
Metodi comparativi per lo studio del comportamento
Metodi comparativi per lo studio del comportamento
II metodo comparativo, utilizzato già da Aristotele per descrivere le differenze di socialità tra carnivori ed erbivori, si basa su confronti tassonomici incrociati che hanno lo scopo di identificare modelli di evoluzione del comportamento. Questi confronti riguardano evidentemente taxa con origine evolutiva comune, determinata tramite gli alberi filogenetici. Grazie ai rivoluzionari progressi nelle tecniche di analisi e di raccolta dei dati è divenuto possibile saggiare con precisione le ipotesi evoluzionistiche negli studi comparativi del comportamento. Lavori recenti hanno prodotto chiari modelli filogenetici di comportamento relativi alle origini storiche, ai cambiamenti in sequenza, alla covariazione con altri tratti, alla instabilità e alla velocità evolutiva. Gli sviluppi più promettenti di questo tiPo di studi riguardano la possibilità di verificare, con esperimenti mirati, l'attendibilità dei modelli filogenetici proposti e di utilizzarli per identificare le specie a rischio di estinzione e tutelare la biodiversità della Terra.
"Ogni volta che un biologo cerca di sapere perché un organismo sia costituito o agisca in un determinato modo, deve ricorrere al metodo comparativo". Konrad Lorenz (1958)
Importanza della filogenesi
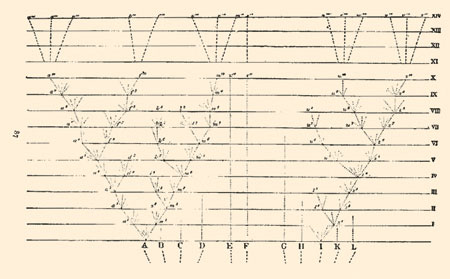

Nella sua opera (1859) The origin of species (trad. it. L'origine delle specie) Charles Darwin ha utilizzato solo un'illustrazione (fig. 1). Essa rappresenta per l'epoca un'immagine rimarchevole dal punto di vista teorico: mostrava un ipotetico 'albero' in cui la diversità degli organismi viventi era organizzata gerarchicamente: "Tutti gli esseri viventi si rassomigliano l'uno con l'altro in modo progressivo, così che è possibile classificarli in gruppi all'interno di altri gruppi. Evidentemente tale classificazione non è arbitraria come quella delle stelle nelle costellazioni". Il tipo di informazioni contenute in questi alberi è alla base di tutti gli studi comparativi del comportamento. La costruzione di un albero di relazioni evolutive comunemente prende il nome di filogenesi, permette di ricostruire i modelli storici di origine e le variazioni lungo la linea evolutiva. Tutti i metodi comparativi, sia nel contesto dello studio del comportamento sia in altre aree appartenenti al campo delle scienze della vita quali l'ecologia (Wanntorp et al., 1990) e la neurobiologia (Campbell, 1976), si basano su informazioni che provengono dalla sistematica filogenetica. Questa dipendenza è nuova, ma importante. l primi studiosi di comportamento animale compresero pienamente l'importanza delle filogenesi, sebbene all'epoca ne fossero disponibili poche. Infatti, K. Lorenz e N. Tinbergen si trovarono di fronte alla necessità non solo di raccogliere dati originali sul comportamento, ma anche di compiere un lavoro di tipo sistematico, nel tentativo di ricostruire gli alberi filogenetici (fig. 2) sulla base di tutte le informazioni disponibili (Burghardt e Gittleman, 1990). Oggigiorno, il campo della sistematica sta andando incontro a una rivoluzione, disponendo di nuove tecniche e di dati originali per la costruzione di alberi, ma anche utilizzando informazioni diverse provenienti dalla morfometria, dalla biologia molecolare e dalla statistica, che permettono descrizioni più dettagliate delle relazioni evolutive tra i taxa, che possono essere utilizzate per verificare ipotesi di evoluzione del comportamento.
La natura della filogenesi
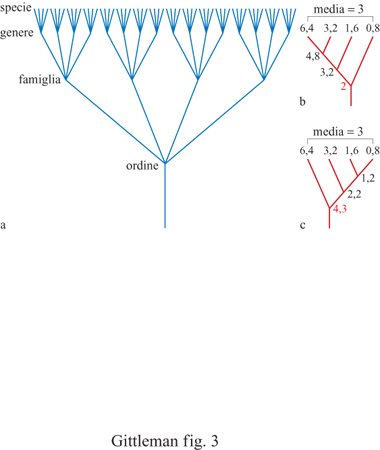
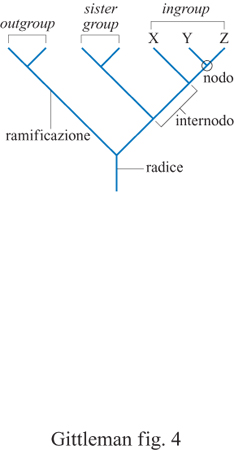
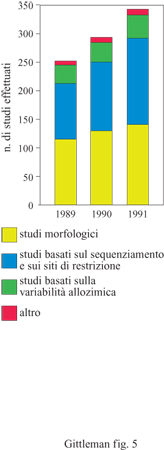
Le filogenesi contengono informazioni sui pro genitori comuni di diversi organismi; tale informazione è espressa come diagramma ramificato. È importante distinguere tra un sistema filogenetico di tassonomia e un sistema linneano. Quest'ultimo, il più comune fino a tempi recenti, fornisce informazioni sulle posizioni relative dei taxa all'interno di una struttura gerarchica nested, cioè a scatole cinesi (ranghi tassonomici come regno, phylum, classe e così via). Sebbene le categorie linneane possano essere utilizzate per dedurre informazioni filogenetiche, non necessariamente si rivelano utili a questo scopo. Se utilizzate per identificare la posizione di un taxon in una struttura gerarchica, esse possono non fornire informazioni o fornime di fuorvianti, circa l'antenato comune. Per esempio, un albero basato su una tassonomia linneana sarebbe tipicamente simmetrico con ramificazioni di uguale lunghezza tra i taxa. Questa è chiaramente una visione non realistica della storia evolutiva. La distinzione è quindi cruciale per i metodi comparativi nello studio del comportamento (fig. 3). Le questioni che coinvolgono l'origine evolutiva, la sua velocità e la sua direzione implicano il principio di una discendenza comune e sono influenzate direttamente dalla topologia dell'albero filogenetico. Gli alberi filogenetici forniscono specifici tipi di informazione. l nodi rappresentano antenati presuntivi; le estremità costituiscono la terminazione delle ramificazioni e tipicamente rappresentano la specie negli studi comparativi. Le ramificazioni degli alberi filogenetici sono le linee che connettono tra loro i nodi, o un nodo con l'estremità. Una radice è la base dell'albero, ovvero il suo ramo più ancestrale. l soggetti o i taxa di uno studio comparato vengono chiamati tecnicamente nel loro insieme ingroup, o gruppo di parentela, che deve contenere più di due taxa per essere in grado di inferire una relazione filogenetica. Il sister group, o gruppo affine, è il taxon più strettamente imparentato a un gruppo di parentela. Da ultimo, poiché è necessario avere un punto di partenza per fare un'inferenza su un ordine evolutivo, un outgroup, o gruppo estraneo, indipendente dal gruppo da studiare, viene di solito utilizzato per rappresentare il progenitore del gruppo di parentela (fig. 4). C'è ancora un ultimo punto da sottolineare: osservando un albero filogenetico è cruciale non accettare come dato di fatto una determinata visione della storia evolutiva; gli alberi filogenetici sono solo stime o ipotesi sulla filogenesi. Questa è il risultato di una serie di eventi storici che normalmente non conosciamo e non possiamo ricostruire. l metodi comparativi, che necessariamente si basano sulla filogenesi, saranno sempre vulnerabili per l'incertezza della ricostruzione storica. Fortunatamente, comunque, le verifiche sui reperti fossili e le verifiche analitiche ottenute con le simulazioni al calcolatore rendono sempre più attendibili le ipotesi filogenetiche (Purvis et al., 1994). Inoltre, il numero di filogenesi che vengono pubblicate sta aumentando in modo sbalorditivo (fig. 5).
Usi e abusi dei dati comparativi
Gli studi comparativi terminano e si prolungano nel tempo. Da una parte, i set di dati relativi a gruppi tassonomici ampi permettono di saggiare in linea generale un'ipotesi e costituiscono l'unica base solida per lo studio filogenetico dei moduli comportamentali. Dall'altra, la grande maggioranza degli studi comparativi si basa su dati raccolti in modo indipendente da ricercatori diversi, con metodi differenti e in condizioni differenti: i dati di cattiva qualità costituiscono il tallone di Achille della ricerca comparativa (Gittleman, 1989). Fortunatamente, con l'accumularsi dei dati, sia sugli alberi filogenetici sia sulla fonte dei dati comportamentali, le informazioni sono ora maggiormente verificate e più attendibili rispetto ai primi lavori. La replicazione di molti studi comportamentali su popolazioni diverse aggiunge validità alla serie di dati raccolti, specie quando i confronti vengono effettuati utilizzando i valori di un singolo carattere per una data specie. Inoltre, gli studi comparativi più recenti prevedono un numero crescente di test a priori e la verifica statistica, mediante simulazioni al calcolatore (Martins, 1996). Senza dubbio, gli studi comparativi moderni tengono conto degli ammonimenti di Tinbergen (1951), secondo i quali la pianificazione e l'attuazione di uno studio di questo tipo devono essere messi a punto con la stessa accuratezza di un qualsiasi altro esperimento (Gittleman e Decker, 1994). Un altro problema latente portato in evidenza dagli studi comparativi è che raramente sappiamo qualcosa sui meccanismi causali che determinano un risultato comparativo, né sappiamo se un saggio comparativo ci permetterà di distinguere tra gli effetti della selezione e quelli della deriva, della selezione indiretta o di una variazione incontrollata dell'ambiente (Leroi et al., 1994). Per verificare un'ipotesi adattativa in modo definitivo è necessario misurare direttamente nei taxa in studio la variabilità genetica, spesso mediante l'uso di procedure sperimentali specifiche. Negli studi comparativi che utilizzano numerosi caratteri su linee filogenetiche estese è raro che si possa compiere tale lavoro. Tuttavia, questo non è un limite del metodo in sé, ma è piuttosto una debolezza dovuta alla ridotta disponibilità di dati e alla mancanza di comprensione dei caratteri analizzati. Quanto più i dati comparativi si arricchiranno di informazioni sui meccanismi causali e di misure di ereditabilità del carattere in esame, tanto più il metodo comparativo diverrà efficace.
Nuovi metodi comparativi
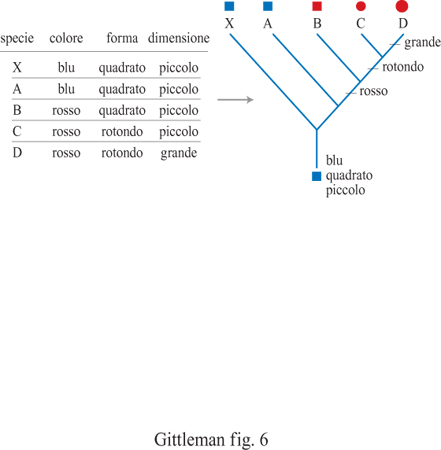
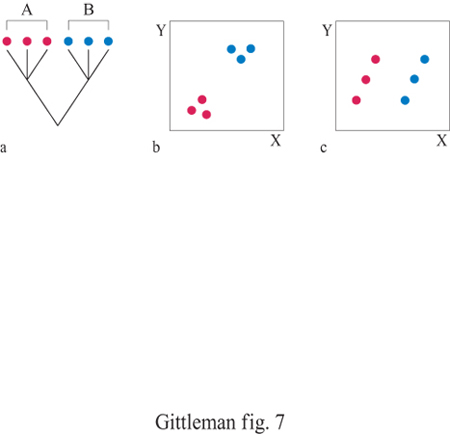
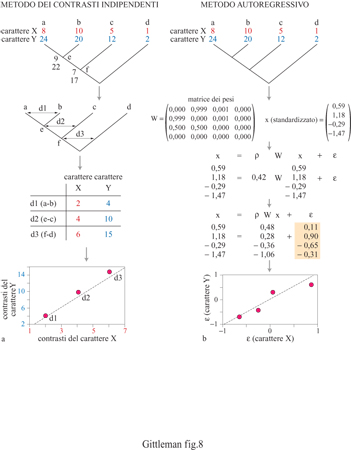
Il metodo comparativo, basato su confronti tassonomici incrociati per analizzare l'evoluzione di moduli comportamentali, risale almeno all'antica Grecia: Aristotele ha utilizzato comparazioni tra carnivori ed erbivori per descrivere le differenze di socialità (Ridley, 1983). Ma, a parte l'accumulo costante di dati sulle caratteristiche degli organismi, c'è realmente qualcosa di nuovo nel metodo comparativo dei nostri giorni? La risposta è senz'altro affermativa: due aspetti degli studi comparativi moderni differiscono sostanzialmente dai metodi utilizzati dai primi ricercatori. Il primo riguarda il metodo stesso. I metodi comparativi moderni sono più sistematici e quantitativi. In generale ci sono due tipi di metodi comparativi, a seconda della natura dei dati da analizzare: quelli di categoria e quelli quantitativi. I primi implicano essenzialmente la sovrapposizione alla filogenesi di caratteri di categoria o discreti (per esempio, il sistema di accoppiamento, come la poliginia oppure la monogamia). Determinando con precisione in quale forma e dove il carattere in esame è presente a livello dei differenti nodi e ramificazioni, è possibile identificarne l'origine evolutiva, il mantenimento, la sequenza delle modifiche e la variazione associata con altri caratteri. Per esempio, la distribuzione dei caratteri di categoria è in grado di mostrare se un carattere è ancestrale rispetto a un altro (fig. 6). Gli ostacoli che possono insorgere con i metodi applicati a categorie derivano dalla difficoltà di assegnare probabilità statistiche ai modelli comparativi e di studiare il grado di variazione del carattere in relazione alla lunghezza del ramo. l metodi quantitativi implicano test statistici di variazione associata tra caratteri quantitativi (per esempio, dimensione del gruppo sociale, dimensione della preda o frequenza di accoppiamento), una volta stabilite le relazioni filogenetiche tra i taxa considerati. Essenzialmente, i metodi comparativi quantitativi esaminano la variazione del carattere e, contemporaneamente, verificano la struttura filogenetica presente nei dati. Inizialmente, il motivo principale che ha fatto utilizzare l'approccio quantitativo era strettamente statistico: i test statistici standard, come la regressione, presuppongono l'indipendenza dei dati ma, frequentemente, le specie correlate non si sono evo Iute seguendo processi indipendenti, bensì condividendo alcuni caratteri con i progenitori comuni (fig. 7). Le variabili quantitative possono essere utilizzate per valutare statisticamente l'evoluzione corre lata lungo una linea filogenetica, utilizzando il metodo dei contrasti indipendenti o l'autoregressione (fig. 8). Anche se entrambi i metodi quantitativi sono statisticamente affidabili nella maggior parte delle condizioni, ciascuno fornisce assunzioni analitiche su come i tratti in esame si evolvano (per esempio, modelli specifici di evoluzione) e sul reale grado di correlazione dei caratteri con la filogenesi (Harvey e Pagel, 1991; Martins, 1996). L'altro aspetto decisivo nel campo dei metodi comparativi, in particolare per lo studio dei problemi del comportamento, è la disponibilità di filogenesi indipendenti dal carattere oggetto di studio. Tradizionalmente, gli alberi filogenetici sono basati su tratti morfologici come la forma del cranio o la dimensione dei denti. Dato che il comportamento spesso varia con la morfologia, ogni saggio comparativo del comportamento basato su una filogenesi morfologica corre il rischio di essere viziato da circolarità. La diffusione dei marcatori molecolari, per esempio allozimi o sequenze di DNA, per stimare relazioni filogenetiche potenzialmente interrompe questa circolarità (Gittleman et al., 1996). Il campo della sistematica molecolare è estremamente promettente per lo studio di numerose problematiche filogenetiche (Avise, 1994). Questi alberi molecolari costituiscono importanti 'ipotesi nulle' per la ricostruzione dei processi storici di evoluzione del comportamento.
Modelli di evoluzione del comportamento
lllustreremo ora alcuni esempi che forniscono un'idea di come i metodi comparativi moderni siano stati applicati a varie problematiche del comportamento. Enfasi particolare verrà data al tipo di studi più che al merito dei modelli di filogenesi del comportamento. Questi nuovi metodi comparativi sono troppo nuovi per poterci dire se l'evoluzione comportamentale di linee indipendenti mostri tendenze ripetitive. Ci sono molti lavori riassuntivi che illustrano l'approccio comparativo e filogenetico allo studio del comportamento (Miles e Dunham, 1993; Gittleman e Decker, 1994; Martins, 1996).
L'origine del comportamento

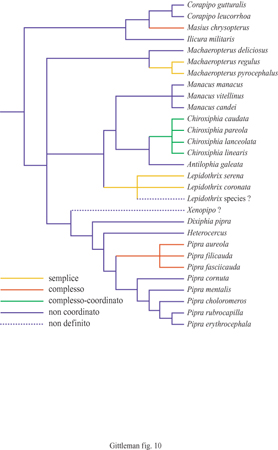
Uno dei problemi più complessi nello studio filogenetico del comportamento è che la fonte primaria dei dati è l'osservazione diretta. Ciò significa che, nel caso in cui i dati comportamentali non vengano raccolti come si presentano o non possano essere in qualche modo registrati, essi vengono persi per sempre. Avere a disposizione una filogenesi apre la prospettiva eccitante di poter ricostruire i modelli storici o le origini di un comportamento. Un esempio di come si possano utilizzare la filogenesi e il metodo comparativo per identificare le origini di un comportamento è lo studio del comportamento sociale dei manachini (Prum, 1994), uccelli neotropicali conosciuti per il dimorfismo sessuale del piumaggio, per il comportamento di aggregazione dei maschi nel confrontarsi davanti a una femmina e per gli elaborati moduli di corteggiamento del maschio. Durante l'accoppiamento, due o più maschi si esibiscono in una serie di moduli comportamentali, display coordinati, per mettersi in mostra (fig. 9). Questi moduli comportamentali, sulla base di una descrizione etologica dettagliata, possono essere classificati in tre categorie (fig. 10): display semplici, nei quali due o più maschi esibiscono versioni sincronizzate di moduli comportamentali normalmente eseguiti da un solo maschio; display complessi, modelli sincronizzati di esibizione attuati da più maschi e che non vengono mai esibiti da maschi soli; display complessi-coordinati, costituiti da un modulo complesso e coordinato esibito come forma di richiamo sessuale da più maschi con diverse probabilità di successo. La distribuzione filogenetica di questi comportamenti di esibizione, sovrapposta a una filogenesi, indica che la mancanza di coordinazione delle esibizioni è la condizione ancestrale e che ci sono cinque origini evolutive indipendenti nei diversi display dei manachini. Inoltre, l'origine delle esibizioni complesse è ricorrente, ossia esse si presentano in tutte le specie di una ramificazione durante il processo evolutivo. La sovrapposizione di dati comportamentali comparativi su un albero filogenetico mette in evidenza che i processi filogenetici del comportamento possono essere ampiamente ricostruiti. Ciò implica, tra l'altro, l'esistenza di una forte componente storica nell'evoluzione del comportamento, in contrasto con l'idea assai diffusa che il comportamento sia estremamente labile (v. oltre).
Sequenze filogenetiche del comportamento
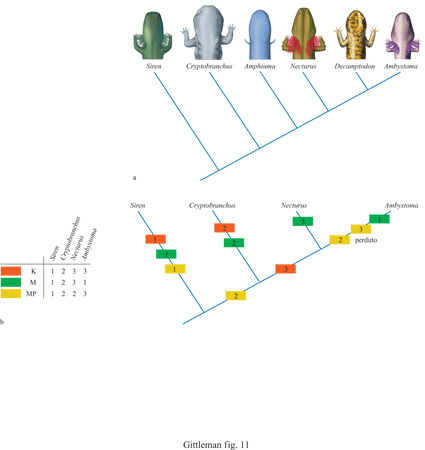
Sulla base di una filogenesi è possibile determinare l'origine di un modello comportamentale e la sequenza di modificazioni avvenute durante la sua storia evolutiva. Dato che il comportamento è così strettamente legato alla fisiologia e alla morfologia, è di particolare interesse studiare la correlazione tra i modelli di sequenze comportamentali e altri caratteri funzionali. Un elegante esempio di come l'analisi filogenetica possa essere applicata alla sequenza di cambiamenti di una determinata caratteristica è fornita da uno studio comparativo sulle salamandre (Lauder e Reilly, 1996). Sono stati raccolti dati dettagliati, tra loro correlati, sui caratteri comportamentali (cattura della preda) e morfologici (dimensione della mascella), e sui moduli motori (movimenti della testa e della mascella). Dato che nelle salamandre la cattura della preda è molto rapida, avendo una durata di 30÷120 ms, alcune di queste informazioni sono state ottenute in modo assai particolareggiato con un'analisi effettuata fotogramma per fotogramma. L'identificazione della variazione di questi caratteri lungo linee filogenetiche ha permesso di evidenziare diverse tendenze. Primo, taxa quali Ambystoma e Siren, simili morfologicamente, si differenziano nel comportamento. Ciò potrebbe essere dovuto a differenze nei moduli motori. Secondo, invece i taxa strettamente correlati Ambystoma e Necturus, che sono morfologicamente differenti, mostrano somiglianze nel comportamento. Quindi, al contrario di quanto si poteva prevedere, le differenze morfologiche non sono sempre legate a differenze di comportamento. Terzo, e forse più sorprendente, tutti e quattro i taxa analizzati mostrano caratteristiche evolutive indipendenti di comportamento, morfologia e moduli motori, indicando che si è verificata una divergenza filogenetica notevole in tutte e tre le classi di caratteri (fig. 11). Questo tipo di studio comparato mostra chiaramente la complessità dell'identificazione di modelli filogenetici in diversi caratteri. C'è anche un altro punto: se la filogenesi venisse ricostruita sulla base di uno di questi caratteri, sarebbe molto diversa da quella basata su un altro carattere. Ciò sottolinea l'importanza e la criticità della filogenesi nello studio comparativo.
Correlazione tra le variazioni del comportamento
La correlazione è una misura della relazione tra variabili. La difficoltà di calcolare la correlazione tra caratteri comparativi insorge nel caso in cui si voglia considerare anche il processo filogenetico che caratterizza i tratti in esame. I metodi comparativi quantitativi sopra menzionati, cioè i contrasti indipendenti e l'autoregressione, danno una soluzione a questo problema analitico e, in sostanza, assolvono allo stesso compito fornendo valori per i caratteri comparativi indipendenti dalla struttura filogenetica, e utilizzabili quindi per lo studio della correlazione tra i caratteri. Dato che i metodi comparativi sono importanti per determinare la filogenesi, è necessario anzitutto valutare se i caratteri in questione siano realmente correlati alla filogenesi. Ci sono numerose analisi statistiche che possono essere applicate in questi casi. Fondamentalmente, è necessaria una qualche misura del rapporto tra distanza filogenetica (o rango tassonomico) e variazione del carattere.
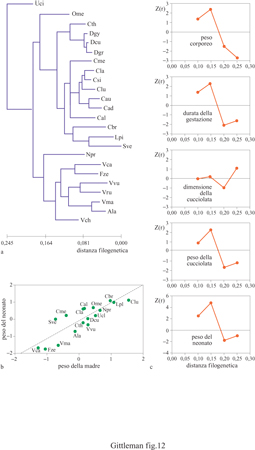
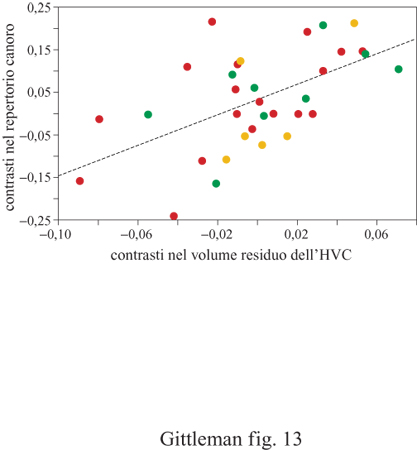
In uno studio sulla dimensione del gruppo sociale, sui modelli di ciclo vitale (come lunghezza della gravidanza, peso alla nascita) e sulla mole corporea dei canidi (fig. 12), è stata analizzata la correlazione tra ognuna di queste variabili e una filogenesi molecolare (Geffen et al., 1996). È risultato che non solo questi caratteri sono correlati in maniera significativa con la filogenesi, ma questo si verifica nella maniera attesa: le correlazioni sono maggiori per le specie più strettamente affini, riflettendo una maggiore somiglianza del carattere, mentre diminuiscono molto rapidamente per le specie più lontane. Oltre a permettere di valutare se i caratteri quantitativi sono correlati con la filogenesi, questa statistica descrittiva fornisce informazioni sulla relativa instabilità dell'evoluzione del carattere, come descritto di seguito. Una volta nota la correlazione filogenetica, il metodo dei contrasti indipendenti, o quello autoregressivo, può essere utilizzato per valutare la variazione associata dei caratteri. Per esempio, nello studio appena menzionato, l'autoregressione è stata applicata per scoprire se nei canidi l'evoluzione della dimensione del gruppo fosse correlata all'investimento energetico per la prole. Sebbene il peso dei neonati aumenti con la mole corporea della madre, la relazione è proporzionale: ciò non supporta l'ipotesi che la dimensione del gruppo si evolva per adeguarsi alla maggiore richiesta energetica imposta dalla maggiore mole dei figli. L'approccio dei contrasti indipendenti può essere utilizzato in modo analogo per studiare la correlazione tra variabili quantitative. Un esempio affascinante è costituito dall'applicazione di questo metodo in uno studio comparativo, insolitamente dettagliato, di neuroanatomia e comportamento (fig. 13). Il problema riguarda la relazione tra il canto degli uccelli e l'anatomia del cervello. I contrasti indipendenti sono stati calcolati in 41 specie di oscini (come luì, scriccioli e passeri) per l'ampiezza del repertorio canoro, una misura della complessità del canto, e per la dimensione relativa del centro vocale alto, un'area che ospita nuclei direttamente connessi alla siringe (De Voogd et al., 1993). L'analisi ha mostrato una stretta relazione tra repertorio canoro e dimensione del cervello, anche con punti differenti di ramificazione filogenetica, suggerendo che la relazione è esistita fin dall'origine evolutiva degli oscini. I metodi comparativi quantitativi sono abbastanza efficienti nella determinazione delle interrelazioni tra caratteri comportamentali e altri tratti funzionali. I futuri sviluppi di questi metodi riguarderanno l'analisi della varianza dell'errore del valore del carattere e problematiche di statistica multivariata.
La relativa instabilità del comportamento
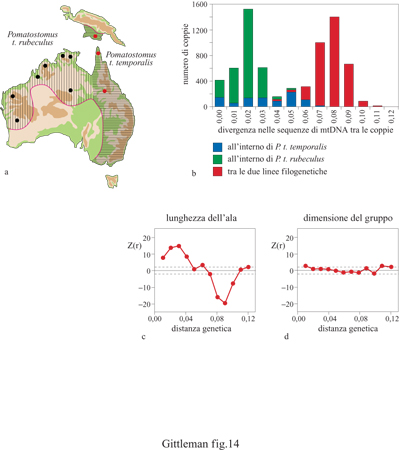
Una delle ragioni per cui il comportamento è stato utilizzato poco frequentemente per la costruzione di filogenesi o, più in generale, per studi che contemplano una prospettiva storica (per esempio, in paleontologia) è la sua relativa instabilità. Si è supposto cioè che se il comportamento cambia di generazione in generazione in risposta alle condizioni ambientali, allora lascerà poche tracce della storia filogenetica. Tuttavia non è dimostrato che in generale il comportamento sia caratterizzato da maggiore instabilità rispetto alla morfologia o alla fisiologia. Anche in questo caso, con alcune informazioni filogenetiche e con le stesse correlazioni statistiche descritte in precedenza nell'esempio sui canidi, è possibile studiare direttamente l'instabilità evolutiva del comportamento. Un lavoro sul garrulo dalla corona grigia, Pomatostomus temporalis (fig. 14), uccello canoro dell'Australia e della Nuova Guinea, che alleva la prole in modo cooperativo, ha fatto luce su questo problema (Edwards e Kot, 1995). Anzitutto è stata determinata in modo particolarmente preciso l'informazione filogenetica, specificando le distanze genetiche del DNA mitocondriale (mtDNA) di 120 individui di 12 popolazioni che rappresentavano linee evolutive distinte (v. figura 14b). Di tutti i soggetti sperimentali è stata misurata la lunghezza dell'ala, e ciascun individuo è stato assegnato a uno di 53 diversi gruppi sociali (v. figura 14c). Sono state poi calcolate le correlazioni tra la distanza filogenetica delle popolazioni e ciascuno di questi caratteri comportamentali (v. figura 14d). Come atteso, sono state rilevate significative correlazioni positive con la lunghezza dell'ala tra gli individui che differivano per meno del 5% nell'mtDNA, mentre la lunghezza dell'ala era correlata negativamente negli individui che differivano del 7÷11 % nell'mtDNA. Invece, non è stata osservata alcuna relazione tra la distanza filogenetica e la dimensione del gruppo. Anche un altro studio indipendente, condotto su otto gruppi di mammiferi non strettamente imparentati (dalla famiglia in su), ha messo in evidenza che i comportamenti costituiscono un segnale filogenetico relativamente piccolo (Gittleman et al., 1996), confermando la presunta maggiore instabilità del comportamento rispetto alla morfologia, sia a livelli tassonomici più bassi che a quelli più alti. Lo studio sul garrulo dalla corona grigia pone un altro punto interessante: in che misura sia possibile identificare i modelli filogenetici al di sotto del livello di specie. Queste ricerche sembrano supportare l'idea che il comportamento sia instabile. Altri lavori mostrano però che la questione dell'instabilità relativa dei caratteri può essere più complessa (de Queiroz e Wimberger, 1993). Tra taxa che vanno dalle vespe alle lucertole sono state costruite filogenesi utilizzando un ampio ambito di caratteri, tra cui quelli comportamentali (per esempio, comportamento alimentare, di costruzione del nido e di accoppiamento) e quelli morfologici (per esempio, peso del corpo e lunghezza del becco). In ciascun taxon la filogenesi comportamentale e quella morfologica sono statisticamente indistinguibili; in altre parole, per alcuni taxa i modelli comportamentali sono altrettanto conservati quanto i tratti morfologici. Queste ricerche, considerate nel loro insieme, mettono in luce il fatto che sappiamo veramente poco del perché alcuni caratteri comportamentali in certi taxa sembrano labili, mentre altri sono relativamente fissati lungo le linee filogenetiche. Ora che gli strumenti comparativi sono stati messi a punto in modo affidabile, il lavoro futuro potrà chiarire queste tendenze. Ciò è di fondamentale importanza in quanto ci permetterà di determinare se differenti tipi di caratteri fenotipici, inclusi quelli comportamentali, possono essere utilizzati nella sistematica. Inoltre, si pone la questione di quanto il comportamento costituisca una forza trainante nell'evoluzione. Come scrisse l'autorevole biologo evolutivo, Ernst Mayr: "Uno slittamento verso una nuova nicchia o zona adattativa è avviato, quasi senza eccezioni, da una variazione del comportamento" (1963). La ricerca comparativa moderna può fmalmente dirimere questa questione.
Velocità di evoluzione del comportamento
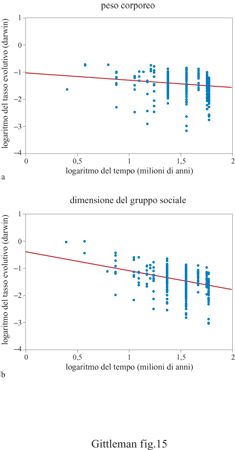
Il comportamento deve evolversi più rapidamente di altri caratteri. Gli individui modificano il loro comportamento di momento in momento, mentre altri tratti quali la dimensione del cervello o il tasso metabolico sono relativamente costanti. Così, ci si potrebbe attendere che la velocità di evoluzione del comportamento sia più alta di quella di altri caratteri, specialmente se misurati su ampie scale temporali (filogenetiche), come nel caso dei confronti tra taxa diversi. È sorprendente che finora pochi studi abbiano verificato questa idea; probabilmente ciò è dovuto al fatto che qualsiasi analisi comparativa richiede informazioni dettagliate e accurate sugli intervalli di tempo tra gli eventi di speciazione (per esempio, i nodi degli alberi). Nuove filogenesi, specialmente quelle molecolari, hanno fornito informazioni sulla storia evolutiva indipendentemente dai caratteri fenotipici e hanno anche identificato defmite finestre temporali (Harvey et al., 1996). Da un punto di vista analitico, ci sono molte grandezze che permettono di misurare la velocità evolutiva (Gingerich, 1993). Una delle prime fu il darwin, proposto da J.B.S. Haldane (1949), usato originariamente per misurare la velocità di evoluzione dei cavalli, e più recentemente, di una serie di organismi: dalla variazione di popolazione nelle lumache di mare, al tempo di generazione dei ratti di laboratorio (Gingerich, 1993). Essenzialmente, questa misura calcola la velocità di evoluzione come l'entità della variazione di un carattere tra il valore iniziale e quello finale, in un lasso di tempo in una determinata finestra temporale. Poiché le velocità evolutive cambiano con la scala temporale (la velocità evolutiva sembra inversamente proporzionale a essa), i darwin sono di solito rappresentati graficamente, in scala logaritmica, in funzione degli intervalli di tempo, ottenuti da distanze filogenetiche e calibrati per ciascun gruppo tassonomico. Questi diagrammi, detti diagrammi dei darwin (fig. 15), mostrano due misure statistiche del tasso evolutivo: il tasso evolutivo netto fornito dalla pendenza della retta e il tasso evolutivo istantaneo per brevi intervalli di tempo, fornito dall'intercetta.
Un recente esempio dell'uso dei darwin per confrontare le velocità evolutive di caratteri comportamentali e morfologici avvalora l'idea che il comportamento si evolva più rapidamente: confrontando i diagrammi dei darwin riportati in figura 15, si può osservare che, nel caso dei primati, la retta relativa alla dimensione del gruppo ha una pendenza e un'intercetta maggiori rispetto a quella relativa al peso corporeo (in questo caso è stata usata una filogenesi molecolare per determinare le relazioni evolutive). Lo stesso dato è stato rilevato per molti gruppi di mammiferi. (Gittleman et al., 1996). Quindi, l'uso di una misura classica della velocità di evoluzione congiunto alla moderna filogenesi molecolare consente alle analisi comparative di affrontare problematiche considerate irrisolvibili fino a epoca recente. Quest'ultimo esempio non utilizza il metodo comparativo per sé, come nei casi precedenti, ma rimane su un terreno comune: la filogenesi, utilizzata, in questo caso, per qualcosa di più rispetto alla deduzione di una discendenza evolutiva, cioè per fornire precisi intervalli di tempo in cui si è verificato il processo evolutivo. Quando le informazioni temporali diverranno più affidabili, gli studi comparativi sui tempi e sulle velocità evolutive saranno più comuni.
Nuove direzioni
Gli studi comparativi saggiano i processi evolutivi del comportamento con accuratezza e rigore. Un'opportuna combinazione delle informazioni comparative su numerosi caratteri di organismi con alberi filogenetici precisi e di metodi statistici rigorosi permette verifiche dettagliate delle diverse ipotesi. In particolare, gli approcci comparativi consentono di esaminare in modo efficace l'origine, la velocità, l'instabilità e la variazione evolutiva di modelli comportamentali. Questo progresso è di grande interesse e sarebbe una piacevole sorpresa anche per Darwin vedere le molteplici applicazioni del pensiero filogenetico. Tutte queste problematiche sono di carattere storico, sostanzialmente di ricostruzione del passato. È lecito però chiedersi se i metodi comparativi possano essere utilizzati per affrontare anche questioni legate al presente (sperimentazione) e magari anche per predire il futuro (conservazione). Vi sono due promettenti linee di ricerca a riguardo. La prima affronta una debolezza tradizionale degli studi comparativi: l'assenza di prove sperimentali. Gli studi filogenetici possono includere controlli sperimentali e manipolazioni così da riportare generiche correlazioni a più specifici meccanismi causali? Uno studio sui pesci gasterosteidi (spinarello) descrive un modo per applicare la filogenesi a un contesto sperimentale (McLennan, 1991). Gli spinarelli hanno una colorazione nuziale molto variabile, che va dall'assenza di colore all'oro, rosso, blu e nero. La filogenesi di 6 taxa di spinarello suggerisce che l'origine e l'elaborazione delle colorazioni nuziali sono correlate a interazioni maschio/ maschio, cure parentali, e interazioni maschio/femmina. Sono stati effettuati esperimenti di scelta sessuale in cui le femmine dello spinarello (Gasterosteus aculeatus) sceglievano il maschio in base al colore, alle condizioni di allevamento o alle affinità comportamentali. L'osservazione ha mostrato in modo chiaro che le femmine, fondamentalmente, scelgono in base al colore del corpo più che ai modelli di cura parentale o alle interazioni maschio/maschio, suggerendo che i modelli filogenetici osservati sono legati alle interazioni maschio/femmina e che molto probabilmente la scelta della femmina si basa sull'intensità del colore del maschio. I modelli filogenetici possono quindi essere saggiati sperimentalmente.
La seconda linea di ricerca lungo la quale ci si sta muovendo riguarda la possibilità di intervenire sugli eventi futuri. Per conservare la biodiversità sulla Terra è necessario identificare in maniera efficace le specie a rischio di estinzione e proteggere quelle con caratteristiche che potrebbero renderle vulnerabili in futuro. Gli studi comparativi possono fornire un aiuto diretto in due modi (Brooks et al., 1992). Anzitutto, gli alberi filogenetici possono essere utilizzati per identificare quali specie sono più insolite di altre. Specie come il panda minore (Ailurus fulgens) o il panda maggiore (Ailuropoda melanoleuca), che sono le uniche due specie esistenti di una sotto famiglia tassonomica (ailurini), sono ovviamente più caratterizzanti rispetto a gruppi di carnivori con molte specie come canidi o felidi. Tale dato da solo non si traduce in un'azione più efficace per proteggere queste specie; ciononostante, un simile uso della filogenesi può costituire un criterio importante per decidere quali specie devono essere immediatamente studiate e, di conseguenza, per capire i possibili problemi. Per esempio, dettagliati studi comportamentali comparativi hanno messo in luce alcune caratteristiche del panda maggiore significativamente differenti da quelle di altri carnivori, che ne limitano la capacità di adattamento a condizioni ambientali variabili (Gittleman, 1994); in particolare, è stato evidenziato un lento sviluppo neonatale sia per l'incremento di peso sia per la maturazione comportamentale associata. Altri caratteri come il tipo di sistema di accoppiamento, le preferenze alimentari o i livelli di aggressività possono mettere in evidenza problemi futuri. In secondo luogo, poiché molte caratteristiche seguono linee filogenetiche, se è a rischio di estinzione una specie relativamente poco conosciuta all'interno di un genere con molte altre descritte in modo più dettagliato, allora le caratteristiche di queste ultime possono essere utilizzate come previsione a breve termine delle caratteristiche a rischio. Dunque i metodi comparativi non sono soltanto in grado di ricostruire il passato, anche sulla base di caratteri complessi come i modelli comportamentali; essi possono essere utilizzati anche per prevedere il futuro.
Bibliografia citata
AVISE, J.C. (1994) Molecular markers, natural history and evolution. New York, Chapman & Hall.
BROOKS, D.R., MAYDEN, R.L., McLENNAN, D.A. (1992) Phylogeny and biodiversity: conserving our evolutionary legacy. Trends Ecol. Evol., 7, 55-59.
BURGHARDT, G.M., GITTLEMAN, J.L. (1990) Comparative behavior and phylogenetic analyses: new wild, old bottles. In Interpretation and expIanation in the study of animaI behavior, a c. di Bekoff M., Jamieson D., Boulder, Westview Press, pp. 192-225.
CAMPBELL, C.B.G. (1976) What animals should we compare? In Evolution, brain, and behavior, a c. di Masterton R.B., Hodos W., Jerison H., New York, John Wiley & Sons., pp. 107-113.
DARWIN, CH. (1859) On the origin of species by means of natural selection; or, the preservation of Javored races in the struggle for Iife. Londra, John Murray. Trad. it. L'origine delle specie, Bologna, Zanichelli, 1982.
DE QUEIROZ, A., WIMBERGER, P.H. (1993) The usefulness of behavior for phylogeny estimation: levels of homoplasy in behavioral and morphological characters. Evolution, 47, 46-60.
DEVOOGD, T.J., KREBS, J.R., HEALY, S.D. PURVIS, A. (1993) Relations between song repertoire size and the volume of brain nuclei related to song: comparative evolutionary analyses amongst oscine birds. Proc. R. Soc. Lond., 254, 75-82.
EDWARDS, S.V., KOT, M. (1995) Comparative methods at the species level: geographic variation in morphology and group size in grey-crowned babblers (Pomatostomus temporalis). Evolution, 49,1134-1146.
GEFFEN, E., GOMPPER, M.E., GITTLEMAN, J.L., LUH, H.-K., MACDONALD, D.W., W AYNE, R.K. (1996) Size, life-history traits, and social organization in the Canidae: a reevaluation. Am. Nat., 147, 140-160.
GINGERICH, P.D. (1993) Quantification and comparison of evolutionary rates. Am. J. Sci., 293-A, 453-478.
GITTLEMAN, J.L. (1989) The comparative approach in ethology: aims and limitations. In Perspectives in ethology, voI. 8, a c. di Bateson P.P.G., Klopfer P.H., New York, Plenum Press, pp. 55-83.
GITTLEMAN, J.L. (1994) Are the pandas successful specialists or evolutionary failures? Bioscience, 44, 456-464.
GITTLEMAN, J.L., ANDERSON, C.G., KOT, M., LUH, H.-K. (1996) Phylogenetic lability and rates of evolution: a comparison of behavioral, morphological and life history traits. In PhyIogenies and the comparative method in animaI behavior, a c. di Martins E., Oxford, Oxford University Press, pp. 166-205.
GITTLEMAN, J.L., DECKER, D.M. (1994) The phylogeny of behavioI. In Behaviour and evolution, a c. di Slater P.J.B., Halliday T.R., Cambridge, Cambridge University Press, pp. 80-105.
HALDANE, J.B.S. (1949) Suggestions as to quantitative measurement of rates of evolution. Evolution, 3, 51-56.
HARVEY, P.H., BROWN, A.J.L., MAYNARD SMITH, J., NEE, S., a c. di (1996) New usesJor new phyIogenies. Oxford, Oxford University Press.
HARVEY, P.H., PAGEL, M.D. (1991) The comparative method in evolutionary biology. Oxford, Oxford University Press.
LAUDER, G.V., REILLY, S.M. (1996) The mechanistic bases of behavioral evolution: a multi variate analysis of musculoskeletal function. In PhyIogenies and the comparative method in animaI behavior, a c. di Martins E., Oxford, Oxford University Press, pp. 104-137.
LEROI, A.M., ROSE, M.R., LAUDER, G.V. (1994) What does the comparative method reveal about adaptation? Am. Nat. 143, 381-402.
LORENZ, K. (1958) The evolution of behavioI. Sci. Am. 199, 67-78.
McLENNAN, D.A. (1991) Integrating phylogeny and experimental ethology: from pattem to process. Evolution, 45, 1773-1789.
MARTINS, E.P., a c. di (1996) PhyIogenies and the comparative method in animaI behavior. Oxford, Oxford University Press.
MAYR, E. (1963) AnimaI species and evolution. Cambridge, Mass., Harvard University Press.
MILES, D.B., DUNHAM, A.E. (1993) Historical perspectives in ecology and evolutionary biology: the use of phylogenetic comparative analyses. Ann. Rev. Ecol. Syst., 24, 587-619.
PRUM, R.O. (1994) Phylogenetic analysis of the evolution of alternative social behavior in the manakins (A ves: Pipiridae). Evolution, 48, 1657-1675.
PURVIS, A., GITTLEMAN, J.L., LUH, H.-K. (1994) Truth or consequences: the effects of phylogenetic accuracy on two comparative methods. J. Theor. Biol., 167, 293-300.
RIDLEY, M. (1983) The Explanation of organic diversity. Oxford, Clarendon Press.
TINBERGEN, N. (1951) The study ofinstinct. Oxford, Oxford University Press. (trad. it. Lo studio dell'istinto. Milano, Adelphi, 1994).
WANNTORP, H.-E., BROOKS, D.R., NILSSON, T., NYLIN, S., RONQUlST, F., STEARNS, S.C., WEDELL, N. (1990) Phylogenetic approaches in ecology. Oikos, 57, 119-132.
Bibliografia generale
BROOKS, D.R., McLENNAN, D.A. Phylogeny, ecology, and behavior. Chicago, University of Chicago Press, 1991.
EGGLETON, P., VANE- WRIGHT, R.I., a c. di, Phylogenetics and ecology. Londra, Academic Press, 1994. HARVEY, P.H., BROWN, A.J.L., MAYNARD SMITH, J., NEE, S., a c. di, New uses for new phylogenies. Oxford, Oxford University Press, 1996.
HARVEY, P.H., PAGEL, M.D. The comparative method in evolutionary Biology. Oxford, Oxford University Press, 1991.
MARTINS, E.P., a c. di, Phylogenies and the comparative method in animaI behavior. Oxford, Oxford University Press, 1996.
RIDLEY, M. The explanation of organic diversity. Oxford, Clarendon Press, 1983.