
Mente
Mente
SOMMARIO: 1. Introduzione. 2. La coscienza. 3. I micrositi sinaptici come operatori probabilistici. 4. La possibilità che un evento mentale non materiale agisca a livello dei micrositi cerebrali secondo un meccanismo analogo a quello dei campi di probabilità della meccanica quantistica. 5. Verifiche delle teorie sul rapporto mente-cervello. 6. Il problema del rapporto mente-cervello. 7. Conclusioni generali. □ Bibliografia.
1. Introduzione.
A prima vista si riscontra, fra i filosofi, un notevole consenso a sostegno di una filosofia della mente monistico-materialistica, ma un esame più approfondito rivela una vasta gamma di opinioni al riguardo. D'altronde le concezioni dei filosofi sono andate mutando nel corso degli ultimi decenni, a mano a mano che si emergeva dalla lunga notte scura del behaviorismo. Per rendersi conto di questo mutamento graduale basta seguire il destino di certi tabù verbali. Allo zenit (o piuttosto al nadir!) dell'influenza del provocatorio libro di Ryle (v., 1949), The concept of mind, parole come ‛mente', ‛coscienza', ‛pensieri', ‛intenzioni', ‛opinioni' non erano permesse. Nell'intento di esorcizzare lo spettro cartesiano estirpandolo dalla macchina cervello-corpo, tutte le parole che avessero un vago sapore di dualismo cartesiano divennero parole ‛sporche', inammissibili in un discorso filosofico raffinato. Fra queste oscenità verbali spiccavano quattro parole inglesi di quattro lettere: mind (mente), self (sé), soul (anima) e will (volontà).
Le opinioni dei filosofi che si considerano monisti-materialisti o fisicalisti si distribuiscono lungo un ampio arco di posizioni e spesso uno stesso filosofo, col trascorrere del tempo, passa da una posizione a un'altra. È interessante il fatto che Armstrong (v., 1968 e 1981) abbia mutato significativamente le proprie concezioni nel periodo intercorso fra le pubblicazioni dei suoi due libri; la stessa cosa è capitata a Dennett (v., 1969 e 1981). Si può percepire il venir meno dell'influenza di Ryle nel corso del ventennio 1960-1980 attraverso il mutamento nell'uso delle parole da parte di questi due filosofi.
Rappresenta un'autentica ossessione per i filosofi l'idea di doversi sbarazzare del concetto di homunculus, il piccolo uomo che sta nel cervello, un residuo di dualismo cartesiano. Dennett (v., 1969, p. 190), per esempio, pensa di dover prendere in considerazione ciò che una persona fa e non la sua mente, per evitare di pervenire a prospettive cartesiane. Ma che cos'è una persona in questo contesto? Nella misura in cui tentiamo di sottoporre il problema del rapporto mente-cervello a un'analisi scientifica, dobbiamo spingere lo sguardo al di sotto della persona intesa come entità globale, fino a giungere agli eventi neurali che si verificano nel cervello e, in ultima analisi, ai più remoti micrositi delle attività sinaptiche.
2. La coscienza.
Nel cap. 4 del suo libro, The nature of mind, Armstrong (v., 1981) fornisce un meditato resoconto filosofico dell'esperienza di coscienza. Suona strana l'affermazione che una persona del tutto priva di coscienza possegga una mente dotata di conoscenze e di opinioni, che non sono definitivamente perdute, ma soltanto latenti. Eppure spesso accade che in queste condizioni riusciamo a risolvere un problema o a scoprire come esprimere una qualche idea in modo appropriato. Tutto ciò affiora alla nostra coscienza nel momento in cui si desta. Armstrong interpreta questo fenomeno come una prestazione di una coscienza minimale, perché un qualcosa di mentale sta effettivamente avvenendo. Costituiscono una classe speciale di attività mentali associate alla percezione quelle attività che insorgono quando ci si desta dal sonno e dai sogni e si diventa consapevoli dell'ambiente circostante. In questo caso la coscienza minimale si tramuta in coscienza percettiva, che sembra caratterizzare lo stato di parziale consapevolezza o, meglio, lo stato di non-attenzione. Tuttavia non si tratta ancora di uno stato di piena coscienza, che consiste in una percezione del mentale. Una tale percezione ‛interna' si chiama, tradizionalmente, ‛introspezione' o ‛consapevolezza introspettiva'. Possiamo quindi chiamare questo terzo tipo di coscienza ‛coscienza introspettiva': si tratta di ‟una consapevolezza, simile alla percezione, degli stati attuali e delle attività della nostra mente [...]. Dal momento che l'introspezione è a sua volta un'attività mentale, può anch'essa divenire oggetto di consapevolezza introspettiva". Ho voluto citare testualmente queste affermazioni di Armstrong, che sono pienamente consone alle mie opinioni, perché, altrimenti, avrei potuto dare l'impressione di aver interpretato tendenziosamente quanto egli ha scritto.
La coscienza introspettiva è ciò che può essere chiamato ‛autocoscienza' (v. Popper ed Eccles, 1977, dialogo II). Quando si presta un'intensa attenzione a una qualche esperienza percettiva, nella consapevolezza così intensificata entra in gioco la coscienza introspettiva. Questo intervento dell'autocoscienza si verifica anche nei confronti di altri stati mentali, come quelli elencati sotto il titolo ‛sensi interni' nella fig. 8: sensazioni, ricordi, pensieri, intenzioni. Noi postuliamo che tutti questi stati e queste attività, di cui siamo introspettivamente consapevoli, abbiano una base unitaria nel sé, che è un'entità individuale continua. Questa unificazione nel sé di diversi stati e attività mentali colti introspettivamente è della massima importanza, perché ci offre l'opportunità di esaminare in maniera coerente il modo in cui il nostro sistema di valori può essere integrato nelle nostre decisioni. Quindi, come sostiene Armstrong, ‟la coscienza introspettiva emerge, pienamente vigile, tipicamente in situazioni ‛problematiche', in situazioni che non si possono superare con comportamenti standard, di routine". È strano che, dopo queste argomentazioni e conclusioni mentalistiche, Armstrong affermi che ‟un fisicalista deve considerare gli stati e le attività oggetto di introspezione alla stregua di stati fisici [...] di un cervello".
La coscienza introspettiva implica un'intensa attenzione rivolta alle attività mentali in esame; pertanto Armstrong ipotizza che essa svolga un ruolo importante nel ricordo degli eventi. Si può in linea di massima convenire sul fatto che gli eventi cui non si presta attenzione non si ricordano; in particolare i ricordi degli eventi forniscono la base della storia passata del sé. Ciò spinge Armstrong ad affermare, addirittura, che ‟senza la coscienza introspettiva non saremmo consapevoli della nostra esistenza fino al momento attuale, il nostro sé non sarebbe presente a se stesso. [...] Possiamo quindi capire perché la coscienza introspettiva risulti in definitiva una condizione imprescindibile perché possa esistere un qualcosa di mentale o persino un qualcosa in generale".
Possiamo quindi chiederci: come stanno le cose per quel che riguarda gli animali superiori, che dimostrano in maniera evidente di possedere una consapevolezza cosciente (v. Griffin, 19812 e 1984)? È sufficiente attribuir loro una coscienza minimale e una coscienza percettiva? Non vi è quasi alcuna prova che gli animali posseggano una coscienza introspettiva o un'autocoscienza, ma sembrano ben dotati di facoltà mentali quali percezioni, sensazioni, ricordi, intenzioni, pensieri, e molte specie hanno una capacità ben sviluppata di ricordare eventi passati. Io suggerirei di postulare che essi abbiano una coscienza percettiva più sviluppata, simile a quella che Armstrong considera ‛di transizione' alla coscienza introspettiva.
Secondo Dennet (v. 1969, p. 99) ‟la caratteristica principale della mente, il ‛fenomeno' che più di qualsiasi altro sembra rappresentare la quintessenza di ciò che è ‛mentale' e non fisico, è la coscienza". Eppure Dennett sottopone la coscienza ad analisi, nel tentativo di mostrare che essa non è una caratteristica singola della mente, ma una congerie di caratteristiche molteplici e incompatibili. L'intento di Dennet è di scacciare ‟il piccolo uomo che sta nel cervello", che non è altro che l'entità indipendente del sé dell'interazionismo dualistico, tornata alla ribalta dopo che Ryle ne aveva esorcizzato il fantasma estirpandolo dalla macchina corpo-cervello. Dennett critica il metodo introspettivo, che è viziato dal preconcetto cartesiano dell'infallibilità del soggetto che riferisce le proprie esperienze interne. Comunque egli scinde la coscienza in due componenti: la consapevolezza in senso intenzionale, in particolare delle percezioni, e la coscienza di un qualche vago residuo, neppure collegata ai pensieri. Ma in che cosa consistono il pensare e il ragionare se non in ‛attività conscie della mente', attività di cui siamo consapevoli? Dice bene Hebb (v., 1980) a proposito del funzionamento inconscio della mente: ‟Per chiunque la propria mente è un qualcosa di estraneo, di creativo, di ignoto: un vero mistero".
Popper (v., 1982, p. 122) dice che l'origine della vita e l'emergere della coscienza animale sono miracoli e che un terzo grande miracolo è l'emergere della mente umana e della ragione.
Questo articolo illustra un tentativo di spiegare, al di là del determinismo e degli schemi indeterministici della meccanica quantistica, il fatto che noi siamo in grado di agire deliberatamente e razionalmente.
3. I micrositi sinaptici come operatori probabilistici.
La nostra indagine concerne il modo in cui funziona la corteccia cerebrale al più remoto microlivello, sicché ci occuperemo in particolare delle sinapsi spinali scoperte nel 1957 da E. G. Gray (v., 1982) e schematizzate nella fig. 4D del precedente articolo mente. Quasi tutte le sinapsi eccitatorie corticali hanno la forma riportata nella figura citata; vi sono circa 1014 sinapsi dei genere nella corteccia cerebrale umana. Il bottone appare come un rigonfiamento terminale dell'assone (la fibra presinaptica) e contiene le vescicole sinaptiche in parte connesse con le proiezioni presinaptiche dense, che sporgono dalla membrana presinaptica nella regione contrapposta alla spina sinaptica, situata al di là della fessura sinaptica (larga circa 20 nm).
Un'analisi strutturale più spinta, resa possibile, in particolare, dalla tecnica del criodecapaggio di Akert e altri (v., 1972 e 1975), ha consentito di tracciare uno schema semplificato di una sinapsi spinale; questo schema è riportato nella fig. 1, dove la sinapsi è disegnata in prospettiva e parzialmente ‛scoperchiata' per rivelarne le strutture più profonde. La disposizione delle vescicole sinaptiche e delle proiezioni presinaptiche dense, che nella fig. 4D del precedente articolo mente appariva relativamente libera, si rivela invece estremamente regolare a un'analisi più approfondita: nell'ingrandimento in alto a sinistra della fig. 1 è mostrato l'impacchettamento preciso delle vescicole sinaptiche disposte in configurazione esagonale, alternate alle proiezioni presinaptiche dense a configurazione triangolare. Questa struttura composta si chiama ‛griglia vescicolare presinaptica' e la si può considerare dotata di proprietà paracristalline (v. Akert e altri, 1975; v. Triller e Korn, 1982). I bottoni delle sinapsi cerebrali hanno un'unica griglia vescicolare presinaptica, come indicato nella fig. 1.
Tramite le tecniche più avanzate è stato possibile studiare l'influenza di una singola fibra presinaptica su un neurone. La fibra, in genere, si ramifica, in modo da fornire parecchi bottoni a un unico neurone, non come illustrato nella fig. 4D del precedente articolo mente, dove è stato raffigurato un solo bottone. Quando è stimolata, la fibra eccitatoria esercita un'azione eccitatoria, che mostra un'ampia fluttuazione di ampiezza del potenziale eccitatorio post-sinaptico (PEPS), registrato a livello intracellulare. Ai fini di questo articolo è sufficiente sapere che, tramite una sofisticata tecnica di analisi delle fluttuazioni, Jack e altri (v., The components..., 1981) hanno dimostrato che un impulso presinaptico evoca la scarica di una singola vescicola da un bottone probabilisticamente, con probabilità pari, in genere, a circa 0,33. Questo numero varia ampiamente nei diversi bottoni a partire da quasi 0, ma resta sempre minore di 1. L'originale studio probabili stico sui bottoni eccitatori di un motoneurone è stato rieffettuato da Korn e Faber (v., 1986), con un procedimento analitico diverso, nel caso dei bottoni inibitori di una cellula di Mauthner di pesce e, più recentemente, nel caso dei bottoni eccitatori delle cellule piramidali di una corteccia cerebrale primitiva, l'ippocampo di mammifero (Andersen e Redman, contributo non pubblicato).
Dato che le sinapsi del cervello dei Mammiferi (v. Akert e altri, 1972 e 1975) nonché della cellula di Mauthner (v. Triller e Korn, 1982) sembrano possedere tutte la medesima struttura di base, schematizzata nella fig. 1, si può ipotizzare che la probabilità di emissione quantica (vescicolare) sia una proprietà olistica della griglia vescicolare presinaptica di un bottone. In effetti questa probabilità non è fissa per uno stesso bottone: con un trattamento fisiologico e/o farmacologico può essere aumentata o diminuita (v. Jack e altri, Modifications ..., 1981; v. Hirst e altri, 1981; v. Korn e Faber, 1986).
Si conoscono solo valori approssimati del numero di vescicole sinaptiche inserite in una griglia vescicolare presinaptica. In genere sembra che questo numero vari fra 30 e 50, almeno secondo le indicazioni di Akert e altri (v., 1972 e 1975). Triller e Korn (v., 1982) forniscono un numero variabile fra 44 e 83 nel caso dei bottoni delle cellule di Mauthner. Pertanto solo una piccolissima parte delle vescicole sinaptiche di un bottone (che sono circa 10.000) è inserita nella zona attiva della griglia vescicolare presi naptica; tutte le altre sono disposte liberamente all'interno del bottone (v. fig. 4D del precedente articolo mente; v. fig. 1).
La fig. 2A illustra bene l'impacchettamento delle molecole di trasmettitore all'interno di una vescicola sinaptica, il movimento della vescicola fino alla griglia vescicolare presinaptica e il suo aggancio a una proiezione presinaptica della griglia presinaptica. Infine nella fig. 2B sono illustrate l'adesione della vescicola alla membrana presinaptica, sotto l'effetto degli ioni Ca2+, e la liberazione di tutte le molecole di trasmettitore nella fessura sinaptica. L'intimo contatto fra la vescicola e la membrana presinaptica (v. fig. 2B, 2) è illustrato anche nella parte sinistra della fig. 1, dove sono raffigurate due piccole protuberanze e una vescicola evidentemente pronta a scaricarsi; nella parte destra della stessa fig. 1, invece, si possono vedere, in seguito alla rimozione delle vescicole e delle proiezioni dense, i siti di attacco delle vescicole (sav), disposti in configurazione esagonale, come evidenziato anche nell'ingrandimento a destra.
4. La possibilità che un evento mentale non materiale agisca a livello dei micrositi cerebrali secondo un meccanismo analogo a quello dei campi di probabilità della meccanica quantistica.
In un suo recente libro il fisico quantistico H. Margenau (v., 1984) ha avanzato l'ipotesi che un evento mentale non materiale, come, per esempio, l'intenzione di muoversi, possa influenzare eventi neurali a livello dei micrositi senza violare le leggi di conservazione della fisica. (Si tenga presente che esistono due distinte accezioni del termine ‛quanto': la prima usata nell'ambito della fisica quantistica, la seconda nelle neuroscienze, per indicare l'unità di trasmissione sinaptica, la vescicola sinaptica). Secondo Margenau (ibid., p. 96) ‟nel caso di sistemi fisici molto complicati, come il cervello, i neuroni e gli organi di senso, i cui componenti sono abbastanza piccoli da essere governati da leggi quantistiche probabilistiche, l'organo fisico è sempre pronto a subire un gran numero di cambiamenti possibili, a ognuno dei quali è associata una probabilità definita; se avviene un cambiamento che richiede energia, o più o meno energia di un altro, l'organismo complesso la fornisce automaticamente. Pertanto, anche se la mente ha qualcosa a che fare con il cambiamento in questione, ovvero anche se esiste un'interazione mente-corpo, non è necessario che sia la mente a fornire energia". In conclusione Margenau sostiene (ibid., p. 97): ‟La mente può essere considerata alla stregua di un campo, nel senso fisico - normalmente inteso - del termine. Ma si tratta di un campo non materiale, il cui corrispettivo più prossimo, in termini analogici, è forse un campo di probabilità. Tale campo non può essere paragonato ai più semplici campi non materiali, che necessitano della presenza di materia (il flusso idrodinamico o acustico) [...] né occupa necessariamente una posizione definita nello spazio. Inoltre, per quanto ne sappiamo finora, non è un campo di energia in senso fisico, nè si deve postulare che contenga energia, per spiegare tutti i fenomeni noti di interazione fra mente e cervello".
I ‛micrositi', il cui ruolo abbiamo incominciato a discutere in via ipotetica nel capitolo precedente, non hanno ancora un'identità precisa, ma gli argomenti esposti in precedenza suggeriscono che, in virtù del meccanismo probabilistico di emissione vescicolare, le griglie vescicolari presinaptiche siano perfettamente adatte a costituire i bersagli degli eventi mentali non materiali, come, per esempio, l'intenzione di eseguire un qualche movimento. Non stiamo ipotizzando che gli eventi mentali promuovano l'attività sinaptica tramite un azione eccitatoria esercitata o sugli elementi presinaptici o su quelli postsinaptici, come suggerito nel precedente articolo mente (cap. 6). Al contrario, secondo l'ipotesi dei micrositi, gli eventi mentali si limitano ad alterare la probabilità di emissione vescicolare, che però è provocata da un impulso presinaptico. Un evento mentale eserciterebbe la propria azione sulla griglia vescicolare presinaptica paracristallina, la quale agisce in maniera globale nel controllare la probabilità di emissione di una vescicola dalla configurazione multivescicolare.
Il primo quesito da porsi concerne le dimensioni dell'effetto che un campo probabilistico quantistico può produrre. Più in particolare bisogna chiedersi se la massa della vescicola sinaptica sia tanto grande da non rientrare nell'intervallo di applicazione del principio di indeterminazione di Heisenberg. Margenau (v., 1977, p. 384) ha adattato la nota disequazione di Heisenberg al caso di calcoli inerenti a situazioni non atomiche:
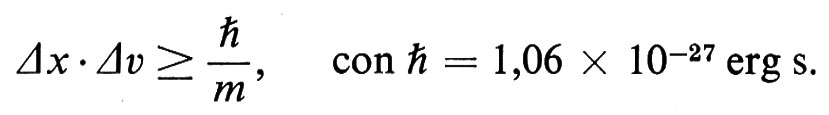
In base a opportuni calcoli si può attribuire alla massa (m) di una vescicola sinaptica di 40 nm di diametro il valore di 3 × 10-17g. Se si assume pari a 1 nm l'indeterminazione Δx relativa alla posizione della vescicola nella griglia vescicolare presinaptica, allora Δv, l'indeterminazione relativa alla velocità, risulta di 3,5 nm al ms, valore prossimo al giusto ordine di grandezza. La membrana presinaptica (v. figg. 1 e 2) è spessa circa 5 nm e il tempo di emissione di una vescicola è di parecchie decine di millisecondi (v. Katz e Miledi, 1965).
Questo calcolo, tuttavia, si basa sul presupposto che la vescicola sinaptica si muova liberamente, il che certamente non avviene quando essa si trova inserita nella griglia vescicolare presinaptica (v. fig. 1). Dato che la griglia è una struttura paracristallina, potrebbe interagire attraverso speciali relazioni di risonanza con un influsso mentale che agisse in maniera analoga a un campo di probabilità. La meccanica quantistica delle strutture microcristalline potrebbe costituire un valido supporto concettuale per la comprensione del modo in cui funziona la griglia vescicolare presinaptica. L'influsso mentale postulato si limiterebbe ad alterare la probabilità di emissione di una vescicola già situata a ridosso della membrana (v. fig. 2B), provocando l'apertura di un canale attraverso la doppia parete (2 × 5nm) delle membrane contrapposte, per il passaggio di una massa di soli 10-18 g circa.
In conclusione si può affermare che i calcoli fatti sulla base del principio di indeterminazione di Heisenberg mostrano che la probabilità di emissione di una vescicola dalla griglia vescicolare presinaptica potrebbe verosimilmente essere modificata da un'intenzione mentale che agisse in maniera analoga a un campo di probabilità quantistico.
Il secondo quesito riguarda l'ordine di grandezza dell'effetto, che consiste semplicemente in un cambiamento della probabilità di emissione di una singola vescicola (v. fig. 1). Questo evento è di parecchi ordini di grandezza troppo piccolo per modificare gli schemi dell'attività neuronica, persino in piccole aree del cervello. Comunque vi sono molte migliaia di bottoni simili su una cellula piramidale della corteccia cerebrale. Secondo l'ipotesi dei micrositi il campo di probabilità dell'intenzione mentale è ampiamente distribuito non solo sulle sinapsi di un singolo neurone, ma anche su quelle di molti altri neuroni che svolgono funzioni simili. Il prossimo capitolo è dedicato a questo argomento, con speciale riferimento alla risposta fornita dal cervello all'intenzione mentale di compiere un movimento volontario.
5. Verifiche delle teorie sul rapporto mente-cervello.
Possiamo chiederci se sia possibile effettuare verifiche sperimentali di previsioni fatte sulla base dell'ipotesi dualistico-interazionistica, da una parte, e della teoria dell'identità, dall'altra (v. mente, fig. 1). Un semplice diagramma (v. fig. 3A) illustra le caratteristiche essenziali della teoria dell'identità. Secondo Feigl (v., 1967), l'identità fra ‛mentale' e ‛nervoso' si ha soltanto nel caso di neuroni o di sistemi di neuroni appartenenti ai centri cerebrali superiori, specie alla corteccia cerebrale. Questi neuroni potrebbero essere chiamati ‛neuroni degli eventi mentalinervosi' (EMN), mentre gli altri neuroni del cervello, e in particolare i neuroni localizzati sui percorsi di ‛entrata' e di ‛uscita', non sarebbero che semplici ‛neuroni degli eventi nervosi' (EN) (v. fig. 3A). In base alla teoria dell'identità si potrebbe prevedere che i neuroni EMN siano peculiari, in quanto, in speciali circostanze, la loro attività coinciderebbe (ecco l'identità) con eventi mentali. Ma naturalmente questa attività si verificherebbe in risposta a stimoli provenienti da altri neuroni, EMN o EN, non essendo in alcun modo determinata o modificata dagli eventi mentali. In ciò consiste la ‛chiusura' del mondo fisico, cui abbiamo accennato precedentemente (v. Popper ed Eccles, 1977).
In effetti sono stati scoperti sperimentalmente, nella corteccia cerebrale, sistemi di neuroni che potrebbero essere costituiti da neuroni EMN, essendo specificamente connessi con stati mentali intenzionali o con stati di attenzione. Le ricerche che hanno portato a questa scoperta sono state condotte su individui addestrati a ‛pensare silenziosamente' in un modo particolare.
La fig. 4 mostra la posizione dell'area motoria supplementare (AMS) dell'emisfero cerebrale sinistro nella parte mediale della corteccia frontale, immediatamente davanti all'area motoria dell'arto inferiore ed estendentesi in profondità sulla faccia mediale. Tramite una tecnica basata sull'uso di Xenon radioattivo, Roland e altri (v., 1980) hanno registrato il flusso ematico regionale (FECr) in un emisfero cerebrale collegato con ben 254 contatori Geiger atti a registrare l'esatta configurazione spaziale della radioemissione indotta dall'iniezione di una piccola quantità di Xenon radioattivo nell'arteria carotide interna. È ormai appurato che un qualsiasi incremento regionale del FECr costituisce un segnale attendibile di un aumento dell'attività neuronica nell'area in questione. Il soggetto era stato addestrato a eseguire una complessa sequenza di movimenti con le dita per tutta la durata (45 s) del conteggio Geiger.
La fig. 5A mostra, come prevedibile, una forte attivazione delle aree controlaterali motoria e sensitiva relative al pollice e alle altre dita, accompagnata, però, da un'attivazione altrettanto forte e bilaterale dell'AMS. La supremazia dell'AMS è evidenziata nella fig. 5B, che si riferisce al caso in cui il soggetto non esegue materialmente alcun movimento, ma si limita a espletare mentalmente il compito motorio appreso, operazione definita ‛pensiero silente'. In queste condizioni sperimentali si è registrato, relativamente alla sola AMS e su entrambi i lati, un aumento del 2o%, altamente significativo, dell'attività neuronica. Il soggetto si trovava in una condizione di riposo assoluto, con occhi e orecchie chiusi. In questo caso l'incremento del FECr indica un aumento dell'attività neuronica dell'AMS sotto l'influsso di un'intenzione mentale concepita dal soggetto. Evidentemente l'intenzione mentale attiva un enorme insieme di neuroni, essenziali, naturalmente, per provocare il movimento desiderato.
Per mezzo di un microelettrodo inserito nel cervello è stato possibile studiare le risposte di singoli neuroni dell'AMS di una scimmia mentre eseguiva un movimento volontario (v. fig. 6, A e C; v. Brinkman e Porter, 1979). Si è cosi registrato un aumento della frequenza di scarica di molti neuroni, circa 50 ms prima della scarica dei neuroni motori corticali, che alla fine hanno provocato il movimento voluto, come puntualmente segnalato dall'elettromiogramma (v. fig. 6B; v. i contributi di Eccles del 1982).
Motivi di ordine etico vietano di eseguire un esperimento del genere su un soggetto umano; comunque anche le registrazioni di campi elettrici e magnetici, effettuate nell'uomo tramite elettrodi applicati allo scalpo durante l'esecuzione di movimenti volontari ripetitivi (v. Deecke e Kornhuber, 1978), indicano che i neuroni dell'AMS costituiscono la sede di una forte attivazione da parte dell'intenzione mentale.
Secondo l'ipotesi dei micrositi, la griglia vescicolare presinaptica offre all'intenzione mentale l'‛opportunità' di cambiare ‛a scelta' la probabilità dell'emissione sinaptica. Ciò varrebbe per tutto il complesso delle sinapsi spinali attivate a un dato momento, probabilmente addirittura migliaia, dato che ce ne sono circa 10.000 su una singola cellula piramidale corticale (v. Szentàgothai, The neuron ..., 1978). Un influsso mentale analogo a un campo di probabilità potrebbe esercitare un'influenza globale sulle sinapsi di un appropriato neurone, aumentando o diminuendo le probabilità dell'emissione vescicolare indotta dagli impulsi in arrivo (v. Margenau, 1984). In tal modo l'efficacia di un'intenzione mentale dipenderebbe dall'integrazione degli eventi probabilistici che si verificano nelle numerose griglie vescicolari presinaptiche di quel neurone.
Per determinare un qualche movimento volontario, come piegare un dito, l'intenzione mentale deve scegliere le giuste cellule piramidali su cui agire modificandone la probabilità di emissione vescicolare. Questa scelta sarebbe effettuata sulla base dell'inventano, appreso, delle cellule dell'AMS responsabili di un particolare movimento. Perché questa scelta sia efficace è necessario che esista uno sbarramento sinaptico di fondo su quelle cellule, in quanto tutto ciò che l'intenzione mentale può fare è modificare la probabilità di emissione vescicolare delle sinapsi attivate. Tanji e Kurata (v., 1982) hanno dimostrato l'ampia convergenza di inputs sensoriali sulle cellule dell'AMS. Tutti i bottoni attivati possono essere considerati siti ove ha luogo la modificazione della probabilità di liberazione vescicolare, provocata dall'intenzione mentale.
Questo può sembrare un metodo alquanto rozzo per iniziare un movimento volontario, ma bisogna considerare che noi abbiamo la capacità di effettuare, a volontà, una immensa gamma di movimenti e ciò richiede una strategia oltremodo complessa con cui selezionare neuroni dell'AMS individuandoli nel contesto di circa 108 cellule piramidali associate in - forse - 30.000 moduli. Tutto ciò di cui abbiamo esperienza diretta è come iniziare mentalmente il movimento specifico. È fondamentale che l'intenzione mentale provochi l'attivazione di blocchi di neuroni dell'AMS nella giusta sequenza temporale per i diversi muscoli implicati nell'atto motorio (v. Brinkman e Porter, 1979 e 1983). Secondo la spiegazione più semplice queste cellule dell'AMS proiettano alle altre aree corticali e sottocorticali in modo che i programmi motori appresi risultino incorporati nell'attivazione finale delle cellule piramidali motorie che scaricano lungo il tratto piramidale.
Riepilogando si può affermare che, per stabilire la validità dell'ipotesi dei micrositi, basta che essa riesca a render conto della capacità di un evento mentale non materiale di mutare la probabilità dell'emissione vescicolare da un singolo bottone facente capo a una cellula piramidale corticale. Se ciò può avvenire nel caso di un singolo bottone, può avvenire anche nel caso di numerosi bottoni facenti capo a quel neurone: tutto il resto avviene di conseguenza, in accordo con le teorie neurofisiologiche del controllo motorio. La chiusura del 1° mondo è stata così violata e tramite un'intenzione mentale noi siamo veramente in grado di effettuare movimenti a volontà.
La fig. 7A illustra una notevole scoperta di Roland (v., 1981): quando il soggetto umano si concentrava mentalmente su un dito cui stava per essere applicato uno stimolo tattile appena percettibile, si verificava un aumento del FECr nell'area tattile del dito del giro postcentrale della corteccia cerebrale, nonché nell'area prefrontale mediana. Questi aumenti dovevano essere stati determinati dall'attenzione mentale, dato che nessuno stimolo tattile era stato applicato durante la registrazione. Quindi la fig. 7A costituisce una chiara dimostrazione del fatto che l'atto mentale dell'attenzione può attivare appropriate regioni della corteccia cerebrale. Analogo risultato si ottiene nel caso che il soggetto presti attenzione alle proprie labbra in attesa di un contatto, ma ovviamente l'area somatosensitiva attivata in questo caso è quella relativa alle labbra.
Il fatto che l'attenzione provochi un incremento della risposta elettrica cerebrale a uno stimolo tattile esercitato su un dito è stato dimostrato da Desmedt e Robertson (v., 1977). In una ricerca molto ingegnosa essi scoprirono che, toccando il dito su cui il soggetto fissava la propria attenzione, si determinava un grande aumento dei potenziali tardivi evocati N 140 e P 500, rispetto ai controlli eseguiti con lo stimolo tattile delle altre dita (cui il soggetto non prestava attenzione). Tutto ciò può essere correlato con l'incremento del FECr prodotto dall'attenzione nell'area tattile relativa al dito (v. fig. 7A). In entrambe queste ricerche si è potuto constatare come un evento mentale, l'attenzione, abbia prodotto risposte neuroniche selettive.
Un risultato analogo si ottiene nel caso in cui il soggetto sia impegnato nella semplice attività di contare (mentalmente) o nell'eseguire (sempre mentalmente) altre operazioni aritmetiche, in uno stato di completo rilassamento, con occhi e orecchie chiusi; in tali condizioni si è riscontrato un incremento del FECr in molte aree corticali, ma non nelle aree sensitive o motorie primarie (v. Roland e Friberg, 1985). Nel caso dell'operazione aritmetica consistente nell'eseguire mentalmente successive sottrazioni del numero 3 a partire da 50, si è verificato un incremento del FECr in una fascia mediale della corteccia frontale, posta davanti all'AMS, e anche in altre aree della corteccia prefrontale di ambedue gli emisferi, nonché nei giri sopramarginale e angolare di entrambi i lobi parietali (v. fig. 7B). In questo caso gli schemi sono più complessi di quelli relativi alla rappresentazione mentale di un movimento (v. fig. 5B). A schemi ancora più complicati hanno dato luogo le esecuzioni di altri esercizi, quali la memorizzazione di una sequenza di parole prive di senso e la raffigurazione mentale dell'atto di percorrere un tracciato.
È prevedibile che in futuro si giungerà a constatare che le innumerevoli operazioni mentali di cui siasno capaci attivano un numero talmente elevato di regioni specifiche della corteccia cerebrale da far sì che la maggior parte della neocorteccia sia sottoposta all'influenza mentale del pensiero (v. Ingvar, 1985). Naturalmente finora non esiste alcun criterio per dimostrare un'influenza diretta. Le aree attivate direttamente possono influenzare a loro volta altre aree, come nel caso dell'AMS (v. fig. 5B) che attiva la corteccia motoria (v. fig. 5A).
L'ipotesi dei micrositi, secondo cui eventi mentali non materiali altererebbero la probabilità di emissione vescicolare dalle griglie vescicolari presinaptiche, può spiegare tutte queste influenze esercitate dalle operazioni mentali. Viceversa i sostenitori della teoria dell'identità dovrebbero postulare che l'attività dei neuroni EMN (v. fig. 3A) fosse generata esclusivamente per via neuronica e fosse spiegabile come risposta a inputs provenienti o da neuroni EN o da altri neuroni EMN appartenenti ai centri cerebrali superiori. In tal modo la chiusura del 1° mondo del cervello viene conservata al prezzo inaccettabile di negare l'azione promotrice svolta da processi mentali quali le intenzioni e l'attenzione, azione illustrata nelle figg. 5, 6 e 7. La differenza fra la teoria dell'identità e l'ipotesi dei micrositi è cruciale; secondo quest'ultima, infatti, i neuroni EMN riceverebbero, oltre agli inputs provenienti da altri neuroni EMN e dai neuroni EN, anche un input proveniente autonomamente da eventi mentali (EM), rappresentato dalle frecce aggiuntive che compaiono nella fig. 3B. Diversa sarebbe la risposta dei neuroni EMN in assenza di eventi mentali quali l'intenzione, l'attenzione o il pensiero silente (v. figg. 5B e 7, A e B).
6. Il problema del rapporto mente-cervello.
Per formulare più precisamente l'ipotesi dualistica dell'interazione mente-cervello, bisogna premettere che il mondo degli eventi mentali (2° mondo) ha un'esistenza altrettanto autonoma quanto quello della materia-energia (1° mondo) (v. fig. 8). Va detto che il solo mezzo di cui disponiamo per conoscere il 1° mondo sono i nostri organi di senso. Gli organi di senso ci forniscono i dati percettivi su cui si basano le nostre azioni, i nostri pensieri e i nostri ricordi, e quindi i dati su cui si fondano tutte le attività umane, comprese la scienza e la tecnologia. L'ipotesi interazionistica qui presentata non ha niente a che vedere con questi problemi ontologici, ma riguarda soltanto il modo in cui gli eventi mentali agiscono sugli eventi neurali; in altre parole concerne la natura delle frecce che compaiono nella fig. 8, puntate verso il basso, attraverso la frontiera (interfaccia) fra 1° e 2° mondo. Secondo l'ipotesi dei micrositi l'influenza mentale modifica la probabilità di emissione vescicolare da un bottone attivato, attraverso un meccanismo analogo a quello seguito dai campi di probabilità della meccanica quantistica.
Dato che si postula che gli eventi mentali possano influenzare soltanto eventi neurali indotti da impulsi presinaptici a sottostare al meccanismo probabilistico di emissione quantica (vescicolare), è prevedibile che l'efficacia degli eventi mentali sarebbe ridotta a zero se fosse ridotta a zero l'attività presinaptica di fondo. In tal caso si verificherebbe una perdita di coscienza irreversibile, a meno che non si verificasse una netta ripresa della scarica degli impulsi nella corteccia cerebrale. Un esempio di questo stato è il coma ‛vigile', che sopraggiunge allorché un trauma che colpisce il mesencefalo blocca il sistema reticolare attivante (v. Hassler, 1978; v. Eccles, 1980, p. 160). In effetti è possibile che il ruolo principale del sistema reticolare attivante sia quello di fornire una base di impulsi eccitatori alla corteccia cerebrale, predisponendo un'enorme quantità di emissioni vescicolari probabilistiche, che costituiscono i bersagli dei campi quantici probabilistici di influenza mentale.
Sicché possiamo supporre che gli eventi mentali interagiscano in maniera globale con gli eventi neurali relativi a schemi spazio-temporali di attività della corteccia cerebrale desta (v. Eccles, How..., 1982). Persino in un singolo modulo corticale, con i suoi 3.000 neuroni circa, dev'esserci un'incessante intensa attività dinamica di complessità inimmaginabile. Benché conosciamo per grandi linee la struttura nervosa di un modulo (v. mente, fig. 5: v. Szentàgothai, The neuron ..., 1978 e 1983), finora lo studio della fisiologia relativa non ha fatto grandi progressi. Tutto ciò che possiamo congetturare è che eventi mentali agenti come un campo nel modo postulato da Margenau (v., 1984) siano in grado di modificare l'attività spazio-temporale di un modulo alterando la probabilità di emissione in molte migliaia di sinapsi attive. Ciò non implica alcuna violazione di leggi di conservazione.
Ci si può chiedere come faccia la scimmia a erigere l'immensa barriera sinaptica che dà luogo all'attività neuronica illustrata nella fig. 6, A e C, che, a sua volta, attraverso i complessi circuiti ben noti, determina l'atto motorio desiderato. L'unica risposta possibile è che questa ‛prestazione' rappresenta la tappa finale di una lunga sequenza di sedute di addestramento. L'apprendimento motorio è essenziale per poter eseguire tutte le azioni che implicano una certa competenza, dipendenti dalla corteccia cerebrale, e ciò vale in particolare per le azioni umane (v. Eccles, 1986). Una memoria di un qualche tipo è necessaria per tutte le esperienze e le azioni coscienti.
Un'ultima considerazione a proposito delle frecce che, nella fig. 8, sono dirette verso l'alto, dalla corteccia cerebrale alla mente, per esempio quelle dirette verso la ‛percezione' nella parte sinistra della figura. È possibile che la scarica di una vescicola da una griglia vescicolare presinaptica causi un evento mentale tramite un'onda di probabilità quantistica diretta in verso opposto? Dato l'altissimo numero di emissioni vescicolari che avverrebbero nelle aree percettive della corteccia cerebrale, potrebbe verificarsi una sommazione di moltissimi eventi mentali ‛unitari' fino al raggiungimento della soglia percettiva.
7. Conclusioni generali.
Un'osservazione di carattere generale è che tutte le ipotesi finora avanzate nel tentativo di fornire una qualche spiegazione di come le esperienze coscienti derivino da, o siano collegate con, eventi neurali si sono concentrate sull'estrema complessità degli eventi neurali che hanno luogo nella corteccia cerebrale attiva; di tal fatta sono le ipotesi avanzate da Feigl (v., 1967) e da Sperry (v., 1976), secondo cui gli eventi mentali sono proprietà configurazionali olistiche del processo cerebrale. Mountcastle (v., 1978) ha elaborato il concetto di ‛sistemi distribuiti': sistemi ‟composti da numerosissimi elementi modulari associati reciprocamente in blocchi disposti in parallelo e in serie", che si pensa forniscano un meccanismo obiettivo di consapevolezza cosciente. Edelman (v., 1978) ha suggerito l'ipotesi che ‟il cervello elabori selettivamente i segnali sensoriali e le informazioni contenute nella propria memoria in maniera fasica (ciclica) e rientrante, capace di generare le condizioni necessarie per gli stati consci". Szentàgothai (v., The local ..., 1978) ha prospettato la possibilità di spiegare scientificamente le funzioni superiori del cervello, compresa anche la coscienza, in termini di ‟sovrastrutture" determinate da ‟modelli dinamici". Secondo Eccles (v., How ..., 1982) ‟l'influenza mentale si esercita su un sistema dinamico, estremamente complesso, di neuroni interagenti".
L'alternativa opposta a queste ipotesi ‛nebulari' è quella qui proposta, vale a dire l'ipotesi secondo cui il luogo fondamentale dell'azione esercitata sul cervello da eventi mentali non materiali è costituito da singoli micrositi, le griglie vescicolari presinaptiche dei bottoni, ognuna delle quali libera le singole vescicole, in risposta a un impulso presinaptico, in maniera probabilistica. È questa probabilità che si suppone modificata da un'influenza mentale agente in modo analogo a un campo di probabilità quantistico, nella maniera descritta precedentemente. Il modo in cui l'azione efficace a livello dei micrositi viene amplificata dai circuiti nervosi convenzionali dipende dai complessi circuiti presi in considerazione (v. Feigl, 1967; v. Sperry, 1976; v. Mountcastle, 1978; v. Edelman, 1978; v. Szentàgothai, The local..., 1978; v. Eccles, How ..., 1982). L'ipotesi dei micrositi può essere considerata un tentativo di avviare uno studio scientifico del ‛circuito riflettente' di Creutzfeldt (v., 1979), considerato il tramite fra il cervello e il mondo simbolico autonomo della mente (il 2° mondo di Popper ed Eccles: 1977). In contrasto con le ipotesi ‛nebulari', l'ipotesi dei micrositi lancia una sfida unica nel suo genere alla neurobiologia molecolare.
Bibliografia.
Akert, K., Peper, K., Sandri, C., Structural organization of motor end plate and central synapses, in Cholinergic mechanisms (a cura di P. G. Wasser), New York 1975, pp. 43-57.
Akert, K., Pfenninger, K., Sandri, C., Moor, H., Freeze etching and cytochemistry of vesicles and membrane complexes in synapses of the central nervous system, in Structure and function of synapses (a cura di B. P. Papas e D. F. Purpura), New York 1972, pp. 67-86.
Armstrong, D. M., A materialist theory of the mind, London 1968.
Armstrong, D. M., The nature of mind, Ithaca, N. Y., 1981.
Brinkman, C., Porter, R., Supplementary motor area in the monkey: activity of neurons during performance of a learned motor task, in ‟Journal of neurophysiology", 1979, XLII, pp. 681-709.
Brinkman, C., Porter, R., Supplementary motor area and premotor area of the monkey cerebral cortex: functional organization and activities of single neurons during performance of a learned movement, in ‟Advances in neurology", 1983, XXXIX, pp. 393-420.
Creutzfeldt, O. D., Neurophysiological mechanisms and consciousness, in Brain and mind, Ciba Foundation Series 69, Amsterdam 1979, pp. 217-233.
Deecke, L., Kornhuber, H. H., An electrical sign of participation of the mesial ‛supplementary' motor cortex in human voluntary finger movement, in ‟Brain research", 1978, CLIX, pp. 473-476.
Dennett, D. C., Content and consciousness, New York 1969.
Dennett, D. C., Brainstorms, Cambridge, Mass., 1981.
Desmedt, J. E., Robertson, D., Differential enhancement of early and late components of the cerebral somatosensory evoked potentials during forced-paced cognitive tasks in man, in ‟Journal of physiology", 1977, CCLXXI, pp. 761-782.
Eccles, J. C., Mente, in Enciclopedia del Novecento, vol. IV, Roma 1979, pp. 91-102.
Eccles, J. C., The human psyche, Berlin-Heidelberg-New York 1980.
Eccles, J. C., How the self acts on the brain, in ‟Psychoneuroendocrinology", 1982, VII, pp. 271-283.
Eccles, J. C., The initiation of voluntary movements by the supplementary motor area, in ‟Archiv für Psychiatrie und Nervenkrankheiten", 1982, CCXXXI, pp. 423-441.
Eccles, J. C., Learning in the motor system, in Progress in brain research (a cura di H.-J. Freund, U. Büttner, B. Cohen e J. Noth), vol. LXIV, Amsterdam 1985, pp. 3-18.
Edelman, G. M., Group selection and phasic reentrant signalling: a theory of higher brain function, in The mindful brain (a cura di G. M. Edelman e V. B. Mountcastle), Cambridge, Mass., 1978, pp. 51-100.
Feigl, H., The ‛mental' and the ‛physical', Minneapolis 1967.
Gray, E. G., Rheabilitating the dendritic spine, in ‟Trends in neurosciences", 1982, V, pp. 5-6.
Griffin, D. R., The question of animal awareness, Los Altos 19812.
Griffin, D.R., Animal thinking, Cambridge, Mass., 1984.
Hassler, R., Interaction of reticular activating system for vigilance and the truncothalamic and pallidal systems for directing awareness and attention under striatal control, in Cerebral correlates of conscious experience (a cura di P. Buser e A. Rongeul-Buser), Amsterdam 1978, pp. 110-129.
Hebb, D. O., Essay on mind, Hillsdale, N. J., 1980.
Hirst, G. D. S., Redman, S. J., Wong, K., Post-tetanic potentiation and facilitation of synaptic potentials evoked in cat spinal motoneurones, in ‟Journal of physiology", 1981, CCCXXI, pp. 97-109.
Ingvar, D. H., Memory of the future. An essay on the temporal organization of conscious awareness, in ‟Human neurobiology", 1985, IV, pp. 127-136.
Jack, J. J. B., Redman, S. J., Wong, K., The components of synaptic potentials evoked in cat spinal motoneurones by impulses in single group 1a afferents, in ‟Journal of physiology", 1981, CCCXXI, pp. 65-96.
Jack, J. J. B., Redman, S. J., Wong, K., Modifications to synaptic transmission at group 1a synapses on cat spinal motoneurones by 4-aminopyridine, in ‟Journal of physiology", 1981, CCCXXI, pp. 111-126.
Katz, B., Miledi, R., The measurement of synaptic delay and time course of acetylcholine release at neuromuscular junction, in ‟Proceedings of the Royal Society" (Series B), 1965, CLXI, pp. 483-495.
Kelly, R. B., Deutsch, J. W. Carlson, S. S., Wagner, J. A., Biochemistry of neurotransmitter release, in ‟Annual review of neuroscience", 1979, II, pp. 399-446.
Korn, H., Faber, D.S., Regulation and significance of probabilistic release mechanisms at central synapses, in New insights into synaptic function (a cura di G. M. Edelman, W. E. Gall e W. M. Cowan), New York 1986.
Margenau, H., The nature of physical reality, Wooldbridge, Conn., 1977.
Margenau, H., The miracle of existence, Woodbridge, Conn., 1984.
Mountcastle, V.B., An organizing principle for cerebral function: the unit module and the distributed system, in The mindful brain (a cura di G. M. Edelman e V. B. Mountcastle), Cambridge, Mass., 1978, pp. 7-50.
Penfield, W., Roberts, L., Speech and brain mechanisms, Princeton 1959.
Popper, K. R., The open universe, London 1982.
Popper, K.R., Eccles, J. C., The self and its brain, Berlin-Heidelberg-New York 1977.
Roland, P.E., Somatotopical tuning of postcentral gyrus during focal attention in man. A regional cerebral blood flow study, in ‟Journal of neurophysiology", 1981, XLVI, pp. 744-754.
Roland, P.E., Friberg, L., Localization in cortical areas activated by thinking, in ‟Journal of neurophysiology", 1985, LIII, pp. 1219-1243.
Roland, P.E., Larsen, B., Lassen, N. A., Shinkøj, E., Supplementary motor area and other cortical areas in organization of voluntary movements in man, in ‟Journal of neurophysiology", 1980, XLIII, pp. 118-136.
Ryle, G., The concept of mind, London 1949.
Sperry, R.W., Mental phenomena as causal determinants in brain function, in Consciousness and the brain (a cura di G. G. Globus, G. Maxwell e I. Savodnik), New York 1976, pp. 163-177.
Szentágothai, J., The local neuronal apparatus of the cerebral cortex, in Cerebral correlates of conscious experience (a cura di P. Buser e A. Rongeul-Buser), Amsterdam 1978, pp. 131-138.
Szentágothai, J., The neuron network of the cerebral cortex. A functional interpretation, in ‟Proceedings of the Royal Society" (Series B), 1978, CCI, pp. 219-248.
Szenthágothai, J., The modular architectonic principle of neural centers, in ‟Review of physiology, biochemistry and experimental pharmacology", 1983, IIC, pp. 1-61.
Tanji, J., Kurata, K., Comparison of movement-related activity in two cortical motor areas of Primates, in ‟Journal of neurophysiology", 1982, XLVIII, pp. 633-653.
Triller, A., Korn, H., Transmission at a central inhibitory synapse. III. Ultrastructure of physiologically identified an stained terminals, in ‟Journal of neurophysiology", 1982, XLVIII, pp. 708-736.