
Neurobiologia
NEUROBIOLOGIA
Organizzazione neuronale cerebrale e cerebellare di János Szentágothai
sommario: 1. Introduzione generale: formazioni laminari del tessuto nervoso. 2. La corteccia cerebellare. a) Sviluppo della corteccia cerebellare. b) I neuroni della corteccia cerebellare. c) Organizzazione della corteccia cerebellare in zone sagittali. d) Modelli concettuali delle funzioni delle reti neuronali cerebellari. 3. La corteccia cerebrale. a) Aspetti dello sviluppo. b) Architettonica della corteccia. c) Caratterizzazione dei sei strati principali della corteccia. d) Tipi cellulari della isocorteccia. e) Connessioni interneuronali. f) Il principio dell'architettonica modulare. g) Sistemi di fibre (e cellule) diffuse. □ Bibliografia.
1. Introduzione generale: formazioni laminari del tessuto nervoso
La presenza costante di strutture laminari nel sistema nervoso dei Vertebrati è dovuta al meccanismo di formazione delle cellule nervose (e gliali) dal neuroepitelio del tubo neurale primitivo che è un epitelio cilindrico pseudostratificato le cui cellule tappezzano le superfici interna ed esterna del tubo. I nuclei cellulari, tuttavia, possono muoversi liberamente nello strato immediatamente sovrastante la superficie ventricolare per circa tre quarti della distanza totale tra la superficie interna e quella esterna del tubo. La zona più interna, in cui hanno luogo le divisioni cellulari, è chiamata ‛zona ventricolare' (o germinativa); la zona successiva, verso l'esterno, è detta ‛zona intermedia' (o mantello); quella superficiale è definita ‛zona marginale'.
Nella maggior parte delle strutture del sistema nervoso centrale la proliferazione cellulare avviene unicamente nella zona ventricolare. Le cellule, una volta divenute postmitotiche, si dirigono verso la zona intermedia e sono destinate a differenziarsi in neuroni (o, in misura minore, sempre in questo stadio, anche in cellule gliali) andando a costituire il ‛mantello' dell'encefalo. Contrariamente a quanto potrebbe apparire logico, saranno le cellule che sono migrate verso il mantello nella fase più precoce a formare lo strato più interno dell'organo in formazione. Le cellule che sono generate in stadi successivi normalmente oltrepassano lo strato in cui si è stabilita la generazione precedente e si dispongono al suo esterno, così che ogni successiva generazione di progenitori di cellule nervose (e probabilmente cellule gliali) va a formare strati sempre più esterni del mantello.
In fasi più avanzate dello sviluppo delle strutture più grandi (gli emisferi in particolare), proprio sopra la zona ventricolare compare uno strato isolato di cellule che si dividono, detto ‛zona subventricolare'. In questo strato l'attività mitotica continua non solo durante l'intera vita fetale, ma anche per un certo periodo della vita extrauterina. È generalmente ritenuto che la maggioranza delle cellule gliali e alcuni dei neuroni più piccoli siano prodotti in questo strato.
La zona ventricolare dà infine origine alle cellule ependimali che tappezzano le superfici ventricolari. Tuttavia, questo è solo uno dei modi attraverso cui hanno origine le cellule che andranno a costituire la parete dell'encefalo. Il miglior esempio dell'altro meccanismo che produce cellule encefaliche in gran numero è osservabile al livello del IV ventricolo; infatti, dopo che la produzione di materiale cellulare della zona ventricolare (zona germinativa) si è esaurita in gran parte delle regioni della parete ventricolare, rimane ancora una popolazione di cellule altamente proliferative situata ai margini postero-laterali del tetto del ventricolo, il cosiddetto ‛labbro romboide'. Qui le cellule aumentano di numero molto rapidamente e quindi si spostano sulla superficie esterna dell'abbozzo cerebellare in direzione rostrale e mediale. Tali cellule continuano a proliferare ricoprendo la zona marginale e formando uno strato ‛granulare esterno' che contribuirà successivamente alla formazione di gran parte del materiale cellulare delta corteccia cerebellare. In generale, le cellule nervose sono prodotte con un meccanismo analogo - produzione intensiva di progenitori di cellule nervose e migrazione in massa in altre regioni del cervello in via di sviluppo - ma la trattazione di questo argomento andrebbe oltre lo scopo di questo lavoro.
Le strutture corticali vere e proprie si sviluppano relativamente tardi nell'evoluzione del sistema nervoso: nella filogenesi dei Vertebrati esse compaiono per prime nel cervelletto dei Pesci e raggiungono la maggiore complessità - certamente nelle strutture anatomiche se non nei loro circuiti neuronali - nel sistema nervoso centrale dei mormiriformi, ordine degli Holostei. (I mormiriformi sono pesci debolmente elettrici, che producono impulsi elettrici mediante organi muscolari situati nella coda e che mediante elettrorecettori, cioè recettori modificati del sistema della linea laterale, controllano i campi elettrici esterni).
La comparsa di reali strutture corticali cerebrali avviene in una fase dell'evoluzione relativamente recente. La corteccia vera e propria compare, anche se in forma piuttosto rudimentale, nel cervello degli Uccelli, ma solo nei Mammiferi raggiunge le caratteristiche essenziali di organizzazione proprie della corteccia. Gli elementi strutturali fondamentali della corteccia, una volta stabiliti, sono tuttavia mantenuti rigidamente nell'intera classe dei Mammiferi.
2. La corteccia cerebellare
La corteccia del cervelletto è uno strato di sostanza grigia trilaminare ripiegato in maniera complessa, che riveste le sottili lamine di sostanza bianca che si distendono a ventaglio dalle più estese masse centrali, in modo molto simile alle pagine di un libro parzialmente aperto. La differenza, tuttavia, risiede nel fatto che le lamine principali e più spesse di sostanza bianca danno origine a lamine (elementari) secondarie e terziarie. Ne risulta, in sezione trasversa, quella caratteristica configurazione della sostanza bianca e grigia, chiamata dagli antichi anatomisti arbor vitae, che somiglia a una foglia di Thuja occidentalis . Il tessuto corticale che corrisponde a una plica elementare, largo circa 1 mm alla base e con una circonferenza di 3-4 mm, è comunemente chiamato folium (lamella); alla base di ciascun folium il tessuto corticale si piega all'indietro nelle due lamelle adiacenti senza che vi sia soluzione di continuità. Di conseguenza, lo strato corticale tra due lamelle contigue presenta sempre una ripiegatura all'interno con la concavità rivolta verso l'esterno. Le arborizzazioni neuronali sono pertanto sottoposte a una continua trasformazione cartesiana, passando da ‛convesse' nella parte più alta delle lamelle, a ‛dritte rettangolari' lungo la loro parte rettilinea, a ‛concave' rispetto all'interno del cervelletto. Tali trasformazioni cartesiane della rete neuronale modificano radicalmente la ‛geometria' (e anche le caratteristiche metriche) delle ramificazioni dendritiche e assoniche, ma non la loro ‛topologia' (da non confondere con il termine topografia). La lunghezza delle lamelle varia enormemente non soltanto tra specie aventi cervelletto di dimensioni molto diverse, ma anche nella stessa specie a seconda della zona del cervelletto in cui sono situate. Le lamelle del lobo mediano (o verme) sono in genere - specialmente nei mammiferi inferiori - le più lunghe, mentre quelle dei lobi laterali (emisferi) sono le più corte. La relativa brevità delle lamelle emisferiche è il risultato di un ripiegamento degli emisferi in due anse successive, di cui la rostrale è il ‛lobo ansiforme' e la caudale il ‛lobo paraflocculare', con una zona intermedia che costituisce il ‛lobo paramediano'. Tale ripiegamento, più evidente nei Carnivori, è chiaramente identificabile nei primati inferiori all'uomo. Nei Mammiferi, il cervelletto raggiunge il suo massimo sviluppo nell'uomo e nei Cetacei. La somiglianza è però molto più apparente che reale, poiché se si prendono in esame le suddivisioni omologhe corrispondenti, le zone maggiormente sviluppate nei Cetacei (lobi paramediano e paraflocculare ventrale), che costituiscono più della metà degli emisferi, sono solo di modeste dimensioni nell'uomo.
I primi studi di anatomia comparata del cervelletto dei Mammiferi e un'accettabile identificazione delle parti omologhe furono effettuati coi lavori pionieristici di Bradley (v., 1905) e ancor più di Bolk (v., 1906). Una svolta fondamentale fu tuttavia determinata dagli studi sugli Uccelli di Larsell (v., 1948), il quale suddivise il verme in 10 lobuli principali contrassegnati da numeri romani (I-X, iniziando dalla parte anteriore). In seguito, grazie soprattutto all'apporto di Jan Jansen, un'analoga suddivisione in lobuli fu adottata anche per gli emisferi.
a) Sviluppo della corteccia cerebellare
La corteccia cerebellare si sviluppa dalla porzione caudale della lamina cerebellare primitiva. La parte anteriore del tetto del IV ventricolo inizia a formarsi, nel ratto, al 13° giorno di vita embrionale. (La seguente descrizione è ripresa da Altman, 1982, perché i suoi studi forniscono i dati più esaurienti e completi di datazione cellulare basati su esperimenti di marcatura con timidina triziata. Ove necessario, i dati corrispondenti per altre specie saranno menzionati a parte).
I neuroni sono generati - come in tutte le altre parti del tubo neurale - nell'epitelio che riveste il lume ventricolare. Si ritiene che le cellule che si differenziano nella porzione rostrale della lamina cerebellare già al 13° giorno di vita embrionale, e che sono state generate circa un giorno prima, diano origine alle cellule del locus ceruleus, mentre quelle che si differenziano subito dopo nelle zone più laterali della lamina anteriore diventino cellule del nucleo vestibolare laterale. Le cellule del cervelletto vero e proprio sono generate nella metà caudale della lamina cerebellare dal 13° giorno in poi. Le cellule dei nuclei cerebellari sono le prime a formarsi, con un livello massimo di produzione al 14° giorno, e sono seguite, con un intervallo di tempo di un giorno, dalle cellule di Purkinje, le quali raggiungono la produzione massima durante il 15° giorno. Le cellule di Golgi del futuro strato granulare compaiono per ultime, a partire dal 19° giorno di vita embrionale. Poiché le cellule di Purkinje vengono generate più tardi dei nuclei, esse (e successivamente le cellule di Golgi) debbono attraversare la massa cellulare dei neuroni nucleari per assumere la loro disposizione definitiva in un'unica fila al di sotto dello strato molecolare che si sviluppa più tardi. Questa penetrazione avviene principalmente intorno al 20° giorno, ed è, come ricordato precedentemente, una caratteristica pressoché generale dello sviluppo corticale. Una fase decisiva dello sviluppo corticale cerebellare si ha al 17° giorno di vita embrionale, allorché un gran numero di cellule viene generato nel margine postero-laterale della lamina cerebellare, il labbro romboide, dando origine allo strato granulare esterno (SGE). Tali cellule si diffondono molto rapidamente su tutta la superficie della lamina cerebellare e raggiungono il margine anteriore del cervelletto primitivo nel 22° giorno. Le cellule di Purkinje si allineano dapprima in parecchi strati, subito dopo in un unica fila tra lo strato granulare esterno e i nuclei cerebellari profondi. È quindi al 22° giorno che i componenti cellulari dell'abbozzo del cervelletto si trovano disposti nelle loro posizioni finali; più esattamente, nelle posizioni dalle quali inizierà l'‛istogenesi cerebellare', cioè il differenziamento e il movimento dei neuroni nelle loro posizioni definitive. La disposizione dei corpi cellulari di Purkinje in un'unica fila (che nel ratto inizia al 4° giorno di vita postnatale con differenze regionali) può esser dovuta alla compressione esercitata da un lato dallo strato granulare esterno in rapido sviluppo (e in seguito dallo strato molecolare in via di formazione) e dall'altro lato dallo strato granulare interno in via di accrescimento. Le cellule dello strato granulare esterno proliferano rapidamente durante il primo periodo di vita extrauterina. Le cellule più profonde dello strato granulare esterno si differenziano per prime e diventano in prevalenza le prime cellule granulari. Dopo che hanno assunto un aspetto a fuso a direzione trasversa, emettono due branche assoniche che saranno più tardi le fibre parallele. Ben presto il pericario inizia la sua discesa attraverso lo strato delle cellule di Purkinje per andare a formare lo strato granulare interno, il che conferisce alle cellule granulari la loro caratteristica forma a T. Secondo gli studi di Rakic (v., 1971), il movimento dei corpi delle cellule granulari è reso possibile dalla loro adesione ai processi verticali della glia di Bergmann e da un meccanismo di scorrimento, dovuto all'interazione delle due superfici cellulari. Le prime cellule granulari entrano in contatto con le prime fibre muschiose (afferenti del cervelletto) non appena arrivano nello strato immediatamente inferiore a quello delle cellule di Purkinje, costituendo subito le prime sinapsi nonostante che non siano ancora sviluppati i dendriti caratteristici delle cellule granulari. Il medesimo processo di differenziamento prosegue in direzione esterna (nel ratto oltre i primi 21 giorni di vita), così che le fibre parallele più precoci sono ammassate nella zona più profonda dello strato molecolare in formazione, le ultime in quella più esterna. Non tutte le cellule dello strato granulare esterno, tuttavia, si trasformano in cellule granulari. Nel 40% inferiore di quello che diverrà lo strato molecolare alcune cellule si differenziano in ‛cellule a canestro' (il 6°-7° giorno nel ratto), mentre nel restante 60% più esterno si sviluppano le ‛cellule stellate' (tra l'8° e l'11° giorno nel ratto). Quindi anche gli interneuroni inibitori dello strato molecolare si sviluppano da componenti cellulari dello strato granulare esterno. L'aspetto più notevole, anche se meno conosciuto, dell'istologia cerebellare è l'arborizzazione dendritica piana, disposta ad angolo retto rispetto al decorso delle fibre parallele, che riguarda prevalentemente i dendriti delle cellule di Purkinje, ma anche, seppur limitatamente, quelli delle cellule a canestro e stellate. La forma e la disposizione spaziale caratteristiche dell'arborizzazione della cellula di Purkinje (v., 1837), somigliante al candelabro a sette braccia (menorah) della liturgia ebraica, è chiaramente interpretabile come un metodo ingegnoso di disporre arborizzazioni dendritiche separate (cioè non compenetrantisi) senza che vi sia perdita di spazio tra di loro, ma non si è riusciti a capire come si attui tale sistemazione. Sono state proposte varie ipotesi per una spiegazione plausibile di questo curioso fenomeno, ma nessuna è del tutto soddisfacente.
Tra i neuroni corticali e le fibre afferenti al cervelletto si stabiliscono, già durante i primi stadi del differenziamento cellulare, primitive sinapsi; però queste sinapsi precoci raramente o forse mai corrispondono, per forma e localizzazione sui neuroni bersaglio, alla loro struttura e localizzazione definitive. Questo è dovuto a quattro distinti processi: 1) una ‛translocazione' delle sinapsi durante il differenziamento e la maturazione del tessuto, come già individuato da Ramón y Cajal (v., 1911) e messo in evidenza da Larramendi (v., 1969); 2) un processo specifico di selezione, per cui solo pochi o talvolta (ad es. nel caso delle fibre rampicanti) uno solo dei molti elementi presinaptici disponibili permane come singola afferenza; 3) una crescita differenziale delle parti neuronali che partecipano alla sinapsi, consistente in divisioni, emissioni di protuberanze, prolungamenti, ecc.; infine 4) modificazioni prodotte dallo sviluppo di nuove parti neuronali che compaiono più tardi e vanno a incunearsi tra gli elementi preesistenti. (I meccanismi menzionati ai punti 3) e 4) sono più evidenti nello sviluppo dei glomeruli sinaptici).
b) I neuroni della corteccia cerebellare
Nella corteccia cerebellare sono presenti cinque differenti tipi di neuroni: cellule di Purkinje (v. Purkinje, 1837), granuli, cellule a canestro, cellule di Golgi e cellule stellate. E stato descritto da Lugaro (v., 1895) un ulteriore tipo di cellule - fusiformi, ampie, orientate orizzontalmente - ma nonostante i recenti studi di Palay e Chan-Palay (v., 1974) miranti a individuarle più chiaramente e in particolare a chiarire quali contatti stabiliscano i loro assoni, i risultati sono troppo ambigui per essere inclusi in questa rassegna.
1. Organizzazione fondamentale delle catene neuronali. - Se si tenta di tracciare la via che seguono gli impulsi che entrano nel cervelletto, si possono individuare due catene sequenziali di neuroni: 1) fibre muschiose → sinapsi glomerulari → granuli → fibre parallele → cellule di Purkirje → cellule nucleari cerebellari (vestibolari); 2) fibre rampicanti → cellule di Purkinje → neuroni nucleari cerebellari (vestibolari). Queste sono le catene funzionali attraverso cui passa l'impulso, ma esistono molte altre catene collaterali di neuroni che possono determinare un'inibizione a feedback (retroazione) o a feedforward (proazione); tali catene collaterali verranno descritte in seguito.
Dati questi complessi collegamenti tra le catene neuronali, non ci sembra ragionevole cercare di descrivere gli elementi neuronali della corteccia cerebellare cercando di seguire un ordine sequenziale basato sulla funzione, ma cercheremo invece di seguire l'ordine imposto dalla specifica organizzazione strutturale considerata. La configurazione generale della corteccia cerebellare è illustrata schematicamente nella fig. 7; la sua struttura microscopica è mostrata nella tav. IB, in cui è stata utilizzata una reazione immunocitochimica per evidenziare i neuroni GABA-ergici che appaiono colorati in marrone. Tutte le altre cellule (specialmente i nuclei) sono colorate col verde metile.
2. Le cellule di Purkinje o strato ganglionare. - Le cellule di Purkinje rappresentano probabilmente il più importante tipo cellulare dell'intero sistema nervoso dei Vertebrati. I corpi cellulari sono disposti in un'unica fila (più correttamente, in uno strato costituito da un'unica fila di cellule) e pertanto costituiscono uno strato intermedio chiaramente separato della corteccia cerebellare. La forma del corpo cellulare ricorda quella di un ravanello o di una barbabietola. Il suo processo dendritico principale si dirige verso la superficie e si divide immediatamente in due rami principali. L'intera arborizzazione dendritica si espande in piano, come una pianta a spalliera, perpendicolarmente all'asse longitudinale delle lamelle. La posizione relativa e la direzione dei due rami dendritici principali dipendono dalla posizione delle cellule di Purkinje: nel punto di convessità delle lamelle i due rami decorrono ad angolo acuto, cosicché il piano delle arborizzazioni dendritiche ha l'aspetto di un ventaglio, mentre diviene quadrangolare (250 × 300 μm) o leggermente rettangolare nelle parti rettilinee della superficie lamellare; nella parte più profonda delle pieghe (nel punto di massima concavità) il piano dendritico assume l'aspetto di un ventaglio rovesciato (v. tav. II). Lo spessore del piano dendritico è di 8-9 μm, ma dato che il piano dei due principali rami dendritici può esser spostato lungo l'asse longitudinale delle lamelle, il piano dendritico di una singola cellula di Purkinje può raggiungere uno spessore doppio o anche triplo. I dendriti primari e secondari sono relativamente lisci e presentano solo spine di tipo particolare (bulbose e non acuminate; v. Larramendi e Victor, 1967), che stabiliscono sinapsi con le fibre rampicanti (v. Palay e Chan-Palay, 1974). I rami dendritici terziari vengono detti ‛rametti spinosi', ma anche i rami discendenti di secondo ordine possono presentare delle spine. Le spine, che hanno una lunghezza di 1,5-2 μm, protrudono da tutti i lati dei rametti dendritici che ne sono letteralmente coperti. È stato calcolato che nel gatto il numero totale di spine per ogni cellula di Purkinje è di circa 80.000. Le spine dei rametti spinosi entrano in contatto quasi esclusivamente con le fibre parallele. Gli assoni delle cellule di Purkinje subito dopo la loro origine dai poli basali dei corpi cellulari (nel tratto corrispondente alla radice del ‛ravanello' o ‛barbabietola') mostrano caratteristiche ultrastrutturali particolan: un rivestimento elettrondenso (v. Chan-Palay, 1972) e ammassi di microtubuli a matassa. La parte non mielinizzata del segmento iniziale dell'assone è in gran parte coperta dai rami terminali degli assoni delle cellule a canestro (v. sotto, 5) che formano i prolungamenti verso il basso dei canestri intorno alle cellule di Purkinje (v. Ramón y Cajal, 1888), definiti ‛pennelli' da Palay e Chan-Palay (v., 1974). Durante la loro discesa attraverso lo strato granulare, e anche dopo che sono penetrati nelle lamine di sostanza bianca, gli assoni emettono dei caratteristici rami collaterali iniziali, che hanno decorso ricorrente; quando questi raggiungono lo strato ganglionare si dividono in due ricchi plessi orientati tangenzialmente (paralleli alla superficie). Le collaterali assoniche delle cellule di Purkinje entrano in contatto coi dendriti principali delle cellule di Purkinje e coi corpi cellulari delle cellule di Golgi e delle cellule a canestro. L'assone principale continua poi il suo decorso e termina nei nuclei cerebellare e vestibolare, dando luogo a un'arborizzazione conica formata da 2-3 diramazioni principali relativamente dritte (v. Chan-Palay, 1977; v. Palkovits e altri, 1977); queste emettono rametti terminali piuttosto brevi in direzione dei dendriti e dei corpi cellulari delle cellule nucleari. Le terminazioni assoniche delle cellule di Purkinie hanno le stesse caratteristiche ultrastrutturali riscontrate nelle sinapsi inibitorie (vescicole sinaptiche ovoidali, schiacciate o pleomorfiche, e contatto ‛simmetrico' con la membrana). Tramite metodi immunocitochimici si possono mettere in evidenza in tutte le terminazioni dell'assone (e anche delle sue collaterali) delle cellule di Purkinje sia decarbossilasi dell'acido glutammico (GAD), sia acido gamma-amminobutirrico (GABA). (Per ulteriori dettagli, che non possono essere discussi nella brevità di queste pagine, e per una rassegna critica dell'abbondante letteratura, v. Ito, 1984).
3. Lo strato granulare. - I granuli sono senz'altro il più numeroso tipo di neuroni presente in tutto il sistema nervoso dei Vertebrati; nell'uomo il loro numero potrebbe essere maggiore di quello di tutti gli altri neuroni dell'organismo. Questo sorprendente dato presuntivo risulta dai calcoli di Zagon e altri (v., 1977) ed è basato sul contenuto in DNA del cervelletto e su precedenti ipotesi di Braitenberg e Atwood (v., 1958). Tale numero non è assurdo se paragonato al numero dei granuli (2,2 × 109) presenti nel cervelletto del gatto, determinato con metodi stereologici più prudenti (v. Palkovits e altri, 1977). Il volume totale dello strato granulare può salire, dal gatto all'uomo, di 10 volte, ma la grandezza e la densità delle cellule presentano variazioni minime o pressoché nulle. Una stima, anche se molto prudente, di tutti gli altri neuroni può essere basata sul numero di 1010 neuroni presenti nella corteccia cerebrale umana (v. von Economo e Koskinas, 1925, e conteggi non pubblicati dell'autore). Dato che il numero totale di tutte le altre cellule nervose in tutti gli altri distretti, centrali e periferici (gangli e plessi), non può essere molto superiore a 1010 il rapporto tra i neuroni granulari del cervelletto e tutti gli altri neuroni nell'organismo umano potrebbe essere di 4-5 × 1010 : 2 × 1010. La maggior parte dei granuli ha 4 dendriti relativamente corti con piccoli bulbi terminali disposti ad artiglio. I corpi cellulari sono arrotondati o, più esattamente, poliedrici, con un diametro di 5-8 μm. Il sottile assone risale attraverso lo strato ganglionare (formato dai corpi delle cellule di Purkinje) nello strato molecolare, dove si divide in due rami a forma di T, chiamati fibre parallele (v. sotto).
Il neurotrasmettitore che è generalmente considerato responsabile della trasmissione dell'impulso dalle fibre parallele alle cellule di Purkinje è il glutammato, ma l'evidenza è per il momento di natura indiziaria (per una discussione al riguardo, v. Ito, 1984).
Le ‛fibre muschiose' (o muscoidi) sono uno dei due tipi di afferenze della corteccia cerebellare. Tranne le afferenze che hanno origine nell'oliva inferiore, tutte le afferenze che provengono da altre parti (e cioè spino-cerebellari, reticolocerebellari, ponto-cerebellari, vestibolo-cerebellari, cuneocerebellari e altre ancora) terminano come fibre muschiose.
Tuttavia, una notevole parte di fibre muschiose ha origine nei nuclei cerebellari; tali afferenze muschiose nucleari, benché leggermente più piccole e di forma bulbosa più semplice (v. Hàmori e altri, 1981), stabiliscono contatti sinaptici sostanzialmente uguali a quelli delle fibre muschiose di diversa provenienza. È noto che le fibre muschiose generalmente si dividono per dirigersi verso due lamelle adiacenti, con svariate ramificazioni e diverse terminazioni sinaptiche ‛a rosetta', intermedie (‛di passaggio') e terminali, nello strato granulare. Le rosette sinaptiche possono essere di forma e grandezza considerevolmente diverse (di norma quelle intermedie sono meno elaborate) e hanno conformazione generalmente sinusoidale con un notevole numero di diramazioni laterali bulbose. Esse sono densamente coperte dalle corte terminazioni dendritiche (a forma di artiglio o di dito) dei granuli. Le rosette muschiose e i loro contatti sinaptici formano i ben noti ‛glomeruli' o ‛isole sinaptiche' dello strato granulare, che possono essere evidenziati chiaramente coi metodi tradizionali di colorazione cellulare. La struttura e i costituenti dei glomeruli sinaptici sono illustrati in forma semidiagrammatica nella fig. 9. Vi sono altri due elementi che partecipano alla formazione del glomerulo. Le branche assoniche dei neuroni di Golgi formano un delicato intrico, in genere alla periferia del glomerulo, e le loro brevi terminazioni entrano in contatto con i dendriti dei granuli a valle delle sinapsi tra fibre muschiose e dendriti granulari; sia l'ultrastruttura delle terminazioni presinaptiche degli assoni delle cellule di Golgi che il contenuto in decarbossilasi dell'acido glutammico (GAD) e in acido gamma-amminobutirrico (GABA) mostrano chiaramente la loro natura inibitoria. Meno spesso, le fibre muschiose entrano in contatto anche con cellule di Golgi, in parte con i loro dendriti principali (v. Eccles e altri, 1967), o - secondo Palay e Chan-Palay (v., 1974) più frequentemente con i loro corpi cellulari. Questi contatti presentano caratteristici ripiegamenti paralleli della membrana postsinaptica, tanto che questa sinapsi è stata chiamata da Palay e Chan-Palay ‛sinapsi a castagna'; questa icastica definizione si attaglia particolarmente ai contatti assosomatici.
Finora non è stato possibile accertare la natura del neurotrasmettitore delle fibre muschiose e non è stato neppure trovato un soddisfacente candidato anche se inizialmente sembrava che vi fossero alcune vaghe indicazioni. In tempi più recenti due peptidi hanno assunto il ruolo di possibili candidati, e cioè la sostanza P e la somatostatina. L'azione di controllo inibitorio intrinseca alle terminazioni assoniche di Golgi è senza dubbio mediata dal GABA.
4. Lo strato molecolare. - Il costituente più importante dello strato molecolare, e cioè le arborizzazioni dendritiche delle cellule di Purkinje, è stato descritto in precedenza. Un altro costituente importante di questo strato è rappresentato dalle fibre parallele, che sono gli assoni dei granuli dello strato granulare che ascendono verticalmente nello strato molecolare e che si dividono in due branche assoniche a forma di T aventi corso esattamente parallelo alla superficie delle lamelle. La lunghezza delle fibre parallele è ancora controversa: nel gatto sono state rilevate lunghezze che variano da 2 a 7 mm. Dato che le lamelle, specialmente nelle circonvoluzioni degli emisferi cerebellari, sono relativamente brevi (2-3 mm nel gatto) e terminano bruscamente da ambedue le parti, le fibre parallele non possono avere una lunghezza maggiore delle lamelle stesse, e cioè 2-3 mm. Le fibre parallele formano sinapsi con tutti i tipi di spine dendritiche che incontrano lungo il loro corso; poiché la maggior parte dei dendriti decorre ad angolo retto rispetto all'asse longitudinale delle lamelle, le ramificazioni dendritiche delle cellule di Purkinje e degli interneuroni inibitori locali incrociano ad angolo retto le fibre parallele. Le sinapsi dello strato molecolare sono quindi in prevalenza ‛di incrocio', un tipo di sinapsi in cui l'elemento presinaptico non è terminale ma assume la forma di un allargamento fusiforme delle fibre parallele che entrano in contatto con le spine dendritiche. Molte spine dendritiche sono come invaginate nella parte ispessita delle fibre parallele. L'ultrastruttura delle sinapsi delle fibre parallele corrisponde a quella che è generalmente accettata come tipica delle sinapsi eccitatorie.
Questa regola generale dell'architettura sinaptica non è rispettata nel caso dell'altro tipo di afferenze principali della corteccia cerebellare, e cioè le ‛fibre rampicanti'. Attualmente sembra stabilito che le fibre rampicanti abbiano origine esclusivamente nell'oliva inferiore (v. Szenthàgotai e Rajkovits, 1959; v. Desclin, 1974). Benché più d'una fibra si avvicini a una determinata cellula di Purkinje formando un morbido gomitolo intorno al corpo cellulare, in genere solo una fibra rampicante sopravvive nel cervelletto adulto, mentre le altre vengono eliminate (durante il processo ricordato nel cap. 2, È a).
Le fibre rampicanti stabiliscono contatti multipli con le spine più brevi e spesse dei dendriti primari e secondari; tali contatti sinaptici possono essere dell'ordine delle centinaia.
Le sinapsi delle fibre rampicanti esercitano un azione sicuramente eccitatoria. Nelle registrazioni intracellulari la risposta delle fibre rampicanti mostra un improvviso picco iniziale seguito da una lunga (10-15 ms) depolarizzazione della membrana che determina una rapida sequenza di picchi (da 2 a 6). Si ritiene generalmente che l'aspartato sia il neurotrasmettitore implicato nelle sinapsi tra fibre rampicanti e cellule di Purkinje.
Le fibre rampicanti hanno collaterali dirette alle cellule di Golgi, principalmente verso i loro corpi cellulari. È probabile, anche se non certo, che altre collaterali prendano contatto con le cellule a canestro e stellate.
5. Interneuroni inibitori locali. - Un altro importante costituente della corteccia cerebellare è rappresentato dagli interneuroni inibitori locali. Nonostante che il principale neurone efferente (la cellula di Purkinje) sia esso stesso inibitorio, vi è una grande quantità di altri neuroni inibitori locali. Questi ultimi sono disposti in circuiti a feedback - che sono stati illustrati già nei classici diagrammi di Ramón y Cajal - o, in grado minore, in vie collaterali a feedforward in parallelo alla catena neuronica principale. Gli interneuroni inibitori sono presenti non soltanto nello strato molecolare, per cui questa descrizione è valida per i vari strati della corteccia.
I neuroni di Golgi sono cellule abbastanza grandi che si trovano in tutto lo strato molecolare, anche se sono preferenzialmente collocati immediatamente al di sotto della fila dei corpi delle cellule di Purkinje. Hanno rami dendritici relativamente densi, orientati in prevalenza verso la superficie della corteccia cerebellare. T dendriti ascendenti penetrano attraverso lo strato molecolare, quelli discendenti si ramificano nello strato granulare dove stabiliscono sinapsi con le fibre muschiose. Alcune cellule di Golgi, in genere più piccole e poste a maggior profondità, non hanno dendriti che entrano nello strato molecolare ma i loro dendriti ricevono afferenze esclusivamente dalle fibre muschiose; per il resto presentano una struttura analoga.
Gli assoni hanno abbondanti ramificazioni, in genere molto vicine a quelle dei dendriti, e sono localizzati esclusivamente nello strato granulare. I rametti preterminali si suddividono in piccole terminazioni o ispessimenti ‛di passaggio' che avvolgono i glomeruli cerebellari. I contatti vengono stabiliti con le terminazioni dendritiche a forma di artiglio dei granuli sempre in prossimità (cioè a valle) dei contatti tra le terminazioni delle fibre muschiose e le terminazioni dendritiche dei granuli stessi. Le cellule di Golgi contengono GABA e decarbossilasi dell'acido glutammico in tutte le loro parti, e pertanto il meccanismo da esse usato per la trasmissione dell'impulso è senz'altro GABA-ergico. Le cellule di Golgi ricevono afferenze sinaptiche molteplici. L'afferenza principale è rappresentata dalle fibre parallele che prendono contatto con i loro dendriti nello strato molecolare; un'altra afferenza deriva direttamente dalle fibre muschiose, che innervano il loro corpo cellulare con le sinapsi ‛a castagna' (v. Palay e Chan-Palay, 1974) o i loro dendriti principali con analoghe sinapsi; un'ultima afferenza sinaptica arriva alle cellule di Golgi (in genere sui corpi cellulari) dalle collaterali delle fibre rampicanti (eccitatorie) e dalle collaterali assoniche delle cellule di Purkinje (inibitorie).
I neuroni a canestro sono situati nel terzo inferiore dello strato molecolare, cioè immediatamente al di sopra della fila delle cellule di Purkinje. Le arborizzazioni dendritiche si espandono a raggera, ma vengono compresse nella direzione dell'asse longitudinale delle lamelle; la maggioranza dei rami è confinata negli spazi che rimangono liberi tra le arborizzazioni dendritiche di cellule di Purkinje adiacenti ed è pertanto orientata ad angolo retto rispetto all'asse longitudinale delle lamelle. I dendriti che si trovano più in basso hanno decorso parallelo allo strato dei corpi delle cellule di Purkinje, mentre quelli superiori risalgono nello strato molecolare assumendo la forma di un semicerchio irregolare o semiellissoide.
L'assone è estremamente caratteristico: dopo un segmento iniziale più sottile, esso si ispessisce e decorre parallelamente alla superficie ad angolo retto rispetto alle fibie parallele in una delle due direzioni del piano trasverso della lamella. Gli assoni hanno una lunghezza di 500-600 μm e presentano ramificazioni laterali secondarie, in direzione longitudinale rispetto alla lamella, grazie alle quali possono stabilire contatti con molte file successive di cellule di Purkinje in direzione longitudinale. Le ramificazioni preterminali discendono verso lo strato delle cellule di Purkinje divenendo considerevolmente più larghe e rigonfie nella loro parte terminale, che si attacca alla metà inferiore dei corpi delle cellule di Purkinje stabilendo con queste numerose sinapsi e alcune anche coi segmenti iniziali degli assoni delle cellule di Purkinje. I rami terminali spessi e rigonfi degli assoni delle cellule a canestro circondano la parte inferiore dei corpi delle cellule di Purkinje con densi ‛canestri' - formati in genere da più di uno strato, in modo simile a quello in cui la rafia avvolge un fiasco di Chianti - dai quali deriva appunto il nome della cellula. Le ramificazioni terminali degli assoni delle cellule a canestro proseguono formando una sorta di ‛barba' lungo il segmento iniziale dell'assone delle cellule di Purkinje per varie decine di μm. Questo assetto sinaptico molto particolare è stato giustamente chiamato ‛a pennello' da Palay e Chan-Palay (v., 1974), e ricorda le sinapsi cosiddette ‛ad assone a cappuccio' dei neuroni di Mauthner dei vertebrati inferiori. Molte delle ipotesi fatte a proposito dei possibili meccanismi di trasmissione sono basate sull'esistenza di una stretta guaina (come messo in evidenza da Palay e Chan-Palay, 1974) formata dal denso sistema di fibre del pennello intorno al segmento assonico iniziale delle cellule di Purkinje (v. tav. III). La trasmissione tra cellula a canestro e cellula di Purkinje è comunque GABA-ergica secondo tutti i criteri accettati per le sinapsi a trasmissione chimica.
Gli assoni delle cellule a canestro hanno anche ramificazioni ascendenti che formano sinapsi con la superficie liscia dei dendriti principali delle cellule di Purkinje.
I due terzi esterni dello strato molecolare sono popolati di cellule stellate, che rappresentano il terzo tipo di interneuroni locali principali. Esse hanno corpi cellulari e ramificazioni dendritiche un po' più ridotti delle cellule a canestro, ma il modello generale delle arborizzazioni dendritiche è simile. I dendriti, scarsamente spinosi, sono compressi in direzione dell'asse della lamella e le proiezioni delle ramificazioni nel piano trasversale della lamella hanno forma ovoidale con diametro longitudinale verticale. Le arborizzazioni dendritiche sono l'immagine speculare di quelle delle più profonde cellule a canestro. Gli assoni delle cellule stellate possono o ramificarsi immediatamente o correre parallelamente alla superficie nel piano trasversale delle lamelle. Le branche terminali secondarie (o terziarie) degli assoni delle cellule stellate prendono contatto coi dendriti lisci più grandi delle cellule di Purkinje, sebbene talvolta possano formarsi sinapsi anche tra gli interneuroni. Nella parte più profonda dello strato molecolare alcune cellule - il cui corpo è situato tra il terzo medio e quello inferiore dello strato molecolare - hanno forma intermedia tra quella delle cellule stellate e quella delle cellule a canestro, nel senso che, in gran parte, le loro branche assoniche secondarie terminano come quelle delle cellule stellate, ma presentano anche alcune branche discendenti che partecipano alla formazione dei canestri pericellulari delle cellule di Purkinje.
L'alto contenuto in taurina della corteccia cerebellare, che si riduce drasticamente interferendo nella genesi di granuli e cellule stellate (ad esempio somministrando raggi X durante il primo periodo di vita postnatale), e anche la sensibilità dei dendriti delle cellule di Purkinje ad applicazioni locali di taurina hanno fatto avanzare l'ipotesi che questo amminoacido fosse il mediatore implicato nella trasmissione delle cellule stellate. Tuttavia l'alta immunoreattività a sieri anti-GAD e anti-GABA degli assoni e dei corpi delle cellule stellate non è facilmente riconducibile a questa ipotesi. La funzione delle cellule stellate è senz'altro inibitoria.
c) Organizzazione della corteccia cerebellare in zone sagittali
La suddivisione funzionale della corteccia cerebellare in zone sagittali - in alternativa all'evidente sistema anatomico di suddivisioni lobulari trasversali - è emersa gradualmente dalle prime osservazioni di Jansen e Brodal (v., 1940) di una grossolana suddivisione delle connessioni cortico-nucleari in tre zone sagittali: una zona mediale, una intermedia e una laterale si protendono da tre ampie zone corticali longitudinali verso i rispettivi nuclei cerebellari, e cioè quello mediale (del tetto o del fastigio), quello intermedio (interposito) e quello laterale. In successivi studi, molti altri autori e in particolare Voogd (v., 1964), usando analisi istologiche più raffinate e metodi di tracciatura anterograda e retrograda, hanno messo in evidenza un sistema di zone molto più complesso, consistente in sette zone indicate con le lettere A, B (nel verme), C1, C2, C3 (corrispondenti alla zona intermedia di Brodal e Jansen) e D1, D2 (corrispondenti alla zona laterale di Brodal e Jansen). Il dato più rilevante relativo alle zone sagittali C1-D2 è che esse si estendono senza soluzione di continuità attraverso le pliche laterali dei lobi ansiforme, paramediano e paraflocculare. Una suddivisione in zone essenzialmente analoga è stata messa in evidenza per le afferenze muschiose (per maggiori dettagli al riguardo v. Ito, 1984).
Nonostante la generale distribuzione zonale e pur nell'ambito di questa struttura schematica, la mappatura corticale delle risposte agli stimoli naturali mostra una distribuzione essenzialmente a chiazze, che a prima vista sembra casuale; tuttavia, un'analisi dettagliata della rappresentazione dell'area recettiva del trigemino, benché a chiazze, mostra l'esistenza di una distribuzione estremamente sistematica e sofisticata su ampie zone delia corteccia cerebellare, vagamente somigliante all'organizzazione periorale dei recettori cutanei nei nuclei sensitivi.
d) Modelli concettuali delle funzioni delle reti neuronali cerebellari
La conoscenza della regolarità quasi cristallina della rete neuronale del cervelletto fornisce un saldo supporto alla costruzione di modelli citoarchitettonici e di teorie sulla funzione cerebellare. Il primo di tali modelli è stato proposto da Braitenberg e Onesto (v., 1961) ed era basato sul presupposto che le fibre parallele agiscano come ‛linee di ritardo' in modo da convertire gli schemi di attività da spaziale in temporale. Secondo un'altra linea di pensiero, proposta da Szentàgothai (v., 1963 e 1965), l'eccitazione si diffonderebbe fondamentalmente lungo l'asse longitudinale delle lamelle, mentre gli interneuroni inibitori locali, grazie alle loro fitte connessioni con le due strisce di tessuto corticale a lato del fascio di eccitazione delle fibre parallele (azioni ‛fuori fascio'), avrebbero la funzione di focalizzare e limitare l'eccitazione a strisce ristrette nell'asse longitudinale delle lamelle. Osservazioni fisiologiche dirette sembrerebbero confermare quest'ipotesi, e pertanto il concetto della corteccia cerebellare come ‛macchina neuronale' è basato essenzialmente su di essa (v. Eccles e altri, 1967). La presenza di due distinti canali di ingresso alla corteccia cerebellare ha portato Marr (v., 1969) a suggerire l'esistenza di una ‛rete di apprendimento' essenzialmente all'interno del modello di funzionamento spaziale di Eccles e altri (v., 1967). Un più probabile modello, alternativo a quello di Marr, è quello introdotto da Albus (v., 1971). Per spiegare anche l'analisi degli schemi temporali, Fujita (v., 1982) ha cercato di combinare il principio di apprendimento di Marr e Albus con la teoria dei filtri adattativi. Questi ‛modelli di apprendimento' non spiegano la coordinazione per se stessa, in quanto rappresentano la funzione cerebellare in termini solo monodimensionali della ‛regolazione del guadagno' adattativo.
Tuttavia, recentemente è stato affermato che dati fisiologici sull'adattamento a breve termine erano a favore della teoria secondo la quale il cervelletto sarebbe un ‛dispositivo' per l'apprendimento (v. Ito, 1984).
I primi modelli avevano in comune il difetto di essere basati quasi esclusivamente su ciò che si presumeva accadesse nella corteccia cerebellare stessa. Benché i nuclei cerebellari (e vestibolari) fossero noti già da tempo come le principali linee in uscita dal cervelletto, solo più tardi venne compresa la loro importanza in qualità di principali centri a livello dei quali i segnali in entrata vengono smistati per la loro elaborazione in segnali in uscita. Dato che la corteccia cerebellare viene considerata al di fuori del contesto delle interazioni cortico-nucleari, e tanto più dell'intero sistema sensomotorio, gli studi teorici che mirano a ottenere un livello di integrazione maggiore rivestono particolare importanza.
Uno di tali studi, basato sull'idea che le microzone cerebellari siano unità funzionali, venne gradualmente prendendo forma dai lavori di Oscarsson (v., 1976), in cui erano analizzate le connessioni afferenti ed efferenti del lobulo V del verme nel cervelletto del gatto. Informazioni più approfondite furono ottenute da Ito e collaboratori (v. Ito, 1984) in studi sulle funzioni del flocculo in relazione ai riflessi vestibolo-oculomotori. Questa ipotesi presuppone l'esistenza di piccoli raggruppamenti di cellule di Purkinje (circa 500 di numero) di pochi millimetri di estensione sui quali converge pertanto un considerevole numero di fibre parallele eccitatorie e di interneuroni locali inibitori. Rispetto all'organizzazione neuronale di base, Ito (v., 1984) distingue due diversi ‛microcomplessi cortico-nucleari'. Nei microcomplessi di tipo V la disposizione neuronale è relativamente semplice, e poiché sia le afferenze muschiose che quelle rampicanti emettono collaterali verso le cellule nucleari cerebellari e vestibolari, il circuito neuronale per la ‛modulazione corticale' sarebbe semplicemente sovrapposto al circuito nucleare ‛riflesso' di base; questo tipo di accoppiamento sarebbe pertanto solo un modello di via collaterale e modificabile del circuito cerebellare. Nei microcomplessi di tipo C il circuito elementare viene complicato da una varietà di collaterali ricorrenti dalle efferenze nucleari; tali collaterali si dirigono in parte su un circuito locale inibitorio, su collaterali delle cellule di origine delle afferenze muschiose e rampicanti in entrata e anche - cosa più importante - direttamente alle rispettive microzone della corteccia cerebellare. Questa disposizione neuronale, d'altra parte, potrebbe servire anche a un gran numero di circuiti interni e più ampi a feedback che provvedono sia circuiti reverberanti che meccanismi per bilanciare il possibile effetto di ‛deriva' esercitato dai feedback eccitatori.
La teoria secondo la quale il sistema nervoso centrale agirebbe come una rete tensoriale e le sue applicazioni ai modelli del cervelletto (v. Pellionisz e Llinàs, 1980; v. Pellionisz, 1985) rappresentano un tentativo più ambizioso non solo di fornire una spiegazione funzionale della rete cerebellare cortico-nucleare, ma anche di considerare il cervelletto come una parte dell'intero sistema sensomotorio. Secondo questa teoria, il cervelletto agisce come un coordinatore che trasforma l'intenzione motoria dismetrica in esecuzione motoria vera e propria. In termini matematici questa è una trasformazione covariante-controvariante, che ha luogo attraverso un tensore metrico alimentato dalla rete cerebellare cortico-nucleare, di vettori multidimensionali espressi nel sistema di coordinate intrinseco al sistema nervoso centrale (SNC). Questo modello ha l'indubbio vantaggio di spiegare in modo particolarmente elegante, in sistemi riflessi ben studiati come quelli vestibolo-oculari, la trasformazione che avviene tra sistemi di riferimento - una caratteristica essenziale dei meccanismi sensomotori - e di avere una notevole capacità predittiva. Mentre l'interpretazione secondo la quale il cervelletto agirebbe come coordinatore multidimensionale è compatibile con gli studi effettuati in precedenza (v. Pellionisz, 1985), l'integrazione di rappresentazioni funzionali finora disparate in un modello del cervelletto generale e verificato richiede uno sforzo teorico e sperimentale tuttora in corso.
3. La corteccia cerebrale
a) Aspetti dello sviluppo
Come abbiamo visto nella parte introduttiva (v. cap. 1), i neuroni della corteccia cerebrale derivano dalle cellule postmitotiche nella zona ventricolare delle primitive vescicole cerebrali. Il processo di differenziamento inizia con un movimento verso l'esterno di queste cellule in direzione della zona marginale esterna della parete vescicolare. Le numerose cellule che si trovano in stadi diversi di migrazione formano una striscia relativamente larga di corpi cellulari sparsi a diversi livelli di profondità, chiamata zona intermedia. Solo un po' più tardi le cellule che sono migrate più esternamente verso la superficie formano uno strato compatto, chiamato ‛lamina corticale', posto immediatamente al di sotto dello strato marginale divenuto ora più sottile. Come abbiamo già visto, nella corteccia cerebellare le cellule che migrano più tardi dalla zona ventricolare, e che quindi arrivano più tardi nel luogo in cui si sono stabilite quelle migrate in precedenza, hanno la tendenza generale a oltrepassare lo strato formato dalle cellule precedenti e a stabilirsi al loro esterno. Le successive generazioni di cellule vanno pertanto ad aggiungersi in sequenza verso l'esterno alle file o strati di generazioni precedenti. Tale schema risulta particolarmente evidente nel caso della lamina corticale, in cui gli strati più profondi della nascente corteccia cerebrale sono quelli formati dalle cellule arrivate prima, e gli strati più superficiali si aggiungono uno dopo l'altro nella parte esterna della lamina.
Lo sviluppo di una nuova tecnica immunocitochimica, basata sulla dimostrazione della localizzazione fine (elettronmicroscopica) di una proteina acida fibrillare della glia (GFA), ha reso possibile stabilire le linee cellulari che danno origine a neuroni e cellule gliali, a cominciare dalle più precoci forme cellulari postmitotiche nella zona ventricolare (ZV nella fig. 14). Basandosi su questi studi, Rakic (v., 1981) ha proposto un nuovo profilo rappresentativo, secondo il quale cellule mitotiche sia positive sia negative alla GFA coesisterebbero nella zona ventricolare (mitotica). Le cellule GFA negative danno origine ai neuroni, che iniziano la loro migrazione verso la zona intermedia, raggiungendo quindi la lamina corticale; le cellule GFA-positive producono le cellule gliali radiali e in seguito - direttamente o indirettamente - gli astrociti protoplasmatici e fibrosi (cioè le cellule gliali epiteliali della parete - o corteccia in via di sviluppo - cerebrale). Le cellule gliali radiali mantengono le loro connessioni sia con la superficie interna (ventricolare) sia con quella esterna (piale) della parete cerebrale. I corpi cellulari rimangono localizzati nella zona ventricolare costituendo infine lo strato cellulare ependimale che riveste la superficie ventricolare. Secondo l'elegante teoria di Rakic (v., 1972 e 1981), le fibre della glia radiale rappresentano il substrato su cui migrano praticamente tutte le cellule (almeno tutte quelle derivate dal neuroepitelio originario). Questa teoria spiega come l'originale mosaico formato dalle cellule appartenenti alla stessa linea cellulare mantenga la propria topologia nella lamella corticale nonostante i radicali cambiamenti di forma cui va incontro la parete cerebrale durante le prime fasi dello sviluppo e anche successivamente. Questo meccanismo è illustrato nello stereodiagramma di Rakic e Goldman-Raltic (v., 1982). Questo diagramma spiega anche l'ipotesi avanzata da Mountcastle (v., 1978), secondo la quale gli stretti e verticali aggregati di cellule corticali - le cosiddette ‛minicolonne' - deriverebbero dalla stessa linea cellulare allineata e in movimento lungo un fascicolo gliale radiale. Una rappresentazione simile è implicata nel termine ‛colonna ontogenetica', introdotto da Edelman (v., 1978), che si riferisce alla stessa struttura. Tale meccanismo morfogenetico potrebbe senz'altro spiegare - almeno in parte - l'estremo ordine che si riscontra nelle connessioni corticali e la strana organizzazione verticale della rete neuronale della corteccia.
Successivamente, al di sopra (verso l'esterno) della zona ventricolare, si forma la cosiddetta ‛zona subventricolare', chiamata inizialmente ‛zona subependimale' e un po' più tardi lamina cellulare subependimale. Qui l'attività mitotica prosegue ancora per diverso tempo dopo che la lamina corticale si è formata e le sue cellule si trovano in avanzato stato di differenziamento e addirittura dopo la loro maturazione in veri e propri neuroni. Probabilmente la maggioranza delle cellule gliali (ectodermiche) di tutta la parete cerebrale si forma qui, come pure una parte significativa dei neuroni più piccoli, le cui proliferazione e migrazione possono proseguire per un considerevole periodo della vita postnatale di molti mammiferi, incluso l'uomo.
b) Architettonica della corteccia
La scoperta della struttura della corteccia cerebrale ebbe inizio dalle osservazioni di uno studioso italiano di medicina, Francesco Gennari, il quale nel 1782 scoprì una stria di colore chiaro (lineola albidior) che decorre, in direzione parallela alla superficie, nel mezzo della sostanza grigia corticale, nella superficie mediale del lobo occipitale del cervello. Tale stria fu ben presto osservata e descritta da altri autori del periodo (v. Vicq d'Azyr, 1786; v. Sömmering, 1788). Questa caratteristica stria bianca presente nel quarto strato della corteccia visiva primaria è in realtà visibile anche a occhio nudo ed è ancora nota col nome di stria di Gennari; è proprio a causa di questa stria che in seguito la corteccia visiva primaria venne chiamata ‛area striata'. Un fondamentale passo avanti fu compiuto solo molto più tardi, quando lo psichiatra francese Baillarger (v., 1840) mise in evidenza in sezioni non colorate della corteccia umana, grazie alla differenza di trasparenza, sei strati paralleli alla superficie. I due strati opachi che egli riuscì a identificare corrispondono a due strati, presenti quasi ovunque, di fibre tangenziali mielinizzate, detti striscia esterna e striscia interna di Baillarger o, secondo la nomenclatura moderna, lamina IV e Vb, rispettivamente. Nozioni assolutamente nuove sulla struttura della corteccia vennero ottenute con l'introduzione dei metodi di colorazione cellulare (prima col colorante naturale carminio; v. Berlin, 1858), grazie ai quali fu possibile individuare la generale disposizione delle cellule corticali in sei (da cinque a sette o più) strati tangenziali (paralleli alla superficie) più o meno distinti. La prima identificazione di tipi cellulari distinti piramidali, fusiformi, granulari (piccoli e poliedrici) - risale a questi primi studi, anche se solo con l'introduzione della ‛reazione nera' (precipitazione di cromato d'argento) di Golgi (v., 1873) fu possibile identificare con chiarezza la forma delle cellule nervose corticali (v. Golgi, 1883).
La mappa del cervello umano più generalmente accettata, anche perché poteva adattarsi a quella di altri mammiferi, primati e subprimati, fu quella pubblicata da Brodmann nel 1908 (e in versioni molto più precise nel 1912 e 1914). Questa mappa distingueva oltre 50 aree corticali, il che rappresenta una soluzione intermedia abbastanza realistica (come fu provato molto più tardi) tra un numero o troppo limitato o eccessivo; quest'ultimo è il caso di von Economo e Koskinas (v., 1925), che proponevano un numero di 107 aree, numero che porterebbe a escludere la possibilità o la ragionevolezza di qualunque tentativo di identificare più di pochi tipi cellulari principali sulla base di criteri strutturali (v. Bailey e von Bonin, 1951). L'interesse nei confronti della citoarchitettonica diminuì nel periodo compreso fra gli anni trenta e i tardi anni cinquanta, anche se la sua utilità per la neuropatologia umana e in certe branche della neurochirurgia, come pure negli studi comparativi, non è mai stata sottovalutata. Tuttavia, quando per lo studio delle funzioni corticali cominciò a diffondersi l'uso dei microelettrodi e anche quello dei traccianti, non solo furono confermate le primitive suddivisioni in aree architettoniche, ma poterono essere individuate suddivisioni territoriali più piccole di uno o più spesso due ordini di grandezza sulla base di finì criteri funzionali e di connessioni.
La fondamentale struttura in sei strati della ‛isocorteccia' cominciò a essere generalmente accettata in seguito agli studi di Brodmann (v., 1908, 1912 e 1914) e grazie anche all'unificazione della nomenclatura, suggerita dai Vogt (v. Vogt e Vogt, 1919) sulla base di un attento e sistematico confronto (sia per differenti regioni cerebrali che per specie differenti) della citoarchitettonica e della mieloarchitettonica. Tuttavia non è stato possibile raggiungere un accordo completo e una nomenclatura unificata, e per il momento è improbabile che ciò accada, specialmente riguardo alla suddivisione in sublamine delle varie regioni corticali. Inoltre, il rapido sviluppo di una gran quantità di tecniche chemoarchitettoniche determinerà probabilmente un'ulteriore confusione, rendendo ancor più difficile una visione sintetica che metta ordine nella valanga di nuovi dati. Ciò nonostante, nel quadro attuale dell'architettonica può essere incluso un dato nuovo; infatti, l'uso dei metodi di colorazione aldeide-fucsina per la lipofuscina (v. Braak, 1971) ha introdotto un criterio nuovo e diverso dai due preesistenti criteri architettonici, puramente morfologici: 1) la dimensione, forma e densità cellulare in citoarchitettonica; 2) il decorso, la densità e lo spessore delle fibre in mieloarchitettonica.
La struttura generale della isocorteccia è mostrata in modo molto elegante e chiaro nella fig. 19, che è un diagramma sintetico di Braak (v., 1984); nella figura i numeri romani si riferiscono alla nomenclatura delle lamine secondo la sequenza ormai tradizionale dall'esterno (superficie piale) all'interno per ciò che riguarda la citoarchitettonica, mentre i numeri arabi si riferiscono alla mieloarchitettonica. Per l'architettonica del pigmento (lipofuscina), sono stati mantenuti i numeri romani ma col prefisso ‛p' (pigmento). Le suddivisioni in seno a uno strato sono indicate da lettere dell'alfabeto latino.
Regioni corticali che si discostano dal consueto schema architettonico a sei strati erano già state individuate da Meynert (v., 1867-1868); a esse era stato dato il nome di ‛cortecce bianche' per lo strato di sostanza bianca che ricopre la loro superficie. I termini ‛isocorteccia' per le cortecce ‛grigie' a sei strati e ‛allocorteccia' per cortecce di altro tipo furono introdotti da Vogt e Vogt nel 1919; in base a studi embriologici e comparativi, si è d'accordo nel ritenere che l'isocorteccia si sia sviluppata di recente, principalmente nei Mammiferi, tanto che è definita ‛neopallio' per distinguerla da parti più arcaiche del cervello. La nomenclatura di queste parti arcaiche è abbastanza controversa e pertanto non verrà riportata in questo articolo. Sarà sufficiente ricordare che le due vescicole telencefaliche che protrudono da ambedue le parti del primitivo allargamento prosencefalico, che si trova all'estremità anteriore del tubo neurale, mantengono in gran parte la loro struttura arcaica nella zona circostante il foro di comunicazione tra la cavità originaria del prosencefalo (il terzo ventricolo) e le cavità delle vescicole emisferiche (ventricoli laterali). Questa zona circolare e i suoi derivati più tardivi sono pertanto nominati globalmente col termine di porzione (o lobo) ‛limbica' (dal latino limbus, margine) della corteccia.
c) Caratterizzazione dei sei strati principali della corteccia
La lamina I, o strato molecolare, è quasi completamente priva di cellule nervose, se si escludono pochi neuroni solamente inibitori, e contiene un numero variabile di assoni a orientamento tangenziale (lamina mieloarchitettonica).
Si tratta di uno strato quasi esclusivamente sinaptico, in cui il tessuto nervoso è formato in prevalenza dalle arborizzazioni dei dendriti apicali delle cellule piramidali. Questo strato (p1) è quasi del tutto privo di pigmento.
La lamina II, o strato corpuscolare, contiene piccoli corpi cellulari raggruppati piuttosto densamente, tra cui è difficile distinguere le cellule piramidali da quelle non piramidali, per ragioni che descriveremo in seguito. Il corrispondente strato mieloarchitettonico 2 è quasi del tutto privo di fibre mielinizzate. Nella rappresentazione pigmentoarchitettonica pII questo strato è in genere fortemente colorato. La presenza di grossolani granuli fortemente colorati e raggruppati fittamente fa presupporre che questa lamina sia ricca anche di cellule non piramidali.
La lamina III, o strato piramidale, è in genere la più spessa della corteccia e contiene in prevalenza corpi cellulari di forma piramidale (triangolari con l'angolo acuto in direzione della superficie); le cellule si dispongono in colonne verticali. La dimensione dei corpi cellulari aumenta gradualmente procedendo verso la parte più profonda dello strato. Nella corrispondente rappresentazione mieloarchitettonica si può osservare in molte aree uno strato abbastanza pronunciato di fibre tangenziali mielinizzate (la linea Kaes-Bechterew del sottostrato 31). La pigmentoarchitettonica mostra un aumento della densità di pigmentazione a grana fine (del tipo delle cellule piramidali) crescente con l'aumentare della profondità. Nelle aree primarie sensitive e motorie si riscontra un andamento inverso.
La lamina IV, o strato granulare, contiene corpi cellulari poliedrici relativamente piccoli. Questo strato è abbastanza sottile nella maggioranza delle regioni corticali, ma nelle aree della corteccia sensitiva primaria si allarga suddividendosi in diversi sotto- o sotto-sotto-strati. In mieloarchitettonica la lamina IV corrisponde alla stria esterna di Baillarger. Nella corteccia visiva primaria la stria di Gennari occupa una parte considerevole dell'intero strato, con l'eccezione di un sottile sotto-strato al di sopra e al di sotto di essa. Nelle aree corticali non sensoriali questa lamina è praticamente priva di pigmento, che è invece molto abbondante nella corteccia visiva, nella quale rende evidenti delle nette suddivisioni.
La lamina V, o strato ganglionare, contiene in prevalenza corpi cellulari piramidali con l'eccezione dello strato immediatamente confinante con la lamina IV, nel quale si riscontra con relativa frequenza la presenza di singoli corpi cellulari poliedrici di grandi dimensioni. Nel sotto-strato più profondo di questa lamina si trovano talvolta corpi cellulari piramidali eccezionalmente grandi, specialmente nelle regioni motorie primarie (cellule giganti di Betz). In mieloarchitettonica la parte più profonda di questo strato 5b contiene la stria interna di Baillarger.
La lamina VI, o strato multiforme, contiene corpi cellulari fusiformi e piramidali irregolari orientati verticalmente. La lamina è così chiamata per gli evidenti mutamenti di orientamento dei principali processi che dipendono dalla loro collocazione in prossimità delle concavità o delle convessità della corteccia. Come abbiamo già spiegato a proposito delle cellule di Purkinje nella corteccia cerebellare (v. tav. II), ciò è la conseguenza di una trasformazione cartesiana di tutti gli elementi tissutali con conseguenze non sempre immediatamente evidenti. La rappresentazione mieloarchitettonica è dominata dalla transizione verso la sostanza bianca; lo strato pigmentoarchitettonico pVIa è particolarmente ricco di pigmento a grana fine.
d) Tipi cellulari della isocorteccia
1. Neuroni piramidali. - Il tipo cellulare più caratteristico presente non solo nella neocorteccia (e anche in alcune parti della allocorteccia) ma addirittura nell'intero sistema nervoso dei Mammiferi è quello dei neuroni piramidali. Essi vengono così chiamati a causa della loro caratteristica forma a triangolo isoscele col vertice in direzione della superficie della corteccia, forma riconoscibile anche con semplici colorazioni cellulari. Il vertice si prolunga in un ‛dendrite apicale' o tronco dendritico, che nella maggioranza dei casi raggiunge il confine tra gli strati superficiali I e II della corteccia dove si divide a formare un ciuffo dendritico terminale. Il dendrite apicale dà origine a molte ramificazioni aventi corso orizzontale o lievemente ascendente. Il corpo cellulare ha la forma di una piramide allungata, anche se talvolta si riscontrano delle distorsioni specialmente negli strati corticali più profondi e in particolare nelle zone più interne dei solchi. La parte principale dell'arborizzazione dendritica ha origine nel corpo cellulare; il cosiddetto ‛dendrite basale' forma un'arborizzazione raggiata di forma sferica. La tav. IV mostra due cellule piramidali della parte profonda della lamina III della corteccia della scimmia colorate secondo il classico metodo di Golgi. L'osservazione al microscopio elettronico mostra che il citoplasma dei corpi cellulari piramidali e anche dei dendriti è relativamente chiaro a causa della relativa scarsità di organelli citoplasmatici (mitocondri, reticolo endoplasmatico rugoso), benché delicati granuli di lipofuscina contribuiscano al quadro pigmentoarchitettonico. Sia i corpi cellulari che i dendriti principali contengono molti microtubuli.
L'assone dei neuroni piramidali ha origine alla base del corpo cellulare e ha decorso discendente verticalmente. Probabilmente la maggioranza degli assoni piramidali si allontana dalla corteccia in direzione della sostanza bianca, mentre in precedenza si era ritenuto che gli assoni delle piccole cellule piramidali della lamina II terminassero localmente. Prima di divenire mielinizzati, gli assoni piramidali emettono un gran numero di collaterali che arborizzano in uno spazio abbastanza ampio intorno alla cellula di origine. L'osservazione di cellule piramidali colorate col metodo di Golgi ha fatto ritenere in un primo tempo che le ramificazioni delle collaterali assoniche fossero non specifiche; tuttavia esse possono coprire grandi distanze (fino a 1,5 mm e anche più). Pertanto, si ritenne che l'ampiezza totale delle arborizzazioni collaterali di un assone piramidale potesse coprire distanze di 3 mm e forse più. Quando è divenuta d'uso comune la tecnica di iniezione intracellulare con l'enzima perossidasi del rafano è risultato che le arborizzazioni delle collaterali assoniche delle cellule piramidali sono molto più specifiche, e in realtà arborizzano abbondantemente in zone ben definite del tessuto corticale circostante. Per tale ragione dovranno presto essere distinte classi completamente diverse di cellule piramidali.
A. Spine dendritiche. L'aspetto più caratteristico dei neuroni piramidali consiste nel fatto che la maggior parte dei loro dendriti sono lateralmente ricoperti di delicate appendici a forma di bacchetta di tamburo note col nome di spine dendritiche. Tali spine furono descritte già dai primi studiosi che usarono il metodo di Golgi - in particolare da Golgi (v., 1883) e Ramón y Cajal (v., 1899) - anche se solo con l'avvento del microscopio elettronico fu possibile riconoscere il loro ruolo di sito recettivo delle sinapsi che si formano nel punto di contatto tra le arborizzazioni terminali degli assoni che innervano quella particolare cellula piramidale. Il numero delle spine dendritiche e la loro presenza o assenza in certe parti dell'albero dendritico e in certe condizioni sperimentali e patologiche resero immediatamente evidente la loro importanza agli studiosi della corteccia cerebrale. La densità (numero per unità di lunghezza del dendrite) delle spine varia considerevolmente a seconda della specie, della regione corticale e del tipo di dendrite: tuttavia non è stato possibile stabilire una chiara relazione con la posizione della specie nell'albero filogenetico. È difficile trovare dati attendibili riguardo al numero delle spine: nelle più piccole cellule piramidali dovrebbe essere compreso tra un valore minimo, e improbabile, di 6.000 e uno massimo di 30.000. Questi dati concorderebbero con la media di 80.000 spine delle cellule di Purkinje. Il numero di sinapsi che ogni cellula piramidale riceve è superiore al numero delle spine; ciò è dovuto in piccola parte al fatto che alcune spine ricevono più terminazioni (2 o 3) da assoni presinaptici, e specialmente al fatto che anche le superfici interspinali del tronco dendritico ricevono molte terminazioni sinaptiche, particolarmente la parte iniziale e priva di spine dei dendriti; lo stesso accade nel caso del corpo cellulare e anche del segmento iniziale dell'assone. Per quello che si può giudicare sulla base della marcatura delle branche assoniche terminali con perossidasi del rafano, il numero di singoli bottoni terminali stabilito da un qualunque assone con una determinata cellula varia tra 1 e 10.
B. Distribuzione laminare. I neuroni piramidali sono presenti praticamente in tutte le lamine, a eccezione della lamina I della corteccia. Al confine tra lamina II e lamina III si riscontra un improvviso aumento della dimensione del corpo delle cellule piramidali, e quindi un aumento graduale di dimensione in tutta la lamina III. Nella lamina IV il numero dei neuroni piramidali varia grandemente a seconda delle aree corticali: nei campi sensitivi primari si trovano scarse cellule piramidali, mentre sono moltissime nelle regioni motorie e premotorie in cui una lamina IV ben distinta è virtualmente assente. La lamina V contiene cellule piramidali relativamente grandi (neuroni di grande taglia sono le cellule piramidali giganti di Betz della corteccia motoria); la varietà della dimensione, forma e orientamento delle cellule piramidali presenti nella lamina VI è talmente grande che diventa difficile decidere se una cellula sia o meno un vero neurone piramidale.
C. Agglomerati. La distribuzione delle cellule piramidali non è casuale, ma esse sono disposte molto sistematicamente in agglomerati (clusters). Il ‛nucleo' di ogni agglomerato è formato da un piccolo gruppo (3-5) di cellule piramidali grandi della lamina V (e VI), i cui corpi sono distribuiti in posizione sfalsata per una considerevole estensione in profondità, ma sufficientemente vicino a una linea radiale (verticale) della corteccia talché i loro dendriti apicali formano un fascio dendritico ascendente.
I dendriti apicali dei neuroni piramidali della più piccola lamina III, che sono un po' più sottili, entrano in contatto con la parte esterna dei fasci formati dai dendriti apicali. Benché molte o addirittura la maggior parte delle cellule della lamina II siano neuroni piramidali, esse non presentano dendriti apicali, dato che i dendriti diretti verso la superficie formano immediatamente delle arborizzazioni talché tali cellule, osservate in sezione verticale, appaiono ‛bicuspidate'. La distanza media tra gli ‛assi' degli agglomerati varia tra 50 e 100 μm. Nonostante la presenza di fasci dendritici, l'osservazione al microscopio elettronico non mette in evidenza la presenza di sinapsi (né di altre interrelazioni strutturali dirette) tra i dendriti apicali appartenenti allo stesso fascio. La spiegazione più prudente di tali agglomerati (fasci) di neuroni piramidali è che le cellule di un agglomerato abbiano un'origine comune da una determinata linea cellulare del primitivo neuroepitelio della zona germinativa ventricolare e si muovano lungo i fasci di glia radiale.
Come abbiamo già visto nel caso della corteccia cerebellare (v. tav. II), le pliche del tessuto corticale (chiamate lamelle nella corteccia cerebellare e ‛giri' o ‛solchi' in quella cerebrale) trovano riscontro nella geometria delle arborizzazioni dendritiche e, in minor misura, anche di quelle assoniche. Questo aspetto dell'architettura corticale è sempre stato messo chiaramente in luce già dagli autori classici, ma è raramente ricordato o discusso esplicitamente. La spiegazione più semplice e forse l'unica realistica di questi radicali mutamenti delle cellule e delle arborizzazioni è considerarli delle trasformazioni cartesiane di forma, come viene fatto molto spesso dagli studiosi che cercano di spiegare i cambiamenti di forma che si riscontrano nel corso della filogenesi. Tale principio fondamentale, adattato alla situazione della corteccia cerebrale, è chiaramente illustrato nel diagramma della fig. 22. In questo diagramma tale distribuzione è chiarissima nella lamina superficiale 1 - che risulta ispessita nella parte più profonda del solco e assottigliata nella parte convessa dei giri - e nella lamina VI, in cui la situazione è opposta (cioè si assottiglia nella parte più profonda del solco e si ispessisce nelle parti convesse della corteccia). Pertanto il decorso delle branche terminali dei ciuffi dendritici apicali è quasi verticale nella parte profonda dei solchi e quasi orizzontale nella parte superiore dei giri. Al contrario, il decorso principale dei dendriti basali diviene più verticale nella parte superiore dei giri e quasi completamente parallelo alla superficie nella parte profonda dei solchi. Logicamente tali cambiamenti sono meglio evidenti nella lamina I e specialmente nella lamina VI (e in grado minore anche nella lamina V).
2. Neuroni non piramidali. - Per ragioni di praticità la tassonomia moderna classifica tutti gli altri neuroni della corteccia cerebrale come ‛non piramidali'. Tale suddivisione, se da un lato costituisce il primo passo nella dicotomia logica della classificazione, ha peraltro poco significato, in quanto ricadono in questa categoria cellule che non hanno praticamente niente in comune. Purtroppo, dato che non vi è accordo su quale dovrebbe essere il passo successivo nella gerarchia tassonomica, la nomenclatura usata in questo articolo è arbitraria e riflette il punto di vista individuale dell'autore.
I neuroni non piramidali possono essere suddivisi in tre categorie pincipali: A, neuroni proiettivi ad assone lungo; B, neuroni proiettivi ad assone corto; C, interneuroni ad assone corto (veri). Le categorie B e C sono distinte in base a un criterio indubbiamente esile, e cioè sulla base del fatto che un neurone o è parte integrante di una catena neuronica attraverso la quale passano gli impulsi diretti a una qualsiasi parte della corteccia (e in tal caso può a ragione esser definito ‛proiettivo', cioè di tipo B), oppure, se costituisce una ‛linea laterale' (in genere inibitoria) per l'inibizione a feedforward o a feedback (disinibizione), viene definito vero interneurone. La catena neuronale principale adibita all'inoltramento dell'impulso può alla fine divenire anch'essa inibitoria - come avviene nella corteccia cerebellare - ma al momento non vi sono prove che questo accada nella isocorteccia. In questo articolo non useremo le definizioni ‛Golgi di tipo 1' (tipo Deiters) e ‛Golgi di tipo 2' per vari motivi: primo, perché i neuroni piramidali sono senz'altro i migliori ma non gli unici esempi di cellule di Golgi di tipo 1; secondo, perché la classica definizione ‛neuroni di Golgi di secondo tipo', anche se molto descrittiva, è di scarso aiuto per distinguere tra le categorie B e C citate in precedenza. Non adotteremo neppure la definizione ‛neuroni a circuito locale', al momento molto usata, perché, più che aiutare nella comprensione, ci sembra possa creare confusione.
A. Neuroni non piramidali proiettivi ad assone lungo. I dati sui neuroni non piramidali proiettivi ad assone lungo sono pochi e non molto attendibili. L'unica regione della corteccia nella quale neuroni stellati chiaramente non piramidali con dendriti spinosi danno origine a efferenze corticali è la corteccia visiva. Queste cellule sono state descritte già da Ramón y Cajal (v., 1899) nella corteccia visiva, e tali osservazioni sono state confermate da molti altri autori e più recentemente con traccianti retrogradi (v. neurobiologia: Trasporto assonale, suppl.). Non abbiamo a disposizione dati attendibili per quanto riguarda altre zone della corteccia sensitiva primaria.
Particolare incertezza si ha nel caso delle cellule della lamina VI, molte delle quali presentano efferenze in prevalenza dirette verso il talamo. Data l'irregolarità della forma e dell'orientamento dei loro corpi cellulari e dei loro dendriti principali, è difficile stabilire se queste cellule non dovrebbero o non potrebbero invece essere considerate neuroni piramidali. La maggior parte delle cellule fusiformi o irregolari di questo strato, avendo dendriti spinosi e un assone che penetra nella sostanza bianca, dovrebbero essere considerate veri neuroni piramidali.
B. Neuroni proiettivi ad assone corto. Le cellule stellate spinose sono risultate essere i principali neuroni bersaglio delle afferenze sensoriali specifiche. Esse sono localizzate nella lamina IV delle aree corticali sensoriali; hanno corpo piccolo, generalmente stellato, e dendriti relativamente brevi (in genere meno di 150 μm) ricoperti di spine e occupano spazi di forma sferica. Il loro assone ha prevalentemente origine sul lato rivolto verso la sostanza bianca e si suddivide immediatamente in numerose branche. Solo alcune delle diramazioni assoniche possono essere seguite per un certo tratto in direzione orizzontale, benché molte diramazioni collaterali abbiano termine a breve distanza dal corpo cellulare. Le principali branche assoniche risalgono o discendono (o tutt'e due) verticalmente verso gli strati superficiali e/o più profondi della corteccia. Il principale bersaglio delle arborizzazioni terminali dei neuroni stellati spinosi, oltre alle brevi branche assoniche orizzontali, è rappresentato dalle cellule delle lamine III, Va e VI. La tav. V cerca di dare una rappresentazione generale di una tipica cellula stellata spinosa della corteccia visiva della scimmia. Le cellule stellate spinose di altre regioni sensitive e sensoriali (somatosensitiva, acustica) sono molto simili in molte specie.
Le arborizzazioni assoniche della maggior parte dei neuroni stellati spinosi si mantengono entro una stretta colonna verticale di tessuto corticale, cosicché qualunque afferenza esse ricevano possono dirigere i loro impulsi solo verticalmente, o nello stretto spazio intorno ai loro corpi o nel tessuto corticale al di sopra o al di sotto.
Le cellule stellate spinose non mostrano alcuna reazione immunocitochimica ad anticorpi anti-GAD e anti-GABA, il che è in accordo con l'asimmetria dei contatti della membrana e con l'aspetto sferico delle vescicole di tutti i bottoni terminali identificati dei neuroni stellati spinosi. Tali neuroni sono pertanto sicuramente cellule proiettive che appartengono alla catena neuronica che continua le afferenze sensoriali specifiche in entrata.
Nelle lamine II/III della corteccia sensoriale e anche nelle lamine IV e V è stata riscontrata la presenza di ‛neuroni bipolari' a orientamento strettamente verticale. I due dendriti principali, che presentano solo scarse ulteriori diramazioni, risalgono o discendono verticalmente e possono estendersi per un buon tratto negli strati confinanti e anche in quelli più lontani della corteccia. I dendriti hanno poche o nessuna spina; l'assone in genere ha origine dal dendrite principale discendente e ha decorso verticale, e solo le sue branche terminali decorrono orizzontalmente, ma terminano a distanze relativamente brevi.
Poiché praticamente tutte le cellule che mostrano forti reazioni con anticorpi contro il polipeptide vasoattivo intestinale (VIP) sono bipolari e hanno aspetto del tutto simile alle cellule bipolari colorate col metodo di Golgi, si ritiene generalmente che tale poli peptide sia il principale trasmettitore delle cellule bipolari. Il VIP, applicato direttamente con microcannule, agisce come una sostanza eccitatoria, per cui si ritiene che queste cellule abbiano funzione eccitatoria. Anche la posizione nella lamina IV di molte cellule bipolari o, se non altro, quella dei loro dendriti che vi entrano e/o la traversano, potrebbe essere interpretata a favore dell'ipotesi che esse, alla stregua delle cellule stellate spinose, ricevano impulsi da afferenze specifiche e li trasmettano in direzione verticale verso strati più superficiali o più profondi della corteccia. Le cellule bipolari non sono GABA-ergiche, ma molte contengono l'enzima colina-acetiltransferasi, il che parrebbe deporre a favore della loro natura colinergica.
C. Interneuroni ad assone corto. Al presente, tutti i veri interneuroni conosciuti della isocorteccia hanno natura inibitoria. Vari tipi di tali interneuroni inibitori sono già stati individuati e descritti in base alle caratteristiche delle loro arborizzazioni (in particolare quelle assoniche) dagli autori classici, e specialmente da Ramón y Cajal (v., 1899). Alcuni tipi sono stati recentemente studiati e definiti con precisione grazie all'uso di tecniche più avanzate, in particolare con la marcatura intracellulare con perossidasi del rafano. Tali tipi sono: a) cellule a canestro, con almeno tre sottotipi diversi (a1-a3); b) cellule a doppio pennacchio (double bouquet); c) cellule asso-assoniche o a candelabro. Vi sono anche altri tipi cellulari che sono stati però definiti meno chiaramente, in parte perché non è stato finora possibile ottenere una buona marcatura intracellulare, in parte perché le loro caratteristiche morfologiche, in particolare lo schema delle loro arborizzazioni assoniche, per quanto tipico, non si presta a stabilire una chiara separazione tipologica. Tali tipi sono i seguenti: d) cellule neurogliformi (‛nane' o a ‛ragnatela'); e) cellule di Golgi di secondo tipo lisce o scarsamente spinose che non presentano un ben definito schema di arborizzazione dei loro assoni (questo è di gran lunga il tipo più comune, probabilmente il più frequente e senz'altro il più ubiquitario tipo di interneurone); f) cellule con assone a ciuffo (axonal tuft cells); e infine g) cellule dendroassoniche.
Cellule a canestro. Le cellule a canestro e due dei loro sottotipi sono state descritte da Ramón y Cajal (v., 1899 e 1911). Dopo essere state dimenticate per lungo tempo, sono state ‛riscoperte' recentemente dagli studiosi della corteccia cerebrale e sono divenute uno dei tipi cellulari meglio definiti in base a svariati criteri.
Le cellule a canestro grandi (a1) sono riconoscibili sia per le loro dimensioni che per il caratteristico decorso orizzontale (parallelo alla superficie o allo strato) dei loro assoni principali. Nella maggior parte delle regioni corticali è adesso possibile distinguere due popolazioni principali di queste cellule (v. Somogyi e altri, 1983; v. Kisvàrday e altri, 1985 e 1987) marcando intracellularmente alcuni esemplari rappresentativi. Le cellule a canestro grandi formano due file, una al confine tra le lamine III e IV, e l'altra al confine tra le lamine IV e V. Il loro albero dendritico è a raggiera e, nell'adulto, i dendriti non hanno spine o ne hanno solo poche. La principale arborizzazione assonica si estende orizzontalmente (per 2 o 3 mm) al livello nel quale è localizzato il corpo cellulare (v. fig. 23) e solo un'unica branca principale si diparte dal segmento assonico iniziale seguendo un percorso verticale, ascendente o discendente a seconda che la cellula si trovi nella riga inferiore o in quella superiore.
Le branche assoniche terminali, che si dividono a formare i bottoni sinaptici, sono in genere orientate verticalmente e hanno decorso ascendente nelle cellule della fila inferiore; pertanto, le cellule della fila superiore entrano preferenzialmente in contatto con le cellule della lamina III e quelle della fila inferiore con quelle della lamina V, cioè con cellule piramidali dello strato V (e VI). Benché il principale bersaglio delle cellule a canestro siano le cellule piramidali - come era stato correttamente ipotizzato da Ramón y Cajal - solo il 30% dei bottoni entra realmente in contatto coi corpi cellulari delle cellule piramidali (v. tav. VI), circa altrettanto con la parte liscia (iniziale) dei dendriti e un altro 30% con le spine dendritiche. Il rimanente 10% circa entra in contatto con interneuroni che sono stati identificati come inibitori.
Le cellule a canestro grandi sono GABA-ergiche e le loro sinapsi hanno caratteristiche ultrastrutturali che corrispondono a quelle che normalmente si riscontrano in sinapsi inibitorie.
Le cellule a canestro ‛a breve raggio' o ‛colonnari' (a2) sono state scoperte da Ramón y Cajal (v., 1899) che le denominò ‛a breve raggio' in opposizione alle cellule a canestro grandi (a1) che hanno assoni a decorso lungo. Cellule di tale tipo esistono realmente, in particolare nella lamina II; tuttavia è stato osservato solo in seguito che benché gli assoni di tali cellule a canestro abbiano davvero decorso breve in direzione orizzontale, in direzione verticale possono arrivare a distanze considerevoli (v. Szentàgothai, 1975). Fu allora introdotto il termine di cellule a canestro ‛colonnari'. Si tratta di cellule a canestro tipiche, con dendriti a estensione relativamente breve, che sono localizzate nella lamina II, o piuttosto al confine tra le lamine II e III. Il loro assone in genere ha decorso verticale discendente ed emette branche terminali laterali di non più di 50-75 μm raggruppate a forma di canestro. Il corso discendente della branca assonica principale continua attraverso l'intera lamina III e anche nella lamina IV senza emettere nessun'altra branca terminale laterale; dopo il suo ingresso nella lamina V comincia nuovamente a emettere rami collaterali terminali aventi caratteristiche simili a quelle osservate nella lamina III. L'arborizzazione assonica di questo tipo di cellula si estende pertanto in forma evidentemente colonnare, in una zona cilindrica di tessuto corticale larga circa 100-150 μm.
Questi neuroni sono anch'essi GABA-ergici; tuttavia sembra che essi contengano anche colecistochinina (CCK), per cui è abbastanza semplice individuare e seguire le arborizzazioni di questo tipo di cellule con colorazioni basate sulla immunoreattività anti-CCK.
Recentemente è stato scoperto ancora un altro tipo di cellule a canestro (v. Kisvàrday e altri, 1985), chiamate clutch cells. Si tratta di tipiche cellule a canestro e probabilmente molte di quelle descritte nella letteratura classica appartengono a questo tipo. Esse si trovano in grande quantità nella lamina IV della corteccia sensitiva; le loro branche assoniche principali hanno in genere decorso parallelo alla superficie, ma attraverso le interconnessioni con le branche assoniche verticali la loro arborizzazione riempie praticamente l'intera lamina IV.
I contatti sinaptici stabiliti dalle loro terminazioni assoniche, anche se sembrano seguire il solito schema e sembrano connettersi principalmente con i corpi delle cellule piramidali, sono distribuiti in modo analogo a quelli delle cellule a canestro grandi: solo un terzo entra in contatto con i corpi delle cellule piramidali, un terzo coi dendriti più grandi (principalmente di cellule piramidali) e un terzo con spine dendritiche. Meno del 10% delle sinapsi viene stabilito con interneuroni presumibilmente inibitori. Le clutch cells sono chiaramente GABA-ergiche, e pertanto hanno natura inibitoria.
Cellule a doppio pennacchio (double bouquet cells). Questo tipo cellulare molto caratteristico è stato descritto per la prima volta da Ramón y Cajal, il quale gli attribuì questo nome principalmente a causa dei due ciuffi di dendriti ascendenti e discendenti verticalmente. L'aspetto più caratteristico, tuttavia, consiste nella lunga banda verticale, costituita dalle loro branche assoniche, che attraversa praticamente l'intero spessore della corteccia, dalla lamina II fino a un buon tratto della lamina VI. La stretta banda di branche assoniche somiglia a una ‛coda di cavallo' (v. tav. VII). Le cellule hanno dimensioni medie, il loro albero dendritico ha di preferenza, ma non necessariamente, orientamento verticale, e talvolta i dendriti hanno decorso ricorrente, cosa che è veramente tipica delle arborizzazioni assoniche. Queste cellule sono localizzate principalmente negli strati II-III.
Le cellule a doppio pennacchio e i loro processi sono fortemente immunoreattivi per il GABA e anche per la CCK. L'immunoreattività alla CCK rende facile seguire al microscopio ottico l'intera arborizzazione di tali cellule, perché solo una minoranza di tutti i neuroni immunoreattivi al GABA sono positivi anche per la CCK. Una valutazione statistica della distribuzione degli ingrossamenti terminali degli assoni delle cellule a doppio pennacchio colorati col metodo di Golgi indica che circa il 50% di essi stabilisce contatti con dendriti privi di spine; poiché questi ultimi appartengono con molta probabilità a interneuroni inibitori si potrebbe pensare che le cellule a doppio pennacchio abbiano funzione specificamente disinibitoria. Tuttavia, quasi il 50% dei bottoni delle cellule a doppio pennacchio è in contatto con spine dendritiche o col segmento iniziale liscio dei dendriti basali dei neuroni piramidali, per cui la funzione di queste cellule potrebbe essere sia inibitoria (nel caso delle cellule piramidali) sia disinibitoria (nel caso degli interneuroni inibitori).
Cellule asso-assoniche o ‛a candelabro'. Questo tipo di neuroni è per ora l'unica categoria ben definita di cellule che è sfuggita all'osservazione di Ramón y Cajal (v., 1899 e 1911). La presenza di queste cellule è stata riscontrata per la prima volta nella corteccia visiva del gatto, ma solo un po' più tardi esse sono state riconosciute come un tipo specifico di neuroni (v. Szentàgothai e Arbib, 1974; v. Szentàgothai, 1975). Una volta richiamata l'attenzione alle caratteristiche strutture terminali verticali dei loro assoni, che presentano da 3 a 7 bottoni in fila strettamente verticale, è stato facile riconoscere queste cellule. La parola ‛candelabro' è stata suggerita dalla peculiare disposizione verticale dei segmenti assonici terminali, che si dirigono verso l'alto da un'arborizzazione assonica disposta in una tipica ‛ghirlanda'. Solo dopo che Somogyi (v., 1977) mise a punto una nuova strategia per individuare in sezioni seriate al microscopio elettronico i dettagli delle arborizzazioni assoniche osservati al microscopio ottico, fu possibile rilevare la specifica collocazione delle terminazioni assoniche sul segmento iniziale degli assoni delle cellule piramidali. Poiché è stato riscontrato che il 1000/ delle terminazioni di tali neuroni finisce sui segmenti assonici iniziali, a questo tipo cellulare è stato attribuito il nuovo nome di cellule ‛asso-assoniche' (v. tav. VIII).
La maggioranza delle cellule asso-assoniche individuate finora è localizzata nella parte superiore della lamina III, e anche le loro terminazioni assoniche si trovano in prevalenza nella lamina III. Tuttavia, poiché anche nelle lamine V e VI sono state osservate arborizzazioni assoniche terminali del tutto simili , è possibile che questo tipo di cellule si trovi in tutte le parti della corteccia in cui sono presenti cellule piramidali.
Le file terminali di bottoni sinaptici asso-assonici, come pure le cellule e i loro dendriti e assoni, mostrano una forte immunoreattività ad anticorpi diretti contro la GAD e il GABA, per cui questo tipo cellulare è senz'altro inibitorio. Anche l'ultrastruttura delle terminazioni sinaptiche è in accordo con i criteri classici: le strutture specializzate di giunzione sulla membrana hanno carattere simmetrico e le vescicole sinaptiche sono ovoidali.
Le cellule asso-assoniche sono uniche per molti versi: esse rappresentano infatti l'unico tipo cellulare con una specificità del 100% verso un preciso bersaglio e risulterebbero particolarmente adatte a bloccare completamente ed efficacemente gli impulsi generati da una cellula piramidale. Tutte le altre terminazioni inibitorie in contatto con una cellula eccitatoria, come ad esempio una cellula piramidale, costituiscono solo una minoranza dei contatti sinaptici. In particolare, tutte le azioni sinaptiche che hanno origine in qualunque parte distale di una cellula, per esempio su un dendrite distale, devono commisurarsi con le altre azioni che avvengono simultaneamente in molti siti diversi dello stesso neurone, prima che possa essere stabilito il bilancio netto tra eccitazione e inibizione nel sito di generazione dell'impulso, cioè il segmento iniziale dell'assone. Le sinapsi inibitorie sono quasi sempre molto opportunamente localizzate in funzione della massima efficacia. Le sinapsi inibitorie sulle spine dendritiche non sono mai sole; esse si trovano sempre sulla parte basale delle spine, cioè ‛a valle' dell'apice della spina stessa che riceve sempre un'innervazione eccitatoria. Le sinapsi asso-somatiche, specialmente quelle sui neuroni piramidali, sono formate quasi esclusivamente da cellule a canestro, cioè sono inibitorie. Il corpo cellulare rappresenta una buona collocazione per determinare un'azione inibitoria, ma il segmento assonico iniziale è una collocazione ancora migliore. Sarebbe peraltro insensato ritenere che una terminazione asso-assonica attiva (cioè una fila di bottoni che appartengono alla stessa terminazione assonica) possa essere sufficiente a bloccare la produzione di un impulso condotto dall'assone di una cellula piramidale, qualunque possa essere il bilancio netto di influenze eccitatorie e inibitorie che agiscono su questa cellula. Se la situazione fosse questa, una sola sinapsi asso-assonica per ogni segmento assonico iniziale sarebbe sufficiente. Invece si è visto subito che c'è una convergenza di 5 a 1 su ogni segmento assonico iniziale, cioè ci sarebbero 5 cellule asso-assoniche per ogni cellula piramidale; sulla base di calcoli che hanno naturalmente un notevole grado di discordanza, una cellula asso-assonica darebbe origine a diverse centinaia di file di bottoni terminali. Tale disposizione avrebbe senso presupponendo che con un sufficiente grado di eccitazione netta sarebbero necessarie fino a 5 cellule asso-assoniche attive simultaneamente per bloccare del tutto la generazione di impulsi da parte di questa cellula piramidale.
Cellule neurogliformi (‛nane' o ‛a ragnatela'). La descrizione migliore e forse anche la più accurata di questo tipo di cellule è stata fornita da Ramón y Cajal (v., 1899); nelle descrizioni più moderne, tuttavia, c'è una certa confusione con alcuni interneuroni più piccoli che dovrebbero essere più correttamente inclusi nelle categorie che descriveremo in seguito. Il termine ‛neurogliforme', ideato da Ramón y Cajal, si riferisce specificamente alle piccole dimensioni dei corpi di queste cellule e ai numerosi dendriti ‛ondulati' (o talvolta ricorrenti) a orientamento radiale che danno all'albero dendritico l'aspetto di una densa sfera del diametro di 100 μm. È stato in alcuni casi sostenuto che queste cellule non hanno assone, ma spesso, pur se con qualche difficoltà dovuta al fatto che l'assone in genere arborizza nello stesso spazio sferico dell'albero dendritico, è stato possibile individuare un assone vero e proprio (anche in cellule simili del talamo e del tronco dell'encefalo). Queste cellule sono probabilmente GABA-ergiche.
Interneuroni lisci o scarsamente spinosi (di tipo comune). Questo è il tipo di interneurone più comune: è presente praticamente in tutti gli strati e in tutte le aree corticali ed è anche quello che presenta la meno tipica disposizione delle arborizzazioni assoniche e dendritiche. Corrisponde quasi perfettamente alla descrizione dei neuroni del secondo tipo fatta da Golgi (v., 1883). L'assone ha in genere origine nella superficie esterna (quella, cioè, rivolta verso la superficie piale) del corpo cellulare, ma le diramazioni assoniche iniziano ben presto ad assumere l'aspetto di una fontana, discendendo obliquamente come a coprire l'intera regione del corpo cellulare con un ombrello. Queste cellule sono GABA-ergiche, ma finora non si è riusciti a identificare le cellule con cui stabiliscono contatti sinaptici le loro diramazioni assoniche.
Cellule con assone a ciuffo. Queste cellule sono state descritte, chiaramente illustrate e così denominate da Ramón y Cajal (v., 1911). Sono cellule presenti caratteristicamente nello strato cellulare esterno della lamina II, per cui le loro arborizzazioni assoniche si trovano principalmente nella lamina I. La loro funzione inibitoria è stabilita solo sulla base di prove indiziarie.
Cellule dendro-assoniche. Questo tipo di cellule è stato identificato e descritto solo recentemente (v. Szentàgothai, 1979), e dato che mancano altre descrizioni, particolarmente al microscopio elettronico, si potrebbe evitare del tutto di parlarne se non fosse per il fatto che questo è l'unico tipo di cellula finora riscontrato solo nella corteccia dei Primati. Hanno la caratteristica di avere arborizzazioni assoniche e dendritiche molto simili (come capita talvolta nel caso delle cellule neurogliformi), ma un'attenta osservazione ha rivelato quasi sempre la presenza di un unico, caratteristico assone. Abbiamo ricordato questo tipo di cellule solo per sottolineare il fatto che con l'enumerazione dei tipi classici di cellule siamo ben lontani dall'averli descritti tutti.
e) Connessioni interneuronali
Fino a pochissimo tempo fa si sapeva ben poco delle connessioni interneuronali della neocorteccia. Già gli autori classici, e in particolare Ramòn y Cajal (v., 1899 e 1911), avevano correttamente ritenuto che le afferenze sensoriali specifiche terminassero nella lamina IV, ed era noto che le cellule piramidali sono gli elementi efferenti della neocorteccia. Tuttavia, solo con l'avvento del microscopio elettronico, quando cioè furono individuate le relazioni sinaptiche tra assoni e spine dendritiche (v. Gray, 1961), è stato possibile cominciare a chiarire le connessioni neuronali di base nella neocorteccia. Da allora si è verificato un impressionante sviluppo dell'istologia corticale, al quale hanno contribuito i seguenti fattori: 1) l'introduzione di nuove tecniche di marcatura (quella anterograda, che utilizza la captazione di amminoacidi triziati e la loro visualizzazione con autoradiografia; quella retrograda, che utilizza la captazione di coloranti fluorescenti e di perossidasi del rafano; quella anteroe retrograda, che utilizza l'iniezione intracellulare di perossidasi del rafano); 2) la combinazione, al livello di microscopio ottico ed elettronico, di vecchie e nuove tecniche; 3) lo sviluppo e l'integrazione nel repertorio delle metodologie neuroistologiche della biochimica enzimatica e dell'immunocitochimica per l'identificazione dei vari trasmettitori; 4) infine, ma non meno importante, la marcatura mediante iniezione intracellulare o intraassonica di perossidasi del rafano di cellule nervose e assoni identificati da un punto di vista fisiologico. Questo sviluppo delle metodologie avvenuto negli ultimi 15 anni ha già cambiato radicalmente le nostre conoscenze che prevediamo continueranno ancora a evolvere a ritmo serrato.
1. Catene neuronali per la conduzione dell'impulso dalle afferenze alle efferenze. - L'uso della degenerazione assonica ha provato che le afferenze talamiche specifiche (dal nucleo genicolato, nel caso del sistema visivo e acustico) hanno termine preferenzialmente nella lamina IV, anche se, in molte aree sensoriali della corteccia, possono talvolta arrivare alla lamina III e anche alla lamina I. Le tecniche della degenerazione assonica hanno messo in evidenza che vi erano terminazioni di afferenze anche in strati più profondi (VI) della corteccia. Una visione completamente nuova, particolarmente nella corteccia visiva, è emersa dalla marcatura intra-assonica di afferenze sensoriali identificate da un punto di vista fisiologico. Da allora si è rapidamente accumulata una tale quantità di dati relativi alle terminazioni di afferenze specifiche (specialmente visive) che anche un breve resoconto riempirebbe tutto lo spazio a nostra disposizione; pertanto ricorderemo solo alcuni fatti relativi alle meglio conosciute afferenze della corteccia visiva del gatto.
Esistono due tipi di afferenze, X e Y, che hanno origine da due ben definiti tipi di cellule della retina e del nucleo genicolato dorsale (v. visione). Le afferenze di tipo X terminano in gran parte nell'area 17 della corteccia visiva, mentre alcune, che si trovano al limite dell'area 17 verso l'area 18, oltrepassano il confine tra le due o in seno alla corteccia stessa o, tramite branche collaterali emesse dall'assone principale, nella sostanza bianca. Le terminazioni assoniche si trovano generalmente confinate nella parte superiore del quarto strato (IVa), benché anche nella lamina VI sia presente una ricca arborizzazione terminale. Le afferenze Y hanno arborizzazioni maggiormente estese e molte entrano nell'area 17 o nella 18 o in ambedue. Le terminazioni assoniche non sono confinate nella lamina IVa ma se ne trovano molte anche nella lamina IVb. Un numero molto maggiore di branche terminali entra nella lamina III, mentre è minore il numero di quelle che terminano nella lamina VI. Le arborizzazioni terminali delle afferenze X sono in genere limitate a un singolo albero e sono densamente raggruppate nella lamina IVa, e solo quelle che raggiungono il confine tra l'area 17 e l'area 18 mostrano raggruppamenti terminali abbastanza fitti e separati. Le terminazioni delle afferenze Y si suddividono in alberi separati. La grande maggioranza delle espansioni terminali entra in contatto con le spine dendritiche delle cellule stellate spinose, ma le terminazioni delle afferenze Y stabiliscono contatti anche con i dendriti basali dei neuroni piramidali della lamina III. Tramite la colorazione simultanea dello stesso materiale con il metodo di Golgi e con la colorazione immunocitochimica delle cellule GABA-ergiche, si è fatta strada la convinzione (v. Freund e altri, 1985) che i due tipi di afferenze (X e Y) abbiano termine su due diversi gruppi di interneuroni inibitori. Per ciò che riguarda la catena eccitatoria (cellule stellate spinose e piramidali) non è stato possibile individuare un'analoga separazione delle linee afferenti.
2. Schema generale delle connessioni degli interneuroni inibitori. - Purtroppo sappiamo molto poco delle principali afferenze che arrivano ai molti tipi di interneuroni mibitori che sono stati identificati. L'unica cosa che per ora sembra stabilita è stata ricordata nel punto precedente, e cioè che nella corteccia visiva del gatto le afferenze visive X e Y hanno termine su due gruppi diversi di interneuroni inibitori. Benché anche quella indagine così particolare non abbia permesso di identificare quale sia il tipo di neurone interessato, è molto probabile che almeno un tipo - per lo meno nel caso delle afferenze Y - sia rappresentato dalle grandi cellule a canestro della fila superiore localizzate al confine tra le lamine III e IV. Gli interneuroni inibitori più piccoli potrebbero essere le vere e proprie cellule a canestro della lamina IV, le cosiddette clutch cells, e/o uno qualunque degli interneuroni inibitori più piccoli che si trovano in grande abbondanza nella lamina IV (e nelle lamine adiacenti III e V), e cioè le cellule ‛scarsamente spinose' e le ‛microgliformi', gruppi ambedue abbastanza ben rappresentati nelle aree sensoriali corticali.
L'ipotesi che praticamente tutti davano per scontata, e cioè che una delle principali fonti di afferenze agli interneuroni inibitori fosse rappresentata da collaterali iniziali (locali) degli assoni delle cellule piramidali, è stata messa in discussione, almeno nel caso delle cellule piramidali della lamina III, dai recenti studi di Kisvàrday e altri (v., 1986) secondo i quali meno del 5% di tutti i bersagli sinaptici delle collaterali delle cellule piramidali sarebbero cellule GABAergiche, cioè interneuroni inibitori. Poiché le cellule piramidali rappresentano il 60% delle cellule nervose della neocorteccia, e poiché praticamente tutti gli assoni delle cellule piramidali presentano un rilevante numero di collaterali locali, bisognerebbe presumere l'esistenza di un feedback eccitatorio di eccezionale ampiezza da cellule piramidali verso altre cellule piramidali; però, poiché la popolazione totale di interneuroni è solo il 20%, il 5% delle terminazioni delle collaterali piramidali rappresenterebbe già una notevole porzione del totale di afferenze agli interneuroni inibitori. In ogni modo è ancora ignoto come si evitino i feedback positivi (runaway) nella rete corticale.
Data l'apparente prevalenza di feedback eccitatorio positivo da cellule piramidali, attraverso collaterali piramidali, ad altre cellule piramidali, le cellule asso-assoniche potrebbero avere l'importantissimo ruolo di bloccare le efferenze delle cellule piramidali ogniqualvolta l'eccitazione supera un determinato limite di sicurezza. In ogni modo, dato che non sappiamo come le cellule asso-assoniche possano essere implicate nell'azione eccitatoria, per il momento non possiamo dare nessuna spiegazione logica di questo meccanismo così importante.
f) Il principio dell'architettonica modulare
Anche se già nel 1938 Lorente de Nò aveva chiaramente individuato il fatto che le connessioni intracorticali avevano prevalentemente orientamento verticale, ci vollero altri venti anni prima che fosse possibile chiarire definitivamente l'organizzazione funzionale verticale (colonnare) della corteccia. Ciò fu possibile grazie agli studi di Mountcastle (v., 1957) e Hubel e Wiesel (v., 1959) che misero in luce la fondamentale somiglianza nelle proprietà dei campi recettivi delle cellule che erano disposte in spazi verticali organizzati a ‛colonne' nel tessuto corticale. (I principi generali di questa disposizione sono stati descritti nell'articolo visione a proposito della corteccia visiva). I dati istologici per la neocorteccia erano in buon accordo con i requisiti necessari a tale principio di organizzazione colonnare (v. Szentàgothai, 1969); inoltre, i progressi compiuti nel tracciare un'esatta mappa delle afferenze visive corticali grazie alle più moderne tecniche per evidenziare le vie da esse seguite (v. Hubel e Wiesel, 1972) dettero un ulteriore contributo al chiarimento ditale organizzazione, almeno nella corteccia visiva. Poiché le vie visive che hanno origine dai due occhi sono separate praticamente fino al primo relè corticale, fu possibile fare una mappa (nella lamina IV) degli spazi alternati connessi con l'occhio ipsilaterale e con quello controlaterale. Risultò allora che gli spazi tissutali monoculari della corteccia visiva primaria non erano in realtà colonnari, ma assomigliavano piuttosto alle strisce di una zebra se guardati dalla superficie, che le linee di separazione tra gli spazi connessi con i due occhi erano orientate verticalmente e che un certo grado di separazione tra le linee continuava a sussistere verso la superficie fino agli strati III e II, e in profondità attraverso gli strati V e VI. Come si poteva logicamente dedurre anche sulla base dell'organizzazione generale delle connessioni tra i neuroni intracorticali, un qualunque schema di impulsi che arriva tramite afferenze specifiche alla lamina IV viene trasferito seguendo uno schema ascendente e discendente in senso verticale, in cui la monocularità diminuisce gradualmente, ma le proprietà del campo recettivo rimangono inalterate.
Nonostante alcuni precedenti tentativi a opera di Hubel e Wiesel (v., 1959) di spiegare un'altra proprietà peculiare dei campi recettivi, e cioè la sensibilità specifica a una determinata direzione dell'orientamento della fonte dello stimolo visivo - la cosiddetta ‛specificità di orientamento' -, presumendo l'esistenza di un sistema di partizioni situate ad angolo retto rispetto al sistema delle strisce monoculari, il problema del rapporto tra ‛colonne della dominanza oculare' e ‛colonne di orientamento' è rimasto controverso. Ulteriori complicazioni si sono determinate con l'introduzione di nuove tecniche di marcatura dei neuroni e anche con alcune recenti osservazioni di fisiologia. L'impiego di 2-desossiglucosio radioattivo (uno zucchero che viene captato dalle cellule nervose in attività ma non metabolizzato) ha offerto qualche possibilità di marcare quelle cellule che sono state attivate in alcune specifiche condizioni sperimentali. Per ora, purtroppo, il livello di risoluzione della marcatura cellulare non è ancora tale da rendere visibili i profili delle attività cellulari in casi così complessi come quello del sistema delle colonne di orientamento.
Un'altra tecnica, risultata adatta a rivelare le discontinuità della corteccia visiva, è la dimostrazione istochimica della succinicodeidrogenasi. Questa tecnica mostra un'altra disposizione verticale che ha la forma di strette colonne verticali ascendenti attraverso la lamina III e un analogo sistema di colonne discendenti verticalmente nello strato infragranulare; sorprendentemente, le colonne discendenti non sono allineate con le colonne ascendenti, ma si trovano spostate dilato di metà della larghezza delle strisce della visione monoculare. Le colonne della succinicodeidrogenasi (o blobs, per usare un termine usato colloquialmente dagli scienziati) mostrano una corrispondenza sistematica con le strisce della visione monoculare.
Vi sono due possibili spiegazioni, radicalmente diverse fra loro, delle colonne di succinicodeidrogenasi. Secondo Hubel queste colonne sarebbero i siti della corteccia visiva in cui hanno preferenzialmente termine (nei Primati) le afferenze specificamente sensibili al colore, e sarebbe possibile osservare una sistematica connessione tra queste colonne e un sistema di brevi strisce localizzato nell'area 18. Secondo la spiegazione più tradizionale, sostenuta da Horton (v., 1984), le colonne di succinicodeidrogenasi potrebbero rappresentare una concentrazione di strutture, probabilmente di sinapsi inibitorie, che servirebbe a spiegare il sistema della sensibilità all'orientamento. Quest'ultima spiegazione concorda con alcune precedenti considerazioni teoriche di Braitenberg e Braitenberg (v., 1979) e anche con l'ipotesi abbastanza pragmatica avanzata dall'autore di questo articolo (v. Szentàgothai, 1983). Per il momento non è possibile dare una descrizione del tutto convincente e coerente dell'architettura funzionale della corteccia.
Nel frattempo anche altri studi su regioni diverse della neocorteccia sembrano indicare l'esistenza di un'organizzazione verticale simile a quella riscontrata nelle aree sensoriali specifiche della corteccia.
Benché fossero già disponibili alcuni dati, peraltro abbastanza generici, relativi all'organizzazione verticale delle connessioni cortico-corticali, le osservazioni di Goldman e Nauta (v., 1977), basate sull'uso di metodi autoradiografici per individuare le connessioni neuronali da varie aree del lobo frontale (aree 4, 9, 12), hanno definitivamente chiarito le basi anatomiche del principio dell'organizzazione colonnare.
A differenza delle ‛colonne' della corteccia sensoriale primaria (per esempio le colonne della dominanza oculare), in cui le terminazioni dirette delle afferenze talamiche sono in gran parte limitate alla lamina IV, mentre gli strati corticali più superficiali e più profondi ricevono principalmente impulsi trasmessi da neuroni locali (le cosiddette cellule stellate spinose), le colonne cortico-corticali si estendono attraverso praticamente tutto lo spessore della corteccia. Viste dalla superficie, queste colonne hanno in genere l'aspetto di strisce parallele, molto simili alle strisce zebrate presenti nella corteccia visiva nei territori della dominanza oculare; tuttavia, i margini dei territori che contengono le terminazioni di fibre provenienti da aree diverse della corteccia sono senz'altro verticali, e i territori di proiezione possono alternarsi con regolarità alle proiezioni di diversa origine corticale. Uno splendido esempio di questa situazione è illustrato nella fig. 25 (v. Goldman-Rakic e Schwartz, 1982), dove le proiezioni del corpo calloso provenienti dall'emisfero controlaterale si alternano con regolarità ai territori di proiezione del bordo posteriore del solco interparietale ipsilaterale. Il grande ordine presente nelle terminazioni assoniche della corteccia non è eguagliato da quello riscontrato nei neuroni corticali che danno origine a queste connessioni. Tuttavia, l'attenta analisi di Schwartz e Goldman-Rakic (v., 1984) effettuata con tecniche di marcatura retrograda ha mostrato che lo stesso ordine, anche se con un minor grado di raffinatezza, è generalmente presente nelle connessioni delle afferenze e delle efferenze.
Se si prendono in considerazione singole afferenze cortico-corticali, invece di intere popolazioni di afferenze messe in evidenza dalle tecniche di tracciamento, l'arborizzazione terminale non corrisponde a delle strisce ma piuttosto a vere e proprie colonne cilindriche.
Seguendo i singoli assoni cortico-corticali colorati col metodo di Golgi, come pure popolazioni assoniche più vaste, è possibile osservare che le terminazioni raggiungono in gran parte la lamina I oltre il confine dei margini verticali dei loro territori di proiezione.
Un aspetto importante, che è stato messo in luce dopo le fondamentali osservazioni di Goldman e Nauta, è la relativa costanza in un ampio ambito filogenetico (dal ratto alla scimmia Rhesus) delle dimensioni delle colonne cortico-corticali (v. Szentàgothai, 1978; v. Bugbee e Goldman-Rakic, 1983; v. Goldman-Rakic, 1984); pertanto l'espansione della superficie corticale che si riscontra nella filogenesi si accompagna a un aumento del numero piuttosto che della larghezza delle colonne di fibre afferenti. Questa relativa costanza delle dimensioni delle colonne si riscontra anche, mutatis mutandis, nel numero di cellule e nelle proporzioni relative esistenti tra i tipi di cellule. Già da lungo tempo era noto che la densità cellulare della corteccia diminuisce via via che si sale la scala filogenetica, mentre i mutamenti dei tipi di cellule corticali e quelli dello schema di arborizzazioni dendritiche e assoniche sono qualitativamente trascurabili. La relativa costanza che si riscontra nella filogenesi dei mammiferi superiori per quel che riguarda il numero di cellule presenti in aree della corteccia di uguale superficie (v. Rockel e altri, 1974) è controbilanciata dal crescente spessore della corteccia, che potrebbe spiegare la diminuzione della densità cellulare. Una conseguenza naturale del maggior numero di colonne (moduli) e di cellule che si riscontra nella scala filogenetica deve essere, fra le altre cose, un aumento delle connessioni intercorticali. Ciò va ad aggiungersi alla frazione di volume tissutale ‛non cellulare' della corteccia, ma probabilmente non è necessario che vi sia una proporzionalità lineare tra il numero delle cellule corticali e quelle delle connessioni cortico-corticali, cosicché in mancanza di appropriati studi specifici è difficile dare una valutazione quantitativa di questo aumento delle connessioni.
Poiché le afferenze cortico-corticali sono senz'altro il principale denominatore comune dell'architettura colonnare della neocorteccia, è logico immaginare le colonne elementari disposte attorno a un'afferenza cortico-corticale centrale. L'autore di questo articolo ha provato a costruire questo ideale modello fin dal 1978. Anche se estremamente semplificati, questi moduli riescono a dare un'impressione abbastanza esatta (almeno per il momento) delle connessioni neuroniche intere intracorticali. La fig. 26 dà una dimostrazione del principio la parte superiore del diagramma mostra le connessioni cortico-corticali disposte a mo' di mosaico. Poiché praticamente tutte le cellule piramidali sopragranulari danno origine ad afferenze cortico-corticali - in gran parte ipsilaterali, ma alcune anche al corpo calloso -, l'intera efferenza distale dei tre strati esterni è diretta ad altre regioni della corteccia. Molte cellule piramidali (o fusiformi) infragranulari presenti nelle lamine V e VI, nonostante si dirigano principalmente verso bersagli subcorticali, mostrano assoni principali o, più spesso, collaterali, diretti all'emisfero opposto, cosicché sembra giusto supporre che più dell'80% delle efferenze corticali rientri direttamente in altre regioni corticali. Esistono connessioni intracorticali adiacenti che convergono su 3-5 colonne elementari, in qualunque direzione tangenziale, cioè collegamenti a ponte fino a 3 mm in questa direzione. Molte di queste connessioni, naturalmente, vengono stabilite dagli assoni orizzontali delle cellule a canestro grandi, ma si trova anche un gran numero di connessioni eccitatorie. La parte inferiore del diagramma della fig. 26 mostra l'ingrandimento di una singola colonna disposta intorno a un unico assone corticale, preso come esempio, situato nell'asse dei cilindri di base. La figura cerca di dare l'impressione dei presunti cambiamenti dinamici della colonna, originariamente cilindrica, che si determinano per la maggiore espansione delle branche terminali delle afferenze cortico-corticali nella lamina I e delle collaterali assoniche di alcune cellule piramidali specifiche della lamina V nello stesso strato. L'eccitazione si propaga anche nella parte più profonda della corteccia tramite collaterali di vari tipi di cellule piramidali. Al contrario, le connessioni tangenziali inibitorie sono prevalenti negli strati corticali intermedi (lamine III, IV, V) che avrebbero la tendenza a restringere o ‛cingere la vita' della colonna. Questo disegno, pertanto, cerca di dare un'impressione generale della distribuzione dinamica di questi cilindri, che da verticali divengono a forma di clessidra (iperboloide di rotazione), e quindi tenta di dare anche un'idea di come potrebbe verificarsi il ‛confinamento' spaziale dell'attività in singole colonne. Questa ipotesi è stata elaborata dall'autore di questo articolo indipendentemente da Edelman e Finkel (v., 1984) e partendo da premesse del tutto diverse.
La disposizione modulare (colonnare) dovrebbe essere considerata nient'altro che un principio ‛puramente architettonico' che si avvale dell'uso ripetitivo di unità tissutali di struttura simile. Questo principio architettonico è diffuso anche in altri centri nervosi di vertebrati superiori (v. Szentàgothai, 1981 e 1983) e l'organizzazione colonnare della corteccia prefrontale si riscontra anche in organizzazioni simili del neostriato (v. Selemon e Goldman-Rakic, 1985). Il principio architettonico modulare non è e non dovrebbe essere considerato un principio funzionale. Da un punto di vista architettonico, e anche da quello dello sviluppo, un modulo cilindrico può essere un'unità, ma circa 5.000 cellule corticali possono essere connesse fra loro con un numero infinito di combinazioni, talché solo due decine o centinaia di neuroni sono attive in un qualunque intervallo di tempo nella stessa colonna e una grande varietà di catene neuronali può essere attivata o inibita simultaneamente.
Benché la fig. 26 indichi l'esistenza di una tendenza generale nella diffusione dell'eccitazione o dell'inibizione in una singola colonna, questo non significa che le connessioni funzionali di una catena neuronale siano sempre le stesse. Al contrario, le circostanze funzionali (afferenze) e gli ‛antecedenti' possono determinare di volta in volta l'attivazione in qualunque colonna di una infinita varietà di catene neuronali.
Il principio modulare e quello che abbiamo cercato di spiegare servendoci della fig. 26, come pure le ipotesi sulle connessioni cortico-corticali suggerite nella parte superiore del diagramma, danno una vivida rappresentazione della organizzazione delle funzioni corticali nel continuo e infinito andare e venire dell'attività corticale da un sito verso altri siti, vicini o lontani, della corteccia. Inoltre, vi sono circuiti esterni alla corteccia - tramite i quali qualunque tipo di attività corticale può venir diretta indietro verso altre parti della corteccia utilizzando un grandissimo numero di catene neuronali attraverso i nuclei superiori del tronco dell'encefalo, il talamo, i nuclei inferiori del ponte e del bulbo, il cervelletto, ecc. Tuttavia, la discussione di questi problemi andrebbe ben oltre lo scopo di questo articolo.
g) Sistemi di fibre (e cellule) diffuse
I sistemi di fibre diffuse che risalgono alla corteccia cerebrale, la cui funzione può essere sia attivante sia disattivante, sono stati descritti per la prima volta nei classici lavori di Moruzzi (v. Moruzzi e Magoun, 1949). Tuttavia, solo con la messa a punto di tecniche per lo studio delle fibre monoamminergiche è stato possibile definire con esattezza la vera natura della maggioranza di queste vie e il loro sito di origine. Anche se questi sistemi di fibre erano in gran parte davvero monoamminergici - noradrenergici (adrenergici), dopamminergici, serotoninergici - l'introduzione di nuove tecniche immunocitochimiche ha permesso di mettere in luce analoghi sistemi diffusi di natura colinergica. Un'ulteriore complicazione è stata causata dal fatto che cellule specifiche, descritte come cellule corticali fusiformi caratteristicamente orientate in senso verticale e contenenti il polipeptide intestinale vasoattivo (VIP), sono risultate anch'esse di natura colinergica, cosicché il sistema colinergico della corteccia non ha soltanto natura estrinseca (ascendente), ma anche (e per una porzione significativa) origine locale.
Il sistema noradrenergico ha origine nel locus coeruleus, un nucleo compatto situato nella parte dorsale del ponte. Le fibre risalgono attraverso varie vie abbastanza diffuse, di preferenza attraverso le parti limbiche e frontali della corteccia, da dove si espandono in tutta la corteccia. Le fibre noradrenergiche della corteccia hanno aspetto varicoso, in genere a carattere terminale, e sono presenti in tutti gli strati. L'innervazione più fitta si trova nelle lamine IV e V, anche se nella lamina IV della corteccia visiva primaria dei Primati può essere abbastanza sparsa. Nella lamina I le fibre noradrenergiche sono piuttosto poche, hanno orientamento tangenziale e divengono più dense al confine tra le lamine I e II. Nell'insieme, il plesso terminale corticale ha andamento continuo in tutta la corteccia e probabilmente la maggioranza delle fibre entra nella corteccia dalle regioni frontali, da dove si diffonde in prevalenza intracorticalmente verso la parte posteriore dell'emisfero.
Le nostre conoscenze riguardo ai contatti sinaptici stabiliti con elementi cellulari corticali sono piuttosto scarse. È generalmente accettato che la prevalenza delle terminazioni sia non sinaptica, per cui gran parte del mediatore può venir rilasciato nello spazio intercellulare. Tuttavia questa convinzione non è sostenuta dai più recenti studi immunocitochimici. È ancora in discussione quale azione eserciti la noradrenalina sulle cellule corticali: potrebbe aumentare la trasmissione attraverso certe sinapsi e deprimerla in altre. Alcuni dati indicano che la plasticità delle funzioni corticali, specialmente nei primi periodi di vita, viene assicurata dalle terminazioni noradrenergiche, e anche la formazione di sinapsi potrebbe essere influenzata dalla noradrenalina, che quindi sarebbe più un ‛modulatore' sinaptico che un vero e proprio ‛mediatore'.
Il sistema serotoninergico ha origine nelle cellule del rafe mediale e dorsale della parte inferiore del tronco dell'encefalo. Le fibre serotoninergiche risalgono attraverso il fascicolo longitudinale mediale e terminano in tutti gli strati della corteccia con una lieve prevalenza per la lamina I, almeno nei mammiferi inferiori, mentre nei Primati lo strato più ricco di terminazioni serotoninergiche è la lamina IV. I contatti sinaptici stabiliti dagli assoni serotoninergici con cellule corticali non sono ancora del tutto chiari, e anche la questione della liberazione sinaptica o non sinaptica del mediatore nel sistema delle afferenze serotoninergiche è tuttora aperta, come nel caso delle terminazioni noradrenergiche. Anche la funzione di questo sistema non è ancora chiara, pur essendo stato osservato che la serotonina diminuisce l'attività spontanea delle cellule corticali.
Il sistema dopamminergico ha origine nell'area segmentale ventrale e nella porzione mediale della sostanza nera del mesencefalo. A differenza degli altri sistemi di fibre diffuse, le cui terminazioni sono largamente sparse in tutta la corteccia, le fibre dopamminergiche terminano quasi esclusivamente nelle aree frontali e limbiche della corteccia. L'effetto funzionale esercitato dalle fibre dopamminergiche è complessivamente inibitorio, ma non è possibile dare in questo articolo una descrizione più completa di questo sistema.
Un ulteriore sistema diffuso presente nella corteccia è quello delle fibre e delle cellule colinergiche, la cui esistenza era nota da molto tempo e che è stato oggetto di studi fin dall'inizio degli anni sessanta. Per poter definire assoni e cellule con esattezza, è stato prima necessario mettere a punto delle metodologie immunocitochimiche affidabili, in particolare quelle basate sulla presenza di colin-acetil-transferasi. Attualmente possiamo distinguere con certezza due sistemi colinergici: quello estrinseco, costituito da fibre e terminazioni ascendenti, e un sistema intrinseco di cellule che è probabilmente lo stesso delle cellule corticali contenenti il polipeptide intestinale vasoattivo.
Il sistema estrinseco ha origine nei neuroni relativamente grandi della parte basale del proencefalo, particolarmente nel nucleo basale (sostanza innominata) e in alcune strutture correlate. Anche alcune regioni segmentali del mesencefalo probabilmente danno origine ad afferenze colinergiche alla corteccia cerebrale. Le fibre colinergiche estrinseche hanno terminazioni diffuse in tutti gli strati corticali, con una lieve prevalenza per la lamina V (nel ratto), mentre sono meno abbondanti nella lamina IV della corteccia somatosensitiva. Questo è l'unico sistema, almeno per ora, nel quale è stato dimostrato che le terminazioni delle afferenze sono in prevalenza sinaptiche, anche se non sono stati identificati con chiarezza gli elementi postsinaptici (generalmente tronchi dendritici). Le sinapsi corrispondono a quelle di secondo tipo, dato che hanno contatti di membrana simmetrici. Poiché questo tipo di sinapsi è in genere associato all'azione inibitoria, questa osservazione contrasta col fatto che l'azione dell'acetilcolina è concordemente ritenuta eccitatoria. Tuttavia, se gli elementi di contatto sono inibitori, un'inibizione esercitata su interneuroni inibitori potrebbe ben rendere conto degli effetti eccitatori descritti.
Il sistema intrinseco corrisponde alle cellule fusiformi locali che sono caratterizzate dall'orientamento verticale dei due dendriti principali e dalla pronunciata reattività nei confronti degli anticorpi anti-VIP. Il sistema assonico di queste cellule presenta un'arborizzazione intricata - generalmente non lontana dai corpi cellulari - di fibre terminali ‛a corona di rosario'.
Purtroppo non è stato finora possibile separare le terminazioni estrinseche da quelle del sistema intrinseco.
bibliografia
Albus, J.S., A theory of cerebellar function, in ‟Mathematical biosciences", 1971, X, pp. 25-61.
Altman, J., Morphological development of the rat cerebellum and some of its mechanism, in The cerebellum. New vistas (a cura di S. Palay e V. Chan-Palay), Berlin-New York 1982, pp. 8-49.
Bailey, P., Bonin, G. von, The isocortex of man, Urbana 1951.
Baillarger, J.-G.-F., Recherches sur la structure de la couche corticale des circonvolutions du cerveau, in ‟Mémoires de l'Académie Royale de Médecine de Paris", 1840, VII, pp. 149-183.
Berlin, R., Beiträge zur Struktur der Grosshirnwindungen, Erlangen 1858.
Betz, W., Anatomischer Nachweis zweier Gehirnzentra, in ‟Zentralblatt für medizinische Wissenschaften", 1874, XII, pp. 578-580, 595.
Bolk, L., Das Cerebellum der Säugetiere, Haarlem-Jena 1906.
Braak, H., Über das Neurolipofuscin in der unteren Olive und dem Nucleus dentatus cerebelli des Menschen, in ‟Zeitschrift für Zellforschung und mikroskopische Anatomie", 1971, CXXI, pp. 573-592.
Braak, H., Architectonics as seen by lipofuscin stains, in Cerebral cortex, vol. I (a cura di A. Petres ed E.G. Jones), New York-London 1984.
Bradley, O.C., The mammalian cerebellum: its lobes and fissures, in ‟Journal of anatomy and physiology", 1905, XXXIX, pp. 99-117.
Braitenberg, V., Atwood, R. P., Morphological observations on the cerebellar cortex, in ‟Journal of comparative neurology", 1958, CIX, pp. 1-27.
Braitenberg, V., Braitenberg, C., Geometry of orientation columns in the visual cortex, in ‟Biological cybernetics", 1979, XXXIII, pp. 179-186.
Braitenberg, V., Onesto, N., The cerebellar cortex as a timing-organ. Discussion of an hypothesis, in Proceedings of the 1st Annual Congress on medical cybernetics, Neaples 1961, pp. 1-19.
Brodmann, K., Beiträge zur histologischen Lokalisation der Grosshirnrinde: die Cortexgliederung des Menschen, in ‟Journal für Psychologie und Neurologie", 1908, VI, pp. 231 ss.
Brodmann, K., Neue Ergebnisse über die vergleichende histologische Lokalisation der Grosshirnrinde mit besonderer Berücksichtigung des Stirnhirns, in ‟Anatomischer Anzeiger", suppl., 1912, XLI, pp. 157-216.
Broodmann, K., Physiologie des Gehirns, in Neue deutsche Chirurgie, vol. XI (a cura di P. von Brun), Stuttgart 1914, pp. 85-462.
Bugbee, N. M., Goldman-Rakic, P.S., Columnar organization of cortico-cortical projection in Squirrel and Rhesus monkeys: similarity of column width in species differing in cortical volume, in ‟Journal of comparative neurology", 1983, CCXX, pp. 355-365.
Chan-Palay, V., The tripartite structure of the undercoat in initial segments of Purkinje cell axons, in ‟Zeitschrift für natomie und Entwicklungsgeschichte", 1972, CXXXIV, pp. 200-234.
Chan-Palay, V., Cerebellar dentate nucleus. Organization, cytology and trasmitters, Berlin-New York 1977.
Desclin, J. C., Histological evidence supporting the inferior olive as the major source of cerebellar climbing fibers in rat, in ‟Brain research", 1974, LXXVII, pp. 365-384.
Eccles, J.C., Ito, M., Szentágothai, J., The cerebellum as a neuronal machine, Berlin-New York 1967.
Economo, C. von, Koskinas, G. N., Die Cytoarchitektonik der Hirnrinde des erwachsenen Menschen, Wien 1925.
Edelman, G. M., Group selection and phasic reentrant signaling: a theory of higher brain function, in The mindful brain (a cura di G. M. Edelman e V. B. Mountcastle), Cambridge, Mass., 1978, pp. 51-100.
Edelman, G. M., Finkel, L. H., Neuronal group selection in the cerebral cortex, in Dynamica spects of neocortical function (a cura di G. M. Edelman, W. E. Gall e W. M. Cowan), New York 1984.
Freund, F.G., Martin, K. A. C., Smith, A. D., Somogyi, P., Glutamate decarboxylase-immunoreactive terminals of Golgi-impregnated axoaxonic cells and of presumed basket cells in synaptic contact with pyramidal neurons of the cat's visual cortex, in ‟Journal of comparative neurology", 1983, CCXXI, pp. 263-278.
Freund, T.F., Martin, K.A.C., Somogyi, P., Whitteridge, D., Innervation of cat visual areas 17 and 18 by physiologically identified X- and Y-type thalamic afferents. II. Identification of post-synaptic targets by GABA immunocytochemistry and Golgi impregnation, in ‟Journal of comparative neurology", 1985, CCXLII, pp.275-291.
Freund, T.F., Martin, K.A.C., Whitteridge, D., Innervation of cat visual areas 17 and 18 by physiologically identified X- and Y-type thalamic afferents. I. Arborization patterns and quantitative distribution of postsynaptic elements, in ‟Journal of comparative neurology", 1985, CCXLII, pp. 263-274.
Fujita, M., Adaptive filter model of the cerebellum, in ‟Biological cybernetics", 1982, XLV, pp. 195-206.
Gennari, F., De peculiari structura cerebri nonnulisque eius morbis, Parma 1782.
Ghez, U.M., Fahn, S., The cerebellum, in Principles of neural science (a cura di E. R. Kandel e J. H. Schwartz), Amsterdam 1981.
Goldman, P.S., Nauta, W. J. H., Columnar distribution of cortico-cortical fibers in the frontal association, limbic and motor cortex of the developing Rhesus monkeys, in ‟Brain research", 1977, CXXII, pp. 393-413.
Goldman-Rakic, P.S., Modular organization of prefrontal cortex, in ‟Trends in neuroscience", 1984, VII, pp. 419-429.
Goldman-Rakic, P.S., Schwartz, M. L., Interdigitation of controlateral and ipsilateral columnar projections to frontal association cortex in Primates, in ‟Science", 1982, CCXVI, pp. 755-757.
Golgi, C., Sulla struttura della sostanza grigia del cervello, in ‟Gazzetta medica lombarda", 1873, XXXIII, pp. 244-246.
Golgi, C., Recherches sur l'histologie des centres nerveux, in ‟Archives italiennes de biologie", 1883, III, pp. 285-317.
Gray, E.G., Ultra-structure of synapses of the cerebral cortex and certain specializations of neuroglial membranes, in Electron microscopy in anatomy (a cura di J.D. Boyd, F.R. Johnson e J.D. Lever), London 1961, pp. 54-73.
Hámori, J., Mezey, E., Szentágothai, J., Electron microscopic identification of cerebellar nucleocortical mossy terminals in the rat, in ‟Experimental brain research", 1981, XLIV, pp. 97-100.
Horton, J. C., Cytochrome oxidase patches: a new cytoarchitectonic feature of monkey visual cortex, in ‟Philosophical transactions of the Royal Society", series B, 1984, CCCIV, pp. 199-253.
Hubel, D.H., Wiesel, T.N., Receptive fields of single neurones in the cat's striate cortex, in ‟Journal of physiology", 1959, CXLVIII, pp. 574-591.
Hubel, D.H., Wiesel, T.N., Laminar and columnar distribution of geniculo-cortical fibers in the macaque monkey, in ‟Journal of comparative neurology", 1972, CXLVI, pp. 421-450.
Ito, M., The cerebellum and neural control, New York 1984.
Jansen, J., On cerebellar evolution and organization from the point of view of a morphologist, in Neurobiology of cerebellar evolution and development (a cura di R. Llinás), Chicago 1969, pp. 881-893.
Jansen, J., Brodal, A., Experimental studies on the intrinsic fibers of the cerebellum. II. The cortico-nuclear projection, in ‟Journal of comparative neurology", 1940, LXXIII, pp. 267-321.
Kisvárday, Z.F., Martin, K.A.C., Freund, T.F., Maglóczyky, Z., Whitteridge, D., Somogy, P., Sunaptic targets of HRP-filled layer III. Pyramidal cells in the cat striate cortex, in ‟Experimental brain research", 1986, LXIV, pp. 541-552.
Kisvárday, Z.F., Martin, K.A.C., Friedlander, M.J., Somogyi, P., Evidence of interlaminar inhibitory circuits in the striate cortex of the cat, in ‟Journal of comparative neurology", 1987, CCLX, pp. 1-19.
Kisvárday, Z.F., Martin, K.A.C., Whitteridge, D., Somogyi, P., Synaptic connections of intracellularly filled clutch cells: a type of small basket cell in the visual cortex of the cat, in ‟Journal of comparative neurology", 1985, CCXLI, pp. 111-137.
Larramendi, L.M.H., Analysis of synaptogenesis in the cerebellum of the mouse, in Neurobiology of cerebellar evolution and development (a cura di R. Llinás), Chicago 1969, pp. 803-843.
Larramendi, L. M. H., Victor, T., Synapses on the Purkinje cell spines in the mouse. An electron microscopic study, in ‟Brain research", 1967, V, pp. 15-30.
Larsell, O., The comparative anatomy and histology of the cerebellum from Monotremes through Apes, vol. II (a cura di J. Jansen), Minneapolis 1970.
Larsell, P., The development and subdivisions of the cerebellum of birds, in ‟Journal of comparative neurology", 1948, LXXXIX, pp. 123-183.
Lorente de Nó, R., The cerebral cortex: architecture, intracortical connections and motor projections, in Physiology of the nervous system (a cura di J. F. Fulton), London-New York 1938, pp. 291-321.
Lugaro, E., Über die Verbindung des nervösen Elementes der Kleinhirnrinde untereinander mit allgemeinen Betrachtungen über die physiologische Bedeutung der Rapporte zwischen den nervösen Elementen, in ‟Untersuchungen zur Naturlehre des Menschen und der Tiere", 1895, XV, pp. 475-514.
Marr, D., A theory of cerebellar cortex, in ‟Journal of physiology", 1969, CCII, pp. 437-470.
Meynert, T., Der Bau der Grosshirnrinde und seine örtlichen Verschiedenheiten, nebst einem pathologisch-anatomischen Corollarium, in ‟Vierteljahrsschrift für Psychiatrie", 1867-1868, I, pp. 77-93, 125-217, 381-403; II, pp. 88-113.
Moruzzi, G., Magoun, H.W., Brain stem reticular formation and activation of the EEG, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 455-473.
Mountcastle, V.B., Modalities and topographic properties of single neurons of cat's sensory cortex, in ‟Journal of neurophysiology", 1957, XX, pp. 408-434.
Mountcastle, V.B., An organizing principle for cerebral function: the unit module and the distributed system, in The mindful brain (a cura di G. M. Edelman e V.B. Mountcastle), Cambridge, Mass., 1978, pp. 7-50.
Oscarsson, O., Spatial distribution of climbing and mossy fiber input into the cerebellar cortex, in Experimental brain research, suppl. 1, Afferent and intrinsic orgnization of laminated structure in the brain (a cura di O. Creutzfeldt), Berlin 1976, pp. 34-42.
Palay, S.L., Chan-Palay, V., Cerebellar cortex. Cytology and organization, Berlin-New York 1974.
Palkovits, M., Magyar, P., Szentágothai, J., Quantitative histological analysis of the cerebellar cortex in the cat. II. Cell numbers and densities in the granular layer, in ‟Brain research", 1971, XXXII, pp. 15-30.
Palkovits, M., Mezey, E., Hámori, J., Szentágothai, J., Quantitative histological analysis of the cerebellar nuclei in the cat. Numerical data on cells and synapses, in ‟Experimental brain research", 1977, XXVIII, pp. 189-209.
Pellionisz, A., Tensorial aspects of the multidimensional approach to the vestibulo-oculomotor reflex and gaze, in Adaptive mechanisms in gaze control (a cura di A. Berthoz e G. Mellvill-Jones), Amsterdam 1985, pp. 231-396.
Pellionisz, A., Llinás, R., Tensorial approach to the geometry of brain function: cerebellar coordination via metric tensor, in ‟Neuroscience", 1980, V, pp. 1125-1136.
Purkinje, J. E., Über die gangliöse Natur bestimmter Hirnteile. Bericht über die Versammlung deutscher Natuforscher und Ärtze in Prag im September, 1837, Bd. 3, Sekt. 5, in Anatomisch-physiologische Verhandlungen, Prag 1837, pp. 177-180.
Rakic, P., Neuron-glial relationship during granule cell migration in developing cerebellar cortex: a Golgi and electron microscopic study in Macacus rhesus, in ‟Journal of comparative neurology", 1971, CXLI, pp. 283-312.
Rakic, P., Mode of cell migration to the superficial layers of fetal monkey neocortex, in ‟Journal of comparative neurology", 1972, CXLI, pp. 283-312.
Rakic, P., Mode of cell migration to the superficial layers of fetal monkey neocortex, in ‟Journal of comparative neurology", 1972, CXLV, pp. 61-84.
Rakic, P., Neuron-glial interaction during brain development, in ‟Trends in neuroscience", 1981, IV, pp. 184-187.
Rakic, P., Early developmental events: cell lineages, acquisition of neuronal positions, and areal laminar development, in ‟Neuroscience research program. Bulletin", 1982, XX, pp. 439-451.
Rakic, P., Goldman-Rakic, P.S., Development and modifibiality of the cerebral cortex, Cambridge, Mass., 1982.
Ramón y Cajal, S., Estructura de los centros nerviosos de las aves, in ‟Revista trimestral de histología normal y patologíca", Barcelona 1888.
Ramón y Cajal, S., Estudio sobra la corteza cerebral humana, in ‟Revista trimestral de microscopía", 1889, IV, pp. 1-63.
Ramón y Cajal, S., Histologie du système nerveux de l'homme et des vertebrés, 2 voll., Paris 1911.
Rockel, A.J., Hiorns, R.W., Powell, T. P. S., Numbers of neurons through full depth of neocortex, in ‟Journal of anatomy", 1974, CXVIII, pp. 371 ss.
Schwartz, M. L., Goldman-Rakic, P.S., Callosal and intrahemispheric connectivity of the prefrontal association cortex in Rhesus monkey: relation between intraparietal and principal sulcal cortex, in ‟Journal of comparative neurology", 1984, CCXXVI, pp. 403-420.
Selemon, L.D., Goldman-Rakic, P.S., Longitudinal topography and interdigitation of corticostriatal projections in the Rhesus monkey, in ‟Journal of neurosciences", 1985, V, pp. 776-794.
Sidman, R.L., Cell proliferation, migration, and interaction in the developing mammalian central nervous system, in The neurosciences. Second study program, (a cura di F. O. Schmitt), New York 1970, pp. 100-107.
Sömmerrings, S. T., Vom Hirn und Rückenmark, Mainz 1788.
Somogyi, P., A specific axo-axonal interneuron in the visual cortex of the rat, in ‟Brain research", 1977, CXXXVI, pp. 345-350.
Somogyi, P., Cowey. A., Combined Golgi and electron microscopic study on the synapses formed by double bouquet cells in the visual cortex of the cat and monkey, in ‟Journal of comparative neurology", 1981, CLXXXV, pp. 547-566.
Somogyi, P., Kisvárday, Z.F., Martin, K. A. C., Whitteridge, D., Synaptic connections of morphologically identified and physiologically characterized large basket cells in the striate cortex of cat, in ‟Neuroscience", 1983, X, pp. 261-294.
Szentágothai, J., New data on the functional anatomy of synapes, in ‟Magyar tudományos akadémia biológiai tudományok osztálnyának közleményei", 1963, VI, pp. 217-227.
Szentágothai, J., The use of degeneration methods in the investigation of short neuronal connections, in Progress in brain research, vol. XIV, Degeneration patterns in the nervous system (a cura di M. Singer e J. P. Schadé), Amsterdam 1965, pp. 1-32.
Szentágothai, J., Architecture of the cerebral cortex, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper, A. A. Ward e A. Pope), Boston 1969, pp. 13-28.
Szentágothai, J., Glomerulal synapses, complex synaptic arrangements, and their operational significance, in The neurosciences. Second study program, (a cura di F. O. Schmitt), New York 1970, pp. 427-443.
Szentágothai, J., The module concept' in cerebral cortex architecture, in ‟Brain research", 1975, XCV, pp. 475-496.
Szentágothai, J., The neuron network of the cerebral cortex: a functional interpretation. The Ferrier Lecture 1977, in ‟Proceedings of the Royal Society", series B, 1978, CCI, pp. 219-248.
Szentágothai, J., Local neuroncircuits of the neocortex, in The neurosciences. Fourth study program (a cura di F. O. Schmitt e F. G. Worden), Cambridge, Mass., 1979, pp. 399-415.
Szentágothai, J., Principles of neural organization, in Advances in physiological science, vol. I, Regulatory functions of the CNS principles of motion and organization (a cura di J. Szentágothai, M. Palovits e J. Hámori), Oxford-Budapest 1981, pp. 1-16.
Szentágothai, J., The modular architectonic principle of the neural centers, in ‟Review of physiological and biochemical pharmacology", 1983, IIC, pp. 1-61.
Szentágothai, J., Arbib, M., Conceptual models of neural organization, Cambridge, Mass., 1974, reprint 1975.
Szentágothai, J., Rajokovits, K., Über den Ursprung der Kletterfasern des Kleinhirns, in ‟Zeitschrift für Anatomie und Entwicklungsgeschichte", 1959, CXXI, pp. 130-141.
Vicq d'Azyr, F., Traité d'anatomie et de physiologie, Paris 1786.
Vogt, O., Vogt, C., Allgemeine Ergebnisse unserer Hirnforschung, in ‟Journal für Psychologie und Neurologie", 1919, XXV, pp. 279-461.
Voogd, J., The cerebellum of the cat. Structure and fiber connexions, Philadelphia 1964.
Zagon, I.S., McLaughlin, P. J., Smith, S., Neural populations in the human cerebellum: estimations from isolated cell nuclei, in ‟Brain research", 1977, CXXVII, pp. 279-282.
Trasporto assonale di Giorgio Macchi, Marina Bentivoglio
sommario: 1. Introduzione e cenni storici. 2. Metodi di studio. a) Trasporto di sostanze marcate con isotopi radioattivi. b) Accumulo di materiali in seguito a lesioni del nervo. c) Osservazione diretta dei movimenti di organelli assonali. 3. Strutture subcellulari implicate nel trasporto. a) Le neurofibrille. b) Il reticolo assoplasmico. 4. Caratteristiche del trasporto assonale e sue componenti. a) Trasporto anterogrado rapido. b) Trasporto anterogrado intermedio. c) Trasporto anterogrado lento. d) Destino del materiale trasportato in via anterograda. e) Trasporto retrogrado. 5. Meccanismi che regolano il trasporto assonale: a) trasporto rapido; b) trasporto lento. □ 6. Caratteristiche del trasporto di alcune sostanze. a) Glicoproteine. b) Lipidi. c) Nucleosidi. d) Trasmettitori sinaptici ed enzimi a essi correlati. e) Costituenti gliali. f) Trasporto retrogrado di proteine speciali e virus. 7. Il trasporto assonale come meccanismo plastico. 8. Il trasporto assonale in patologia. 9. Il trasporto assonale negli studi di neuroanatomia sperimentale. a) Metodi anterogradi. b) Metodi retrogradi. 10. Considerazioni conclusive. □ Bibliografia.
1. Introduzione e cenni storici
Con il termine ‛trasporto assonale' vengono comunemente designati tutti i complessi meccanismi che si svolgono nella cellula nervosa, deputati al trasferimento di sostanze dalle loro sedi di sintesi, nel caso di sostanze endogene, o di assunzione, nel caso di sostanze sia endogene sia esogene, alle loro sedi di utilizzazione.
In tutte le cellule può essere identificata una complessa sequenza di eventi deputata al trasferimento di macromolecole e organelli dai siti di sintesi a quelli di destinazione. Tuttavia tali meccanismi sono particolarmente sviluppati nelle cellule nervose. In queste ultime, infatti, i prolungamenti dendritici e assonali possono essere anche migliaia di volte maggiori, come lunghezza e volume, del corpo cellulare, mentre i siti di sintesi sono per lo più limitati a quest'ultimo. È inoltre da tenere presente che, una volta differenziati, i neuroni non vanno incontro a ulteriori moltiplicazioni e durante la loro vita cellulare, spesso lunga decenni, devono provvedere a una continua ricostituzione di tutte le componenti molecolari. Per quanto sia stato recentemente stabilito che trasferimenti di sostanze hanno luogo anche lungo i prolungamenti dendritici delle cellule nervose (v. Grafstein e Forman, 1980), la maggior parte dei fenomeni di trasporto si verifica in realtà lungo l'assone. Questo ‛traffico intracellulare' o, meglio, ‛intra-assonale' non solo è deputato alla ricostituzione di componenti strutturali utilizzate nel metabolismo cellulare, ma fornisce anche tutti i materiali specificamente richiesti nello svolgimento delle funzioni sinaptiche.
La prima formulazione del concetto di trasporto assonale è legata all'individuazione del corpo cellulare come ‛centro trofico' della cellula nervosa. Molti dati sperimentali avevano da tempo evidenziato che l'integrità dell'assone dipendeva dal soma, poiché i nervi, isolati dai corpi cellulari di origine delle loro fibre, andavano rapidamente incontro a processi degenerativi. Sulla base di tale evidenza, S. Ramón y Cajal (v., 1911 e 1928), osservatore finissimo del tessuto nervoso, ha sottolineato la capacità del corpo cellulare di promuovere gli scambi nutritivi dei processi cellulari con l'ambiente circostante, e ha avanzato l'ipotesi che il ‛neuroplasma' potesse essere utilizzato come via di diffusione di sostanze che, elaborate nel nucleo cellulare, esercitassero un'azione trofica periferica. Ramòn y Cajal aveva inoltre già intuito alcuni aspetti dinamici della fisiologia neuronale, quali la derivazione dell'assone, durante lo sviluppo, dal corpo cellulare, e il sostegno fornito alla rigenerazione assonale da materiali forniti dal corpo cellulare stesso. Queste ultime ipotesi erano fondate sull'osservazione che, in seguito a interruzione dell'assone, il segmento separato dal corpo cellulare andava incontro a degenerazione, mentre la parte prossimale della fibra rigenerava formando un cono di crescita, simile a quello dello sviluppo embrionale, che la guidava nuovamente verso la periferia.
La conferma sperimentale dell'esistenza di un movimento intra-assonale è successivamente derivata dai classici lavori sperimentali culminati nello studio descritto da P. A. Weiss e H. B. Hiscoe (v., 1948). In tali ricerche venivano riportati gli effetti provocati dalla costrizione cronica del nervo periferico di mammiferi e, in particolare, le modificazioni osservate nei siti prossimali alla sede della costrizione. Il segmento prossimale dell'assone andava infatti incontro a fenomeni degenerativi, caratterizzati da un progressivo rigonfiamento, che vennero ascritti al blocco di una ‛colonna assoplasmica' il cui movimento, dal corpo cellulare alle regioni sinaptiche, fosse stato interrotto dalla riduzione del canale di scorrimento. Inoltre, una volta rimossa la costrizione, la fibra nervosa tendeva a riprendere il cammino verso la periferia (a una velocità di 1-2 mm/die negli esperimenti di Weiss e Hiscoe). Tali osservazioni sperimentali hanno così indotto a ritenere che vi fosse un movimento lento e costante di materiali lungo l'assone, diretto verso la periferia e, quindi, gli organi bersaglio. Per definire questo movimento Weiss e Hiscoe introdussero originariamente il termine ‛flusso assoplasmatico', che suggeriva tuttavia un trasferimento ‛passivo' lungo l'assone. Attualmente viene invece adottata la denominazione di ‛trasporto' assonale, a indicare un processo attivo sulla base del quale vengono veicolate le varie sostanze.
L'introduzione di sostanze marcate con isotopi radioattivi e di molti moderni metodi di indagine (v. cap. 2) ha portato a un notevolissimo progresso delle conoscenze sulle dinamiche del trasporto assonale. Sono state così identificate diverse velocità di migrazione ed è stato scoperto che il trasporto è bidirezionale (e avviene quindi anche in senso retrogrado) lungo le fibre nervose. Le indagini sperimentali sui fenomeni di trasporto assonale hanno dato inoltre un notevole impulso agli studi sulla struttura delle componenti subcellulari del neurone, e dell'assone in particolare, e agli studi sui neurotrasmettitori e su tutte le sostanze coinvolte nei meccanismi sinaptici. Infine il trasporto assonale ha fornito approcci sperimentali moderni ed efficaci negli studi relativi all'origine e alla terminazione delle connessioni interneuronali (v. cap. 9) e ha esercitato quindi un'influenza profondamente innovatrice anche in questo campo della neurobiologia.
2. Metodi di studio
Nello studio dei fenomeni implicati nel trasporto assonale, e nell'identificazione delle sostanze trasportate, vengono comunemente utilizzati tre principali gruppi di metodiche (v. Grafstein e Forman, 1980). Il primo gruppo è rappresentato dal trasporto di sostanze traccianti che possano essere identificate all'interno della cellula nervosa: a tale scopo sono stati per lo più utilizzati materiali marcati con isotopi radioattivi; più recentemente è stato inoltre introdotto l'uso di sostanze che possono essere visualizzate mediante metodiche istochimiche, quali l'enzima horseradish peroxidase (HRP; v. cap. 4). Un secondo gruppo di metodi è invece basato sull'accumulo, lungo le fibre nervose, del materiale trasportato: tale accumulo viene generalmente causato mediante costrizione o interruzione della fibra stessa. Altri studi, infine, sono basati sull'osservazione diretta dei movimenti di organelli subcellulari e dell'assoplasma, in neuroni o in singole fibre in vitro. Il maggior numero di ricerche effettuate con tali metodiche è stato diretto all'identificazione delle componenti del trasporto anterogrado. Tuttavia anche il trasporto retrogrado può essere indagato mediante le stesse tecniche. Le dinamiche del trasporto retrogrado, inoltre, sono state anche ampiamente studiate mediante l'uso di sostanze esogene (v. cap. 4).
a) Trasporto di sostanze marcate con isotopi radioattivi
L'uso di traccianti radioattivi costituisce probabilmente il metodo più diffuso per lo studio del trasporto assonale: è stato infatti largamente applicato all'indagine di vari sistemi neuronali, sia centrali che periferici, in vertebrati e invertebrati.
La maggior parte di tali studi è basata sulla somministrazione di precursori, che vengono successivamente incorporati nelle sostanze trasportate. Alcuni amminoacidi triziati, quali 3H-leucina, -prolina e -metionina, vengono utilizzati come precursori negli studi sulle proteine. Il fucosio e la glucosammina vengono utilizzati per lo studio delle glicoproteine e dei glicolipidi, il glicerolo e la colina per i fosfolipidi, l'acetato, l'acido mevalonico e il colesterolo per gli altri lipidi. Nelle indagini sul trasporto assonale di neurotrasmettitori vengono utilizzate le molecole stesse del neurotrasmettitore marcato. Poiché i terminali assonici non sembrano essere sede di processi di sintesi (v. cap. 3), negli studi in via retrograda non vengono generalmente somministrati precursori, bensì le macromolecole stesse.
In seguito all'iniezione del materiale marcato, le osservazioni sperimentali sul trasporto vengono essenzialmente condotte mediante metodiche biochimiche e autoradiografiche, che consentono di determinare la localizzazione subcellulare, le caratteristiche, la composizione e la velocità di migrazione del materiale trasportato. Nelle indagini basate sui traccianti radioattivi la velocità di trasporto viene solitamente desunta dagli spostamenti del materiale marcato lungo l'assone. A tale scopo le fibre nervose vengono suddivise in segmenti e viene determinato il profilo della radioattività in ciascuno di essi. La velocità di migrazione del materiale trasportato più rapidamente è indicata dalla prima onda di radioattività, mentre la velocità delle altre componenti viene determinata sulla base degli spostamenti dei successivi picchi di radioattività.
Negli studi in via anterograda la velocità di migrazione può essere anche determinata sulla base dei tempi di accumulo della sostanza nei terminali assonici. Tale parametro è particolarmente utile nelle misurazioni del trasporto in assoni brevi, nei quali è difficile ottenere profili di radioattività differenziati in vari segmenti.
Poiché l'iniezione intraoculare di sostanze marcate fornisce un approccio sperimentale assai conveniente e consente una marcatura selettiva delle vie ottiche, queste ultime rappresentano il modello sperimentale più largamente adottato (v. Taylor e Weiss, 1965). Un altro modello molto diffuso è inoltre rappresentato dall'iniezione di isotopi nella mucosa olfattiva, allo scopo di studiarne il trasporto lungo i nervi olfattivi. Questo modello è particolarmente utilizzato nei Pesci, nei quali i nervi olfattivi sono molto lunghi (v. Gross e Kreutzberg, 1978). Molti esperimenti sono stati anche condotti sul nervo sciatico. In quest'ultimo modello sperimentale il precursore marcato puo venire iniettato nei gangli delle radici dorsali per lo studio di assoni sensitivi, oppure nelle corna anteriori del midollo spinale per lo studio del trasporto in assoni motori.
Come tutte le metodiche sperimentali in campo biologico, anche tali tecniche presentano alcuni inconvenienti e problemi di interpretazione dei dati (v. Grafstein e Forman, 1980). Ad esempio, le velocità di migrazione vanno incontro a variazioni considerevoli da un nervo all'altro, soprattutto per quanto riguarda le componenti del trasporto lento, ed è quindi spesso difficile paragonare fra loro i dati sperimentali ottenuti in sistemi diversi. Anche le velocità di incorporazione del precursore e della sua liberazione dal corpo cellulare possono presentare una notevole variabilità. Deve inoltre essere tenuto presente che in alcuni casi il precursore può venire trasportato come tale (e non, quindi, in seguito a incorporazione in macromolecole) e che si possono verificare anche fenomeni di diffusione passiva intra-assonale del precursore.
L'uso di precursori è strettamente collegato al tasso di sintesi proteica e, naturalmente, non è applicabile allo studio di materiali non coinvolti in processi di sintesi, quali, ad esempio, le proteine endogene trasportate dai terminali al corpo cellulare. Per ovviare a tali inconvenienti è stato proposto l'uso di altri traccianti che si legano in modo covalente alle proteine, e quindi, una volta marcati con isotopo radioattivo, possono consentire uno studio aspecifico del trasporto intracellulare di queste ultime (v. Fink e Gainer, 1980).
b) Accumulo di materiali in seguito a lesioni del nervo
Gli studi sull'accumulo di materiali nel nervo sono basati sull'osservazione dei parametri di accumulo di sostanze all'estremità distale e prossimale del nervo, in seguito a interruzione della sua continuità. L'accumulo viene provocato mediante legatura o schiacciamento del nervo, mediante raffreddamento di un suo segmento (v. Brimijoin, 1975), oppure mediante compressione (v. Dahlin e altri, 1984). Tali procedimenti sono ovviamente efficaci quando determinano un blocco dei fenomeni di trasporto. In questo gruppo di metodiche le velocità di migrazione delle varie componenti vengono calcolate sulla base del rapporto fra la quantità di materiale accumulato e la lunghezza dell'assone.
Un esempio classico di tale modello sperimentale è rappresentato dallo studio dell'accumulo di acetilcolinesterasi (v. Lubinska e Niemierko, 1971) alle estremità distale e prossimale di un nervo leso. L'accumulo in entrambi i segmenti indica che il trasporto della sostanza è bidirezionale. Vengono comunemente studiati con la tecnica dell'accumulo anche vari costituenti neuronali, che vengono poi identificati sia mediante tecniche biochimiche e istochimiche sia mediante indagini ultrastrutturali al microscopio elettronico. È stato inoltre recentemente proposto (v. Larsson e altri, 1984) uno studio citofluorimetrico delle sostanze trasportate e dei loro parametri di accumulo, basato sulla possibilità di rendere fluorescenti (ad esempio mediante immunofluorescenza) le varie sostanze in esame. Tale tecnica renderebbe possibile lo studio simultaneo di profili di accumulo di molteplici sostanze in un singolo nervo.
Nella valutazione dei dati ottenuti con tali metodi deve essere comunque tenuto presente che il fenomeno di accumulo può anche essere in parte dovuto a una risposta locale all'evento lesivo. Inoltre la lesione stessa può provocare alterazioni metaboliche: infatti il tasso di accumulo procede in modo lineare per alcune ore, ma poi decresce, probabilmente a causa dell'insorgere di fenomeni degenerativi.
c) Osservazione diretta dei movimenti di organelli assonali
L'osservazione diretta viene effettuata al microscopio mediante particolari accorgimenti tecnici (quali, ad esempio, il contrasto di fase o l'ottica di Nomarski). Sono stati utilizzati a tale scopo neuriti in colture tessutali, o singoli assoni immediatamente dopo dissezione da nervi di vertebrati o di invertebrati. L'osservazione è ovviamente limitata agli organelli di maggiori dimensioni. I movimenti delle varie particelle subcellulari sono stati anche ‛filmati' nel corso dell'osservazione . Analisi quantitative, statistiche e temporali ditali movimenti (v. Forman e altri, 1977), consentite dall'osservazione diretta, possono costituire un utile complemento alle metodiche di studio sopra elencate.
3. Strutture subcellulari implicate nel trasporto
Come è noto, il corpo cellulare del neurone contiene nel citoplasma la cosiddetta sostanza di Nissl, intensamente basofila e quindi facilmente evidenziabile in microscopia ottica, in seguito a opportuna colorazione. I corpi di Nissl sono essenzialmente costituiti dalle strutture deputate ai processi di sintesi, molto attivi nel corpo cellulare, del materiale che dovrà successivamente venire convogliato verso la periferia. È quindi preminente, nel corpo cellulare, il reticolo endoplasmico rugoso o granulare, caratterizzato da gruppi di cisterne orientate parallelamente fra loro e da una grande quantità di ribosomi. Tali elementi costituiscono l'equivalente ultrastrutturale, in microscopia elettronica, delle strutture evidenti come corpi di Nissl in microscopia ottica. L'apparato di Golgi rappresenta, come è noto, un'importante sede per la differenziazione delle membrane neuronali e partecipa alla formazione delle vescicole che vengono poi liberate nei terminali sinaptici, poiché i materiali vanno incontro a un processo di ‛assembiaggio' nell'apparato di Golgi e ne vengono generalmente emessi sotto forma di vescicole. L'apparato di Golgi svolge inoltre un importante ruolo nell'addizione di gruppi prostetici a proteine coniugate.
L'assone contiene molte delle strutture presenti nel corpo neuronale, ma, a differenza di quest'ultimo, è privo di reticolo endoplasmico rugoso e di ribosomi liberi. L'assoplasma è infatti strutturalmente simile al citoplasma di tutte le altre cellule, ma ne differisce per il fatto che, non contenendo ribosomi, non può essere sede di processi di sintesi proteica. A eccezione di una piccola componente proteica fornita dalle cellule di Schwann, tutte le proteine dell'assone derivano essenzialmente dal corpo cellulare e vengono ivi sintetizzate. Le proteine vengono incorporate in componenti strutturali dell'assone vicino al loro sito di sintesi e vengono trasportate lungo l'assone entro tali organelli, che fungono da vettori del trasporto. L'assone è costituito quindi da mitocondri, reticolo endoplasmico liscio o agranulare e corpi multivescicolari; ma, soprattutto, il citoscheletro dell'assone è costituito dalle cosiddette ‛neurofibrille', essenzialmente rappresentate da tre elementi: i microtubuli, i neurofilamenti e i microfilamenti. Le principali caratteristiche delle componenti neurofibrillari e del reticolo assoplasmico verranno qui esposte in dettaglio, in quanto tali elementi costituiscono la base strutturale dei meccanismi di trasporto assonale.
a) Le neurofibrille
La struttura fibrillare delle cellule nervose, conosciuta da tempo, solo negli ultimi decenni è stata oggetto di indagini accurate. Con l'avvento della microscopia elettronica e di adeguate procedure di fissazione, che hanno consentito la scoperta del citoscheletro, si è potuto appurare che le neurofibrille sono costituite da tubuli del diametro di 25 nm (microtubuli) e da filamenti del diametro di 10 nm (neurofilamenti). Tali strutture sono costituite da polimeri allungati, disposti parallelamente all'asse longitudinale dell'assone. I neurofilamenti appaiono per lo più segregati dai microtubuli, cosi che in assoni sezionati trasversalmente si osservano spesso gruppi distinti di microtubuli e neurofilamenti disposti in parallelo fra di loro. Il numero e la distribuzione dei microtubuli e dei neurofilamenti sono soggetti ad ampie variazioni da un assone all'altro. I microtubuli sono spesso associati a organelli legati a membrane, mentre i neurofilamenti sono generalmente ben separati dalle strutture membranose.
I microtubuli sono costituiti da proteine denominate tubuline, divise in due classi principali (α- e β-tubuline), con un peso molecolare di circa 55.000. La loro struttura è identificabile in gruppi di 13 protofilamenti, organizzati in forma cilindrica o tubulare. Questi ultimi sono a loro volta costituiti da copolimeri lineari di unità di α- e β-tubulina, che si alternano lungo il protofilamento.
I neurofilamenti sono invece costituiti da subunità formate da tre proteine principali (le cosiddette ‛triplette della componente lenta': v. cap. 4, §c), il cui peso molecolare varia da 65.000 a 80.000 (P68), da 145.000 a 170.000 (P150) e da 185.000 a 220.000 (P200). Le frazioni proteiche P68 e P150 sono intimamente associate fra loro. La componente P200 sarebbe essenzialmente deputata a promuovere l'associazione di complessi brevi di P68 e P150 per costituire filamenti più lunghi. I filamenti sono substrutture presenti in diversi tipi di cellule, ma le proteine dei neurofilamenti vengono sintetizzate solo nei neuroni: quindi i filamenti di altre cellule hanno costituenti proteici diversi. Le proteine dei neurofilamenti non sono omogenee, in quanto presentano differenze regionali, di specie e anche differenze all'interno di uno stesso nervo. Tuttavia nei Mammiferi ogni proteina di una specie ha una mappa amminoacidica simile a quella delle proteine di specie diverse e vi è quindi una notevole reattività crociata interspecie. I neurofilamenti hanno una lunghezza indefinita. Sono distinguibili dai filamenti gliali grazie alloro maggior diametro (i filamenti gliali hanno un diametro di circa 8 nm) e per la presenza di alcune protuberanze e appendici.
I microfilamenti sono essenzialmente costituiti da actina e formano un cosiddetto ‛lattice' entro l'assone.
b) Il reticolo assoplasmico
Il reticolo assoplasmico (reticolo endoplasmico assonale liscio o agranulare) rappresenta un importante veicolo della componente rapida del trasporto assonale anterogrado. Il reticolo assoplasmico può essere suddiviso in due componenti principali: il reticolo assoplasmico centrale e il reticolo assoplasmico subassolemmale. Il reticolo assoplasmico centrale è costituito da una rete di canali appiattiti e varicosi del diametro di 20-40 nm, interconnessi gli uni con gli altri e delimitati da membrane dello spessore di 60-100 Å, che si estendono longitudinalmente nell'assoplasma; esso rappresenta probabilmente un sistema tubulare continuo che connette il corpo cellulare con i terminali dell'assone. Il reticolo assoplasmico centrale è in connessione con la porzione subassolemmale.
4. Caratteristiche del trasporto assonale e sue componenti
Il trasporto assonale veicola tutti i costituenti assonali e dendritici finora identificati (v. tab. I), quindi coinvolge una grande varietà di materiali.
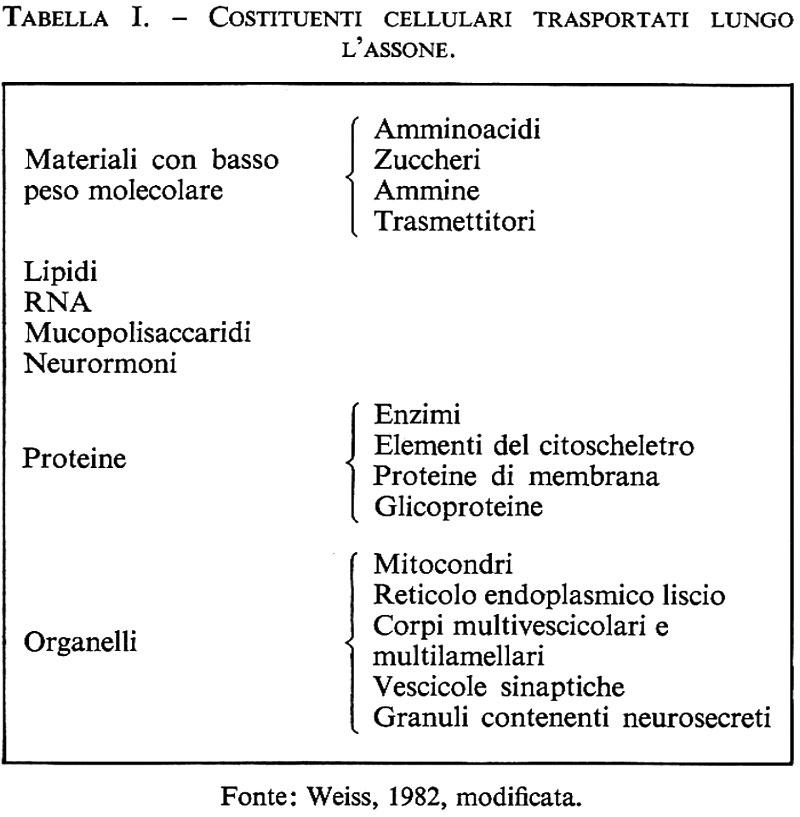
Come già precedentemente accennato, il trasporto di molecole lungo gli assoni è bidirezionale e si svolge quindi dal corpo cellulare verso la periferia (trasporto anterogrado) e viceversa (trasporto retrogrado). Le differenti velocità di migrazione delle varie componenti rappresentano una caratteristica fondamentale del trasporto assonale e sono state sperimentalmente dimostrate e investigate in dettaglio, soprattutto per quanto riguarda il trasporto anterogrado. Si distinguono cosi classicamente un trasporto anterogrado lento e un trasporto anterogrado rapido, mentre i meccanismi del trasporto retrogrado sembrerebbero per lo più basati su un flusso rapido. Sono state inoltre individuate recentemente anche velocità di migrazione intermedie.
Come verrà successivamente precisato, le varie componenti del trasporto assonale differiscono fra loro non so lo per quanto riguarda la velocità di migrazione, ma anche per i tipi di materiale trasportato. Inoltre, secondo alcune ipotesi attualmente molto accreditate (v. cap. 5), ciascuna delle componenti sarebbe collegata a strutture cellulari specifiche. Tali dati sono riassunti nella tab. II. Oltre al tipo di suddivisione presentato nella tab. II, viene anche largamente seguita, per quanto riguarda il trasporto anterogrado, una classificazione in 5 gruppi proposta da M. Willard e altri (v., 1974). Secondo tale classificazione il trasporto anterogrado rapido Costituisce il I gruppo, le due Componenti del trasporto intermedio costituiscono rispettivamente il II e il III gruppo, mentre le due componenti a e b del trasporto anterogrado lento rappresentano rispettivamente il IV e il V gruppo.
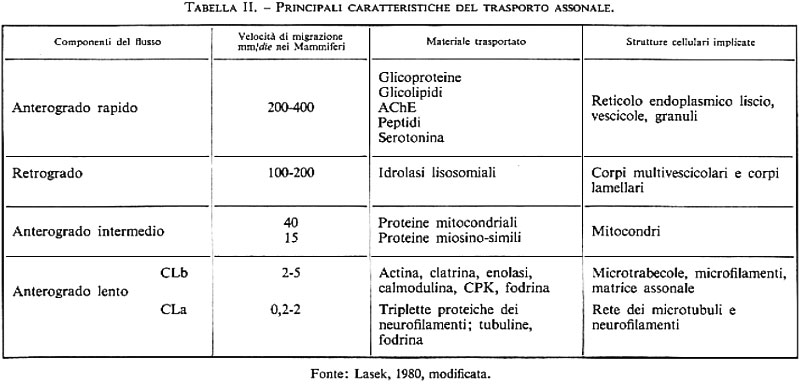
In termini molto generali, che verranno successivamente precisati, la funzione principale delle proteine trasportate lungo la componente rapida sarebbe quella di consentire le interazioni fra il neurone e l'ambiente circostante. Le proteine trasportate lungo il flusso anterogrado rapido sono quindi associate principalmente a membrane e sono destinate all'assolemma e ai terminali sinaptici. Le proteine trasportate nella componente lenta sono invece essenzialmente deputate alla costituzione e al mantenimento delle diverse componenti del citoscheletro e degli enzimi coinvolti nel metabolismo del neurone. La componente lenta è quindi principalmente destinata alla costituzione della struttura interna dell'assone ed è essenzialmente deputata a funzioni intrinseche intraassonali.
a) Trasporto anterogrado rapido
La Componente rapida del trasporto assonale anterogrado è coinvolta nel mantenimento dei sistemi di membrane dell'assone e dei terminali assonici; essa convoglia i costituenti delle membrane e delle vescicole sinaptiche, gli enzimi implicati nel metabolismo dei trasmettitori, i fattori che mediano le interazioni trofiche fra i neuroni e le loro cellule bersaglio, i neurormoni (v. anche neurosecrezione), ecc. Sono probabilmente trasportati nella fase rapida anche i materiali coinvolti nelle interazioni specifiche fra membrane pre- e postsinaptiche. Una delle principali funzioni di tale componente rapida anterograda consisterebbe inoltre nella regolazione dei meccanismi di rapida emissione di materiali dal terminale presinaptico.
Per assolvere le funzioni sopraelencate, vengono trasportati nella fase rapida del trasporto assonale molti costituenti di membrana (glicoproteine, fosfolipidi, glicolipidi) e, in particolare, proteine specifiche associate a membrane (inclusi gli enzimi specifici, quale la dopammina-β-idrossilasi). Vengono anche trasportati nella fase rapida altri materiali, quali i trasmettitori contenuti nelle vescicole sinaptiche, e molte sostanze a basso peso molecolare, quali amminoacidi, glicidi, nucleosidi e ioni.
La velocità massima del trasporto anterogrado rapido è pari a circa 400 mm/die, ma la maggior parte del materiale trasportato in tale componente ha una velocità di migrazione di circa 300 mm/die. La velocità del trasporto è costante negli assoni di ciascun nervo, indipendentemente dalla loro lunghezza, dal loro diametro e dal loro grado di mielinizzazione.
La componente rapida del trasporto anterogrado è principalmente rappresentata da materiale associato a strutture cellulari membranose. Le proteine della componente rapida sono associate al reticolo endoplasmico liscio. Quest'ultimo e i granuli (elettrondensi alle indagini ultrastrutturali) contenenti il materiale trasportato vengono sintetizzati nel corpo cellulare dal reticolo endoplasmico rugoso e si muovono lungo l'assone come singole unità (v. anche neurosecrezione). Quindi il movimento coordinato di migrazione lungo l'assone è dovuto, nella componente rapida, a strutture complesse, rappresentate da organelli legati a membrane all'interno dell'assone stesso.
Il trasporto rapido può essere influenzato da vari fattori. Ad esempio, la componente rapida del trasporto è strettamente dipendente dai processi metabolici che si svolgono lungo i vari segmenti dell'assone. Le varie fasi del metabolismo ossidativo rappresentano quindi un fattore critico per il trasporto rapido, che richiede un apporto continuo di ATP dalla fosforilazione ossidativa e viene completamente bloccato dall'anossia.
La velocità del trasporto rapido è inoltre dipendente dalla temperatura. Vi è una temperatura critica (11-13 °C nei nervi dei Mammiferi e 5-10 °C nei poichilotermi) al di sotto della quale il trasporto va incontro a un blocco (v. Grafstein e Forman, 1980). L'attività elettrica dell'assone non sembra invece costituire un fattore critico per la velocità di migrazione della fase rapida; tuttavia l'attività elettrica sembra influenzare la quantità di materiale trasportato. La fase rapida del trasporto assonale è inoltre dipendente dalla concentrazione extracellulare di ioni. In particolare, tale componente viene inibita da basse concentrazioni di calcio e questa inibizione può essere contrastata dall'aggiunta di ioni potassio e magnesio.
Nonostante il trasporto rapido sia preposto a veicolare elementi derivati dalla sintesi proteica, esso non è tuttavia strettamente dipendente da quest'ultima, come attestato dall'evidenza sperimentale che le componenti rapide non vengono bloccate da inibitori della sintesi proteica. La fase rapida del trasporto assonale viene invece bloccata da agenti che danneggiano specificamente i microtubuli, quali la colchicina, la vinblastina, la vincristina, e da anestetici generali e locali. Questi ultimi hanno però un effetto di blocco a dosi più alte di quelle terapeutiche. La fase rapida viene inoltre bloccata da sostanze che agiscono sull'ingresso intracellulare del sodio, come la batracotossina.
Un importante ruolo nel dare inizio ai meccanismi di trasporto rapido verrebbe svolto da strsstture vescicolari avvolte da membrane. È stato infatti ipotizzato (v. Stone e altri, 1984) che, in seguito alla sintesi nel reticolo endoplasmico rugoso e al successivo trasferimento al reticolo endoplasmico liscio, le proteine destinate al trasporto rapido procedano verso l'apparato di Golgi sotto forma di ‛vescicole di transizione' e di tubuli. Le proteine della componente rapida verrebbero poi emesse dall'apparato di Golgi sotto forma di vescicole avvolte da membrane, e così verrebbero convogliate agli elementi del reticolo assonale che fungono da vettori del trasporto (v. fig. 5). Le membrane delle vescicole, esaurita la loro funzione, possono poi andare incontro a un processo di fusione, mediante un meccanismo calcio-dipendente, e formare così i costituenti stessi del reticolo assonale. Il trasporto intracellulare avverrebbe quindi tramite ondate successive di vescicole, che si formano in una regione e si fondono con la successiva (v. cap. 5). La fusione di vescicole e membrane verrebbe regolata dal calcio in tutte le sue fasi.
Tali meccanismi sono simili a quelli che riguardano le proteine destinate a essere secrete in sistemi non neuronali, nel senso che le proteine destinate a processi di secrezione (quali, ad esempio, gli enzimi digestivi del pancreas o le immunoglobuline delle cellule immunocompetenti) rimangono associate a membrane durante tutta la loro vita intracellulare.
b) Trasporto anterogrado intermedio
L'evidenza di una componente del trasporto assonale con velocità di migrazione intermedia è derivata dall'identificazione di un gruppo di polipeptidi, il III gruppo di Willard e altri (v., 1974), che viene trasportato in vari sistemi a una velocità di circa 3-20 mm/die. Una proteina miosino-simile, che si lega all'actina, è l'unica proteina di questo gruppo finora identificata.
c) Trasporto anterogrado lento
Al contrario della componente rapida, nella quale vengono essenzialmente trasportati organelli legati a membrane, la componente lenta è principalmente implicata nel trasporto di materiali citoplasmatici. La componente lenta del trasporto anterogrado include infatti microtubuli, neurofilamenti e altre proteine assoplasmatiche.
Il trasporto lento di proteine è stato suddiviso in due sottocomponenti: la più lenta, denominata componente lenta a (CLa), corrisponde al V gruppo di Willard e altri (v., 1974), mentre una componente b (CLb), relativamente più veloce, corrisponde al IV gruppo nella classificazione di tali autori. Ciascuna di queste due componenti lente è collegata a differenti strutture subcellulari (v. Black e Lasek, 1980; v., anche, tab. II), che sarebbero specificamente associate all'una o all'altra componente (v. cap. 5, § b). Nella CLa sono essenzialmente coinvolti i microtubuli e i neurofilamenti, mentre le strutture cellulari implicate nella CLb sono principalmente rappresentate dal sistema microtrabecolare e dalla matrice assoplasmatica.
CLa. - Tale sottocomponente rappresenta l'onda più lenta del materiale trasportato e ha, nei Mammiferi, una velocità di migrazione di 0,2-2 mm/die. La CLa è essenzialmente basata su cinque proteine. Due di esse costituiscono subunità della tubulina e hanno un peso molecolare di 53.000 e 57.000. Le altre tre proteine sono costituenti dei neurofilamenti e vengono sempre trasportate insieme lungo l'assone, all'interno del quale sono strutturalmente associate fra loro. Per queste loro caratteristiche tali proteine sono state denominate ‛tripletta della componente lenta' (v. Hoffman e Lasek, 1975).
CLb. - La sottocomponente b del trasporto anterogrado lento ha una velocità di migrazione di tre o quattro volte maggiore della precedente. La CLb ha una struttura molto più complessa della CLa, poiché sono in essa coinvolte almeno 20 proteine maggiori e molte proteine minori (v. Black e Lasek, 1980). Uno dei polipeptidi costituenti la CLb è l'actina, che è presente in tutte le parti del neurone (corpo cellulare, dendriti, assone e sinapsi). Come precedentemente accennato (v. cap. 3, È a), i filamenti di actina sono elementi costitutivi del citoscheletro, e in particolare delle strutture microtrabecolari. Le proteine solubili della CLb sono invece presumibilmente conglobate nella matrice assoplasmatica (v. Lasek, 1980), costituita a sua volta da enzimi solubili implicati nel metabolismo intraassonale, che circondano i microfilamenti e le microtrabecole.
La temperatura non sembra esercitare alcun effetto rilevante sulla componente lenta, a differenza della componente rapida, sulla quale le basse temperature esercitano un effetto di rallentamento o di blocco (v. §§ a). Un'altra differenza fra le due componenti è rappresentata dall'evidenza sperimentale che la sintesi proteica costituisce un fattore critico per la componente lenta e non per quella rapida (v. § a). Il trasporto della componente lenta si arresta infatti in seguito a blocco della sintesi proteica. Al contrario, l'effetto di agenti, come la colchicina, che danneggiano i microtubuli e provocano un arresto della componente rapida, è assai meno cospicuo sulla componente lenta.
d) Destino del materiale trasportato in via anterograda
Esiste una popolazione molto eterogenea di proteine, generalmente chiamate ‛proteine sinaptiche', che sono solubili nell'assoplasma dei terminali neuronali, oppure sono associate a varie componenti subcellulari, quali le vescicole sinaptiche, le membrane plasmatiche presinaptiche, i mitocondri, i microtubuli, i microfilamenti. Come è noto, vi è un continuo ricambio e quindi un continuo avvicendamento di tali proteine sinaptiche, e sono stati chiamati in causa molteplici meccanismi cellulari (v. Droz, 1973) per spiegare tale fenomeno.
La degradazione delle proteine sinaptiche sarebbe essenzialmente basata su quattro meccanismi. Il primo di questi è l'emissione dai terminali, mediante la deplezione delle vescicole sinaptiche. Tale processo è dovuto a fusione della vescicola con la membrana plasmatica, seguita da emissione del neurotrasmettitore mediante un meccanismo di esocitosi. Con un meccanismo simile si verifica anche la liberazione dei neurormoni nelle cellule neurosecretorie (v. neurosecrezione). Inoltre un ruolo quantitativamente cospicuo nella degradazione delle proteine sinaptiche verrebbe svolto da processi catabolici locali, che hanno luogo cioè nel terminale presinaptico stesso. Un terzo meccanismo è rappresentato dal trasporto assonale retrogrado, le cui dinamiche verranno esposte in dettaglio nel paragrafo successivo. È stato infatti sperimentalmente dimostrato che una frazione proteica viene riconvogliata in senso distoprossi male verso il corpo cellulare; le proteine verrebbero poi ivi idrolizzate dagli enzimi lisosomiali presenti nel pericario. I criteri sulla base dei quali vengono ‛selezionate' le proteine che devono essere convogliate lungo la componente retrograda del flusso assonale non sono stati finora identificati. Infine il quarto meccanismo è rappresentato da processi di degenerazione. Questi ultimi sono soprattutto implicati nella scomparsa di costituenti sinaptici nel corso dei processi fisiologici di invecchiamento. Le alterazioni degenerative dei terminali presinaptici sono caratterizzate, dal punto di vista morfologico, da un addensamento di strutture membranose, mitocondri e lisosomi, a causa del quale i terminali appaiono allargati e rigonfi. I terminali degenerati, fagocitati dalle cellule gliali adiacenti, verrebbero infine disintegrati da citolisosomi.
Le proteine sinaptiche, eliminate mediante uno o più dei meccanismi sopra elencati, vengono poi sostituite e rinnovate mediante la migrazione assonale di macromolecole sintetizzate nel corpo cellulare, e trasportate ai terminali secondo i vari meccanismi descritti nei paragrafi precedenti. Un contributo quantitativamente minoritario al rinnovo delle proteine sinaptiche viene apportato anche da altri meccanismi, quali l'endocitosi di macromolecole presenti nello spazio extracellulare e lo scambio di molecole polipeptidiche con le cellule gliali o le strutture postsinaptiche (v. Droz, 1973).
e) Trasporto retrogrado
L'identificazione della bidirezionalità del trasporto assonale (v. Lubinska, 1975) ha messo in luce l'esistenza di componenti del flusso deputate a veicolare materiali dalla periferia al corpo cellulare. Il flusso retrogrado ha ricevuto inoltre molteplici conferme sperimentali dall'osservazione che traccianti proteici, iniettati nel muscolo (v. Kristensson e Olsson, 1973) o nel sistema nervoso centrale (v. cap. 9), venivano poi identificati nei corpi cellulari che innervavano l'area di iniezione. Il più noto di tali traccianti esogeni è la glicoproteina HRP (v. cap. 2), una perossidasi del peso molecolare di 40.000, composta da più di 20 isoenzimi. L'HRP può essere facilmente visualizzata, in microscopia ottica, mediante una reazione istochimica specifica: il prodotto di reazione dell'HRP è infatti facilmente riconoscibile sotto forma di granuli scuri. L'applicazione dell'HRP, il cui prodotto di reazione è elettrondenso, si è inoltre dimostrata molto efficace nelle indagini ultrastrutturali sulle dinamiche del trasporto retrogrado. Inoltre molti dati sperimentali sul trasporto retrogrado sono derivati dagli studi sul trasporto anterogrado, attuati mediante marcatura radioisotopica, nei quali è stata indagata la componente che, giunta alla periferia, veniva nuovamente veicolata verso il corpo cellulare.
I meccanismi e le funzioni specifiche del trasporto retrogrado sono tuttora poco conosciuti. Tuttavia viene concordemente ascritta al trasporto retrogrado, nella letteratura, la funzione di ‛informare' il corpo cellulare di ciò che avviene alla periferia. Ad esempio, il flusso retrogrado è parte di un circuito riverberante (a feedback), che tiene costantemente informato il corpo cellulare sullo stato funzionale delle sinapsi. È particolarmente dimostrativo, a tale proposito, il modello sperimentale allestito da J. M. Aletta e D. J. Goldberg (v., 1982) su un neurone serotoninergico di Aplysia californica. Tale cellula, di grandi dimensioni, si biforca in due branche assonali dirette verso la periferia. In seguito alla sezione di una di queste due branche, le vescicole contenenti il neurotrasmettitore, normalmente destinate alla branca rimossa, vengono dapprima veicolate verso la branca intatta, che riceve, quindi, neurotrasmettitore in eccesso. Tuttavia, in tempi immediatamente successivi, si verifica una rapida regolazione nel corpo cellulare, in base alla quale l'emissione di vescicole viene ridotta fino a soddisfare solo il fabbisogno della branca residua. Ciò indica non solo che il corpo cellulare riceve continuamente dalla periferia un segnale che disciplina l'emissione del neurotrasmettitore, ma anche che tale meccanismo viene regolato con molta precisione da ciascuna delle branche periferiche dell'assone.
Per quanto riguarda il materiale trasportato, il flusso disto-prossimale sembrerebbe assolvere due principali funzioni. La prima di esse è rappresentata dal ‛ritorno' al corpo cellulare di alcuni costituenti (proteine, glicoproteine, fosfolipidi, enzimi, quali l'acetilcolinesterasi e la dopammina-β-idrossilasi) precedentemente veicolati verso la periferia. Gli studi condotti sulle frazioni di materiale che vengono riconvogliate in senso retrogrado indicano percentuali molto variabili, dal 10 al 70%, delle quantità precedentemente avviate nel flusso anterogrado (v. Grafstein e Forman, 1980). In secondo luogo, il flusso retrogrado sembrerebbe svolgere un importante ruolo nell'assunzione di macromolecole e di alcune molecole di piccole dimensioni dallo spazio extracellulare. Ad esempio, la componente retrograda costituisce un'importante via per il trasferimento di fattori di accrescimento, così come per la penetrazione di tossine e di virus neurotropi (v. cap. 6, § f). Non sono stati finora identificati i meccanismi che determinano l'assunzione di materiale dallo spazio extracellulare, ma sembrerebbero a tale proposito importanti alcune caratteristiche, quali la carica elettrica e il peso molecolare delle sostanze, nonché il grado di attività sinaptica del neurone (v. Kristensson, 1978) e, ovviamente, la presenza di recettori di membrana.
Il significato biologico del trasporto retrogrado sembra quindi essere collegato a tre funzioni principali (v. Kristensson, 1981): a) il trasferimento di membrane dai terminali assonici al corpo cellulare, al fine di destinarle a una degradazione nei lisosomi (vedi oltre), come parte di meccanismi di ricambio (turnover) delle membrane stesse; b) il trasferimento di fattori macromolecolari di accrescimento dal campo periferico di innervazione al corpo cellulare; c) un meccanismo a feedback di regolazione dell'attività metabolica del corpo cellulare in relazione all'attività sinaptica.
L'assunzione di molecole dal terminale presinaptico avviene mediante pinocitosi, un processo di introflessione della membrana plasmatica, mediante il quale il materiale fluido viene incorporato, dallo spazio extracellulare, in vescicole intracellulari. Nel corso di tale processo di endocitosi le sostanze incorporate si possono legare alle membrane plasmatiche, all'interno delle vescicole, mediante un meccanismo di adsorbimento alle membrane stesse, oppure possono rimanere, nelle vescicole, allo stato di materiale fluido. È inoltre da tenere presente che, in seguito a lesioni dell'assone, le alterazioni della permeabilità di membrana possono consentire un ingresso diretto di sostanze e una loro rapida diffusione all'interno dell'assone stesso (v. Kristensson e Olsson, 1976).
La velocità di migrazione della componente retrograda è molto variabile e dipende dalla sostanza, dalla specie dell'animale, dai sistemi neuronali esaminati. Il trasporto sarebbe comunque collegato a un flusso rapido, ma con velocità inferiori a quelle della componente anterograda rapida, e generalmente pari a valori fra i 70 e i 200 mm/die. È stato inoltre recentemente dimostrato anche un trasporto retrogrado lento (v. Fink e Gainer, 1980). La componente retrograda lenta ha una velocità di 3-8 mm/die e vi sarebbe principalmente adibita una proteina, dal peso molecolare di 60.000, simile all'albumina serica (v. Gainer e Fink, 1982).
Il trasporto retrogrado, come la componente anterograda rapida, è sensibile alla temperatura (ed è quindi rallentato o bloccato dalle basse temperature) ed è dipendente dall'apporto energetico. Il trasporto retrogrado è inoltre sensibile a molti degli agenti che bloccano la componente anterograda rapida, quali la colchicina e la vinblastina.
Per quanto riguarda le strutture impegnate nel trasporto, le indagini ultrastrutturali sul trasporto dell'HRP (v. Kristensson e Olsson, Uptake..., 1971) hanno dimostrato che il materiale trasportato in via retrograda viene incorporato in organelli avvolti da membrane e in corpi multivescicolan. Una volta raggiunto il corpo cellulare, la sostanza trasportata (rappresentata in tali studi dalla proteina HRP) è stata identificata nel citoplasma, nei dendriti e anche in prossimità della membrana nucleare, ma non all'interno del nucleo stesso. Nel citoplasma il tracciante HRP è sempre stato osservato incorporato in organelli, e non è mai stato identificato, quindi, libero. È stato inoltre osservato che le vescicole pinocitotiche contenenti il tracciante si fondono con i lisosomi presenti nel citoplasma del pericario neuronale; il destino ultimo del materiale trasportato, o almeno di una parte di esso, verrebbe quindi affidato a una degradazione catabolica a opera di enzimi lisosomiali.
Nonostante che il reticolo endoplasmico liscio sia stato più volte correlato ai meccanismi di trasporto retrogrado, recenti indagini ultrastrutturali hanno dimostrato che quest'ultimo non si svolge entro le cisterne del reticolo endoplasmico liscio, bensì in organelli da esso segregati (v. LaVail e altri, 1980). Ciò costituirebbe quindi un'importante differenza strutturale fra le dinamiche del trasporto retrogrado e quelle del trasporto anterogrado, nel quale, come precedentemente accennato, il reticolo endoplasmico svolge un ruolo fondamentale. Tali differenze sono anche confermate da questa osservazione: il reticolo endoplasmico liscio è comunque deputato anche al trasporto anterogrado delle stesse sostanze che possono poi essere veicolate, al di fuori di esso, in senso retrogrado.
5. Meccanismi che regolano il trasporto assonale
Nonostante gli studi dettagliati sulle varie componenti strutturali dell'assone e su molti aspetti dinamici del trasporto assonale, rimangono ancora da chiarire i meccanismi ultimi che determinano la progressione delle molecole lungo l'assone. È infatti tuttora aperto, nella letteratura, un vivace dibattito sulle modalità di movimento delle varie sostanze lungo l'assone, tanto che sono state avanzate molte ipotesi e sono stati elaborati vari modelli teorici. Le principali teorie a tale proposito verranno qui esposte nelle loro formulazioni essenziali.
a) Trasporto rapido
Sono stati ipotizzati essenzialmente tre gruppi di meccanismi di regolazione della componente rapida del trasporto (v. Grafstein e Forman, 1980). Il primo gruppo di teorie prevede l'applicazione ripetuta di forze propulsive agli organelli trasportati, da parte di strutture ancorate all'assone lungo tutta la sua estensione. Secondo tale ipotesi, le vescicole potrebbero legarsi ai microtubuli ed essere quindi trasportate mediante una modalità ‛saltatoria', strettamente dipendente da processi energetici. È stata inoltre avanzata l'ipotesi che i meccanismi propulsivi agiscano sui microtubuli tramite variazioni nella conformazione dei poli elettrolitici anionici di questi ultimi, legati alla tubulina, oppure mediante movimenti delle protuberanze dei microtubuli. È esemplificativa di tale gruppo di teorie l'‛ipotesi unitaria' avanzata da S. Ochs (v., 1982), fondata su un modello del ‛filamento di trasporto', che costituirebbe il vettore unico delle varie fasi del trasporto, incluse le componenti del trasporto lento. Tale ipotesi, derivata dalla constatazione di una stretta dipendenza del trasporto rapido dall'apporto energetico fornito dalla fosforilazione ossidativa, prevede che le molecole trasportate siano legate a un vettore comune, convogliato lungo i microtubuli tramite le appendici di questi ultimi. Tali protuberanze infatti, con l'apporto di ATP, varierebbero la loro conformazione, conferendo ai filamenti di trasporto una spinta propulsiva. M. H. Ellisman (v., 1982) ha inoltre ipotizzato che il trasporto sia dovuto a interazioni focali fra i sistemi di membrana dell'assone e i ponti microtrabecolari della matrice assoplasmatica.
In base a un secondo gruppo di ipotesi, del tutto opposte alle precedenti, il trasporto verrebbe regolato da movimenti di materiale fluido intraassonale. Secondo tali ipotesi, infatti, l'assoplasma verrebbe considerato come un gel percorso da correnti dinamiche. È un esempio di tale indirizzo teorico l'‛ipotesi della microcorrente', avanzata da D. G. Weiss e G. W. Gross (v., 1982) e basata su un principio idrodinamico. Secondo tali autori, il trasporto lungo l'assone sarebbe dovuto a correnti vettrici, generate da forze non specifiche, che si eserciterebbero su tutti i costituenti citoplasmatici. Il microtubulo costituirebbe quindi la struttura vettrice che orienta e stabilizza gli enzimi generatori di energia. La bidirezionalità del flusso sarebbe condizionata da microflussi in direzioni opposte, determinati dalla polarità intrinseca dei microtubuli.
Il terzo gruppo di ipotesi assegna invece un ruolo centrale al reticolo endoplasmico liscio, poiché il trasporto si svolgerebbe essenzialmente entro un sistema continuo, rappresentato dal reticolo endoplasmico stesso, lungo l'assone. In seguito alla fase iniziale del trasporto, i materiali, emessi dall'apparato di Golgi, verrebbero rapidamente introdotti entro un sistema continuo, costituito dal reticolo endoplasmico che si estende lungo l'assone (v. Hammerschlag e Stone, 1982). I costituenti di membrana, intimamente associati al reticolo endoplasmico liscio, verrebbero smistati ai rispettivi siti operativi nei terminali o nell'assolemma, seguendo canali diversi nei tubuli oppure nelle cisterne subassolemmali (v. Droz e Rambourg, 1982).
b) Trasporto lento
Sono state avanzate molte teorie anche per spiegare le componenti lente del trasporto assonale. È stato così ipotizzato che il trasporto lento si basi su meccanismi di tipo peristaltico (v. Weiss, 1967), oppure che entrino in gioco meccanismi simili a quelli chiamati in causa nel trasporto rapido. Riguardo alle componenti del trasporto lento è stata elaborata la cosiddetta ‛teoria centrale del trasporto assonale' (v. Hoffman e Lasek, 1975; v. Lasek, 1980). Secondo tale teoria, attualmente molto accreditata, le proteine si muoverebbero lungo l'assone come costituenti di strutture citologiche. Ciascuna componente (CLa e CLb) del trasporto lento corrisponderebbe a una o più strutture cellulari che si muovono lungo l'assone. Tale rapporto sarebbe specifico, nel senso che a ogni struttura cellulare corrisponderebbe un'unica componente del trasporto assonale e a ciascuna componente del trasporto assonale corrisponderebbero strutture subcellulari specifiche. Secondo tale teoria il trasporto sarebbe dovuto a ponti di miosina che, dai microtubuli o dai neurofilamenti, interagiscono con filamenti di actina ancorati all'assolemma. Quindi nella CLa tutto il sistema di microtubuli e neurofilamenti verrebbe lentamente convogliato lungo l'assone dalla pressione esercitata contro la membrana assonale. I materiali trasportati nella CLb, e cioè non solo le proteine strutturali, quale l'actina, ma anche enzimi metabolici solubili, si muoverebbero come complessi strutturali del sistema di microfilamenti e microtrabecole.
6. Caratteristiche del trasporto di alcune sostanze
a) Glicoproteine
Le glicoproteine sono tipicamente associate alle componenti rapide del trasporto e sono per lo più legate a strutture membranose. Le proteine fucosilate, principalmente trasportate entro organelli, sono primariamente destinate alle terminazioni nervose. Una parte di esse verrebbe tuttavia trasportata nell'assolemma. È stato infatti ipotizzato (v. Marchisio e altri, 1975) che il meccanismo di base del trasporto sia duplice: un trasporto assoplasmico, che veicola glicoproteine legate a vescicole e alle membrane del reticolo endoplasmico liscio, e un trasporto assolemmale, coinvolto nel trasporto di glicoproteine alle porzioni terminali e preterminali della membrana assonale, dove esse possono essere accumulate.
b) Lipidi
Il trasporto di fosfolipidi si svolge sia in senso anterogrado che in senso retrogrado ed è intimamente legato al flusso rapido di proteine, con il quale condivide alcune caratteristiche fondamentali, quali la distribuzione subcellulare e la sensibilità alla temperatura e a inibitori della sintesi proteica (v. Miani, 1963). Sulla base di tali dati è stato quindi ipotizzato che fosfolipidi e proteine vengano veicolati insieme. Tuttavia, al contrario delle proteine, che vengono avviate al trasporto anterogrado subito dopo la loro sintesi, i lipidi possono rimanere disponibili per il trasporto durante un periodo, relativamente prolungato, successivo alla loro sintesi. I lipidi vengono infatti prelevati da un pool nel corpo cellulare e, nonostante vengano veicolati rapidamente, possono accumularsi lentamente nell'assone e nei terminali. Gangliosidi e glicolipidi mostrano anch'essi le caratteristiche, precedentemente elencate per i fosfolipidi, di un trasporto rapido, intimamente collegato a quello delle proteine, con un prolungato tempo di emissione dal corpo cellulare. Vi sono però indicazioni che nel trasporto di alcuni costituenti lipidici, come il colesterolo, siano anche coinvolte delle componenti lente.
c) Nucleosidi
I nucleosidi svolgerebbero un importante ruolo nelle comunicazioni fra i neuroni e i loro bersagli sinaptici, ed è stato ipotizzato che essi possano agire come neurotrasmettitori in alcuni sistemi. Lo studio dei nucleosidi è stato principalmente basato sull'uso di adenosina e uridina marcate con trizio (v. Wise e altri, 1978; v. Repérant e altri, 1982). È stato così stabilito che il trasporto dei nucleosidi è bidirezionale. Inoltre, il trasporto anterogrado di tali sostanze può dare luogo a fenomeni di successivo trasporto trans-sinaptico e, in particolare, a un trasferimento a cellule gliali. Il trasferimento transneuronale può successivamente dare luogo a un trasporto secondario, sia in via anterograda che in via retrograda, da parte di neuroni adiacenti.
d) Trasmettitori sinaptici ed enzimi a essi correlati
I neurotrasmettitori vengono generalmente trasportati lungo le componenti rapide e sono veicolati in senso anterogrado, dal corpo cellulare alla sinapsi, dove vengono utilizzati. Tuttavia, come dimostrato dall'applicazione esogena di neurotrasmettitori triziati (v. cap. 9, È b), tali sostanze possono essere trasportate anche in senso retrogrado. Quest'ultima componente sarebbe però, in condizioni fisiologiche, molto esigua dal punto di vista quantitativo. La noradrenalina viene trasportata a una velocità di 70-240 mm/die nel nervo sciatico di mammiferi. Il trasporto nel sistema nervoso centrale sarebbe più lento che nei nervi periferici. L'acetilcolina è trasportata per circa il 40% legata a organelli. Sembra che il trasporto di tale neurotrasmettitore avvenga, almeno nelle fasi iniziali, mediante incorporazione in vescicole, ma in parte esso si muoverebbe anche per diffusione intraassonale. La velocità di migrazione, nei Vertebrati, è di circa 100 mm/die, ma sono state anche segnalate velocità di migrazione di 200-250 mm/die. La velocità di migrazione della serotonina si aggirerebbe intorno ai 130 mm/die, ma tale parametro sembra essere molto variabile. La velocità di migrazione della dopammina sarebbe invece, nel midollo spinale di mammiferi, simile a quella osservata per la noradrenalina. Il trasporto della dopammina viene studiato mediante la somministrazione di dopa, il precursore, marcato con isotopo; è prerequisito per il trasporto che si verifichi la decarbossilazione da dopa a dopammina.
Per quanto riguarda gli enzimi correlati al metabolismo dei neurotrasmettitori, le loro velocità di migrazione, studiate prevalentemente mediante la tecnica dell'accumulo, sono generalmente interpretate in termini di trasporto lento o rapido, ma non sempre rientrano nell'ambito di queste due grandi categorie (v. Grafstein e Forman, 1980). La maggior parte di tali enzimi viene veicolata sia in frazioni solubili che legata a organelli, e le due frazioni potrebbero quindi essere veicolate a velocità diverse. L'acetilcolinesterasi è una delle prime sostanze delle quali sia stato individuato un trasporto bidirezionale. Il trasporto anterogrado di tale enzima avrebbe una velocità simile a quella del trasporto rapido di proteine, mentre la quantità di enzima veicolata in senso retrogrado è generalmente inferiore a quella delle proteine trasportate in direzione disto-prossimale. L'enzima coinvolto nella sintesi dell'acetilcolina, la colinacetiltransferasi, viene invece veicolato mediante le componenti del flusso lento. La dopammina-β-idrossilasi è correlata, come l'acetilcolinesterasi, al flusso rapido, e il suo trasporto è bidirezionale. Le componenti anterograda e retrograda della dopammina-β-idrossilasi sono trasportate in compartimenti separati. È stato determinato che la frazione che ritorna in senso disto-prossimale al corpo cellulare è per lo più in forma inattiva dal punto di vista enzimatico, e sarebbe quindi destinata essenzialmente ai meccanismi di degradazione catabolica. La tirosina-idrossilasi viene invece veicolata a velocità più lente della dopammina-β-idrossilasi, ed è stata identificata soprattutto in forma solubile. Anche l'altro enzima correlato al metabolismo della dopammina, la dopadecarbossilasi, è soprattutto veicolato in forma solubile, con velocità inferiori a quelle dei due enzimi precedentemente elencati, ma superiori alle velocità di migrazione delle componenti lente vere e proprie.
e) Costituenti gliali
Come indicato nell'esauriente trattazione di B. Grafstein e D. S. Forman (v., 1980), alcuni studi hanno dimostrato che, oltre ai costituenti assonali, possono essere veicolate mediante trasporto assonale anche alcune sostanze tipicamente presenti nelle cellule gliali. Tali osservazioni presentano un particolare interesse, in quanto il trasferimento a cellule gliali di materiali sintetizzati nell'assone può costituire una peculiare modalità di interazione fra glia e neurone. Un esempio di tali sostanze è rappresentato dalla proteina S-100, che, tipicamente associata alle cellule gliali o alle cellule di Schwann, è stata anche identificata in cellule nervose. Secondo N. Miani e altri (v., 1972), la proteina S-100 viene veicolata lungo l'assone mediante le componenti del trasporto lento. Anche altre proteine costituenti la mielina, sintetizzate primariamente in cellule di Schwann o oligodendrogliali, sono state isolate da frazioni proteiche trasportate lungo l'assone. Le proteine mieliniche sono per lo più associate al flusso lento, ma una piccola parte di esse è stata anche identificata nelle componenti del trasporto rapido.
f) Trasporto retrogrado di proteine speciali e virus
Virus. - È noto da tempo che un trasporto neuronale selettivo lungo i tronchi nervosi periferici può costituire la via di ingresso e di diffusione di virus neurotropi (v. Goodpasture, 1925). Ciò è stato ipotizzato per il virus della rabbia, della poliomielite, per virus erpetici. Molti recenti studi, soprattutto in ambito neuropatologico, sono stati rivolti all'indagine del trasporto assonale del virus dell'herpes simplex. Quest'ultimo può infatti infettare molti tipi di neuroni dopo assunzione da parte dei terminali, e si diffonde mediante meccanismi di trasporto assonale retrogrado rapido, come è stato possibile verificare dalla sua distribuzione, corrispondente alle interconnessioni, ben note, fra determinati circuiti neuronali. Ad esempio, dopo iniezione nel nucleo striato del ratto, il virus è stato ritrovato solo nei corpi cellulari che inviano allo striato i loro assoni (v. Bak e altri, 1977). È stato inoltre dimostrato un trasporto anterogrado sia del virus dell'herpes simplex sia di quello dell'herpes zoster, e tale trasferimento anterogrado (dai gangli sensitivi al loro territorio di innervazione) è stato chiamato in causa nella patogenesi delle infezioni cutanee e nella loro distribuzione selettiva ai dermatomeri innervati (v. Grafstein e Forman, 1980). Tuttavia non è stato ancora chiarito il ruolo che il trasporto retrogrado di virus neurotropi svolge nella patologia umana, anche se esso sembrerebbe coinvolto nella distribuzione di virus che, in seguito alla fase viremica, possono superare la barriera ematoencefalica (v. neuropatologia: Malattie virali del sistema nervoso centrale; v. virus).
Tossina tetanica. - La tossina tetanica raggiunge per via ematica i terminali dell'assone e viene da questi assunta e trasportata ai corpi cellulari. Il trasporto retrogrado della tossina tetanica è apparentemente non specifico e avrebbe una velocità pari a 60-300 mm/die. Il trasporto retrogrado inizierebbe mediante un legame selettivo alla membrana terminale del nervo e assunzione per pinocitosi. Quando raggiunge il corpo cellulare la tossina viene apparentemente trasferita, almeno in parte, ai terminali presinaptici che su di esso fanno sinapsi e dove la tossina esercita la sua azione. L'attività tossica non sembra costituire un prerequisito per il trasporto, poiché può essere avviato al trasporto anche un frammento antigenico della tossina, che non ne ha, però, l'attività tossica. L'assunzione della tossina tetanica dal sangue e la sua successiva diffusione tramite il trasporto assonale suggeriscono che tale meccanismo possa essere comune all'ingresso nel sistema nervoso di altre tossine batteriche e alla loro successiva distribuzione.
Fattore di accrescimento delle cellule nervose (Nerve Growth Factor, NGF). - L'NGF è una proteina che, nel sistema nervoso periferico, è essenziale per lo sviluppo e la sopravvivenza dei neuroni simpatici e di alcuni neuroni sensitivi. L'NGF, trasportato in via retrograda lungo l'assone, svolgerebbe infatti in tali neuroni un importante ruolo trofico e di regolazione metabolica, sia durante lo sviluppo che a maturazione ultimata. L'NGF causa una notevole ipertrofia dei neuroni simpatici in seguito a somministrazione in animali neonati (v. Levi-Montalcini, 1966; v. neurogenesi). Inoltre, in seguito al trasporto e alla localizzazione nei corpi delle cellule gangliari, l'NGF induce selettivamente un aumento di sintesi dell'enzima tirosinaidrossilasi e dopammina-β-idrossilasi (v. Paravicini e altri, 1975). Il trasporto retrogrado di NGF sembra essere altamente specifico. L'NGF che raggiunge terminali adrenergici o sensitivi periferici per somministrazione locale o endovenosa viene assunto con grande selettività e trasportato in via retrograda mediante un trasporto rapido ai corpi delle cellule gangliari. La specificità sarebbe dovuta al meccanismo di assunzione e non di trasporto, tanto è vero che la velocità di migrazione è simile a quella della tossina tetanica (v. Stöckel e altri, 1975). L'assunzione sarebbe mediata da recettori di membrana e l'NGF verrebbe quindi trasportato in vescicole e compartimenti tubulari.
L'interruzione del trasporto assonale retrogrado di NGF (per distruzione dei terminali adrenergici mediante neurotossine specifiche, per interferenza con il sistema di microtubuli o per somministrazione di anticorpi specifici) esercita degli effetti caratteristici sia in stadi precoci dello sviluppo che in animali adulti (v. Thoenen e altri, 1982). In roditori neonati il blocco del trasporto assonale di NGF provoca la degenerazione della maggior parte dei neuroni simpatici dei gangli paravertebrali. Nell'animale adulto l'interruzione dell'apporto di NGF provoca principalmente un'alterazione della sintesi degli enzimi specifici coinvolti nella produzione di noradrenalina e una generale riduzione di volume dei corpi delle cellule nervose bersaglio della sua azione. Tali dati dimostrano che l'NGF svolge fisiologicamente un importante ruolo per la sopravvivenza dei neuroni bersaglio durante lo sviluppo e che, a maturazione avvenuta, l'NGF svolge un ruolo trofico, mantenendo entro i limiti fisiologici il livello degli enzimi deputati alla sintesi dei neurotrasmettitori specifici.
È stato infine recentemente segnalato che nel sistema nervoso centrale si verifica assunzione di NGF da parte dei terminali di assoni colinergici che innervano la corteccia cerebrale, e un trasporto retrogrado selettivo ai loro corpi cellulari di origine, localizzati nel telencefalo basale (v. Seiler e Schwab, 1984). La specificità di tale trasporto indica la presenza di recettori di membrana per l'NGF anche in tali neuroni colinergici centrali.
7. Il trasporto assonale come meccanismo plastico
In varie situazioni fisiologiche, quale lo sviluppo, o patologiche, in seguito a eventi traumatici, la cellula nervosa ha la capacità di mettere in atto una serie di risposte, di meccanismi di adattamento, che vengono generalmente compresi nel termine ‛plasticità neuronale'. Il trasporto di sostanze lungo l'assone, assolvendo una serie di compiti essenziali per la sopravvivenza e la funzionalità del neurone, nonché per le sue interazioni con l'ambiente circostante, svolge anche un ruolo preminente nei meccanismi della plasticità neuronale. Pur essendo state negli ultimi anni oggetto di numerose indagini sperimentali, la regolazione e le dinamiche della plasticità neuronale sono tuttora largamente sconosciute. Tuttavia, per quanto riguarda l'implicazione del trasporto assonale nelle reazioni plastiche della cellula nervosa, due problemi sono stati affrontati in dettaglio: le relazioni fra assone e reazioni degenerative del corpo cellulare (in seguito a lesioni dell'assone o in condizioni patologiche dei suoi campi periferici di innervazione) e i fenomeni di rigenerazione assonale.
Per quanto riguarda il primo punto, è noto che in seguito a interruzione dell'assone si producono delle alterazioni nel corpo cellulare che possono riflettersi in variazioni morfologiche, quale la rarefazione dei corpi di Nissl, con diminuzione (‛cromatolisi') o aumento della basofilia del citoplasma. La cromatolisi riflette delle alterazioni dei fenomeni di sintesi proteica: la dispersione della sostanza di Nissl corrisponde al disgregarsi dei ribosomi collegati al reticolo endoplasmico rugoso, come pure dei gruppi di ribosomi liberi nel citoplasma. Poiché la cellula nervosa è, come è noto, grandemente influenzata dal suo campo di innervazione, si possono verificare fenomeni degenerativi nel corpo cellulare anche per lesioni molto periferiche. Ad esempio, la distruzione o l'atrofia dei muscoli scheletrici causa ipoplasia delle corna anteriori del midollo spinale. Pur non essendo le lesioni assonali o le alterazioni dei territori bersaglio del neurone dei modelli sperimentali esattamente equivalenti, in entrambi i casi si pone il problema di quale sia il ‛segnale' che il corpo cellulare riceve dalla sua periferia e che dà l'avvio alla reazione degenerativa. È opinione comune e accreditata che tale messaggio venga veicolato tramite i meccanismi del trasporto assonale retrogrado; ne è prova sperimentale indiretta il fatto che il cosiddetto ‛segnale della cromatolisi' e il trasporto assonale retrogrado di macromolecole, quale l'HRP, raggiungono il corpo cellulare con velocità simili (v. Kristensson, 1978). La natura del ‛segnale' è tuttavia del tutto sconosciuta e sono state suggerite (v. Lubinska, 1975) due possibilità. La prima è che esso venga affidato agli organelli già fisiologicamente trasportati in senso retrogrado e venga quindi da questi convogliato. La seconda possibilità è che vi sia un materiale specifico (la cosiddetta wound substance degli autori anglosassoni) che, dall'assone leso, solleciti una risposta del corpo cellulare all'evento lesivo. Quest'ultima ipotesi non ha comunque ricevuto finora conferme sperimentali, poiché non sono state finora identificate con certezza sostanze specifiche presenti solo in situazioni postlesionali.
Occorre tuttavia tener presente che un'interruzione del nervo causa un prematuro rientro di proteine verso il corpo cellulare e che la composizione di queste ultime sembra essere diversa da quella delle proteine degli assoni normali. Inoltre è stato più volte verificato il ruolo di fattori trofici specifici, quale l'NGF (v. cap. 6, § f), che mediano il trasferimento delle interazioni trofiche dalle cellule effettrici al neurone che le innerva. Ciò è ulteriormente comprovato dall'osservazione sperimentale che la reazione cromatolitica di cellule gangliari all'assotomia viene impedita dalla preventiva somministrazione esogena di NGF (v. Purves e Nja, 1976). Inoltre il trasporto retrogrado sarebbe non solo implicato nella segnalazione al corpo cellulare della lesione periferica, ma anche preposto a informarlo quando i terminali hanno ristabilito la reinnervazione dell'organo bersaglio (v. Kristensson, 1978).
Per quanto riguarda i fenomeni di crescita dell'assone, nel corso sia dello sviluppo sia della rigenerazione postlesionale, è stato definitivamente accertato che il trasporto assonale costituisce un meccanismo fondamentale, poiché fornisce i materiali necessari all'allungamento dell'assone stesso, che viene guidato dal cono di crescita (growth cone degli autori anglosassoni). Il cono di crescita dell'assone rappresenta una struttura chiave anche nella formazione di connessioni sinaptiche appropriate (v. Landis, 1983). Esso consiste in un allargamento protoplasmatico (il cono propriamente detto) e in sottili espansioni periferiche, che gli conferiscono motilità. Il sistema di microtubuli, responsabile del supporto strutturale dell'assone e quindi del trasporto di molecole essenziali per il suo allungamento, si estende fino alla base del cono di crescita e ne assicura la progressione, mentre la motilità del cono di crescita sarebbe essenzialmente affidata a filamenti di actina. Il fatto che tutte le proteine richieste per la rigenerazione dell'assone vengano fornite dalle fasi lente del trasporto assonale, e la constatazione, confermata in vari sistemi, che l'allungamento assonale procede, nel corso della rigenerazione, con un ritmo equivalente alla velocità di migrazione del trasporto lento, suggeriscono un rapporto di causa-effetto tra i due fenomeni. Le proteine delle componenti lente del trasporto assonale (e in particolare della CLb) costituirebbero quindi il fattore che determina il tasso di allungamento dell'assone (v. Lasek e altri, 1981; v. Veraa e Grafstein, 1981).
La rigenerazione assonale, studiata nel sistema nervoso periferico, si svolge essenzialmente in tre fasi (v. Veraa e Orafstein, 1981; v. Frizell e Sjöstrand, 1981). La prima fase è rappresentata dal cosiddetto periodo di latenza, immediatamente postlesionale; si verificano quindi l'accrescimento dell'assone e, ad accrescimento ultimato, la maturazione dell'assone stesso. Il periodo di latenza precede l'inizio dei fenomeni di rigenerazione vera e propria e rappresenta presumibilmente l'intervallo di tempo necessario alla formazione del cono di crescita, tramite la riorganizzazione del citoscheletro all'estremità distale dell'assone leso. Il periodo di latenza è variabile; ad esempio, i fenomeni di rigenerazione negli assoni motori di ratto iniziano entro i due giorni successivi alla lesione. Durante tale intervallo di tempo i neurofilamenti e i microtubuli si accumulano nel punto di sezione dell'assone e, come risultato della riorganizzazione di queste proteine del sistema microtrabecolare, si costituisce il cono di crescita. Quest'ultimo non contiene di per sé microtubuli e neurofilamenti, ma tali strutture rimangono alla base del cono di crescita e lo seguono nel suo avanzamento. La fase di accrescimento è così contrassegnata dall'allungamento dell'assone. Una volta raggiunto l'organo bersaglio e ultimata la sua reinnervazione, inizia il periodo di maturazione, durante il quale l'assone neoformato, dapprima molto sottile, riacquista il suo diametro normale e completa la sua rimielinizzazione.
Alcune interessanti indagini sperimentali sono state rivolte allo studio dell'accrescimento del nervo in seguito a lesioni apportate in tempi successivi. E stato così dimostrato che l'accrescimento del nervo conseguente a una seconda lesione risulta accelerato rispetto alla rigenerazione verificatasi in seguito al primo evento lesivo (v. McQuarrie e Grafstein, 1973), come se la crescita dell'assone venisse stimolata dall'essere già stato sottoposto in precedenza a una lesione. L'accelerazione dell'accrescimento dell'assone è cospicua se la prima lesione precede la seconda di due settimane, ma l'effetto di stimolo viene esercitato anche se l'intervallo di tempo è di soli due giorni. L'interpretazione di questo fenomeno rimane tuttora nel campo delle ipotesi; sono stati chiamati in causa fattori estrinseci, quale l'ambiente che la prima lesione crea intorno all'assone rigenerato. E tuttavia opinione comune e accreditata che il fenomeno non sia rapportabile a fattori esterni alla cellula nervosa, bensì a un effetto stimolante che il primo evento lesivo esercita sul metabolismo neuronale. Questo stimolo non sarebbe correlabile all'evento lesivo di per sé, ma al fatto che il neurone abbia già sopperito in precedenza a fenomeni di rigenerazione. Ciò è comprovato, fra l'altro, dall'osservazione sperimentale che un fenomeno di accelerazione della crescita assonale si verifica anche in fibre che, pur non essendo state precedentemente lesionate, sono andate incontro a modificazioni plastiche. Ad esempio, è stato più volte dimostrato che l'assone può far fronte alla denervazione delle masse muscolari o di un altro territorio periferico mediante un meccanismo definito sprouting collaterale, che consiste in una proliferazione dei terminali dell'assone, finalizzata a reinnervare i campi deafferentati. Se una fibra nella quale si sono verificati fenomeni di sprouting viene successivamente sezionata, essa rigenera con una velocità maggiore dei normali tassi di accrescimento assonale (v. Sparrow e Grafstein, 1983).
Come è stato precedentemente accennato, i fenomeni di accrescimento e rigenerazione dell'assone, essenzialmente basati sull'apporto delle sue componenti strutturali, sono principalmente correlati alle dinamiche delle fasi lente del trasporto assonale. Tale evidenza sperimentale lascia tuttavia aperti molti quesiti, due dei quali presentano un particolare interesse pratico e speculativo.
Il primo quesito riguarda la partecipazione delle componenti del trasporto anterogrado rapido ai fenomeni di rigenerazione. Infatti, pur non essendo le velocità di migrazione delle componenti rapide specificamente rapportabili alla velocità dei fenomeni di rigenerazione, è comunque presumibile che i due fenomeni siano in qualche modo correlati. Il rapporto fra trasporto rapido e rigenerazione assonale non è stato in realtà finora chiarito, ma è stato osservato che nella fase di accrescimento si verificano notevoli rimaneggiamenti del trasporto rapido (v. Frizell e Sjöstrand, 1981). Durante l'accrescimento del nervo aumenta infatti il trasporto di alcune sostanze, quali i precursori di membrana (ad esempio le glicoproteine), mentre altre componenti del trasporto coinvolte in funzioni molto specializzate, quali gli enzimi del metabolismo dei neurotrasmettitori, mostrano una diminuzione del trasporto. Tali variazioni, che riguardano la quantità dei materiali ma non la loro velocità di migrazione, stanno a indicare che i neuroni, durante la rigenerazione, ‛danno la priorità' alle componenti delle proteine strutturali. Il trasporto di enzimi correlati al metabolismo dei neurotrasmettitori si normalizza poi durante la fase di maturazione, una volta, quindi, che sia avvenuta la reinnervazione dell'organo bersaglio.
Il secondo quesito riguarda la provenienza dei materiali del citoscheletro direttamente coinvolti nell'allungamento dell'assone. Infatti, pur essendo questi ovviamente forniti, in origine, dal corpo cellulare, è da presumere che un carico proteico appositamente destinato alla rigenerazione non potrebbe essere immediatamente disponibile a grande distanza dal pericario, dato che verrebbe affidato alle fasi lente del trasporto. Ad esempio, nel caso della sezione di assoni motori di ratto alla distanza di 80 mm dal corpo cellulare, un carico di proteine specificamente finalizzate alla rigenerazione giungerebbe a destinazione più di un mese dopo la lesione, mentre invece la rigenerazione ha inizio, come è stato precedentemente specificato, entro i due giorni immediatamente successivi. Evidentemente, quindi, i materiali necessari al citoscheletro durante l'allungamento dell'assone vengono tratti proprio dal pool di proteine dei neurotubuli e dei microfilamenti che si accumulano alla base del cono di crescita (v. Veraa e Grafstein, 1981). Ciò porta all'interessante conclusione che il neurone è sempre pronto a fronteggiare l'eventualità di una lesione assonale e a produrre i materiali necessari a sostenere la propria rigenerazione. Sulla base di tali dati è quindi possibile ipotizzare che gli assoni possiedano capacità rigenerative intrinseche anche nel sistema nervoso centrale, e che sia quiridi l'ambiente circostante a ostacolare la possibile rigenerazione dei neuroni centrali.
8. Il trasporto assonale in patologia
Il trasporto assonale può andare incontro ad alterazioni in una grande varietà di condizioni patologiche, sia cliniche che sperimentali. I modelli sperimentali di alterazioni del trasporto assonale hanno fornito preziosi strumenti di indagine dei meccanismi fisiologici che regolano il trasporto e dei meccanismi patologici che con esso interferiscono. Per quanto riguarda i quadri clinici di neuropatia, molti dati su alterazioni della velocità di trasporto sono stati ottenuti misurando l'accumulo di enzimi (in particolare la dopammina-β-idrossilasi e l'acetilcolinesterasi) su prelievi di nervo surale. Mediante queste e altre metodiche è stato osservato che la velocità del trasporto assonale è ridotta in molte condizioni patologiche, quali l'iperuricemia, la malattia di Déjerine-Sottas e, soprattutto, la neuropatia diabetica. Quest'ultima è stata anche oggetto di indagini sperimentali, sia in ceppi mutanti diabetici che nel diabete sperimentale indotto, ad esempio, dalla somministrazione di streptozotocina. Sono state così dimostrate notevoli alterazioni delle fasi lente del trasporto assonale in corso di neuropatia diabetica, e sono state segnalate anche alterazioni delle componenti rapide.
Il modello clinico e sperimentale più studiato di patologia del trasporto assonale è rappresentato dalle neuropatie tossiche e, in particolare, dalle condizioni patologiche caratterizzate dalla cosiddetta ‛degenerazione neurofibrillare' e dalla presenza, quindi, di conglomerati patologici di neurofibrille (neurofibrillary tangles degli autori anglosassoni). Tale processo degenerativo consiste nell'accumulo di neurofilamenti e può verificarsi in vari distretti neuronali. I neurofilamenti possono infatti accumularsi nel corpo neuronale, oppure nelle porzioni prossimali o distali dell'assone. Un esempio di degenerazione neurofibrillare localizzata nel corpo neuronale è fornito dalle alterazioni presenti in neuroni della corteccia cerebrale, e in alcune strutture sottocorticali, nelle demenze senili e presenili tipo Alzheimer. Nell'assone l'accumulo di neurofilamenti dà luogo a caratteristici rigonfiamenti, che costituiscono l'alterazione morfologica comune alle cosiddette neuropatie giganto-assonali (v. Griffin e altri, 1983).
L'accumulo di neurofilamenti può essere provocato dalla somministrazione di molti tipi di neurotossine chimiche. L'intossicazione di β, β′-imminodipropionitrile (IDPN) è uno dei modelli sperimentali più studiati e produce un quadro caratteristico di patologia assonale, nel quale i microtubuli sono segregati dai neurofilamenti. Il trasporto assonale di questi ultimi viene così a essere profondamente alterato, e il blocco dei loro meccanismi di trasporto causa un accumulo di neurofilamenti nelle porzioni prossimali dell'assone. L'IDPN agisce quindi selettivamente sulle componenti del trasporto, mentre il trasporto assonale rapido rimane per lo più inalterato. Tale sostanza, inoltre, alterando il movimento prossimo-distale delle proteine dei neurofilamenti, produce non solo rigonfiamenti prossimali, ma anche un'atrofia dei segmenti distali dell'assone.
Simile all'effetto dell'TDPN è quello esercitato dal 2,5esandione, una sostanza tossica implicata nelle polineuropatie da collanti, che produce anch'essa un rigonfiamento giganto-assonale dovuto all'accumulo di neurofilamenti. Le alterazioni causate da quest'ultima tossina sono essenzialmente localizzate nei segmenti distali dell'assone. Anche l'acrilammide esercita un effetto di blocco sul trasporto lento, causando una riduzione delle proteine trasportate, e agisce prevalentemente sui segmenti distali dell'assone. Tuttavia, a differenza dell'IDPN, l'acrilammide produce anche alterazioni delle componenti rapide del trasporto. Infine, l'intossicazione da alluminio, che produce anch'essa un accumulo di neurofilamenti, agisce sia a livello del corpo cellulare che delle parti prossimali dell'assone e fornisce quindi un modello sperimentale di alterazione patologica simile al quadro clinico della sclerosi laterale amiotrofica.
Un curioso fenomeno, del tutto diverso dalle situazioni precedentemente elencate, ma che può anch'esso fornire un modello sperimentale di malattia del motoneurone, è rappresentato dal trasporto assonale retrogrado di una sostanza neurotossica, la doxorubicina o adriamicina. Tale composto, un antibiotico citotossico che ha la caratteristica di essere fluorescente, è dotato di una forte capacità di legarsi agli acidi nucleici e al DNA in particolare, ed è usato nel corso di terapie antineoplastiche (v. chemioterapia antineoplastica; v. sangue: Leucemie). È stato recentemente dimostrato (v. Bigotte e Olsson, 1982; v. Koda e Van der Kooy, 1983) che tale sostanza viene assunta dai terminali assonici e trasportata in via retrograda ai corpi cellulari, dove può essere visualizzata mediante osservazione al microscopio in fluorescenza. La doxorubicina produce infatti una caratteristica fluorescenza di colore rosso-arancione, e ha una localizzazione, all'interno del nucleo, simile a quella della cromatina nucleare. Il trasporto assonale retrogrado di tale sostanza è stato dimostrato sia nel sistema nervoso periferico che nel sistema nervoso centrale. La doxorubicina apparentemente non agisce in modo primario sui meccanismi del trasporto assonale, ma, una volta raggiunto il corpo cellulare, produce dopo un certo intervallo di tempo un effetto citotossico specifico, causando fenomeni di cromatolisi e morte cellulare. Oltre che per somministrazione locale, cioè in aree di innervazione centrale o periferica, questo antibiotico citotossico può raggiungere per via ematica i terminali assonici in territori situati al di fuori della barriera ematoencefalica, quali ad esempio la neuroipofisi e l'area postrema (v. Bigotte e Olsson, 1983), dove produce alterazioni caratteristiche del nucleo e del nucleolo dei corpi cellulari e degenerazioni assonali. Poiché la diffusione intraneuronale della sostanza è specificamente correlata al suo trasporto assonale retrogrado, ne è stato proposto l'uso in tecniche neuroanatomiche (v. cap. 9, § b), ma le sue proprietà neurotossiche (un cosiddetto trasporto assonale ‛suicida') le conferiscono un particolare interesse per future applicazioni in modelli sperimentali di patologia neuronale selettivamente localizzata in determinati circuiti.
9. Il trasporto assonale negli studi di neuroanatomia sperimentale
È stato ampiamente dimostrato, nei capitoli precedenti, come la scoperta del trasferimento di sostanze lungo gli assoni delle cellule nervose e lo studio delle caratteristiche di tale fenomeno abbiano largamente contribuito alle attuali conoscenze sulla struttura dell'assone e sulle sue interazioni con l'ambiente circostante e, inoltre, all'identificazione del materiale trasportato e delle dinamiche che ne regolano il trasporto. Deve essere tuttavia sottolineato che, come è stato brevemente accennato nel cap. 1, le metodiche basate sul trasporto assonale hanno portato un notevolissimo contributo anche agli studi di neuroanatomia sperimentale.
Fino a qualche tempo fa lo studio delle connessioni del sistema nervoso si avvaleva di metodiche di colorazione diretta del corpo cellulare e dei suoi prolungamenti, quale il metodo di Golgi, e delle tecniche di degenerazione. Queste ultime sono basate sull'identificazione delle modificazioni patologiche del corpo cellulare e dell'assone in seguito a lesioni sperimentali. L'identificazione della sofferenza retrograda dei corpi cellulari, conseguente a interruzione dell'assone, ha rappresentato, per lungo tempo, uno strumento di studio dei corpi cellulari di origine delle proiezioni afferenti alle strutture in cui veniva effettuata la lesione. D'altra parte, i metodi di colorazione argentica delle fibre, selettiva per la degenerazione anterograda degli assoni a valle della lesione, sono stati ampiamente utilizzati per studiare la distribuzione delle proiezioni efferenti dalla zona lesa. Le metodiche sopra elencate presentano tuttavia una serie di inconvenienti (quale, ad esempio, l'interruzione di fibre ‛di passaggio' nell'area in cui viene praticata la lesione sperimentale) che possono inficiare, in molti casi, la validità delle osservazioni sperimentali. Inoltre la degenerazione retrograda, pur avendo fornito uno strumento prezioso per tracciare le prime ‛mappe' di distribuzione dei corpi cellulari di origine di vari sistemi, non è una metodica molto sensibile. È infatti assai problematica, in un preparato istologico, l'identificazione di un piccolo numero di corpi cellulari in degenerazione, come pure della degenerazione di corpi cellulari sparsi, distribuiti lungo un ampio territorio.
Verso la fine degli anni sessanta e, soprattutto, agli inizi degli anni settanta alcuni studiosi, sulla base dei numerosi dati che concordemente indicavano la precisione e la selettività di tutte le dinamiche correlate al trasporto assonale, hanno proposto di utilizzare quest'ultimo meccanismo nello studio dell'organizzazione dell'origine e della distribuzione dei circuiti neuronali. L'introduzione delle metodiche basate sul trasporto di molecole esogene, che potessero essere utilizzate come ‛traccianti' nelle ricerche di neuroanatomia sperimentale, ha profondamente innovato gli studi in tale settore della ricerca neurobiologica. È prerequisito di tali metodiche che i traccianti possano essere efficacemente identificati all'interno del sistema nervoso. Per gli studi in via anterograda è stato così inizialmente proposto l'uso di sostanze marcate con isotopo radioattivo e, in particolare, degli amminoacidi (v. Lasek e altri, 1968; v. Cowan e altri, 1972). Per quanto riguarda gli studi in via retrograda, un apporto decisivo è stato fornito dalle ricerche di Kristensson e Olsson (v., Uptake..., 1971). Infatti, nel corso di esperimenti finalizzati alla dimostrazione del trasporto retrogrado di proteine, tali autori hanno utilizzato l'HRP (v. oltre), identificandone quindi per primi il movimento disto-prossimale e la localizzazione nel corpo cellulare, mediante una semplice metodica istochimica. Sulla base di queste osservazioni è stato quindi proposto (v. LaVail e LaVail, 1972) che molecole esogene, e l'HRP in particolare, potessero essere utilizzate in studi neuroanatomici in via retrograda.
Tutte le metodiche introdotte in neuroanatomia sperimentale sono basate sulla somministrazione di sostanze esogene nelle aree del sistema nervoso centrale o periferico di cui si vogliono studiare le afferenze e le efferenze. La via più comune di somministrazione è rappresentata dall'iniezione, mediante microsiringhe, di soluzioni contenenti la sostanza. Dopo un periodo di sopravvivenza, necessario perché il trasporto assonale abbia luogo, si procede al sacrificio dell'animale mediante procedure di fissazione adeguate. I successivi procedimenti istologici sono rappresentati dall'autoradiografia, nel caso in cui i traccianti siano legati a isotopi radioattivi, da procedimenti istochimici (come nel caso dell'HRP), o dall'osservazione in fluorescenza, nel caso in cui i traccianti siano essi stessi fluorescenti o siano legati a molecole fluorescenti. Sono state inoltre recentemente proposte varie metodiche di rivelazione basate su tecniche immunoistochimiche e, quindi, sul trattamento delle sezioni istologiche mediante anticorpi specifici antitracciante. Ovviamente, per l'applicazione di tali metodiche, che avrebbero il vantaggio di una grande sensibilità, soprattutto per quanto riguarda la visualizzazione morfologica dei neuroni marcati, è necessario che il tracciante sia dotato di proprietà antigeniche.
Il principio che è alla base di tutte le metodiche neuroanatomiche di trasporto assonale retrogrado è che i traccianti, veicolati negli assoni dall'area di iniezione, vadano a localizzarsi selettivamente e specificamente nei corpi cellulari di origine. Se ne deduce quindi che i corpi cellulari marcati dal tracciante inviano proiezioni all'area in cui quest'ultimo è stato somministrato. D'altro canto, nelle metodiche in via anterograda l'origine delle fibre e/o dei terminali assonici marcati viene ascritta ai corpi cellulari presenti nell'area di iniezione. Ovviamente, l'uso dei traccianti implica che la loro distribuzione sia selettivamente legata alle dinamiche del trasporto assonale e non a meccanismi di altro genere, come una diffusione ematica e quindi aspecifica. Poiché il trasporto assonale è un fenomeno comune a tutte le cellule nervose, tali metodiche si prestano a uno studio ubiquitario dei circuiti neuronali, nel sistema nervoso centrale e periferico. Alcune particolari metodiche di trasporto assonale, quali ad esempio l'assunzione e il trasporto retrogrado di possibili neurotrasmettitori, marcati con isotopi (v. Cuénod e altri, 1982; v. § b), sono invece fondate sul principio che l'assunzione e il trasporto di determinate sostanze siano specifici solo per i circuiti in cui esse sono fisiologicamente implicate, e che esse consentano quindi l'evidenziazione selettiva delle vie afferenti nelle quali le molecole agirebbero come neurotrasmettitori.
a) Metodi anterogradi
La metodica neuroanatomica più diffusa di trasporto assonale anterogrado si basa sull'osservazione che taluni amminoacidi, iniettati in prossimità di un pericario neuronale, vengono assunti dalla cellula, incorporati nella sintesi proteica, e quindi in macromolecole, e trasportati come tali lungo l'assone, sia nelle fasi lente che in quelle rapide del trasporto, per accumularsi finalmente nei terminali. Il metodo di marcatura degli amminoacidi consiste nella loro coniugazione con isotopi, e la rivelazione del tracciante è quindi basata sulla procedura autoradiografica. Il trizio è l'isotopo usato più comunemente.
Dalla prima segnalazione di Lasek e altri (v., 1968) sulla distribuzione centrale di fibre sensitive primarie nel midollo spinale, seguita dal classico studio di Cowan e altri (v., 1972) su vari circuiti del sistema nervoso centrale, l'uso degli amminoacidi triziati si e conquistato una larga popolarità per la sua affidabilità ed efficacia. Il vantaggio principale dell'uso di amminoacidi sta nel fatto che essi possono essere assunti e incorporati in macromolecole proteiche solo nel corpo cellulare, dove cioe avvengono i fenomeni di sintesi proteica, mentre altri traccianti, quale ad esempio l'HRP, possono essere assunti da ogni distretto del neurone. Il trasporto assonale anterogrado di amminoacidi consente quindi di ovviare al problema dell'assunzione da parte di fibre di passaggio nell'area di iniezione, e costituisce una tecnica elettiva per una metodica di marcatura anterograda senza contemporaneo trasporto retrogrado.
La 3H-leucina, un amminoacido essenziale, si è fin dall'inizio imposta come precursore amminoacidico di elezione negli studi neuroanatomici. Anche la 3H-prolina è stata però ampiamente utilizzata, e un altro amminoacido, la 3H-lisina, è stato talvolta aggiunto alle soluzioni iniettate (v. tab. III).
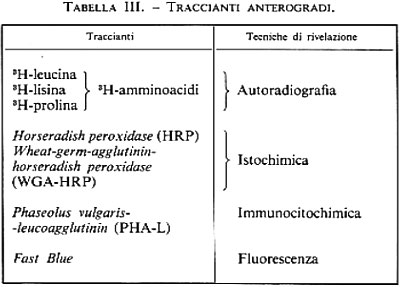
La metodica comporta alcuni delicati aspetti di carattere interpretativo, quale la valutazione dell'estensione dell'area di iniezione, che dipende notevolmente dai tempi di sopravvivenza dell'animale e di esposizione autoradiografica. Tuttavia molteplici dati della letteratura indicano che l'area di massima assunzione del tracciante è rappresentata dalla zona centrale, caratterizzata da neuroni intensamente marcati, che rimane relativamente costante nel tempo. Un secondo ma non meno importante aspetto interpretativo, anch'esso dipendente dai tempi di sopravvivenza e di esposizione, riguarda la marcatura dei terminali assonici. Manipolando i tempi di sopravvivenza si possono infatti mettere in evidenza la fase veloce e quella lenta del trasporto assonale anterogrado di amminoacidi triziati. Si può quindi aumentare l'intensità della marcatura delle fibre, con tempi di sopravvivenza relativamente prolungati, o dei terminali dell'assone (evidenziabili, dati i meccanismi del trasporto rapido, già con tempi di sopravvivenza brevi). Ne deriva che tali parametri temporali hanno una grande importanza nella rilevazione qualitativa e quantitativa dei dati sperimentali. È d'altra parte importante sottolineare che il trasporto assonale anterogrado di amminoacidi triziati ha subito, nel corso degli anni, minori modificazioni metodologiche di altre tecniche neuroanatomiche, quale l'HRP, e ciò consente quindi l'applicazione di un protocollo sperimentale ben collaudato e largamente standardizzato. È importante sottolineare inoltre che il trasporto anterogrado di amminoacidi triziati è l'unico metodo nel quale sia possibile ovviare all'inconveniente dell'assunzione di tracciante da parte di fibre di passaggio.
Nello studio di circuiti monosinaptici è ovviamente indesiderabile che si verifichi un trasferimento transneuronale del tracciante, poichè ciò potrebbe confondere l'interpretazione dei risultati ottenuti. In realtà, in condizioni normali, cioè con le quantità che vengono comunemente iniettate, il passaggio transneuronale di amminoacidi triziati è quantitativamente irrilevante e non ostacola quindi il rilevamento di dati concernenti i sistemi monosinaptici. Tuttavia, con grandi quantità di tracciante radioattivo, tale trasporto transneuronale può essere proporzionalmente aumentato, fino a diventare ben visualizzabile, ed essere utilizzato quindi per lo studio di vie multisinaptiche. E questo il protocollo adottato nello studio di vari sistemi neuronali. In particolare, sono stati ottenuti dei risultati particolarmente brillanti nello studio delle vie ottiche. Infatti, dopo iniezione intraoculare di una grande quantità di tracciante radioattivo è stato possibile visualizzare il campo terminale delle fibre del tratto ottico che, dopo una sinapsi nel talamo, raggiungono la corteccia visiva tramite il sistema talamo-corticale. È stato così dimostrato il correlato morfologico dei dati funzionali relativi alle colonne di dominanza oculare (v. Wiesel e altri, 1974) e quindi l'organizzazione modulare della corteccia visiva, poiché è stato visualizzato che i campi terminali relativi a un occhio sono organizzati in colonne fra loro segregate (v. anche visione).
I continui progressi della tecnica dell'HRP, sia nella evidenziazione istochimica del trasporto (v. oltre) che nella sua efficacia, hanno fatto si che l'uso di tale enzima si sia affiancato in questi ultimi anni a quello degli amminoacidi triziati come valido mezzo di indagine in via anterograda. La fortuna e la diffusione dell'HRP come tracciante anterogrado vanno ricercate soprattutto nella semplicità del metodo, che non richiede le lunghe e laboriose manipolazioni della tecnica autoradiografica. Inoltre, l'uso dell'HRP consente di rilevare i risultati relativi sia al trasporto anterogrado dell'enzima, e quindi alle efferenze di una data zona, sia al suo trasporto retrogrado, e quindi alle afferenze alla stessa area di iniezione. L'introduzione dell'HRP combinata a lectine (v. oltre) ha ulteriormente incentivato l'uso dell'HRP come tracciante anterogrado (v. tab. III), poiché ha fornito una metodica estremamente sensibile.
Alcune lectine stesse, come la Phaseolus vulgaris-leucoagglutinin (PHA-L), sono state ultimamente proposte come traccianti anterogradi di grande sensibilità, soprattutto per quanto concerne l'evidenziazione morfologica dei neuroni marcati. La marcatura mediante PHA-L fornirebbe infatti risultati comparabili a quelli del metodo di Golgi. La rivelazione della PHA-L è basata su metodiche immunocitochimiche (v. tab. III).
Merita infine segnalare l'osservazione sperimentale che la sostanza fluorescente Fast Blue, generalmente utilizzata come tracciante retrogrado (v. oltre), marca anche efficacemente fibre e terminali in via anterograda (v. Rosina, 1982). Quantunque sia presto per giudicarne l'effettiva validità, è interessante segnalare che tale metodica presenta notevoli possibilità applicative. É infatti una procedura di grande semplicità tecnica, soprattutto per quanto riguarda le prospettive di efficaci combinazioni con altri traccianti fluorescenti in tecniche di marcatura multipla retrograda e anterograda.
b) Metodi retrogradi
L'uso della macromolecola HRP rappresenta il più diffuso metodo neuroanatomico basato sul trasporto assonale retrogrado. L'HRP è stato il primo tracciante retrogrado introdotto in letteratura per lo studio del sistema nervoso centrale (v. LaVail e LaVail, 1972) e, data la facile applicabilità del protocollo sperimentale, i brevi periodi di sopravvivenza necessari (variabili in genere da uno a tre giorni) e la rapidità della metodica istochimica, la tecnica che se ne serve ha conquistato un'immediata e vastissima popolarità.
Subito dopo l'introduzione dell'HRP come tracciante nelle ricerche di neuroanatomia sperimentale sono iniziati molteplici studi volti a incrementarne l'efficacia e la sensibilità. Inoltre, si è intrapresa una serie di studi destinati alla ricerca di un secondo tracciante che potesse essere combinato con l'HRP in metodiche di marcatura retrograda multipla (v. oltre). Come precedentemente accennato (v. cap. 4), il metodo di rivelazione dell'HRP è basato su una reazione enzimatica, catalizzata dall'HRP stessa, in cui, in presenza di H2O2 e di un cromogeno come substrato, quest'ultimo precipita, svelando la presenza dell'enzima nel corpo cellulare e/o in altri distretti del neurone. Con i substrati più comunemente adottati il cromogeno precipita sotto forma di granuli scuri. Sulla base della reazione istochimica già largamente utilizzata in letteratura (v. Graham e Karnovsky, 1966), è stata in un primo tempo utilizzata come substrato la diamminobenzidina. Successivamente, nel tentativo di incrementare la sensibilità del metodo, sono stati proposti molti altri substrati, il più diffuso ed efficace dei quali è la tetrametilbenzidina (v. Mesulam, 1978). Quest'ultima sostanza è particolarmente efficace anche nel rivelare il trasporto assonale anterogrado (e quindi la marcatura di fibre e terminali) dell'HRP. L'introduzione di modifiche dell'originaria reazione istochimica, delle metodiche di perfusione, e di molti altri dettagli del protocollo sperimentale dell'HRP, hanno fatto sì che il metodo acquistasse una notevole sensibilità e consentisse un grande progresso della ricerca neuroanatomica.
Pur essendo state tutte le fasi del protocollo sperimentale dell'HRP sottoposte a un rigoroso controllo, rimane il problema della possibilità di assunzione del tracciante da parte di vari distretti del neurone e, in particolare, da parte di fibre danneggiate nel corso della procedura sperimentale. Il problema di fibre di passaggio nell'area di iniezione rimane quindi sempre da considerare nella valutazione dei dati sperimentali.
Per quanto riguarda il secondo problema, cioè la ricerca di un altro tracciante da combinare con l'HRP in metodiche di marcatura retrograda multipla, molti studi sono stati indirizzati alla messa a punto di molecole modificate della stessa HRP, che potessero però essere visualizzate con tecniche diverse da quella istochimica. È infatti essenziale, nell'individuare la marcatura di due traccianti in uno stesso corpo cellulare, che essi siano ben distinguibili l'uno dall'altro (v. oltre). A tale scopo è stato proposto l'uso di HRP coniugata con isotopo radioattivo (3H-HRP), oppure inattivata dal punto di vista enzimatico e quindi coniugata con isotopo (v. Hayes e Rustioni, 1979). Tutte le molecole di HRP coniugate con trizio possono essere ovviamente evidenziate mediante procedura autoradiografica (v. tab. IV). Infine è stato a più riprese segnalato, nella letteratura, che il trasporto retrogrado di altre macromolecole, le lectine, è molto efficace. Al fine quindi di incrementare ulteriormente l'efficacia del trasporto retrogrado dell'HRP, come anche del suo trasporto anterogrado, è stato proposto di coniugare l'enzima con l'agglutinina di germe di grano (wheat-germ-agglutinin, WGA) (v. tab. IV).

Sulla base di tutte queste modifiche, l'HRP si pone attualmente come tracciante versatile e affidabile e, nelle sue varie forme coniugate, come possibile strumento in molteplici metodiche neuroanatorniche. È inoltre da sottolineare che l'HRP è, a tutt'oggi, il tracciante di elezione per gli studi che contemplano un'indagine ultrastrutturale al microscopio elettronico.
Il metodo che utilizza traccianti fluorescenti rappresenta uno dei più recenti capitoli delle tecniche neuroanatomiche in via retrograda (v. fig. 11). Uno dei principali vantaggi è costituito dalla semplicità della tecnica di rivelazione, che comporta solo l'osservazione in fluorescenza e non prevede, quindi, alcun tipo di manipolazione istologica. Per quanto riguarda la loro utilizzazione nello studio delle connessioni interneuronali, le sostanze fluorescenti trovano la loro principale applicazione in metodiche di marcatura retrograda multipla. I traccianti fluorescenti sono stati tuttavia largamente utilizzati anche in altre strategie sperimentali. Infatti, al contrario dell'HRP, che viene catabolizzata dopo un certo periodo di tempo, molte delle sostanze fluorescenti non solo non decadono nel tempo, ma continuano anzi ad accumularsi, almeno per molte settimane, nei corpi cellulari marcati in via retrograda. Tale proprietà ne consente l'uso, impossibile con l'HRP, per applicazioni che prevedono un lungo tempo di sopravvivenza, o nello studio di sistemi la cui organizzazione varia secondo una determinata sequenza temporale, come avviene, ad esempio, nel corso dello sviluppo o di fenomeni rigenerativi.
L'identificazione di traccianti fluorescenti ha preso l'avvio dalla ricerca di sostanze da combinare con l'HRP (v. Kuypers e altri, 1977) e ha poi seguito un'evoluzione diversa e autonoma. Una delle maggiori sorprese degli studi attuati in tale campo è stata la scoperta del grande numero di sostanze fluorescenti esogene che, iniettate nel sistema nervoso centrale, vengono assunte e trasportate lungo l'assone ai corpi cellulari (v. Bentivoglio e altri, 1979 e 1980; v. anche Kuypers e altri, 1979). I traccianti fluorescenti più comunemente usati e le loro principali caratteristiche sono elencati nella tab. IV. La maggior parte di essi è costituita da sostanze a basso peso molecolare che si legano agli acidi nucleici. I meccanismi che presiedono al trasporto assonale di tali sostanze sono tuttora del tutto sconosciuti. Dagli studi neuroanatomici finora effettuati in vari sistemi sembra che i traccianti fluorescenti vengano convogliati secondo le dinamiche del trasporto retrogrado rapido. Tuttavia il fatto che la loro visualizzazione (e cioè l'intensità della fluorescenza dei corpi cellulari marcati) venga favorita da tempi di sopravvivenza assai più prolungati di quelli necessari per la visualizzazione dell'HRP suggerisce che anche delle componenti lente possano essere implicate nell'accumulo disto-prossimale delle sostanze fluorescenti.
Un'importante caratteristica di tali traccianti è che essi sono distinguibili l'uno dall'altro sulla base del loro diverso colore e/o della loro diversa localizzazione subcellulare. Un delicato problema metodologico relativo al loro impiego è però rappresentato dal fatto che alcuni di essi possono fuoriuscire dai neuroni marcati dopo periodi di sopravvivenza relativamente brevi, ponendo quindi problemi di marcatura transneuronale. Alcuni accorgimenti metodologici sono necessari per ovviare a tale inconveniente. È stato segnalato inoltre che alcuni di essi non sono trasportati efficacemente nel sistema nervoso periferico, mentre altri, in seguito a trasporto lungo i nervi periferici, marcano i corpi cellulari di origine con una localizzazione subcellulare diversa da quella osservata nel sistema nervoso centrale (ad esempio citoplasmatica invece che nucleare).
Le metodiche di marcatura retrograda multipla sono basate sull'evidenziazione di traccianti diversi in uno stesso corpo neuronale in seguito a trasporto retrogrado in diverse branche dell'assone. Tali tecniche sono state messe a punto principalmente per lo studio delle proiezioni collaterali, dirette da una cellula a diverse aree di terminazione mediante biforcazioni dell'assone. Fino a qualche anno fa lo studio morfologico di tale problema era essenzialmente basato sull'evidenziazione morfologica diretta delle biforcazioni assonali, con metodi di colorazione diretta del corpo cellulare e delle sue ramificazioni, quali il metodo di Golgi o, più recentemente, l'iniezione intracellulare di sostanze coloranti. Tali metodiche hanno però l'evidente limitazione di non poter rivelare con precisione l'organizzazione topografica e quantitativa e le aree di terminazione delle popolazioni di origine di assoni biforcati. I metodi di marcatura retrograda multipla sono invece basati su un principio teoricamente semplice: se un corpo cellulare si connette con aree diverse del nevrasse mediante biforcazioni del suo assone, due (o più) traccianti iniettati in uno stesso animale, nelle diverse aree di terminazione delle collaterali assoniche, possono essere da queste ultime assunti e trasportati in via retrograda per localizzarsi entro lo stesso corpo cellulare. Per converso, quindi, la presenza di due (o più) traccianti in uno stesso corpo neuronale implica che quest'ultimo proietta, mediante branche collaterali del suo assone, ai siti di iniezione dei traccianti.
La realizzazione tecnica di metodiche che consentissero uno studio di questo tipo ha tuttavia richiesto molti tentativi e un grosso impegno di ricerca, poichè non si è rivelato affatto semplice reperire due traccianti che fossero ‛compatibili' dal punto di vista metodologico e ben distinguibili l'uno dall'altro nelle loro metodiche di rivelazione.
Le tecniche attualmente più diffuse contemplano combinazioni di istochimica dell'HRP e autoradiografia e, soprattutto, l'uso di traccianti fluorescenti. Questi ultimi, infatti, ben si adattano a metodi di marcatura multipla, poiché le combinazioni di traccianti che producono un diverso colore nel corpo cellulare hanno offerto una soluzione tecnica relativamente semplice ai problemi di marcatura retrograda multipla. Inoltre, poiché alla possibilità di distinguere colori diversi si aggiunge anche quella di distinguere la diversa localizzazione subcellulare dei traccianti, il trasporto assonale retrogrado di sostanze fluorescenti può contemplare anche l'uso di più di due traccianti. Su tale base sono state infatti messe a punto metodiche di triplice marcatura retrograda e, teoricamente, il numero dei traccianti compatibili potrebbe aumentare ulteriormente.
Deve essere inoltre ricordato che i metodi di marcatura retrograda multipla, pur essendo stati messi a punto per lo studio delle proiezioni collaterali, hanno in realtà offerto anche un validissimo strumento di indagine delle interrelazioni reciproche fra diverse popolazioni cellulari, come la loro eventuale segregazione o sovrapposizione.
L'identificazione di singoli neuroni che controllano mediante ramificazioni dell'assone aree anche molto distanti del nevrasse e la possibilità di studiare simultaneamente l'organizzazione di più popolazioni neuronali efferenti hanno aperto la strada a un nuovo capitolo della neuroanatomia, consentendo anche un rilevante approfondimento delle interrelazioni funzionali fra diversi gruppi cellulari.
La combinazione dei metodi di trasporto assonale retrogrado con tecniche che consentono l'identificazione dei neurotrasmettitori implicati nei circuiti in esame ha costituito un ulteriore recente sviluppo della neuroanatomia e della neurochimica. Molti studi, infatti, sono stati recentemente rivolti alla messa a punto di metodiche che consentissero lo studio dell'organizzazione delle connessioni interneuronali, fornendo, al contempo, delle indicazioni sui neurotrasmettitori in esse implicati. Le metodiche finora proposte si basano essenzialmente su due tipi di approccio sperimentale. Il primo è rappresentato dal trasporto retrogrado selettivo di neurotrasmettitori somministrati per via esogena. Il secondo indirizzo sperimentale si basa invece sulla combinazione di metodiche di trasporto retrogrado non selettivo (quali quelle dell'HRP e dei traccianti fluorescenti) con tecniche istochimiche o immunocitochimiche.
Per quanto riguarda il primo punto, è stato verificato sperimentalmente (v. Cuénod e altri, 1982) che alcuni neurotrasmettitori triziati, iniettati nel sistema nervoso centrale, vengono assunti e trasportati selettivamente nei circuiti in cui agiscono fisiologicamente come neurotrasmettitori. Tali studi sono stati condotti in varie specie animali (piccione, ratto, gatto) e hanno finora riguardato la glicina, il GABA, l'aspartato e/o il glutammato, l'acetilcolina, la dopammina e la serotonina. Nonostante che nell'uso di alcune di queste molecole si siano verificati ‛falsi negativi' o ‛falsi positivi', tale tecnica ha per ora fornito risultati molto interessanti, quale, ad esempio, il dato che molte vie efferenti dalla corteccia cerebrale utilizzano aspartato e/o glutammato come trasmettitore. Molti aspetti del trasporto di neurotrasmettitori esogeni sono tuttora da chiarire. Tuttavia sembra accertato che la selettività del trasporto sia da ascrivere alla presenza di meccanismi di assunzione molto specifici (il cosiddetto high affinity uptake), che costituiscono una delle proprietà caratteristiche delle molecole neurotrasmettitoriali. Probabilmente, inoltre, la selettività risiede anche nei meccanismi di trasporto, poiché tali molecole, generalmente di piccole dimensioni, verrebbero convogliate in senso disto-prossimale legate a organelli (quali vescicole, corpi multivescicolari o lisosomi) e potrebbero quindi avere vettori (i cosiddetti carriers) specifici. Il trasporto retrogrado di neurotrasmettitori si svolge a una velocità pari a 150-200 mm/die.
Del tutto diverso è l'indirizzo sperimentale nel quale i corpi di origine di determinate proiezioni neuronali vengono marcati mediante traccianti aspecifici, mentre gli eventuali trasmettitori (o gli enzimi correlati al loro metabolismo) presenti nel corpo neuronale vengono invece identificati mediante metodiche specifiche. Le difficoltà ditali tecniche risiedono generalmente nella combinazione delle due diverse metodologie (cioè quella per il tracciante e quella per il neurotrasmettitore), che possono contemplare, ad esempio, perfusioni o procedimenti istochimici non compatibili. Inoltre, come nei metodi di marcatura retrograda multipla, è necessario che le due sostanze, in questo caso il tracciante e il neurotrasmettitore o la molecola a esso correlata, siano simultaneamente visualizzabili in uno stesso corpo neuronale.
Per quanto riguarda l'HRP, sono state proposte metodiche combinate con l'istochimica dell'acetilcolinesterasi, come pure metodiche combinate con tecniche immunocitochimiche. L'HRP e la sostanza che svela la presenza del neurotrasmettitore sono in genere distinguibili sulla base del diverso colore dei precipitati delle relative reazioni istochimiche. Nuove prospettive in questo campo sono state introdotte dai traccianti fluorescenti, poiché questi ben si prestano a combinazioni con metodiche di istofluorescenza per le monoammine e di immunofluorescenza. In tali casi la distinzione fra tracciante e neurotrasmettitore, o molecola a esso correlata, si basa sul loro diverso colore all'osservazione in fluorescenza. I traccianti fluorescenti si stanno anche rivelando efficaci nelle combinazioni con metodiche istochimiche. In quest'ultimo caso la doppia marcatura viene rilevata mediante una doppia osservazione del preparato al microscopio, in campo chiaro e in fluorescenza.
Tutti questi approcci sperimentali, ancora in corso di studio, hanno aperto interessanti prospettive sia nelle metodiche di trasporto retrogrado che nello studio della distribuzione dei neurotrasmettitori, e sembrano assai promettenti.
10. Considerazioni conclusive
Da quanto è stato esposto nei vari capitoli risulta chiaro come il trasporto assonale, cioè la dinamica di traslazione delle sostanze che si muovono dalla periferia al pirenoforo, e viceversa, rappresenti oggi una preziosa metodica di indagine della vita normale, cioè della fisiologia, e patologica, cioè della fisiopatologia, del neurone.
La scoperta del trasporto rapido e di quello lento di molecole e l'identificazione di compartimenti distinti dell'assone a essi correlati, il reticolo endoplasmico liscio e l'apparato neurofilamentoso, hanno consentito di separare due funzioni distinte dell'assone. Una di esse è infatti legata precipuamente al turnover metabolico delle membrane, e quindi all'attività specifica della singola cellula. L'altra è essenzialmente correlata all'omeostasi neuronale, e quindi al mantenimento della struttura interna del neurone, e in particolare del citoscheletro.
La prima di queste due grandi categorie funzionali del neurone è legata alla fase rapida del trasporto assonale e consente di utilizzare i materiali trasportati, quali le componenti proteiche e fosfolipidiche di membrana e i trasmettitori delle vescicole sinaptiche, nelle interazioni specifiche fra membrane pree postsinaptiche e nella regolazione dei meccanismi di rapida emissione e riassunzione di materiali nel terminale presinaptico. La seconda categoria, legata alla fase lenta del trasporto assonale, consente al materiale citoplasmatico trasportato di rinnovare i costituenti neuronali e di regolare, attraverso le strutture filamentose e trabecolari, il metabolismo intraassonale del neurone, incluso quello sinaptico.
Il rinnovo delle proteine sinaptiche, mediato dal trasporto anterogrado attraverso molteplici meccanismi, quali la deplezione delle vescicole, i processi catabolici locali e i processi di degenerazione, rappresenta uno dei momenti fondamentali della vita e della funzione del neurone. D'altra parte, al trasporto retrogrado verrebbe invece assegnato, mediante un meccanismo a feedback, il compito di ‛informare il centro, e quindi il corpo cellulare, di ciò che avviene alla periferia e, di conseguenza, di ciò che è necessario convogliare verso i terminali. Il trasferimento di membrane dai terminali assonici al corpo cellulare in funzione di un meccanismo di ricambio delle membrane stesse e il flusso retrogrado di fattori macromolecolari di accrescimento dalla periferia al centro rappresenterebbero due elementi biologici di rilievo nella funzione di regolazione attribuita al flusso retrogrado.
L'importante ruolo svolto dal trasporto assonale nei fenomeni rigenerativi dell'assone costituisce un ulteriore meccanismo di rilevante significato biologico. Sembra a tale proposito accertato che la fase lenta del trasporto assonale sia essenziale nella costituzione del cono di crescita dell'assone e delle sue componenti strutturali, neurofilamentose e neurotubulari. L'apporto della fase rapida del trasporto assonale ai fenomeni rigenerativi è tuttora in fase di studio, ma è comunque stato accertato che nei momenti iniziali dei fenomeni rigenerativi si verificano modifiche del trasporto rapido di componenti con funzioni molto specializzate, quali gli enzimi correlati al metabolismo dei neurotrasmettitori, che recupererebbero poi la normalità in seguito alla reinnervazione dell'organo bersaglio.
Per quanto riguarda le alterazioni cui può andare incontro il trasporto assonale nella patologia spontanea, la lesione delle strutture neurotubulari e neurofilamentose è prevalente nelle neuropatie tossiche, dismetaboliche ed eredo-degenerative, mentre al trasporto assonale retrogrado può essere attribuito un particolare valore nella genesi di lesioni cellulari reversibili o irreversibili cosiddette primitive.
Quanto all'utilizzazione del trasporto assonale anterogrado e retrogrado nello studio delle connessioni interneuronali, è sufficiente ricordare che a esso si deve una vera e propria rivoluzione nella conoscenza dell'organizzazione anatomica dei circuiti neuronali. L'utilizzazione di traccianti di vario tipo trasportati lungo l'assone, e suscettibili di essere istologicamente visualizzati e localizzati nel sistema nervoso, ha infatti consentito risultati sorprendenti nel rivelare le connessioni anche di singole unità cellulari, nello studio delle loro interrelazioni anatomofunzionali e delle loro diramazioni collaterali e nello studio neurochimico dei circuiti neuronali.
bibliografia
Aletta, J. M., Goldberg, D. J., Rapid and precise down regulation of fast axonal transport of transmitter in an identified neuron, in ‟Science", 1982, CCXVIII, pp. 913-916.
Bak, I. J., Markham, C. H., Cook, M. L., Stevens, J. G., Intraaxonal transport of herpes simplex virus in the rat central nervous system, in ‟Brain research", 1977, CXXXVI, pp. 415-429.
Bentivoglio, M., Kuypers, H. G. J. M., Catsman-Berrevoets, C.E., Dann, O., Fluorescent retrograde labeling in rat by means of substances binding specifically to adenine-thymine rich DNA, in ‟Neuroscience letters", 1979, XII, pp. 235-240.
Bentivoglio, M., Kuypers, H. G. J. M., Catsman-Berrevoets, C. E., Loewe, H., Dann, O., Two new fluorescent retrograde neuronal tracers which are transported over long distances, in ‟Neuroscience letters", 1980, XVIII, pp. 25-30.
Bigotte, L., Olsson, Y., Retrograde transport of doxorubicin (adriamycin) in peripheral nerves of mice, in ‟Neuroscience letters", 1982, XXXII, pp. 217-221.
Bigotte, L., Olsson, Y., Toxic effects of adriamycin on the central nervous system, in ‟Acta neuropathologica" (Berl.), 1983, LXI, pp. 291-299.
Black, M. M., Lasek, R. J., Slow components of axonal transport: two cytoskeletal networks, in ‟Journal of cell biology", 1980, LXXXVI, pp. 616-623.
Brimijoin, S., Stop-flow: a new technique for measuring axonal transport, and its application to the transport of dopamine-beta-hydroxylase, in ‟Journal of neurobiology", 1975, VI, pp. 379-394.
Cowan, W. M., Gottlieb, D. I., Hendrickson, A. E., Price, J. L., Woolsey, T. A., The autoradiographic demonstration of axonal connections in the central nervous system, in ‟Brain research", 1972, XXXVII, pp. 21-51.
Cuénod, M., Bagnoli, P., Beaudet, A., Rustioni, A., Wiklund, L., Strett, P., Transmitter-specific retrograde labeling of neurons, in Cytochemical methods in neuroanatomy, New York 1982, pp. 17-44.
Dahlin, L.B., Rydevik, B., McLean, W. G., Sjöstrand, J., Changes in fast axonal transport during experimental nerve compression at low pressure, in ‟Experimental neurology", 1984, LXXXIV, pp. 29-36.
Droz, B., Renewal of synaptic proteins, in ‟Brain research", 1973, LXII, pp. 383-394.
Droz, B., Rambourg, A., Axonal smooth reticulum and fast orthograde transport of membrane constituents, in Axoplasmic transport (a cura di D. G. Weiss), Berlin 1982, pp. 384-389.
Ellisman, M. H., A hypothesis for rapid axoplasmic transport based upon focal interactions between axonal membrane systems and the microtrabecular crossbridges of the axoplasmic matrix, in Axoplasmic transport (a cura di D. G. Weiss), Berlin 1982, pp. 390-396.
Ellisman, M. H., Lindsey, J. D., Organization of axoplasm. Membraneous and fibrillar components possibly involved in fast neuroplasmic transport, in Axoplasmic transport (a cura di D. G. Weiss), Berlin 1982, pp. 55-63.
Ellisman, M. H., Porter, K. R., Microtrabecular structure of the axoplasmic matrix: visualization of cross-linking structures and their distribution, in ‟Journal of cell biology", 1980, LXXXVII, pp. 464-479.
Fink, D. J., Gainer, H., Axonal transport of proteins: a new view using in vivo covalent labeling, in ‟Journal of cell biology", 1980, LXXXV, pp. 175-186.
Forman, D.S., Padjen, A. L., Siggins, G.R., Axonal transport of organelles visualized by light microscopy: cinemicrographic and computer analysis, in ‟Brain research", 1977, CXXXVI, pp. 197-213.
Frizell, M., Sjöstrand, J., Changes in axonal transport in relation to the nerve cell body response during peripheral nerve regeneration, in Posttraumatic peripheral nerve regeneration: experimental basis and clinical implications (a cura di A. Gorio, H. Millesi e S. Mingrino), New York 1981, pp. 35-44.
Gainer, H., Fink, D. J., Evidence for slow retrograde transport of serum albumin in rat sciatic nerve, in ‟Brain research", 1982, CCXXXIII, pp. 404-408.
Goodpasture, E.W., The axis-cylinders of peripheral nerves as portals of entry to the central nervous system for the virus of herpes simplex in experimentally infected rabbits, in ‟American journal of pathology", 1925, I, pp. 11-28.
Grafstein, B., Forman, D.S., Intracellular transport in neurons, in ‟Physiological reviews", 1980, LX, pp. 1167-1283.
Graham, R.C., Karnovsky, M., The early stages of absorption of injected horseradish peroxidase in the proximal tubules of mouse kidney in ultrastructural cytochemistry by a new technique, in ‟Journal of histochemistry and cytochemistry", 1966, XIV, pp. 291-302.
Griffin, J. W., price, D. L., Hoffman, P. N., Neurotoxic probes of the axonal cytoskeleton, in ‟Trends in neurosciences", 1983, VI, pp. 490-495.
Gross, G. W., Kreutzberg, G.W., Rapid axoplasmic transport in the olfactory nerve of the pike. I. Basic transport parameters for proteins and amino acids, in ‟Brain research", 1978, CXXXIX, pp. 65-76.
Hammerschlag, R., Stone, G. C., Fast axonal transport as endomembrane flow, in Axoplasmic transport (a cura di D. G. Weiss), Berlin 1982, pp. 406-413.
Hayes, N. L., Rustioni, A., Dual projections of single neurons are visualized simultaneously: use of enzymatically inactivated [3H]HRP, in ‟Brain research", 1979, CLXV, pp. 321-326.
Hoffman, P. N., Lasek, R. J., The slow component of axonal transport. Identification of major structural polypeptides of the axon and their generality among mammalian neurons, in ‟Journal of cell biology", 1975, LXVI, pp. 351-366.
Ishikawa, H., Tsukita, S., Morphological and functional correlates of axoplasmic transport, in Axoplasmic transport (a cura di D. G. Weiss), Berlin 1982, pp. 251-259.
Koda, L. Y., Van der Kooy, D., Doxorubicin: a fluorescent neurotoxin retrogradely transported in the central nervous system, in ‟Neuroscience letters", 1983, XXXVI, pp. 1-8.
Kristensson, K., Retrograde transport of macromolecules in axons, in ‟Annual review of pharmacology and toxicology", 1978, XVIII, pp. 97-110.
Kristensson, K., Retrograde signaling of nerve cell body in response to trauma, in Postttraumatic peripheral nerve regeneration: experimental basis and clinical implications (a cura di A. Gorio, H. Millesi e S. Mingrino), New York 1981, pp. 27-34.
Kristensson, K., Olsson, Y., Retrograde axonal transport of protein, in ‟Brain research", 1971, XXIX, pp. 363-365.
Kristensson, K., Olsson, Y., Uptake and retrograde axonal transport of peroxidase in hypoglossal neurones. Electron microscopical localization in the neuronal perikaryon, in ‟Acta neuropathologica" (Berl.), 1971, XIX, pp. 1-9.
Kristensson, K., Olsson, Y., Diffusion pathways and retrograde axonal transport of protein tracers in peripheral nerves, in ‟Progress in neurobiology", 1973, I, pp. 85-109.
Kristensson, K., Olsson, Y., Retrograde transport of horseradish peroxidase in transected axons. 3. Entry into injured axons and subsequent localization in perikarvon, in ‟Brain research", 1976, CXV, pp. 201-213.
Kuypers, H. G. J. M., Bentivoglio, M., Van der Koody, D., Catsman-Berrevoets, C.E., Retrograde transport of bisbenzimide and propidium iodide through axons in their parent cell bodies, in ‟Neuroscience letters", 1979, XII, pp. 1-7.
Kuypers, H. G. J.M., Catsman-Berrevoets, C.E., Padt, R.E., Retrograde axonal transport of fluroescent substances in the rat's forebrain, in ‟Neuroscience letters", 1977, VI, pp. 127-133.
Landis, S.C., Neuronal growth cones, in ‟Annual review of physiology", 1983, XLV, pp. 567-580.
Larsson, P.A., Goldstein, M., Dahlstrom, A., A new methodological approach for studying axonal transport: cytofluorometric scanning of nerves, in ‟Journal of histochemistry and cytochemistry", 1984, XXXII, pp. 7-16.
Lasek, R. J., Axonal transport: a dynamic view of neuronal structures, in ‟Trends in neurosciences", 1980, III, pp. 87-91.
Lasek, R. J., Joseph, B. S., Whitlock, D.G., Evaluation of a radioautographic neuroanatomical tracing method, in ‟Brain research", 1968, VIII, pp. 319-336.
Lasek, R. J., McQuarrie, I.G., Wujek, J.R., The central nervous system regeneration problem: neuron and environment, in Posttraumatic peripheral nerve regeneration: experimental basis and clinical implications (a cura di A. Gorio, H. Millesi e S. Mingrino), New York 1981, pp. 59-70.
LaVail, J. H., LaVail, M. M., Retrograde axonal transport in the central nervous system, in ‟Science", 1972, CLXXVI, pp. 1416-1417.
LaVail, J. H., Rapisarda, S., Sugino, I. K., Evidence against the smooth endoplasmic reticulum as a continuous channel for the retrograde axonal transport of horseradish peroxidase, in ‟Brain research", 1980, CXCI, pp. 3-20.
Levi-Montalcini, R., The nerve growth factor: its mode of action on sensory and sympathetic nerve cells, in ‟Harvey lectures", 1966, LX, pp. 217-259.
Lubinska, L., On axoplasmic flow, in ‟International review of neurobiology", 1975, XVII, pp. 241-296.
Lubinska, L., Niemierko, S., Velocity and intensity of bidirectional migration of acetylcholinesterase in transected nerves, in ‟Brain research", 1971, XXVII, pp. 329-342.
McQuarrie, I.G., Grafstein, B., Axon outgrowth enhanced by a previous nerve injury, in ‟Archives of neurology", 1973, XXIX, pp. 53-55.
Marchisio, P.C., Gremo, F., Sjöstrand, J., Axonal transport in embryonic neurons. The possibility of a proximo-distal axolemmal transfer of glycoproteins, in ‟Brain research", 1975, LXXXV, pp. 281-285.
Mesulam, M.-M., Tetramethyl benzidine for horseradish peroxidase neurohistochemistry: a non-carcinogenic blue reaction-product with superior sensitivity for visualizing neural afferents and efferents, in ‟Journal of histochemistry and cytochemistry", 1978, XXVI, pp. 106-117.
Miani, N., Analysis of the somato-axonal movement of phospholipids in the vagus and hypoglossal nerves, in ‟Journal of neurochemistry", 1963, X, pp. 859-874.
Miani, N., Derenzis, G., Michetti, F., Correr, S., Olivieri-Sangiacomo, C., Caniglia, A., Axonal transport of S-100 protein in mammalian nerve fibres, in ‟Journal of neurochemistry", 1972, XIX, pp. 1387-1394.
Ochs, S., Calcium and the mechanism of axoplasmic transport, in ‟Federation proceedings", 1982, XLI, pp. 2301-2306.
Paravicini, U., Stöckel, K., Thoenen, H., Biological importance of retrograde axonal transport of nerve growth factor in adrenergic neurones, in ‟Brain research", 1975, LXXXIV, pp. 279-291.
Peters, A., Palay, S.L., Webster, H., The fine structure of the nervous system. The cells and their processes, New York 1970.
Purves, D., Nja, A., Effect of nerve growth factor on synaptic depression after axotomy, in ‟Nature", 1976, CCLX, pp. 535-536.
Ramón y Cajal, S., Histologie du système nerveux de l'homme et des Vertébrés, vol. I, Paris 1909, vol. II, Paris 1911, Madrid 1952.
Ramón y Cajal, S., Degeneration and regeneration of the nervous system, Cambridge 1928.
Repérant, J., Vesselkin, N. P., Kenigfest, N. B., Miceli, D., Rio, J. P., Bidirectional axonal and transcellular transport of [3H]adenosine from the lamprey retina, in ‟Brain research", 1982, CCXLVI, pp. 305-310.
Rosina, A., Rapid anterograde movement of the fluorescent tracer Fast Blue: a new method for tracing central connections, in ‟Neuroscience letters", 1982, XXXIII, pp. 217-221.
Seiler, M., Schwab, M. E., Specific retrograde transport of nerve growth factor (NGF) from neocortex to nucleus basalis in the rat, in ‟Brain research", 1984, CCC, pp. 33-39.
Sparrow, J. R., Grafstein, B., Prior collateral sprouting enhances axonal regeneration, in ‟Brain research", 1983, CCLXIX, pp. 133-136.
Stöckel, K., Schwab, M., Thoenen, H., Comparison between the retrograde axonal transport of nerve growth factor and tetanus toxin in motor, sensory and adrenergic neurons, in ‟Brain research", 1975, CIC, pp. 1-16.
Stone, G.C., Hammerschlag, R., Bobinski, J.A., Involvement of coated vescicles in the initiation of fast axonal transport, in ‟Brain research", 1984, CCXCI, pp. 219-228.
Taylor, A.C., Weiss, P.A., Demonstration of axonal flow by the movement of tritium-labeled protein in mature optic nerve fibers, in ‟Proceedings of the National Academy of Sciences", 1965, LIV, pp. 1521-1527.
Thoenen, H., Barde, Y.-A., Edgar, D., Factors involved in the regulation of the survival and differentiation of neurons, in Repair and regeneration of the nervous system (a cura di J. G. Nicholls), Berlin 1982, pp. 173-185.
Veraa, R. P., Grafstein, B., Cellular mechanisms for recovery from nervous system injury: a conference report, in ‟Experimental neurology", 1981, LXXI, pp. 6-75.
Weiss, D.G., General properties of axoplasmic transport, in Axoplsmic transport in physiology and pathology (a cura di D. G. Weiss e A. Gorio), Berlin 1982, pp. 1-14.
Weiss, D.G., Gross, G. W., The microstream hypothesis of axoplasmic transport: characteristics, predictions and compatibility with data, in Axoplasmic transport (a cura di D. G. Weiss), Berlin 1982, pp. 362-382.
Weiss, P.A., Neuronal dynamics, in ‟Neuroscience research program bulletin", 1967, V, pp. 371-400.
Weiss, P.A., Hiscoe, H.B., Experiments on the mechanism of nerve growth, in ‟Journal of experimental zoology", 1948, CVII, pp. 315-395.
Wiesel, T. N., Hubel, D. H., Lam, D. M. K., Autoradiographic demonstration of ocular-dominance columns in the monkey striate cortex by means of transneuronal transport, in ‟Brain research", 1974, LXXIX, pp. 273-279.
Willard, M., Cowan, W. M., Vagelos, P.R., The polypeptide composition of intra-axonally transported proteins: evidence for four transport velocities, in ‟Proceedings of the National Academy of Sciences", 1974, LXXI, pp. 2183-2187.
Wise, S.P., Jones, E. G., Berman, N., Direction and specificity of the axonal and transcellular transport of nucleosides, in ‟Brain research", 1978, CXXXIX, pp. 197-217.