
memoria
memoria
La memoria non costituisce un sistema unitario ma è divisibile in sottocomponenti distinte cui corrispondono sistemi neurali diversi. Nel sistema della memoria a breve termine sembrano essere implicate le strutture delle aree associative posteriori, mentre per la memoria di lavoro, quella che mantiene l’informazione per tempi brevi per eseguire su di essa operazioni, il correlato neurale dell’esecutivo centrale si trova nella corteccia prefrontale. Per la memoria a lungo termine, si distinguono il sistema della memoria dichiarativa (o esplicita) – a sua volta suddivisa in memoria semantica (di fatti e conoscenze generali) e memoria episodica (degli eventi) – e quello della memoria non dichiarativa (o implicita). L’acquisizione, la conservazione e il recupero di tracce di memoria a lungo termine dichiarativa dipendono dall’integrità dell’ippocampo e di altre strutture del lobo temporale mediale e di aree neocorticali: lesioni a queste strutture provocano amnesia ma lasciano intatta la memoria a lungo termine implicita. Per quest’ultima non c’è un sistema unico: forme diverse di memoria coinvolgono regioni cerebrali diverse. [➔ corteccia cerebrale; depressione a lungo termine; ippocampo; movimento, L’apprendimento motorio; neurogenesi; plasticità neurale; potenziamento a lungo termine; sonno] La m. è la capacità di immagazzinare informazioni per poterle utilizzare successivamente. Essa non costituisce un sistema unitario ma è divisibile in sottocomponenti distinte, ognuna delle quali svolge un compito specifico, risultando quindi distinta da un punto di vista funzionale. Alla distinzione funzionale corrisponde una distinzione anatomica fra le aree cerebrali che sottendono alle specifiche sottocomponenti. I risultati sperimentali hanno portato alla proposta dell’esistenza di sistemi di m. multipli e dissociabili.
Sistemi di memoria
Un sistema di m. è un insieme di processi portati avanti in aree cerebrali specifiche che permettono di immagazzinare o richiamare uno specifico tipo di informazione mnestica. Il suo operare può essere dissociato da quello di un diverso sistema sulla base di evidenze neuropsicologiche concordanti, per es., sulla base degli effetti di lesioni cerebrali selettive. Le prime evidenze sulla esistenza di sistemi di m. separati sono infatti venute da studi su pazienti con lesioni cerebrali specifiche. Più recentemente, a partire dall’inizio degli anni Novanta del 20° secolo, sono state utilizzate le tecniche di neuroimaging: soggetti normali sono impegnati in diversi compiti di m. e di stimolazione transcranica magnetica, attraverso la quale si disturba temporaneamente l’attività neuronale in una specifica area e si osserva come questo interferisce con lo svolgimento di un particolare compito di memoria. Agli studi sull’uomo si affiancano gli studi su modelli animali, sia utilizzando lesioni selettive sia registrando l’attività neuronale in specifiche aree. Sono stati distinti tre grandi sistemi di m.: due di m. a lungo termine, il sistema della m. dichiarativa e quello della m. non dichiarativa, e il sistema della m. a breve termine e della m. di lavoro.
Memoria a breve termine (MBT)
La MBT può contenere una quantità limitata di informazioni per breve tempo, in genere pochi secondi, dopo di che tali informazioni vengono dimenticate. La MBT può essere ulteriormente distinta in MBT verbale e MBT visuospaziale. La prima consente, per es., di tenere a mente un numero di telefono mentre andiamo verso l’apparecchio telefonico per digitarlo: questa m. è implicata anche nell’apprendimento del suono delle parole (rappresentazione fonologica) in una lingua straniera. La MBT verbale, detta anche circuito fonologico, comprende un magazzino fonologico a breve termine, che trattiene l’informazione acustica in deposito per pochi secondi, e una componente articolatoria, il ‘ripasso articolatorio’, che partecipa alla ripetizione subvocalica, e che concorre al mantenimento in m. dell’informazione (per es., quando ripetiamo, mentalmente, sottovoce o ad alta voce, un numero di telefono mentre camminiamo verso l’apparecchio telefonico). La MBT visuospaziale, detta anche taccuino visuospaziale, mantiene per pochi secondi l’informazione visuospaziale ed è coinvolta nella formazione delle immagini mentali. Le conoscenze sulla MBT visuospaziale sono inferiori a quella sulla MBT verbale, ma si ritiene che anch’essa comprenda due componenti, un magazzino visuospaziale e un tracciatore di posizione.
Memoria di lavoro (MdL)
Nella MDL si raggruppano i processi associati al mantenimento dell’informazione per tempi brevi, allo scopo di eseguire su di essa operazioni mentali. Un esempio classico è la necessità di mantenere in m. il riporto in una operazione di somma, ma anche giocare a scacchi o eseguire altri compiti che richiedono una strategia, ossia una sequenza di mosse che deve essere pianificata, simulata, valutata in termini delle conseguenze e rivista prima dell’azione. Il modello di MDL cui facciamo riferimento è quello di Alan Baddeley, che prevede tre componenti: i due sistemi di MBT sopra descritti (il circuito fonologico e il taccuino visuospaziale), e un sistema di controllo e di coordinamento, chiamato esecutivo centrale. Secondo questo modello, l’esecutivo centrale controlla e coordina i due magazzini di MBT, costruendo così un sistema operativo essenziale per poter eseguire due compiti contemporaneamente, per seguire il succedersi delle parole in modo da comprendere il significato di una frase, per formulare un ragionamento, per procedere correttamente in un discorso, per elaborare e mettere in atto una strategia, per monitorare un corso di eventi. Il circuito fonologico sembra essere localizzabile nella corteccia parietale inferiore sinistra; mentre per il taccuino visuospaziale le strutture interessate sembrano trovarsi nella aree parietotemporooccipitali di destra; il correlato neurale dell’esecutivo centrale si trova nella corteccia prefrontale (➔ funzioni cerebrali superiori).
Memoria a lungo termine (MLT)
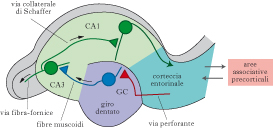
La MLT ha una enorme capacità e il ricordo può perdurare per tempi molto lunghi, giorni, mesi, addirittura anni. Si distinguono due tipi di MLT, dichiarativa e non dichiarativa. La MLT dichiarativa (o esplicita) è l’insieme delle conoscenze cui abbiamo accesso consapevole, ed è quella a cui noi facciamo comunemente riferimento quando parliamo di m. e di ricordo. A sua volta, come proposto da Endel Tulving, la m. dichiarativa può essere ulteriormente suddivisa in m. riguardante gli eventi, detta m. episodica (per es., dove mi trovavo a pranzo una settimana fa, in che occasione ho conosciuto una certa persona), e in m. riguardante i fatti, le conoscenze relative al mondo, detta m. semantica (per es., ricordarsi qual è la capitale della Francia). Le m. episodiche sono contestualizzate, ossia hanno precisi connotati spaziotemporali (in che luogo sono accaduti gli eventi ricordati e quando). Le m. semantiche non posseggono invece una componente contestuale. Per es., ricordarsi chi ha scritto Orgoglio e pregiudizio implica la m. semantica, ma ricordarsi quando lo abbiamo letto per la prima volta oppure l’aspetto del volume e dove era collocato nella libreria fa parte della m. episodica. La MLT non dichiarativa (o implicita) raggruppa tutte le conoscenze alle quali non abbiamo accesso coscientemente, come le abilità motorie e percettive (m. procedurale), l’acquisizione di regole, i comportamenti appresi attraverso processi di condizionamento, abitudine o sensibilizzazione e il priming (metodo di studio della m. basato sulla presentazione, prima della prova, di stimoli che richiamano gli elementi da ricordare). Essa si esprime nella prestazione del soggetto, non in parole, e richiede prove ripetute per formarsi. Strutture della MLT dichiarativa. Lo studio delle strutture coinvolte nella m. dichiarativa inizia con le osservazioni di William B. Scoville e Brenda Milner negli anni Cinquanta del 20° secolo. Il caso più interessante è risultato quello del paziente H. M., al quale vennero asportate bilateralmente le strutture del lobo temporale mediale (LTM). Dopo l’operazione, H. M. mostrava un quoziente di intelligenza intatto, una normale padronanza del linguaggio, un vocabolario ricco e una normale m. di lavoro, ma la capacità di formare nuove m. a lungo termine per fatti ed eventi (amnesia anterograda) era scomparsa. H. M. mostrava anche amnesia retrograda limitata, ossia perdita di m. per fatti ed eventi accaduti prima dell’intervento, che si estendeva all’indietro per qualche anno. La m. di fatti ed eventi avvenuti diversi anni prima dell’intervento era intatta. Dopo l’intervento, H. M. era ancora in grado di apprendere e memorizzare nuove informazioni: per es., aveva imparato a disegnare osservando la mano allo specchio, un tipico apprendimento di abilità che conduce a un tipo di m. non dichiarativa. Quindi, l’unica forma di MLT danneggiata in H. M. era la m. dichiarativa. Osservazioni successive hanno confermato quanto visto in H. M. e hanno individuato nell’ippocampo e nelle aree corticali circostanti (le cortecce peririnali, entorinali e paraippocampali) le strutture dell’LTM cruciali per la m. dichiarativa. L’ippocampo è bidirezionalmente connesso con la corteccia entorinale (CE), che costituisce sia l’ingresso attraverso il quale l’ippocampo riceve informazioni dalle aree corticali associative, sia la principale uscita dell’ippocampo stesso. All’interno dell’ippocampo, il flusso principale dell’elaborazione dell’informazione va dal giro dentato, che riceve informazioni dalla CE, al campo CA3, le cui cellule, oltre a proiettare al campo CA1, sono altamente interconnesse tra loro costituendo una ‘matrice di autoassociazione’, al campo CA1, che riproietta alla CE, ma esistono anche connessioni dirette fra la CE e i campi CA1 e CA3. È sufficiente lesionare l’ippocampo per avere deficit di m. dichiarativa. Questo è stato osservato, per es., nel paziente R.B., in cui un’ischemia aveva causato la perdita selettiva dei neuroni del campo CA1; R. B. mostrava un’amnesia anterograda come H. M. e un’amnesia retrograda meno grave di H. M. Anche lesioni alle aree corticali connesse con l’ippocampo, quali la CE, provocano deficit di m. dichiarativa. Del resto questo non è sorprendente, dato che la CE funge sia da ingresso sia da principale uscita per l’ippocampo. Quindi, lesioni che distruggono il flusso dell’informazione fra le diverse strutture dell’LTM, e fra di esse e le aree corticali associative, provocano deficit di memoria.
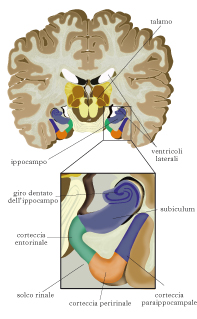
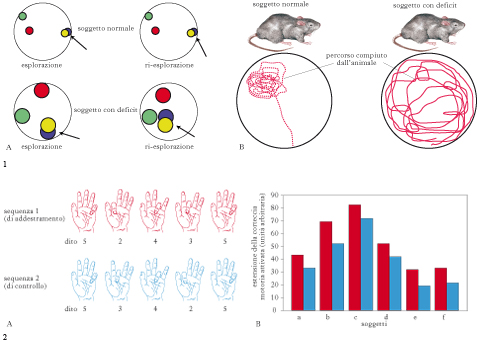
Differenti compiti delle strutture dell’LTM. Le strutture dell’LTM non hanno ruoli completamente equivalenti. Per es., anche se l’ippocampo svolge un ruolo nel riconoscimento di uno stimolo, molti studiosi suggeriscono che un ruolo cruciale nel riconoscimento della semplice familiarità venga svolto dalle aree corticali circostanti e, in partic., dalla corteccia peririnale. Giudicare un oggetto familiare è diverso dal collocarlo in un contesto spaziotemporale preciso: per es., io potrei ricordarmi di aver già visto un certo tavolo (giudizio di familiarità) ma non ricordarmi dove e in che circostanza l’ho visto. Anche per la m. semantica è stato suggerito che il ruolo dell’ippocampo possa non essere dominante, ma su questo non c’è ancora consenso. Infine, la m. di riconoscimento dei volti sembra essere sostenuta dall’attività di aree al di fuori dell’ippocampo. Viceversa, l’ippocampo svolge il ruolo dominante nella m. episodica, per es. nell’acquisizione e nel recupero delle relazioni spaziali fra oggetti in un ambiente e nella formazione di una mappa spaziale o nell’acquisizione e nel recupero della collocazione spaziotemporale delle memorie. In partic., l’ippocampo destro si attiva mentre i soggetti sono impegnati in compiti di navigazione spaziale, l’ippocampo sinistro si attiva invece fortemente in compiti che impegnano la m. verbale a lungo termine. In un lavoro recente (2008), è stato mostrato nell’uomo che un gruppo di neuroni ippocampali e della CE si attiva durante l’osservazione di brevi scene cinematiche e che il ricordo spontaneo di queste scene, effettuato tramite descrizione verbale, era preceduto dalla riattivazione di questi stessi neuroni, che sembrerebbe essere il correlato neuronale dell’esperienza soggettiva dell’emergere del ricordo. Per completezza di informazione, ricordiamo che danni alla m. dichiarativa si osservano anche in seguito a lesioni diencefaliche (corpi mammillari, sindrome di Korsakoff).
Ruolo dell’ippocampo nella formazione della MLT. L’ippocampo e le altre strutture dell’LTM sono cruciali per la formazione iniziale di una traccia di MLT dichiarativa (codifica e consolidamento), alla cui formazione contribuiscono informazioni che vengono da aree associative differenti, importanti anche per il ruolo dell’attenzione nella codificazione dell’informazione che deve essere memorizzata. Tuttavia, in precedenza abbiamo detto che lesioni all’LTM producono una amnesia retrograda limitata; questo suggerisce che l’ippocampo e le altre strutture dell’LTM siano un sito di immagazzinamento temporaneo per le tracce di MLT dichiarativa ma non il sito definitivo: quando recuperiamo un’informazione acquisita da molti anni, la recuperiamo da un magazzino che non è l’ippocampo. Il ruolo dell’ippocampo sarebbe quello di consentire i processi che conducono alla formazione di una traccia di MLT dichiarativa, associando insieme le diverse componenti che costituiscono tale traccia (per es., le caratteristiche visive e uditive di una scena). La traccia di m., poi, sarebbe lentamente trasferita nel magazzino definitivo, costituito da aree associative neocorticali, che fanno quindi parte del sistema della MLT esplicita. Vi sono quindi un consolidamento iniziale, ‘locale’, della traccia, che avviene nel sito di formazione, l’ippocampo (la cui mancanza conduce alla rapida perdita della traccia), e un consolidamento di sistema, ossia quello che permette il progressivo trasferimento della traccia di m. alla neocorteccia. Questo processo nell’uomo può richiedere molti mesi o addirittura anni ed è molto probabile che durante il suo verificarsi le tracce di m. possano essere modificate, arricchite di nuovi elementi oppure, al contrario, semplificate. L’ippocampo sarebbe importante, durante questa lunga fase, anche per il recupero della traccia di memoria. Secondo alcuni studiosi, il recupero, anche di m. da molto tempo consolidate, non si affrancherebbe mai dall’ippocampo (teoria della traccia multipla) e, in soggetti con lesioni ippocampali, la qualità del recupero delle m. più antiche, pur presente, non sarebbe equivalente a quella di soggetti normali; secondo altri, invece, le m. più antiche diventerebbero completamente indipendenti dall’ippocampo. Va anche considerato che nel giro dentato dell’ippocampo la neurogenesi (→) è attiva anche nell’adulto. Il ruolo di tale processo nella MLT dichiarativa è stato oggetto di numerosi studi, ma è ancora dibattuto se essa partecipi al processo di formazione delle tracce di m., di consolidamento di sistema o di creazione di gruppi di m. temporalmente contigue.
Ruolo delle aree associative della neocorteccia. Se le aree associative della neocorteccia sono il magazzino definitivo della m. esplicita, danni a queste aree dovrebbero danneggiare il richiamo della m. esplicita acquisita prima del danno (amnesia retrograda). Questo è esattamente ciò che si osserva e lesioni in aree associative differenti danno origine a difetti specifici della memoria. Per la m. semantica, si ritiene che le nozioni vengano conservate in maniera diffusa nelle aree associative posteriori, temporo-parieto-occipitali, mentre per la m. episodica le zone corticali specializzate per la conservazione delle tracce sembrano essere le aree associative prefrontali.
Ruolo della corteccia prefrontale (CPF) nella m. dichiarativa. Questo ruolo è ancora intenso oggetto di studio. Pazienti con lesioni localizzate alle aree associative prefrontali non mostrano l’amnesia anterograda tipica dei soggetti con lesioni all’LTM, ma mostrano la cosiddetta amnesia della fonte, ovvero l’incapacità di ricordare quando e dove un fatto è stato appreso o un evento si è verificato. In effetti, la CPF si attiva durante la codifica e il recupero di m. episodiche e la stimolazione transcranica magnetica, che ne disturba transientemente la funzione, disturba la prestazione di soggetti impegnati in compiti di m. episodica. Le interazioni fra ippocampo e CPF sono inoltre cruciali per la m. prospettica, la capacità di ricordarsi cosa dobbiamo fare e quando dobbiamo farlo, che fa riferimento ad alcune delle strutture implicate nella m. episodica.
Strutture della MLT non dichiarativa
Non c’è un sistema unico della MLT non dichiarativa: forme diverse di m. non dichiarativa coinvolgono regioni cerebrali diverse. Per es., m. emotive come quelle che si formano con un condizionamento classico che associa uno stimolo neutro a uno stimolo doloroso (condizionamento alla paura) coinvolgono l’amigdala; l’apprendimento di abilità motorie coinvolge i nuclei della base, la corteccia motoria e il cervelletto, che sembra anche coinvolto nei processi di condizionamento. L’acquisizione di regole e procedure sembra coinvolgere i nuclei della base e le loro connessioni con la CPF (vie corticostriatali). In partic., il nucleus accumbens potrebbe partecipare a processi di associazione fra azione e ricompensa. L’apprendimento di abilità percettive, per es. visive o acustiche, si correla a cambiamenti nelle aree sensoriali corticali corrispondenti e coinvolge probabilmente proiezioni dalle aree associative superiori alle aree sensoriali primarie e secondarie, proiezioni dette top-down. Anche per la MLT non dichiarativa la traccia di m. subisce un processo di consolidamento locale nella struttura in cui si è inizialmente formata, senza il quale verrebbe rapidamente perduta.
Consolidamento e riconsolidamento della MLT
Sia per la MLT dichiarativa sia per quella non dichiarativa si ipotizza che una traccia di m. si formi e si consolidi in seguito alla modificazione dell’efficacia sinaptica nei circuiti neurali delle strutture che fanno parte del sistema di m. corrispondente. Inoltre, sia per la MLT dichiarativa sia per quella non dichiarativa il sonno (➔) facilita i processi di consolidamento. A partire dal 2000, è stata avanzata l’ipotesi che, per un tempo molto lungo dopo che una traccia di m. è diventata stabile, la sua riattivazione la renderebbe di nuovo labile, soggetta quindi a interferenza e a distruzione. Per ritornare in uno stato stabile, accessibile a un ulteriore richiamo, sarebbe necessario un nuovo processo di consolidamento, detto appunto riconsolidamento. Nell’uomo è stato mostrato, per es., che se una traccia di m. non dichiarativa (apprendimento motorio) viene riattivata dopo diverse ore dalla sua iniziale formazione, quando ormai è ben stabilita, essa è cancellata da un altro apprendimento motorio se questo segue immediatamente la riattivazione, ma ciò non avviene se esso avviene a distanza di ore dalla riattivazione. Nei roditori la maggior parte delle evidenze sul riconsolidamento viene da studi sulla m. emotiva: se la traccia di m., una volta consolidata, viene riattivata (per es., il soggetto è posto di nuovo nel contesto in cui ha subito l’esperienza spiacevole), essa ritorna labile e il blocco del nuovo processo di consolidamento ne determina la cancellazione. La possibilità di interferire con il riconsolidamento di una traccia di m. emotiva riattivata potrebbe fornire nuove opportunità per progettare approcci terapeutici a disturbi nei quali la persistenza intrusiva di tracce di m. può essere responsabile di comportamenti maladattivi, quali le sindromi postraumatiche o le fobie. Nicoletta Berardi
Meccanismi cellulari e molecolari
La formazione e il consolidamento di tracce di memoria a lungo termine (MLT) dipendono dalla capacità dei circuiti neurali di modificarsi in maniera duratura, funzionalmente e morfologicamente, in risposta all’attività elettrica che l’esperienza induce nei circuiti stessi. Questa capacità prende il nome di plasticità neurale (➔) a lungo termine. La modifica avviene a livello delle sinapsi, la cui efficacia può essere potenziata, e allora si parla di potenziamento a lungo termine (LTP, Long Term Potentiation), o ridotta, e allora si parla di depressione a lungo termine (LTD, Long Term Depression). Le prime evidenze in questo senso sono state ottenute in un mollusco marino, Aplysia, da Eric Kandel. La semplicità dei circuiti neurali interessati dall’apprendimento in Aplysia ha favorito il raggiungimento di relazioni causali fra la modificazione dell’efficacia sinaptica e la formazione e la durata di tracce di memoria.
Cellule di posizione nella MLT spaziale
Si ritiene che nella MLT spaziale siano coinvolte cellule specializzate dell’ippocampo e della corteccia entorinale, le cosiddette cellule di posizione e le cellule griglia (grid cells). Le cellule di posizione si attivano nel momento in cui il soggetto si trova in una zona precisa all’interno di un ambiente, detta campo di posizione (CDP). Esse formano i loro CDP in seguito all’esperienza di esplorazione dell’ambiente compiuta dall’animale, determinando la formazione di una mappa spaziale. I CDP rimangono stabili nel tempo, in corrispondenza con la capacità dell’animale di navigare nell’ambiente anche a distanza di tempo dalla prima esplorazione: l’esperienza ha causato un cambiamento a lungo termine nei circuiti dell’ippocampo e tale cambiamento si correla con la formazione di una traccia di MLT spaziale. Se si previene la plasticità sinaptica di tipo LTP nell’ippocampo, per es. interferendo con uno dei fattori necessari perché essa venga indotta, i CDP non rimangono stabili, l’animale non forma una mappa stabile dell’ambiente, e non sarà capace in seguito di navigare al suo interno: il blocco della plasticità sinaptica previene il cambiamento nei circuiti dell’ippocampo e la formazione di una traccia di MLT dipendente dall’ippocampo. Evidenze correlative di questo tipo sono state ottenute interferendo con l’induzione o il mantenimento di LTP in strutture appartenenti ad altri sistemi di memoria e studiando la formazione delle corrispondenti tracce di MLT.
Modificazioni neuronali nell’apprendimento motorio
Un gruppo di esperimenti interessanti è stato realizzato in relazione all’apprendimento motorio. Nell’uomo, l’esercizio in un compito di destrezza manuale determina un cambiamento nella corteccia motoria primaria (M1): l’estensione della zona attivata dalla sequenza motoria esercitata diventa maggiore di quella attivata da una sequenza per la quale non è stata fatta pratica. Questo suggerisce che si sia verificato un cambiamento nell’efficacia sinaptica di un gruppo di connessioni: neuroni che non venivano reclutati per eseguire tale sequenza ora lo sono. È stato proposto che le connessioni interessate dal fenomeno di potenziamento siano un gruppo di connessioni, orizzontali, le quali uniscono neuroni reclutati per l’esecuzione di movimenti differenti, in questo caso delle dita. Questa ipotesi è stata verificata nel roditore. È stato dimostrato che l’apprendimento motorio determina il potenziamento della trasmissione sinaptica nelle connessioni orizzontali della corteccia motoria nella zona di rappresentazione dell’arto esercitato e che tale potenziamento indotto dall’apprendimento occlude l’LTP indotto attraverso la stimolazione elettrica. Questa è una evidenza molto forte a favore del fatto che l’esperienza determina l’acquisizione di nuove abilità motorie, modificando l’efficacia sinaptica in specifici circuiti motori attraverso fenomeni di plasticità sinaptica di tipo LTP.
Cambiamenti a lungo termine nelle strutture neurali
Anche per memorie acquisite con condizionamento classico o con condizionamento operante nel primo decennio del 21° secolo è stato dimostrato che l’apprendimento determina il potenziamento della trasmissione sinaptica nelle strutture neurali coinvolte nell’apprendimento stesso, e che tale potenziamento occlude LTP indotto elettricamente. Possiamo quindi affermare che l’induzione e il mantenimento di plasticità sinaptica a lungo termine indotta dall’esperienza in precisi circuiti neurali sembrano essere l’evento causato dall’apprendimento, necessario per la formazione di tracce di MLT. Per ogni tipo di memoria, i cambiamenti plastici avvengono nelle specifiche strutture neurali che fanno parte del sistema di memoria corrispondente. Questa è una importante conclusione a cui solo in tempi più recenti è stato possibile giungere grazie alla migliore conoscenza dei sistemi di memoria, all’avanzamento delle metodologie utilizzate negli esperimenti e all’uso di approcci causali. I passi che conducono a cambiamenti a lungo termine nei circuiti neurali in risposta all’esperienza coinvolgono numerosi fattori. Alcuni di questi agiscono a livello dell’induzione della plasticità, e la loro mancanza impedisce la formazione di una traccia di memoria; altri agiscono a livello del consolidamento del cambio plastico, quali i fattori che conducono alla sintesi di nuove proteine o alla trascrizione genica, entrambe necessarie affinché il cambio di efficacia sinaptica si mantenga stabile nel tempo, e la loro mancanza determina il rapido decadimento della traccia (mancanza di consolidamento). Alcuni di questi fattori risultano mutati nelle sindromi di ritardo mentale o di deficit cognitivi nell’uomo; questo ha condotto alla creazione di modelli animali specifici per deficit cognitivi umani su base genetica su cui sperimentare nuove strategie terapeutiche.
Modelli animali di memoria
I meccanismi cellulari e molecolari della memoria non possono essere studiati sul cervello umano. Per questo motivo vengono utilizzate, come sistemi modello, altre specie animali con un sistema nervoso più semplice e accessibile. L’obiettivo degli studi sui modelli animali di memoria è di comprendere le modificazioni durature e persistenti che avvengono nelle aree cerebrali e nei circuiti nervosi coinvolti in un compito di apprendimento e memoria, nel corso della transizione temporale dalla memoria a breve termine a quella a lungo termine. Vi è un generale accordo sul fatto che l’engramma (➔) dipenda da modificazioni a lungo termine della trasmissione sinaptica a livello delle connessioni coinvolte o anche da una vera e propria crescita e riorganizzazione di queste connessioni. Il presupposto dell’utilizzazione dei modelli animali di memoria è che le basi fondamentali della plasticità neuronale siano ben conservate nei sistemi nervosi degli animali più diversi. Tra gli invertebrati, due sono i modelli che hanno fornito maggiori risultati e informazioni per i meccanismi della memoria e dell’apprendimento: il moscerino della frutta Drosophila melanogaster e il mollusco marino Aplysia californica.
I lavori su Drosophila
Questi lavori, iniziati alla fine degli anni Sessanta del secolo scorso, hanno portato alla caratterizzazione di un numero crescente di mutazioni di gene singolo che interferivano con specifici compiti di apprendimento e memoria. Gli studi molecolari e comportamentali dei mutanti che non ricordano hanno dimostrato l’importanza della via di trasduzione del segnale che, attraverso il nucleotide ciclico CAMP, collega eventi di membrana alla trascrizione genica nucleare, mediata dal fattore di trascrizione CREB (CAMP Responsive Element Binding protein) nei neuroni coinvolti nei compiti di memoria. Le conclusioni raggiunte in Drosophila sono in significativo accordo con gli studi sulla memoria effettuati in Aplysia e nei mammiferi.
I lavori su Aplysia
Tra le semplici e stereotipate risposte comportamentali di Aplysia, studiate dal premio Nobel Eric Kandel, è stato approfondito il riflesso di ritrazione delle branchie, una forma di apprendimento associativo a lungo termine, ascrivibile a un semplice circuito sinaptico ben identificato che coinvolge pochi essenziali neuroni. La sensibilizzazione a breve termine coinvolge meccanismi locali, a livello della sinapsi glutammatergica tra i neuroni sensoriali e i motoneuroni che innervano le branchie. Lo stimolo sensibilizzante nocivo attiva gli interneuroni modulatori che rilasciano serotonina, attivando nel neurone sensoriale un meccanismo di segnalazione intracellulare che coinvolge il rilascio di CAMP, l’attivazione della proteina chinasi A (PKA) e una cascata di fosforilazioni di varie proteine bersaglio e determinando, infine, un aumento della quantità di neurotrasmettitore rilasciato dal neurone sensoriale sui motoneuroni. La durata della sensibilizzazione a breve termine è determinata dalla durata delle modificazioni postraduzionali transitorie indotte dalla serotonina. La transizione della sensibilizzazione da breve a lungo termine coinvolge modificazioni dell’espressione genica a livello nucleare, indotte dalla fosforilazione del fattore di trascrizione CREB da parte della PKAserotonina dipendente. Tra i prodotti della trascrizione attivati da CREB e responsabili per il mantenimento a lungo termine della sensibilizzazione, vi sono proteine responsabili dell’attivazione costitutiva della PKA e proteine responsabili per la crescita di nuovi contatti sinaptici. Si conclude che le modificazioni a breve termine dell’efficacia sinaptica coinvolgono modificazioni postraduzionali di proteine sinaptiche preesistenti, mentre le modificazioni a lungo termine implicano attivazione dell’espressione genica, sintesi di nuove proteine e formazione di sinapsi.
I lavori sui mammiferi
Le conclusioni raggiunte dagli studi compiuti sugli invertebrati sono state estese e generalizzate ai circuiti sinaptici delle aree coinvolte in processi di memoria nel cervello dei mammiferi. Tra queste, l’ippocampo, nel quale è stato molto studiato il fenomeno dell’LTP (Long Term Potentiation, ➔ potenziamento a lungo termine): una brevissima stimolazione ad alta frequenza di una delle vie sinaptiche eccitatorie ippocampali ne aumenta la efficacia della trasmissione sinaptica per un periodo considerevole di tempo (giorni o settimane). L’LTP ippocampale coinvolge sia meccanismi presinaptici (aumentato rilascio di neurotrasmettitore) sia postsinaptici (aumentato numero di recettori postsinaptici). Pur con le debite differenze, l’LTP ippocampale nei mammiferi e la sensibilizzazione a lungo termine in Aplysia hanno molti punti in comune, soprattutto per ciò che concerne la transizione tra breve e lungo termine e il dialogo tra sinapsi ed espressione genica. In entrambi i casi, queste forme di plasticità sinaptica a lungo termine sembrano costituire il substrato per l’apprendimento o l’acquisizione di nuove memorie. Il topo è un modello sperimentale di elezione per studi sulla memoria, in quanto permette di collegare il livello molecolare e genetico con il livello funzionale e comportamentale. È possibile infatti studiare gli effetti fenotipici sull’apprendimento e sulla memoria, introducendo geni esogeni nel genoma del topo (topi transgenici), oppure inattivare l’espressione di un gene endogeno dal genoma del topo (topi knock-out). In modo più selettivo, è possibile introdurre singole mutazioni in un gene endogeno di interesse (topi knock-in), ottenere un knock-out tessuto-specifico, oppure un knock-out condizionale. Dagli studi sui modelli animali di memoria, si conclude che i meccanismi di plasticità sinaptica forniscono il substrato e il fondamento cellulare e molecolare per i processi di memorizzazione delle informazioni e delle esperienze in tutte le specie animali dotate di un sistema nervoso, indipendentemente dal dettaglio anatomico e funzionale dei vari tipi di memoria.