
Navigazione animale
Navigazione animale
Nel corso dell'evoluzione la capacità di spostarsi da un luogo a un altro, fondamentale per la sopravvivenza della maggior parte degli animali, si è diversificata per far fronte alle esigenze degli ambienti che venivano di volta in volta colonizzati. La capacità di navigazione è frequente tra gli Artropodi e i Vertebrati, che coprono lunghe distanze per sfuggire periodicamente a condizioni climatiche sfavorevoli, raggiungere i luoghi più adatti per la riproduzione, cercare sorgenti di cibo su ampie superfici o anche tornare ai luoghi di riposo notturno o diurno. Molte specie sono giunte a coprire distanze considerevoli, sia in terra che in aria o in acqua, e nello stesso tempo hanno acquisito meccanismi di orientamento spaziale che vengono, nell'insieme, classificati come 'navigazione animale'. Con questa locuzione si intende la capacità di un animale di raggiungere una meta spazialmente definita e circoscritta, anche relativamente lontana. Di solito, nella vasta letteratura sull'argomento, non si includono tra i fenomeni di navigazione gli spostamenti verso un traguardo che, già alla partenza, può essere individuato attraverso un contatto sensoriale diretto.
I movimenti migratori, con cui molti animali raggiungono mete specifiche, sono oggetto di ricerche relative alle loro caratteristiche spaziotemporali e ai processi che li governano. Tra i meccanismi di navigazione sono d'importanza capitale le 'bussole biologiche', che elaborano informazioni di origine astronomica o desunte dal campo magnetico terrestre, e le 'mappe mentali', relative alla disposizione dei punti di riferimento. Moltissime specie animali possiedono organi di senso specializzati per la rilevazione di informazioni fisiche utili per l'orientamento, le quali variano dal gradiente geomagnetico alla luce polarizzata fino alle sensazioni propriocettive generate dal movimento stesso. Nei Vertebrati, e soprattutto nei Mammiferi superiori, vi sono anche strutture cerebrali specialmente deputate alla sintesi di tali informazioni sensoriali; grazie a tali strutture neurali (prima fra tutte, la formazione ippocampale), gli animali conservano in memoria, e quindi possono riconoscere, i luoghi già visitati in precedenza e programmare lunghi spostamenti da un luogo all'altro.
Il raggiungimento della meta può essere caratterizzato da tattiche di ricerca casuale o sistematica, oppure dipendere da informazioni, di origine genetica o acquisite, relative alle direzioni da tenere e alle distanze da coprire. La capacità di ritornare al luogo di partenza può basarsi su informazioni assunte all'andata o prescindere da esse. In quest'ultimo caso si parla di vera navigazione se l'animale sa calcolare la propria posizione rispetto all'obiettivo e lo raggiunge con l'ausilio di una o più bussole biologiche. Il calcolo della posizione può avvenire in base a una mappa mentale, comprendente i soli luoghi noti o ulteriormente estesa per un processo di acquisizione di informazioni a distanza. I termini impiegati per descrivere fenomeni e meccanismi della navigazione animale sono tratti dall'esperienza umana, sicché si parla di navigazione, mappa, bussola e inversione di rotta. Nella grande maggioranza dei casi i meccanismi di navigazione sono usati per raggiungere un luogo o un'area già noti all'animale; si parla allora di homing, o 'ritorno a casa', anche se non si tratta del ritorno a una residenza abituale.
Metodi di studio
Uno dei più usati metodi di studio della navigazione animale è il sistema di cattura-contrassegno-ricattura, cioè l'applicazione su animali catturati di contrassegni individuali, quali anelli, targhette, tatuaggi, o anche transponder magnetici impiantati sottocute, e il successivo ritrovamento dei soggetti che li portano. Tuttavia questo metodo è poco efficiente, perché la percentuale degli esemplari risegnalati di solito è molto bassa. Per gli animali migratori i dati che si ottengono riguardano per lo più l'ubicazione della località di partenza e di arrivo, talvolta anche dei luoghi di sosta, mentre altre informazioni sulle rotte seguite e i tempi impiegati rimangono sconosciute. Altri metodi sono l'osservazione diretta, i rilevamenti radar sugli Uccelli in migrazione, le registrazioni idrofoniche dei Cetacei e la telemetria basata sulla ricezione di ultrasuoni o di segnali radio emessi da trasmittenti fissate sugli animali. Di largo impiego è la telemetria satellitare che usa stazioni radioriceventi montate su satelliti orbitanti e capaci di calcolare la posizione geografica dell'emittente. Recentemente si è diffuso il metodo di applicare agli animali apparecchi registratori che immettono in memorie elettroniche i dati relativi alla direzione di moto e, talora, anche parametri fisici ambientali (temperatura, pressione, luminosità) e fisiologici (attività, frequenza cardiaca, temperatura corporea). Se all'apparecchio registratore è collegato un apparecchio GPS (Global position system), possono essere registrate, a brevi intervalli di tempo, anche la latitudine, la longitudine e l'altitudine a cui si trova l'animale e si possono correlare i suoi movimenti con informazioni ambientali e fisiologiche.
Infine, certi animali tenuti in cattività mostrano l'intenzione di fuggire verso direzioni determinate. Anche la capacità di compensare i dislocamenti prodotti dall'uomo, presente in animali migratori e non, è stata impiegata come metodo d'indagine. Sfruttando questi fenomeni in situazioni sperimentali, è stato raccolto un gran numero di dati relativi a soggetti tenuti per lo più in arene o gabbie circolari, nelle quali i movimenti sono registrati con vari metodi. In queste condizioni è possibile manipolare le informazioni spaziali, per esempio modificando il campo magnetico, per identificare i riferimenti necessari all'animale per orientarsi.
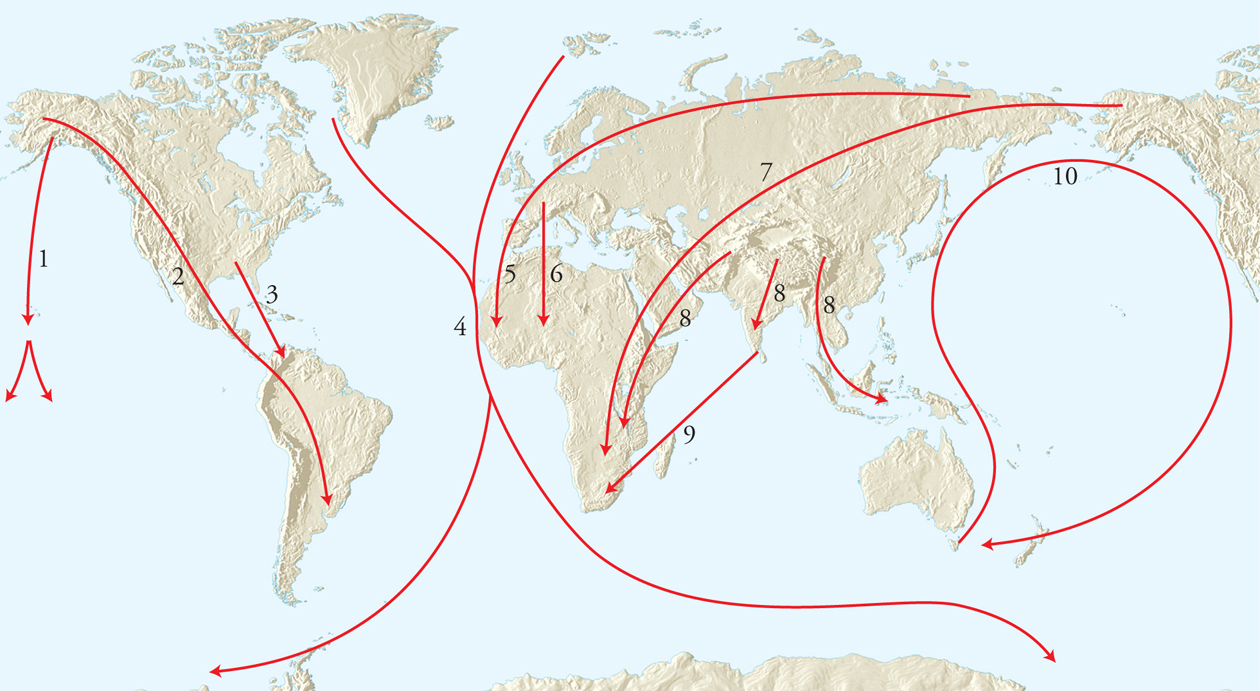
Il maggior numero di ricostruzioni degli spostamenti su grandi distanze è stato ottenuto studiando principalmente i volatili (fig. 2), ma anche animali terrestri o acquatici. Le rotte sono condizionate da fattori geomorfologici e dalle caratteristiche fisiologiche ed ecologiche della specie: certi Uccelli evitano di sorvolare catene montuose o ampi bracci di mare, mentre altri volano a lungo sul mare, non solo per raggiungere le isole oceaniche, ma anche per abbreviare il percorso e sfruttare venti favorevoli.
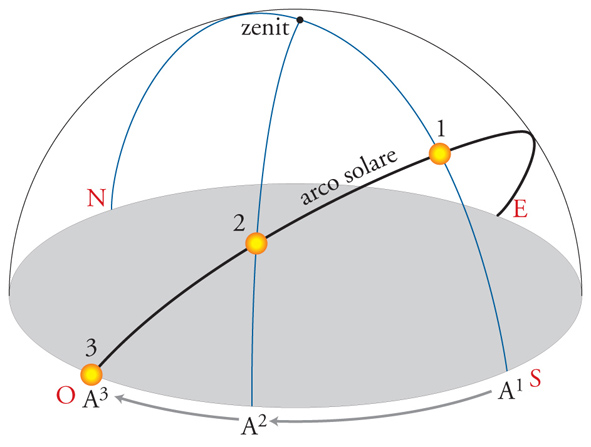
La navigazione animale presuppone un processo attivo di elaborazione di informazioni di varia origine, cui seguono scelte direzionali. Queste sono basate sulla capacità di determinare le direzioni geografiche, per mezzo delle cosiddette bussole biologiche, e sul possesso di rappresentazioni mentali, le mappe cognitive, relative ad aree più o meno estese. Tra gli animali è molto comune un meccanismo di orientamento attraverso il Sole che va sotto il nome di 'bussola solare'. L'animale, per prendere una direzione determinata, assume un angolo di orientamento con l'astro, e poiché la posizione del Sole varia nel corso del giorno, l'animale deve conoscerne la posizione rispetto alle direzioni cardinali alle varie ore del giorno. L'informazione dell'ora gli viene fornita da un ritmo biologico circadiano, detto orologio biologico, sincronizzato in base all'alternanza giorno-notte. Un'apparente complicazione è rappresentata dal fatto che l'animale non si orienta rispetto alla posizione del Sole in cielo, ma rispetto alla sua proiezione sull'orizzonte, cioè al suo azimut. L'azimut solare però varia con velocità diversa a seconda dell'ora del giorno, della stagione e della latitudine. Si noti, per esempio, che nella fig. 3 il tempo impiegato dall'azimut del Sole per passare da A1 a A2 è uguale a quello da A2 ad A3 e quindi che la sua velocità di variazione è stata diversa nei due tratti. Ancora poco conosciuto è il processo che permette agli animali di tener conto delle variazioni della velocità con cui varia l'azimut solare.
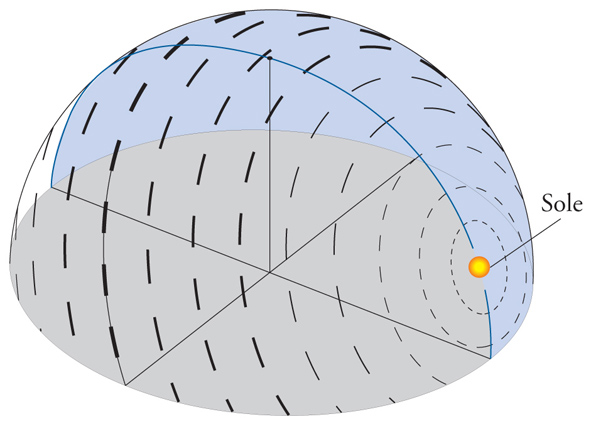
Un'ulteriore fonte di calibrazione della bussola solare rispetto ai punti cardinali è fornita dalla capacità di molte specie animali (come Uccelli, api e formiche) di percepire la percentuale di polarizzazione e l'orientamento del piano di polarizzazione della luce proveniente dalla volta celeste. Infatti, questi parametri variano regolarmente e simmetricamente nel cielo, a partire dalla posizione del Sole, il quale determina due emicalotte simmetriche di luce polarizzata, separate dal meridiano e dall'antimeridiano solare, e quindi dall'azimut del Sole. Questo fatto è esemplificato nella fig. 4, che contiene una rappresentazione schematica della polarizzazione della luce nella volta celeste. La direzione di polarizzazione è fornita dalla direzione delle barre che sono disposte concentricamente rispetto al Sole, rappresentato a un'altezza di 24° sull'orizzonte. In alcune specie di Passeriformi è stata provata l'esistenza di un meccanismo di orientamento, probabilmente molto diffuso tra gli Uccelli che migrano di notte, basato sulle stelle, che ha preso il nome di 'bussola stellare'. La dimostrazione della capacità di orientamento stellare è relativamente agevole perché questi animali, anche se chiusi in gabbie circolari, nelle notti serene tentano di fuggire nella direzione di migrazione; se vengono studiati in un planetario, invece che sotto il cielo stellato, la direzione dei tentativi di fuga può essere fatta variare a piacere, ruotando la volta stellata. L'orientamento stellare appare basato sul riconoscimento delle costellazioni, e in particolare di quelle circumpolari. Il fringillide nordamericano Passerina cyanea rimane ben orientato nel planetario, anche se singole stelle, compresa quella polare, vengono oscurate. Se però vengono 'spente' sia la stella polare sia le stelle che si trovano a meno di 35° da essa, allora gli uccelli sono incapaci di orientarsi. Gli esperimenti con il planetario sono serviti anche a dimostrare che la configurazione delle costellazioni e la loro disposizione intorno al Polo Nord celeste viene appresa, nelle prime settimane di vita, attraverso l'osservazione dell'apparente rotazione della volta celeste. Uccelli tenuti fin dalla nascita in un planetario con le stelle in rotazione imparano che la direzione nord è indicata dalla stella intorno a cui ruotano le costellazioni, qualunque essa sia; se, invece, le stelle del planetario restano immobili, essi non imparano a orientarsi.
Il campo magnetico terrestre costituisce una sorgente d'informazione direzionale per molti animali. Quando in laboratorio la manipolazione sperimentale del campo magnetico induce variazioni prevedibili dell'orientamento di un animale, se ne deduce che l'animale in esame è in grado di stabilire l'orientamento del campo magnetico ed è lecito affermare che possiede un equivalente biologico della bussola magnetica. In molti altri casi la capacità di orientamento magnetico è solamente sospettata per il fatto che gli animali possono mantenere una rotta costante anche in apparente assenza di altri fattori orientanti, come nel caso degli Uccelli migratori notturni che continuano a volare nella direzione corretta sotto una spessa coltre di nubi. Gli esperimenti più significativi sull'orientamento magnetico sono stati condotti in laboratorio, su Passeriformi che migrano di notte. L'osservazione prolungata di molti esemplari, tenuti in gabbie circolari e privati di altri riferimenti visivi, consente di stabilire la direzione statisticamente preferita con un'approssimazione accettabile e di constatare gli effetti indotti dalle manipolazioni del campo magnetico. Nella maggior parte dei casi la bussola biologica risulta funzionare come una bussola a inclinazione. L'animale infatti non appare capace di distinguere il senso delle linee di forza del campo magnetico (che siamo soliti rappresentare con un vettore), ma solo la loro inclinazione rispetto alla superficie terrestre. L'inclinazione del vettore è nulla all'equatore magnetico, ma va crescendo in senso opposto verso i poli: mentre l'estremità del vettore che indica il nord punta in basso nell'emisfero boreale, in quello australe è volta verso l'alto. Gli animali sensibili esclusivamente all'inclinazione del vettore non possono orientarsi nella zona dell'equatore magnetico.
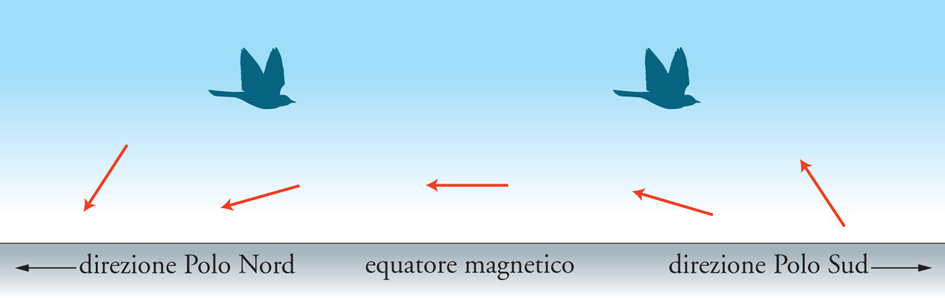
Se consideriamo il caso di Uccelli che in autunno si trasferiscono dall'emisfero boreale a quello australe, osserviamo che inizialmente possono orientarsi verso sud, andando nella direzione in cui il vettore magnetico punta in alto (fig. 5). Arrivati però all'equatore magnetico, devono ricorrere a bussole non magnetiche per proseguire verso sud, finché non trovano di nuovo un grado di inclinazione magnetica sufficiente per orientarsi. Se continuassero a volare nella direzione in cui il vettore punta verso l'alto, tornerebbero indietro verso nord. La necessaria inversione di orientamento sembra venga automaticamente indotta dal passaggio attraverso l'equatore magnetico.
La coesistenza di più tipi di bussola nello stesso animale pone il problema della loro importanza relativa e delle eventuali calibrazioni reciproche. Negli Uccelli migratori la navigazione è basata sull'interazione reciproca di più bussole. Il sistema è caratterizzato da una plasticità sufficiente per l'adattamento alle varie situazioni nel corso del viaggio migratorio, relative ai parametri sia magnetici (inclinazione, declinazione, intensità) sia astronomici (costellazioni visibili e caratteristiche del corso del Sole a latitudini diverse, copertura del cielo, e così via).
Per esempio, il passero delle savane, Passerculus sandwichensis, un migratore notturno, in autunno si sposta dal Canada e dagli Stati Uniti centro-settentrionali per svernare negli Stati Uniti meridionali e nella parte settentrionale dell'America Centrale. Uccelli catturati in natura a pochi giorni dalla nascita e allevati in assenza di riferimenti visivi importanti, ma tenuti a un ciclo normale di luce e buio, in laboratorio tentano di spostarsi secondo la giusta direzione di migrazione servendosi del campo magnetico terrestre. La direzione presa con questa bussola magnetica primaria può essere modificata se entra in conflitto con informazioni di origine celeste: infatti, quando vengono esposti a un cielo notturno, naturale o artificiale, i passeri deducono, in base alla rotazione del cielo, la direzione del nord e ciò serve loro a ricalibrare la bussola magnetica, se discordante. L'esposizione al cielo diurno dimostra poi che, anche senza vedere direttamente il Sole, gli Uccelli possono ricalibrare la bussola magnetica grazie alla luce polarizzata. La sola vista del Sole in un cielo depolarizzato è invece inefficace. La calibrazione della bussola magnetica avviene nel corso di tutta la migrazione e probabilmente ha la funzione di evitare errori di rotta dovuti alla declinazione magnetica, che può essere considerevole in certe regioni. Nel corso della migrazione l'informazione direzionale dedotta dal cielo durante il crepuscolo serale prevale su quella di origine magnetica o stellare.
Dal comportamento di molti animali si deduce che essi hanno rappresentazioni mentali della disposizione di luoghi e di punti di riferimento. Si parla allora di possesso di una mappa cognitiva. Rappresentazioni mentali di questo tipo sono acquisite attraverso l'esperienza e di solito hanno un'estensione coincidente con quella dell'area che l'animale ha avuto modo di visitare. Perché si possa parlare di una mappa, è necessario che l'animale non si limiti a percorrere vie segnate da una serie di punti di riferimento familiari, ma sappia anche trovare soluzioni spaziali nuove, per esempio individuando con sicurezza la via diretta tra due luoghi che aveva precedentemente visitato muovendosi dalla sua residenza abituale con escursioni di andata e ritorno indipendenti. La capacità di acquisire una mappa è certamente dimostrata in molti Vertebrati, mentre è discutibile se anche alcuni Artropodi la posseggano.
Grazie alla mappa cognitiva, l'animale può muoversi nella zona che essa copre in base alla sola conoscenza dei punti di riferimento, siano essi di natura visiva o di tipo chimico o acustico. Più complesso è l'uso della mappa in associazione a una bussola. In questo caso la mappa è mentalmente orientata rispetto ai punti cardinali e l'animale, riconosciuta la propria posizione rispetto all'obiettivo, può calcolare la direzione da tenere anche mediante una delle sue bussole. La migliore dimostrazione di questo fenomeno è fornita dai colombi viaggiatori esposti a un ritmo di illuminazione sfasato: anche in un'area familiare essi tendono a deviare dalla direzione attesa, come previsto in base al meccanismo di funzionamento della bussola solare.
I meccanismi di navigazione conosciuti sono numerosi e notevolmente diversi. La fonte da cui le informazioni provengono può essere utilmente adottata come criterio di classificazione. Si tenga però presente che molti animali si basano su più meccanismi di navigazione, di cui possono servirsi alternativamente o contemporaneamente, a seconda delle situazioni. L'orientamento a base genetica è ben conosciuto in seguito a studi sui Passeriformi che migrano di notte. Giunto il momento della loro prima migrazione, gli Uccelli, se tenuti in cattività, divengono attivi anche di notte e manifestano una tendenza alla fuga, la cui durata risulta essere all'incirca pari a quella del viaggio che essi farebbero se fossero liberi. Le direzioni prescelte in cattività corrispondono a quelle di migrazione in natura e possono cambiare nel corso dei tentativi di fuga, in corrispondenza dei cambiamenti di rotta in natura. Dato che l'informazione genetica, in questo caso, fornisce la direzione, il verso e la lunghezza del viaggio, essa può essere espressa tramite un vettore: si parla infatti di 'navigazione vettoriale'. Tale definizione si applica, per esempio, al caso delle api, che volano verso sorgenti di cibo segnalate dalle compagne. Come è noto, un'operaia che ha raccolto cibo in abbondanza quando ritorna nell'arnia esegue sul favo la cosiddetta 'danza scodinzolante' o 'danza a otto', dalla quale le compagne possono dedurre la distanza, la direzione e (grazie all'odore che è rimasto sul corpo della bottinatrice) la qualità della sorgente del cibo.
L'orientamento su tracce precedenti, solitamente tracce chimiche depositate nel viaggio di andata, viene utilizzato da molte specie di Molluschi e di formiche. Questo metodo è di interesse limitato e non implica i fenomeni di memoria che caratterizzano invece tutti i meccanismi di orientamento basati sul viaggio di andata. Il caso più semplice è quello dell'inversione del percorso, in cui un animale ripercorre il tragitto di andata basandosi su stimoli e punti di riferimento di cui conserva memoria, come accade per esempio per la migrazione riproduttiva dei salmoni. All'epoca della riproduzione, i salmoni del genere Oncorhynchus compiono migrazioni dette 'anadrome', perché implicano la risalita controcorrente di corsi d'acqua. Gli adulti, prima di morire, depongono le uova nella parte superiore dei corsi dei fiumi, ovvero in torrenti e ruscelli. I giovani di alcune specie iniziano, poco dopo la nascita, la migrazione verso i luoghi di residenza, mentre quelli di altre specie attendono uno o più anni prima di compiere un viaggio che li condurrà, a seconda dei casi, a un lago in cui si immette il loro fiume, oppure alle acque marine costiere nei pressi della foce o anche a fondi marini lontani migliaia di chilometri. Questi pesci, una volta adulti, tornano a riprodursi nei torrenti o nei laghi dove sono nati. Le caratteristiche delle migrazioni in mare variano da specie a specie e i relativi meccanismi di orientamento sono poco conosciuti. La risalita delle acque dolci è stata invece oggetto di numerose osservazioni ed esperimenti. Il salmone può nuotare controcorrente coprendo distanze anche superiori a 3000 km e fare una serie di svolte corrette tra fiumi confluenti o tra i vari immissari di un medesimo lago. Queste scelte sono frutto principalmente, ma non esclusivamente, di discriminazioni olfattive, basate sulla memoria degli odori che i salmoni hanno percepito dopo la nascita e nel corso della successiva migrazione. Infatti, in acquario, i salmoni sanno distinguere acque di origine diversa e mostrare preferenza per quelle provenienti dai luoghi da raggiungere. Inoltre, se privati dell'olfatto, hanno una capacità di homing ridotta. Infine, se si allevano giovani salmoni in presenza di sostanze odorose artificiali, all'epoca della migrazione risultano fortemente attratti da quelle medesime sostanze.
Tra i meccanismi basati sul viaggio di andata, il più complesso è quello detto 'di integrazione del percorso'. L'animale tiene conto della direzione e della lunghezza di ogni tratto del percorso di andata ed è in grado, in ogni momento, di calcolare la sua posizione rispetto al punto di partenza. Da questo calcolo esso deduce la lunghezza e la direzione del percorso di ritorno, sicché abbiamo di nuovo un caso di navigazione vettoriale. Questo meccanismo è particolarmente utile agli animali che compiono percorsi di andata tortuosi, per esempio alla ricerca di sorgenti puntiformi di cibo, ma hanno poi, o possono avere, bisogno di fare ritorno alla propria tana per la via più breve. È in gran parte ignoto come gli animali valutino la direzione e la lunghezza dei vari tratti del tragitto di andata ma, in base ai pochi casi ben studiati, si può distinguere un metodo fondato su riferimenti e informazioni esterne, allotetiche, e un altro basato su informazioni interne, idiotetiche, costituite da registrazioni centrali dei movimenti o dei programmi di movimento. I Mammiferi sanno usare le informazioni idiotetiche per tenere continuamente aggiornato il calcolo della propria posizione rispetto al punto di partenza, ma per lunghi percorsi il calcolo risente di un progressivo accumulo di errori, sia in rapporto alla distanza che alla direzione. Anche Mammiferi notturni come i topi e i criceti privilegiano perciò, quando è possibile, l'uso di riferimenti esterni.
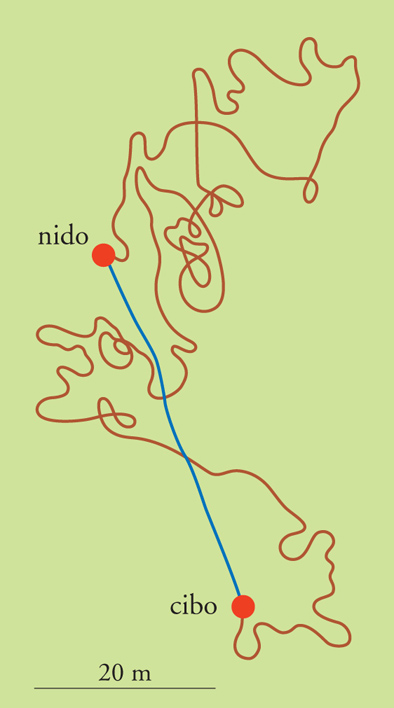
Le formiche deserticole del genere Cataglyphis sono un chiaro esempio del fenomeno dell'integrazione del percorso. Le operaie compiono veloci escursioni solitarie nelle ore più calde in cerca di cibo e tornano poi al nido senza l'ausilio di tracce odorose. Mentre la ricerca della preda può comportare un percorso lungo e tortuoso, il ritorno a casa avviene per la via più breve possibile (fig. 7). Al momento di iniziare il ritorno, la formica ha già calcolato la direzione da tenere e la distanza dal nido: infatti, se un'operaia, appena trovato il cibo, viene dislocata altrove, parte nella stessa direzione che avrebbe tenuto se non fosse stata disturbata e, una volta coperta la distanza che avrebbe dovuto portarla a casa, inizia un'esplorazione sistematica dei dintorni alla ricerca del nido. Le informazioni derivanti dal viaggio di andata (direzione e lunghezza dei singoli tratti) sono usate per calcolare direzione e distanza del viaggio di ritorno, e la direzione è calcolata sulla posizione del Sole e sulla polarizzazione celeste. Invece, il calcolo delle distanze coperte si basa probabilmente sulle immagini che si formano nella retina durante la locomozione. È interessante notare che il meccanismo di integrazione del percorso è operante anche durante il viaggio di ritorno: se una formica che sta marciando verso il nido è costretta a una deviazione a causa di un ostacolo, corregge la rotta successiva in modo da compensare la deviazione. Questo avviene anche se la formica è stata dislocata su un terreno che le è sconosciuto.
Le forme più complesse di navigazione, classificate come 'navigazione vera', sono quelle che possono prescindere dalle informazioni accumulate nel viaggio di andata e che, in molti casi, consentono di raggiungere l'obiettivo anche partendo da aree sconosciute. La navigazione vera prevede che l'animale possa calcolare in qualunque momento la propria posizione in base a riferimenti locali o a parametri fisici o chimici ambientali e che si diriga verso la meta usando una o più bussole. Il meccanismo va sotto il nome di 'mappa e bussola'; con questa definizione si intende che l'animale, per dirigersi verso la sua meta, stabilisce la propria posizione nella mappa cognitiva, ne deduce la direzione del suo obiettivo e vi si dirige poi con l'aiuto di una bussola. Il caso più semplice è quello in cui la mappa è acquisita con l'esperienza diretta; allora ha l'estensione dei luoghi noti e l'animale, trasportato e rilasciato più lontano, è incapace di orientarsi. In altri casi il meccanismo di mappa e bussola è operante anche al di fuori del territorio noto, grazie a un processo di acquisizione di informazioni a distanza. Si può, per esempio, immaginare che la memoria di punti di riferimento visibili da lontano e mai visitati possa servire a orientarsi quando ci si trova vicino a essi.
Un esempio emblematico in tal senso è quello del colombo viaggiatore, una razza domestica del colombo selvatico, Columba livia, che l'uomo ha selezionato per la capacità di ritornare a casa dopo essere stato trasportato altrove e rilasciato. I colombi, addestrati a ritornare da luoghi situati in varie direzioni, acquisiscono due mappe, entrambe funzionanti in associazione con la bussola solare. La prima è una mappa topografica dei luoghi familiari, che si estende sull'area conosciuta dal colombo; la seconda, detta 'mappa olfattiva', è incentrata, come la prima, sulla colombaia, ma è molto più estesa. La mappa olfattiva consente il ritorno da luoghi sconosciuti, come è dimostrato dal fatto che, privati dell'olfatto (ostruzione delle narici, anestesia della mucosa olfattiva, sezione dei nervi olfattivi), i colombi sono incapaci di orientarsi su aree mai sorvolate prima. La mappa olfattiva è acquisita nell'area della colombaia durante i primi mesi di vita, associando gli odori portati dai venti con la direzione da cui essi provengono. In questa maniera il colombo viene informato sugli odori prevalenti nelle aree che circondano la colombaia e può servirsene per stabilire la propria posizione rispetto a casa. Se, per esempio, un vento da nord porta un odore specifico, predominante in zone poste a nord della colombaia, il colombo, quando viene rilasciato in queste zone, vi riconosce l'odore associato con la direzione nord e dovrà semplicemente volare nella direzione opposta, per portarsi verso casa. L'acquisizione della mappa olfattiva per mezzo dell'associazione tra le direzioni dei venti e gli odori è stata dimostrata con esperimenti di allevamento dei colombi in voliere speciali, che impediscono, riducono o alterano questa associazione. Ciò ha prodotto i disturbi nella navigazione prevedibili in base al trattamento effettuato. Più diretta è la prova fornita dai risultati ottenuti con colombi esposti a venti odorosi artificiali.
Indagini sulle basi neurali del meccanismo di homing del colombo hanno messo in evidenza il coinvolgimento di due regioni cerebrali. La formazione ippocampale è essenziale per l'orientamento basato su riferimenti topografici familiari e dunque per la memoria visiva dei luoghi, mentre la corteccia piriforme, un'area che riceve numerose proiezioni dai bulbi olfattivi, ha un ruolo determinante nell'orientamento olfattivo. La lesione delle due regioni ha infatti effetti diversi sulla capacità di homing, perché, mentre i colombi privati della corteccia piriforme sono incapaci di ritornare da località sconosciute e non mostrano invece alcun deficit se rilasciati da luoghi familiari, i colombi sottoposti a lesioni della formazione ippocampale sono sempre capaci di volare in direzione di casa, ma, una volta giunti in prossimità di essa, trovano difficoltà a localizzare la colombaia non essendo in grado di riconoscerla sulla base di riferimenti visivi. Oltre a essere coinvolta nell'apprendimento e nell'uso di mappe topografiche, la formazione ippocampale ‒ in particolare la porzione che si trova nell'emisfero sinistro ‒ svolge un ruolo importante nell'apprendimento associativo mediato dalla bussola solare, cioè nell'imparare che un certo stimolo proviene da una direzione determinata.
Il meccanismo di navigazione olfattiva del colombo viaggiatore presuppone che ci siano sostanze disperse nell'atmosfera che caratterizzano regioni differenti e che queste siano sufficientemente stabili nel tempo, nonostante i fenomeni dinamici dell'atmosfera. Questa precondizione è stata dimostrata con raccolte ripetute e analisi gas-cromatografiche di campioni dell'atmosfera in una vasta regione della Baviera. Ne è risultato che, mentre le quantità assolute delle sostanze prese in considerazione variano notevolmente, i loro rapporti quantitativi sono sufficientemente stabili da poter consentire l'orientamento di un colombo teorico in una simulazione al computer.
L'applicazione di metodi di monitoraggio satellitare alle rotte di animali marini (Uccelli oceanici, Cetacei, Pinnipedi, tartarughe e Pesci) ha richiamato l'attenzione sui meccanismi di navigazione globale che questi animali possiedono. Molte specie compiono lunghe migrazioni con le quali, a dispetto dell'uniformità dell'ambiente e della deriva operata dalle correnti marine o dai venti, riescono a raggiungere con rotta più o meno diretta obiettivi lontanissimi e anche minuscoli, come certe isole oceaniche. Uno degli esempi più evidenti è rappresentato dalle femmine delle tartarughe marine, che vanno a deporre le loro uova sulla medesima spiaggia ogni due o più anni e fanno poi ritorno ai loro abituali luoghi di residenza. Questi, secondo la specie o la popolazione, possono essere molto vicini o lontani migliaia di chilometri. La telemetria satellitare ha permesso di ricostruire interi percorsi: alcuni si svolgono lungo la costa e potrebbero essere basati su mappe cognitive, altri invece corrono in mare aperto, o in acque profondissime, e quindi in assenza di riferimenti geomorfologici. Anche sui movimenti degli Uccelli marini, e in particolare sui voli di foraggiamento degli albatros nidificanti, esiste un'estesa documentazione che permette confronti con le capacità delle tartarughe.
Il meccanismo di determinazione della posizione geografica dei migratori oceanici è sconosciuto, ma sono state formulate in proposito ipotesi interessanti. La determinazione della latitudine geografica o magnetica sembra essere alla portata delle capacità neurosensoriali di molti animali: non sarebbe certamente sorprendente se l'altezza delle costellazioni circumpolari o quella del Sole al momento della culminazione venissero impiegate per determinare la latitudine geografica, mentre per la latitudine magnetica sarebbero disponibili l'intensità totale del campo magnetico terrestre e la sua inclinazione. Più difficile è immaginare quale possa essere un secondo gradiente le cui isolinee siano più o meno parallele ai meridiani geografici e forniscano, dunque, la longitudine. In certe aree, però, i soli parametri magnetici potrebbero essere sufficienti a stabilire la propria posizione, perché le isolinee dell'intensità e dell'inclinazione si intersecano con un angolo sufficientemente ampio.
L'ipotesi di una navigazione magnetica è indirettamente sostenuta dai recenti progressi nella ricerca sui meccanismi fisiologici e i componenti anatomici della percezione magnetica. Nelle tartarughe marine, esperimenti di laboratorio dimostrano reazioni orientate all'esposizione a campi magnetici artificiali che sono in accordo con l'ipotesi di un meccanismo di navigazione magnetica. Questi risultati non trovano però conferma negli esperimenti condotti su adulti in migrazione, che possono compiere traversate oceaniche e raggiungere i loro obiettivi anche quando la percezione magnetica viene sperimentalmente disturbata. Vi è tuttavia un aspetto delle capacità di orientamento delle tartarughe marine che le distingue da altri navigatori oceanici, perché esse hanno difficoltà a compensare i dislocamenti passivi. Se le femmine che hanno iniziato da poco il loro ciclo di nidificazioni vengono trasferite sperimentalmente in un punto lontano in mare aperto, esse talora sono capaci di ritrovare la spiaggia di nidificazione, impiegando per lo più molto tempo. Sembrano perciò incapaci di 'vera navigazione'. Al contrario, molte specie di Uccelli oceanici sono capaci di compensare dislocamenti sperimentali in luoghi lontani e sconosciuti. Negli albatros e nelle procellarie i movimenti migratori spontanei e i voli di ritorno dopo dislocamento sperimentale non risultano disturbati da trattamenti che interferiscono con la percezione del campo magnetico terrestre. La ricerca si è perciò indirizzata allo studio di meccanismi di navigazione olfattiva. La scoperta che le procellarie sono attratte dal solfuro di dimetile, che si accumula in aria al di sopra del mare dove c'è elevata produttività, ha aperto la via a nuovi esperimenti e ipotesi sul ruolo che gli stimoli olfattivi potrebbero avere nel dirigere i movimenti degli Uccelli oceanici. Attualmente si ritiene che questi animali siano in grado di percepire al di sopra dell'oceano un 'paesaggio olfattivo' e orientarsi in base a esso.
Bibliografia
Able, Able 1996: Able, Kenneth P. - Able, Mary A., The flexible migratory orientation system of the Savannah sparrow (Passerculus sandwichensis), "Journal of experimental biology", 199, 1996, pp. 3-8.
Bonadonna 2003: Bonadonna, Francesco - Benhamou, Simon - Jouventin, Pierre, Orientation in 'featureless' environments: the extreme case of pelagic birds, in: Avian migration, edited by Peter Berthold, Eberhard Gwinner, Edith Sonnenschein, Berlin-Heidelberg, Springer, 2003, pp. 367-377.
Bonadonna 2005: Bonadonna, Francesco e altri, Orientation in the wandering albatross: interfering with magnetic perception does not affect orientation performance, "Proceedings of the Royal Society B", 272, 2005, pp. 489-495.
Bingman, Able 2002: Bingman, Verner P. - Able, Kenneth P., Maps in birds: representational mechanisms and neural bases, "Current opinion in neurobiology", 12, 2002, pp. 745-750.
Bingman, Cheng 2005: Bingman, Verner P. - Cheng, Ken, Mechanisms of animal global navigation: comparative perspectives and enduring challenges, "Ethology, ecology and evolution", 17, 2005, pp. 295-318.
Dittman, Quinn 1996: Dittman, Andrew H. - Quinn, Thomas P., Homing in Pacific salmon: mechanisms and ecological basis, "Journal of experimental biology", 199, 1996, pp. 83-91.
Emlen 1975: Emlen, Stephen T., Migration: orientation and navigation, in: Avian biology, edited by Donald S. Famer and James R. King, New York, Academic Press, 1975, pp. 129-219.
Hedenström 2003: Hedenström, Anders - Alerstam, Thomas - Åkesson, Susanne, Ecology of long-distance movements: migration and orientation performance, "Oikos", 103, 2003, pp. 243-448.
Johnsen, Lohmann 2005: Johnsen, Sönke - Lohmann, Kenneth J., The physics and neurobiology of magnetoreception, "Nature reviews. Neuroscience", 6, 2005, pp. 703-712.
Lohmann 2004: Lohmann, Kenneth J. e altri, Geomagnetic map used in sea-turtle navigation, "Nature", 428, 2004, pp. 909-919.
Luschi 2001: Luschi, Paolo e altri, Testing the navigational abilities of ocean migrants: displacement experiments on green sea turtles (Chelonia mydas), "Behavioral ecology and sociobiology", 50, 2001, pp. 528-534.
Nevitt 1995: Nevitt, Gabrielle A. - Veit, Richard R. - Kareiva, Peter, Dimethyl sulphide as a foraging cue for Antarctic Procellariiform seabirds, "Nature", 376, 1995, pp. 680-682.
Papi 2000: Papi, Floriano e altri, Open-sea migration of magnetically disturbed sea turtles, "Journal of experimental biology", 203, 2000, pp. 3435-3443.
Papi 2006: Papi, Floriano, Navigation of marine, freshwater and coastal animals: concepts and current problems, "Marine and freshwater behaviour and physiology" (in corso di stampa).
Papi, Luschi 1996: Papi, Floriano - Luschi, Paolo, Pinpointing 'Isla Meta': the case of sea turtles and albatrosses, "Journal of experimental biology", 199, 1996, pp. 65-71.
Papi, Wallraff 1992: Papi, Floriano - Wallraff, Hans G., Birds, in: Animal homing, edited by Floriano Papi, London-New York, Chapman & Hall, 1992, pp. 263-319.
Pardi 1979: Pardi, Leo, Orientamento degli animali, in: Enciclopedia del Novecento, Roma, Istituto della Enciclopedia Italiana, 1979, IV, pp. 965-988.
Quinn 2005: Quinn, Thomas P., The behaviour and ecology of Pacific salmon and trout, Bethesda, American Fisheries Society, 2005.
Thinus-Blanc 1987: Thinus-Blanc, Catherine, The cognitive map concept and its consequences, in: Cognitive processes and spatial orientation in animal and man, edited by Paul Ellen and Catherine Thinus-Blanc, Dordrecht, Nijhoff, 1987, I, pp. 1-19.
Wallraff 2003: Wallraff, Hans G., Zur olfaktorischen Navigation der Vögel, "Journal für Ornithologie", 144, 2003, pp. 1-32.
Wallraff 2005: Wallraff, Hans G., Avian navigation: pigeon homing as a paradigm, Berlin, Springer, 2005.
Wehner 1992: Wehner, Rüdiger, Arthropods, in: Animal homing, edited by Floriano Papi, London-New York, Chapman & Hall, 1992, pp. 45-144.
Wiltschko, Wiltschko 1995: Wiltschko, Roswitha - Wiltschko, Wolfgang, Magnetic orientation in animals, Berlin-New York, Springer, 1995.