
Le origini dell'uomo e l'evoluzione dei primi ominidi
Le origini dell'uomo e l'evoluzione dei primi ominidi
Sebbene le forme mioceniche ancestrali siano poco note rispetto agli australopitecini, è stata attualmente documentata la comparsa dei primi ominidi, risalente ad almeno 4,5 milioni di anni fa, con i fossili di Ardipithecus ramidus; inoltre la loro diffusione geografica si è rivelata maggiore di quanto supposto in precedenza. Man mano che tra gli ominidi primitivi viene riconosciuta una diversità specifica più ampia, sia negli australopitecini sia nel genere Homo, il quadro delle relazioni evolutive appare sempre più complesso. Inoltre, sia la concentrazione delle ricerche sul rinvenimento e l'analisi degli australopitecini sia la scarsità generale di esemplari fossili appartenenti al genere Homo ostacolano la comprensione delle origini e della diversificazione del genere a cui apparteniamo. Per una chiarificazione dell'evoluzione dei primi ominidi è necessario il rinvenimento di ulteriori esemplari fossili ma forse, e soprattutto, è necessaria l'applicazione di più sofisticati metodi di analisi filogenetica e sistematica.
Il concetto di specie nella famiglia
Hominidae La famiglia dell'uomo, designata in zoologia con il nome Hominidae, comprende tutte le specie bipedi di Primati, viventi o estinte, che si sono differenziate a partire dalla loro separazione da un antenato comune con le scimmie antropoidi africane, in una data compresa tra 6 e 10 milioni di anni fa. La famiglia comprende due generi ben conosciuti, Homo e Australopithecus, e forse un terzo, Ardipithecus. Mentre la maggior parte dei ricercatori concorda nel riconoscere due o tre distinti generi di ominidi, non si può dire lo stesso per la collocazione tassonomica delle specie di ominidi estinte. Poiché il concetto biologico definisce la specie come un gruppo di organismi riproduttivamente isolato rispetto ad altri gruppi simili, come possiamo identificare le specie fossili, dal momento che non potremo mai conoscerne i comportamenti riproduttivi? Come principio generale, ci si riferisce a specie attuali di Primati, di cui è nota l'appartenenza a gruppi riproduttivi isolati, per stabilire l'intervallo di variabilità di specifici caratteri morfologici dello scheletro. Questo criterio è poi applicato a insiemi di fossili: se l'intervallo di variabilità dei caratteri morfologici di un campione fossile è uguale o minore a quello di una specie vivente, è meglio collocare i fossili in un'unica specie; se invece si riscontra nell'insieme dei fossili un'ampia variabilità che permette una suddivisione del campione in tipi morfologici differenti, allora tali fossili appartengono probabilmente a più specie.
Valutare il significato delle differenze morfologiche non è sempre semplice e in paleoantropologia rimane intensa la controversia riguardo al numero di specie di ominidi esistite. Per decidere in modo documentato circa il numero di specie nei resti fossili è essenziale possedere una vasta conoscenza della variazione interna a ogni specie vivente e di quella delle specie tra loro. Le specie costituiscono gli elementi fondamentali della classificazione, sono entità reali e distinte in natura; è quindi di estrema importanza un'accurata ricognizione delle paleospecie nella ricostruzione storica dell'evoluzione. La paleoantropologia è stata per molti versi influenzata da due scuole di denominazione delle specie: gli splitters (gli 'spaccacapelli') e i lumpers (gli 'ammucchiatori'). Gli splitters danno un eccessivo rilievo a tratti morfologici minori, creando perciò troppe specie. llumpers, d'altro canto, tendono a trascurare le differenze meno importanti; di conseguenza concepiscono un numero troppo basso di specie fossili. Se la verità, molto probabilmente, sta nel mezzo o quasi, recenti considerazioni suggeriscono, per una serie di ragioni, che il numero delle specie di ominidi sarà sempre sotto stimato (Tattersall, 1992).
lnnanzitutto i campioni fossili sono frammentari, quindi si può semplicemente non disporre di prove sufficienti per definire una nuova specie. In secondo luogo, gli osservatori possono differire sensibilmente nell' interpretazione del significato dei tratti anatomici per definire le specie. In terzo luogo, e soprattutto, studi di vasta portata sui gruppi superstiti di Primati, quali i lemuri, suggeriscono che la correlazione tra specie e morfologia scheletrica è limitata. Specie di lemuri facilmente distinte in base al comportamento, al colore della pelle e del pelo e così via, presentano differenze scheletriche di scarsa entità; perciò, se calcolato unicamente sulla base delle ossa fossili (il comportamento e le parti molli non fossilizzano), il numero delle specie di lemuri risulterebbe sempre sotto stimato e ciò costituisce un problema serio per chi si occupa di paleospecie.
L'evoluzione degli ominidi non si differenzia da quella di altri organismi. La divergenza e la ramificazione sembrerebbero la regola e quanti più ominidi si troveranno, tanto più l'albero diverrà intricato. Da un punto di vista teorico, poiché il mutamento morfologico e la speciazione non vanno necessariamente di pari passo, è più che probabile che alcune specie restino non identificate. Anche se non si saprà mai esattamente quante specie di ominidi siano esistite, l'avanzamento della conoscenza dei principi evolutivi e della variazione anatomica permette ora di discutere e interpretare i taxa dei fossili di ominidi entro un quadro razionale (Kimbel e Martin, 1993).
Gli australopitecini e gli inizi del Miocene
Il nome del genere Australopithecus, inizialmente coniato da R.A. Dart, un anatomista dell'università Witwatersrand di Johannesburg, nel 1925, combina il vocabolo latino australis, che significa 'meridionale', con quello greco per 'scimmia', pithekos. Sebbene Australopithecus non sia una scimmia, né sia confrnato all'Africa australe, il Codice internazionale di nomenclatura zoologica, che riconosce questa denominazione seppur imprecisa come una designazione generica valida, garantisce la continuità d'uso di questo nome (Meikle e Parker, 1994).
Tutte le specie di Australopithecus sono qui collettivamente designate come australopitecini. Il termine formale Australopithecinae, una sottofamiglia distinta nella gerarchia linneana, fu proposto nel 1939 da W.K. Gregory e H. Hellmann per distinguere tale gruppo dalle specie più tarde di Homo alle quali era stato a volte attribuito il rango di sotto famiglia degli Homininae. In considerazione del fatto che queste distinzioni a livello di sotto famiglia non sono universalmente accolte, per comodità e semplicità d'esposizione qui si userà, per riferirsi alle specie di Australopithecus, il termine informale di australopitecini.
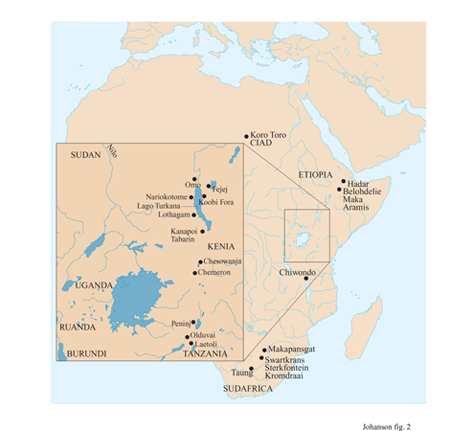
l resti dei più antichi ominidi, oggetto di questo saggio, sono stati trovati in Africa e si situano lungo un intervallo geologico che va approssimativamente da 4 a l milione di anni fa (qui indicato come periodo plio-pleistocenico). Attualmente si riconoscono sette specie distinte di Australopithecus: anamensis, afarensis, africanus, aethiopicus, boisei, robustus e bahrelghazali. Gli australopitecini avevano un cervello relativamente piccolo, presentavano dimorfismo sessuale e denti grossi dallo smalto spesso, presumibilmente non fabbricavano utensili in pietra ed erano bipedi. A eccezione di un esemplare trovato a Koro Toro (Ciad), A. bahrelghazali (Brunet et al., 1996), tutti gli australopitecini sono stati scoperti in due principali regioni geografiche: l'Africa orientale, prevalentemente in siti associati alla Rift Valley, e l'Africa meridionale, in caverne dolomitiche (fig. 2).
Per quanto un numero crescente di ritrovamenti di australopitecini fornisca conoscenze sempre più approfondite sull'adattamento, lo stile di vita, la diversità tassonomica e l'anatomia di questi primi ominidi, si è ancora ben lungi dal comprenderne le esatte origini nel Miocene. La radiazione evolutiva delle scimmie del Miocene (avvenuta da 5 a 20 milioni di anni fa) costituisce senza dubbio la sorgente da cui scaturiscono gli ominidi e le scimmie antropoidi attuali. Tuttavia, le lacune della documentazione paleontologica, specie nel tardo Miocene (da 5 a Il milioni di anni fa), ci impediscono di individuare precise relazioni evolutive tra le scimmie antropoidi del Miocene da un lato e gli ominidi e le scimmie antropoidi africane dall' altro (Pilbeam, 1996).
La ricerca di un antenato comune agli ominidi e alle scimmie africane viventi resta una delle maggiori sfide per la ricerca paleoantropologica; ogni indizio tende tuttavia a suggerire che tale creatura fosse una scimmia indifferenziata, forse simile allo scimpanzé (Pilbeam, 1996). l più antichi ominidi avrebbero quindi un aspetto più somigliante alle scimmie antropomorfe rispetto agli ominidi successivi. Ovviamente, quando i resti fossili diverranno ancor più completi, si potrà arrivare a un punto della loro storia evolutiva, immediatamente successivo alla divergenza tra le scimmie e gli ominidi, in cui risulterà difficile separarli; per il momento, ciò non è ancora possibile.
Negli anni Settanta e Ottanta era ampiamente accettato che Ramapithecus del Miocene indiano e pakistano e la sua controparte africana, Kenyapithecus, rappresentassero gli ominidi più antichi. Caratteri quali lo spesso smalto dei denti, le dimensioni ridotte dei canini, una presunta arcata dentaria parabolica (in seguito dimostratasi frutto di una erronea ricostruzione), la faccia corta e via dicendo, erano paragonati ai tratti di Australopithecus. Risalente a circa 12÷14 milioni di anni fa, Ramapithecus/Kenyapithecus fu ben presto accolto come un protoominide, coerentemente con la visione, da molti condivisa, secondo la quale scimmie e ominidi si sarebbero separati 15÷20 milioni di anni fa.
L'acceso dibattito sulla posizione evolutiva di questo taxon giunse a una conclusione con la scoperta, avvenuta nel 1981 in Pakistan, di una faccia quasi completa di Ramapithecus, ora ampiamente riconosciuto (Pilbeam, 1982) come appartenente al genere Sivapithecus. Dal momento che questo esemplare presenta un insieme di caratteri craniali comuni agli oranghi, Ramapithecus (Sivapithecus) è ora considerato un progenitore di queste scimmie antropoidi odierne.
Gli sviluppi dell'antropologia molecolare, tra cui le ricerche sulle proteine del sangue e, più specificamente, le comparazioni tra il DNA delle scimmie e quello dell'uomo, dimostrano un incredibile grado di similarità tra gli uomini d'oggi e le scimmie antropoidi africane, in particolare gli scimpanzé (Sarich, 1992). Una implicazione fondamentale di questi studi è che l'antenato comune sia molto più recente, forse tra 6 e lO milioni di anni fa, ma questa creatura ancora elude i cacciatori di fossili.
È interessante osservare in proposito che durante il Miocene, la cosiddetta Età delle Scimmie, queste Cercopithecoidae erano relativamente rare di numero e scarsamente diversificate. Grosso modo 10 milioni di anni fa le scimmie subirono una radiazione adattativa, forse in risposta alla generale tendenza di un abbassamento della temperatura e un'aridificazione, che produsse ambienti come boschi e savane meno attraenti per loro, adattate meglio a climi più caldi e umidi come le folte foreste tropicali. Le scimmie cominciarono dunque a proliferare, forse grazie alloro elevato livello di flessibilità comportamentale, alla maggiore tolleranza ambientale e a certe preferenze alimentari. La capacità di mangiare e digerire foglie e frutti acerbi può aver dato loro un vantaggio sulle scimmie antropoidi, che basavano la loro dieta su frutti più maturi. Inoltre, le scimmie si riproducono assai più rapidamente e ciò può aver determinato il loro vantaggio demografico rispetto alle scimmie antropoidi, che si riproducono più lentamente. È da notare che questa tendenza al declino delle scimmie antropoidi persiste ancora oggi quando le specie esistenti si vengono a trovare in aree sempre più ristrette. La sola scimmia che continua a prosperare è la 'Scimmia Nuda', o 'Scimmia Bipede', ovvero - come siamo stati talvolta chiamati - la 'Terza Scimmia'.
Gli australopitecini dell'Africa meridionale
L'annuncio di Dart del ritrovamento di Australopithecus africanus ('la scimmia meridionale d'Africa'), nel 1925, fornì una sorprendente conferma a quanto predetto da Charles Darwin nel 1871, e cioè che i nostri antenati più lontani si sarebbero probabilmente trovati in Africa, poiché in quel continente abitano i nostri parenti più prossimi, gli scimpanzé e i gorilla.
Un esemplare immaturo, forse al di sotto di cinque anni d'età alla morte, il fossile di Taung, dal cervello relativamente piccolo (405 cm) suscitò una particolare attenzione per la posizione del foro occipitale. Nei quadrupedi questo foro è collocato verso la parte posteriore del cranio, ma nei bipedi, come nell'esemplare di Taung, è collocato sotto la scatola cranica. Il 'Bambino di Taung', bipede, era perciò un ominide. In Inghilterra il singolo reperto di Dart non convinse gli scienziati, affascinati da una visione eurocentrica delle origini umane sorretta dal cranio di Piltdown, più tardi rivelatosi uno dei più fortunati falsi scientifici. Alcuni studiosi europei dubitarono della legittimità di usare un esemplare immaturo per istituire una nuova specie e si spinsero fino al punto di ipotizzare che se l'individuo di Taung avesse raggiunto la maturità, esso si sarebbe evoluto in una scimmia.
La scoperta dei ricchi depositi fossiliferi nelle caverne dei siti di Makapansgat (a partire dal 1948) e Sterkfontein (a partire dal 1936), nella regione sudafricana del Transvaal, fornì numerosi esemplari adulti di A. africanus, confermando la concezione di Dart, secondo il quale queste creature erano veri ominidi e non scimmie antropomorfe fossili. Un'ulteriore comprensione della differenziazione degli australopitecini si ebbe con la scoperta di esemplari fossili di ominidi nelle caverne di due altri siti, Kromdraai (dal 1938) e Swartkrans (dal 1948), sempre nella regione del Transvaal. Gli ominidi di questi ultimi due siti erano di costituzione più robusta, con creste variamente sviluppate sul cranio e poderose mascelle con molari e premolari massivi. L'appellativo iniziale di Paranthropus robustus, conferito a questi ominidi da R. Broom, è ancora utilizzato da alcuni ricercatori, anche se la maggior parte di essi vi si riferisce con il nome di Australopithecus robustus.
l nostri antenati australopitecini non occupavano in realtà le caverne dei siti del Transvaal. Le loro ossa, come quelle di altri animali, caddero nelle caverne sotterranee attraverso qualche orifizio e vi si accumularono naturalmente. La formazione di brecce ossifere nelle caverne si protrasse per un lungo periodo di tempo, attraverso un insieme molto complesso di processi geologici che rendono difficile l'interpretazione della stratigrafia delle caverne. L'attribuzione cronostratigrafica degli ominidi si rivelò difficoltosa a causa della carenza di sedimenti adatti, come le ceneri vulcaniche, al fine di ottenere datazioni assolute dei depositi delle caverne.
Una cronologia locale per i depositi che contengono australopitecini è stata ottenuta usando una forma di datazione relativa. Essa si basa sull'assunto che i siti più antichi presentano una percentuale maggiore di animali estinti e una minor frequenza di forme viventi, mentre una relazione inversa caratterizza i siti più recenti. Applicando questo principio, i siti - dal più antico al più giovane - risultano così ordinati: Makapansgat, Sterkfontein, Taung, Kromdraai e Swartkrans. Una conclusione che può essere tratta da questa sequenza è che A. africanus è più antico di A. robustus. Comparando le specie animali trovate nei siti del Transvaal con quelle dei siti dell' Africa orientale, datati con prove radiometriche, si possono ottenere età geologiche approssimative per i depositi di australopitecini. Le stime d'età biostratigrafiche, che generalmente confermano la sequenza relativa presentata sopra, sono: Makapansgat, 2,8 milioni di anni; Sterkfontein, 2,5 milioni di anni; Taung, 2,3 milioni di anni; Kromdraai e Swartkrans, da 1,8 a 1 milione di anni fa (Delson, 1988).
Nonostante le peculiarità del deposito e la mancanza di una datazione geologica precisa, i siti del Sudafrica hanno fornito un vasto insieme di fossili di australopitecini, rappresentativi di quasi tutte le parti dello scheletro. Collezioni complessive di circa 300 esemplari a Swartkrans e di quasi 400 a Sterkfontein collocano gli australopitecini sudafricani tra le specie di ominidi fossili meglio rappresentate.
Sebbene erroneamente, le due specie sudafricane sono talvolta indicate come 'gracile' (A. africanus) e 'robusta' (A. robustus). Come taglia fisica esse non erano probabilmente molto differenti (in media, entrambe pesavano 36 kg e misuravano 1,2 m in altezza), così che i termini 'gracile' e 'robusto' non si riferiscono in realtà che allo sviluppo delle mascelle e alle dimensioni dei denti (McHenry, 1992). Sotto molti aspetti questi due taxa rappresentano delle variazioni sul tema; tuttavia numerosi tratti anatomici, specie nel cranio, li rendono facilmente distinguibili l'uno dall'altro. In generale, il cranio di A. africanus manca di creste ectocraniche pronunciate, mentre in A. robustus spesso sono presenti strutture che, in alcuni casi, si sviluppano a formare creste cervi cali e sagittali, segno di un grosso sviluppo della muscolatura masticatoria e nucale. In visione laterale, i crani 'robusti' hanno una fronte molto bassa e una faccia piatta. Gli zigomi, prominenti, nascondono l'area nasale accentuando la topografia 'a scodella' dello scheletro facciale. Al contrario, le forme 'gracili' presentano una fronte leggermente 'a duomo'; l'osso alveolare sporgente conferisce alla faccia un profilo più prognato. l 'robusti' si distinguono, inoltre, per le mascelle dalla costituzione più pesante e in particolare per le mandibole marcatamente voluminose. Sebbene vi sia qualche sovrapposizione tra le due specie riguardo alle dimensioni dei denti, gli australopitecini 'robusti' hanno molari e premolari molto grossi. Essi sono ancora caratterizzati da incisivi e canini relativamente piccoli a confronto con i grossi denti posteriori, mentre queste due serie di denti sono più armoniosamente proporzionate negli australopitecini 'gracili'.
Se, in base all'usura dei denti, entrambe le specie del Sudafrica erano verosimilmente vegetariane, la costituzione più pesante del cranio nelle forme 'robuste' suggerisce che queste avessero una considerevole potenza masticatoria. Forse A. robustus fu un vegetariano più tardo e più specializzato, che consumava grandi quantitativi di cibo coriaceo e di bassa qualità. Le superfici occlusive dei denti posteriori nei 'robusti' mostrano un'alta frequenza di infossature e scalfitture profonde, il che implica che la loro dieta conteneva una notevole quantità di graniglia (Grine, 1988). Tranne alcuni arte fatti in osso levigato, rinvenuti a Swartkrans e probabilmente usati per scavare tuberi e radici, vi sono poche prove dell 'uso di utensili da parte degli australopitecini. Dart suggerì che certe ossa lunghe, mandibole di antilopi e coma venissero usati come attrezzi e armi dagli ominidi di Makapansgat. Si ritiene ora che la presunta 'cultura osteodontocheratica' (dell' osso, del dente, del como) altro non sia che un accumulo naturale, prodotto da iene, istrici e leopardi (Brain, 1981).
Le testimonianze attuali suggeriscono che A. africanus e A. robustus furono confrnati al Sudafrica; l'estensione della loro migrazione verso nord è sconosciuta. Tuttavia, è significativo che nessuna di queste specie sia stata riportata alla luce nei siti ricchi di ominidi dell'Africa orientale.
La scienza paleoantropologica
La scoperta degli ominidi del periodo plio-pleistocenico in Sudafrica fu determinante fino al 1959, quando una singola scoperta nella gola di Olduvai, in Tanzania settentrionale, bastò a indirizzare l'attenzione degli antropologi verso la Rift Valley africana. Dopo decenni di laboriose ricerche effettuate a Olduvai, il17 luglio 1959 M. Leakeytrovò sul fianco di una collina un cranio parzialmente esposto dall' erosione superficiale. Inizialmente chiamato Zinjanthropus boisei (Zinj è un antico nome arabo per indicare l'Africa orientale e C. Boisei era un benefattore), l'australopitecino 'iperrobusto', dalla pronunciata cresta sagittale, dalla grande faccia concava, dagli enormi molari atti a frantumare e triturare, inaugurò una nuova epoca degli studi sull'evoluzione umana (Tobias, 1968).
Lo 'Zinj' stimolò lo studio approfondito di una singola località (la gola di Olduvai); ciò portò alla realizzazione di un nuovo approccio che integrava le competenze di discipline diverse in uno studio strategico, specificamente rivolto a ottenere una conoscenza per quanto possibile estesa delle origini dell'uomo. Questo impegno multidisciplinare è formalmente chiamato 'paleoantropologia'. Incorporando un insieme di discipline diverse, quest'ultima si spinge oltre le normali esplorazioni e scoperte per adottare un approccio attentamente articolato e mirato, al fine di rispondere alle domande sul passato evolutivo dell'uomo.
La maggior parte dei fossili trovati in Africa orientale è strettamente legata alla Rift Valley, che presenta straordinarie associazioni tra i fossili e gli strati sedimentari in cui questi si depositarono, fossilizzarono e infine vennero scavati, o portati alla luce dall'erosione. Nella Rift Valley il ricco contesto geologico degli ominidi fossili e della fauna che li accompagna consente datazioni geologiche precise, che a loro volta permettono una calibratura degli eventi evolutivi del passato. Inoltre, la conoscenza del contesto geologico aumenta notevolmente la comprensione dei paleoambienti in cui i nostri antenati vissero, morirono e si fossilizzarono. Il contesto geologico dei siti dell'Africa orientale contrasta fortemente con quello dei siti di australopitecini sudafricani, dove è meno facile ricavare informazioni sulla cronologia, le associazioni e i paleoambienti. Sin dagli anni Cinquanta la 'Nuova Paleoantropologia' (Johanson, 1996) è divenuta più sofisticata sia sul campo, sia in laboratorio. Si distinguono per la cruciale importanza del loro contributo innovativo la Orno Research Expedition nell 'Etiopia meridionale, ancora in corso, le investigazioni attorno al Lago Turkana (Coppens et al., 1976), lo Hadar Research Project (Johanson, 1996) e il Middle Awash Group (White et al., 1994). Questi progetti principali, iniziati verso la fine degli anni Sessanta, posero le fondamenta dell'approccio metodologico e teorico che oggi sono alla base della paleoantropologia.
Specialisti di scienze biologiche e della terra, sotto la direzione di un paleoantropologo, lavorano in modo integrato per ricavare dal passato il maggior quantitativo possibile di informazioni. l palinologi, estraendo granuli di polline fossile dagli strati antichi, forniscono conoscenze sulle associazioni vegetali del passato. l paleontologi, studiando gruppi di mammiferi che vanno dai roditori agli elefanti, contribuiscono con preziosi dettagli alla conoscenza della fauna che viveva accanto ai primi ominidi e perciò arricchiscono la visione delle antiche comunità animali. Lo studio dei resti materiali da parte degli archeologi aiuta a concretizzare i modi di vita e gli adattamenti culturali dei primi ominidi. Tecnologie avanzate come il telerilevamento sono state di grande aiuto nel rinvenire nuovi depositi fossiliferi; i GPS (Global Positioning Systems, sistemi di posizionamento globale) permettono una localizzazione precisa dei siti fossiliferi.
l geologi non solo hanno sviluppato sofisticati metodi di datazione per stabilire un preciso quadro di riferimento cronologico dell'evoluzione umana, ma contribuiscono, con approfonditi studi sul campo, alla comprensione del contesto in cui sono stati trovati fossili nei siti di interesse paleoantropologico. Infatti, fossili e manufatti sono di scarsa utilità se non vengono accuratamente riferiti a un preciso contesto geologico.
In laboratorio, l'anatomia comparata classica resta essenziale per analizzare e valutare la posizione tassonomica degli esemplari fossili e il posto che essi verosimilmente occupano nella filogenesi degli ominidi. Le innovazioni teoriche della biologia evolutiva hanno enormemente ampliato il tradizionale quadro filo genetico entro il quale le specie di ominidi erano comprese. Il perfezionamento di sofisticate tecnologie d'indagine permette persino di esaminare la struttura interna di ossa e denti. Ciò è stato spesso di grande aiuto nell' analisi dell' anatomia funzionale e nello sviluppo di modelli biomeccanici che forniscono informazioni sugli adattamenti locomotori e sui vincoli imposti alla struttura ossea. L'uso degli elaboratori per ricostruire e analizzare la forma dei fossili è una delle tecnologie più recenti, che accresce la nostra capacità di vedere oltre i meri frammenti fossili.
Gli australopitecini dell'Africa orientale
I primi australopitecini
l depositi fossiliferi dell'Africa orientale hanno restituito le specie più antiche e più primitive di Australopithecus; datate ora a oltre 4 milioni di anni fa, esse sono considerevolmente più antiche di quelle sudafricane. Risulta dunque, sulla base delle attuali testimonianze, che gli ominidi fecero la loro prima apparizione in Africa orientale e migrarono da qui verso sud. Sfortunatamente, a causa della pressoché completa assenza di siti plio-pleistocenici tra l'Africa orientale e quella meridionale, non vi sono testimonianze fossili che permettano di stabilire collegamenti tra le due aree. Nonostante questa sia l'estensione geografica a noi nota di Australopithecus (con l'eccezione di un solo esemplare proveniente dal Ciad), si può tranquillamente presumere che questo genere di primi ominidi fosse più ampiamente diffuso di quanto sinora documentato.
La regione del Lago Turkana (chiamato un tempo Lago Rodolfo), nel Kenya settentrionale, è senza alcun dubbio una delle aree africane più ricche di fossili. l primi fossili di vertebrati vi vennero raccolti negli anni Trenta in depositi, ora riferiti alla formazione di Shungura, situati a nord del lago, in Etiopia, poco a ovest del fiume Orno; i primi ominidi vennero tuttavia recuperati in quest' area solo alla fine degli anni Sessanta. l fossili di australopitecini ritrovati nella formazione di Shungura sono rappresentati principalmente da denti isolati oltre che da una delle più massicce mandibole di A. boisei, risalente a 2,1 milioni di anni fa. A metà degli anni Sessanta con il ritrovamento di due frammenti venne riconosciuto per la prima volta il potenziale della zona a sud-ovest del Lago Turkana per le scoperte di ominidi. Il reperto più antico, un frammento di mandibola provvisto di un solo molare, consunto, proviene da Lothagam ed è stato datato a oltre 5 milioni di anni fa. Sfortunatamente la porzione conservata del reperto non basta a determinare se si tratti di un ominide o di una scimmia antropomorfa. Da depositi più recenti, risalenti a circa 4 milioni di anni fa, venne recuperato a Kanapoi un omero distale, indubbiamente appartenente a un ominide.
L'identità tassonomica dell'omero di Kanapoi rimase incerta finché non si rinvennero nel sito altri nove reperti dentali, craniali e postcraniali di ominidi (Leakey et al., 1995). A seguito di un attento studio comparativo, questi reperti di 3,9÷4,1 milioni di anni d'età, nonché l'omero, furono assegnati a una nuova specie, A. anamensis. Il nome della specie deriva dal termine indigeno anam, che significa 'lago', e fu scelto in chiaro riferimento alla prossimità di Kanapoi sia con il Lago Turkana sia con il paleolago Lonyumun, che esisteva anticamente nell'area. L' olotipo della specie consiste di una piccola mandibola in cui le file di denti sono parallele, la parte anteriore della mascella è marcatamente sfuggente e il sostegno interno, il toro mandibolare, è fortemente sviluppato e allungato posteriormente. Per tutti questi aspetti, come anche per i dettagli della dentatura, i caratteri di A. anamensis ricordano quelli di alcune scimmie antropoidi del Miocene ed esso è pertanto l'ominide più primitivo finora descritto. Significativo è il fatto che una tibia incompleta indichi con certezza che si tratta di una forma bipede, fornendo così la più antica tra le testimonianze sinora note della postura eretta dell'uomo e della sua locomozione.
Recenti scoperte di ominidi ancora più antichi, ma per il momento enigmatici, datati a 4,4 milioni di anni di età, sono state fatte nel sito di Aramis, in Etiopia (White et al., 1994). Aramis si trova nell' area del medio corso del fiume Auasc, nella regione dell'Afar, appena 75 km a sud di Radar, dove venne scoperto lo scheletro di 'Lucy'. l resti dentali, craniali e postcraniali di diciassette individui mostrano una serie di tratti anatomici molto più primitivi e simili a quelli delle scimmie rispetto agli altri australopitecini noti: essi giustificano così l'istituzione di una nuova specie, chiamata Australopithecus ramidus. Il nome della specie deriva dal termine ramid, che nella lingua della locale tribù afar significa 'radice'. Dopo l'annuncio iniziale, il materiale di Aramis fu trasferito a un nuovo genere, Ardipithecus, da T.D. White, G. Suwa e B. Asfaw (1995). Ardi significa, in lingua afar, 'terreno' o 'suolo': si conferisce così una duplice enfasi al fatto che questo taxon rappresenti per gli ominidi una specie basale.
A. ramidus differisce dagli altri ominidi per i canini relativamente grandi, i molari e i premolari piccoli, una base cranica primitiva, il primo molare inferiore da latte dm1 (deciduous molar) molto simile a quello degli scimpanzé e uno smalto dentario sottile. Due elementi in particolare, il dm1 simile a quello delle scimmie e il sottile smalto dentario, possono rivelarsi significativi nella valutazione della posizione filogenetica di A. ramidus nell'albero degli ominidi. Meticolose comparazioni, metriche e morfologiche, del dm1 confermano la sua vicinanza ai caratteri riscontrati nei moderni scimpanzé. Se questo dente fosse stato trovato isolato, avrebbe potuto costituire la prova dell'antichità degli scimpanzé. Lo smalto dentario sottile è tipico delle scimmie antropomorfe africane, mentre uno smalto più spesso caratterizza tutti gli ominidi, specie A. boisei in cui è estremamente resistente. Tuttavia, uno smalto dentario spesso può costituire una risposta ad alimenti coriacei da masticare a lungo e non è immediato stabilire in quale misura tale carattere sia veramente significativo per definire le affinità filo genetiche (Pilbeam, 1996).
È cosa certa che i resti di un notevole scheletro completo, raccolti nel 1994 e nel 1995 ad Aramis (White et al., 1995), aiuteranno a definire la posizione di A. ramidus nella filogenesi umana. Lo scheletro conserva ancora le sottili ossa dei polsi e quelle delle dita, lunghe e arcuate. Finora gli scopritori sono stati cauti nell'affermare che A. ramidus sia una forma bipede, sebbene la base del cranio porti un foro occipitale situato in avanti, condizione che usualmente si considera in rapporto con la locomozione bipede. Studi approfonditi del bacino, degli arti inferiori e delle ossa del piede potrebbero forse rivelare un modo di locomozione diverso da quello di ogni altro primate, vivente o estinto. Le scoperte di Aramis hanno aperto un nuovo terreno e sono tra le prime a spalancare una finestra su depositi più antichi di 4 milioni di anni fa. È indubbio che A. ramidus arricchirà le nostre conoscenze su un periodo assai remoto dell' evoluzione umana. La zona del medio corso del fiume Auasc è un terreno davvero fertile per i cercatori di ominidi fossili che hanno già contemplato l'ipotesi di scrutare in strati geologici risalenti a circa 5 milioni di anni fa. Se in orizzonti così antichi dovessero essere trovati ominoidi fossili, sarebbe difficile decidere quali siano i nostri antenati e quali quelli delle scimmie antropomorfe africane.
La comparsa di Australopithecus afarensis
Resti di A. afarensis sono stati recuperati in numerosi siti: a Radar, Maka, Fejej, Belohdelie e Orno in Etiopia, nella zona a est del Lago Turkana in Kenya e a Laetoli in Tanzania. Per ironia della sorte, il primo reperto in assoluto di questa specie, nonché il primo esemplare adulto di Australopithecus, un frammento di mascella, raccolto da L. Leakey nel 1935, fu catalogato come una scimmia. Nel 1981 esso furiscoperto nelle collezioni del Museo di storia naturale di Londra e correttamente identificato come appartenente alla specie di ominidi fossili cui ora si attribuisce il nome di A. afarensis.
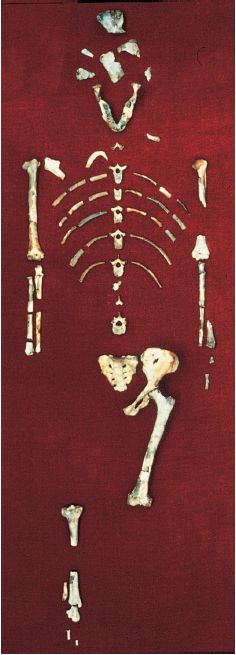
La più grande e completa raccolta di fossili di A. afarensis proviene da Radar, sito posto nella regione dell'Afar, in Etiopia. La collezione di ominidi fossili di Radar ora ammonta a quasi 325 reperti, incluso il notevole scheletro incompleto comunemente noto come 'Lucy' (fig. 9). I fossili di A. afarensis raccolti nella formazione di Radar, che ha uno spessore di circa 200 metri, rientrano in un intervallo temporale di 400.000 anni, con un'età compresa tra 3 milioni e 3,4 milioni di anni fa. Un attento campionamento geologico e l'applicazione di sofisticate tecnologie di datazione mediante argo, attraverso fusione laser di singoli cristalli, fanno di Radar una delle più calibrate sequenze di ominidi fossili (Walter, 1994; Walter e Aronson, 1993).
Oltre allo scheletro di 'Lucy' (AL. 288-1), che ha 3,2 milioni di anni, Radar si distingue per la scoperta, avvenuta nel 1975 nello stesso sito (AL. 333), di oltre 200 frammenti di ominidi, appartenenti a un singolo orizzonte geologico. A differenza di altri strati della formazione di Radar, contenenti un miscuglio di fossili di mammiferi che rispecchia il normale accumulo dei sedimenti, lo strato di AL. 333 contiene solo fossili di ominidi e ciò fa pensare a un evento deposizionale di tipo catastrofico. La catastrofe, la cui esatta natura resta ignota, potrebbe essere stata causata da un'alluvione improvvisa che uccise una comunità di ominidi, forse durante il sonno o il riposo, nel letto di un fiume in secca. Questo insieme unico, al quale talvolta si allude come alla 'Prima Famiglia', contiene almeno 13 individui, nove adulti e quattro fra adolescenti e bambini, e ci fornisce una visione istantanea senza precedenti delle variazioni scheletriche all'interno di una singola specie di ominidi fossili. Si pensa che la sostanziale differenza tra individui grandi e piccoli rifletta uno spiccato dimorfismo sessuale, con i maschi molto più grossi delle femmine. Le stime indicano che in media i maschi pesavano 45 kg e misuravano circa 1,5 m di altezza, mentre le femmine pesavano solo 29 kg ed erano alte l,l m (McRenry, 1992).

A metà degli anni Settanta, mentre erano in corso le ricerche a Radar, fu intrapresa una nuova campagna di scavi nel sito di Laetoli, in Tanzania (Leakey e Rarris, 1987), leggermente più antico di Radar sotto il profilo geologico. Circa 30 reperti, rappresentati per lo più da denti e mascelle, furono recuperati dai depositi di Laetoli, risalenti a 3,6 milioni di anni fa. Questo sito è però più noto per la straordinaria scoperta, nel 1978, di una pista di orme lunga 27 metri che degli ominidi impressero in una cenere vulcanica 3,6 milioni di anni fa. Tale pista (fig. 10), composta da 69 impronte, fu tracciata da due individui di A. afarensis mentre avanzavano nel paleopaesaggio. Fortunatamente, dopo che essi ebbero lasciato le loro impronte nella cenere vulcanica leggermente umida, uscì il sole che indurì i calchi in seguito ricoperti e conservati da un'altra eruzione vulcanica.
Studi dettagliati delle impronte rivelano un'anatomia delle parti molli molto simile alla nostra, che fornisce una prova cruciale della locomozione bipede di A. afarensis. Le impronte rivelano un forte colpo di tallone e una profonda impressione lasciata da un alluce non divergente che indica la non opponibilità dell'alluce stesso. Sono evidenti anche la rotondità alla base dell'alluce e gli archi, trasversale e longitudinale, del piede.
Dopo un attento studio anatomico gli ominidi di Radar e Laetoli vennero assegnati ad Australopithecus afarensis, una specie bipede, dal cranio e dai denti primitivi e dallo spiccato dimorfismo sessuale (Johanson et al., 1978). Un notevole numero di caratteri craniali, mandibolari e mascellari distingue A. afarensis dalle altre specie di australopitecini e specialmente da A. africanus (Johanson, 1989). L'istituzione della specie A. afarensis, le sue implicazioni filo genetiche e la sua presunta locomozione aprirono, in paleoantropologia, un dibattito che dura da un quarto di secolo. Per esempio, alcuni studiosi suggerirono che gli ominidi di Radar e Laetoli fossero semplici varianti settentrionali di A. africanus (Tobias, 1980). Altri hanno perfino messo in discussione l'unità di A. afarensis, ipotizzando per Radar la presenza di più di un taxon (Olson, 1985; Senut e Tardieu, 1985). Anche il modello del dimorfismo sessuale, che per alcuni indica due specie, e la capacità locomotoria di A. afarensis sono stati oggetto di discussione (Susman et al., 1984).
Fortunatamente, ben 75 altri reperti di ominidi recuperati a Radar a partire dal 1990 aiutano a risolvere tali questioni (Kimbel et al., 1994). Uno degli esemplari più importanti è il primo cranio, quasi completo, di A. afarensis adulto, recuperato da uno strato di 3 milioni di anni fa. L'esemplare, il cranio di australopitecino più antico e più completo mai rinvenuto, ha grandi canini e reca i segni di forti muscoli, che suggeriscono trattarsi di un maschio. La maggior parte dei caratteri craniofacciali di questo eccezionale reperto conferma una precedente ricostruzione di A. afarensis, basata sui frammenti di individui differenti (Kimbel et al., 1984).
L'esemplare A.L. 444-2 si distingue per le grandi dimensioni e ha il più grande cranio di Australopithecus mai trovato, anche se il cervello aveva un volume di soli 550 cm³. Numerosi dettagli anatomici del cranio distinguono A. afarensis dalle altre specie di australopitecini. Per esempio, il cranio di Radar ha una cresta sagittale molto bassa e posizionata posteriormente; le fibre dei muscoli masticatori, i temporali, sono orientate orizzontalmente, forse in risposta alla pressione delle forze sviluppate dalla grossa, e assai logora, dentatura anteriore. Le fibre dei temporali nei 'robusti' (A. robustus e A. boisei) sono dirette più verticalmente, con una cresta sagittale situata in avanti, e ciò riflette un'enfasi sulla forza masticatoria verticale. La condizione di A. afarensis ricorda quella degli scimpanzé e dei gorilla. Un'ulna virtualmente completa (A.L. 438-1), anatomicamente identica a quella di 'Lucy' ma più lunga del 22%, conferma il modello dello spiccato dimorfismo sessuale precedentemente documentato dal campione di Radar. Un omero breve, di robusta costituzione, specie se in combinazione con un lungo avambraccio, può riaprire il dibattito sui comportamenti da arrampicatore e su quelli alimentari di A. afarensis. A questo riguardo alcuni ricercatori (Susman et al., 1984) hanno suggerito che, se pure A. afarensis poteva essere bipede a terra, esso camminava ancora con un'andatura ondeggiante, a ginocchia flesse. Non è irragionevole pensare che questi primi ominidi si siano avventurati nuovamente sugli alberi per sfuggire al pericolo, costruire rifugi per dormire, o per cercare cibo. Quasi tutti i ricercatori accettano oggi la conclusione che i reperti di Radar siano riferiti a un'unica specie, A. afarensis, ma una piccola minoranza continua a postulare la presenza di due taxa. Anche se è sempre possibile che a Radar vi sia una seconda specie, si è dimostrato impossibile precisare un insieme di caratteri anatomici che differenzino i due taxa. La crescente fiducia che a Radar vi sia una sola specie di Australopithecus deriva dal grande aumento del campione dentario e mandibolare del sito. Sebbene il campione allargato mostri maggiori variazioni dimensionali, mancano variazioni anatomiche significative, il che rinforza quindi la tesi che nei depositi di Radar vi sia una singola specie di Australopithecus.
Manca per ora un consenso sull'identità dell'antenato di A. afarensis. Secondo alcuni ricercatori i tratti primitivi, scimmieschi, di A. ramidus ne fanno, per gli ominidi più tardi, un antenato meno probabile rispetto ad A. anamensis (Leakey et al., 1995). Per contro, virtualmente tutti i tratti anatomici a noi noti di quest'ultima specie ne fanno un antenato ideale della specie A. afarensis. Gli stati dei caratteri della morfologia della mascella e dei denti di A. anamensis sono solo leggermente più primitivi di quelli riscontrati in A. afarensis.
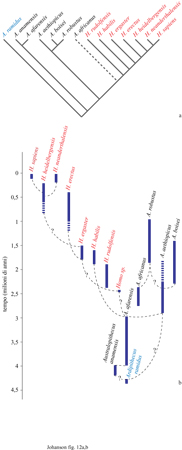
Dopo che una singola specie, A. afarensis, visse per circa un milione di anni, l'albero genealogico degli ominidi si diversificò in due distinti percorsi evolutivi (fig. 12). Un ramo portò a Homo, un ceppo - forse associato ai primi utensili in pietra - in cui la selezione favorì l'espansione del cervello e la riduzione dell'apparato masticatorio. La tendenza adattativa ed evolutiva dell'altro ramo enfatizzò invece il sistema masticatorio, avendo come esito una progressiva specializzazione nel regime vegetariano e la comparsa degli australopitecini 'robusti'.
Uno degli aspetti più degni di nota dell'albero genealogico degli ominidi presentato (v. figura 12) riguarda la posizione di A. africanus. Generalmente gli alberi filo genetici degli ominidi pongono questa specie come l'ultimo antenato comune a tutti gli ominidi cronologicamente successivi (McRenry, 1996). Con la nuova prospettiva offerta da A. afarensis, è stata rivista anche la relazione evolutiva tra A. africanus e Homo (Johanson e White, 1979). Un ampio studio dei caratteri cranio dentali di questi primi ominidi ha evidenziato in A. africanus una moltitudine di caratteri derivati già indirizzati nella direzione degli australopitecini 'robusti'. Tali caratteri - come l'ingrossamento della dentatura posteriore, il crescente rafforzamento del corpo mandibolare, l'ampliamento delle aree di inserzione dei muscoli masticatori - preannunciano tutti la specializzazione masticatoria tipica dei 'robusti'. A. africanus, perciò, costituisce un'eccellente specie intermedia tra il più indifferenziato A. afarensis e i più specializzati 'robusti'. Inoltre, questa ipotesi predice che quando si troveranno fossili intermedi tra A. afarensis e Homo essi non somiglieranno ad A. africanus.
Nel 1985, la scoperta a ovest del Lago Turkana di un cranio relativamente completo, soprannominato 'Teschio nero' (Black skull), ha fatto ampia luce sul ritmo del cambiamento morfologico, sul carattere di tale variazione e sulla possibilità di molteplici linee filetiche nell'evoluzione degli australopitecini (Walker et al., 1986). Questo esemplare, KNM - WT 17000, ha circa 2,5 milioni di anni e presenta un'interessante mescolanza di tratti anatomici primitivi (tipo A. afarensis) e derivati (tipo A. boisei).
Soprattutto, il 'Teschio nero', la cui colorazione è dovuta a minerali di manganese scuri, ha una faccia larga, denti posteriori estremamente ingrossati e una grande cresta sagittale a flangia, somigliando così ai più tardi australopitecini 'robusti' e specialmente ad A. boisei. Tuttavia, questo esemplare possiede anche un certo numero di caratteri primitivi, apparentemente ereditati dal suo presunto antenato A. afarensis: tra i più evidenti vi sono la faccia prognata, la ridotta capacità cranica (410 cm³), l' arretramento della cresta sagittale e del complesso della cresta temporo-cervicale sulla parte posteriore del cranio.
Gli scopritori (Walker et al., 1986) hanno sottolineato i tratti del tipo A. boisei riferendo l'esemplare a questa specie. Altri ricercatori (Kimbel et al., 1988), hanno accuratamente comparato 32 caratteri craniali riscontrati in KNMWT 17000 con altre specie conosciute di Australopithecus e hanno raggiunto una diversa conclusione. Se da un lato l'esemplare presenta alcuni tratti morfologici esclusivi, dall'altro esso ha in comune esclusivamente con A. boisei solo due caratteri craniali, mentre i restanti 30 sono variamente condivisi con altre specie di australopitecini. Questa specifica commistione di caratteri unici, avanzati e primitivi, suggerì che l'esemplare rappresentasse una nuova specie, ancestrale e distinta rispetto ad A. boisei.
L'attribuzione specifica, A. aethiopicus, fu tratta dal nome dato a una mandibola di 2,5 milioni di anni proveniente dalla formazione di Shungura in Etiopia meridionale (Arambourg e Coppens, 1968). L'antichità di questa specie (2,5 milioni di anni) ne fa un antenato possibile di A. robustus, di A. boisei, o di entrambe le specie. Forse ancor più interessante è la conclusione che A. africanus non può essere l'antenato di tutti i più tardi 'robusti', dal momento che è privo dei tratti primitivi che A. afarensis e A. aethiopicus condividono.
Un'ipotesi plausibile suggerisce che A. africanus abbia dato origine ad A. robustus nell'Africa meridionale e che A. aethiopicus sia stato il precursore di A. boisei neII'Africa orientale. Se ulteriori rinvenimenti dovessero confermare questa ipotesi, allora due rami paralleli di australopitecini si sarebbero evoluti separatamente in specie 'robuste'. Presumibilmente in risposta ai cambiamenti ambientali e alle conseguenti specializzazioni alimentari, i due australopitecini 'robusti' si sarebbero diversificati da un antenato comune, A. afarensis, sviluppando adattamenti simili in modo indipendente.
Per quanto concerne la sequenza delle variazioni morfologiche, è interessante notare che nel ceppo 'robusto' dell'Africa orientale l'ingrossamento dentario e mandibolare, come anche l'ipertrofia dei muscoli masticatori, precede la riduzione del prognatismo facciale. Inoltre, l'ampia porzione anteriore dell'arco dentale superiore probabilmente ospitava degli incisivi allargati, tipici di A. afarensis, e non incisivi ridotti, caratteristici di A. boisei. Il cranio KNMWT 17000 fornisce preziosi elementi sulle modalità dei cambiamenti evolutivi in cui la selezione naturale favorisce l'espansione del sistema masticatorio prima della flessione della base cranica, con conseguente verticalizzazione della faccia e dentatura anteriore ridotta.
Origini del genere Homo
Attualmente i resti fossili del genere Homo, a cui apparteniamo, sono relativamente poco conosciuti. Sebbene sia stato ampiamente sostenuto che A. africanus costituisce il candidato più probabile come antenato di Homo, si può obiettare che motivi validi possono essere avanzati a sostegno di una connessione evolutiva diretta tra A. africanus e gli australopitecini 'robusti' (v. sopra). Se questa ipotesi si dimostrerà infine corretta, allora Homo non ha predecessori riconoscibili. Una possibilità è che A. afarensis sia stato un antenato comune non solo agli australopitecini successivi, ma anche a Homo. Per il momento, è impossibile verificare questa ipotesi a causa della sostanziale mancanza di reperti fossili nella linea di Homo, dal più recente reperto documentato di A. afarensis al primo reperto di Homo. Le pretese concernenti un Homo di 2,5 milioni di anni fa e fondate su un osso temporale proveniente da Cameron, in Kenya (Hill et al., 1992), e su una mandibola proveniente da Chiwondo, in Malawi (Bromage et al., 1995), sono state contestate per la dubbia datazione geologica e per la carenza di caratteri anatomici diagnostici che permettano una chiara attribuzione dei reperti al genere Homo.
Tradizionalmente, la ricerca rivolta alla comprensione dell' evoluzione di Australopithecus ha in larga misura offuscato quella rivolta alla conoscenza del genere Homo. Recentemente, tuttavia, la situazione sta cambiando (Wood, 1991) e si inizia a intravedere una migliore percezione dell'antichità e della varietà del nostro genere. A partire dal 1964, quando L.S.B. Leakey, P.V. Tobias e IR. Napier dettero l'annuncio del ritrovamento di Homo habilis, quasi tutti gli ominidi plio-pleistocenici non riferibili agli australopitecini furono collocati in questa specie o in Homo erectus (l'Uomo di Giava e l'Uomo di Pechino).
Molte voci autorevoli hanno messo in dubbio queste attribuzioni tassonomiche, sostenendo da un lato che H erectus era una specie esclusivamente non africana e, dall'altro, che il primo Homo africano presentava una maggiore diversità tassonomica di quanto precedentemente supposto. Una mandibola proveniente dal Lago Turkana fu assegnata da C.P. Groves e V. Mazak (1975) a una nuova specie, Homo ergaster, che include anche uno scheletro straordinariamente completo noto come KNM-WT 15000 (Walker e Leakey, 1993) e due crani (KNM-ER 3773 e 3883), anch'essi provenienti dal bacino del Lago Turkana. Un'ulteriore diversificazione in specie iniziali del genere Homo è atte stata dall'assegnazione, avvenuta a opera di V.P. Alexeev (1986), di un ben conosciuto cranio (KNM-ER 1470) del Lago Turkana a una nuova specie chiamata Homo rudolfensis.
La peculiarità di Homo habilis fu corroborata dal recupero nella gola di Olduvai, nel 1986, di uno scheletro parziale, di proporzioni minute (Johanson et al., 1987). Questo esemplare, noto come 'Olduvai Rominid 62' , ha mascella e denti simili ad altri esemplari di Olduvai assegnati a H habilis. Tuttavia, gli arti superiori relativamente lunghi e quelli inferiori corti sono più simili a quelli dei primi australopitecini, come 'Lucy'. D'altra parte, Homo ergaster ha una struttura fisica molto simile a quella dell 'uomo moderno per taglia e proporzioni.
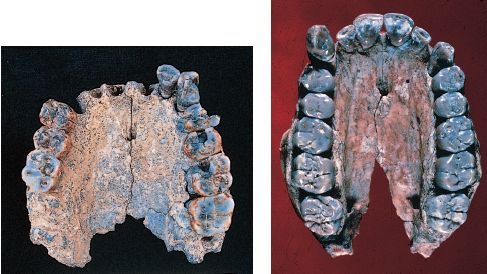
La scoperta a Radar in Etiopia di una mascella risalente a 2,3 milioni di anni (fig. 15) rappresenta la testimonianza certa più antica del genere Homo in associazione con indubbi artefatti in pietra (Kimbel et al., 1996; Kimbel et al., 1997). Il reperto, un palato quasi completo contenente dieci denti, possiede una serie di tratti anatomici comuni a Homo e distinti da Australopithecus. Essi includono l'assenza di prognatismo, una profonda volta palatale e un'arcata dentaria ampia e tondeggiante. Sfortunatamente, la limitatezza delle testimonianze fossili del primo Homo non permette di assegnare l'esemplare di Radar ad alcuna delle specie già esistenti, né di determinare se essa rappresenti una nuova specie di Homo. Per la soluzione di questo problema tassonomico occorre attendere l'eventuale ritrovamento di un materiale più completo.
Conclusioni
In questa breve panoramica sull'evoluzione dei primi ominidi è stata posta l'attenzione sugli australopitecini, con solo qualche breve considerazione sul primo Homo. In linea di massima, questa situazione riflette la natura del materiale fossile per i primi ominidi di cui si dispone attualmente: mentre la scoperta di nuovi fossili di australopitecini procede di buon passo, la comprensione delle origini, della diversificazione e degli adattamenti nel genere Homo continua a essere ostacolata dalla penuria dei reperti fossili. È auspicabile che la ricerca paleoantropologica dei prossimi anni attribuisca un'alta priorità al recupero di fondamentali, e indispensabili, fossili di ominidi da depositi fossiliferi più antichi di 2 milioni di anni.
Tuttavia, la scoperta di nuovi esemplari fossili non fornirà risposte complete se per questi ritrovamenti non verranno adottati migliori metodi di analisi filo genetica. Per quasi un secolo, il campo della paleoantropologia è stato spinto dal desiderio di fare ulteriori scoperte, nella speranza che una maggior quantità di fossili avrebbe fornito tutte le risposte alle domande sull'evoluzione umana. Una delle principali insufficienze di questa nozione è spesso consistita in una serie di assunzioni a priori, che hanno fortemente influenzato l'interpretazione e vincolato il significato evolutivo dei fossili stessi. È solo abbandonando questi preconcetti e combinando metodi più solidi di analisi filo genetica con una valutazione allargata (per esempio, al contesto geologico, all'ecologia, all'adattamento) delle testimonianze fossili che potremo cogliere la vera natura del passato evolutivo dell'uomo.
Bibliografia citata
ALEXEEV, V.P. (1986) The origin ofthe human race, tradotto dal russo da H. Campbell Chreighton. Mosca, Progress Publishers.
ARAMBOURG, C., COPPENS, Y. (1968) Découverte d'un australopithécien nouveau dans les gisements de l'Orno (Ethiopia). South African Journal of Science, 64, 58-59.
BRAIN, C.K. (1981) The hunters or the hunted? An introduction to african cave Taphonomy. Chicago, Univ. of Chicago Press.
BROMAGE, T.G., SCHRENK, F., ZONNEVELD, F.W. (1995) Paleoanthropology of the Malawi Rift: an early hominid mandible from the Chiwondo beds, northern Malawi. Journal of Human Evolution, 28, 71-108.
BRUNET, M., BEAUVILAIN, A., COPPENS, Y., HEINTZ, E., MOUTAYE, A.H.E., PILBEAM, D. (1996) Australopithecus bahrelghazali, une nouvelle espèce d'Hominidé ancien de la région de Koro Toro (Tchad). Cr. Acad. Sci., 322, 907-913.
COPPENS, Y., HOWELL, F.C., LEAKEY, R.E.F. (1976) Earliest man and environments in the Lake Rudolf basin: stratigraphy, paleoecology and evolution. Chicago, Univ. of Chicago Press.
DELSON, E. (1988) Chronology of South African Australopith site units. In Evolutionary history ofthe 'Robust' Australopithecines, a c. di Grine F.E., New York, A. de Gruyter, pp. 317-324.
GRINE, F.E. a c. di (1988) Evolutionary history of the 'Robust' Australopithecines. New York, A. de Gruyter.
GROVES, C.P., MAZAK, V. (1975) An approach to the taxonomy of the Hominidae: gracile Villafrancman hominids of Africa. Caso Miner. Geol., 20, 225-247.
HILL, A., WARD, S.C., DEINO, A., CURTI S, G., DRAKE, R. (1992) Earliest Homo. Nature, 355, 719-722.
JOHANSON, D.C. (1989) The current status of Australopithecus. In Hominidae: Proceedings of the 2nd international congress of human paleontology, a c. di Giacobini G., Milano, Jaca Books, pp. 77-96.
JOHANSON, D.C. (1996) The strategy of paleoanthropology: early African hominids. Yearbook of physical anthropology, 39, 1-10.
JOHANSON, D.C., MASAO, F.T., ECK, G.G., WHITE, T.D., W ALTER, R.C., KIMBEL, W.H., ASFAW, B., MANEGA, P., NDESSOKIA, P., SUWA, G. (1987) New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania. Nature, 327, 205-209.
JOHANSON, D.C., WHITE, T.D. (1979) A systematic assessment of early African hominids. Science, 202, 321-330.
JOHANSON, D.C., WHITE, T.D., COPPENS, Y. (1978) A new species ofthe genus Australopithecus (Primates: Hominidae) from the Pliocene of Eastern Africa. Kirtlandia, 28, 1-14.
KIMBEL, W.H., JOHANSON, D.C., RAK, Y. (1994) The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia. Nature, 368, 449-451.
KIMBEL, W.H., JOHANSON, D.C., RAK, Y. (1997) Systematic assessment of a maxilla of Homo from Hadar, Ethiopia. Am. J. Phys. Anthropol., 103, 235-262.
KIMBEL, W.H., MARTIN, L.B., a c. di (1993) Species, species concepts and primate evolution. New Y ork, Plenum Press.
KIMBEL, W.H. et a!. (1996) Late Pliocene Homo and Oldowan tools from the Hadar Formation (Kada Hadar Member), Etmopia. Journal of human evolution, 31, 549-561.
KIMBEL, W.H., WHITE, T.D., JOHANSON D.C. (1984) Cranial morphology of Australopithecus afarensis: a comparative study based on composite reconstruction of the adult skull. Am. J. Phys. Anthropol., 64, 337-388.
KIMBEL, W.H., WHITE, T.D., JOHANSON, D.C. (1988) Implications of KNM- WT 17000 for the evolution of 'Robust' Australopithecus. In Evolutionary history of the 'Robust' Australopithecines, a c. di Grine F.E., New York, A. de Gruyter, pp. 259-268.
LEAKEY, M.D., HARRIS, 1M., a c. di (1987) Laetoli: A Pliocene site in Northern Tanzania. Oxford, Clarendon Press.
LEAKEY, M.G., FEIBEL, C.S., McDOUGALL, I., WALKER, A.C. (1995) New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya. Nature, 376, 565-57l.
McHENRY, H. (1992) How big were early hominids? Evolutionary anthropology, 1, 15-20.
McHENRY, H. (1996) Homoplasy, clades and hominid phylogeny. In Contemporary issues in human evolution, a c. di Meikle W.E., Howell F.C., Jablonski N.G., San Francisco, California Academy of Sciences, pp. 77-89.
MEIKLE, W.E., PARKER, S.T. (1994) Naming our ancestors: an anthology of hominid taxonomy. Prospect Heights, Waveland Press.
OLSON, T.R. (1985) Taxonomic affinities of the immature hominid cranium from Hadar and Taung. Nature, 316, 539-540.
PILBEAM, D. (1982) New hominoid skull material from the Miocene of Pakistan. Nature, 295, 232-234.
PILBEAM, D. (1996) Genetic and morphological records of the Hominoidea and hominid origins: a synthesis. Mol. Phylogenet. Evol., 5, 155-168.
SARICH, V.M. (1992) Immunological evidence on primates. In The Cambridge Encyclopedia of Human Evolution, a c. di Jones S., Martin R., Pilbeam D., New York, Plenum Press, pp. 303-306.
SENUT, B., TARDIEU, C. (1985) Functional aspects of Plio-Pleistocene hominid limb bones: implications for taxonomy and phylogeny. In Ancestors: The hard evidence, a c. di Delson E., New York, Alan R. Liss, pp. 193-201.
SUSMAN, R.L., STERN, J.T., JUNGERS, W.L. (1984) Arboreality and bipedality in the Hadar hominids. Folia Primatol., 43, 113-156.
TATTERSALL, I. (1992) Species concepts and species identification in human evolution. Journal of Human Evolution., 22, 341-349.
TOBIAS, P.V. (1968) The cranium and maxillary dentition of Australopithecus (Zinjanthropus) boisei, Olduvai Gorge. VoI. 2, Cambridge, Cambridge University Press.
TOBIAS, P.V. (1980) 'Australopithecus afarensis' and A. africanus: critique and an alternative hypothesis. Palaeontologia africana, 23, 1-17.
WALKER, A.C., LEAKEY, R.E.F. (1993) The Nariokotome Homo erectus skeleton. Cambridge, Harvard University Press.
WALKER, A.C., LEAKEY, R.E.F., HARRIS, J.M., BROWN, F.H. (1986) 2.5 Myr Australopithecus boisei from west of Lake Turkana, Kenya. Nature, 322, 517-522.
WALTER, R.C. (1994) Age of Lucy and the First Family: singlecrystal 40Ar/39Ar dating of the Denen Dora and lower Kada Hadar members of the Hadar Formation, Ethiopia. Geology, 22, 6-10.
WALTER, R.C., ARONSON, J.L. (1993) Age and source ofthe Sidi Hakoma Tuff, Hadar Formation, Ethiopia. Journal of Human Evolution, 25, 229-240.
WHITE, T.D., SUWA, G., ASFAW, B. (1994) Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia. Nature, 371, 306-312.
WHITE, T.D., SUWA, G., ASFAW, B. (1995) Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia. Nature, 375, 88.
WOOD, B.A. (1991) Koobi Fora research project. Vol. 4: Hominid cranial remains. Oxford, Clarendon Press.
Bibliografia generale
AIELLO, L., DEAN, M.C. An introduction to human evolutionary anatomy. London, Academic Press, 1990.
JOHANSON, D.C., EDGAR, B. From Lucy to language. New York, Simon and Schuster Editions, 1996.
MEIKLE, W.E., HOWELL, F.C., JABLONSKI, N.G. Contemporary issues in human evolution. San Francisco, California Academy of Sciences, Memoir 21, 1996.
TATTERSALL, I. The fossil trail: how we know what we think we know about human evolution. New York, Oxford University Press, 1995.