
Le cure parentali
Le cure parentali
Le cure parentali comprendono l'insieme dei comportamenti dei genitori che servono a promuovere lo sviluppo e la crescita della prole e a incrementarne le probabilità di sopravvivenza e il successo riproduttivo finale. I vantaggi che derivano alla prole dalle cure dei genitori possono comportare per questi ultimi un certo costo in termini di riduzione della probabilità di sopravvivenza o della fertilità futura. A causa di questi costi, la selezione naturale agisce sui genitori assicurando, grazie a specifici meccanismi di riconoscimento, che essi si limitino a prendersi cura dei propri piccoli. Le cure parentali possono far sorgere conflitti tra i due genitori, tra i piccoli e forse anche tra questi e i genitori. In alcune specie esistono individui che aiutano i genitori a crescere i piccoli, traendone spesso un vantaggio indiretto in termini di fitness. L'acme dell'altruismo nelle cure parentali è rappresentato dalla completa sterilità delle operaie caratteristica degli insetti eusociali. Le cure parentali possono essere sfruttate anche da parassiti, che ottengono, con l'inganno, di far allevare i propri piccoli da altri individui della stessa o di altre specie.
Il concetto di cure parentali
La riproduzione è una componente necessaria della vita di un organismo. Tutti gli animali appartenenti a specie che si riproducono sessualmente hanno avuto due genitori che si sono riprodotti con successo, poiché è questo l'unico sistema per passare da una generazione alla successiva. Tuttavia, il riprodursi con successo non implica necessariamente l'esistenza delle cure parentali. Molti animali, specialmente tra le specie marine, si limitano a diffondere i propri prodotti sessuali (uova e spermatozoi) nell'ambiente circostante, dove le uova fecondate potranno svilupparsi con maggiore o minore successo senza mai venirne a contatto con i propri genitori. Tale strategia è utilizzata, per esempio, dalle aringhe, e si situa a uno dei due estremi di un continuum che comprende una grande varietà di strategie riproduttive, che vanno dal rilascio di molti minuscoli prodotti sessuali senza alcun contatto tra genitori e prole al contatto protratto nel tempo in cui le cure parentali prolungate sono rivolte a pochi individui di grandi dimensioni. Quest'ultimo caso è tipico dei mammiferi, incluso l'uomo, e degli uccelli, ma è comune anche nei vertebrati inferiori e in alcuni invertebrati.
Le cure parentali, nel caso per esempio di molti invertebrati, possono presentarsi anche sotto un'altra forma: quella di procurare il cibo per i piccoli che, tuttavia, non verranno mai a contatto con i propri genitori. Tale strategia riproduttiva si riscontra in molte api e vespe solitarie. In queste specie le femmine costruiscono celle dove accumulano provviste di cibo, sulle quali poi depongono le proprie uova. Quindi richiudono le celle e lasciano che la prole si sviluppi da sola. Una volta schiuse, le larve cresceranno nutrendosi di queste provviste. A causa della quantità di energia necessaria per portare a termine tale lavoro, le madri possono produrre solamente pochi piccoli 'ben riforniti'. Possiamo dunque distinguere le cure parentali indirette, che consistono nel procurare il cibo ai piccoli senza mai venire direttamente a contatto con essi, da quelle dirette, in cui genitori e prole interagiscono per un lungo periodo di tempo.
Non sempre è facile decidere se un certo comportamento sia o meno da comprendere tra le cure parentali, poiché in alcuni casi i piccoli possono trarre beneficio dalla semplice presenza di un adulto. In molti pesci, per esempio, i maschi difendono un territorio provvisto di siti appropriati per la deposizione delle uova e in tal modo attirano le femmine. Incidentalmente, il comportamento di difesa del territorio da parte del maschio consente la protezione delle uova e delle larve che ne derivano. Al fine di escludere casi simili, possiamo comprendere nella definizione in senso ampio di cure parentali la produzione di grandi uova ricche di tuorlo, le cure rivolte all'uovo fecondato entro o fuori il corpo del genitore, la costruzione di nidi, l'approvvigionamento dei piccoli (prima e dopo la nascita) e il supporto fornito loro dopo che hanno raggiunto l'indipendenza alimentare (Clutton-Brock, 1991).
In senso più stretto, le cure parentali comprendono la cura delle uova e dei piccoli (compreso il loro nutrimento) dopo la deposizione delle uova o dopo la nascita e, a volte, l'u1teriore supporto fornito alla prole dopo che ha raggiunto l'indipendenza alimentare.
L'espressione cure parentali è essenzialmente descrittiva e indica un comportamento genitoriale dal quale la prole può trarre qualche beneficio. Tuttavia esiste anche un concetto che insiste maggiormente sulla necessità di prendere in considerazione l'intero arco di vita degli organismi: si tratta del concetto di investimento parentale, introdotto da R.L. Trivers (1972). Esso sottolinea gli effetti del comportamento parentale in termini di fitness misurata generalmente come successo riproduttivo; il comportamento del genitore deve garantire alla prole un beneficio dimostrabile, ovvero incrementare le sue probabilità di svilupparsi, di crescere e, una volta raggiunta l'età adu1ta, di riprodursi, e le cure prodigate devono avere dei costi per i genitori e non devono costituire un effetto incidentale della loro semplice presenza o di comportamenti adottati per proprio vantaggio. Tale definizione prevede che i costi e i benefici vengano misurati in termini di fitness, prendendo in considerazione il successo riproduttivo dell' individuo nell' intero arco della sua vita. l vantaggi per la prole possono essere rappresentati da un incremento nelle sue probabilità di sopravvivenza fino all'età adulta e da un aumentato successo riproduttivo durante la fase fertile della sua vita. Tra i costi che i genitori devono pagare, quelli considerati più frequentemente sono la riduzione della fertilità e l'aumento della mortalità, che possono essere provocati dallo sforzo effettuato a vantaggio della prole. l concetti di cura parentale e investimento parentale sono, dunque, strettamente connessi tra loro e in questa sede saranno trattati insieme. Essi sono però differenti poiché, se l'ambiente è ricco di risorse e povero di predatori, cure parentali anche molto generose possono risultare virtualmente prive di costi per i genitori.
Le cure parentali come soluzione evolutiva ai problemi ecologici
Le cure parentali si sono evolute molte volte e in modo indipendente in diversi gruppi di animali. In alcuni di questi come gli uccelli e i mammiferi, esse appartengono al modello riproduttivo caratteristico e originario. In altri, come gli invertebrati, gli anfibi, i pesci e i rettili, costituiscono un'eccezione alla regola e si sono evo Iute indipendentemente in ciascuno di questi gruppi. Il fatto che le cure parentali si siano originate ripetutamente suggerisce che la loro comparsa abbia costituito una risposta a problemi di natura ecologica. Gli esempi che seguono descrivono diverse strategie di cure parentali all'interno di un unico gruppo e illustrano i problemi ecologici che molto probabilmente hanno portato alla loro evoluzione. Il dormiglione (Cancer pagurus), un granchio molto apprezzato per le sue carni, produce, come la maggior parte degli altri decapodi marini, più di un milione di piccole uova, con un diametro di 0,3 mm, che durante lo sviluppo vengono trasportate sul corpo della madre. Dalle uova si sviluppano minuscole larve, che vengono rilasciate in mare aperto e che per lungo tempo si nutrono e crescono nel plancton. Pochissime (meno dello 0,1 %) sono quelle che sopravvivono a questa fase così delicata della propria vita e che arrivano a far parte del benthos sul fondo del mare, dove si stabiliranno durante la fase giovanile; naturalmente ancora meno numerosi sono i granchi che sopravvivono sufficientemente a lungo da potersi riprodurre. Se si eccettua il breve periodo durante il quale le uova vengono trasportate sul corpo della madre, abbiamo qui un esempio di riproduzione in assenza di cure parentali. Il granchio delle bromeliacee (Metopaulias depressus) ha invece sviluppato una strategia completamente diversa, caratterizzata da numerose cure materne (Diesel e Schuh, 1993). Questa specie ha colonizzato le foreste montane della Giamaica centrale, dove si è adattata a vivere su grandi bromeliacee epifite, piante che raccolgono piccole quantità d'acqua piovana nei piccioli delle proprie foglie, allargati alla base a formare una sorta di piccola coppa. Le piccole piscine che si vengono a formare rappresentano una risorsa essenziale per la riproduzione e la sopravvivenza dei granchi in un habitat, per il resto, completamente secco. La madre granchio produce solamente circa 50 uova di grandi dimensioni, dal diametro di 1,2 mm, e le larve vengono poste in una di queste piscine, che può contenere circa 250 mI d'acqua. Grazie alla ricca riserva di tuorlo fornita loro dalla madre, le larve, che non sono ancora in grado di nutrirsi autonomamente, possono crescere affidandosi a questa sola risorsa (sono cioè lecitotrofiche). Tuttavia, la loro sopravvivenza è gravemente minacciata dalle condizioni ambientali sfavorevolmente acide (pH da 3 a 4), dal basso contenuto di calcio e di ossigeno nell' acqua e dalla predazione da parte delle voraci ninfe di zigottere. Una volta completata la metamorfosi, i giovani granchi possono essere predati dai ragni e per crescere hanno bisogno di una maggiore quantità di cibo di quanta gliene possa offrire la base fogliare allargata di una bromeliacea. Le condizioni biotiche e abiotiche piuttosto ostili hanno favorito perciò l'evoluzione di una vasta gamma di cure parentali, facilitata dalla possibilità di controllare agevolmente la prole, che si trova aggregata all'interno del territorio materno. La madre difende le larve dai predatori e provoca un aumento della concentrazione di ossigeno disciolto aerando l'acqua e ripulendo la piccola piscina dai detriti organici. Vi porta inoltre gusci di chiocciola ricchi di carbonato di calcio, utili per neutralizzare l'acqua e incrementarne il contenuto di calcio, condizioni entrambe essenziali per il successo della muta delle larve e per la formazione del loro nuovo carapace. l giovani granchi possono rimanere in questa sorta di asilo nido anche per due mesi e, troppo piccoli per andare a caccia, vengono riforniti dalla madre di prede come millepiedi e coleotteri, catturati in qualche altro punto della pianta.
Esperimenti condotti sul campo hanno dimostrato che la sopravvivenza e la crescita della prole sono sensibilmente incrementate quando i piccoli sono sottoposti a cure materne così generose e complesse. l costi del comportamento di cura parentale sono di solito più difficili da individuare rispetto ai suoi benefici. Nel granchio delle bromeliacee una cura prolungata della prole comporta un ritardo nella muta e nella crescita della madre e, poiché nelle femmine la fertilità aumenta con le dimensioni corporee, intense cure parentali provocano una riduzione della fertilità materna futura.
Come mostrato da questo esempio, le cure parentali possono costituire, nel corso dell'evoluzione, un'innovazione molto utile per colonizzare nuovi ambienti che, in quanto tali, presentano condizioni meno favorevoli allo sviluppo degli stadi precoci della prole. A stabilire quale ambiente possa essere definito inospitale è la storia naturale tipica di un determinato gruppo di animali. l crostacei decapodi vivono tipicamente in ambienti marini e le cure parentali, che sono evolute indipendentemente nei gamberi e in diversi granchi d'acqua dolce, si sono dimostrate fondamentali perché essi potessero colonizzare con successo sia habitat d'acqua dolce che habitat di terra. Negli onischi delle cantine, crostacei isopodi caratterizzati da un lungo passato evolutivo di vita terricola, le cure parentali sono essenziali. Un esempio è dato da Hemilepistus reamuri, una specie che ha colonizzato ambienti desertici ostili, caratterizzata da cure biparentali, cioè fornite da entrambi i genitori. In maniera simile, nei pesci teleostei le cure parentali si sono evolute diverse volte in risposta alla colonizzazione di habitat d'acqua dolce (Baylis, 1981). Condizioni ambientali avverse rispetto all'ambiente originario e rischi di predazione e di parassitismo costituiscono alcuni dei fattori principali che favoriscono lo sviluppo delle cure parentali.
Il carburante per l'ontogenesi
Uno dei problemi principali che gli animali, dagli invertebrati ai mammiferi, si trovano ad affrontare, è quello di fornire alla propria prole una quantità di risorse alimentari sufficiente a garantirne la crescita. Tale esigenza ha generato molteplici soluzioni nel corso dell'evoluzione. Un modello, comune e ampiamente diffuso, di investimento parentale precedente alla deposizione delle uova è quello nel quale le sostanze nutritive vengono direttamente fornite all'uovo o all'embrione dall'apparato riproduttivo materno. Ciò può realizzarsi, per esempio, dotando ciascun uovo di una grande quantità di tuorlo e rendendo così possibile il successivo sviluppo lecitotrofico delle larve, oppure fornendo alla prole sostanze nutritive derivate dal corpo dei genitori o provenienti dall'esterno.
Come accennato in precedenza, i genitori di molte specie procurano il cibo ai propri piccoli senza avere con loro alcuna interazione diretta. Per esempio, le femmine degli sfecidi appartenenti al genere Philanthus scavano nel terreno una galleria dotata di diverse celle-nido, dove depositano api immobilizzate. In seguito, depongono un uovo sopra questo cibo e chiudono le celle. La vespa vasaia, Euodynerus foraminatus, riesce persino a regolare la quantità di cibo in base al grado di competizione atteso all'interno di un determinato nido. Infatti, deposita assieme alle uova razioni maggiori di risorse alimentari quando è probabile che i piccoli dovranno affrontare un'elevata competizione per i partner sessuali e per i nidi, e che potranno trarre vantaggio dal raggiungimento di maggiori dimensioni corporee.
Altre specie fanno affidamento su risorse alimentari abbondanti, ma reperibili secondo criteri imprevedibili dal punto di vista spaziale e temporale. Un esempio è costituito dai coleotteri del genere Necrophorus. Quando un genitore trova una carogna, per esempio di un topo, la seppellisce, la difende e la lavora, trasformandola in una palla. La femmina vi depone poi accanto le proprie uova e quando, al termine del periodo di incubazione, queste si schiudono, nutre le larve (in alcune specie coadiuvata anche dal padre) rigurgitando pezzi di cibo prelevati dalla carogna (Trumbo, 1996). Senza l'aiuto di un genitore, o di entrambi, i piccoli non sarebbero in grado di monopolizzare questa risorsa alimentare essenziale per la sopravvivenza e lo sviluppo. Troppi competitori intra- e interspecifici utilizzano questa stessa risorsa e alcuni di essi depositano accanto alla carogna le larve anziché le uova, fornendo così alla propria prole un importante vantaggio in termini di tempo e di dimensioni, che le larve dei necrofori non potranno mai recuperare. Le cure parentali, nei necrofori, sono dunque essenziali per salvaguardare dai competitori le ricche ma effimere risorse alimentari destinate alla prole.
Negli anfibi, gruppo principalmente acquatico, si è evoIuta, in diverse occasioni indipendenti tra loro, una straordinaria gamma di cure parentali (Crump, 1996). Le fasi di sviluppo tipiche degli anuri includono uno stadio di uovo e uno stadio di girino, che costituisce un retaggio della loro lunga storia evolutiva legata all'ambiente acquatico. Il girino, infatti, è altamente adattato a nutrirsi e a crescere in acqua. Molte specie di anuri tropicali hanno evoluto complessi sistemi di cure parentali al fine di incrementare le probabilità di sopravvivenza dei piccoli in condizioni sfavorevoli nelle prime fasi del loro sviluppo.
Una condizione particolarmente avversa è costituita dall'abbondanza e dalla varietà di predatori caratteristica degli eco sistemi acquatici tropicali. I predatori minacciano gravemente la sopravvivenza delle uova e delle larve, che sono altamente nutritive e quasi completamente prive di difese. In risposta a tale pericolo, si sono evo Iute due diverse strategie: la cura e la difesa di uova e di girini nell'habitat acquatico, e il trasferimento degli stadi precoci di sviluppo dall'acqua ad ambienti maggiormente terricoli.
Leptodactylus ocellatus rappresenta un buon esempio di specie che hanno adottato la prima strategia. Le femmine producono un muco schiumo so con il quale formano il nido e ove depongono le uova; in seguito se ne prendono cura e lo difendono in maniera aggressiva dai potenziali predatori. Quando nascono, i girini rimangono gli uni accanto agli altri, muovendosi in una densa formazione aggregata, anch'essa difesa dalla madre. Nella pipa acquatica (Pipa pipa) la femmina trasporta le uova incapsulate nello spessore della cute del proprio dorso. Le giovani rane, ben protette in tale tasca cutanea, superano questa fase ontogenetica e completano la metamorfosi, per poi uscire e dare inizio alla propria vita indipendente.
Altri anuri tropicali si sono evoluti nella seconda direzione, adattandosi a modalità riproduttive maggiormente terricole. In questi casi, tuttavia, è necessario evitare la disidratazione di uova e girini, che non possono svilupparsi in assenza di acqua. I genitori del dendrobate pigmeo (Dendrobates pumilio) raggiungono tale scopo umettando ripetutamente le uova, quelli di Phyllomedusa duellmani dotandole di capsule che contengono esclusivamente acqua, quelli della rana africana (Chiromantis petersi) producendo un nido schiumo so su un albero; successivamente potranno trasportare i girini sul dorso fino a una sorgente. Nella rana terricola della Nuova Guinea (Cophixalus parkeri) sono principalmente i maschi a badare alla prole fino alla schiusa delle uova, per un periodo di circa 85÷100 giorni. In tal modo riducono sensibilmente il pericolo di predazione da parte di artropodi, eventuali episodi di cannibalismo e possibili infezioni micotiche. L'unica specie nota che si prenda cura dei piccoli è la rana giamaicana delle grotte (Eleutherodactylus cundalli), in cui l'intera covata, che può comprendere fino a 70 individui completamente sviluppati, viene trasportata sul dorso della madre dalla grotta in cui è nata fino alla foresta, dove potrà procurarsi il cibo in modo autonomo (Diesel et al., 1995). L'acme delle cure parentali nelle rane è stato raggiunto dalle specie che hanno evoluto un sistema di alimentazione della prole mediante sostanze prodotte dal corpo stesso del genitore. I maschi del rinoderma di Darwin (Rhinoderma darwinii), per esempio, inghiottono i girini appena usciti dall'uovo nella propria sacca vocale, dove si svilupperanno, nutrendosi di sostanze provenienti dai tessuti patemi, e da dove usciranno sotto forma di giovani rane. In maniera simile, in alcune specie del genere Gastrotheca, le uova si sviluppano entro piccole tasche dermiche che hanno evoluto una struttura simile a quella della placenta e che permette e regola lo scambio di gas, acqua ed escreti nel corso del lungo periodo di sviluppo che i piccoli trascorrono ben protetti dal corpo materno.
Altre specie che hanno colonizzato habitat terricoli depongono le uova in piccole pozze che si raccolgono nelle cavità degli alberi o nelle piante che accumulano acqua. Tuttavia, le risorse nutritive contenute in questi 'asili' non sono sufficienti per garantire che lo sviluppo dei girini abbia successo. La raganella arboricola giamaicana Osteopilus brunneus ha risolto tale problema in un modo molto elegante: le femmine depongono più di 100 uova in una di queste piccole piscine e, in seguito, vi fanno ritorno portando ai girini numerose uova (in alcuni casi anche 200), inizialmente fecondate ma diventate sterili in un secondo stadio del loro ciclo vitale (Crump, 1996). Un sistema di approvvigionamento dei piccoli ancora più complesso si è evoluto nel dendrobate pigmeo, le cui femmine trasportano e depositano ciascun girino in una piscinetta diversa. Le madri sono in grado di ricordare la postazione assegnata a ciascun girino e, in seguito, fanno ripetutamente ritorno in ognuno dei siti per fornire a ciascun piccolo una certa quantità di uova non fecondate. In entrambe queste specie la sopravvivenza dei girini dipende interamente dal cibo portato loro dalla madre.
La grande varietà di cure parentali evoluta negli anuri che si riproducono in ambiente terricolo dimostra una forte selezione per la cura delle uova, dei girini e persino delle giovani rane in ambienti in cui tali stadi di sviluppo sono fortemente minacciati. Un'altra strategia evolutiva consiste nel ridurre, nelle specie terricole, gli stadi di sviluppo poco adatti a tale ambiente. Tutte le specie appartenenti al genere Eleutherodactylus hanno perso completamente lo stadio di girino, evolvendo un percorso di sviluppo in cui dalle uova nascono direttamente giovani rane che hanno già portato completamente a termine la propria metamorfosi. Almeno una di queste specie (E. jasperi) è andata addirittura oltre, evolvendo l' ovoviviparità.
L'evoluzione delle cure parentali
Nonostante siano stati condotti numerosi studi, di natura tanto descrittiva quanto sperimentale, sul significato delle cure parentali nelle specie attualmente viventi, ben poco si sa dell'origine di questi comportamenti e dei motivi per cui in alcune specie essi sono prerogativa del sesso maschile e in altre di quello femminile. Data la loro grande varietà e le numerose linee evolutive indipendenti da cui derivano, è certo che le cure parentali si sono evolute seguendo molti percorsi diversi. A titolo di esempio, descriveremo solo uno dei loro possibili scenari d'origine. Per la maggior parte degli ovipari eterotermi l'origine delle cure parentali è probabilmente legata alla necessità di difesa, da parte di uno dei genitori, di territori provvisti di risorse disponibili in quantità limitata ma essenziali per la sopravvivenza e la riproduzione, come, per esempio, aree ricche di cibo, luoghi in cui trovare riparo o rifugio, siti adatti alla deposizione delle uova e aree deputate alle cerimonie di corteggiamento (Baylis, 1981). Se le uova o le larve, o entrambe, vengono a trovarsi entro il territorio di uno dei genitori, viene soddisfatto un importante prerequisito per l'evoluzione delle cure parentali: la possibilità, per il genitore, di controllare facilmente la prole (Clutton-Brock, 1991). l piccoli possono allora beneficiare di una certa protezione semplicemente per il fatto di trovarsi entro il territorio dei genitori. In una situazione di questo tipo si potrebbero essere trovati gli antenati del già citato granchio delle bromeliacee, le cui femmine difendevano probabilmente una bromeliacea, usandola come territorio di caccia e unica fonte d'acqua in un ambiente per il resto completamente secco. Le larve e i piccoli granchi, venendo si a trovare all'interno della base fogliare allargata di una di queste piante, potrebbero aver visto originariamente aumentate le proprie probabilità di sopravvivenza semplicemente grazie alla presenza della madre. In questa situazione, la madre non doveva probabilmente pagare alcun costo per proteggere la propria prole, ma in tal modo potrebbero essere state gettate le basi per l'evoluzione di uno dei più complessi sistemi di cure parentali. Scenari simili potrebbero spiegare l'origine delle cure parentali mostrate dalle rane che si riproducono in ambiente terricolo e dai pesci teleostei d'acqua dolce.
Per spiegare la distribuzione delle cure parentali tra maschi e femmine nei taxa caratterizzati da fecondazione esterna, sono state suggerite tre ipotesi: la sequenza temporale di emissione dei gameti, il grado di certezza di paternità e il livello di associazione del genitore con gli stadi embrionali. Queste ipotesi non si escludono reciprocamente e si basano tutte sul modello dell'abbandono del partner, mediante il quale si cerca di prevedere quale dei due genitori abbandonerà per primo la prole, lasciando all'altro la scelta tra il rimanere a badarvi da solo o l'abbandonarla a sua volta (Trivers, 1972).
R.L. Trivers suggerì che la fecondazione esterna, tipica per esempio dei pesci teleostei e delle rane, possa aver favorito l'evoluzione delle cure parentali. Nei pesci teleostei le cure paterne si sono evo Iute probabilmente come conseguenza della territorialità maschile. Se i maschi difendono un territorio che comprende i siti di deposizione delle uova, sono in grado di prendersi cura della propria prole potendo, tuttavia, continuare a corteggiare altre femmine e ad accoppiarsi con esse. Di conseguenza, il costo delle cure paterne in termini di ridotte opportunità di accoppiamento è basso (Trivers, 1972). Nei pesci le cure paterne sono molto più comuni di quelle materne. Si può ipotizzare che l'evoluzione delle cure parentali nei pesci abbia seguito una sequenza temporale in cui ciascuno stato è ancestrale al successivo: nessuna cura parentale, cure esclusivamente maschili, cure biparentali, cure esclusivamente femminili. Rimane tuttavia da comprendere, per esempio per quanto riguarda le rane, il motivo per cui, all'interno di una stessa famiglia o di uno stesso genere, alcune specie mostrino cure paterne e altre cure materne. l modelli teorici disponibili sono ancora troppo grossolani per spiegare come si siano evoluti tali ribaltamenti nelle strategie riproduttive. In tal senso potrebbero rivelarsi utili studi comparativi basati su una conoscenza maggiormente approfondita della storia naturale e dell' ecologia di queste specie nonché su una nozione più precisa delle affinità evolutive esistenti tra di esse.
Presupposti per le cure parentali nei vertebrati superiori
Le cure parentali costituiscono un lusso che i genitori riservano di solito esclusivamente ai propri figli; infatti, in genere, gli individui che non hanno figli reagiscono in maniera aggressiva agli eventuali approcci da parte di piccoli estranei. Avvicinandosi incautamente a una femmina che non sia sua madre, un piccolo può anche rischiare la morte. Per esempio, incidenti di questo genere si verificano nelle dense colonie di pinnipedi, nelle quali i piccoli possono essere maltrattati fino alla morte dalle femmine gravide o anche da quelle che stanno allattando i propri figli (Trillmich, 1981). Sembra dunque che le madri siano in grado di riconoscere la prole e il riconoscimento è un presupposto fondamentale affinché le cure parentali possano essere selettive: gli adulti devono stabilire un legame specifico con i propri piccoli e respingere quelli degli altri. Allo stesso tempo, quando hanno una propria prole, per poter assicurare le dovute cure, i genitori devono sviluppare verso i piccoli una certa sensibilità. Una forma molto semplice di 'riconoscimento' è osservabile in alcuni invertebrati. Come già detto, nei necrofori i genitori preparano per i figli la carogna di un piccolo vertebrato, quale per esempio un topo, facendone una palla che poi seppelliscono nel terreno. La femmina depone le uova accanto a questa ricca scorta di cibo e successivamente padre e madre attendono insieme in prossimità della carcassa che i piccoli compaiano. Sono necessari circa due giorni e mezzo affinché le uova si sviluppino in larve al primo stadio, che si avvicinano immediatamente alla carogna. Le femmine accettano le larve che si presentano da 8÷12 ore prima fino a 24 ore dopo il momento atteso. Queste larve vengono nutrite dai due adulti, che esse siano o no la loro vera prole. l giovani che invece compaiono al di fuori di questo intervallo di tempo vengono uccisi e consumati. l meccanismi fisiologici che regolano questo intervallo di accettazione sono ancora sconosciuti anche se risultano correlati a cambiamenti nei livelli ormonali giovanili della femmina.
Un esempio davvero estremo di repentino cambiamento motivazionale e fisiologico è quello della rana australiana Rheobatrachus silus (Tyler, 1983). In questa specie le femmine ingeriscono le uova appena fecondate e 'covano' i piccoli all'interno del proprio stomaco per un periodo di circa 5÷6 settimane, terminato il quale 'partoriscono' oralmente. Durante questo periodo le madri digiunano, probabilmente a causa dei segnali di sazietà inviati dallo stomaco notevolmente dilatato. Sembra che i piccoli siano protetti dalla digestione operata dai succhi gastrici della madre attraverso la secrezione di prostaglandine che inibiscono la produzione di acidi e riducono la motilità viscerale.
Negli uccelli e nei mammiferi con prole inetta, ovvero i cui piccoli rimangono immobili nel nido per un certo periodo dopo la nascita, i genitori possono inizialmente utilizzare, come indizio sufficiente per riconoscere la prole, semplicemente il luogo in cui essa si trova. Finché la prole rimane incapace di muoversi, è dunque possibile, in queste specie, scambiare tra loro i piccoli appartenenti a nidi diversi senza disturbare in alcun modo le cure parentali. Nella rondine delle rocce, poco prima dell'involo, i piccoli sviluppano richiami individuali che vengono appresi dai genitori. In queste famiglie, dunque, il riconoscimento della prole dipende inizialmente dalla sua posizione, ma in seguito i genitori imparano a riconoscere individualmente i propri figli indipendentemente da dove essi si trovino. In maniera simile i piccoli imparano a riconoscere i propri genitori. Ancora una volta si può individuare un intervallo di tempo critico entro il quale, in questo caso, si impara a riconoscere i singoli individui. Tale intervallo coincide esattamente con determinate fasi di sviluppo dei piccoli, per esempio con il periodo in cui iniziano a mostrare una certa capacità di locomozione.
La regolazione dell'intervallo di accettazione è stata studiata approfonditamente nella pecora (Levy et al., 1996). In questo caso il problema del riconoscimento si presenta del tutto diverso, poiché la prole è precoce, ovvero possiede alla nascita un apparato sensoriale e locomotore già ben sviluppato. I piccoli si muovono e camminano da subito; di conseguenza, se una madre non fosse in grado di riconoscere i propri figli, rischierebbe di confonderli con figli non suoi. Quando è in procinto di partorire, la femmina si allontana dunque dal gruppo in cui vive, garantendosi in tal modo un periodo sicuro nel quale imparare a riconoscere i figli. Un tale comportamento le permette, infatti, di apprendere le caratteristiche individuali del proprio piccolo immediatamente dopo la sua nascita, senza che vi sia alcun rischio di confonderlo con altri. Le pecore che non hanno figli, e persino quelle gravide, sono notevolmente aggressive verso gli agnellini. Immediatamente prima e al momento del parto, esse diventano invece molto sensibili nei confronti dei piccoli. Tale cambiamento è dovuto a modificazioni dei livelli di ormoni steroidei e alla stimolazione meccanica dell'apparato genitale durante il parto. Questi eventi stimolano il rilascio di ossitocina (lo stesso ormone che stimola la produzione del latte) che, a sua volta, induce nella madre un'immediata attrazione per il liquido amniotico. Madre e neonato sperimentano dunque immediatamente un contatto ravvicinato, poiché la madre inizia a leccare il piccolo ripulendolo dal liquido amniotico, asciugandone la pelliccia e cominciando in tal modo a prendersi cura di lui. Questo contatto rappresenta per la madre un'ottima occasione per apprendere l'odore del proprio o dei propri agnellini. Gli eventi che si susseguono nel corso del parto producono modificazioni neurofisiologiche nel bulbo olfattivo della madre, che le permettono di riconoscere con precisione le caratteristiche olfattive di ciascuno dei suoi piccoli. In tal modo viene stabilito, entro poche ore dalla nascita, un legame selettivo tra madre e figlio. In seguito, si svilupperà anche un riconoscimento di tipo acustico.
Una volta formato si il legame specifico tra madre e figlio, gli altri piccoli vengono sistematicamente respinti, persino se viene dato loro l'odore del vero figlio di quella femmina. Tuttavia, le madri possono essere indotte ad apprendere l'odore del piccolo di un'altra femmina, e dunque ad accettarlo, per mezzo di una stimolazione cervico-vaginale artificiale, che induca nuovamente l'iniziale responsività del sistema neuronale coinvolto in tale apprendimento. Le madri così stimolate accettano i piccoli che avevano respinto violentemente solo pochi minuti prima.
Una volta portato a termine questo apprendimento, simile all'imprinting, il legame altamente specifico stabilito si tra madre e figlio garantisce la selettività delle cure materne. L'intero processo si ripeterà l'anno successivo, quando la madre darà alla luce nuovi agnellini. Rispetto alle pecore che affrontano il primo parto, quelle multipare sembrano imparare molto più velocemente e sono distratte meno facilmente dagli eventi che possono verificarsi in prossimità del parto. Ciò suggerisce che i meccanismi neuronali coinvolti possono essere in qualche modo sensibilizzati dal primo parto.
Modelli di cure parentali nei vertebrati superiori
Gli uccelli
Gli uccelli rappresentano l'unico gruppo di animali in cui la monogamia sociale (sebbene non necessariamente genetica) costituisce la regola. In particolare nelle specie canore, come la cinciallegra (Parus major), ma anche in molti grandi uccelli marini, come i gabbiani, i genitori si occupano insieme della cova delle uova e provvedono a nutrire i nidiacei per un lungo periodo di tempo, fino alloro involo. Il maschio e la femmina possono contribuire in misura diversa alle necessità familiari, ma l'allevamento della prole è essenzialmente un'impresa comune. È molto frequente che all'involo segua un lungo periodo in cui i genitori continuano a prendersi cura dei piccoli, che sono autonomi solo per metà. Spesso, durante questo periodo, è per lo più il maschio a badare alla prole, permettendo in tal modo alla femmina di recuperare le risorse necessarie per poter deporre una seconda covata. Le cure parentali successive all'involo hanno ricevuto fino a poco tempo fa una scarsa considerazione, ma esse giocano un ruolo importante nel facilitare i piccoli verso una transizione graduale all'indipendenza alimentare e possono rivelarsi vitali per la loro sopravvivenza. Questo comportamento si riscontra tipicamente nelle specie di uccelli con prole inetta.
Uccelli come i polli, i fagiani e i trampolieri hanno, invece, una prole precoce che può essere allevata tanto da uno solo quanto da entrambi i genitori. Solamente alcuni gruppi di questi uccelli, e principalmente le oche e i cigni, allevano i figli in coppia. Nella maggior parte delle altre specie con prole precoce, i maschi non contribuiscono affatto alle cure parentali e le fennnine crescono i piccoli interamente da sole. Sebbene questi piccoli si nutrano spesso del tutto autonomamente fin dalla nascita, le cure che la madre deve rivolgere loro possono essere ugualmente molto impegnative. l piccoli richiedono inizialmente periodi di cova molto frequenti, perché i costi energetici che devono affrontare per la termoregolazione devono essere mantenuti entro limiti ragionevoli in un ambiente la cui temperatura è nettamente inferiore rispetto a quella corporea che negli uccelli è di circa 40,5 CC. Solo eccezionalmente i ruoli sono invertiti e sono i maschi a crescere da soli i piccoli, come avviene per esempio nella jacana e in alcune specie limicole come il piro-piro maculato (Actitis macularia).
I mammiferi
l mammiferi sono caratterizzati, tra le altre cose, dall'allattamento. Nel corso della vita di una femmina, il periodo dell' allattamento è quello più dispendioso dal punto di vista energetico. Il motivo per cui il latte tipico di una certa specie viene prodotto esclusivamente dalle femmine non è del tutto chiaro. Tale disparità fisiologica tra i due sessi nelle capacità di provvedere alla prole ha portato molto spesso all' evoluzione di sistemi di cure parentali esclusivamente fennninili e, di conseguenza, alla diffusione della poliginia come strategia riproduttiva tipica dei mammiferi. Le strutture sociali di tipo monogamico, nelle quali anche il maschio contribuisce alle cure parentali, costituiscono in questo caso più l'eccezione che la regola. Tra le poche specie di mannniferi che mostrano cure parentali da parte di entrambi i genitori vi sono gli sciacalli, i gibboni e i topolini delle risaie. Persino l'uomo, per il quale, nelle culture occidentali, la monogamia costituisce una regola morale, è moderatamente poliginico, e nella maggior parte delle società studiate dagli antropologi la poliginia viene accettata.
Anche nel caso dei mammiferi, come negli uccelli, vi sono specie con prole inetta e specie con prole precoce. La maggior parte dei roditori e dei carnivori appartiene al primo gruppo, mentre la prole precoce è tipica degli ungulati e dei caviomorfi, quali il porcellino d'India e le specie affini. l piccoli del primo tipo nascono sprovvisti di pelo, con gli occhi e le orecchie ancora chiusi; la loro sopravvivenza e la loro crescita dipendono in larga misura dalla presenza di un nido o di una tana che li protegga dai predatori e dal calore fornito loro dalla madre per la termoregolazione. Le figliate degli animali con prole inetta sono costituite da un maggior numero di individui rispetto a quelle delle specie precoci e, almeno inizialmente, crescono più velocemente, perché per potersi muovere e termoregolare richiedono un minimo sforzo metabolico. l piccoli delle specie precoci nascono, invece, con il pelo già completamente cresciuto e con gli occhi e le orecchie aperti; essi sono da subito in grado di muoversi e di termoregolarsi autonomamente dai genitori. In molti casi cominciano a seguire la madre immediatamente dopo il parto e, come accade nel caso delle antilopi delle steppe africane, la loro sicurezza dipende in larga parte dalla loro capacità di fuga e dal comportamento difensivo messo in atto dalla madre. l piccoli dei primati presentano una strategia intermedia. l nostri bambini nascono con abilità sensoriali già ben sviluppate, ma hanno bisogno di un tempo piuttosto lungo per poter sviluppare un'abilità locomotoria tale da permettere loro di seguire autonomamente gli spostamenti compiuti dagli adulti. In molti mammiferi, e in particolare nelle specie di maggiori dimensioni, le cure parentali proseguono ancora a lungo dopo lo svezzamento dei piccoli. Può trattarsi semplicemente della condivisione di un territorio, come accade per esempio negli orsi bruni, i cui piccoli possono continuare a utilizzare il territorio materno per un periodo di tempo molto lungo prima di doversi, al momento della dispersione, trasferire altrove. Ma queste cure prolungate possono anche manifestarsi sotto forma di interazioni comportamentali molto elaborate, come nel caso di molte specie di primati che vivono in gruppi, in cui le figlie ereditano il rango della madre e sono da essa attivamente sostenute nelle situazioni di competizione. Questo comporta la formazione di legami di solidarietà familiare in cui tutte le figlie di una determinata madre, e tutti i loro piccoli, si sostengono a vicenda e possono in tal modo arrivare a dominare tutte le altre femmine del gruppo. Una struttura sociale di questo tipo è stata osservata in molti macachi, babbuini e cercopitechi, le cui fennnine rimangono nel gruppo in cui sono nate mentre i maschi se ne allontanano quando raggiungono la maturità sessuale o poco prima.
L'investimento parentale
Sebbene sia facilmente dimostrabile come, negli uccelli e nei mannniferi, i piccoli traggano beneficio dalle cure parentali che ricevono - poiché, se ne vengono privati, essi non si sviluppano adeguatamente - è molto più difficile individuare i meccanismi che producono i già menzionati costi di tali comportamenti per i genitori. Naturalmente la nutrizione dei piccoli implica dei costi energetici piuttosto ovvi; basti pensare al portare cibo al nido nel caso degli uccelli e alla produzione del latte nei mannniferi. Si potrebbe pensare che, una volta terminato il periodo in cui è necessario prendersi cura della prole, i genitori siano in grado di riprendersi e di ritornare nelle stesse condizioni in cui vivevano prima dell'evento riproduttivo. È stato tuttavia dimostrato, in diversi casi, che un genitore che si sia preso cura della prole vede notevolmente ridotte le proprie ulteriori possibilità di riproduzione. Uno dei possibili meccanismi alla base di tale tendenza è quello per cui i genitori che provvedono a procurare il cibo necessario ai propri piccoli presentano un accresciuto tasso di mortalità. Essi devono infatti trascorrere una maggiore quantità di tempo in luoghi potenzialmente pericolosi, dove possono imbattersi nei predatori.
Oltre a questi effetti diretti sulla mortalità, sono stati di recente dimostrati sperimentalmente alcuni meccanismi fisiologici che possono essere responsabili della riduzione del futuro successo riproduttivo dei genitori. Uno di essi consiste in un'alterazione delle funzioni immunitarie (Sheldon e Verhulst, 1996). Per esempio, nel diamante mandarino (Taeniopygia guttata) il sistema immunitario dei genitori è risultato essere tanto più indebolito quanti più nidiacei essi dovevano nutrire e, naturalmente, questo può rendere i genitori maggiormente suscettibili a parassiti e malattie. La ridotta resistenza alle infezioni e ai parassiti può, a sua volta, aumentare il tasso di mortalità e ridurre la fertilità. Infine, prendersi cura dei piccoli può ridurre la fertilità dei genitori al momento del successivo evento riproduttivo, in quanto, come è stato dimostrato per la cinciarella (Parus caeruleus), questo comportamento può interferire con la normale muta del piumaggio. l genitori che si trovano a dover badare alla propria prole a stagione inoltrata (per esempio, nel caso di una seconda covata) devono poi accelerare il processo di muta del proprio piumaggio e devono fado proprio nel periodo dell' anno in cui la disponibilità di cibo va gradualmente diminuendo. Una simile circostanza può avere come conseguenza la crescita di penne meno sviluppate e quindi di un piumaggio non sufficientemente isolante. Gli adulti di questa specie si troveranno allora a dover subire nel corso del gelido inverno gli effetti negativi di tali condizioni. Rispetto ai propri conspecifici dotati di un buon piumaggio, essi dovranno costantemente spendere una maggiore quantità di energia per la propria termoregolazione e si troveranno, dunque, ad affrontare la successiva stagione riproduttiva in condizioni tutt'altro che ottimali.
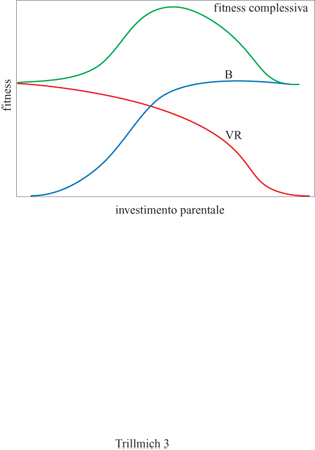
Come accennato in precedenza, a questo compromesso tra sforzo riproduttivo e riduzione del successo riproduttivo futuro (fig. 3) è stato dato il nome di investimento parentale (Trivers, 1972). l genitori devono risolvere il problema di come poter aumentare la fitness della prole mediante le cure parentali cercando, tuttavia, di ridurre al minimo la diminuzione delle proprie capacità di riprodursi con successo nel futuro. Il maggior guadagno netto in termini di fitness si ha quando la somma del successo riproduttivo riferito alla prole attuale e del rendimento riproduttivo atteso per il futuro raggiunge il valore massimo. Che la riproduzione abbia un certo costo sembra inevitabile, poiché le risorse disponibili per i genitori non sono illimitate e, se pure lo fossero, essi non avrebbero le illimitate capacità fisiologiche necessarie per servirsene in modo da incrementare il proprio rendimento energetico nel corso delle cure parentali. Questa situazione porta a un bilanciamento tale per cui destinare una maggiore quantità di risorse a una funzione determinata, come la cura della prole, ha come conseguenza necessaria la riduzione delle risorse disponibili per un'altra funzione quale la crescita e il mantenimento delle funzioni corporee del genitore o delle sue difese immunitarie.
I conflitti suscitati dalle cure parentali
Considerando il tipo di dinamica coinvolta nell'investimento parentale, è lecito attendersi che la preziosa risorsa costituita dalle cure parentali susciti conflitti tra le diverse parti in causa. La teoria prevede che conflitti di questo tipo sorgano tanto tra i due partner, quanto tra essi e la loro prole e infine tra i piccoli stessi. Come si manifestano tali conflitti nella vita reale degli animali e come influenzano lo svolgimento delle cure parentali?
Il conflitto tra genitori
Quando osserviamo una coppia di storni che nutre i propri nidiacei, siamo colpiti dalla grande frequenza con cui entrambi i genitori ritornano al nido e riempiono il becco spalancato dei piccoli con ciò che hanno portato con sé. Entrambi sembrano lavorare al massimo delle proprie possibilità, specialmente quando i piccoli sono un po' cresciuti e hanno bisogno di una grande quantità di cibo per il metabolismo e la crescita. Tuttavia, tanto le considerazioni di natura teorica quanto i risultati del lavoro sperimentale dimostrano che, negli uccelli, i genitori, quando lavorano insieme, forniscono meno cibo ai propri piccoli che non quando se ne occupano singolarmente. Se si incolla un pezzetto di piombo alle penne della coda di un maschio di storno, esso, trovandosi in qualche misura ostacolato nei movimenti, riduce il proprio contributo alla nutrizione dei nidiacei. La femmina aumenta allora la propria frequenza di imbeccata, ma mai al punto da compensare completamente il mancato contributo del maschio. Ciò suggerisce che essa, quando il suo partner era ancora in grado di dare il proprio normale contributo, non stesse fornendo il massimo sforzo possibile. Quando uno dei due partner viene a mancare del tutto, l'aumentato impegno del genitore rimasto solo è ancora più evidente. Improvvisamente esso arriva quasi a raddoppiare i propri sforzi per fornire ai nidiacei una quantità di cibo sufficiente. Il fatto che i piccoli nutriti da entrambi i genitori non presentino in genere il massimo tasso di crescita suggerisce che, in circostanze normali, i due partner non lavorino al massimo delle proprie possibilità per permettere ai propri piccoli la massima crescita. Ciascuno lavora invece solamente quanto basta perché i nidiacei possano crescere a un tasso medio. Si tratta del risultato di una strategia evolutivamente stabile, frnalizzata a impedire che uno dei due membri della coppia parentale venga sfruttato dall'altro. È più facile comprendere la ragione per cui i genitori limitano i propri sforzi nel nutrire i piccoli se si ricorda che le cure parentali hanno dei costi: la selezione naturale favorisce in entrambi i partner un impegno limitato nel crescere la covata attuale, allo scopo di non mettere eccessivamente a repentaglio il loro successo riproduttivo futuro. Se uno dei due rispondesse alla diminuzione degli sforzi effettuati dall'altro compensando con un corrispondente aumento della propria mole di lavoro, frnirebbe per essere sfruttato dal compagno che, nel caso estremo, potrebbe addirittura abbandonare del tutto la prole attuale e cercarsi altrove un secondo partner con cui produrre un'altra covata. La strategia evolutivamente stabile consiste dunque, molto semplicemente, nel fare una quantità di lavoro che sia appena al di sotto del livello richiesto per la crescita ottimale della prole e nel compensare solo in parte l'eventuale riduzione del contributo del proprio partner, per far sì che esso continui a lavorare.
Gli studi di N.B. Davies (1992) sulla passera scopaiola (Prunella modularis) esemplificano forse più chiaramente come gli uccelli possiedano la capacità decisionale che serve loro per regolare gli sforzi frnalizzati alla nutrizione dei piccoli in base ai vantaggi attesi in termini di fitness da una determinata covata. In questa specie, in primavera, i maschi e le femmine delimitano i propri territori indipendentemente gli uni dalle altre. In seguito, i maschi riaggiustano i confini del proprio territorio in base a quelli delle femmine in modo che un territorio maschile comprenda al proprio interno più territori femminili. Il maggior successo riproduttivo è realizzato dai maschi poliginici. l loro approcci vengono respinti dalle femmine, poiché esse ottengono un maggior successo riproduttivo accoppiandosi con due maschi. Infatti, le covate nutrite da due maschi e una femmina sono quelle caratterizzate dal maggiore successo di involo. Le femmine cercano dunque di stabilirsi con più di un maschio (poliandria), mentre ciascuno di essi difende le proprie femmine dagli altri maschi, sorvegliandole con un comportamento aggressivo specialmente durante il periodo della deposizione delle uova. l maschi regolano quindi il proprio contributo alla nutrizione dei nidiacei in base all'accesso che hanno avuto alla femmina per potersi accoppiare con lei durante il periodo fertile. Se vengono tenuti lontani dalla femmina durante questo periodo, non daranno, in seguito, alcun contributo all'allevamento della prole e potranno addirittura tentare di distruggere la nidiata allo scopo di ottenere un nuovo accoppiamento. Questo esempio mostra quanto elevata sia la capacità degli uccelli di regolare finemente la quantità di cure parentali da prestare e come possa essere intenso, all'interno di una famiglia, il conflitto provocato tra i due sessi dalla necessità di prendersi cura della prole.
Il conflitto tra piccoli
Analoghi conflitti di interessi sono stati descritti, in molte specie, anche tra i fratelli appartenenti a una stessa covata. Questo fenomeno va dalla moderata competizione per il cibo al cannibalismo vero e proprio, quale è stato osservato per esempio nei girini di alcuni anfibi anuri, tra i quali si può sviluppare un morfotipo cannibale di grandi dimensioni che, sebbene preferisca nutrirsi di girini con cui non è imparentato direttamente, può cibarsi anche dei propri sfortunati fratelli più piccoli. Un piccolo che possa monopolizzare le cure parentali o accaparrarsi la maggior parte di questa risorsa preziosa crescerà meglio e avrà, in seguito, maggiori probabilità di entrare a far parte della fetta di popolazione che si riproduce con successo. Di solito i conflitti tra parenti stretti sono limitati, perché essi hanno in comune una buona parte del patrimonio genetico. È dunque probabile che i geni che favoriscono strategie comportamentali che riducono le probabilità di sopravvivenza di un parente stretto finiscano per limitare anche la propria diffusione (Hamilton, 1964). Tuttavia, nonostante l'influenza moderatrice della parentela, intense lotte tra fratelli sono state osservate in molti uccelli da preda, sule ed egrette, e anche in mammiferi come i maiali, gli arctocefali delle Galapagos e le iene (Mock e Parker, 1997). Sembra che i maialini abbiano addirittura evoluto armi specializzate, come canini funzionali e incisivi simili a canini, che, presenti fin dalla nascita, puntano lateralmente e vengono utilizzati per disputarsi il possesso dei capezzoli anteriori della madre, che producono la maggior quantità di latte.
Il conflitto tra fratelli è stato studiato soprattutto negli uccelli. In alcune specie di aquila la nidiata è composta da due uova, ma il piccolo che nasce per primo uccide quasi sempre il proprio fratellino minore, sia attaccandolo direttamente sia impedendogli di partecipare alle imbeccate genitoriali. Questo conflitto è più intenso poco dopo la schiusa, ovvero in una fase in cui l'apporto di cibo necessario non è ancora tale da rischiare di limitare la crescita del giovane aquilotto. In questi casi, il secondo uovo sembrerebbe costituire più che altro una forma di assicurazione nell'eventualità in cui il primo non dovesse schiudersi. Presumibilmente, per specie che vivono a lungo e che maturano lentamente come queste, è più probabile riuscire ad allevare con successo un unico piccolo, che sia per così dire 'di prima qualità', piuttosto che anche uno solo di due piccoli di qualità inferiore. In specie in cui l'aggressività tra fratelli è molto meno forte, come nelle egrette, la disponibilità di cibo, che i piccoli valutano in base alla frequenza delle imbeccate che ricevono dai propri genitori, influenza enormemente i risultati delle rivalità tra nidiacei. Quando il cibo è abbondante, gli ultimi nati resistono più facilmente agli attacchi dei propri fratelli maggiori perché questi ultimi sono sufficientemente sazi da permettere loro più spesso di nutrirsi indisturbati. Anche le tecniche adottate dai genitori per nutrire i nidiacei influenzano notevolmente l'intensità dei contrasti tra fratelli. Un esempio evidente in tal senso è fornito dal confronto tra l'egretta comune e l'airone blu (Ardea herodias). Nell'egretta, la cui prole viene nutrita con boli derivati dalla predazione di piccoli pesci, i nidiacei di dimensioni maggiori possono arrivare al becco del genitore e prenderne direttamente il cibo, monopolizzandolo e privandone così i fratelli più piccoli. Gli aironi blu si nutrono invece di pesci più grandi e rigurgitano la preda sul fondo del nido dove i nidiacei devono darsi da fare per staccarne piccoli pezzi. In tale situazione è impossibile monopolizzare il pasto e la competizione per il cibo è dunque molto meno forte in questi nidi che non in quelli delle egrette (Mock e Parker, 1997).
Il conflitto tra genitori e figli
Un forte conflitto tra fratelli sembra contrario agli interessi dei genitori, i quali dovrebbero presumibilmente trarre maggiore beneficio dal produrre una nidiata numerosa piuttosto che dal crescere singoli individui al suo interno. Di conseguenza, sembrerebbe ragionevole attendersi che essi interferiscano nei contrasti tra nidiacei. Tuttavia, le osservazioni condotte fino a oggi non sono riuscite a fornire molte prove di tali interferenze neanche quando i contrasti avevano esiti letali.
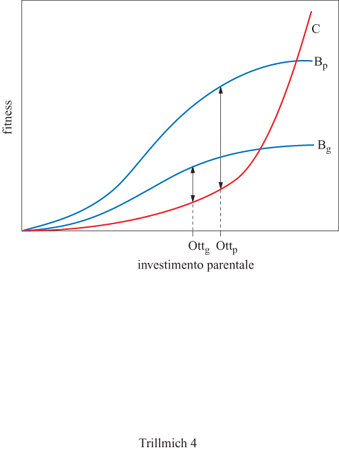
Esiste però un aspetto delle cure parentali quotidiane, molto meno drammatico, per il quale è stata ipotizzata l' esistenza di un conflitto tra genitori e figli. Si tratta dell'interazione tra genitori che portano il cibo e piccoli che lo reclamano (Kilner e Johnstone, 1997). Quando, negli storni, i genitori nutrono i propri nidiacei, questi ultimi cercano di avvicinarsi loro il più possibile, emettono intense vocalizzazioni, allungano il collo perché il proprio becco si trovi il più possibile vicino al loro e lo spalancano anche per farsi notare di più. La colorazione gialla, particolarmente accesa, presente sul bordo del loro becco, diventa un segnale molto efficace. Non è del tutto chiaro cosa stia accadendo: i piccoli stanno segnalando ai genitori i propri bisogni reali, oppure ciascun individuo sta cercando di ottenere qualcosa di più della giusta razione di cibo? Teoricamente, potrebbe sorgere un conflitto tra genitori e prole in merito alla quantità di cure parentali prodigate a un particolare nidiaceo (fig. 4). Questo avviene perché la selezione naturale favorisce i genitori che si prendono cura di tutti i piccoli allo stesso modo, avendo con ciascuno di essi lo stesso grado di parentela. Ciascun nidiaceo, invece, darà maggior importanza alle cure parentali a lui stesso prodigate che a quelle rivolte a uno qualunque dei suoi fratelli. Infatti, un piccolo condivide con ciascuno dei propri fratelli, per via della discendenza comune, solamente la metà dei propri geni, ma è ovviamente in grado di trasmettere alla generazione successi va il proprio patrimonio genetico per intero. D'altro canto, genitori e figli danno la stessa importanza ai costi che un determinato investimento parentale comporta per gli adulti. Infatti, tali costi implicano un minore successo riproduttivo del genitore, la cui futura prole ha lo stesso valore per entrambe le parti in causa (in condizioni di monogamia) che con essa condividono, in ogni caso, la metà dei geni. Tale differenza nella tendenza evolutiva di genitori e figli nel valutare i benefici delle cure parentali ha condotto Trivers (1974) a ipotizzare l'esistenza di un conflitto tra le due parti (v. figura 4).
In alternativa, è stato proposto che i genitori possano non essere in possesso di tutte le informazioni necessarie circa lo stato dei piccoli perché, pur essendo in grado di vedere e stimare le loro dimensioni e forse le loro condizioni, hanno difficoltà a sapere quanto siano davvero affamati. E allora, quando osserviamo il nostro nido di storni, vi è da chiedersi se stiamo assistendo a una forma di comunicazione tra genitori e prole, oppure a un conflitto tra le due parti, in cui ciascun nidiaceo sta cercando di ottenere più di quanto gli spetti e i genitori devono fare attenzione a smascherare e a vanificare i loro espedienti. Sono stati studiati molti casi di presunto conflitto tra genitori e figli ma i risultati di questi lavori rimangono ambigui. Un'ipotesi che ha ricevuto esaurienti conferme sperimentali è quella secondo cui il comportamento di richiesta di cibo da parte dei piccoli indica effettivamente i loro bisogni e influenza, di conseguenza, il comportamento di nutrizione da parte dei genitori. Poiché il comportamento di richiesta di cibo da parte dei piccoli comporta dei costi, sarebbe poco conveniente per loro utilizzarlo al fine di perpetrare un inganno. Tale segnale dovrebbe dunque essere un indicatore attendibile della fame dei nidiacei. Tuttavia, i piccoli utilizzano molteplici segnali (acustici, visivi, prossimità al genitore e grado di allungamento del collo) per comunicare il proprio desiderio di essere nutriti e non è ancora chiaro che relazione vi sia tra questi loro comportamenti reali e i modelli teorici relativamente semplificati oggi disponibili (Mock e Parker, 1997).
È stato suggerito che il conflitto tra genitori e figli sia difficile da osservare perché gli adulti, a causa della propria posizione di potere rispetto ai piccoli, possono imporre alla prole le proprie decisioni. Tuttavia, come già accennato, osservazioni dettagliate condotte per esempio sull'egretta e sulla sula dai piedi azzurri (Sula nebouxii) non hanno fornito alcuna prova del fatto che i genitori interferiscano realmente nei conflitti tra fratelli, anche quando questi possono avere esiti letali (Mock e Parker, 1997). Un'interferenza sarebbe invece attesa, in questi casi, se genitori e prole fossero in conflitto circa la soglia di cibo sotto la quale il tasso di crescita dei nidiacei si ridurrebbe al punto da rendere necessario lasciar morire uno dei fratelli minori per permettere la sopravvivenza dei fratelli maggiori. In caso contrario, ovvero se genitori e prole si trovassero d'accordo sulla necessità di ridurre il numero di individui presenti nella covata, le lotte tra nidiacei sarebbero tollerate, al fine di permettere l'eliminazione del fratello più debole.
Molti comportamenti osservati che sono stati ascritti all'esistenza di un conflitto tra genitori e prole possono forse essere meglio interpretati come forme di comunicazione circa le proprie condizioni fisiche e quelle dell' ambiente (Bateson, 1994; Kilner e Johnstone, 1997). Durante la fase di svezzamento nei mammiferi, per esempio nel cercopiteco verde (Cercopithecus aethiops), in presenza di cibo abbondante, le madri in condizioni fisiche eccellenti sono in grado di far crescere i propri piccoli molto velocemente, a tassi di crescita quasi massimi, per poi svezzarli presto senza che possa essere osservato alcun conflitto tra madre e prole. Quando le risorse alimentari sono più scarse o quando la madre si trova in condizioni fisiche peggiori, per esempio a causa dell'età o di un'infestazione da parassiti, paradossalmente i piccoli vengono svezzati molto più tardi. Ciò potrebbe essere causato da una maggiore difficoltà nel trasferimento di energia e di sostanze nutritive dalla madre ai piccoli, che porterebbe questi ultimi a svilupparsi più lentamente posticipando il raggiungimento dell'indipendenza. Ma le madri in condizioni fisiche decisamente cattive, o che si trovano in un ambiente estremamente povero di cibo, svezzano anch'esse i propri piccoli precocemente. Possono addirittura smettere di prendersene cura quando essi sono ancora talmente piccoli da avere, in assenza di cure materne, poche, o nessuna, possibilità di sopravvivenza. Nell'insieme, queste osservazioni suggeriscono che, finché è permesso dai limiti imposti dall'ambiente, le madri e i piccoli adattino le proprie interazioni alle condizioni fisiche materne e ai condizionamenti ambientali. Ciononostante, è chiaro che esiste un conflitto tra gli interessi della madre e quelli della prole e questo diventa ancora più evidente nei casi in cui la femmina smette di prendersi cura di piccoli ancora fortemente dipendenti da lei. In questo caso, la madre di cercopiteco preferisce perdere un piccolo pur di evitare di pagare un costo eccessivamente elevato in termini di investimento parentale che potrebbe concretizzarsi in una fertilità futura ridotta o addirittura in un aumentato rischio di mortalità.
Nei casi in cui le condizioni della madre e della prole variano in maniera imprevedibile con il variare delle condizioni ambientali, la comunicazione circa lo stato del genitore e quello della prole dovrebbe essere onesta ma costosa per evitare il rischio dell'inganno. In caso contrario, un figlio perfettamente in grado di badare a se stesso potrebbe far credere alla madre di essere invece ancora molto piccolo e dipendente da lei, allo scopo di ottenere un po' più di latte e di protezione e quindi poter crescere più in fretta. La madre dovrebbe dunque costringere i propri piccoli a segnalare in maniera il più possibile inequivocabile di avere ancora bisogno delle sue cure e può farlo, per esempio, mostrando di respingerli. Un simile comportamento è particolarmente evidente negli arctocefali delle Galapagos (Arctocephalus galapagoensis) in cui le femmine spesso abbandonano il proprio neonato se il figlio che hanno avuto l'anno precedente è ancora troppo piccolo per sopravvivere da solo. Tuttavia, in questi casi, le madri si comportano di solito in maniera molto aggressiva verso i propri figli, minacciando li, mordendo li e allontanandoli violentemente dal neonato. Solo i giovani di dimensioni minori, che non superano i 14 kg (la metà di una femmina adulta), resistono a queste aggressioni e continuano a lottare per avere accesso al latte materno, comunicando in tal modo il proprio urgente e reale bisogno di ulteriori cure. I figli di maggiori dimensioni, invece, di fronte a una simile aggressività materna desistono immediatamente svezzandosi all'istante. In questi casi sembra dunque che i figli segnalino correttamente alla madre il proprio bisogno di aiuto poiché quelli più piccoli non sono davvero in grado di procurarsi il cibo da soli. Con una massa corporea ridotta, non possono nuotare bene come gli altri e non riescono a nutrirsi abbastanza per mantenere il proprio peso, né tantomeno per assicurarsi un'ulteriore crescita (Horning e Trillmich, 1997).
Il comportamento di helping
Sorprendentemente, in alcune specie esistono individui che collaborano all'allevamento di figli che non hanno generato. Tale fenomeno è chiamato helping e comprende comportamenti quali il nutrire o il proteggere i piccoli. Se le cure parentali suscitano tanti conflitti, perché mai un individuo dovrebbe spendere le proprie energie a favore di piccoli altrui? I migliori esempi di questo fenomeno vengono ancora una volta dagli uccelli (Brown, 1987). Sono state proposte principalmente due spiegazioni per l'helping; la prima, applicabile per esempio al comportamento della ghiandaia della Florida (Aphelocoma coerulescens), chiama in causa una ridotta disponibilità di habitat utilizzabili per la riproduzione. La ghiandaia della Florida si riproduce esclusivamente nella boscaglia e la popolazione di questi uccelli occupa spesso interamente l'habitat disponibile. In tali casi, i giovani non hanno alcuna opportunità di allontanarsi dal territorio dei genitori, sistemarsi in un territorio proprio e lì riprodursi. Devono invece rimanere a casa e attendere pazientemente che si presenti loro l'opportunità di impadronirsi di uno dei territori confinanti o di parte del territorio d'origine. Mentre aspettano che si liberi un posto possono incrementare la propria fitness aiutando i propri genitori. Nutrendo i propri fratelli o sorvegliandoli dai predatori, gli aiutanti permettono ai propri genitori di allevare un maggior numero di nidiate oppure nidiate comprendenti più figli di quanto questi potrebbero fare senza il loro aiuto. Di conseguenza, gli aiutanti ottengono un guadagno in fitness indiretta, ovvero la fitness ottenuta non attraverso la propria riproduzione ma attraverso l'incremento del successo riproduttivo dei consanguinei (Hamilton, 1964). Tuttavia, non appena tali aiutanti hanno la possibilità di scegliere se riprodursi 'personalmente' o continuare ad aiutare i genitori, essi abbandonano questi ultimi e creano una famiglia propria. Ciò è risultato particolarmente evidente in un'occasione particolare in cui una malattia aveva drasticamente ridotto la popolazione. Dopo tale crollo demografico, non fu possibile individuare neanche un aiutante e tutti gli individui sopravvissuti, che in precedenza erano stati aiutanti, si stabilirono nei territori rimasti liberi e si dedicarono alla propria riproduzione. La causa principale del comportamento di helping delle giovani ghiandaie della Florida è dunque ecologica e risiede nella disponibilità limitata di territori utilizzabili per la riproduzione.
Il comportamento di helping può anche derivare da una scarsa disponibilità di partner sessuali. Questo avviene molto frequentemente nel martin pescatore, le cui femmine covano la prole più a lungo dei maschi, essendo inoltre le sole a provvedere all'incubazione notturna. Sono dunque molto più esposte alla predazione rispetto ai maschi e, di conseguenza, diventano spesso più rare di questi. In tale situazione i figli maschi che non riescono a trovare una partner possono aiutare i genitori quando questi ultimi si riproducono di nuovo, ottenendo così, anche in questo caso, un incremento della propria fitness indiretta. Ancora più sorprendentemente, in questa specie, a dare una mano nella crescita dei piccoli possono essere anche maschi che non sono in alcun modo imparentati con la coppia genitoriale e che vengono chiamati aiutanti secondari. Poiché evidentemente essi non ne traggono alcun beneficio dal punto di vista genetico, un simile comportamento è particolarmente difficile da spiegare. Tuttavia, è stato scoperto che questi maschi tentano di ottenere l'accesso alle femmine che stanno aiutando, per potersi in seguito accoppiare con esse. Sembra anche che il maschio appartenente alla coppia genitoriale intuisca che il proprio status viene in tal modo minacciato, poiché (a meno che la nidiata non sia troppo numerosa per poter essere cresciuta senza un aiuto addizionaie) esso respinge in maniera aggressiva gli aiutanti con i quali non è imparentato. D'altra parte, gli aiutanti secondari contribuiscono in misura molto minore, rispetto a quelli primari, alla nutrizione dei piccoli. Evidentemente esiste un meccanismo per il quale essi mettono in atto solamente un minimo indispensabile di cure parentali. Gli aiutanti secondari lavorano solamente quel tanto che basta per essere tollerati dal partner della femmina e molti preferiscono nutrire direttamente questa (una tipica forma di corteggiamento) anziché i suoi piccoli.
Dunque, anche con il comportamento di helping, gli animali ottengono vantaggi in termini di fitness individuale, facilitando la riproduzione dei propri parenti stretti (kin selection o selezione di parentela) o incrementando le possibilità di riprodursi 'personalmente' in seguito.
Il parassitismo di covata
Poiché le cure parentali rappresentano una risorsa assai preziosa, non stupisce che alcuni animali sfruttino per la propria prole gli sforzi prodigati da altri individui. Negli uccelli il successo dei parassiti di covata dipende dall'abilità nel fare accettare le proprie uova all'animale ospite. Si tratta di un fenomeno molto frequente all'interno di una stessa specie. Per esempio, circa il 15% dei nidi di storno contiene uova che non sono state deposte dalla femmina che le cova e che in seguito nutrirà i nidiacei, ma da altre femmine di storno che sono riuscite, di nascosto, a mettere le proprie uova in quel nido. In tal modo, queste ultime possono sfruttare, per incrementare la propria fitness, le cure parentali di un'altra coppia di genitori.
Esistono diversi uccelli, tra i quali i cuculi, i molotri, gli indicatori e alcuni tessitori, che praticano il parassitismo di covata interspecifico. Queste specie, come per esempio il cuculo europeo (Cuculus canorus), sfruttano le cure parentali prodigate da individui appartenenti ad altre specie. A tal fine, devono per prima cosa risolvere il problema di come far accettare alla specie ospite le proprie uova, che hanno un aspetto differente dalle sue. Una soluzione consiste nel produrre uova che siano mimetiche rispetto a quelle della specie parassitata, altrimenti il parassitismo può funzionare esclusivamente nei casi in cui la specie ospite non abbia evoluto la capacità di distinguere le proprie uova e quindi di rifiutare quelle del parassita. Una volta superato tale ostacolo, il piccolo cuculo appena uscito dall 'uovo spinge fuori dal nido i propri fratellastri e si appresta a sfruttare appieno la risposta dei nuovi genitori adottivi alle proprie vivaci richieste di cibo. Esso inoltra infatti le proprie richieste con tale intensità che sembra simulare la presenza di un'intera nidiata. In tal modo, induce nei genitori che ha ingannato l'intera gamma dei comportamenti di cura, di riscaldamento, di nutrizione e di difesa dai predatori. In altre specie il parassitismo ha sugli ospiti effetti negativi più moderati. Il parassita infatti non distrugge la covata dell'ospite, ma approfitta delle cure parentali a essa prodigate. Tale diversa modalità implica tuttavia un altro problema: i genitori possono confrontare i piccoli della specie parassita con i propri; in questa situazione i primi devono competere con i secondi per accaparrarsi le cure parentali, come accade per esempio nelle vedove africane (o ploceidi) che parassitano gli astrildidi. l nidiacei di quest'ultimo gruppo sono facilmente riconoscibili perché l'interno della loro bocca presenta delle papille di forma e colore assai caratteristici e variabili da specie a specie, e i genitori nutrono esclusivamente i piccoli che esibiscono tali segnali specie-specifici. Diversamente da quelli del cuculo, i nidiacei delle vedove non spingono fuori dal nido i piccoli della specie parassitata. Copiano invece le loro linee e macchie colorate così esattamente da essere accettati dagli adulti come figli propri. La selezione a favore della similarità (o, meglio, dello spiccato mimetismo) ha prodotto nei nidiacei della specie parassita copie incredibilmente dettagliate dei disegni presenti sul palato e sul becco dei piccoli della specie ospite. Per esempio, il nidiaceo della vedova paradisea (Steganura paradisea) è talmente simile a quello della melba (Pytilia melba) che persino gli omitologi hanno impiegato un bel po' di tempo per rendersi conto che non tutti i piccoli presenti in un nido di melba appartenevano davvero a tale specie. Nel corso dell'evoluzione, la specie parassita ha copiato fedelmente i segnali della specie ospite, al fine di evitare che i genitori parassitati possano discriminare i diversi piccoli durante le fasi di nutrizione. Ciò rimanda alla fondamentale importanza dei meccanismi di riconoscimento tra genitori e prole. Di fatto, nelle vedove, lo spiccato mimetismo nei confronti delle specie ospiti ha condotto all'evoluzione e alla speciazione parallela di parassiti e parassitati, così che ciascuna specie ospite ha una specifica specie di vedova come parassita (Nicolai, 1974). L'eusocialità Gli insetti eusociali, tra i quali sono particolarmente note le formiche, le vespe (v. il saggio di S. Turillazzi e M.J.W. Eberhard, in questo volume) e le api, rappresentano il culmine dell'altruismo (Bourke, 1997). In essi si sono evoIute vere e proprie caste di femmine sterili, che aiutano la madre nell'allevamento dei fratelli. L'evoluzione ha dunque portato, in queste specie, alla totale perdita delle potenzialità riproduttive da parte di alcuni individui, poiché il vantaggio che essi ottengono aiutando la madre è così grande che possono permettersi di rinunciare a mettere al mondo una prole propria. Si tratta sostanzialmente, in questo caso, di cure parentali che non vengono mai messe in atto dai veri genitori! Quello dell'evoluzione delle cure parentali in queste società è un campo di studio talmente vasto che non possiamo fare altro che rimandare il lettore interessato, per esempio, al meraviglioso libro di E.O. Wilson (1971) su questo argomento.
Bibliografia citata
BATESON, P. (1994) The dynamics of parent-offspring relationships in mammals. Trends in Ecol. Evol., 9, 399-403.
BAYLIS, J.R. (1981) The evolution of parental care in fishes with reference to Darwin's ruIe of male sexual selection. Environ. Biol. Fishes, 6, 223-251.
BOURKE, A.F.G. (1997) Sociality and kin selection in insects. In Behavioural ecology: an evolutionary approach. 4a ed., a c. di Krebs lR., Davies N.B., Oxford, Blackwell, pp. 203-227.
BROWN, J.L. (1987) Helping and communal breeding in birds: ecology and evolution. Princeton, Princeton University Press.
CLUTTON-BROCK, T.H. (1991) The evolution of parental care. Princeton, Princeton University Press.
CRUMP, M.L. (1996) Parental care among the Amphibia. In Parental care: evolution, mechanisms and adaptive significance, a c. di Rosenblatt J.S., Snowdon C.T., San Diego, Academic Press.
DAVIES, N.B. (1992) Dunnock behaviour and sodal evolution. Oxford-New York, Oxford University Press.
DIESEL, R., BAURLE G., VOGEL, P. (1995) Cave breeding and froglet transport: a novel pattern of anuran brood care in the Jamaican frog, Eleutherodactylus cundalli. Copeia, 354-360.
DIESEL, R., SCHUH, M. (1993) Maternal care in the bromeliad crab Metopaulias depressus (Decapoda): maintaining oxygen, pH and calcium levels optimal for the larvae. Behav. Ecol. Sociobiol., 32, 11-15.
HAMILTON, D.W. (1964) The genetical evolution of social behaviour I and II. J. Theoretical Biol., 7, 1-52.
HORNING, M., TRILLMICH, F. (1997) Ontogeny of diving behaviour in the Galápagos dur seal. Behaviour, 134, 1211-1257.
KILNER, R., JOHNSTONE, R.A. (1997) Begging the question: are offspring solicitation behaviours signals of need? Trends Ecol. Evol., 12, 11-15.
LÉVY, F., KENDRICK, K.M., KEVERNE, E.B., PORTER, R.H., ROMEYER, A. (1996) Physiological, sensory and experential factors of parental care in sheep. In Parental care: evolution, mechanism and adaptive significance. Advances in the study of behavior, a c. di Rosenblatt J.S., Snowdon C.T., vol. 25, pp. 385-422, San Diego, Academic Press.
MOCK, D.W., PARKER, G.A. (1997) The evolution of sibling rivalry. Oxford-New York, Oxford University Press.
NICOLAY, J. (1974) Mimicry in parasitic birds. Sci. Am. 231, 92-98.
SHELDON, B.C., VERHULST, S. (1996) Ecological immunology: costly parasite defences and trade-offs in evolutionary ecology. Trends Ecol. and Evol., 11, 317-321.
TRILLMICH, F. (1981) Mutual mother-pup recognition in Galápagos fur seal and sea lions: cues used and functional significance. Behaviour, 78, 21-42.
TRIVERS, R.L. (1972) Parental investment and sexual selection. In Sexual selection and the descent of man 1871-1971, a c. di Campbell B., Chicago, Aldine Publ. Co., pp. 136-179.
TRUMBO, S.T. (1996) Parental care in invertebrates. In Parental care: evolution, mechanism and adaptive significance. Advances in the study of behavior, a c. di Rosenblatt J.S., Snowdon C.T., vol. 25, pp. 385-422, San Diego, Academic Press.
TYLER, M.J., a c. di (1983) The gastric brooding frog. Kent, Croom Helm Ltd.
WILSON E.O. (1971) The insect societes. Cambridge, Mass., Belknap Press of Harward University Press.
Bibliografia generale
ROSENBLATT J.S., SNOWDON, C.T, a c. di, Parental care: evolution, mechanism and adaptive significance (Advances in the study of behavior, vol. 25). San Diego, Academic Press, 1996.