
Mammifero
Mammifero
I Mammiferi (termine composto del latino mamma, "mammella", e fero, "porto"; alla lettera "fornito di mammelle") sono una classe di Vertebrati amnioti omeotermi (latino scientifico Mammalia). Le caratteristiche essenziali dei Mammiferi risiedono nella presenza delle ghiandole mammarie, produttrici di latte per nutrire la prole, e nei dispositivi di viviparità basati sulla placenta, che consente un intimo rapporto fra embrione e utero. Il carattere più appariscente della classe è comunque dato dalla presenza di formazioni cornee, i peli, di diverso tipo e robustezza: i peli, come le penne che caratterizzano la classe degli Uccelli, sono costituiti da cheratina, ma gli zoologi sono incerti sulla loro origine evolutiva. Sia i peli sia le penne servono per mantenere costante la temperatura corporea, nonostante le fluttuazioni di temperatura che si verificano nell'ambiente circostante; di regola i peli costituiscono rivestimenti continui, pur essendo completamente assenti in alcuni gruppi, quali i Cetacei. I Mammiferi sono caratterizzati inoltre da una estrema varietà dimensionale: dal minuscolo toporagno, probabilmente il rappresentante della classe più vicino agli antichi progenitori del Mesozoico, all'enorme balenottera azzurra, il più voluminoso mammifero mai vissuto.
l. Principali caratteristiche anatomiche
Notevoli trasformazioni dello scheletro hanno accompagnato l'evoluzione dei Mammiferi. Rispetto ai Rettili, complessivamente, i Mammiferi hanno sviluppato migliori capacità motorie grazie alla modificazione tanto dell'assetto degli arti, che si sono portati su piani paralleli a quello di simmetria, quanto dei loro rapporti con lo scheletro assile. Il confronto tra gli scheletri dei Rettili terapsidi e quelli dei Mammiferi rivela che esiste una continuità fra i due gruppi, mentre i reperti paleontologici non possono fornire alcuna indicazione per quello che riguarda analogie nei tessuti molli caratteristici dei Mammiferi, quali ghiandole mammarie o placenta. Nella colonna vertebrale le vertebre sono anfipiane, come negli altri Tetrapodi, separate da dischi intervertebrali cartilaginei. Le vertebre toraciche si articolano con le costole superiori che, anteriormente, sono unite direttamente allo sterno. Tutti i Mammiferi, a eccezione dei Cetacei e dei Sirenidi, posseggono due paia di arti e cinti sempre ben sviluppati. È sempre presente uno sterno, spesso diviso in più pezzi. Nel bacino, l'ileo, l'ischio e il pube sono saldati; esiste una sinfisi ventrale dei due pubi e talora anche degli ischi. La conformazione e la robustezza degli arti, delle dita e delle unghie sono molto variabili e dipendono dal sistema di locomozione: i plantigradi poggiano sul suolo l'intero autopodio, i digitigradi le falangi, gli unguligradi le estremità delle dita. Lo scheletro degli arti dei Mammiferi attualmente viventi presenta differenze esteriori notevoli; esiste tuttavia un elevato grado di omologia, in quanto essi derivano da una struttura ancestrale comune. Nel cranio, che si articola alla colonna vertebrale mediante due condili occipitali, il numero delle ossa è molto ridotto rispetto a quello dei Rettili. Due antichi elementi ossei dell'articolazione mascellomandibolare si sono localizzati nell'orecchio medio trasformandosi in martello e incudine, mentre su ogni lato del cranio la cavità posteriore dell'orbita (fossa temporale), che contiene i muscoli masticatori, si è ampliata per contenere un encefalo molto sviluppato; sul tetto del cavo orale si è generalizzata la presenza di una paratia ossea orizzontale (palato secondario) in grado di separare le vie aeree da quelle alimentari. La regione della faccia è variamente sviluppata in relazione al maggior sviluppo assunto dalla scatola cranica per contenere l'encefalo. Le ossa mobili del cranio sono la mandibola, che si articola con l'osso temporale, l'osso ioide, provvisto di corna anteriori e posteriori, e gli ossicini dell'udito, l'incudine e il martello. I movimenti della mandibola dipendono dallo sviluppo dei muscoli della masticazione, principalmente il massetere, il temporale e gli pterigoidei. Il differenziamento dei denti è un'altra importante caratteristica dei Mammiferi. Mentre i denti dei Rettili sono generalmente conici e di grandezza uniforme, quelli dei Mammiferi variano per forma e grandezza, e ciò li rende adatti alla masticazione di differenti tipi di cibo. Per es. l'uomo ha denti adatti sia a tagliare (gli incisivi) sia a frantumare (i molari). Quasi tutti i Mammiferi hanno due successive dentizioni (si dicono perciò difiodonti): a una dentatura da latte succede la dentatura definitiva.
Per quello che riguarda il sistema nervoso, i Mammiferi hanno gli emisferi cerebrali più sviluppati degli altri Vertebrati di taglia equivalente e sembrano quelli più capaci di apprendimento. Inoltre, i sensi sono molto sviluppati e la durata relativamente lunga delle cure parentali consente alla prole di apprendere importanti informazioni per la sopravvivenza mediante l'osservazione dei comportamenti dei genitori. Il cervelletto è esteso e costituito dal verme e dagli emisferi. Sono presenti il ponte di Varolio che circonda il midollo allungato, la commessura del corpo calloso che unisce i due emisferi, i corpi quadrigemini nel mesencefalo e 12 paia di nervi cranici. Gli occhi sono protetti da palpebre mobili, con o senza membrana nittitante; nell'orecchio interno è presente la chiocciola; l'orecchio esterno è provvisto di un padiglione sostenuto da cartilagine, il quale funziona come raccoglitore di suoni. Nel sistema circolatorio vi è una netta separazione della circolazione polmonare dalla circolazione sistemica e il cuore è tipicamente costituito da quattro compartimenti che impediscono il mescolamento del sangue ricco di ossigeno con quello povero di ossigeno (v. cuore). Le emazie, generalmente discoidali, sono prive di nucleo. L'attivo metabolismo dei Mammiferi è supportato da un sistema respiratorio efficiente, coadiuvato da un caratteristico setto completo muscolotendineo, il diaframma, che separa nel celoma la cavità toracica (pleurica) da quella addominale (peritoneale) e aiuta la ventilazione dei polmoni. I bronchi si ramificano senza anastomizzarsi nei polmoni e questi, rivestiti dalle pleure, sono liberi di essere compressi ed espansi nelle cavità pleuriche nel corso dei movimenti di espirazione e inspirazione. La laringe è provvista di corde vocali per l'emissione dei suoni. I Mammiferi hanno reni compatti a superficie lobata o liscia, ovoidali con capsule surrenali sovrapposte a essi; gli ureteri a eccezione di quelli dei Monotremi, che hanno una cloaca, sboccano in una vescica urinaria comunicante con l'esterno mediante l'uretra, in cui nel maschio si aprono anche i deferenti. I testicoli sono protetti da una borsa cutanea, lo scroto e, poiché la fecondazione è interna, i maschi posseggono sempre un organo erettile. Tutti i Mammiferi tranne i Monotremi sono vivipari. Alla nascita gli individui sono completi e assumono il nutrimento dalla madre. L'apparato riproduttore femminile è costituito dalle tube, dove viene fecondato l'uovo, dall'utero (doppio, bicorne, bipartito, semplice) in cui si sviluppa l'embrione, e dalla vagina che si apre, insieme all'uretra, nel seno urogenitale. Nei Mammiferi placentati il rivestimento dell'utero e le membrane extraembrionali che derivano dall'embrione formano la placenta, attraverso la quale il nutrimento passa nel circolo embrionale.
Omeotermia
Presumibilmente l'omeotermia costituisce uno dei più importanti adattamenti evolutivi sia negli Uccelli sia nei Mammiferi. Tutti gli omeotermi sono endotermi: la fonte di calore è costituita dall'ossidazione del glucosio e di altre molecole energetiche, processi che si verificano all'interno delle cellule. Nei Mammiferi i neuroni che presiedono alla termoregolazione sono concentrati nella regione ipotalamica dell'encefalo dove sono presenti due aree regolatrici: un centro conservatore del calore, che controlla la vasocostrizione del letto vascolare superficiale, il raddrizzamento dei peli, il controllo del riflesso del brivido e la termogenesi ormonale; un centro dissipatore del calore che controlla la vasodilatazione e la sudorazione (o la respirazione ansimante). Le terminazioni nervose dei neuroni che funzionano da termorecettori sono situate nella cute. Vi sono specifici corpuscoli devoluti alla termocezione: per es. il corpuscolo di Ruffini è specializzato per trasdurre l'energia termica e potenzia l'emissione di segnali verso il sistema nervoso centrale allorché la temperatura aumenta; i bulbi di Krause sono invece recettori del freddo, nel senso che la frequenza del segnale da loro trasmesso aumenta con la diminuzione della temperatura. I recettori del caldo hanno un'azione eccitatoria sul centro ipotalamico che promuove la dissipazione del calore e una inibitoria su quello che tende a mantenerlo; i recettori del freddo esercitano invece un'azione esattamente inversa sui due centri nervosi. La temperatura corporea è dunque controllata da meccanismi di feedback, nell'ambito dei quali l'ipotalamo funziona come un termostato, innescando nell'organismo sia risposte finalizzate all'aumento della produzione del calore, sia risposte di segno contrario. Quando la temperatura corporea di un mammifero si innalza sopra il valore indicato dal termostato, i vasi sanguigni prossimi alla superficie si dilatano aumentando il flusso del sangue alla pelle. Se l'aria è più fredda della superficie del corpo, il calore si trasmette dalla superficie del corpo direttamente all'esterno. Gli animali che vivono in climi caldi hanno aree superficiali generalmente più grandi di quelli che vivono al freddo. Il calore viene disperso dalla superficie per mezzo dell'evaporazione della saliva, della traspirazione e utilizzando vari adattamenti di tipo comportamentale. Gli adattamenti al caldo torrido negli animali grossi come il cammello derivano dalla buona capacità di tollerare sia le variazioni della temperatura sia quelle del contenuto idrico totale: un cammello può sopportare una perdita di acqua fino al 25% del suo peso corporeo, mentre un uomo, quando raggiunge una carenza di acqua pari al 12% del suo peso, diventa incapace di deglutire e ha bisogno di cure per sopravvivere. Per l'adattamento al caldo degli animali di piccola taglia si rendono necessarie sia strategie per la conservazione dell'acqua, sia particolari adattamenti comportamentali, come, per es., la vita notturna. Quando, a causa del freddo, la temperatura corporea scende, i vasi sanguigni prossimi alla superficie cutanea subiscono una costrizione riducendo la perdita di calore. Aumenta inoltre la produzione di energia a causa dell'attività muscolare e della stimolazione nervosa e ormonale. Gli adattamenti al freddo intenso sono essenzialmente basati su sistemi di isolamento termico che vengono attuati per mezzo di peli oppure di uno spesso strato sottocutaneo di grasso; per es. alcuni grossi Mammiferi artici, come le foche o gli orsi polari, sono talmente ben isolati che, quando sono sulla terraferma, per disperdere il calore devono bagnare la pelliccia o dormire, cosa che, in alcune specie di foche, riduce la quantità di calore del 25%. Inoltre, al diminuire della temperatura i peli si drizzano per azione dei muscoli erettori, trattenendo l'aria che isola la superficie; nell'uomo il 'ricordo' evolutivo di questo fenomeno è rappresentato dalla cosiddetta pelle d'oca (v. brivido). Numerosi animali conservano invece il calore facendo scendere la temperatura delle loro estremità, le quali sono adattate a possedere una temperatura interna diversa da quella del resto del corpo. In molti animali artici l'abbassamento della temperatura interna delle estremità è mantenuto in termini accettabili da un sistema di scambio di calore controcorrente: per es., nei delfini le arterie che conducono verso le estremità delle pinne il sangue con una temperatura relativamente elevata sono in stretto contatto con le vene che riconducono il sangue caratterizzato da una temperatura inferiore verso il tronco, in modo che il sangue raffreddato, di ritorno dalle vene, assuma calore dal sangue proveniente attraverso le arterie.
Sottoclassi dei Mammiferi
Vi sono oggi circa 4000 specie di Mammiferi, appartenenti a tre sottoclassi: Prototeri, Metateri, Euteri. I Prototeri comprendono solo l'ordine dei Monotremi, limitato geograficamente all'Australia e alla Nuova Guinea, con due famiglie e tre specie, riferibili a due forme fondamentali, l'ornitorinco e l'echidna. Sono i soli Mammiferi che depongono le uova, simili a quelle dei Rettili e contenenti abbastanza tuorlo per nutrire l'embrione che si sta sviluppando. La femmina dei Monotremi presenta speciali ghiandole sul ventre che producono latte; dopo la nascita il piccolo succhia il latte dal pelo della madre, in quanto essa non ha capezzoli. In questo ordine la commistione fra caratteri dei Rettili e quelli dei Mammiferi suggerisce che i Monotremi derivino da un ramo molto antico nella genealogia dei Mammiferi. I Metateri comprendono l'ordine dei Marsupiali, un gruppo rappresentato da circa 240 specie. Essi completano il loro sviluppo in una tasca, il marsupio. L'uovo contiene una piccola quantità di tuorlo che serve per il nutrimento dell'embrione all'inizio del suo sviluppo nel tratto riproduttivo della madre. I Marsupiali nascono in un momento molto precoce del loro sviluppo. Il canguro rosso, per es., nasce dopo solo 33 giorni dalla fecondazione, e ha la grandezza di un'ape; gli arti anteriori sono delle semplici gemme, mentre quelli posteriori sono abbastanza forti da permettergli di arrivare, in pochi minuti, dal tratto riproduttivo al marsupio della madre, nel quale il piccolo fissa la sua bocca a un capezzolo completando lo sviluppo. Il parto prematuro rende la femmina nuovamente disponibile all'accoppiamento, ma lo sviluppo dello zigote rimane bloccato sino al momento nel quale il marsupio è di nuovo libero per ospitare il nascituro. Queste peculiarità riproduttive sono verosimilmente correlate con l'ambiente che presenta fonti alimentari precarie, rendendo necessarie lunghe migrazioni in cerca di cibo, con rischi per la prole. I Metateri sono prevalentemente concentrati in Australia, dove i Marsupiali si sono sviluppati e hanno occupato nicchie ecologiche altrove proprie degli Euteri, seguendo il loro stesso ventaglio adattivo, con l'esclusione degli adattamenti all'ambiente acquatico e a quelli correlati con il volo. L'opossum dell'America Settentrionale e Meridionale è attualmente l'unico marsupiale che vive fuori dell'Australia, ma nel Terziario l'America Meridionale (Patagonia) ha avuto una fauna marsupiale differenziata. Secondo alcune recenti evidenze fossili probabilmente i Marsupiali sono originati in quella che è attualmente l'America Settentrionale e si sono spostati al Sud quando le terre erano ancora riunite: questa ipotesi è, tuttavia, ancora oggetto di studio. La maggior parte dei Mammiferi viventi corrisponde agli Euteri, definiti anche Placentati, con un termine che non tiene conto che anche alcuni Marsupiali hanno una sia pur primitiva placenta. I piccoli degli Euteri nascono inetti ma non prematuri, per cui non è necessario il marsupio. Le specie attuali di Euteri compongono 16 ordini, il più numeroso dei quali è quello dei Roditori con circa 1700 specie; seguono i Chirotteri (pipistrelli) con 850 specie e gli Insettivori (talpe e toporagni) con 400 specie.
Evoluzione
Nell'albero filogenetico delle varie specie, prima i Mammiferi e in seguito gli Uccelli derivano dai Rettili. I Mammiferi comparvero nel periodo Triassico dell'Era Mesozoica (i fossili più antichi risalgono a 220 milioni di anni fa) e i loro progenitori sono i Rettili terapsidi, che scomparvero durante il regno dei dinosauri. L'Era Mesozoica, da 65 a 240 milioni di anni fa, fu caratterizzata da una situazione climatica stabile, dall'aumento del livello del mare, da un'intensa attività orogenetica che conferì alla Terra gran parte delle caratteristiche attuali e, infine, dalla presenza dei Rettili che dominarono tutti gli habitat terrestri, ritornando anche a occuparne alcuni acquatici. I Mammiferi, rimasti in ombra durante il trionfo dei Rettili, erano, per la maggior parte, di piccolissima taglia, si nutrivano presumibilmente di insetti ma possedevano un certo numero di innovazioni importanti: mantenevano una temperatura corporea costante ed elevata, erano coperti di pelo che fungeva da isolante per le temperature più basse e i loro piccoli venivano partoriti anziché nascere dalle uova. L'omeotermia, pur avendo in termini di richieste energetiche un costo elevato (il tasso metabolico di un Mammifero è circa 10 volte quello di un Rettile di dimensioni simili che possieda la medesima temperatura), offriva presumibilmente un alto numero di vantaggi: infatti la capacità di mantenere un alto grado di efficienza anche a temperature molto basse permetteva ai primi Mammiferi di uscire in cerca di cibo e di un partner durante le ore notturne, cioè quando gli animali dominanti, i Rettili, erano probabilmente inattivi; questo rese possibile la conquista di nuovi ambienti. Il regno dei grandi Rettili, durato 125 milioni di anni, si estinse improvvisamente, alla fine del Mesozoico, per ragioni che costituiscono un mistero irrisolto. Il cataclisma risparmiò in parte i piccoli Mammiferi. I geologi, sulla base degli enormi cambiamenti che si possono osservare nei fossili, indicano la fine del periodo Cretaceo come il punto di confine fra il Mesozoico e la successiva Era Cenozoica. In questo periodo la vita sulla terra e nel mare subì infatti grandi trasformazioni e si verificarono forti variazioni ambientali. Durante il Cenozoico, iniziato 65 milioni di anni fa, in tali condizioni di forti variazioni ambientali, i Mammiferi si trovarono al centro di imponenti fenomeni di 'radiazione adattiva', un'evoluzione divergente che si verifica proprio quando una specie colonizza aree a forte diversificazione ambientale e si forma rapidamente una serie di specie affini, adatte ad habitat differenti. In questa era, il successo evolutivo degli Uccelli, degli Insetti e dei Mammiferi fu accompagnato da un'altra grande innovazione biologica, il fiore. Durante il Cretaceo le piante da fiore, le Angiosperme, diventarono infatti più abbondanti e rimpiazzarono le Gimnosperme. Secondo alcuni scienziati, questo cambiamento nella vegetazione contribuì al declino dei dinosauri. Durante i primi 10-15 milioni di anni comparvero gli esponenti di quasi tutti gli ordini di Mammiferi moderni. Le relazioni filogenetiche fra Marsupiali e Mammiferi placentati sono talvolta oscure, ma si pensa che essi siano maggiormente correlati fra di loro rispetto ai Monotremi. Reperti fossili indicano che Placentati e Marsupiali si sono separati da un progenitore comune circa 80-100 milioni di anni fa. Una radiazione adattiva, che è avvenuta durante il Tardo Cretaceo e l'inizio del Terziario, ha prodotto l'ordine dei Mammiferi placentati che conosciamo oggi. Nella prima metà del Cenozoico i Primati sono rappresentati da piccoli Mammiferi placentati arboricoli; la loro struttura dentale suggerisce che siano derivati dagli Insettivori del Tardo Cretaceo. Una specie fossile, chiamata Purgatorius unio, riportata alla luce nel Montana in strati al limite tra il Cretaceo e l'Era Terziaria, viene considerata da molti il primate più antico sinora noto. I Primati vengono comunemente distinti in due sottordini: le Proscimmie (le forme ancestrali) e gli Antropoidei (le piccole scimmie, le grandi scimmie e l'uomo). Le Proscimmie ricordano gli ancestrali Primati arboricoli: i lemuri del Madagascar, i loridi e i tarsioidei dell'Africa tropicale e dell'Asia meridionale sono esempi di Proscimmie. I primi Antropoidei che appaiono tra i reperti fossili sono Primati scimmieschi, evoluti dal gruppo ancestrale delle Proscimmie circa 40 milioni di anni fa in Africa e forse ancora prima in Asia. Dato che l'Africa e l'America Meridionale si erano già separate, non si sa se i progenitori delle piccole scimmie abbiano raggiunto l'America Meridionale trasportati da tronchi d'albero o da detriti provenienti dall'Africa, oppure migrando dall'America Settentrionale verso la parte meridionale del continente. È comunque certo che le piccole scimmie del Nuovo Mondo, le platirrine, e quelle del Vecchio Mondo, le catarrine, si sono differenziate intraprendendo percorsi evolutivi separati da molti milioni di anni. Le grandi scimmie, che vivono attualmente nelle regioni tropicali del Vecchio Mondo sono divise in quattro generi: Hylobates (gibboni), Pongo (orangutan), Gorilla e Pan (quest'ultimo genere comprende gli scimpanzé). Testimonianze fossili dei progenitori diretti delle scimmie antropomorfe odierne (gibboni, orangutan, scimpanzé e gorilla) risalgono a circa 20 milioni di anni. Il Pleistocene, cominciato 2,5 milioni di anni fa, fu caratterizzato da glaciazioni intermittenti che determinarono estinzioni vastissime e migrazioni, le quali modificarono radicalmente la distribuzione degli esseri viventi sulla terra. La vegetazione tropicale, una volta estesa su gran parte del pianeta, si ritirò negli habitat caldi e umidi equatoriali. I giganteschi mammut e molti altri grandi Mammiferi scomparvero con l'ultima glaciazione che ebbe termine circa 10.000 anni fa. Si estesero le zone semiaride e aride, e la flora e la fauna sopravvissute cominciarono il periodo più recente di radiazione adattiva.
Studi di filogenesi dei Mammiferi placentati
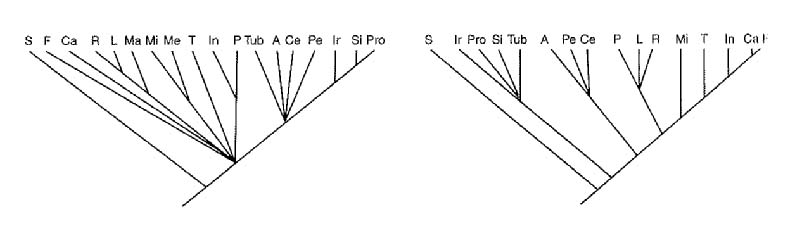

Le relazioni evolutive fra la maggior parte dei Mammiferi placentati non sono ancora ben stabilite, anche se tutte le forme viventi sono state ampiamente studiate e le forme ancestrali sono ben documentate da numerosi reperti. I reperti fossili suggeriscono che tale gruppo sia stato interessato da una radiazione adattiva esplosiva circa 60 milioni di anni fa, allorché comparvero le linee corrispondenti sia agli ordini attuali sia ad altri gruppi estinti. La filogenesi dei Mammiferi placentati tuttavia non risulta facilmente ricostruibile sulla base della documentazione paleontologica proprio a causa della sua notevole accelerazione. Con lo sviluppo delle tecnologie molecolari, dagli anni Settanta del 20° secolo è iniziata una raccolta di dati di natura biochimica riguardanti prevalentemente le molecole proteiche strategiche per l'evoluzione. Si sono cioè misurate le differenze fra le varie specie paragonando la sequenza degli aminoacidi delle proteine. La quantità degli aminoacidi diversi fornisce la misura dei tempi dell'evoluzione indipendente, a partire da un progenitore comune. Nella fig. 8 sono riportati differenti diagrammi relativi alla sistematica dei Mammiferi placentati, ricavati analizzando caratteri morfologici e biochimici e facendo uso di metodi sistematici diversi. Come si può notare, i diversi metodi portano a risultati piuttosto discordanti. È da tenere presente che la sistematica tradizionale presenta le seguenti caratteristiche: considera ogni tipo di carattere, dà grande spazio alla velocità dell'evoluzione e, in molti casi, tiene conto dei dati ecologici. Lo schema di classificazione chiamato cladistico dà, invece, maggior risalto alle affinità filogenetiche, in quanto considera solo i caratteri derivati condivisi, non tiene conto della velocità dell'evoluzione e impiega raramente i dati ecologici. Entrambe le classificazioni danno grande rilievo alle documentazioni paleontologiche. Lo studio delle relazioni evolutive fra le specie fa ricorso anche all'analisi diretta dei geni. In particolare, molto usata è stata una tecnica, definita ibridazione molecolare, che ha permesso di confrontare le sequenze di basi di numerosi geni di specie diverse e di stabilirne le omologie. Il suo presupposto è costituito dalla possibilità di separare i due filamenti di DNA tramite il calore e di riassociarli nuovamente con il raffreddamento. Trattando in questo modo il DNA di due diverse specie si possono ottenere doppie eliche interspecifiche o ibride. A causa della diversità della sequenza delle basi, i due filamenti ibridi risultano legati in modo relativamente debole e per separarli è necessario meno calore che per il DNA omogeneo. La correlazione fra la termolabilità di un DNA ibrido e lo sbilanciamento delle basi che si fronteggiano è tanto stretta che è possibile valutare la percentuale delle basi non bilanciate misurando la temperatura che consente un determinato grado di dissociazione dei due filamenti. Il limite di questo metodo è tuttavia dovuto al fatto che può essere applicato solo a specie relativamente affini, poiché i filamenti di un'elica ibrida che presentino una diversità fra le basi superiore al 20% non possono appaiarsi stabilmente. Con queste metodiche si sono potuti risolvere alcuni problemi evolutivi dibattuti per molto tempo: per es. è stato accertato che, sebbene il panda gigante o maggiore e il panda rosso o minore, entrambi asiatici, abbiano in comune un certo numero di caratteristiche strutturali e di comportamento, essi non sono strettamente affini. Esperimenti di ibridazione del DNA uniti ad analisi di proteine enzimatiche, comparazioni immunologiche e studi della morfologia dei cromosomi hanno chiaramente indicato che il panda rosso è più affine al procione americano che al panda maggiore: diverge dai procioni dopo che questi si sono separati dagli orsi mentre il panda gigante diverge dagli orsi molto recentemente e si può considerare sostanzialmente un orso asiatico con alcune differenze determinate prevalentemente dalla dieta a base di bambù. La tecnica degli acidi nucleici ibridi ha riservato sorprese ancor più interessanti; per es. si è osservata una piccolissima differenza di sequenze di basi fra uomo e scimpanzé, rispetto alle differenze con altre scimmie. Un altro parametro usato per stabilire i rapporti filogenetici consiste nel confrontare i cromosomi di diverse specie. I riarrangiamenti cromosomici infatti hanno avuto conseguenze evolutive importanti. Quando vengono confrontati i cromosomi di organismi evolutivamente lontani si osservano riarrangiamenti radicali, mentre se si osservano i cromosomi di specie molto vicine si può notare invece una somiglianza notevole. La fig. 10 mostra lo schema dei cromosomi di uomo, scimpanzé, gorilla e orango con il caratteristico bandeggio che si ottiene dopo alcune particolari colorazioni differenziali. Sia i risultati ottenuti confrontando le sequenze del DNA sia quelli ottenuti confrontando le bande dei cromosomi suffragano complessivamente l'ipotesi che scimpanzé e uomo siano derivati da un progenitore comune, in un'epoca più recente rispetto a quella che si era comunemente creduta basandosi su altri parametri. Si può concludere tuttavia che la filogenesi dei Mammiferi placentati è ancora molto incompleta sia perché i dati utilizzati sono discontinui e filogeneticamente eterogenei sia perché i programmi che consentono di elaborare al computer schemi di filogenesi tengono poco conto dei fenomeni piuttosto frequenti di evoluzione parallela. L'evoluzione parallela determina infatti l'affermazione di un medesimo carattere in diverse linee filogenetiche. Sostanzialmente, dopo più di 20 anni di studi intensi, molti quesiti restano sostanzialmente aperti. In realtà, non esiste nessun mezzo che, da solo, possa dare informazioni complete sui tempi dell'evoluzione: anche se la documentazione fossile manterrà una funzione insostituibile, è possibile prevedere che, in futuro, i metodi molecolari accresceranno il loro ruolo nel definire i rapporti filogenetici sia tra i vari gruppi animali sia tra i Mammiferi e che la sistematica dovrà sempre più allargare l'ambito delle discipline che le sono di supporto.
Bibliografia
g. brum, l. mckane, g. karp, Biology. Exploring life, New York, Wiley, 19942 (trad. it. Biologia. La continuità della vita, Bologna, Zanichelli, 1998).
n.a. campbell, Biology, Menlo Park (CA), Benjamin/Cummings, 1996 (trad. it. Bologna, Zanichelli, 1996).
a.j.f. griffiths, d.t. suzuki, r.c. lewontin, An introduction to genetic analysis, New York, Freeman, 19966 (trad. it. Genetica. Principi di analisi formale, Bologna, Zanichelli, 1996).
w.k. purves, g.h. orians, h. craig heller, Life. The science of biology, Sunderland (MA), Sinauer Associates, 19954 (trad. it. Corso di biologia, Bologna, Zanichelli, 1995).