai cambiamenti nelle frequenze cromosomiche. Sebbene non fosse possibile dimostrare rigorosamente che la selezione per alcune configurazioni cromosomiche specifiche fosse adattativa in particolari ambienti, Dobzhansky riuscì a riprodurre cambiamenti stagionali nelle frequenze cromosomiche modulando i parametri fisici del laboratorio.
Dobzhansky pubblicò i suoi studi in Genetics and the origin of species (1937) e in una serie di articoli dagli anni Trenta agli anni Cinquanta con il titolo di Genetics of natural populations. Egli mostrò chiaramente che le popolazioni naturali ospitavano una gran quantità di variabilità criptica, maggiore di quanto si fosse pensato, confermando di fatto i precedenti studi di Četverikov. Inoltre, fornì un metodo diretto per stabilire la dimensione della variabilità per mezzo dei metodi citologici osservativi, mentre Četverikov aveva bisogno di allevare diverse generazioni di mosche per svelare la variazione criptica. Da ultimo, creò un legame tra la tradizione di studi sul campo della storia naturale (dalla quale provenivano Darwin e Wallace) e la tradizione di laboratorio dei genetisti, colmando una distanza che negli anni Venti del Novecento era divenuta uno dei principali ostacoli allo sviluppo di una visione unitaria dell'evoluzione: i naturalisti pensavano che i genetisti di laboratorio lavorassero con scenari artificiali, mentre i genetisti ritenevano che i naturalisti fossero incapaci di testare le proprie ipotesi e fossero quindi obbligati a teorie speculative. In parte grazie alle sue ricerche, Dobzhansky poté scrivere nel 1947 che l'evoluzione per selezione naturale poteva essere soggetta a prove quantitative e sperimentali: gli esperimenti controllati potevano allora sostituire la speculazione rispetto a ciò che la selezione naturale è in grado o meno di fare. Inoltre, non ci si poteva più accontentare della mera verifica dell'esistenza della selezione naturale. Il meccanismo della selezione naturale poteva essere studiato in casi concreti e, di conseguenza, la genesi dell'adattamento, probabilmente il problema centrale della biologia, rientrava nell'ambito del metodo sperimentale.
Molti degli esperimenti sul campo e in laboratorio di Dobzhansky erano progettati in collaborazione con Wright; questi proponeva i modelli e il tipo di dati da raccogliere e Dobzhansky portava il problema sul campo, raccogliendo gli esemplari ed effettuando le analisi citogenetiche.
Rispetto al problema chiave della biologia evolutiva, l'adattamento, Dobzhansky, come Wright, enfatizzava il fatto che esso fosse il risultato dell'interazione di molteplici geni, e non la caratteristica di singoli alleli. La selezione agiva sulla totalità del complesso cromosomico, non sui singoli geni come aveva evidenziato Fisher. Dobzhansky sostenne che la selezione naturale poteva agire in diversi modi. I tipi cromosomici potevano essere caratterizzati dalla mortalità differenziale, o dalla longevità differenziale, o dalla fecondità o dalle differenze nell'attività sessuale, o dalla combinazione di due o più di queste e altre variabili. Durante questa seconda fase della sintesi (1940-1960) la teoria evoluzionistica è stata inserita, metodologicamente, tra le scienze tradizionali.
Specie e speciazione
Un altro dei problemi a cui si dedicarono i protagonisti della sintesi fu la natura delle specie e il processo di speciazione. I tentativi, sia precedenti che successivi a Darwin, di definire le specie in un qualsiasi modo abbastanza generale da includere un vasto insieme di organismi (dai procarioti unicellulari agli eucarioti pluricellulari) erano caratterizzati da un approccio idealistico e tipologico. In parte come risposta a questa visione prevalente, Darwin assunse una posizione maggiormente nominalista: i limiti tra le specie sono in larga misura creazioni umane e artificiali. La distinzione tra specie e varietà era per Darwin necessariamente sfumata, poiché molte cosiddette specie erano solamente varietà in corso di diversificazione, ma ancora non abbastanza diverse da poter essere catalogate come vere specie separate. Tale confusione portò alcuni critici di Darwin a sottolineare come il il titolo del suo libro fosse fuorviante, perché non dimostrava 'l'origine delle specie' ma evidenziava solamente come esse si modificassero entro certi limiti. Negli anni Quaranta del Novecento, tuttavia, naturalisti come Mayr, Simpson, Julian Huxley e George L. Stebbins avevano formulato un'idea più definita della specie, nota come 'concetto biologico di specie'. Questo concetto prevede che le specie siano entità reali in Natura, definite in generale dalla loro incapacità di incrociarsi con altri gruppi simili, e che esse condividano un pool genico comune insieme ad altri caratteri morfologici e fisiologici. Il concetto biologico di specie è 'multidimensionale', nel senso che non esiste alcun singolo criterio sufficiente a distinguere una specie da un'altra, e quindi i tassonomisti e i biologi evoluzionisti devono valutare tutte le caratteristiche ecobiologiche di una popolazione per determinarne lo statuto di specie o di semplice varietà. Sebbene per i microbiologi e per molti botanici il concetto biologico di specie non fosse di grande aiuto nella loro ricerca tassonomica, esso è rimasto un importante elemento della sintesi evoluzionistica dai primi anni Quaranta del Novecento in poi.
Il processo di speciazione emerse come controversia durante gli anni Cinquanta, opponendo i sostenitori della speciazione allopatrica e della speciazione simpatrica. Sviluppata da Darwin e poi fortemente rielaborata a cavallo tra XIX e XX sec. da autori quali John Thomas Gulick e David Starr Jordan, la speciazione allopatrica si rifà all'idea che una condizione necessaria per la differenziazione di una specie in due o più discendenti sia l'isolamento geografico. Mayr, Huxley e altri, a partire dal 1940 avevano sostenuto che fintanto che una popolazione condivide un pool genico comune caratterizzato da incrocio incondizionato tra i suoi membri, non si può avere diversificazione tra specie. Tuttavia, se alcune porzioni della popolazione venissero a trovarsi separate fisicamente dalle altre, o a causa della formazione di una barriera (fiume, una catena montuosa), o per mezzo della migrazione di alcuni individui in un nuovo territorio separato dall'originario, allora ci si dovrebbe aspettare una diversificazione in seguito a mutazioni casuali e a diverse pressioni selettive nei due ambienti. L'ipotesi alternativa, la speciazione simpatrica, sosteneva invece che tale rigoroso isolamento non fosse necessario per la speciazione, poiché alcuni esempi, come le zecche, stavano a dimostrare che due o più specie potevano nascere in una stessa area geografica limitata. La speciazione simpatrica era giudicata impossibile dai principali architetti della sintesi, come Mayr, poiché, fintanto che gli organismi fossero stati a contatto tra loro e liberi di incrociarsi, non si sarebbero mai accumulate differenze genetiche tra una parte e l'altra della popolazione sufficienti a dar luogo all'isolamento riproduttivo. Il successivo dibattito si concentrò sulla definizione dell'isolamento geografico: per esempio, due popolazioni di talpe possono essere effettivamente isolate pur vivendo a pochi metri di distanza, poiché la loro mobilità e le abitudini ecologiche escludono praticamente ogni contatto. Gli umani, d'altro canto, sono rimasti un'unica specie per almeno un milione di anni, anche se distribuiti geograficamente sull'intero Pianeta. Il concetto biologico di specie e i meccanismi di speciazione sviluppati durante la sintesi misero la parola fine a qualsiasi ipotesi che identificasse i diversi gruppi razziali umani con differenti specie.
Altri aspetti della sintesi evoluzionistica includevano la sistematica (tassonomia e il problema della specie), la biogeografia e la paleontologia. La questione della specie fu affrontata direttamente nel volume del 1942 di Mayr, Systematics and the origin of species. Le specie, sia animali sia vegetali, erano definite come popolazioni interfeconde con un insieme condiviso di caratteri anatomici, fisiologici, ecologici e comportamentali (se rilevanti). La biogeografia era rappresentata da un gran numero di ricercatori da Darwin in poi, particolarmente negli anni Venti e Trenta del Novecento, quando iniziò ad allontanarsi dalla semplice descrizione verso ricerche generalmente più dinamiche e causali, nei lavori di Emmett R. Dunn, Simpson e Philip J. Darlington. Le distribuzioni geografiche furono correlate con la speciazione, l'isolamento, il flusso genico e l'ibridazione interspecifica. La paleontologia fu portata nella sintesi da Simpson, che applicò i metodi statistici (compresa l'allometria) all'analisi delle tendenze dei dati fossili e alle distribuzioni delle popolazioni estinte. Gli aspetti botanici della sintesi furono rappresentati da George L. Stebbins e da Edgar Anderson i quali, al contrario di molti botanici, sottolinearono l'estensione (con qualche eccezione, in particolare la loro capacità di ibridazione) dell'applicazione alle piante del concetto biologico di specie come enunciato da Mayr e Huxley.
La denominazione 'sintesi evoluzionistica' fu mutuata dal titolo del libro di Huxley del 1942, Evolution, the modern synthesis. Come il volume di Dobzhansky di cinque anni prima, quello di Huxley fornì il riassunto più chiaro e aggiornato di ciò che rappresentava, o non rappresentava, la nuova biologia evoluzionistica. La sintesi moderna escludeva tanto quanto includeva. Una delle caratteristiche della sintesi fu il progressivo consolidamento intorno a un rigido insieme di idee riguardo ciò che sarebbe dovuto essere incluso o escluso dalla teoria evoluzionistica vera e propria. Nella sintesi nel periodo 1930-1950, quindi, furono incorporati la genetica mendeliana, la teoria darwiniana della selezione, il problema della specie, l'isolamento geografico (e il problema della speciazione) e la paleontologia. Venivano invece escluse le vecchie nozioni del neolamarckismo, dell'ortogenesi e le macromutazioni (così come proposte da De Vries), ritenute senza alcun ruolo nel processo evolutivo. Venne inoltre escluso, per quanto ritenuto comunque di un certo rilievo, il collegamento con l'embriologia, che era stata considerata a pieno titolo parte della teoria evoluzionistica dalla generazione precedente di Haeckel e Weisman. Era chiaro che la selezione poteva agire sia sui caratteri embrionali sia su quelli adulti, ma non era disponibile alcun metodo per studiare il processo noto al tempo come 'genetica dello sviluppo'. Uno dei risultati della sintesi fu quindi che lo sviluppo venne considerato una scatola nera; sebbene gli architetti della sintesi avessero riconosciuto che alcuni fattori potevano alterare l'espressione genica nel corso dello sviluppo, furono obbligati nella realtà dei fatti a trattare i geni come se non avessero alcun ruolo in questo processo. A partire dagli anni Ottanta del Novecento, tuttavia, grazie ai nuovi strumenti della biologia molecolare, e in particolare della genomica, un maggiore interesse è stato dimostrato per la relazione tra geni, sviluppo ed evoluzione. All'inizio del XXI sec. uno dei campi di ricerca biologica in maggiore crescita, noto cripticamente come 'evo-devo' (evolution-development), ha attratto l'attenzione di una nuova generazione di ricercatori; i biologi tornano a distanza di un secolo ai problemi centrali del paradigma haeckeliano.
L'evoluzione del comportamento animale
Darwin aveva sollevato il problema dell'evoluzione del comportamento animale nella sua opera On the origin of species, con un intero capitolo dedicato all'istinto. L'argomento occupò anche gran parte dei successivi The descent of man (1971) e The expression of emotions in man and animals (1872). Wallace, scopritore con Darwin della selezione naturale, si dedicò all'evoluzione della 'mente' umana e della morale, escludendola infine dai processi materiali come la variabilità ereditaria e la selezione naturale. Sebbene a cavallo tra XIX e XX sec. molti scienziati lavorassero sull'evoluzione del comportamento, lo studio sistematico in questo campo, poi battezzato 'etologia', non ebbe seguito fino agli anni Quaranta del Novecento.
Etologia
L'etologia è distinta dalla psicologia animale, una disciplina importante ma separata, nata nell'Ottocento. La psicologia animale era in gran parte incentrata sullo studio dell'apprendimento negli animali e quindi si occupava principalmente delle cause prossime. Gli studiosi di psicologia animale tendevano a pensare che l'organismo nascesse come una sorta di tabula rasa, su cui le esperienze fatte dall'animale venivano trascritte. Per quanto gli istinti fossero riconosciuti come importanti, non fu loro accordato il posto principale nello studio del comportamento. La psicologia animale fu profondamente influenzata dalle teorie comportamentistiche di Ivan Petrovič Pavlov, John Watson e Burrhus F. Skinner. L'etologia, invece, considerava il comportamento come una componente della costituzione fisiologica, e particolarmente neuropsicologica, dell'organismo, con un interesse specifico per la comprensione del suo significato adattativo e quindi evolutivo.
Uno dei pionieri dell'etologia negli anni Trenta del Novecento fu lo zoologo austriaco Konrad Lorenz. Studiando diversi animali, Lorenz sottolineò il ruolo adattativo dei comportamenti innati (o istintivi). Egli fondò la sua teoria sul ruolo che gli stimoli esterni hanno su quelle che definì 'pulsioni' dell'organismo. Man mano che l'organismo percepisce ‒ i morsi della fame, la vista di una preda o di un predatore, o di un possibile compagno ‒ questi stimoli si sommano aumentando la pulsione. In risposta all'aumento della pulsione, l'animale incrementa il livello di reattività nel suo ambiente, cercando (ma non consciamente in senso umano) il modo di rilasciare il surplus energetico accumulato. Quando l'animale trova una modalità adatta nel suo ambiente, reagisce di conseguenza, e la pulsione viene ridotta o eliminata. Tale modello spiegava non solo le risposte specifiche a stimoli specifici (alla vista di un compagno si reagisce con il rituale dell'accoppiamento), ma anche come stimoli specifici potessero provocare comportamenti alternativi o fuori luogo (l'animale cui è negato il rituale dell'accoppiamento reagisce cercando cibo). Lorenz sostenne inoltre che le reazioni istintive possono comunque avere anche una componente di apprendimento. Nel corso delle sue ricerche, osservò che i pulcini delle oche selvatiche creano un legame immediato con il primo organismo con cui vengono a contatto, che di solito è la madre (un processo detto 'imprinting').
Le piccole oche seguono da vicino la madre da cui hanno l'imprinting, senza mai staccarsi. L'imprinting ha un ovvio valore adattativo, dal momento che fornisce protezione e nutrimento ai giovani animali. Lorenz sottolineò la natura istintiva, meccanica, di questo processo, mostrando che, sostituendosi all'oca madre vicino alla covata nel momento della schiusa delle uova, l'imprinting avrebbe riguardato lui stesso. Secondo la teoria di Lorenz, la selezione naturale aveva creato la possibilità di avere un imprinting, mentre l'ambiente ne definiva lo specifico oggetto.
Lorenz era un fervente darwiniano che enfatizzava il ruolo della lotta e della 'sopravvivenza del più adatto' nelle società umane come in quelle animali. Sostenne anche che i diversi gruppi etnici e razziali, come specie animali differenti, ereditano diverse forme di funzioni mentali, simili agli 'istinti', alcuni dei quali sono più adattativi di altri. Queste idee lo portarono molto vicino alle teorie naziste sulla razza, che appoggiò nei suoi scritti del periodo nazionalsocialista (1933-1945). Nel dopoguerra l'etologia si sviluppò ulteriormente grazie agli studi dello zoologo olandese Nikolaas Tinbergen. Mentre Lorenz fu un aperto oppositore degli esperimenti sugli animali ‒ preferendo l'osservazione nei loro habitat naturali ‒ Tinbergen fece largo uso dei metodi sperimentali. Senza negare il valore della ricerca sul campo, Tinbergen dimostrò che la sperimentazione poteva rendere maggiormente rigorosi modelli generali come quelli sviluppati da Lorenz. I suoi primi lavori riguardarono una vespa velenosa (Philantus triangulum) che costruisce un elaborato intrico di tunnel nel terreno, rifornendolo con api che la femmina punge e paralizza per cibare le proprie larve. Per verificare se la vespa usasse la vista o l'olfatto per localizzare le api, Tinbergen pose un'ape paralizzata sul fondo di una provetta in un terrario chiuso, con un ventilatore sopra la provetta. Se la vespa avesse usato l'olfatto, ragionò Tinbergen, si sarebbe diretta verso l'estremità aperta della provetta, mentre nel caso avesse utilizzato la vista, sarebbe andata verso l'altro estremo. Tinbergen osservò che le vespe si dirigevano immediatamente controvento verso l'apertura della provetta, supportando l'ipotesi che fosse l'olfatto, e non la vista, che per primo permetteva alla vespa di localizzare la preda. Questi esperimenti erano semplici e chiari e allinearono l'approccio maggiormente naturalistico di Lorenz alla nuova enfasi che la biologia del XX sec. nel suo complesso poneva sulla sperimentazione come componente necessaria alla ricerca biologica.
Al contrario di Lorenz e Tinbergen, l'etologo americano Daniel Lehrman si oppose all'idea che i comportamenti ritualizzati, altamente specifici, fossero direttamente ereditati, o istintivi. Lehrman sottolineava che solamente i processi fisiologici ‒ per esempio, alcune reti neuronali, o la produzione di specifici ormoni ‒ possono essere ereditati; questi, in combinazione con i comportamenti casuali, con tentativi ed errori, che normalmente vengono esperiti dagli organismi, sarebbero alla base dell'apprendimento di alcuni comportamenti, a prima vista istintivi. Lehrman studiò sperimentalmente come i pulcini di tortora, subito dopo la schiusa, imparano a beccare dal gozzo della madre che contiene un liquido nutriente ('latte del gozzo'). Gli uccelli che covano producono un ormone, la prolattina, che stimola la produzione del latte, riempiendo e gonfiando il gozzo. Lehrman ipotizzò che i pulcini nascessero solo con il riflesso casuale dell'azione del beccare. Quando beccano accidentalmente il gozzo della madre, viene rilasciato il latte, che ricompensa non solo i pulcini ma anche il genitore in quanto, secondo Lehrman, il gozzo rigonfio risulta fastidioso. Lehrman verificò la sua ipotesi iniettando prolattina in due gruppi di uccelli: uno, che aveva già allevato almeno una covata (e quindi, presumibilmente, aveva imparato la risposta di rilascio) e uno che non aveva mai avuto questa esperienza. I risultati confermarono l'ipotesi di Lehrman: mettendo pulcini adottivi in entrambi i nidi, gli uccelli iniettati che già avevano cresciuto la prole iniziavano da subito a dar da mangiare ai pulcini, mentre gli uccelli 'inesperti' dovevano sentire lo stimolo dei pulcini prima di reagire. Per quanto dopo un decennio fosse poi dimostrato che il modello di Lehrman era eccessivamente semplificato, i suoi studi risultarono fondamentali per capire che i comportamenti, anche quelli cosiddetti istintivi, possono essere considerati una combinazione di risposte innate e apprese, passibili di studio sperimentale.
Negli anni Cinquanta e Sessanta del Novecento gli etologi affrontarono i comportamenti stereotipati che si trovano in molti organismi, dai platelminti (vermi) agli insetti (Drosophila, mantidi, mosconi e una miriade di specie di vespe) fino a un vasto gruppo di vertebrati. Il comportamento per l'accoppiamento, i comportamenti aggressivi o agonistici (minacce ostentate) e il comportamento rispetto al territorio vennero messi al centro dell'indagine etologica nel suo divenire un settore importante della biologia evolutiva. Ernst Mayr, tra gli altri, sottolineò l'importanza della comprensione del comportamento non solo nella storia evolutiva di una specie ma, come anche Oskar Heinroth, nella classificazione e nella definizione delle specie come taxa naturali.
L'etologia è stata applicata anche agli esseri umani, particolarmente negli scritti divulgativi di Lorenz (Das sogenannte Böse. Zur Naturgeschichte der Aggression, Il cosidetto male. Per una storia naturale dell'aggressione, 1963; Er redete mit dem Vieh, den Vögeln und den Fischen, L'anello di Re Salomone, 1952), di Robert Ardrey (The territorial imperative, 1966), e di Desmond Morris (African genesis, 1961). Questi studi diedero l'impressione che gli esseri umani si comportassero in base agli stessi 'istinti' ‒ come la territorialità e l'aggressione innata ‒ degli altri animali e che fenomeni come guerra, stupro o espansione territoriale fossero connaturati all'essere umano. Un corollario di questa teoria era che le tendenze animali negli umani non fossero eliminabili, ma solamente controllabili per mezzo dell'educazione e altre influenze 'civilizzatrici'. Queste affermazioni, basate soprattutto sull'analogia, furono duramente contestate da alcuni etologi, che criticarono l'estrapolazione acritica dagli animali agli umani, e dai sociologi e gli psicologi, che considerarono queste ipotesi contrarie alle prove disponibili che dimostravano l'importanza dell'apprendimento e la flessibilità, piuttosto che la fissità, del comportamento umano.
Sociobiologia
Nel corso degli anni Settanta del XX sec., la vecchia etologia veniva progressivamente allargata in un nuovo campo, la sociobiologia, che combinava lo studio del comportamento nel più ampio contesto della socialità umana, utilizzando concetti e metodi della genetica di popolazione, dell'ecologia, della statistica e delle simulazioni matematiche. La pietra miliare della sociobiologia è considerato il monumentale volume di Edward O. Wilson, Sociobiology, the new synthesis (1975). Biologo evoluzionista al Museum of Comparative Zoology di Harvard e specialista delle formiche, Wilson tentò di coordinare tutti gli aspetti della biologia animale in un unico paradigma darwiniano.
Il volume rappresentava una dettagliata rassegna della letteratura sul comportamento animale, la genetica, le strutture delle popolazioni e l'ecologia, in riferimento all'evoluzione di comportamenti complessi. Organizzato per gruppi tassonomici (in gran parte secondo i phyla), Sociobiology presentava un considerevole numero di prove a favore dell'evoluzione graduale di molti comportamenti animali ritualizzati (istintivi) come caratteri adattativi specie-specifici. Wilson mise insieme i nuovi risultati della genetica del comportamento animale, dell'ecologia evoluzionistica, dell'analisi strutturale delle popolazioni, della demografia e dei modelli di accrescimento delle popolazioni, con le proprie teorie sulla biogeografia delle isole (nella quale le piccole sottopopolazioni all'interno di specie ad ampia distribuzione sono considerate semiisolati insulari). Sintesi della letteratura disponibile in quel momento, il libro di Wilson raccolse larghi consensi.
Nell'ultimo capitolo del libro Wilson, tuttavia, applicava la sua 'nuova sintesi' all'evoluzione del comportamento umano. Nella prospettiva sociobiologica, il comportamento è una caratteristica innata di tutte le popolazioni umane (poiché tutti gli umani sono membri della stessa specie). Lo scopo di Wilson era quindi di spiegare ciò che considerava gli universali del comportamento umano, caratteristici della specie nel suo insieme. I comportamenti sono per una parte significativa controllati geneticamente e sono stati conservati dalla selezione naturale come utili adattamenti. Tra i caratteri considerati da Wilson ci sono la selezione del partner, la territorialità, il comportamento materno, l'indottrinamento, la religiosità e l'aggressività.
Wilson e i suoi seguaci, particolarmente Richard Dawkins in Inghilterra e Irven DeVore negli Stati Uniti, hanno anche invocato il controverso principio della 'selezione parentale', secondo cui la selezione opera sulla fitness del gruppo di consanguinei piuttosto che su quella del singolo individuo. La selezione parentale è stata utilizzata per spiegare il comportamento di una madre o di un altro stretto consanguineo che mettono a repentaglio la propria vita per salvare un bambino o un altro membro del gruppo parentale: quanto più DNA (geni) è condiviso dagli individui di un gruppo di consanguinei, tanto più grande sarà la possibilità che un individuo metta a rischio il proprio benessere a vantaggio di un parente. Il valore adattativo della selezione parentale sta dunque nel fatto che aiutando uno stretto parente (per esempio, un fratello o una sorella, con cui viene condiviso il cinquanta per cento dei geni) l'individuo sta in realtà aiutando i propri geni a perpetuarsi.
La teoria sociobiologica umana sottolineò in particolare la divisione del lavoro tra uomini e donne che Wilson e altri affermavano essere praticamente universale nelle società passate e presenti. Secondo questa ipotesi, i genotipi maschili erano stati selezionati per l'aggressività, le abilità di caccia, l'alta mobilità e la promiscuità sessuale. I genotipi femminili, d'altro canto, erano stati selezionati per la raccolta di cibo, la passività, la nutrizione e la cura dei bambini. Il concetto di 'scelta femminile' (cioè che le femmine scelgono i partner tra i maschi in competizione, un'idea basata sui modelli animali risalenti a Darwin) era invocata dai sociobiologi per suggerire che le femmine preservano i migliori genotipi della popolazione selezionando con cura quei maschi, tra i tanti disponibili, che faranno da padri alla propria discendenza. L'applicazione forse più controversa delle teorie sociobiologiche alle differenze maschio-femmina scaturì, alla fine degli anni Novanta del Novecento, dalla pubblicazione di A natural history of the rape (2000) dello studioso di comportamento animale Randy Tornhill, che interpretava lo stupro come naturale conseguenza (per quanto chiaramente indesiderabile) della selezione per l'aggressività maschile e per la competizione per le femmine. Gli studi di modelli animali (in particolare relativi alle anatre selvatiche, nelle quali apparentemente è stato descritto lo 'stupro di gruppo' di molti maschi contro una femmina) costituivano un significativo supporto delle affermazioni riguardo una presunta base biologica della pulsione allo stupro tra i maschi di ogni specie.
L'avvento della sociobiologia nel 1975, e la sua costante elaborazione, provocò notevoli controversie tra i biologi, i sociologi e gli antropologi. Di poco successiva all'accalorata discussione sulla razza e sulla presunta base genetica del Q.I. (correlata ai lavori dello psicologo di Berkeley Arthur Jensen e del fisico di Stanford William Shockley), che ebbe luogo tra il 1969 e il 1975, la sociobiologia ‒ in particolare le sue applicazioni all'uomo ‒ fu considerata l'ennesima forma di determinismo biologico, l'idea che il comportamento e la cultura umani siano in gran parte controllati dai geni. Le teorie sociobiologiche umane furono criticate perché considerate speculative, dal momento che Wilson e i suoi seguaci non avevano mai trovato possibili geni per le pratiche culturali. Gli specialisti (i genetisti di popolazione, gli antropologi, i sociologi, le femministe) ritennero i dati citati e le conclusioni nelle loro rispettive discipline esagerati, ingenui o semplicemente sbagliati. Un forte movimento antisociobiologia nacque negli Stati Uniti e in Europa, accusando Wilson e i suoi sostenitori di costruire una teoria in favore dello status quo politico, in particolare rispetto alla posizione delle donne nella società (Wilson sostenne che, nonostante l'istruzione e le accresciute opportunità nel mondo del lavoro, le donne avrebbero sempre avuto un ruolo minore degli uomini in campi come la scienza, la politica e il militare). Wilson e i suoi risposero che le critiche erano politiche, mentre i sociobiologi seguivano semplicemente le evidenze scientifiche. Non è ancora stata raggiunta una piena risoluzione della controversia, tuttavia per i biologi evoluzionisti e gli antropologi gli argomenti, e i dati riportati, in favore della sociobiologia umana erano ancora problematici. Negli anni Novanta, all'interno dei domini psicologici e psichiatrici una nuova teoria, nota come 'psicologia evoluzionistica', con tutte le caratteristiche della vecchia sociobiologia, ha iniziato ad attirare notevolmente l'attenzione. Il problema fondamentale che la maggior parte dei critici trovava nell'approccio sociobiologico al comportamento umano era la tendenza al riduzionismo nel correlare comportamenti complessi con l'azione diretta dei geni, mentre il percorso tra il gene e il comportamento è lungo e complicato, e in gran parte ancora da capire. Per parte loro, i sociologi e gli antropologi sostenevano che i fenomeni sociali andassero studiati a livello sociale e che non fosse possibile alcun tipo di correlazione lineare con elementi come i geni a un livello di organizzazione molto inferiore. I critici consideravano dunque particolarmente ingenua la conclusione di Wilson secondo cui tutte le materie umanistiche e le scienze sociali sarebbero arrivate a essere sussunte in un paradigma darwiniano e sociobiologico.
Genetica ed evoluzione umana
L'applicazione di alcuni aspetti della genetica all'evoluzione umana cominciò nella prima parte del secolo, principalmente con le analisi della distribuzione dei gruppi sanguigni nelle diverse popolazioni. Si scoprì che la distribuzione degli alleli dei gruppi sanguigni del più comune e diffuso sistema A-B-0 è molto differente tra i diversi gruppi umani: per esempio, gli africani hanno una frequenza maggiore (0,7) del tipo sanguigno 0 rispetto ai caucasici (0,6), mentre questi ultimi hanno una frequenza maggiore del tipo A (0,3) rispetto ai primi (0,18). Differenze più marcate si notano nel fattore Rh, per il quale gli africani hanno una frequenza di 0,73, i caucasici 0,16 e gli asiatici 0,04. I profili di distribuzione dei principali alleli dei gruppi sanguigni fornirono un mezzo per ricostruire le parentele ancestrali, i percorsi migratori e l'evoluzione delle popolazioni umane. Gli studi pionieristici in questo campo furono condotti da Ludwig e Hermann Hirschfeld nel 1918 e da Luigi Luca Cavalli-Sforza a partire dagli anni Sessanta del Novecento. Per diversi aspetti i risultati andavano in parallelo con le scoperte degli antropologi che usavano gli strumenti tradizionali (tracciando le migrazioni per mezzo dei manufatti, la linguistica, le tecniche edilizie e simili). A complicare le cose c'era ovviamente il costante incrocio tra le popolazioni umane, che in alcuni casi rendeva impossibile determinare se la presenza di certi alleli fosse il risultato di relazioni ancestrali o fosse stata introdotta da contatti recenti. Gli studi sui gruppi sanguigni mostrarono comunque le possibilità aperte dalla genetica per la comprensione sia delle migrazioni umane sia delle linee evolutive.
Con l'avvento della biologia molecolare, a partire dagli anni Cinquanta del Novecento con gli studi di Linus C. Pauling sulla struttura delle proteine, nuovi mezzi si resero disponibili per la ricerca sulle relazioni genetiche tra popolazioni. Se le proteine sono il diretto prodotto dei geni (come era stato stabilito dalle ricerche di George W. Beadle e Edward L. Tatum negli anni Quaranta), allora le differenze tra queste molecole possono essere utilizzate ‒ per inferenza a livello genetico ‒ per distinguere una popolazione dall'altra. Le tecniche cromatografiche (utilizzando colonne di polveri di cellulosa, o carte molto assorbenti) ed elettroforetiche (per mezzo di uno strato di gel al quale viene applicato un determinato voltaggio, creando un gradiente di carica elettrica) furono impiegate a partire dagli anni Quaranta e Cinquanta per separare composti di macromolecole come le proteine (e successivamente gli acidi nucleici). Le bande separate su una colonna cromatografica o di una elettroforesi su gel rappresentano ognuna una singola proteina, per esempio l'emoglobina, separata da un omogenato ottenuto dalla lisi delle cellule. La presenza o l'assenza di una banda può indicare se una proteina è presente o meno; piccole differenze nella posizione lungo la colonna o nella banda possono distinguere le diverse varianti di una stessa proteina. Quando Pauling, chimico del Caltech, applicò l'analisi elettroforetica alle emoglobine umane nel 1949, diede perciò inizio alla scoperta di un intero insieme di varianti, tra cui la più nota è l'emoglobina degli eritrociti falciformi. In questo caso la sostituzione di un singolo amminoacido in una delle due catene polipeptidiche causa un cambiamento drastico nella struttura e nella funzione (trasporto dell'ossigeno) della molecola. Successivamente, la scoperta che ciò rappresentava una mutazione in una coppia di basi del DNA degli individui affetti dalla malattia detta anemia falciforme fornì la prima prova della base molecolare di una malattia genetica umana. Il fatto che l'anemia falciforme fosse diffusa soprattutto, ma non solo, nelle popolazioni provenienti dall'Africa centroccidentale, appoggiò l'idea che le variazioni genetiche, come i gruppi sanguigni, fossero distribuite in modo disomogeneo tra le popolazioni umane e quindi potessero essere usate come misure delle parentele e delle linee evolutive. Il caso dell'emoglobina falciforme indicava anche che alcune differenze genetiche possono essere deleterie in alcuni ambienti e adattative in altri. In Africa centrale e occidentale, dove la malaria è endemica, gli individui eterozigoti per il gene falciforme (recessivo) hanno una resistenza alla malattia maggiore degli individui normali, che si riflette in una maggiore frequenza del gene mutante nelle popolazioni di quelle aree.
Un problema cui si sono dedicati i genetisti molecolari nella seconda metà del XX sec. è stato quello della razza. Argomento di contesa sin dal XVI sec., ha visto numerosi tentativi di distinguere biologicamente razze e gruppi etnici; i primi, fondati sulla distinzione di caratteri fisici come l'angolo facciale e il volume cranico non produssero risultati netti; da più parti si sperava che la genetica ne portasse di migliori. Il lavoro preliminare sui gruppi sanguigni sembrava promettente, ma esaminare soltanto tale caratteristica non corrispondeva al criterio sociale usuale di determinazione della razza, vale a dire il colore della pelle. La tecnica che si è dimostrata più utile al riguardo è stata il sequenziamento del DNA, per mezzo del quale le somiglianze e le differenze genetiche potevano essere confrontate a livello molecolare. I risultati di questi confronti dimostrarono tuttavia che tutti gli umani hanno il 99,8 % ca. del DNA in comune e che lo 0,2 % di differenze non è in relazione ai criteri standard come il colore della pelle, il tipo di capelli. La razza è dunque un concetto biologicamente privo di significato. Gli esseri umani si sono incrociati lungo tutta la loro storia evolutiva e pertanto l'idea che ci siano state razze 'pure', con una costituzione genetica unica e distinta, è considerata tipologica. Pur rimanendo un fondamentale costrutto sociopolitico, la razza non può essere considerata biologicamente reale.
Più costruttiva è stata l'utilizzazione dei dati molecolari, in particolare delle sequenze nucleotidiche del DNA, per tracciare l'evoluzione umana. I due maggiori paradigmi sull'origine geografica di Homo sapiens vennero sottoposti a verifica dopo il 1960, per mezzo delle sequenze di DNA ottenute dalle popolazioni odierne. Entrambe le ipotesi concordavano sull'evoluzione di Homo sapiens dalla specie ancestrale Homo erectus, erano invece in questione quando e dove ciò fosse avvenuto. L'ipotesi della 'sostituzione' o dell'origine africana sosteneva che Homo erectus migrò verso l'Europa, l'Asia e l'Australia all'incirca uno o due milioni di anni fa; successivamente, intorno ai 100.000 anni fa, la popolazione africana evolvette in Homo sapiens, intraprendendo una seconda migrazione e sostituendo le popolazioni di Homo erectus nelle altre aree (fig.14). L'ipotesi del differenziamento regionale affermava invece che Homo erectus migrò dall'Africa verso l'Europa e l'Asia uno o due milioni di anni fa, andando incontro parallelamente alla speciazione di Homo sapiens in ognuna di queste regioni, poiché vi era un continuo contatto (e flusso genico) tra le diverse popolazioni. Se l'ipotesi del differenziamento regionale fosse risultata corretta, le differenze genetiche fra i tre maggiori gruppi di discendenza (asiatico, caucasico, negroide) sarebbero state maggiori, poiché la divergenza sarebbe stata più lontana nel tempo; se fosse stata esatta l'ipotesi della sostituzione, le differenze genetiche tra i moderni gruppi umani sarebbero state considerevolmente minori. La strategia seguita fu quindi quella di confrontare le somiglianze e le differenze del DNA mitocondriale (usato perché, diversamente dal DNA nucleare, esso viene ereditato solamente dalla madre ed è quindi meno soggetto agli effetti dell'incrocio), tra rappresentanti odierni delle popolazioni asiatiche, africane ed europee. I risultati ottenuti si sono rivelati maggiormente compatibili con l'ipotesi della sostituzione.
Un nuovo campo di ricerca, noto come 'evoluzione molecolare' e relativo alla ricerca sull'evoluzione delle molecole a partire dalle loro forme ancestrali, è emerso a partire dagli anni Settanta del Novecento. L'emoglobina, per esempio, è considerata il frutto dell'evoluzione da elementi ancestrali simili ai citocromi, a causa di una duplicazione di un gene seguita dalla differenziazione di una delle due copie in una nuova direzione. L'introduzione dei computer ha considerevolmente aiutato questi studi, velocizzando il processo della comparazione tra le coppie di basi o delle sequenze amminoacidiche, alla ricerca di omologie e possibili scenari di divergenze.