
Memoria: studi sperimentali
Memoria: studi sperimentali
di Daniele Bovet
Memoria: studi sperimentali
sommario: 1. Definizione. 2. Cenni storici. 3. Classificazione e filogenesi dei vari tipi di memoria. 4. Parametri di tempo della memoria: a) introduzione; b) memoria senso-motoria a breve e a lungo termine; c) approccio sperimentale del processo di consolidamento; d) determinazione genetica dei tipi di memoria. 5. Memoria verbale nella sperimentazione umana: a) metodi di studio; b) pluralità dei tipi di memoria nell'uomo. 6. Ipotesi sulla natura dell'engramma mnesico: a) varietà nell'approccio sperimentale; b) correlati elettrofisiologici; c) modifiche strutturali a livello di neuroni sinaptici; d) ipotesi sulla funzione degli acidi nucleici; e) dati farmacologici: effetti dei farmaci psicotropi. 7. Rapporti fra comportamento acquisito e strutture genetiche nell'animale: a) il fenomeno dell'imprinting. 8. La struttura della memoria nell'uomo: a) patologia della memoria; b) ipermnesia; c) contenuto della memoria; d) il cervello come elaboratore. □ Bibliografia.
1. Definizione
Per il biologo la memoria rappresenta ‛la conservazione e l'accumulo del passato nel presente'. Distingueremo dunque: 1) la memoria propriamente detta, ritenzione o conservazione, legata al processo di apprendimento di ciascun essere sia che si tratti di un semplice condizionamento sia del richiamo di situazioni complesse, sempre tuttavia riferite a risposte acquisite e alla possibilità di riattivarle; 2) la memoria genetica o memoria innata, corrispondente al comportamento istintivo, morfogeneticamente determinata ed estranea a qualsiasi esperienza individuale; 3) fra queste due memorie che rappresentano i termini estremi, ne esiste una terza, detta memoria epigenetica, che si riferisce al processo dell'imprinting e cioè a un tipo particolare di apprendimento strettamente legato alle tendenze innate della specie.
Secondo C. Florès (v., 1964) il termine di memoria permette di raggruppare un insieme di attività che vengono a integrare dei processi sia biofisiologici sia psicologici, i quali possono prodursi nel momento attuale soltanto in quanto alcuni avvenimenti anteriori, vicini o lontani nel tempo, hanno modificato in modo persistente lo stato dell'organismo. Le tre fasi del processo consistono nell'acquisizione, nella ritenzione e nella riattivazione o attualizzazione. In senso più restrittivo la memoria rappresenta, in psicologia umana, ‟una funzione psichica consistente nella riproduzione di uno stato di coscienza passata caratterizzato dal fatto che esso viene riconosciuto dal soggetto" (A. Lalande, Vocabulaire technique et critique de la philosophie, Paris 1968, p. 606).
In contrasto con la memoria logica, che implica un ordine razionale e cioè una rappresentazione collettiva e stabile, esiste una memoria ‛autistica' che compare nell'uomo durante il sogno, durante il sonno paradosso (REM) o ancora nel corso di un delirio. Quest'ultima sembra attingere alla memoria sensomotoria e a quella sociale e appare strettamente legata alla memoria affettiva.
Nel campo della bionica dell'informatica, il termine di memoria (M) sta a indicare quella parte dell'elaboratore che ha la funzione di immagazzinare l'informazione mettendola in relazione da una parte con le unità di ingresso (I) e di uscita (U) e dall'altra con l'unità logico-aritmetica (P) responsabile del trattamento e della messa in codice.
2. Cenni storici
L'approccio sperimentale del problema della memoria il solo che debba essere qui trattato - ha seguito due vie parallele. La prima, basata sull'osservazione del comportamento, si ricollega al problema dell'apprendimento. La seconda, relativa ai meccanismi dell'accumulo dell'informazione, riguarda invece la natura della traccia mnemonica.
La ricerca sui processi mnemonici è basata su procedimenti sperimentali che ne hanno permesso l'analisi quantitativa nell'animale e nell'uomo. Tre tappe a questo riguardo meritano di essere ricordate: 1) nel 1885 la pubblicazione di H. Ebbinghaus, Über Gedächtnis, dedicata alla memoria verbale nell'uomo; 2) nel 1898 la monografia di E. L. Thorndike, in realtà la sua tesi di dottorato all'Università di Colombia: Animal intelligence: an experimental study of the associative processes in animal; 3) infine, nel 1903, l'introduzione da parte di I. P. Pavlov del concetto di riflesso condizionato ripreso nel 1909 con la conferenza Le scienze naturali e il cervello, e la descrizione delle leggi che presiedono alla sua formazione (la psicologia sperimentale e la psicofisiologia degli animali).
Come spesso accade nella storia della scienza, questi studi si sviluppano inizialmente in modo indipendente e l'analisi delle più complesse forme di memoria verbale precede quella del condizionamento e dei fenomeni di semplici associazioni sensomotorie.
Gli studi di Pavlov aprirono la strada, nell'Unione Sovietica, a quelli di V. Bechterev sulle reazioni di fuga e di evitazione e alle teorie delle rappresentazioni psicomorfologiche di Anokhin (1935), di E. A. Asratyan (1953) e di A. R. Luna (1957). Nella direzione indicata da Thorndike e da Pavlov, il behaviorismo elaborato da J. B. Watson (1914) negli Stati Uniti permetterà, con la sperimentazione animale, una precisa analisi dei processi di ritenzione nell'apprendimento strumentale (E.Ch. Tolman, 1925; K. S. Lashley, 1932; B. F. Skinner, 1938; C. L. Hull, 1943).
Il capitolo aperto da Ebbinghaus con le ricerche sulla memoria verbale fu ampliato da G. E. Müller e F. Schumann (1889), A. Binet e V. Henri (1894), H. Munsterberg (1894), A. Jost (1897), Müller e A. Pilzecker (1900). Questi autori hanno esteso la ricerca a campi molto vicini ed egualmente importanti quali, per non citare che i principali, i processi dell'abitudine, dell'attenzione, dell'interferenza e del consolidamento.
In quanto ‛persistenza del passato' si ritiene oggi che la memoria abbia un substrato organico; si parla infatti di una modificazione strutturale della corteccia cerebrale alla quale viene dato il nome di traccia o engramma mnesico.
Dal punto di vista neurofisiologico il problema ha sollevato molte discussioni, ricollegandosi con quello delle ‛localizzazioni cerebrali'. I dati realmente positivi in grado di dimostrare come l'apprendimento modifichi la struttura del cervello non sono a tutt'oggi molto numerosi e provengono da campi assai diversi. Ci sono voluti quasi due secoli perché il problema posto dalle prime ricerche di M. V. C. Malacarne (1780) nel cane venisse affrontato con rinnovato impegno dagli studiosi contemporanei. Vanno qui menzionati i primi significativi contributi di Lashley e più recentemente di R. W. Gerard (1950), A. Fessard (1958), F. Morrell (1961), F. O. Schmitt (v., 1962) ed E. R. John (v., 1967).
3. Classificazione e filogenesi dei vari tipi di memoria
L'estensione stessa del concetto di memoria ha incitato gli autori a distinguere fra diversi tipi di processi mnemonici. Sono stati considerati vari criteri di ordine anatomico (rapporto con le strutture cellulari e nervose interessate), filogenetico (posto nella scala zoologica della specie considerata) e ontogenico (relazione con lo stadio di sviluppo del soggetto). Si distinguono oggi: a) il processo di assuefazione; b) la memoria di associazione pura, corrispondente, per esempio, ai processi di condizionamento classico; c) la memoria sensomotoria o di riconoscimento, che corrisponde ai processi di apprendimento discriminativo ‛per prove ed errori' del comportamento detto strumentale o operativo; d) la memoria di ricostruzione; e) la memoria di evocazione. È interessante notare come le varie forme di memoria costituiscano, in rapporto con l'evoluzione strutturale delle specie, una successione la cui complessità cresce progressivamente nei diversi rami dell'albero genealogico del regno animale.
Da un punto di vista filogenetico il processo di assuefazione, e cioè il decrescere delle risposte a uno stimolo specifico in seguito alla ripetizione dello stimolo stesso, viene considerato come un processo mnemonico elementare in quanto rappresenta la forma più semplice di un comportamento modificato dall'esperienza. Riveste un interesse particolare il fatto che l'assuefazione non dipende che in parte dalle caratteristiche proprie alla specie e si rivela comune a qualsiasi massa protoplasmatica isolata. Oltre che nei Protozoi Rizopodi (ameba) e Ciliati (paramecio) e nei plasmodi dei Mixomiceti, la si ritrova nelle reazioni dei leucociti circolanti dei Vertebrati superiori e perfino nelle cellule isolate delle colture di tessuti embrionali provenienti da specie assai diverse. L'assuefazione rappresenta anche il tipo di comportamento adattativo prevalente nei Metazoi dotati di un sistema nervoso elementare, composto di cellule isolate (Poriferi), o a rete diffusa (Celenterati).
In modo molto schematico può essere suggerita l'esistenza di una correlazione fra la costituzione di un sistema nervoso cordonale e la comparsa delle prime manifestazioni di una memoria associativa nei Platelminti. Un sistema nervoso ganglionare appare nei Nematelminti, negli Anellidi, nei Molluschi, negli Artropodi e negli Echinodermi; il comportamento strumentale raggiunge un grado di complessità particolarmente elevato nei Cefalopodi, fra i Molluschi, e negli Insetti fra gli Artropodi. Per esempio, il sistema di comunicazione descritto in alcuni Imenotteri potrebbe corrispondere a una forma primitiva di memoria di ricostituzione.
Nel phylum dei Cordati, al quale appartengono le diverse classi di Vertebrati, lo sviluppo dei processi mnemonici segue l'evoluzione del sistema nervoso tubolare.
Secondo P. D. McLean (1970) e J. Altman (1966) i rapporti fra strutture e funzioni nei Vertebrati, oltre quelli evolutivi, rispecchiano tre diversi livelli comportamentali, cioè riflesso, istintivo e adattativo, corrispondenti alle diverse strutture, spinomidollari, paleoencefaliche e neoencefaliche (v. tab. I). Al primo livello corrisponde la memoria genetica, al secondo la forma elementare di memoria di associazione, al terzo infine la memoria sensomotoria di ricostruzione e di evocazione.
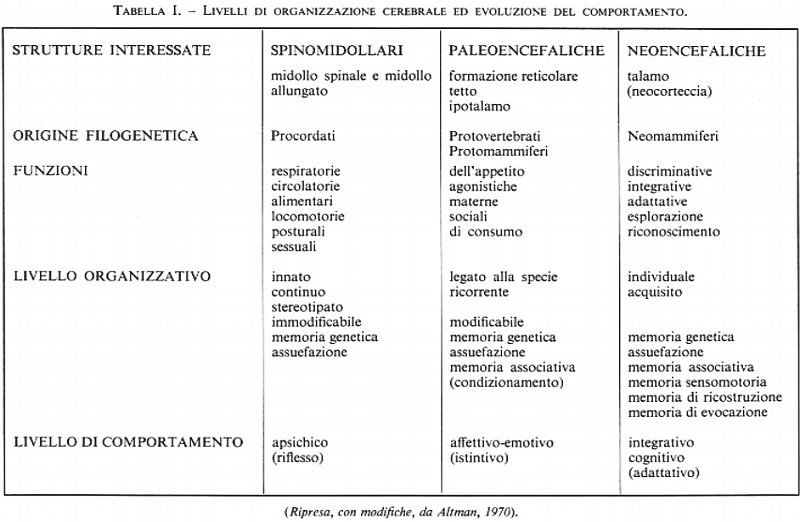
Nella specie umana, il campo dell'apprendimento verbale si ricollega solo nella sua fase iniziale a strutture reazionali di natura motoria e sensomotoria e al processo d'imitazione. Nel loro sviluppo, con l'apprendimento del linguaggio, le relazioni referenziali inizialmente associative acquistano di fatto un carattere simbolico e rappresentativo in rapporto con le attitudini specifiche che caratterizzano l'intelligenza umana.
4. Parametri di tempo della memoria
a) Introduzione
Da un punto di vista concettuale, apprendimento e memoria costituiscono i due aspetti di un'unica funzione. Come un ricordo si collega obbligatoriamente a una precedente esperienza, così non sarebbe possibile concepire un'acquisizione che non si riallacci a un processo di ritenzione.
È tuttavia possibile distinguere le due operazioni da un punto di vista metodologico (A. W. Melton, 1963). Consideriamo un ‛apprendimento' come una serie di reazioni consecutive alla modificazione delle prestazioni quando la variabile è rappresentata dal numero (n) delle prove fornite dal soggetto successivamente; consideriamo invece ‛memoria' le situazioni in cui il parametro sul quale si ferma l'attenzione dello sperimentatore è rappresentato dalla durata dell'intervallo trascorso fra le due sequenze n e n+1 successive.
Una prova di memoria comporta quindi al minimo due test consecutivi: il primo, che corrisponde generalmente all'ultimo test di apprendimento, mostra la somma di quanto è stato appreso in precedenza; il secondo, che corrisponde al test di ritenzione, fornisce una misura delle proporzioni relative fra materiale ritenuto e materiale dimenticato nell'intervallo fra le due prove.
La registrazione corrisponde al processo di entrata e di immagazzinamento dell'informazione; la ritenzione, la conservazione e il recupero corrispondono invece alla disponibilità del materiale immagazzinato.
Lo studio dei parametri di tempo rappresenta un primo ed efficace metodo nelle ricerche sulla memoria. La definizione corrente di memoria, che si riferisce a una modificazione più o meno permanente del comportamento, appare quindi insufficiente. Verranno perciò distinti, in base alla rapidità con la quale si stabiliscono e alla resistenza all'oblio, due tipi di memoria, la memoria a breve termine e la memoria a lungo termine (Gerard, 1949; v. Hebb, 1949).
Il particolare interesse di questa ipotesi - oggi general- mente accettata - risiede nel fatto che essa presenta il vantaggio di potersi applicare a numerosi casi di ritenzione sia nell'uomo che nell'animale.
b) Memoria sensomotoria a breve e a lungo termine
Memoria a lungo termine. - Considerate nel quadro del concetto delle due memorie, la maggior parte delle osservazioni raccolte sulla memoria sensomotoria nell'animale presenta le caratteristiche della memoria a lungo termine; esse corrispondono nell'uomo all'esperienza comune che riguarda per esempio l'apprendimento di uno sport o di un gioco.
Per i processi di ritenzione di comportamenti relativamente elementari, gli esperimenti condotti sull'animale hanno messo in evidenza il carattere progressivo, cumulativo e durevole della conservazione dell'informazione che non presenta, nella maggioranza dei casi, che scarsi segni di degradazione. Per i riflessi condizionati e del comportamento strumentale, i concetti d'inibizione attiva formulati da Pavlov e quelli di estinzione nel pensiero di Skinner, si ricollegano a dei processi attivi di formazione di un nuovo tipo di risposta. Essi risultano fondamentalmente diversi dall'oblio in quanto non presuppongono la deteriorazione della traccia mnesica iniziale.
Memoria a breve termine. - L'analisi dei dati che si riferiscono alla memoria a breve termine ha suscitato contributi sperimentali assai ingegnosi come: a) la tecnica della ‛risposta ritardata' (delayed respons; W. Hunter, 1913). Il procedimento consiste nell'addestrare un animale di laboratorio a scegliere fra numerose soluzioni possibili quella di dirigersi verso un punto della gabbia dove esso ha potuto osservare che è stato depositato e nascosto un frammento di cibo. Lasciato libero di muoversi, dopo un tempo prefissato, la sua scelta starà a testimoniare della precisione del suo ricordo. I risultati, che possono variare a seconda delle condizioni prescelte, indicano per esempio che i ratti non ricordano la posizione del cibo nascosto più di 10 secondi, il gatto più di 3 minuti, il cane più di 10 minuti; b) la tecnica di apprendimento dei ‛comportamenti di alternanza' (L. Petrinovich e R. Bolles, 1957) o di ‛eliminazione' (S. J. Lachman, 1965), che consiste nell'osservare in quale misura l'animale, posto al bivio di un labirinto a due strade, compie una scelta corretta quando la soluzione dipende dalle risposte già date nel corso di un'esperienza immediatamente precedente. Se lo si pone all'ingresso del labirinto dopo un breve intervallo di tempo, il ratto, dopo alcuni tentativi, supererà senza sbagli la prova di semplice alternanza (SDSDSD); è invece incapace di risolvere il problema rappresentato da una doppia alternanza (SSDDSSDDSSDD) che i carnivori superano con relativa facilità; c) la tecnica che I. Garcia (1955) e S. H. Revusky (1970) designano con il nome di ‛memoria associativa', che si riallaccia al comportamento particolare dell'animale il quale, messo in presenza di un alimento di cui ha precedentemente sperimentato gli effetti tossici, presenta un atteggiamento di totale rifiuto. L'esperimento, che si propone di dimostrare nel ratto la capacità di associare due stimoli distanziati nel tempo, consiste nel far seguire, a circa un'ora d'intervallo, la presentazione di un alimento di un sapore insolito (la saccarina) da una somministrazione, per via intraperitoneale, di una sostanza che provoca chiari sintomi di intossicazione. Dopo poche prove, a volte anche dopo una sola, l'animale risponde con un tipico comportamento di rifiuto se lo stesso alimento gli viene nuovamente proposto.
c) Approccio sperimentale del processo di consolidamento
Uno degli aspetti più sorprendenti della memoria nei Mammiferi consiste nel fatto che i ricordi di avvenimenti recenti sono soggetti a fattori di disturbo che non influiscono invece su ricordi più remoti.
Un individuo che ha subito una commozione cerebrale è spesso incapace di ricordare gli avvenimenti che hanno immediatamente preceduto l'incidente mentre è in grado di rievocare in modo pressoché normale eventi passati. Il fenomeno, noto come amnesia retrograda, è stato osservato in primo luogo in clinica ma si è potuto riprodurlo in laboratorio con tecniche assai diverse: l'elettroshock, l'anestesia generale da etere, l'ipossia e la depressione irradiante (spreading depression) provocata dall'applicazione diretta di una soluzione di cloruro di potassio sulla superficie della corteccia cerebrale del ratto.
A parte il loro interesse clinico, le osservazioni che riguardano l'oblio di avvenimenti immediatamente precedenti lo shock hanno avuto il merito di aprire un campo nuovo ed estremamente fecondo.
Si deve a C. P. Duncan (1949) la prima esperienza con la quale venne dimostrato come un elettroshock somministrato a un ratto possa esercitare un effetto negativo sulla ritenzione di un condizionamento di evitazione.
In forme diverse le prove sono state ripetute in varie specie animali, come lo hamster e il topo, e con altre forme di apprendimento quali la discriminazione visiva, la formazione di risposte emozionali, il comportamento operante in una gabbia a leva, l'apprendimento del labirinto, il comportamento di fuga e di evitazione nella gabbia bipartita.
Dall'insieme dei dati risulta evidente che l'amnesia è strettamente in rapporto con la durata dell'intervallo trascorso tra la fine del periodo di apprendimento e il momento in cui viene praticato l'elettroshock. Nei Roditori, il periodo critico durante il quale si manifesta l'effetto amnesico dello shock corrisponde alla durata massima di 25-40 minuti. Il procedimento permette di distinguere nettamente fra gli effetti che l'elettroshok esercita sull'apprendimento vero e proprio (memoria a breve termine) e quelli sul consolidamento di tale apprendimento nell'intervallo che intercorre fra due sessioni successive (memoria a lungo termine).
d) Determinazione genetica dei tipi di memoria
Un'ulteriore conferma dei due tipi di memoria è portata, in modo forse inatteso, dalle differenze che possono riscontrarsi in alcuni ceppi geneticamente omogenei di animali nei quali può prevalere l'una o l'altra forma di memoria. Il punto di partenza di tali studi deve ricercarsi nell'osservazione dell'estrema varietà di comportamento individuale all'interno di una stessa specie animale. Questo fatto, empiricamente noto a chiunque si sia professionalmente occupato di animali (allevatori, domatori, cavallerizzi), ha rappresentato anche per i ricercatori un problema di non lieve entità quando si è trattato, in psicobiologia, di studiare quantitativamente i parametri dell'apprendimento. È per confermare queste differenze così come per disporre di un materiale omogeneo di esperienza che, già nel 1927, R. C. Tryon ha cercato di affrontare il problema selezionando, in una popolazione di ratti, gli individui meno dotati e quelli più dotati per l'apprendimento di un labirinto. In cinque generazioni, Tryon ha ottenuto due tipi di animali dalle caratteristiche comportamentali fondamentalmente diverse.
Più recentemente I. Hirsch (1962) e G. E. McClearn (1964) hanno adottato un approccio diverso mettendo a confronto alcuni ceppi geneticamente omogenei nel topo. È stato così possibile a D. Bovet, F. Bovet-Nitti e A. Oliverio (1966) compiere un'analisi più approfondita e mettere in luce in primo luogo come le prestazioni degli animali si rivelino molto diverse a seconda del ceppo considerato e, in una seconda serie di esperimenti, come alcuni di essi possano essere dotati di memoria a breve termine e gli altri di memoria prevalentemente a lungo termine.
5. Memoria verbale nella sperimentazione umana
a) Metodi di studio
Le osservazioni cliniche e i dati sperimentali riguardanti la memoria verbale costituiscono un materiale largamente adoperato nell'analisi dei processi di ritenzione normale o patologica nell'uomo. Per quanto riguarda le tecniche utilizzate vengono distinte in primo luogo le varie categorie di materiali presi in considerazione (v. tab. II) e in secondo luogo i criteri adoperati nel processo di ritenzione (v. tab. III).


L'evocazione, il riconoscimento e il riapprendimento rappresentano i tre test sperimentali della ritenzione verbale. Nessuno di essi è perfetto (v. Woodworth, 1938).
L'evocazione è l'indice meno adeguato: esso dipende dalle condizioni del momento come dalla latenza; accade spesso che un elemento di cui non ci si arriva a ricordare in quel dato momento sia stato tuttavia memorizzato, come prova il suo ulteriore riapparire.
Un elemento di cui non ci si ricorda può invece spesso venire riconosciuto. In una serie di prove, il numero dei termini che il soggetto riconosce, cioè distingue fra quelli che ancora non conosce, è generalmente quattro volte superiore a quello che egli è in grado di evocare.
Il riapprendimento si dimostra anche un indice sensibile della memoria. Una lista di sillabe sprovviste di senso o una poesia appresa precedentemente possono essere dimenticate nel senso che il soggetto non ricorda il minimo elemento e pur tuttavia egli è in grado di riapprenderle con una facilità e una rapidità sorprendenti.
Ricerche di laboratorio hanno stabilito che l'instaurarsi o la scomparsa di un ricordo si compiono a un ritmo che potrebbe graficamente tradursi in curve a S, paragonabili a quelle ottenute per l'evoluzione della memoria sensomotoria nell'animale e nell'uomo o ancora nell'evoluzione sensoriale (H. Piéron, 1927).
La ritenzione di una serie di dati è quantitativamente apprezzabile secondo la proporzione degli elementi ritenuti dopo ogni presentazione del materiale da acquisire. Il progresso di questo fissaggio, apprezzabile numericamente, si compie in un primo tempo lentamente (tempo di latenza) poi sempre più rapidamente, presentando poi un appiattimento terminale (curva a 5 più o meno simmetrica).
Lo stesso accade per la cancellazione dei ricordi. Secondo Ebbinghaus questa non è che parziale poiché, in prove di riapprendimento successive, vi è economia nel numero delle presentazioni necessarie fino alla riacquisizione completa del materiale. L'indebolirsi del ricordo, che può essere all'inizio molto lento, si accelera poi per attenuarsi progressivamente; l'ultima parte della curva a 5 ha l'andamento di un ramo di iperbole.
b) Pluralità dei tipi di memoria nell'uomo
L'idea di una differenziazione fra memoria a breve e a lungo termine è stata generalmente accettata e anche ampliata per la molteplicità delle tracce mnemoniche (v. tab. IV).
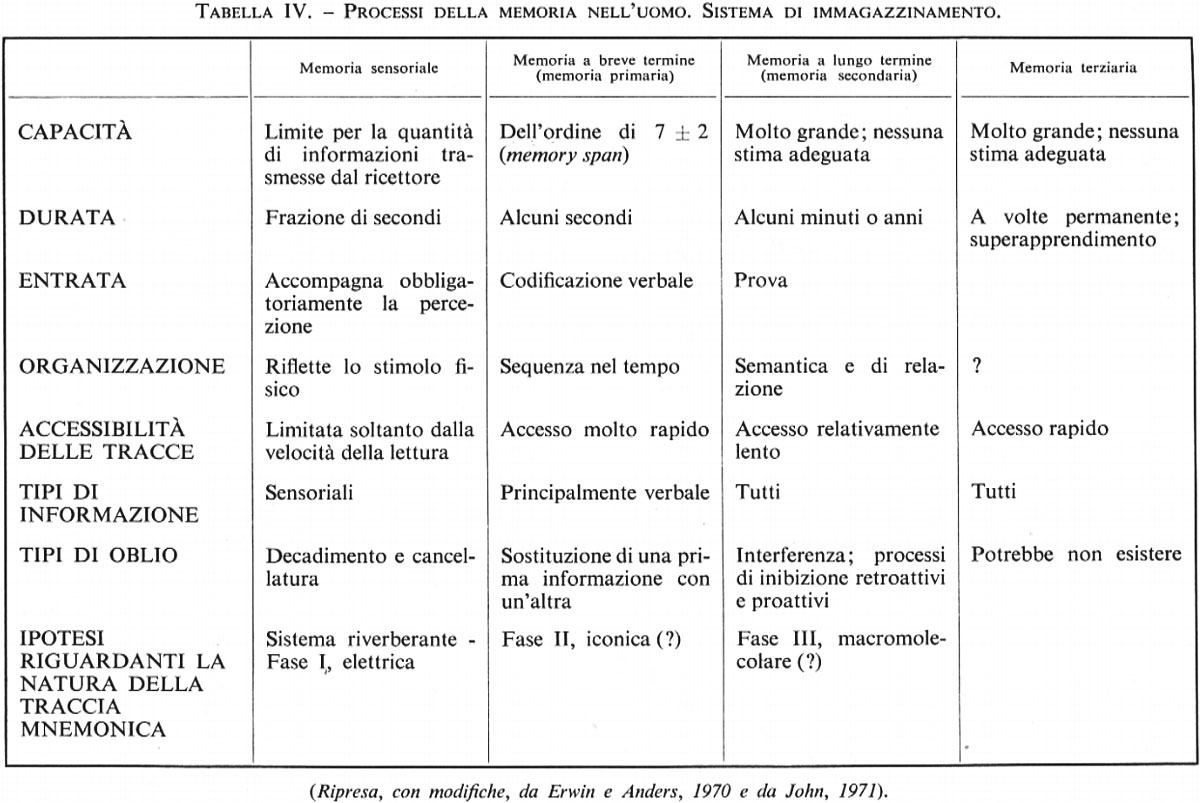
Nella memoria breve si distingue attualmente la fase iniziale che costituisce la memoria immediata o memoria sensoriale o iconica. Lo studio verte principalmente sulla memoria visiva nell'uomo. Soggetti ai quali siano stati mostrati, in rapide sequenze, diversi elementi in serie, interrogati immediatamente dopo danno prova di una memoria quasi perfetta. In un esperimento di G. Sperling (1960) venivano presentate combinazioni di lettere o di numeri per un breve tempo di 1/20 di secondo (50 ms). Se ai soggetti veniva semplicemente richiesto di riferire ciò che avevano visto, generalmente indicavano quattro elementi giusti su una serie di 8 o 12. In altre prove veniva indicata con un segnale visivo la riga da riferire. Se il segnale veniva presentato immediatamente prima o immediatamente dopo la proiezione, la precisione delle risposte si dimostrava superiore all'80%. Se il segnale veniva invece posticipato al secondo dopo la proiezione, la risposta non era superiore a quella che il soggetto era in grado di fornire in assenza di ogni segnale. Tali esperienze hanno messo in luce l'esistenza di un'immagine visiva, virtualmente perfetta all'inizio ma che non persiste normalmente più di 1/4 di secondo (250 ms).
L'analisi dei processi di memorizzazione verbale porta a distinguerne le fasi successive. Secondo una terminologia coniata fin dal 1890 da W. James, la memoria a breve termine corrisponde, nel linguaggio della psicologia corrente, alla memoria primaria e la memoria a lungo termine a quella secondaria (D. A. Norman, 1969).
Nella pratica clinica è più corrente l'impiego di termini contrapposti quali ‛memoria recente' e ‛memoria remota', perdita di ricordi ‛recenti' e ‛remoti', ‛amnesia retrograda' e ‛anterograda'.
Per ciascuno di noi è chiara la distinzione che è possibile operare fra la memoria a breve termine, che permette di ricordare un numero telefonico appena rilevato nell'elenco, per un lasso di tempo dell'ordine di qualche secondo o di un minuto, o il tipo di memoria che s'impiega in una conversazione, e la memoria a lungo termine di un allievo che legge e rilegge una poesia, del lettore che consulta un trattato di anatomia o di storia e le cui acquisizioni, riallacciate a un bagaglio culturale, potranno essere evocate e utilizzate dopo un periodo dell'ordine di un'ora, di un giorno, di un mese o di più anni.
Viene designato con l'espressione di ‛contenuto immediato della memoria a breve termine', (corrispondente a memory span degli autori anglosassoni) il numero d'informazioni (cifre, sillabe o parole) - al quale si attribuisce un valore di 7±2 - che possono essere memorizzate dopo una sola presentazione e che corrisponderebbe al limite massimo della memoria a breve termine. È stato fatto osservare che questo stesso valore di 7±2 rappresenta anche il limite del numero di oggetti (macchie disposte senz'ordine) che l'uomo è capace di discriminare e riconoscere senza ricorrere a un processo di numerazione e il limite che raggiunge l'animale di laboratorio nelle prove di discriminazione simultanee o successive.
Al contrario della memoria verbale a breve termine, che si distingue per il suo carattere al tempo stesso limitato e immediato, la memoria a lungo termine è il risultato di associazioni frequentemente ripetute che si organizzano secondo un ordine semantico o razionale e appaiono così vaste che attualmente non è possibile porre loro un limite.
Nell'analisi della fase iniziale della ritenzione, Müller e Pilzecker (1900) hanno esaminato le prestazioni fornite con un test eseguito mezz'ora dopo l'apprendimento. Il confronto fra due gruppi di soggetti, di cui gli uni erano restati inoccupati durante l'intervallo mentre gli altri erano stati sottoposti a una seconda prova di memoria, ha dimostrato che la capacità di richiamo che era del 56% dopo il riposo si abbassava al 26% nei soggetti sottoposti a un periodo continuo di sforzi mentali. Müller e Pilzecker hanno interpretato questi fatti in base a due ipotesi che si sono poi dimostrate estremamente feconde: il gioco di processi di consolidamento e il ruolo delle reazioni d'interferenza. In base al concetto del consolidamento, il processo fisiologico corrispondente alla formazione del tracciato mnemonico, lungi dal prender fine con la fase di apprendimento propriamente detta, continua attivamente nel periodo immediatamente successivo. Esso costituisce quindi un fenomeno dinamico che assume il suo pieno significato nel quadro di una concezione dualistica della memoria. Nella memoria a lungo termine sarebbe d'altra parte possibile distinguere una memoria terziaria caratterizzata dalla durata - probabilmente permanente - della traccia e dalla sua rapida accessibilità.
In contrasto con le prove sperimentali negli animali, nei quali è stato dimostrato che per l'instaurarsi di tali mutamenti biochimici sono necessari dei tempi riducibili a minuti, per l'uomo è molto più difficile determinare i limiti di una tale trasformazione. Come è stato indicato da V. Cavazzuti (v., 1973) ‟i tempi sono più elastici, più prolungati, meno definiti, sottoposti a differenze psicologiche individuali."
6. Ipotesi sulla natura dell'engramma mnesico.
a) Varietà nell'approccio sperimentale
L'interesse che rivestono le ricerche volte a mettere in luce le basi organiche della memoria proviene dal fatto che esse si iscrivono nel quadro di una visione riduzionistica e organicistica della biologia del comportamento (v. meccanicismo e vitalismo).
All'interrogativo sulla natura dell'engramma mnesico, neurofisiologi, neurochimici e chimici molecolari hanno risposto formulando due ipotesi che, lungi dall'opporsi, appaiono complementari. Secondo la prima di queste ipotesi la traccia mnesica sarebbe costituita dai potenziali d'azione che formano l'influsso nervoso. I processi di apprendimento corrisponderebbero quindi alla formazione di circuiti nervosi specifici (meta circuiti) dovuti allo stabilirsi di nuovi legami sinaptici attraverso vie innate, geneticamente determinate. In accordo con il concetto dei ‛circuiti riverberanti' avanzato da R. Lorente de No, le informazioni mnesiche si troverebbero così codificate in scariche di potenziali di azione. Secondo l'altra ipotesi, invece, l'attivazione del meccanismo chimico della trasmissione sinaptica potrebbe rappresentare l'elemento privilegiato e fondamentale della formazione della traccia mnesica. Nella memoria a lungo termine, la messa in riserva e il consolidamento mnesici appaiono codificati dalla rete di neuroni attraverso un processo di attivazione e di facilitazione permanente della trasmissione sinaptica, strettamente collegata alla sintesi proteica che prelude a una modificazione morfologica del neurone.
I dati sui quali si basano le due diverse ipotesi sono vari: a) le ricerche relative alle modificazioni dell'attività elettrica cerebrale osservate durante il condizionamento e l'apprendimento; b) lo studio delle modificazioni strutturali, anatomiche e neurochimiche che accompagnano la registrazione dell'informazione o semplicemente l'impoverimento o l'arricchimento dell'ambiente; c) l'approccio farmacologico e lo studio di sostanze in grado di modificare al tempo stesso il metabolismo cerebrale e i processi di ritenzione e mnemonici; d) infine i tentativi del trasferimento chimico dell'informazione, quali si hanno quando lo sperimentatore, somministrando a soggetti non addestrati frazioni di organi d'individui precedentemente condizionati, cerca di modificarne il comportamento.
b) Correlati elettrofisiologici
L'analisi dell'attività elettrica del cervello costituisce un primo approccio allo studio della genesi della traccia mnesica.
Si è constatato in primo luogo che uno stimolo sensorio inconsueto - per esempio la brusca illuminazione del campo visivo - provoca un'alterazione del tracciato elettroencefalografico con una reazione di allarme che corrisponde a un'accelerazione del ritmo e a una diminuzione dell'ampiezza del tracciato (ritmo alfa). Si è poi constatato che la risposta allo stimolo visivo può a sua volta essere associata e ritenuta secondo un processo del tutto paragonabile a quello di una risposta motoria. Per il solo fatto di essere stato associato a varie riprese con uno stimolo luminoso effettivo (Durup e A. Fessard, 1935) uno stimolo sonoro per sé inefficace provoca l'apparizione di una reazione di desincronizzazione o di arresto sul tracciato elettroencefalografico.
In seguito le esperienze che mettono in evidenza modificazioni del tracciato nel corso di prove di apprendimento si sono moltiplicate; esse costituiscono la dimostrazione più evidente della realtà di una ‛traccia' mnesica.
Ricerche parallele sono state effettuate sull'attività elettrica dei singoli neuroni cerebrali tramite una tecnica che permette di eliminare le irregolarità di un ritmo più o meno fluttuante. Anche in questo caso la presentazione di due segnali (luminoso e uditivo) genera la formazione di un legame temporaneo di condizionamento classico e questo a livello di un unico neurone.
c) Modifiche strutturali a livello di neuroni sinaptici
Che la memoria a lungo termine si basi su trasformazioni organiche diverse da quelle che intervengono nella fase iniziale dell'acquisizione e del condizionamento è oramai un dato solidamente stabilito. Secondo un'ipotesi non recente e più volte sostenuta, si ammette infatti che la fissazione dell'influsso nervoso potrebbe consistere in un mutamento morfologico. La traccia mnesica, legata inizialmente ai fattori intracellulari che intervengono normalmente nella liberazione e nel metabolismo dei mediatori chimici a livello di giunzione sinaptica, si tradurrebbe successivamente in vere modifiche morfologiche: aumento di superfici di aderenza e anche crescita di ramificazioni laterali e formazione di nuovi contatti. Un certo numero di dati sperimentali sta infatti a testimoniare della plasticità del sistema nervoso.
Già le osservazioni del 1913 di Dolly riguardavano le alterazioni morfologiche delle cellule di Purkinje osservate nell'animale in seguito a una stimolazione elettrica prolungata.
Un insieme di ricerche assai suggestive (Morrell, 1964) ha messo in evidenza i cambiamenti di ordine biochimico che sopravvengono nei tessuti nervosi nel corso dell'evoluzione di focolai epilettogeni secondari che si formano nell'emisfero centrolaterale in seguito a una lesione irritativa primaria. Una maggiore densità di RNA caratterizzerebbe la zona divenuta ipereccitabile. Morrell identifica le modificazioni osservate con una forma elementare di apprendimento.
Per quanto riguarda invece gli effetti delle privazioni sensoriali, una ricca letteratura indica che nei Mammiferi (coniglio, gatto) la totale privazione di stimoli visivi provoca fenomeni soltanto in parte reversibili (L. Goodman, 1932; A. Riesen, 1966). L'animale, portato alla luce, si comporta inizialmente come se fosse totalmente cieco e resta a lungo molto al disotto del normale nelle prove di discriminazione. La ricerca di modificazioni strutturali ha permesso di riconoscere una deformazione specifica delle cellule piramidali della corteccia ottica.
Un aspetto più generale del problema è stato studiato da M. Rosenzweig e collaboratori (1962, 1966 e 1968). Gli autori hanno prelevato cervelli di ratti vissuti in un ambiente complesso e ‛arricchito' e li hanno paragonati a quelli di controlli lasciati crescere in condizioni uniformi e ‛impoverite'. Sono state osservate differenze significative indicanti che l'effetto dell'esperienza si traduce, indipendentemente dall'età, ma in modo transitorio, in un aumento dei peso della corteccia cerebrale, in un aumento delle proteine totali, della colinesterasi e dell'esochinasi e, da un punto di vista morfologico, in un ispessimento della corteccia cerebrale occipitale.
d) Ipotesi sulla funzione degli acidi nucleici
Una teoria diversa, basata sull'esistenza di un engramma mnesico codificato a livello molecolare, è stata formulata in seguito alle scoperte che hanno messo in luce la funzione degli acidi nucleici nella trasmissione ereditaria.
Secondo un'ipotesi più volte formulata (W. C. Halstead, 1951; H. Hydén, 1960; v. Schmitt, 1962) l'engramma che costituisce la memoria corrisponderebbe a una messa in codice delle molecole dell'acido desossiribonucleico (DNA) o dell'acido ribonucleico (RNA) o della sequenza di amminoacidi proteici. Sono stati anche chiamati in causa altri tipi di molecole, le lipoproteine, lipidi e ‛peptidi'.
L'ipotesi macromolecolare della registrazione ha portato alcuni autori a supporre che fosse possibile trasferire un'esperienza o un apprendimento da un animale addestrato a un animale non addestrato somministrando a quest'ultimo un estratto cerebrale del primo. I primi esperimenti di questo tipo sono stati eseguiti sulle planarie, nelle quali sarebbe stata osservata la persistenza di un comportamento condizionato in prove di rigenerazione e perfino la trasmissione del contenuto dell'apprendimento allorché un verme ‛cannibale' ne divora un altro. Questi esperimenti, che non sono stati riprodotti, sono stati oggetto di vivaci polemiche in campo scientifico e hanno trovato maggior eco fuori dei laboratori che non alloro interno.
Anche nel ratto sono stati effettuati tentativi per dimostrare la possibilità di trasferire un apprendimento con la semplice iniezione, in animali non addestrati, di un estratto di cervello prelevato in altri soggetti precedentemente sottoposti a prove di condizionamento (F. R. Babich e altri, 1965; G. Ungar e altri, 1967). Ma quest'approccio, non convincente d'altronde anche concettualmente, non ha condotto a un risultato conclusivo.
e) Dati farmacologici: effetti dei farmaci psicotropi
Secondo l'immagine proposta da Cl. Bernard per il meccanismo d'azione del curaro, le molecole dei farmaci possono costituire uno ‟scalpello chimico" che, nelle mani di un ricercatore, può rivelarsi un prezioso strumento d'indagine.
Benché la memoria non costituisca ancora un campo ben definito della farmacologia del sistema nervoso centrale e benché per il momento non disponiamo, specialmente per quanto riguarda la terapia dei suoi aspetti psicopatologici, di farmaci veramente efficaci, le osservazioni compiute sugli effetti di diverse sostanze sui processi di apprendimento costituiscono già un insieme di ricerche assai suggestive.
In linea generale, i farmaci come la caffeina o le amfetammine aumentano la soglia di vigilanza, accrescono e migliorano il livello delle prestazioni. È in questo senso che vanno interpretati gli effetti di piccole dosi di nicotina facilitanti la ritenzione nell'animale e nell'uomo. Le sostanze psicotomimetiche e allucinogene (cocaina, cannabinolo, LSD) proprio in ragione degli stati confusionali che generano possono provocare l'evocazione di ricordi anche remoti.
Un metodo originale per mettere in evidenza l'azione di determinate sostanze è stato quello di somministrarle, invece che prima, nel periodo immediatamente successivo alla seduta di apprendimento (G. L. McGaugh, 1959). In queste condizioni, la narcosi provocata dall'etere etilico o dal protossido d'azoto (v. Jarvik, 1964) oppure la somministrazione di oppiacei possono determinare un'amnesia analoga a quella da elettroshock.
In senso opposto, la somministrazione di uno psicostimolante può migliorare in condizioni particolari la ritenzione. Nei due casi, l'inibizione o invece il potenziamento dei processi mnesici starebbero a indicare l'effetto dei farmaci sui circuiti elettrici neuronici, che proseguono nel si- sistema nervoso anche dopo la prova di apprendimento, nel periodo immediatamente successivo e durante il processo di consolidamento.
In stretta connessione con le ipotesi riguardanti la genesi della traccia mnesica è stato anche possibile mettere in evidenza nell'animale gli effetti inibitori esercitati sui processi di apprendimento da due gruppi di agenti farmacologici: i modificatori della permeabilità cellulare neuronica e gli inibitori della sintesi proteica. Al primo gruppo di sostanze appartengono l'uabaina e il cloruro di litio posti direttamente a contatto con la superficie di neuroni per iniezione cerebrale. È stato riconosciuto che le due so- stanze, mentre si opponevano al processo normale di formazione e di restaurazione della conduzione nervosa, bloccavano la fase iniziale di un apprendimento elementare nel pesce (R. F. Mark e M. E. Watts, 1970).
Se il consolidamento della traccia mnesica implica la partecipazione diretta di una sintesi di macromolecole, l'inibizione di questa sintesi dovrebbe impedirla. Osservazioni di laboratorio hanno chiaramente stabilito che la somministrazione d'inibitori in grado di bloccare la sintesi degli acidi ribonucleici (RNA) (actinomicina D) o la sintesi di proteine ribosomiali (puromicina, cicloesimide) (v. chemioterapia antineoplastica) impedisce anche la formazione della memoria a lungo termine (B. W. Agranoff e altri, 1966; J. J. Brink e altri, 1966).
7. Rapporti fra comportamento acquisito e strutture genetiche nell'animale
a) Il fenomeno dell'imprinting
Secondo il concetto associazionistico e tradizionale della memoria, sul quale riposa la maggior parte delle interpretazioni organicistiche che siamo attualmente in grado di formulare, lo stimolo viene considerato come un elemento fisico che ‛entra', percorre un sistema concepito come un ‛centro di accumulo' e riappare quando viene evocato'.
L'uomo e l'animale tuttavia non sono in grado di apprendere e ricordare qualsiasi cosa e la memoria va egualmente concepita nel quadro di un insieme di meccanismi solidalmente funzionanti rappresentanti la base strumentale delle attività percettive e motorie della specie (S. Ehrlich, 1970).
Per quanto riguarda il comportamento animale, diverse osservazioni hanno permesso di riconoscere da una parte i limiti delle potenzialità dei meccanismi di associazione, e dall'altra il carattere funzionale e adattativo di molte forme di ritenzione. Ci limiteremo ad alcuni esempi: a) i limiti assai ristretti nei quali sono suscettibili di stabilirsi i processi di condizionamento e di apprendimento associativi (Bolles, 1971); b) l'apparente facilità con la quale si stabiliscono, dopo una sola prova (one trial learning), alcune forme particolari di evitazione passiva (M. E. Jarvik e W. B. Essmann, 1964); c) infine l'esistenza, in special modo negli Uccelli, di processi di ritenzione, quali l'imprinting, in cui l'innato e l'acquisito appaiono come strettamente collegati.
A questo proposito si nota l'esistenza di memorie orientate, particolari per alcune specie e rispondenti a forme di comportamento strettamente adattative. È il caso per esempio dei punti di riferimento che le posizioni delle stelle rappresentano per la navigazione notturna di alcuni uccelli migratori (F. Sauer e E. Sauer, 1960) e dell'attitudine all'enumerazione di alcuni uccelli nidicoli (O. Koehler, 1954).
L'imprinting rappresenta il prototipo di memoria epigeneticamente programmata la cui acquisizione è strettamente delimitata a un breve periodo sensibile e i cui effetti invece appaiono assai durevoli se non irreversibili. Descritta in primo luogo da D. A. Spalding (1873) e da O. Heinroth (1910) nell'Uccello, questa forma di apprendimento è stata anche osservata in varie classi di Vertebrati. Comportamenti molto simili sarebbero stati riscontrati anche negli Invertebrati: Molluschi, Crostacei e Insetti.
Un'esperienza di K. Lorenz (1935) riguarda l'acquisizione da parte dei paperi dell'oca grigia di un caratteristico comportamento d'inseguimento. Si divide una covata di uova in due parti: l'una viene affidata alla madre e l'altra, posta in incubatrice, non vedrà che il solo sperimentatore. Dopo aver contraddistinto i paperi, li si pone tutti insieme sotto una tettoia che poi viene sollevata: quelli del primo gruppo si precipitano verso la madre mentre quelli del secondo gruppo raggiungono la persona che ha avuto cura di loro nei primi giorni di vita.
Nel riprendere l'esperienza in condizioni più precise, E. H. Hess (1957) ha potuto anche notare l'importanza delle condizioni sperimentali impiegate. In rapporto ai processi di apprendimento normale, quello dell'imprinting presenta un certo numero di caratteristiche innate: a) la breve durata del periodo sensibile che, in alcuni casi, è limitato alle prime ore della vita; b) il carattere generalmente unico e irreversibile dell'oggetto sul quale si fissa la scelta dell'uccellino; c) la brevità del periodo di apprendimento che può essere limitato a qualche minuto e a un percorso brevissimo; d) l'assenza di motivazione e di rinforzo o di ricompensa immediata; e) infine la lunga durata - in alcuni casi l'irreversibilità - dell'imprinting.
Un'abbondante letteratura illustra le caratteristiche degli oggetti prescelti e il significato che le relazioni stabili- te potranno rivestire nelle scelte sessuali dell'adulto (W. M. Schein, 1963; F. Schutz, 1965; K. Immelmann, 1966).
8. La struttura della memoria nell'uomo
a) Patologia della memoria
Lo studio delle perturbazioni cliniche della memoria nell'uomo ha portato un importante contributo alla conoscenza delle localizzazioni cerebrali e delle basi anatomiche della memoria verbale. Citeremo solo le sindromi meglio caratterizzate, cioè le amnesie, che si distinguono in amnesie organiche e amnesie non organiche. Dal punto di vista della loro sintomatologia si riconosce l'esistenza di: a) ‛amnesie anterograde', frequenti nei traumi cranici, che consistono in una lacuna mnemonica di tutto quanto è avvenuto dal momento del trauma in poi per un certo tempo, anche se lo stato di coscienza si trovava solo leggermente attutito; b) ‛amnesie retrograde', che consistono nella cancellazione delle tracce più recenti e che si estendono per ore, raramente per giorni, dopo l'evento che le ha provocate.
Nel primo caso si parla di disturbi della formazione della traccia mnesica, nel secondo di alterazione della conservazione delle tracce mnesiche stesse.
L'amnesia lacunare o semplice corrisponde a un'alterazione dell'evocazione mnemonica che interessa indistintamente i vari tipi di memoria per un periodo più o meno esteso.
Di più larga osservazione appaiono le amnesie semplici che si osservano nella paralisi progressiva e nei vari tipi di demenza senile o presenile, dipendenti peraltro verosimilmente da un vero deficit di tracce mnemoniche.
Negli equivalenti epilettici protratti, l'ammalato può compiere azioni assai complesse per poi entrare improvvisamente in uno stato di amnesia completa di tutto quanto è accaduto durante lo stato morboso.
L'amnesia antero-retrograda o amnesia dei fatti recenti è stata descritta da S. Korsakov nel 1889. Il quadro sindromico conclamato è costituito da sintomi caratteristici delle psicosi: amnesia anterograda, confabulazione e falsi riconoscimenti. Si distinguono l'eziologia nutrizionale, principalmente alcolica, l'eziologia vascolare, dovuta a un rammollimento bilaterale che interrompe il circuito limbico, e le eziologie tumorali dovute a processi espansivi mediani e profondi. L'eziologia alcolica è di gran lunga la più frequente e sarebbe legata a una carenza di vitamine del gruppo B per difetto di assorbimento gastrointestinale. Le lesioni, di natura emorragica, con necrosi cellulare e reazione gliale, sarebbero localizzate nell'ipotalamo e nei corpi mammillari.
Un altro importante contributo recato dalla clinica è rappresentato dall'evidenza della funzione dei lobi temporali nei processi di memorizzazione. Le osservazioni del neurofisiologo V. Bechterev (1900) sono state ampiamente confermate ed estese nel campo neurochirurgico da W. Penfield e B. Milner (1958). L'ablazione dei lobi temporali, realizzata a scopo terapeutico in pazienti affetti da epilessia, provoca una perdita delle capacità di ricordare un materiale verbale quando viene effettuata nel lato dominante (sinistro nei soggetti destri) mentre risulta senza conseguenze se eseguita dal lato opposto.
b) Ipermnesia
Le esperienze riferite precedentemente sulla memoria verbale indicano che nell'uomo sono praticamente illimitate le possibilità di apprendimento e l'entità del contenuto della memoria a lungo termine. Numerose osservazioni aneddotiche e cliniche, tuttavia, sottolineano l'esistenza di grandi variazioni individuali. Ai casi di amnesie sperimentali o provocate corrispondono casi di ipermnesie.
Ipermnesie permanenti circoscritte si osservano spesso in soggetti deficienti o idioti: ricordi del giorno, della settimana e del mese in cui cadevano determinate festività per un numero di anni quasi incredibile, memorizzazione dell'intero elenco telefonico, ecc.
Le ipermnesie transitorie possono verificarsi in casi particolari e in stati di lieve confusione mentale. Un esempio ci è dato dalla ‛visione panoramica dell'esistenza' descritta da soggetti in stato d'intossicazione o da soggetti che si son trovati nelle condizioni di aver la sensazione di morte imminente.
Le ipermnesie permanenti (M. Torre, 1965) sono generalmente circoscritte a determinati temi. Ben nota infine è la memoria di alcuni giocatori di scacchi che s'impegnano in più partite senza nemmeno guardare le scacchiere.
Notissimi sono i casi di alcuni individui particolarmente abili nei calcoli. Lo psicologo A. R. Luna (1968) ha riferito il caso di un soggetto apparentemente incapace di dimenticare, in grado di imparare, dopo una sola presentazione, un materiale estremamente complesso fra cui numeri, frasi senza senso e frasi in lingua straniera.
L'esistenza di ricordi antichi e tuttavia precisi pone il problema della forza e della persistenza della memoria remota. Il meccanismo dell'oblio e del mancato ricordo viene attribuito sia a un processo di selezione nella fase di consolidamento, sia a un difetto del meccanismo di evocazione.
Un certo numero di osservazioni ha permesso di stabilire il grado di precisione dei ricordi che si possono evocare in soggetti mediante tecniche diversissime fra loro come l'analisi psicanalitica, lo stato d'ipnosi, gli effetti degli allucinogeni o ancora la stimolazione della corteccia cerebrale.
Penfield (1963) ha riferito come, per effetto della stimolazione elettrica, un paziente può rivivere un episodio insignificante del suo lontano passato e inoltre come sia spesso in grado di ricordare tale episodio con una precisione di dettagli generalmente assai superiore a un ricordo normale. Vari casi suggeriscono che la ricchezza d'informazioni del materiale accumulato nel corso dell'esistenza di un soggetto supera di gran lunga quella dei ricordi che egli da solo è in grado di evocare.
In modo più generale le considerazioni sul contenuto della memoria non debbono far dimenticare la possibilità di un rinnovamento e di una trasformazione nella traccia mnesica. W. R. Russell (1959) ha formulato l'ipotesi del continuo rafforzarsi di tracce mnesiche già formate attraverso l'azione di correnti neuroniche casuali e senza l'intervento di un richiamo cosciente.
c) Contenuto della memoria
Nell'ammettere la possibile esistenza nel cervello di una registrazione permanente degli avvenimenti del passato, alcuni autori hanno recentemente proposto diverse valutazioni dell'ampiezza della memoria umana. J. von Neumann (1958) ha suggerito che la capacità di memorizzare nell'uomo sarebbe dell'ordine di 2,8×1020 bits d'informazioni. Ammettendo che un interruttore del tipo ‛aperto-chiuso' corrisponda a un singolo bit, ciò starebbe a significare che ognuno dei 10 miliardi circa dei neuroni del sistema nervoso è associato con l'equivalente di 30 miliardi di unità di capacità di memoria.
Le stime alle quali pervengono altri autori (J. Griffith, 1970; D. E. Wooldridge, 1963) sono relativamente più modeste.
Alcune approssimazioni suggeriscono che la quantità d'informazioni che un soggetto può acquisire tramite i suoi ricettori e ritrasmettere in un termine breve non supera i 25 bits per secondo nelle condizioni di maggiore impegno. In relazione alla durata totale della vita, 2×109 secondi o 60 anni, la capacità globale raggiungerebbe i 50 miliardi di bits, corrispondenti a 4 o 5 ‛interruttori' per cellula. Altre stime forniscono valori che variano fra 1011 e 1014 bits.
Questi dati sono probabilmente superiori a quelli reali se si tiene conto del ritmo circadiano e dell'erosione dei ricordi nel corso degli anni.
d) Il cervello come elaboratore
Nel senso più generale del termine, la parola memoria si applica all'insieme dei processi che concorrono a conservare gli effetti dell'esperienza per permetterne l'evocazione in termini di comportamento (v. tab. V).
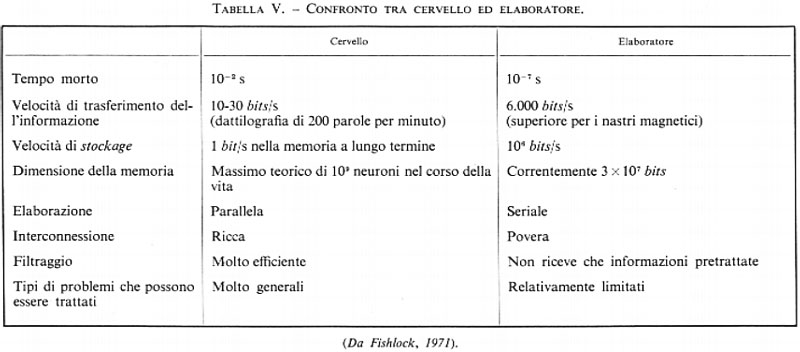
La conservazione dell'informazione rappresenta da questo punto di vista una proprietà comune ai sistemi biologici e ai sistemi fisici. Se consideriamo l'accumulo delle informazioni in un elaboratore digitale, noi osserviamo che questo è organizzato secondo una gerarchia dei livelli di memoria che vanno dal più semplice al più complesso e al più rapido. I modelli più semplici e anche i più familiari sono costituiti da una scheda perforata (che accumula 80 caratteri corrispondenti a 960 bits) e da un nastro magnetico in cui ognuno dei 9 canali paralleli contiene 320 bits per centimetro. I modelli più complessi sono formati dai tamburi e dai dischi magnetici.
Per quanto riguarda il numero d'informazioni accumulate, i valori precedentemente indicati di 1011 e 1014 bits per un cervello umano possono essere paragonati a quelli dei grandi calcolatori moderni (1010 bits) e alla massa d'informazioni di una grande enciclopedia (2×108 bits). Sotto altri aspetti i due tipi di memoria, cervello ed elaboratore, posseggono caratteristiche fondamentalmente diverse. L'elaboratore viene in soccorso del cervello accelerando la rapidità delle operazioni e aumentando la capacità della memoria. In compenso la macchina si rivela inferiore per la sua ridotta capacità di riconoscere e di correggere gli errori e per la minore varietà e generalità dei problemi che è in grado di affrontare.
bibliografia
Agranoff, B. W., Molecules and memories, in ‟Perspectives in biology and medicine", 1965, II, pp. 3-47.
Altman, J., Organic foundations of animal behavior, New York 1970.
Asratyan, E. A., Compensatory adaptations, reflex activity and the brain, Oxford 1965.
Benedetti, G., Neuropsicologia, Milano 1962.
Bovet, D., Bovet-Nitti, F., Oliverio, A. (a cura di), Attuali orientamenti della ricerca sull'apprendimento e la memoria, Accademia Nazionale dei Lincei, quaderno n. 109, Roma 1968.
Bovet, D., Fessard, A., Florès, C., Frijda, N. H., Inhelder, B., Piaget, J., La mémoire, Paris 1970.
Brogden, W. J., Animal studies on learning, in Handbook of experimental psychology (a cura di S. S. Stevens), New York 1951, pp. 568-612.
Canestrari, R., Problemi di psicologia, Bologna 1972.
Cavazzuti, V., Psicologia della memoria umana, Bologna 1973.
Erwin, F. R., Anders, T. R., Normal and pathological memory, in The neurosciences: second study program (a cura di F. O. Schmitt), New York 1970, pp. 163-176.
Fishlock, D., Man modified: an exploration of the man/machine relationship, London 1971.
Florès, C., La mémoire, in Traité de psychologie expérimentale (a cura di P. Fraisse e J. Piaget), t. IV, Paris 1964 (tr. it.: La memoria, in Trattato di psicologia sperimentale, Torino 1973, t. IV, pp. 267-439).
Gaito, J., Macromolecules and behavior, New York 1966.
Harlow, H. F., McGaugh, J. L., Thompson, R. F., Psychology, San Francisco 1971 (tr. it.: Psicologia come scienza del comportamento, Milano 1973, pp. 378-410).
Hebb, D. O., The organization of behavior, New York 1949 (tr. it.: L'organizzazione del comportamento, Milano 1949).
Hess, E. H., Imprinting and the ‛critical period' concept, in Roots of behavior (a cura di E. L. Bliss), New York 1962.
Hess, E. H., Imprinting in birds, in ‟Science", 1964, CXLVI, pp. 1128-1139.
Hydén, H., Lorenz, K., Magoun, H. W., Penfield, W., Pribram, K. H., On the biology of learning, New York-Chicago 1969.
Jarvik, M. E., The influence of drug upon memory, in Animal behaviour and drug action (a cura di H. Steinberg), London 1964, pp. 44-62.
John, E. R., Mechanism of memory, New York 1967.
John, E. R., Brain mechanism of memory, in Psychobiology (a cura di J. L. McGaugh), New York 1971, pp. 199-283.
Konorski, J., Integrative activity of the brain, Chicago 1967.
Lashley, K. S., Brain mechanisms and intelligence, Chicago 1929.
Luria, A. R., Higher cortical function in man, New York 1966.
McGaugh, J., Herz, M. J., Memory consolidation, New York 1972.
Morrell, F., Electrical signs of sensory coding, in The neurosciences: a study program (a cura di G. C. Quarton, T. Melnechuk, F. O. Schmitt), New York 1967, pp. 452-469.
Moruzzi, G., The functional significance of the ascending reticular system, in ‟Archives italiennes de biologie", 1958, XCVI, pp. 17-28.
Moruzzi, G., Active processes in the brain stem during sleep, Harvey Lectures n. 58, 1962-1963, pp. 233-297.
Moruzzi, G., Sleep and interactive behaviour, in ‟Archives italiennes de biologie", 1969, CVII, pp. 175-216.
Penfield, W., Roberts, L., Speech and brain mechanisms, Princeton 1959.
Piéron, H., Vocabulaire de psychologie, Paris 1968 (tr. it.: Dizionario di psicologia, Firenze 1973).
Razran, G., Mind in evolution. An east-west synthesis of learned behavior and cognition, Boston 1971.
Richter, D., Aspects of learning and memory, London 1966.
Schmitt, F. O., Molecular specificity and biological memory, Baltimore 1962.
Slukin, W., Imprinting and early learning, Chicago 1965.
Smith, C. U. M., Brain towards and understanding, New York 1972.
Torre, M., Psichiatria, Torino 1969.
Woodworth, R. S., Experimental psychology, New York 1938.
Young, J. Z., The memory system of the brain, Berkeley 1966.