
linguaggio
linguaggio
Le società animali richiedono sempre un sistema di comunicazione: nella specie umana tale sistema è costituito principalmente dal linguaggio. Indipendentemente dalle teorie sulla sua natura, vi è comune accordo nel considerare il linguaggio una proprietà specie-specifica. Una caratteristica fondamentale che lo distingue da altri sistemi di comunicazione, usati sia dall’uomo sia da altre specie, è di essere ‘discreto’, ossia formato da elementi bene definiti, al contrario della comunicazione animale che si svolge tramite segnali che possono essere scomposti in parti più elementari, sprovvisti singolarmente di significato. Ampio è il dibattito sull’origine del linguaggio tra chi considera che le sue proprietà più specifiche (per es., l’utilizzo di processi ricorsivi) non provengano dal processo evolutivo della comunicazione e chi invece sostiene che la sua comparsa sia stata graduale, a partire da altri sistemi di comunicazione, come quello gestuale. L’evidenza sperimentale di una base neurologica risale a poco più di 150 anni fa. Alcuni contributi (Paul Broca, Karl Wernicke) furono decisivi per determinare la specializzazione di aree cerebrali del linguaggio; oggi sia gli studi clinici sia i risultati sperimentali ottenuti con le tecniche di neuroimaging propendono per un modello di elaborazione del linguaggio che prevede sia l’integrazione sia la segregazione di specifici aggregati neuronali. [➔ afasia; Broca, area di; fonazione; fonemi; imaging cerebrale; neurone, Neuroni specchio; specializzazione emisferica]
Ogni società, umana e animale, è provvista di sistemi di comunicazione che permettono di trasmettere informazioni da un individuo (l’emittente) a un altro (il ricevente). Nella specie umana il sistema specifico di comunicazione, che si affianca ad altri sistemi di comunicazione condivisi con altre specie animali, è il linguaggio. Noam Chomsky, in Le strutture della sintassi (1957), delinea il l. come un nucleo astratto, indipendente da altre forme di comunicazione, di operazioni computazionali specifiche della specie umana: il l. prevede una serie di meccanismi che dispongono gli elementi di alcuni sottoinsiemi linguistici (tratti distintivi, fonemi, sillabe, morfemi e parole) in maniera grammaticalmente corretta, in modo tale da formare elementi di ordine superiore (sintagmi, frasi) che possono essere espressi con mezzi diversi: il l. gestuale, la scrittura e, infine, le parole, sia parlate sia scritte. Più specificamente, nell’ambito della grammatica generativa proposta da Chomsky, Marc D. Hauser e collaboratori sostengono, in un articolo comparso su Science nel 2002, che il l. è una facoltà allargata, presente anche in altre specie animali, e divisa a sua volta in una componente dedicata alla produzione e recezione dei suoni con valore linguistico e una componente costituita dal sistema concettuale e da una facoltà del l. in senso stretto, specifica del l. e presente solo nell’uomo. Questa seconda componente è costituita principalmente, se non unicamente, dalla facoltà ‘ricorsiva’, ossia la proprietà che hanno certe regole formali di potersi riapplicare indefinitamente sui loro stessi risultati.
Linguaggio umano e comunicazione animale
Solo la specie umana ha, secondo i proponenti della grammatica generativa, accesso ai processi ricorsivi, usati specificatamente per la conoscenza linguistica: si pensi per es. alla frase semplice «Anna ama Paolo». Utilizzando il verbo sapere la frase diventa complessa: «Io so che Anna ama follemente Paolo» e con un’ulteriore espansione «Anna, che studia all’Università, ama follemente Paolo». I limiti alla lunghezza della frase sono dati dalle proprietà delle componenti più periferiche, come la memoria a breve termine. Ne consegue che la distanza fra le parole di un enunciato è specificata (dipendenza sintattica non rigida) nei termini della posizione relativa che può essere espansa attraverso l’uso di regole ricorsive (si guardi, per es., la distanza fra le parole Anna e Paolo nelle frasi precedenti). Tra l’altro, recentemente è stato riscontrato, in uno studio di soggetti sani sottoposti a risonanza magnetica funzionale (fRM), che l’apprendimento di sequenze di simboli non verbali ordinati secondo una sintassi non rigida attiva le stesse aree cerebrali (area di Broca) che risultano attive durante l’elaborazione degli aspetti sintattici del linguaggio. La comunicazione animale è invece basata su regole che determinano una distanza fissa fra gli elementi del tipo AB; AB; AB… (dipendenza sintattica rigida). Solo alcuni passeriformi (Sturnus vulgaris) sono in grado, dopo un lungo addestramento, di riconoscere strutture acustiche di tipo ricorsivo (2006). In una prospettiva diversa il l. può essere considerato come un sistema di comunicazione specie-specifica, che consente di formare un numero praticamente infinito di segni, entità dotate di significato, morfemi e parole, usando un numero molto limitato di suoni linguistici che non hanno un significato, ma che, combinati insieme secondo regole specifiche, per ogni lingua parlata, permettono di costruire e distinguere parole e frasi.
Linguaggio e comunicazione
Una caratteristica fondamentale che distingue il l. da altri sistemi di comunicazione, usati sia dall’uomo sia da altre specie, è di essere ‘discreto’ per cui i suoi elementi si distinguono per il fatto di essere ben definiti: per es., in italiano, i suoni /p/ e /b/, per quanto molto simili dal punto di vista acustico e articolatorio, hanno, sia per chi li produce sia per chi li ascolta, un contrasto netto (patto ha un significato ben diverso da batto). Questa caratteristica del l. è detta percezione categorica (➔ fonemi) ed è stata a lungo considerata specifica del l. e specie-specifica; solo in tempi più recenti è stato riscontrato che la percezione categorica si può sviluppare, dopo un lungo addestramento, anche in alcuni animali, quali cincillà e macachi.
Origine del linguaggio
L’approccio filogenetico allo studio di alcune proprietà specifiche della specie umana, quali l’opposizione del pollice o la visione dei colori, ha permesso di evidenziarne una graduale evoluzione a partire da specie prossime. Il caso è invece differente per il l.: fin da Ippocrate si è sostenuto che il l. sia unicamente umano e anche Thomas Huxley, uno dei più accaniti promotori della teoria dell’evoluzione, si dimostrava molto scettico riguardo all’idea che il l. potesse essersi sviluppato gradualmente a partire dai sistemi di comunicazione animale. In tempi più recenti Chomsky si è vivacemente opposto alla teoria che il l. possa essere considerato come il punto più alto di un lungo processo evolutivo della comunicazione. Per Chomsky, infatti, la facoltà ricorsiva, specifica del l., si sarebbe sviluppata mediante un processo di exaptation («preadattamento»). Tale termine sta a indicare un cambiamento di funzione a parità di struttura anatomica: nel corso di una mutazione genetica si è sviluppato un modulo funzionale indipendente, dedicato alla soluzione di compiti quali la navigazione o la quantificazione numerica che, nella specie umana, si è applicato al linguaggio. Per altri autori, invece, la comparsa del l. è stata graduale, a partire da altri sistemi di comunicazione: secondo la teoria gestuale della comunicazione, la parola e il l. sono emersi a partire dai gesti prodotti dalle mani, dalla posizione del corpo o da espressioni facciali. Infatti, anche la produzione di suoni non verbali da parte dei primati a scopo comunicativo è accompagnata da vivaci movimenti dei muscoli facciali e del corpo, lasciando aperta la possibilità che la comunicazione acustica sia stata originariamente un complemento della comunicazione visiva dei gesti e delle azioni che producono rumore. Secondo questa prospettiva, pertanto, la comunicazione vocale è inserita in un modello interno di tipo multisensoriale che elabora informazioni sia visive sia acustiche a scopo comunicativo in zone adiacenti della corteccia cerebrale. D’altra parte ancora oggi gesti manuali sincronizzati con la produzione di parole fanno parte della comunicazione linguistica, come per ricordarne una comune origine. A favore di un’origine primitivamente gestuale del l. è la scoperta, alla fine del secolo scorso, nella scimmia, del sistema dei neuroni specchio (➔ neurone), posto all’interno della corteccia cerebrale frontale e parietale. Tale sistema si attiva sia durante l’esecuzione sia durante la visione di un movimento, così da crearne una rappresentazione interna che ne permette la comprensione. È quindi possibile che, grazie a tale sistema, il l. si sia evoluto da gesti, all’inizio con funzione di pantomima, che, a causa del progressivo aumento del vocabolario, si sono convenzionalizzati; infine sarebbe comparso un processo di grammaticalizzazione, necessario per esprimere la differenza fra episodi differenti nel tempo e nello spazio e per generare espressioni riguardanti il futuro. Una domanda a cui praticamente non si può dare risposta è quella riguardante il momento in cui si è iniziato a usare il l.: non esistono ovviamente fossili linguistici, né tantomeno documenti scritti in protolinguaggi (la scrittura si è sviluppata solo 5.000 anni fa); d’altra parte il cervello, base neurologica del l., non fossilizza. Una prospettiva interessante riguardante l’età di comparsa del l., o per lo meno delle asimmetrie emisferiche che sottendono il l., è quella derivante dallo studio nei crani fossili dei petalia, protrusioni asimmetriche di un lobo cerebrale che lasciano avvallamenti altrettanto asimmetrici nel tavolato interno delle ossa craniche. Tali avvallamenti, più profondi a livello dell’emicranio sinistro, potrebbero essere legati alla comparsa di asimmetrie emisferiche conseguenti alla comparsa della specializzazione emisferica sinistra per il linguaggio. L’esame dei calchi ricavati dai crani degli ominidi ha infatti rilevato una struttura dei petalia simile a quella attuale solo in Homo erectus o, secondo altri, solo in Homo sapiens, collegata probabilmente a una mutazione genetica critica per l’apparizione del linguaggio.
Parlare con la testa: basi neurologiche del linguaggio
L’evidenza sperimentale di una base neurologica e la costruzione di un modello anatomofunzionale sottostante il l. risalgono a poco più di 150 anni fa, quando, specie in Francia e Germania, si diffuse rapidamente il concetto che le funzioni cognitive e morali, in analogia a sistemi e funzioni biologiche quali la circolazione sanguigna o la respirazione, avevano una base fisica e potevano essere sperimentalmente indagate. Di conseguenza, i frenologi (Franz J. Gall, Johann G. Spurzheim) postularono che nella corteccia cerebrale fossero rappresentate in maniera puntuale almeno trenta facoltà, dalla conoscenza metafisica alla memoria per le parole. Pur senza fornire alcuna prova scientifica, la frenologia esercitò un ruolo fondamentale nella ricerca delle basi neurologiche del l., considerandolo non come una facoltà unica, inaccessibile all’indagine sperimentale, ma composto di facoltà distinte, sottese da precisi luoghi anatomici. Il primo fondamentale contributo sperimentale fu fornito dal neurologo francese Paul Broca, attraverso il metodo anatomoclinico che consente di stabilire una connessione fra lesioni di aree specifiche cerebrali – evidenziabili all’esame anatomopatologico o, più recentemente, in vivo, grazie alle tecniche di neuroimaging – e disturbi cognitivi. Applicando il ‘principio di trasparenza’, secondo il quale il deficit cognitivo sotto esame non riflette un abbassamento cognitivo globale, ma rappresenta l’espressione di un sistema cognitivo intatto privo di quella componente sottesa dalla zona cerebrale danneggiata, è stato possibile costruire una geografia e un’architettura funzionale delle funzioni cognitive.
Simmetria emisferica e linguaggio
In una seduta della Société d’anthropologie di Parigi, Broca riportò (1861) l’osservazione anatomoclinica di un paziente che, molti anni prima, aveva perso l’uso della parola, con una capacità espressiva limitata alla sillaba tan; al contrario, la comprensione uditiva e le altre facoltà cognitive erano clinicamente nella norma. All’esame anatomopatologico (il paziente era deceduto per cause extraneurologiche), Broca riscontrò gli esiti di una vasta lesione vascolare alla base della terza circonvoluzione frontale (in seguito denominata area di Broca), che ritenne quindi essere la base neurologica «della memoria delle parole articolate». Negli anni successivi Broca, sulla base di altri casi analoghi, dimostrò un ruolo funzionale asimmetrico dei due emisferi cerebrali; più specificatamente l’emisfero sinistro controlla, in oltre il 90% della specie umana, sia il l. sia la preferenza manuale, un altro tratto specie-specifico.
Il modello di Wernicke-Lichtheim
Nel 1874 Karl Wernicke propose un modello di organizzazione neurologica del l., secondo il quale nell’area di Broca sono depositati i programmi motori necessari per l’articolazione delle parole; di conseguenza, una sua messa fuori uso – per una causa vascolare, traumatica o di altro genere – porta a un disturbo di produzione del l., mentre la comprensione linguistica rimane clinicamente nella norma (afasia di Broca). Nell’area di associazione acustica, situata nella parte posteriore della circonvoluzione superiore temporale sinistra – e in stretta connessione con l’area acustica primaria –, avviene invece il processo di decodificazione del l., per cui gli stimoli uditivi sono trasformati in unità linguistiche, i fonemi. Una lesione in tale zona si evidenzia con un deficit selettivo della comprensione del l., sia parlato sia scritto (afasia sensoriale o di Wernicke). Wernicke ipotizzò l’esistenza di una connessione fra le due aree, per effetto di un arco riflesso, attivo nel processo di acquisizione del linguaggio. Tale modello, che incontrò un immediato e costante favore fra i clinici, fu completato pochi anni dopo da Ludwig Lichtheim che propose un modello a due stadi, uno inferiore in cui avviene la elaborazione degli stadi più periferici del l. (codificazione e decodificazione delle parole), e uno superiore (centro dei concetti, di cui però Lichtheim non specificava la localizzazione all’interno della corteccia cerebrale), in cui avviene il processo di elaborazione semantico-lessicale della lingua, collegato con il centro verboacustico e il centro verbomotorio.
Superamento dei modelli classici. Nel modello di Wernicke-Lichtheim la produzione del l. è considerata un atto essenzialmente motorio, separato dal processo di comprensione che implica esclusivamente il coinvolgimento delle vie uditive. Tale parcellizzazione del sistema si è rivelata, alla luce di dati ricavati da fonti diverse (dati neurofisiologici e di neuroimaging) fondamentalmente errata. Anche la nozione che l’area di Broca svolga un ruolo esclusivamente di controllo neuromotorio sulla produzione del l. è stata profondamente ridimensionata: numerosi studi clinici hanno, infatti, evidenziato che una lesione di tale area porta a un’incapacità specifica di elaborazione sintattica, sia in produzione sia in comprensione (agrammatismo). La sua stimolazione con elettrodi applicati a pazienti non afasici, in corso di interventi neurochirurgici, determina un transitorio deficit nella comprensione di frasi grammaticalmente complesse. Infine, è stata dimostrata da Marco Tettamanti e collaboratori (2009) un’attivazione di tale area nell’apprendimento di sequenze sia verbali sia non verbali, ordinate da una sintassi non rigida, ipotizzando un ruolo specifico di tale area nell’elaborazione di specifiche regole sintattiche.
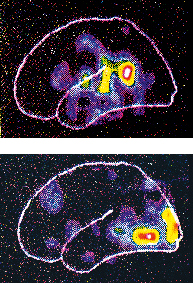
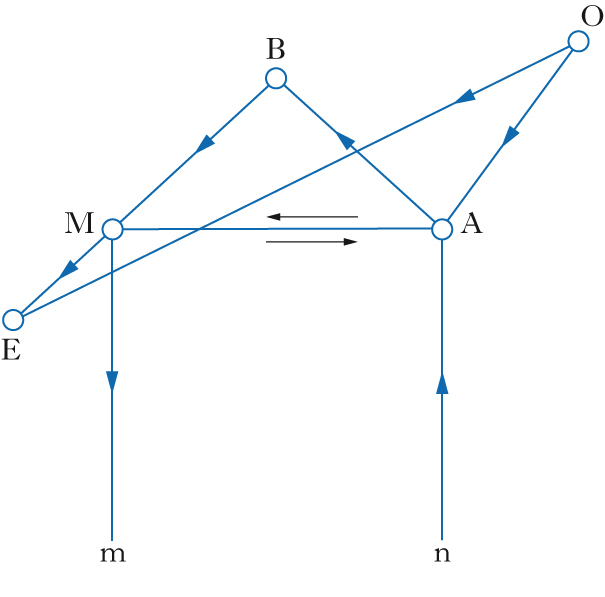
Modelli basati su integrazione/segregazione di aree cerebrali. Uno studio fRM condotto da Friedemann Pulvermüller e collaboratori (2006) su soggetti normali ha dimostrato che, durante l’ascolto di fonemi occlusivi bilabiali (/p/) e dentali (/t/), si registra un’attivazione contemporanea a livello sia delle aree uditive del lobo temporale sia delle aree motorie precentrali, con un’ulteriore specificazione a seconda del fonema elaborato: in corrispondenza della rappresentazione motoria delle labbra per /p/ e della lingua per /t/. Tali dati ben si accordano con quelli ricavati dalla scoperta del sistema dei neuroni specchio da parte di Giacomo Rizzolatti: accanto a neuroni visuomotori ‘canonici’, che rispondono alla presentazione di uno stimolo visivo, nella corteccia premotoria frontale (area 5) della scimmia è stata identificata una particolare classe di neuroni che si attivano sia durante l’esecuzione sia durante l’osservazione di azioni compiute da un’altra scimmia o dallo sperimentatore. Successivamente, sia nella scimmia che nell’uomo, è stato identificato un sistema specchio più ampio, che interessa, oltre alle regioni frontali, i lobi parietali, e che si attiva durante l’osservazione di azioni e di suoni corrispondenti eseguiti sia con gli arti sia con i muscoli orofacciali. Sulla stessa linea, Darren R. Gitelman e collaboratori (2005) hanno osservato, in uno studio fRM, che aree cerebrali differenti si attivano a seconda del compito, fonologico (decidere se due parole rimano fra loro), semantico (se le parole sono sinonimi) o di somiglianza ortografica. Indipendentemente dal tipo di consegna, si è osservato un pattern comune di attivazione per le tre prove nella regione temporo-occipitale sinistra; di contro si è osservato un pattern di attivazione selettiva della corteccia frontale anteriore durante il compito semantico-lessicale, di quella frontale posteriore nei compiti di analisi fonologica, e infine di quella parietale in compiti di analisi ortografica. Tali dati quindi sono in favore di un modello di elaborazione del l. che prevede sia l’integrazione sia la segregazione di specifici aggregati neuronali.
Studi sul lessico. A livello sia clinico che sperimentale sono state evidenziate aree specifiche per l’elaborazione di specifiche categorie grammaticali e lessicali: per es., Daniela Perani e collaboratori (1999) hanno riscontrato un’attivazione neuronale in corrispondenza dell’area dorsolaterale frontale e temporolaterale sinistra in compiti di elaborazione di verbi, mentre l’elaborazione di nomi si accompagna a un’attivazione emisferica diffusa, prevalente a sinistra.
Le regioni della voce
Nell’uomo, il mezzo più naturale di esprimere il l. è la voce, suono vocale specie-specifico: essa si distingue infatti sia dagli altri suoni emessi dal tratto vocale dall’uomo e da altri animali, sia dai ‘gesti acustici’ prodotti a scopo comunicativo con altri mezzi, quali il bussare a una porta, applaudire o battere il pugno sul tavolo. La sua produzione necessita di un raffinato sistema di controllo sui muscoli laringei, articolatori e respiratori, attraverso la formazione di mappe neurali delle relazioni fra muscoli, comandi motori e feedback uditivo (➔ fonazione). Oltre a trasmettere suoni con valore linguistico (fonemi, parole), la voce riveste un ruolo fondamentale nel processo di distinzione e riconoscimento di conspecifici: ogni persona infatti ha la sua voce, in qualche modo analoga alle caratteristiche fisiognomiche che rendono ogni faccia unica, permettendo quindi di distinguere le voci estranee, familiari e infine attribuendo loro una identità. Christopher Petkov e collaboratori (2008), attraverso la tecnica di fRM, hanno localizzato, a livello del solco temporale superiore, una regione specializzata nell’elaborazione delle caratteristiche acustiche della voce umana, non presenti in altri suoni sia naturali sia artificialmente prodotti; tale regione è inoltre sensibile alle proprietà di voci differenti. La scoperta della ‘regione della voce’ nell’uomo ha ovviamente innescato una serie di ricerche volte a evidenziare nei primati strutture neurologiche analoghe, specifiche per l’analisi di segnali acustici usati a scopo comunicativo, seppure con alcuni caveat: l’analisi della vocalizzazione umana potrebbe basarsi infatti prevalentemente su indici di tipo linguistico. Studi fRM eseguiti su macachi hanno inoltre dimostrato un’attivazione dell’area cerebrale corrispondente al piano superotemporale durante l’ascolto di suoni vocalici prodotti da membri della stessa specie.
Tale regione sembra essere specifica per l’analisi di suoni a scopo comunicativo: si è infatti evidenziato che essa si attiva anche all’ascolto di suoni non vocali (suoni ripetitivi, come battere con le mani su oggetti), usati dai macachi a scopo comunicativo. È quindi possibile ipotizzare che le regioni cerebrali specifiche per il riconoscimento vocale nell’uomo si siano sviluppate mediante una modificazione di un’area cerebrale attiva nei processi di comunicazione vocale. In conclusione, i modelli attuali di elaborazione neurologica del l. ipotizzano la presenza di una complessa rete neuronale che elabora on line le componenti sensorimotorie e lessicali del l., mediante meccanismi sia generali sia specifici per singoli compiti. Gianfranco Denes
Le reti del linguaggio
L’uso del linguaggio richiede un flusso di informazioni che interessa un grande numero di aree cerebrali tra loro connesse, il cui insieme costituisce le reti del linguaggio. La mancanza di facoltà omologhe al linguaggio in specie non umane ha impedito la costruzione di modelli animali, pertanto le conoscenze sulle basi neurali del linguaggio si sono basate su altre fonti, in primo luogo lo studio di soggetti con afasia (➔). Queste ricerche hanno innanzitutto mostrato il ruolo dominante nel linguaggio dell’emisfero sinistro, mettendo in evidenza due principali aree corticali connesse al linguaggio, la prima, l’area di Broca (➔), localizzata nella regione frontale, e la seconda, l’area di Wernicke (➔), nel giro temporale superiore (GTS) in posizione posteriore.
Il modello di Wernicke-Lichtheim-Geschwind
Queste osservazioni hanno portato a un primo modello di reti del linguaggio, formulato da Karl Wernicke e successivamente rivisto prima da Ludwig Lichtheim e poi da Norman Geschwind. In questo modello si ipotizza che le aree di Wernicke e di Broca svolgano, rispettivamente, il ruolo di analizzare le ‘immagini acustiche’ delle parole e quello di articolarle; il fascicolo arcuato trasporta informazioni dall’area di Wernicke a quella di Broca per il passaggio dalla comprensione alla articolazione della parola e si presume che ciascuna di queste aree interagisca con aree associative polimodali (➔ funzioni cerebrali superiori), tra le quali assume importanza cruciale il giro angolare (area di Brodmann 39). Geschwind sosteneva che il giro angolare fosse importante per l’associazione tra il nome udito e l’oggetto visto o toccato. Questo modello ha costituito la base per la classificazione delle afasie e ha rappresentato uno schema per le successive ricerche sulle reti del linguaggio. Tuttavia, numerosi studi hanno dimostrato che esso presenta notevoli limitazioni: non rende conto della varietà delle sindromi afasiche, non é sufficientemente specifico rispetto ai sottosistemi linguistici ed esclude aree che contribuiscono all’elaborazione linguistica.
Modelli di reti più complessi
Oggi é chiaro che la differenza di ruolo delle aree di Wernicke e di Broca non è così netta come inizialmente ipotizzato. Per es., soggetti con afasia di Broca non hanno la comprensione del linguaggio completamente intatta, ma mostrano difficolta nella comprensione di frasi che richiedono analisi sintattica la quale, a sua volta, implica la memoria di lavoro (circuito di ripasso articolatorio). Anche l’anatomia del fascicolo arcuato é molto più complessa di come inizialmente pensato, in quanto non connette esclusivamente le sole aree di Wernicke e di Broca. Molte altre aree corticali e sottocorticali dell’emisfero sinistro vanno incluse nelle reti del linguaggio. Tra queste: le cortecce associative polimodali frontali, temporali e parietali, che potrebbero essere l’interfaccia fra concetti e linguaggio; parti della regione dell’insula (area corticale sita nella profondità della scissura che separa il lobo temporale da quello frontale), che potrebbero essere importanti per l’articolazione del linguaggio; aree prefrontali che consentirebbero l’apporto della memoria di lavoro. Per es., il GTS, il giro frontale inferiore e la corteccia parietale inferiore permetterebbero la decodifica fonologica, mentre le aree del giro temporale mediale opererebbero la comprensione a livello di parola. Anche la corteccia supplementare motoria giocherebbe un ruolo, con le zone anteriori attive in relazione alla selezione del lessico, quelle posteriori nel controllo motorio. Le aree inferotemporali sarebbero importanti non solo per il riconoscimento visivo di categorie di oggetti ma anche per la loro denominazione. Se sono lesionate zone circoscritte delle aree inferotemporali, la difficolta di denominazione é limitata a categorie di oggetti, per es., luoghi o persone ma non oggetti di uso comune. Lo schema attuale (2010) delle reti del linguaggio prevede: un sistema di implementazione, che analizza i segnali uditivi (e visivi) afferenti in modo da attivare la loro conoscenza concettuale é che provvede alla costruzione dei fonemi (➔) e delle forme grammaticali, e al controllo dell’articolazione delle parole; un sistema di mediazione, che agisce come intermediario fra il sistema di implementazione e il sistema concettuale, e che é rappresentato da un gruppo di aree associative ancora non completamente definito.
Linguaggio ed emisferi cerebrali
Nonostante l’emisfero dominante per il linguaggio sia il sinistro, il ruolo dell’emisfero destro non é nullo: infatti, esso sembra cruciale per il controllo della prosodia. Pazienti con lesioni anteriori destre non riescono a intonare le parole e le frasi (deficit nel controllo motorio della prosodia); pazienti con lesioni destre posteriori non riescono a capire il tono emotivo dei discorsi (deficit nella comprensione della prosodia). La dominanza dell’emisfero sinistro nell’elaborazione del linguaggio é evidente già alla nascita, suggerendo che l’emisfero sinistro sia predisposto a diventare l’emisfero linguistico nella maggior parte dei soggetti umani. Tuttavia, lo sviluppo delle reti del linguaggio dipende dall’esperienza. Infatti, la specializzazione dell’emisfero sinistro diventa progressivamente maggiore con l’esperienza linguistica; in soggetti con lesione precoce dell’emisfero sinistro si può osservare uno sviluppo di reti del linguaggio nell’emisfero destro, che consentono un soddisfacente, anche se non ottimale, apprendimento del linguaggio. In soggetti precocemente sordi, il linguaggio dei segni (➔) attiva una rete di aree che comprende le aree del linguaggio di sinistra e una rete di aree a destra: questo suggerisce che le aree del linguaggio di sinistra accettino un ingresso visuomotorio, non solo uditivo, e che reti neurali a destra possano essere reclutate per ruoli linguistici.
Il canto degli uccelli
Gli uccelli canori mostrano la comunicazione vocale di gran lunga più elaborata nel mondo animale, a parte l’uomo. I loro canti sono particolarmente complessi, appresi e trasmessi culturalmente da una generazione all’altra, spesso arricchiti da invenzioni e improvvisazioni. Esperti nel campo del linguaggio umano e nel canto degli uccelli hanno spesso messo in evidenza aspetti paralleli fra i due tipi di linguaggio ed é possibile che tale studio comparato fornisca indizi per comprendere l’apprendimento della comunicazione nell’uomo. Il canto degli uccelli consiste di sequenze ordinate di suoni, il più piccolo elemento dei quali é chiamato nota ed é considerato analogo alle unita fonetiche nell’uomo (➔ fonemi). Le note sono raggruppate in ‘sillabe’, che rappresenterebbero l’unita di base del canto, così come lo sono nel linguaggio umano.
Il ruolo dell’apprendimento nel canto degli uccelli
Come nell’uomo, il canto deve essere appreso. Gli uccelli non imparano a cantare e i bambini a parlare se non vengono esposti ai segnali comunicativi degli adulti della propria specie (➔ imprinting). La maggior parte degli animali non necessita dell’esposizione ai segnali di comunicazione della propria specie per essere in grado di metterli in atto, suggerendo che tali segnali rappresentano comportamenti innati, che si basano su circuiti specie-specifici e geneticamente specificati e che quindi non richiedono l’esperienza per maturare. Linguaggio e canto degli uccelli non sono perciò comportamenti innati: tuttavia, neonati e piccoli uccelli mostrano alla nascita predisposizioni percettive, presumibilmente innate, per segnali vocali specie-specifici. Nei neonati, queste predisposizioni consistono nella capacità di discriminare i fonemi di tutte le lingue, punto di partenza per acquisire la fonologia (➔ fonazione; fonemi) di qualunque lingua; nei piccoli degli uccelli é stata documentata la predisposizione a utilizzare come modello il canto della propria specie e la presenza di caratteristiche specie-specifiche del canto essenziali per innescare l’apprendimento. Che lo sviluppo del repertorio vocale maturo rifletta un apprendimento é stato chiaramente mostrato non solo dalla immaturità del canto di uccelli cresciuti in isolamento, detto canto isolato, ma anche dal fatto che gli uccellini apprendono il canto della specie con le inflessioni del tutore al cui canto sono stati esposti e quelli cresciuti in una zona mostrano un ‘dialetto’ diverso da quelli cresciuti in un’altra zona. Il fatto che gli uccellini cresciuti in isolamento producono un canto immaturo, ma che ha comunque alcune caratteristiche della specie, rinforza l’idea che ci sia una predisposizione innata alla comunicazione vocale tipica della propria specie. L’apprendimento al linguaggio richiede sia la percezione e la memorizzazione dei suoni da riprodurre (apprendimento percettivo), sia l’apprendimento a riprodurre il suono memorizzato attraverso la propria vocalizzazione (apprendimento sensorimotorio). Entrambi i processi dipendono dall’udito. Per gli infanti, il compito consiste nell’immagazzinare in memoria i modelli dei fonemi e la prosodia che caratterizzano la loro lingua madre; gli uccelli immagazzinano le note, le sillabe e le caratteristiche prosodiche che caratterizzano il canto della loro specie. A questo punto, sia gli infanti sia gli uccellini devono esercitarsi a riprodurre i modelli appresi, procedendo per prove ed errori, attraverso il feedback uditivo. Il paragone della ontogenesi dei repertori comunicativi negli infanti e negli uccellini mostra perciò paralleli impressionanti.
Il periodo critico dell’apprendimento
Lo sviluppo del canto richiede l’esperienza dell’ascolto del tutore entro un periodo critico (➔) dello sviluppo precoce. Se l’uccellino viene deprivato dell’ascolto di almeno un tutore conspecifico durante questo periodo, produrrà un canto isolato anche se l’ascolto del tutore si fosse reso poi possibile. Se l’uccellino viene privato dell’udito dopo aver fatto esperienza del canto di un tutore, ma prima che si chiuda il periodo critico per l’apprendimento sensorimotorio, il canto prodotto sarà anormale, in quanto, non potendo udire il proprio canto, l’uccellino non riesce a esercitarsi – per prove ed errori – a riprodurre i prototipi appresi (e nemmeno le caratteristiche specie-specifiche innate), correggendo i gesti articolatori che conducono alla produzione del suono finche il suono emesso non uguagli il suono in memoria. Se un uccellino privato dell’ascolto di un conspecifico viene esposto all’ascolto del canto registrato, imparerà a cantare. Se ciò viene però fatto in corrispondenza della chiusura del periodo critico per l’apprendimento percettivo, non é sufficiente a permettere la formazione di prototipi da riprodurre e quindi l’uccellino produrrà il canto isolato. Se però esso viene messo insieme a un tutore durante lo stesso periodo, si ha lo sviluppo di un canto normale; quindi, la durata del periodo critico dipende dall’esperienza stessa, in questo caso dall’esperienza sociale. Altre evidenze che sottolineano l’importanza delle interazioni sociali nell’apprendimento del canto sono state ottenute in specie di uccelli diversi. Per es., il passero dalla corona bianca fornisce feedback uditivi che influenzano il repertorio dei piccoli e l’apprendimento di un canto alieno richiede l’interazione con il tutore non conspecifico, mentre la sua voce registrata non risulta sufficiente. Nell’uomo, é stato dimostrato che l’esperienza con un parlante in una lingua non materna determina, in bambini di 9÷10 mesi, la ricomparsa della capacità di discriminare i fonemi in questa lingua, perduta con lo sviluppo dei prototipi nella propria lingua; l’esperienza con un video dello stesso parlante risulta invece inefficace. Nel 2003, é stato dimostrato che la interazione sociale con altri individui della propria specie influenza la lallazione non solo in termini di maggiori emissioni vocali ma anche di maggiore maturità dei suoni emessi.