
NEURONI SPECCHIO
Neuroni specchio
L’uomo è un animale eminentemente sociale la cui vita dipende dalla capacità di capire cosa fanno gli altri, comprendendone le intenzioni e interpretandone i sentimenti. Senza questa capacità gli esseri umani non riuscirebbero a interagire gli uni con gli altri, né tanto meno a creare forme di convivenza sociale. Secondo il punto di vista tradizionale, le azioni degli altri, come pure le loro intenzioni e le loro emozioni, sono comprese mediante un processo inferenziale sostanzialmente simile a quello che usiamo per individuare le cause di fenomeni puramente fisici. In base a questo punto di vista, quando osserviamo una persona agire, il nostro sistema nervoso capta, mediante la vista e le altre modalità sensoriali, una serie di informazioni che un complesso apparato cognitivo elabora e paragona con precedenti esperienze simili. Alla fine di questo processo l’osservatore ha capito cosa fanno gli altri e quali sono le loro intenzioni.
Diversa è invece la teoria secondo la quale il meccanismo che ci permette di capire le azioni degli altri è radicalmente diverso da quelli impiegati per spiegare i processi fisici. Secondo questa impostazione, noi capiamo gli altri perché ci mettiamo ‘nei loro panni’, ci immaginiamo nella loro situazione e ‘simuliamo’ quello che faremmo se davvero fossimo in quel frangente. Volendo fare un paragone tra i due sistemi di comprensione sopra richiamati, potremmo dire che il primo ricalca l’atteggiamento tipico del detective (pensiamo, per es., alle sottili analisi di uno Sherlock Holmes), mentre il secondo rimanda alla comprensione in prima persona cara a molti fenomenologi e, in particolare, a Maurice Merleau-Ponty.
È possibile che, in certe condizioni, l’osservatore adotti effettivamente un atteggiamento alla Sherlock Holmes. Così come è possibile che in altre condizioni adotti un atteggiamento simulativo. Tuttavia, la mancanza di sforzo e la facilità con le quali normalmente viene compreso il comportamento degli altri suggeriscono che possa esistere un diverso meccanismo, indubbiamente più diretto e immediato di quelli citati, che ci permetterebbe di capire gli altri senza la mediazione di processi inferenziali o la simulazione attiva del comportamento altrui.
Un’importante scoperta neurofisiologica avvenuta negli anni Novanta del 20° sec. ha messo in luce l’esistenza di un tale meccanismo di comprensione, grazie al quale le azioni eseguite dagli altri, captate dai sistemi sensoriali, sono automaticamente trasferite al sistema motorio dell’osservatore, permettendogli così di avere una copia motoria del comportamento osservato, quasi fosse lui stesso a eseguirlo. I neuroni che compiono questa trasformazione dell’azione da un formato sensoriale a uno motorio sono stati chiamati neuroni specchio. La loro scoperta ha reso possibile una nuova concezione del sistema motorio, aprendo inoltre la via all’indagine neurofisiologica di campi prima appannaggio esclusivo di discipline umanistiche.
Nelle pagine che seguono, dopo avere esposto i dati essenziali sulle proprietà dei neuroni specchio, esamineremo le ricerche sul ‘sistema specchio’ condotte negli ultimi anni mostrando la loro importanza, oltre che nell’ambito delle neuroscienze, anche nel campo della cognizione sociale.
Il sistema dei neuroni specchio nella scimmia
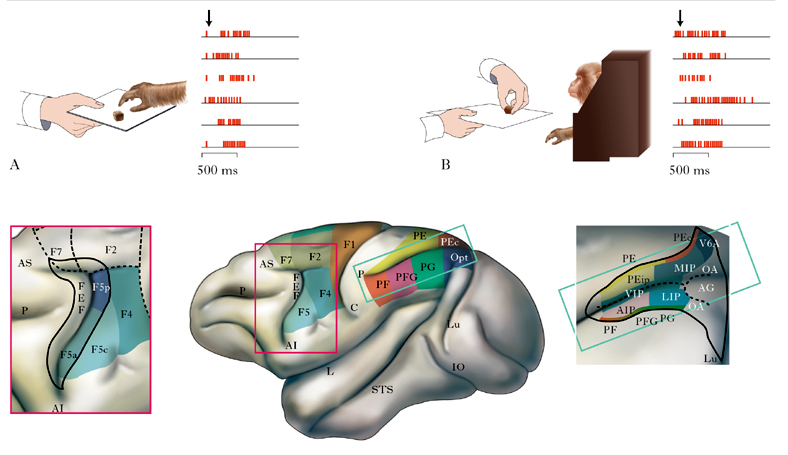
I neuroni specchio sono stati originariamente scoperti nella corteccia premotoria ventrale del macaco, una specie, come l’uomo, eminentemente sociale. Essi formano una particolare classe di cellule nervose che si attivano sia quando la scimmia esegue uno specifico atto motorio, per es. afferrare un pezzo di cibo, sia quando essa osserva un altro individuo (scimmia o uomo) eseguire un atto motorio identico o simile (fig. 1, parte superiore: il neurone specchio si attiva sia quando la scimmia afferra un oggetto (A) sia quando vede un altro individuo afferrarlo (B); sono indicate le corrispondenti scariche elettriche). I neuroni specchio non rispondono alla semplice presentazione del cibo o di altri oggetti che pure interessano l’animale, né si attivano all’osservazione di un’azione mimata senza la presenza dell’oggetto. Affinché il neurone specchio si attivi (o, per riprendere un’espressione usata spesso in fisiologia, ‘spari’) è necessaria l’effettiva interazione della mano con un oggetto bersaglio dell’azione (Rizzolatti, Craighero 2004).
Essendo neuroni motori, i neuroni specchio sparano, come gli altri neuroni presenti nella corteccia premotoria, in associazione a un atto motorio (per es., afferrare), mentre non si attivano per i singoli movimenti delle dita che formano tale atto motorio. Sulla base delle loro proprietà motorie, i neuroni specchio sono stati suddivisi in varie classi: tra queste le più comuni sono quelle dei neuroni dell’afferrare, del manipolare, dello strappare, del tenere. I neuroni specchio non si attivano soltanto durante movimenti eseguiti con la mano: vi sono, infatti, neuroni specchio che sparano anche quando la scimmia compie o osserva atti motori eseguiti con la bocca.
Dopo la scoperta dei neuroni specchio nella corteccia premotoria, altri studi hanno dimostrato la loro presenza nel lobo parietale inferiore, in particolare nelle aree PFG e AIP. La parte inferiore della fig. 1 mostra la visione laterale del cervello di macaco con la parcellazione anatomo-funzionale della corteccia motoria frontale – aree indicate con la lettera F seguite da numeri arabi – e della corteccia parietale posteriore – aree indicate con la lettera P seguita da una (o più) lettere, secondo la convenzione stabilita dal neuroscienziato austriaco Constantin von Economo (1876-1931). Il riquadro verde di destra mostra le aree poste all’interno del solco intraparietale (IP): AIP, area intraparietale anteriore; LIP, area intraparietale laterale; MIP, area intraparietale mediale; PEip, area PE intraparietale; VIP, area intraparietale ventrale. Altre abbreviazioni: AS, solco arcuato superiore, AI, solco arcuato inferiore; C, solco centrale; L, solco laterale; STS, solco temporale superiore; Lu, solco lunato; Opt, area occipito-parietale; FEF, campi oculari frontali. Il riquadro rosso di sinistra mostra un ingrandimento della porzione posteriore del lobo frontale con indicata la suddivisione dell’area F5 nelle sue subaree costituenti: F5c (convessità), F5a (anteriore), F5p (posteriore). Queste ultime due aree sono poste nel labbro posteriore del solco arcuato.
Queste aree ricevono informazioni visive dalla corteccia del solco temporale superiore (STS) e sono riccamente connesse alla corteccia premotoria. STS, pur rispondendo all’osservazione di atti motori compiuti da altri, non deve essere considerato parte del sistema specchio, in quanto i suoi neuroni non posseggono proprietà motorie. Pertanto, il sistema dei neuroni specchio è costituito, nella scimmia, da due principali settori: la corteccia premotoria e il lobo parietale inferiore (Rizzolatti, Luppino 2001).
È lecito chiedersi perché il sistema motorio contenga neuroni che rispondono alla visione di atti motori eseguiti da altri e quale possa essere la loro funzione. L’ipotesi generalmente accettata è che i neuroni specchio sono necessari per una comprensione immediata dell’azione altrui. Per avere un’idea del meccanismo neurofisiologico alla base di questa comprensione si consideri l’esempio seguente. Un individuo, poniamo che si chiami Mario, afferra una tazzina di caffè. Quando compie quest’azione Mario sa cosa sta facendo. Tale sua conoscenza corrisponde all’attivazione di una serie di neuroni motori che preparano l’atto dell’afferrare la tazzina. Quando Mario osserva Anna che afferra una tazzina, gli stessi neuroni motori che si sono attivati durante l’atto motorio di Mario si attivano di nuovo, dando così a quest’ultimo la rappresentazione motoria (detta anche atto motorio potenziale) dell’atto compiuto da Anna. Mario quindi capisce cosa Anna sta facendo perché l’atto motorio potenziale generato dall’osservazione di Anna corrisponde a quello che Mario genera volontariamente quando prepara o esegue lo stesso atto. Ecco in breve il meccanismo specchio.
Questa interpretazione del meccanismo specchio è stata avanzata, inizialmente in via teorica, usando come modello la teoria motoria del linguaggio dello psicologo statunitense Alvin M. Liberman (1917-2000). Secondo Liberman, la comprensione di un fonema non si basa sulle sue proprietà acustiche ma sulla capacità che esso ha di evocare in chi lo ascolta il programma motorio necessario per la sua produzione. Il meccanismo specchio appare essere la controparte neurofisiologica di tale idea. L’atto motorio evoca in colui che lo osserva il programma motorio richiesto per eseguirlo.
In questi ultimi anni una serie di esperimenti di neurofisiologia ha avvalorato questa spiegazione. Analizziamo come si è arrivati a tale risultato. Il più delle volte nel campo delle neuroscienze la via più diretta per stabilire la funzione di un sistema neurale è quella di distruggerlo e di esaminare i deficit che ne conseguono. Nel caso dei neuroni specchio, però, questo metodo non poteva essere applicato dato il numero delle aree coinvolte nel sistema. La distruzione del sistema specchio poteva produrre, infatti, un deficit cognitivo generalizzato, rendendo così impossibile la definizione delle sue funzioni specifiche.
Si è quindi adottata una strategia diversa. Si è pensato che, se i neuroni specchio realmente mediano la comprensione degli atti motori osservati, la loro attivazione dovrebbe riflettere il significato dell’azione anche là dove l’informazione visiva è parziale o addirittura del tutto assente. Se l’animale capisce l’atto motorio di chi agisce, i neuroni specchio dovrebbero attivarsi indipendentemente da quale sia la via che porta l’animale a capire ciò che osserva.
Due esperimenti hanno dimostrato che tale attivazione effettivamente avviene. Nel primo si è esaminata la capacità dei neuroni specchio di riconoscere un’azione sulla base del suono che la caratterizza. L’attività dei neuroni è stata registrata in due condizioni sperimentali: mentre la scimmia osservava un atto motorio accompagnato da un suono distintivo (come, per es., rompere un pezzo di carta o sbucciare un’arachide) e mentre ascoltava lo stesso suono senza vedere l’atto motorio. I risultati hanno mostrato che la maggior parte dei neuroni specchio rispondeva sia all’osservazione dell’atto motorio sia alla presentazione del suono. I neuroni che avevano tali proprietà sono stati chiamati neuroni specchio audiovisivi.
Il secondo esperimento è stato condotto allo scopo di stabilire se i neuroni specchio si sarebbero attivati anche quando la scimmia non vedeva lo sperimentatore compiere l’intero atto motorio che selettivamente attivava il neurone, ma aveva elementi sufficienti per capire cosa stesse facendo. Anche in questo esperimento due erano le condizioni fondamentali: nella prima, la scimmia vedeva lo sperimentatore afferrare un oggetto (condizione di visione); nella seconda, la scimmia vedeva lo sperimentatore muovere il braccio verso l’oggetto per afferrarlo, ma non l’afferramento dell’oggetto che era nascosto da un pannello (condizione di non visione). Dai risultati è emerso che più della metà dei neuroni specchio dell’area premotoria si attivava nella condizione in cui la scimmia non vedeva l’effettivo afferramento dell’oggetto. Questo avveniva però solo nel caso in cui la scimmia sapeva che dietro lo schermo c’era del cibo, in caso contrario l’atto dello sperimentatore non evocava alcuna risposta. Ciò mostra come sia la rappresentazione mentale del significato dell’atto motorio dello sperimentatore, e non le contingenze visive, a determinare la scarica dei neuroni specchio.
Neuroni specchio-simili nella corteccia motoria
L’idea che il sistema motorio generi atti motori potenziali osservando quelli compiuti da altri ha avuto un ulteriore sviluppo negli ultimi anni. Si è osservato, infatti, che esistono neuroni nella corteccia premotoria dorsale e, addirittura, nell’area motoria primaria che si attivano durante l’osservazione di movimenti di raggiungimento diretti verso un bersaglio.
Prove dell’esistenza di questi neuroni derivano da due studi condotti su scimmie condizionate a compiere specifici movimenti. Nel primo, le scimmie imparavano ad azionare una leva il cui movimento determinava lo spostamento di un cursore, sullo schermo di un computer, da una posizione centrale a una periferica. Le scimmie erano premiate quando riuscivano a mettere il cursore in una posizione precisa, definita da un pallino colorato, posizione che variava da prova a prova. È risultato che sia nell’area motoria primaria sia nella premotoria dorsale vi erano neuroni che si attivavano non solo quando la scimmia eseguiva il compito, ma anche quando, immobile, vedeva il cursore (mosso dallo sperimentatore, il quale era però invisibile alla scimmia) spostarsi nella direzione corretta. Questi neuroni appaiono differire dai neuroni specchio ‘classici’ in quanto non richiedono, a differenza di questi ultimi, l’osservazione di un’interazione tra un effettore biologico e il suo bersaglio. Una spiegazione della loro attivazione potrebbe essere che la scimmia, nel vedere il cursore muoversi, si formi un’immagine mentale del proprio movimento. È più probabile, però, che il cursore che appare e si muove rappresenti per la scimmia un analogo (anche se astratto) di una mano che si muove e che il suo movimento verso il bersaglio evochi nella scimmia la rappresentazione del movimento della mano.
Anche nel secondo studio le scimmie erano allenate, usando ancora una leva, a muovere un cursore verso bersagli che in questo caso, però, comparivano continuamente uno dopo l’altro sullo schermo in diverse posizioni. Anche in questo studio la scimmia eseguiva il compito o osservava il movimento del cursore, mosso dallo sperimentatore senza che questi fosse visibile. La condizione di osservazione prevedeva tre varianti: osservazione del cursore che raggiungeva il bersaglio, osservazione solo del bersaglio e osservazione solo del cursore che si muoveva sullo schermo. La registrazione dei neuroni ha mostrato che l’osservazione del cursore che raggiungeva il bersaglio determinava una scarica simile a quella registrata durante l’esecuzione; l’osservazione del cursore senza bersaglio o dei bersagli senza cursore non determinava invece alcuna risposta o causava risposte molto deboli.
Questi due studi mostrano come atti motori potenziali vengano generati non solo nel circuito specchio classico, ma anche nella corteccia premotoria dorsale e nella corteccia motoria e come in queste due aree non sia necessaria l’interazione mano-oggetto che può essere sostituita da oggetti che si muovono in maniera finalistica (Tkach, Reimer, Hatsopoulos 2007).
Il meccanismo specchio nell’uomo
La scoperta dei neuroni specchio nella scimmia ha ovviamente subito posto il problema della loro possibile esistenza nell’essere umano. Molteplici esperimenti hanno dimostrato che un meccanismo specchio è presente anche nell’uomo. Prove in tal senso sono state ottenute sia mediante tecniche non invasive di neurofisiologia, sia grazie a esperimenti di brain imag-ing (visualizzazione dell’attività cerebrale in vivo).
È noto da tempo che quando una persona compie dei movimenti, per es. chiude e apre la mano, i ritmi elettrici delle regioni centrali della corteccia cerebrale, registrati mediante elettroencefalografia (EEG), aumentano di frequenza o, per usare il gergo tecnico, si desincronizzano. La desincronizzazione dei ritmi cerebrali indica che la corteccia cerebrale aumenta la sua attività. Studi sia di EEG sia di magnetoencefalografia (MEG, una tecnica simile all’EEG ma basata sulla registrazione di campi magnetici generati dall’attività elettrica cerebrale) hanno mostrato che la semplice osservazione di un’azione determinava una desincronizzazione della corteccia analoga a quella che normalmente si registra durante l’esecuzione dell’azione, quasi che l’osservatore la stesse eseguendo. Nell’uomo, proprio come nella scimmia, l’azione osservata ‘raggiunge’ le aree motorie e le attiva.
Altre prove dell’esistenza del sistema specchio nell’uomo provengono da studi di stimolazione magnetica della corteccia motoria attraverso lo scalpo (TMS, Transcranic Magnetic Stimulation) e simultanea registrazione delle risposte dei muscoli controllati dalla corteccia stimolata. La logica alla base degli esperimenti è la seguente: se l’osservazione di un determinato atto motorio (per es., afferrare) causa, mediante il meccanismo specchio, un aumento dell’eccitabilità della zona di corteccia motoria che controlla i muscoli coinvolti in quell’atto motorio, l’eccitabilità di quella zona dovrebbe aumentare durante l’osservazione dell’atto motorio rispetto a una condizione di riposo. I risultati degli esperimenti di TMS hanno confermato quest’ipotesi: l’osservazione di un atto motorio eseguito da un altro individuo determina un aumento selettivo dell’attività dei muscoli coinvolti nell’esecuzione dell’atto motorio osservato.
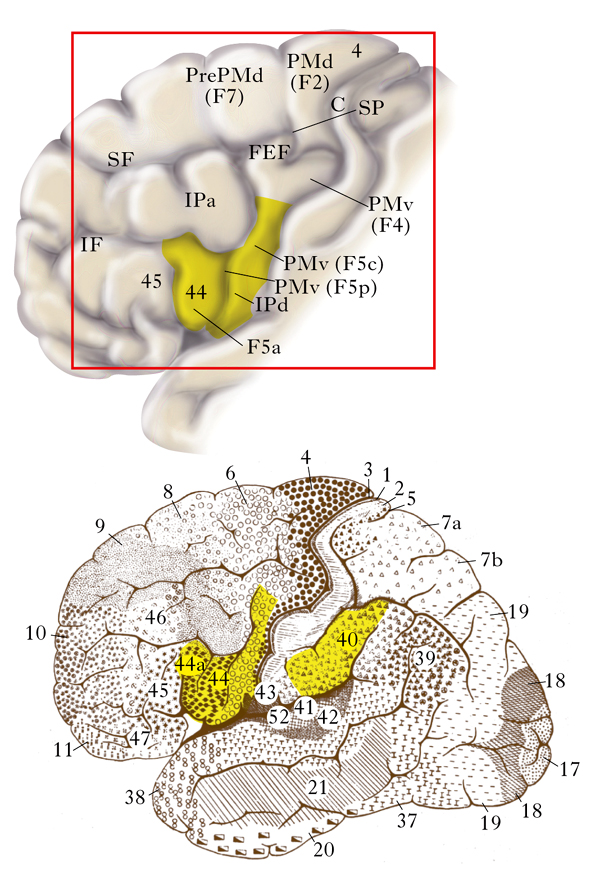
Gli esperimenti di neurofisiologia hanno mostrato l’esistenza del meccanismo specchio nell’uomo; tuttavia, per la loro natura, non sono stati in grado di dare informazioni sulla sua localizzazione. Ciò è stato ottenuto usando tecniche di brain imaging: la tomografia a emissione di positroni o PET (Positron Emission Tomography) e la risonanza magnetica funzionale o fMRI (functional Magnetic Resonance Imaging). Mediante tali tecniche si è mostrato che il sistema specchio che codifica gli atti motori è costituito nell’uomo, come nella scimmia, da due grandi regioni: il lobulo parietale inferiore e l’area premotoria ventrale più la parte posteriore del giro frontale inferiore (Rizzolatti, Sinigaglia 2006). Nella fig. 2 è rappresentata, in basso, la visione laterale del cervello di uomo con indicate le aree citoarchitettoniche secondo la suddivisione di Brodmann. In giallo sono evidenziate le aree che costituiscono il sistema specchio parieto-frontale. Nel riquadro rosso è riportata la visione ingrandita del lobo frontale. In giallo, il nodo frontale del sistema specchio frontale con indicate le possibili omologie tra uomo e scimmia (F5a, F5p e F5c) della porzione posteriore del lobo frontale.
Organizzazione
Il problema dell’organizzazione del sistema specchio parieto-frontale è stato affrontato in genere in esperimenti di fMRI in cui si facevano vedere a soggetti normali filmati che mostravano movimenti transitivi, caratterizzati da interazioni specifiche tra effettori biologici (mano, bocca, piede) e oggetti. Questi studi hanno mostrato che la corteccia premotoria è organizzata in maniera ‘somatotopica’: gli atti motori eseguiti con i diversi effettori risultano infatti localizzati in settori spazialmente separati, anche se parzialmente sovrapposti. L’osservazione degli atti motori eseguiti con il piede determina un’attivazione della parte dorsale dell’area premotoria ventrale, quella di atti motori compiuti con la bocca un’attivazione della parte ventrale della stessa area, mentre quella di atti motori compiuti con la mano un’attivazione in una zona intermedia. Sia l’osservazione di atti motori fatti con la mano sia quella di atti compiuti con la bocca interessano anche la parte posteriore della circonvoluzione frontale inferiore, dove nell’emisfero di sinistra è localizzata l’area di Broca.
Recentemente si è studiata la rappresentazione dei movimenti di raggiungimento di oggetti con la mano, isolandola dai movimenti di afferramento. I risultati hanno mostrato che l’osservazione dei movimenti di raggiungimento determina l’attivazione di una parte della corteccia premotoria distinta da quella che codifica l’afferramento; più precisamente, la massima attivazione è stata riscontrata attorno al solco frontale superiore nell’area premotoria dorsale.
La rappresentazione dei comportamenti motori nel lobo parietale è più complessa. Per quanto riguarda gli atti motori transitivi, si è visto che essi sono rappresentati nella regione attorno al solco intraparietale, con una tendenza dei movimenti della bocca a essere rappresentati verso la parte anteriore di questa regione, quelli del piede caudalmente e quelli della mano in una posizione centrale.
Un interessante dato è emerso da un recente esperimento di fMRI in cui i soggetti osservavano quattro atti motori (afferrare, tirare, lasciare cadere e spingere) eseguiti con la mano, il piede o la bocca. I risultati hanno mostrato che la logica dell’organizzazione del lobo parietale non è solo somatotopica, ma dipende anche dal significato dell’atto motorio. Gli atti motori positivi (quelli diretti verso colui che agisce, come afferrare o tirare) determinano un’attivazione del settore ventrale della regione intraparietale, mentre quelli negativi (diretti via da colui che agisce, come lasciare cadere o spingere) di un settore dorsale della stessa regione. L’interpretazione di questi dati è che la regione motoria della mano sia il fulcro di un sistema che risponde agli atti motori anche quando questi sono eseguiti con altri effettori. La zona ventrale generalizza gli atti motori positivi interpretandoli secondo il loro significato motorio indicato dalla codifica dei movimenti della mano e lo stesso fa la zona dorsale per gli atti motori negativi.
Altri studi hanno esaminato l’organizzazione del sistema specchio del lobo parietale durante l’osservazione di atti motori diversi da quelli che portano all’interazione con oggetti. Lo studio citato prima sui movimenti di raggiungimento ha così mostrato che l’osservazione di tali movimenti attiva una regione dorsale rispetto a quella dei movimenti di interazione distale posta nel lobulo parietale superiore. Anche i movimenti intransitivi hanno una rappresentazione parietale specifica; essi, indipendentemente se simbolici o privi di significato, determinano un’attivazione della parte posteriore del giro sopramarginale che si estende nel giro angolare. Infine, l’osservazione di atti motori eseguiti con strumenti attiva la parte più rostrale del giro sopramarginale, ventralmente all’area di rappresentazione della regione di interazione manuale con gli oggetti.
Sistema specchio ed esperienza motoria
Un interessante problema è il rapporto tra conoscenze motorie degli individui e sistema specchio. Gli atti motori maggiormente rappresentati nel sistema specchio sono quelli compiuti più di frequente dall’osservatore? Questo sistema è modificabile nell’adulto con tipi di esercizi motori specifici? Alcune indagini di fMRI hanno di recente affrontato questi problemi.
In un primo studio è stata esaminata l’intensità di attivazione del sistema specchio in ballerini di danza classica, in esperti di capoeira e in persone che non avevano mai danzato. Lo scopo dell’esperimento era quello di stabilire se le aree cerebrali di pertinenza del sistema specchio si attivavano in maniera diversa secondo l’esperienza nella danza dei vari individui. I risultati hanno mostrato che l’osservazione di passi di capoeira attiva il sistema dei neuroni specchio maggiormente negli esperti di capoeira rispetto ai ballerini classici e ai principianti. Viceversa, l’attivazione nei ballerini classici era maggiore quando osservavano un balletto classico rispetto alla capoeira.
In una ricerca successiva, ci si è chiesti se la differenza nell’attivazione nei tre gruppi dipendesse dal fatto che gli esperti di capoeira avessero una maggiore esperienza visiva di questa disciplina o se, invece, l’attivazione dipendesse dalla loro conoscenza motoria dei vari passi richiesti. Nella capoeira alcuni passi sono eseguiti sia da uomini sia da donne, mentre altri sono eseguiti solo da uomini o solo da donne. Partendo da questa osservazione sono stati presentati, durante un esperimento di fMRI, filmati di passi di danza eseguiti da ballerini e da ballerine. I risultati hanno mostrato che il sistema specchio si attivava di più quando i ballerini osservavano passi di danza eseguiti da individui dello stesso sesso. È quindi la conoscenza motoria e non l’esperienza visiva che rappresenta il fattore principale responsabile dell’attivazione del sistema specchio (Rizzolatti, Sinigaglia 2006).
Allo scopo di approfondire i risultati di questi studi, in un ulteriore esperimento è stata esaminata, settimana per settimana, la relazione tra l’apprendimento dei passi di danza e l’intensità dell’attivazione del sistema specchio. I risultati hanno dimostrato che a mano a mano che i ballerini imparavano i passi di danza vi era un incremento dell’attivazione del loro sistema specchio e che questo era in relazione con le capacità da parte dei ballerini di eseguirli.
Il sistema specchio codifica dunque gli atti motori compiuti da altri su quelli propri del patrimonio motorio dell’osservatore, e più questo è sviluppato più il sistema diventa efficace. Ma che cosa accade quando a osservare azioni manuali sono persone che non hanno né mani né braccia? Ci dobbiamo aspettare un’assenza di attivazione del sistema specchio, in quanto l’osservatore manca del programma motorio per le azioni manuali, oppure dobbiamo aspettarci un’attivazione di un programma motorio analogo della bocca o del piede che permette di capire lo scopo dell’atto motorio osservato? Per rispondere a interrogativi simili è stato condotto un esperimento di fMRI in cui a due individui aplasici nati senza braccia e mani sono stati presentati filmati di atti motori eseguiti con la mano. I risultati dello studio hanno messo in luce un’attivazione del circuito specchio parieto-frontale dimostrando in tal modo che atti motori non appartenenti al repertorio motorio di un determinato individuo in termini di effettori vengono riconosciuti dall’osservatore grazie al fatto che lo scopo dell’atto motorio osservato recluta programmi motori analoghi eseguiti da altri effettori (per es., bocca, piede).
Sistema specchio e imitazione
Quando osserviamo gli altri, oltre a capire cosa fanno, siamo spesso in grado di ripetere immediatamente la loro azione o di apprendere nuovi comportamenti motori. Il meccanismo specchio con la sua capacità di trasformare immediatamente un atto motorio descritto in formato visivo in un atto motorio potenziale rappresenta un meccanismo neurale ideale per l’imitazione. Imitazione è un termine che ha diversi significati, due dei quali sono particolarmente rilevanti. Il primo, usato soprattutto in psicologia, definisce imitazione la capacità di replicare immediatamente un atto motorio noto compiuto da un altro; il secondo, proprio degli etologi, definisce imitazione la capacità di compiere azioni nuove dopo averle viste fare da un altro individuo, eseguendole nella stessa maniera in cui questi le compie.
Vi sono convincenti prove sperimentali, basate su studi di risonanza funzionale, che il sistema specchio è coinvolto sia nella ripetizione immediata di azioni fatte da altri sia nell’apprendimento di nuovi pattern motori. Vi è, però, un’importante differenza tra i due casi. La ripetizione immediata di un’azione osservata attiva il sistema specchio, senza il coinvolgimento di aree corticali di ordine superiore. Gli atti motori osservati reclutano direttamente i corrispondenti atti motori potenziali nell’osservatore, che è così in grado di imitarli.
L’apprendimento per imitazione avviene, invece, mediante un meccanismo più complesso. Durante l’apprendimento per imitazione oltre al sistema specchio si attiva, infatti, anche il lobo prefrontale e, in particolare, l’area 46, nota per essere coinvolta nella memoria di lavoro. Il meccanismo di base dell’apprendimento per imitazione sarebbe il seguente. Il sistema specchio trasforma gli atti motori elementari osservati da un formato visivo in un formato motorio. Gli atti motori codificati in questo formato raggiungono il lobo prefrontale che li combina nel pattern motorio desiderato. Questo nuovo pattern motorio viene ritrasmesso al sistema specchio per la sua esecuzione (Fabbri-Destro, Rizzolatti 2008).
Capire le intenzioni degli altri
Esperimenti nella scimmia
Abbiamo parlato di comprensione di atti motori tralasciando volutamente il termine azione. Volendo definire questo termine, potremmo dire che per azione si intende la manifestazione esterna di un’intenzione ad agire internamente generata. Tradizionalmente, il problema dell’intenzionalità, di come questa si generi e come si traduca in movimenti, è stato trattato in ambito filosofico. Di recente, però, alcuni suoi aspetti sono diventati materia di indagine della neurofisiologia, per quanto riguarda sia le intenzioni di colui che agisce sia la comprensione dell’intenzione che sottende le azioni altrui.
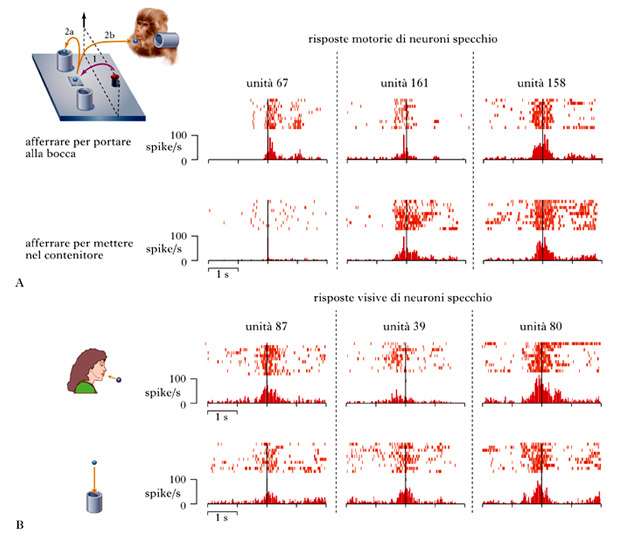
Un problema studiato in questi anni è stato quello di capire se l’intenzione alla base di un’azione (come, per es., afferrare del cibo per mangiarlo) è presente a livello neurale fin dal primo atto motorio (afferrare) di questa azione. A tal fine sono stati condotti esperimenti su scimmie condizionate a prendere del cibo con due scopi diversi: in un caso la scimmia afferrava il cibo per metterlo in un contenitore, nell’altro per mangiarlo. Nella fig. 3A è rappresentato schematicamente il paradigma sperimentale: la scimmia afferra l’oggetto (1) e, a seconda della condizione sperimentale, o lo mette in un contenitore posto sul pianetto (2a) o lo porta alla bocca (2b). In alcune prove mette l’oggetto nel contenitore posto sulla sua spalla. Sono state scelte queste due azioni in quanto implicavano atti motori iniziali (raggiungere e afferrare) identici, e differivano soltanto nello scopo finale. L’ipotesi da verificare era se le diverse intenzioni che sottendono le due azioni si manifestassero nel sistema motorio già all’inizio dell’azione, quando la scimmia compiva gli atti motori iniziali comuni. L’ipotesi alternativa era che l’attività neurale responsabile di questi atti non fosse influenzata dallo scopo finale dell’azione. Sono stati quindi registrati nel lobulo parietale inferiore (LPI) singoli neuroni che codificano l’atto motorio dell’afferrare, e la loro attività è stata studiata nelle due condizioni sopra descritte. Dall’analisi dei risultati è emerso che circa due terzi dei neuroni studiati si attivavano con intensità diversa secondo lo scopo finale dell’azione. Alcuni di questi codificavano l’afferrare per portare alla bocca, altri l’afferrare per mettere nel contenitore. Nella fig. 3A sono rappresentate le risposte di tre neuroni motori registrati durante le condizioni di ‘afferrare per portare alla bocca’ (grasp to eat) e ‘dell’afferrare per mettere nel contenitore’ (grasp to place). Come si può vedere, l’unità 67 si attiva durante l’azione ‘afferrare per portare alla bocca’ ma non durante l’azione ‘afferrare per mettere nel contenitore’; l’unità 161 presenta un comportamento inverso; l’unità 158 si attiva durante entrambe le azioni. La scarica del neurone è allineata con l’istante in cui la scimmia tocca l’oggetto da afferrare.
Appare così chiaro che nell’LPI esistono neuroni (neuroni azione-specifici) che codificano gli atti motori in base alle intenzioni dell’agente. Come mai vi è questa organizzazione? Il vantaggio è evidente. I neuroni azione-specifici di un certo tipo sono connessi con altri neuroni che, quando attivati, portano all’esecuzione di una determinata azione. Avviene quindi che, quando i neuroni azione-specifici scaricano, reclutano automaticamente l’intera catena neuronale necessaria per l’esecuzione dell’azione: di conseguenza non devono ogni volta essere assemblati ex novo i neuroni necessari per compiere un’azione. Tale organizzazione permette una fluida esecuzione dell’azione desiderata (Fogassi, Ferrari, Gesierich et al. 2005).
Molti dei neuroni dell’afferrare dell’LPI si attivano anche quando la scimmia osserva lo sperimentatore eseguire un atto motorio: sono cioè neuroni specchio. Ci si è chiesti, quindi, se i neuroni azione-specifici dal punto di vista motorio fossero anche azione-specifici per l’osservazione. A tale scopo sono stati registrati neuroni dell’LPI e sono state studiate le loro proprietà nelle stesse condizioni dello studio precedente. Questa volta, però, la scimmia non era attiva, ma osservava lo sperimentatore prendere un oggetto o del cibo per metterlo in un contenitore oppure per portarlo alla bocca.
I risultati mostrano che la maggior parte dei neuroni studiati si attiva in modo diverso a seconda che l’atto motorio dell’afferrare, compiuto dallo sperimentatore, appartenga a una o all’altra azione. Nella fig. 3B sono mostrate le risposte visive dei neuroni specchio parietali registrati mentre la scimmia osserva lo sperimentatore afferrare il cibo per mangiarlo o per riporlo nel contenitore. Anche nella condizione ‘osservazione’, la risposta del neurone è selettiva per le due azioni: l’unità 87 risponde alla visione dello sperimentatore che afferra il cibo per portarlo alla bocca, l’unità 39 presenta un comportamento opposto, mentre l’unità 80 risponde ugualmente all’osservazione delle due azioni. La scarica del neurone è allineata con l’istante in cui lo sperimentatore tocca l’oggetto da afferrare. Come spiegare questa diversa attivazione? La situazione sperimentale conteneva sempre elementi che permettevano alla scimmia di capire cosa lo sperimentatore avrebbe fatto in seguito. Un elemento che dava informazioni sull’intenzione dello sperimentatore era, per es., l’oggetto su cui egli agiva. Il cibo significava ‘mangiare’, mentre un oggetto non edibile ‘mettere nel contenitore’. Un altro indicatore era rappresentato dalla ripetitività dell’azione: se lo sperimentatore prendeva del cibo e lo metteva più volte nel contenitore, la scimmia poteva supporre che questa fosse l’intenzione dello sperimentatore anche nelle prove successive (Fogassi, Ferrari, Gesierich et al. 2005).
L’aspetto più interessante dell’esperimento non era, però, la ragione per cui i vari neuroni azione-specifici si attivavano selettivamente durante l’osservazione delle azioni altrui, ma il significato funzionale di questa attivazione. Abbiamo visto che la scarica del neurone motorio azione-specifico dell’afferrare facilita l’attività dei neuroni della catena nel quale è inserito, innescandola. Lo stesso avviene quando il medesimo neurone si attiva durante l’osservazione. Le conseguenze però sono diverse. L’osservatore, infatti, grazie all’attivazione della catena ha, nel caso dell’osservazione, una rappresentazione motoria dell’intera azione che l’agente intende fare. In questa maniera può comprendere la sua intenzione e predire il suo comportamento futuro.
Esperimenti nell’uomo
Una prima prova a favore dell’esistenza di un meccanismo di comprensione delle intenzioni altrui basato sul sistema specchio è stata fornita da uno studio di fMRI. In questo esperimento tre serie di filmati sono state presentate a soggetti volontari. Nella prima serie, definita contesto, venivano mostrati alcuni oggetti (una teiera, una tazza, un piatto con alcuni biscotti) sistemati come se una persona dovesse iniziare la colazione o come se l’avesse appena terminata. Nella seconda serie, definita azione, i soggetti vedevano la mano di una persona che afferrava una tazza senza alcun contesto; nella terza e ultima serie, definita intenzione, si vedeva la stessa mano che afferrava la tazza nei contesti prima e dopo la colazione. I contesti suggerivano ai soggetti quale potesse essere l’intenzione dell’agente che aveva afferrato la tazza: bere o sparecchiare la tavola.
Particolarmente interessante è stato il paragone tra le condizioni intenzione e azione: quando i soggetti comprendevano l’intenzione dell’azione osservata vi era un marcato aumento dell’attività sistema specchio e, in particolare, della parte posteriore del giro frontale inferiore di destra. È altresì interessante notare che l’osservazione dei video in cui la tazza era afferrata per bere determinava un’attivazione maggiore rispetto alla condizione in cui la tazza era afferrata per sparecchiare la tavola. Questi risultati sono in accordo con quelli ottenuti nella scimmia in cui vi sono molti più neuroni dedicati all’afferrare per mangiare rispetto a quelli che codificano l’afferrare per mettere nel contenitore (Rizzolatti, Sinigaglia 2006).
Un secondo esperimento di fMRI, basato sulla tecnica di inibizione da stimoli ripetuti, ha dimostrato che il sistema specchio dell’emisfero di destra si attiva quando il soggetto prevede il risultato di un’azione come quella di aprire una scatolina indipendentemente dal modo in cui tale apertura viene eseguita. Questi dati sono stati interpretati come un’ulteriore prova a favore del fatto che il sistema specchio dell’emisfero destro rivesta un ruolo centrale nel processo di comprensione delle intenzioni alla base delle azioni degli altri (Hamilton, Grafton 2008).
Per approfondire il ruolo del sistema specchio nella comprensione delle intenzioni, è stato recentemente condotto un esperimento di fMRI in cui si è indagata l’attivazione corticale in azioni intenzionali rispetto ad altre in cui lo scopo desiderato sfugge per cause esterne a colui che compie l’azione. Ai soggetti sono stati presentati filmati di azioni fatte con differenti effettori, ognuno in doppia versione: uno nel quale l’attore raggiungeva lo scopo dell’azione (azione intenzionale; per es., afferrava un pezzo del gioco degli scacchi), l’altro in cui l’attore eseguiva un’azione simile ma senza raggiungere lo scopo (azione non intenzionale; per es., muoveva la mano per prendere un pezzo degli scacchi, ma inavvertitamente buttava giù alcuni pezzi). I risultati hanno mostrato che entrambi i tipi di azione attivavano il sistema specchio con la differenza che le azioni non intenzionali attivavano anche le aree ‘attenzionali’, quelle cioè che si attivano quando l’attenzione dell’individuo è attratta da uno stimolo. Questo significa che quando un individuo osserva un’azione non intenzionale, il cervello descrive l’evento senza un aumento di attivazione del sistema specchio, come invece accade nel caso in cui il soggetto deve capire le intenzioni degli altri. La stranezza dell’evento viene segnalata mediante una maggiore attivazione delle aree attenzionali.
In conclusione, i dati dell’uomo, come quelli della scimmia, mostrano che l’intenzione che sottende l’azione eseguita da altri è compresa grazie al sistema specchio. Questo, ovviamente, non implica che il meccanismo specchio sia l’unico che ci permette di capire le intenzioni degli altri: esistono anche altri meccanismi che ci fanno raggiungere il medesimo obiettivo mediante, per es., ragionamenti inferenziali. Tuttavia, quello che il sistema specchio offre è una conoscenza diversa, esperienziale, basata sull’attività di quei circuiti che adoperiamo quando siamo noi stessi a eseguire l’azione osservata in prima persona.
Sistema specchio e linguaggio
La scoperta del sistema specchio ha suscitato interesse e discussioni anche tra gli studiosi del linguaggio, in particolare in relazione al problema dell’evoluzione del linguaggio. Fin dai primi studi di brain imaging sul sistema specchio si è visto che l’osservazione di atti motori manuali attiva la parte posteriore del giro frontale inferiore dell’emisfero di sinistra, cioè la regione dove si trova l’area di Broca, universalmente nota come un’area del linguaggio che riveste un ruolo importante nella produzione dello stesso. La sua attivazione, durante l’osservazione di azioni manuali, ha suscitato molta sorpresa. Tale attivazione, tuttavia, non è poi così incredibile se si considera, come molti sostengono, che il linguaggio umano non si è evoluto dalle vocalizzazioni, dalle grida degli animali, ma dalla comunicazione gestuale, simile a quella che si può osservare nei primati superiori non umani oggi esistenti (Corballis 2002). Le vocalizzazioni degli animali, come anche certi tipi di vocalizzazione umana, avrebbero come loro ruolo funzionale quello di informare sullo stato emotivo di chi le emette, ma scarse capacità, proprio per la loro intrinseca connotazione emozionale, di rappresentare il mondo esterno. Questo tipo di capacità di comunicare (la cosiddetta capacità referenziale), fondamentale nella comunicazione umana, deriverebbe invece dalla comunicazione gestuale. Soltanto più tardi nell’evoluzione suoni specifici si sarebbero associati ai gesti, portando infine allo sviluppo del linguaggio parlato.
Un altro motivo per cui la scoperta del sistema specchio ha suscitato enorme interesse è dovuto al fatto che essa permette di proporre un meccanismo in grado di chiarire un aspetto fondamentale della comunicazione, quello che riguarda la creazione di un legame diretto tra colui che manda un messaggio (mittente) e colui che lo riceve (destinatario). Un problema che ogni teoria della comunicazione deve risolvere è infatti quello della cosiddetta parità tra il mittente e il destinatario, spiegando come un segnale arbitrario emesso da un certo individuo possa assumere un preciso significato per il suo destinatario. Ciò vale sia per il linguaggio basato sui suoni sia per quello basato sui gesti. Ora, il meccanismo specchio consente di risolvere tale problema dal momento che esso ‘trasforma’ un’azione compiuta da un individuo in una rappresentazione di quella stessa azione nella corteccia motoria di chi la osserva. In altri termini, crea un legame diretto, non arbitrario, tra i due individui. Le azioni compiute da un individuo diventano messaggi che l’osservatore comprende senza che vi sia alcun accordo preliminare tra di loro, cosa che tra l’altro implicherebbe un ricorso all’infinito.
Un ulteriore aspetto interessante del sistema specchio riguarda il modo in cui riusciamo a decifrare i suoni linguistici. Come già ricordato, Liberman ha ipotizzato che la nostra capacità di percepire i fonemi non si basa sul loro aspetto acustico, ma sul fatto che essi sono in grado di evocare nell’ascoltatore un pattern motorio simile a quello che serve per emetterli. Ciò spiega perché riusciamo a percepire e considerare come identici, in significato, fonemi acusticamente molto diversi tra loro. Basti pensare a parole pronunciate con voce bassa o acuta, rauca o sintetizzata artificialmente. La teoria di Liberman, nota come teoria motoria del linguaggio, prevede che l’uomo possegga un meccanismo che trasforma i fonemi uditi nel loro equivalente motorio.
Questa ipotesi è stata saggiata sperimentalmente in uno studio di TMS. Gli autori hanno registrato i potenziali motori evocati (MEP, Motor Evoked Potential) dei muscoli della lingua in soggetti cui era stato chiesto di ascoltare stimoli acustici. Questi erano costituiti da parole o da pseudoparole contenenti una doppia f (baffo) o una doppia r (birra) e suoni bitonali. La f è una consonante labiodentale e non richiede, per essere pronunciata, un particolare coinvolgimento della lingua, mentre la r è una consonante linguopalatale e la sua pronuncia comporta un marcato coinvolgimento di tale organo. I risultati dell’esperimento hanno mostrato che l’ascolto delle parole e delle pseudoparole contenenti la doppia r determinava un significativo aumento dei MEP registrati dai muscoli della lingua rispetto al caso dei suoni bitonali e delle parole e delle pseudoparole contenenti la doppia f (Rizzolatti, Sinigaglia 2006).
Questi dati mostrano che nell’uomo esiste un sistema specchio ‘dedicato’ per i suoni del linguaggio (echo mirror system): quando un individuo ascolta stimoli verbali vi è un’automatica attivazione dei suoi centri motori responsabile dell’emissione dei fonemi presenti nelle parole ascoltate. Possedendo la copia motoria del fonema ascoltato l’ascoltatore lo distingue e percepisce.
Sistema specchio ed emozioni
Fino a questo punto abbiamo descritto meccanismi attraverso i quali comprendiamo le azioni altrui prive di un chiaro contenuto emozionale, le cosiddette azioni fredde. Nella vita di tutti i giorni, però, siamo chiamati a capire anche cosa provano gli altri e a inferire, dal loro comportamento, le loro emozioni. Dunque, le questioni su cui i ricercatori si sono concentrati sono state quelle di definire le strutture cerebrali che mediano la comprensione delle emozioni degli altri e di stabilire se il meccanismo specchio giochi un ruolo anche nel riconoscimento delle emozioni.
È ragionevole assumere che, come per la comprensione delle azioni, anche per il riconoscimento delle emozioni esistono due meccanismi distinti: il primo basato su un’elaborazione cognitiva degli aspetti sensoriali del comportamento emotivo; il secondo dipendente da un’attivazione diretta delle strutture coinvolte nella produzione delle emozioni. Si tratta di due meccanismi di riconoscimento delle emozioni radicalmente differenti. Mediante il primo, l’osservatore riconosce l’emozione espressa dall’altro ma non la prova, non la vive, semplicemente la inferisce. Grazie al secondo meccanismo, invece, il riconoscimento dell’emozione si basa su una reale esperienza emozionale analoga a quella di chi la vive e la esprime.
In questi ultimi anni, una serie di esperimenti è stata condotta allo scopo di verificare se le stesse strutture corticali che si attivano quando un individuo prova un’emozione si attivino anche quando questi osserva un’emozione simile in un’altra persona. Due sono state le emozioni particolarmente studiate: il disgusto e il dolore.
Il disgusto, pur non essendo riconosciuto da tutti come un’emozione di base, appare presente negli esseri umani indipendentemente da sesso, razza e classe sociale, tanto che già Charles R. Darwin lo considerava una delle emozioni fondamentali della specie umana. Studi di fMRI hanno mostrato che i centri nervosi che si attivano maggiormente quando una persona prova disgusto sono l’insula, l’amigdala e la corteccia del cingolo anteriore.
Ma cosa avviene quando noi osserviamo una persona che esprime disgusto? Quali aree si attivano? Esiste un meccanismo specchio per il disgusto? Per rispondere a questi quesiti, è stato condotto un esperimento di fMRI che si basava su due condizioni sperimentali. Nella prima, ai soggetti era chiesto di annusare odori piacevoli, neutri o disgustosi, mentre nella seconda essi dovevano osservare l’espressione facciale di individui che, avendo annusato le stesse sostanze, esprimevano le emozioni relative. In accordo con i dati precedenti si è trovato che la stimolazione olfattiva con stimoli disgustosi (odore di uova marce) provocava attivazioni principalmente nell’insula, nell’amigdala e nella corteccia del cingolo anteriore. La stimolazione visiva con facce che esprimevano disgusto attivava varie aree corticali visive, l’insula anteriore di sinistra (ma non l’amigdala) e il cingolo anteriore. Il risultato più importante dell’esperimento è stato che le attivazioni dovute alla stimolazione con stimoli olfattivi disgustosi e quelle dovute a espressioni facciali che esprimevano disgusto evidenziavano una localizzazione identica nell’insula anteriore e nel cingolo. Appare, quindi, che tanto l’insula quanto il giro del cingolo contengono neuroni che si attivano sia durante l’esposizione a un odore disgustoso sia durante l’osservazione del disgusto in altri. Queste strutture appaiono dotate di un meccanismo specchio che trasforma stimoli visivi con contenuto emotivo in pattern viscero-motori tipici dell’emozione osservata (Gallese, Keysers, Rizzolatti 2004).
Oltre che con il disgusto, attivazioni nell’insula anteriore e nella corteccia anteriore del cingolo sono state ottenute studiando soggetti che provavano dolore o immaginavano che un’altra persona provasse un dolore simile. Come lo studio sul disgusto anche questo studio di fMRI si componeva di due condizioni sperimentali. Nella prima si applicava alla mano dei soggetti (tutti di sesso femminile) uno stimolo elettrico leggermente doloroso mediante due piccoli elettrodi; nella seconda, i soggetti vedevano gli stessi elettrodi applicati alla mano del loro fidanzato. Ai soggetti era detto che il fidanzato avrebbe ricevuto lo stesso stimolo doloroso che loro avevano sentito. I risultati hanno mostrato che gli stessi siti dell’insula anteriore e della corteccia del cingolo che si attivavano durante la sensazione dolorosa, si attivavano anche quando i soggetti pensavano che i loro fidanzati provassero dolore (Sing-er, Seymour, O’Doherty et al. 2004). Come il disgusto anche il riconoscimento del dolore in altri è legato all’attivazione delle strutture che entrano in funzione quando è l’individuo che osserva a provare dolore in prima persona. Il riconoscimento del dolore altrui risulta quindi mediato da un meccanismo specchio.
L’ipotesi che il riconoscimento delle emozioni degli altri avvenga attraverso l’attivazione delle stesse strutture che si attivano quando noi proviamo quelle emozioni era già stata avanzata da Antonio R. Damasio e dai suoi collaboratori prima degli esperimenti appena descritti (Damasio 2003). Sulla base di studi neuro-psicologici essi avevano ipotizzato che il riconoscimento delle emozioni altrui dipendesse dall’attivazione di un circuito che codifica le emozioni degli altri come se fossero proprie (as if loop). In particolare, Damasio e i suoi collaboratori attribuivano un ruolo fondamentale nell’as if loop, oltre che all’insula, alle aree somatosensoriali, suggerendo che il riconoscimento delle emozioni altrui necessitasse dell’attivazione delle aree corticali in cui il corpo è rappresentato. Sebbene un contributo sensoriale alla comprensione dell’emozione sia certamente possibile, l’attivazione dell’insula rostrale e, di contro, l’assenza di attivazione nella corteccia somatosensoriale primaria quando si prova dolore oppure disgusto indica che il ruolo fondamentale nel riconoscimento delle emozioni è svolto dai sistemi viscero-motori e non dalle aree sensoriali.
Il meccanismo specchio e l’autismo
L’autismo è una sindrome dello sviluppo del bambino caratterizzata da un comportamento sociale atipico, da disturbi della comunicazione, sia verbale sia non verbale, e da comportamenti ripetitivi. Alcuni anni fa è stata avanzata l’ipotesi che alla base dell’incapacità dei bambini con autismo di entrare in relazione con gli altri (di capire gli altri) vi fosse un malfunzionamento del sistema specchio. Dati sperimentali sia neurofisiologici (Ramachandran, Oberman 2006) sia di brain imaging (Iacoboni, Dapretto 2006) hanno dimostrato che effettivamente nei soggetti affetti da autismo vi è un deficit di attivazione del sistema specchio rispetto a soggetti con sviluppo tipico durante l’osservazione di azioni compiute da altri.
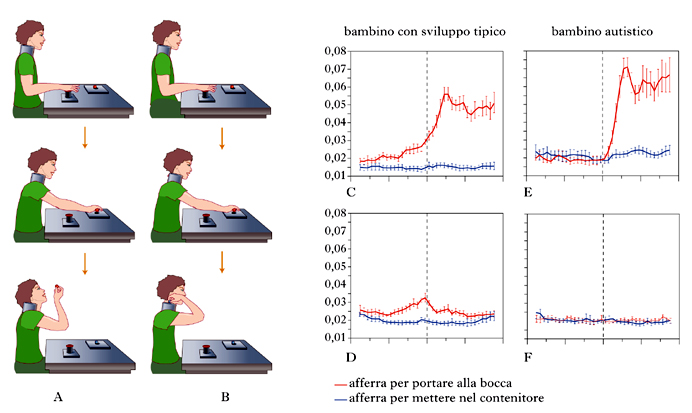
Recenti studi hanno esaminato come l’intenzione di chi agisce sia compresa da bambini con sviluppo tipico e da bambini con autismo. Abbiamo già ricordato come il sistema dei neuroni specchio sia organizzato in catene motorie che permettono all’individuo di trasformare le sue intenzioni in azioni. Si è studiata quindi l’attività di queste catene nei bambini con sviluppo tipico e nei bambini con autismo durante l’esecuzione di azioni e durante l’osservazione delle stesse azioni compiute da altri (fig. 4). A tale scopo è stato usato un paradigma sperimentale simile a quello impiegato nella scimmia: ai bambini dei due gruppi è stato chiesto di afferrare un pezzettino di cibo e portarlo alla bocca (fig. 4A, dall’alto in basso) o prendere un pezzettino di carta e metterlo in un contenitore (fig. 4B). Nella condizione ‘osservazione’, ai bambini era richiesto di osservare lo sperimentatore che eseguiva le medesime due azioni. Mentre poi il bambino effettuava queste prove veniva registrata l’attività elettromiografica del muscolo miloioideo, coinvolto nell’apertura della bocca.
Si è visto che nei bambini a sviluppo tipico il muscolo miloioideo presentava un incremento di attività nella condizione ‘portare alla bocca’ già durante la fase di raggiungimento, ovvero ben prima che i soggetti toccassero il cibo. Di contro, il muscolo rimaneva del tutto silente nella condizione sperimentale che prevedeva il mettere un pezzettino di carta nel contenitore (fig. 4C). Come interpretare questa anticipata attivazione del miloioideo? La risposta è semplice. Il muscolo miloioideo si attiva con anticipo perché l’intenzione alla base del portare alla bocca è già presente nelle fasi iniziali dell’azione (raggiungimento del cibo), e quest’intenzione recluta fin dall’inizio dell’azione gli atti motori necessari per soddisfarla. Il passo successivo è stato quello di vedere se la stessa catena motoria che si attiva in esecuzione si attiva anche durante l’osservazione delle due azioni sopra descritte. Gli esperimenti sui bambini a sviluppo tipico hanno dimostrato che effettivamente accade questo. Durante l’osservazione dell’azione ‘mangiare’ vi è un’evidente attivazione del miloioideo, mentre questa è assente nella condizione mettere nel contenitore (fig. 4D). In altre parole, quando il bambino osserva lo sperimentatore portare il cibo alla bocca, replica l’azione osservata e la rivive, per così dire, al suo interno (Cattaneo, Fabbri-Destro, Boria et al. 2007).
Lo stesso esperimento è stato eseguito anche su un gruppo di bambini con autismo ‘ad alto funzionamento’ (con quoziente di intelligenza nella norma). I risultati hanno mostrato che, a differenza dei bambini con sviluppo tipico, i bambini con autismo non mostravano alcuna attività elettromiografica nel muscolo miloioideo durante la fase di raggiungimento e afferramento del cibo. Il muscolo si attivava solo quando il bambino iniziava a sollevare il cibo per portarlo alla bocca (fig. 4E). Evidente era anche il deficit durante l’osservazione del portare il cibo alla bocca. In questa condizione il muscolo miloioideo nei bambini con autismo rimaneva sempre silente (fig. 4F).
Come interpretare questi dati? È chiaro che l’osservazione dell’azione eseguita da un altro non riesce ad attivare nei bambini con autismo quella copia motoria che compare nei bambini con sviluppo tipico. Questo non significa, però, che il bambino con autismo non capisca cosa fanno gli altri. Anzi, se interrogato, può rispondere correttamente. Come spiegare, dunque, questo paradosso?
In un recente esperimento sono state fatte vedere a bambini con sviluppo tipico alcune fotografie in cui una mano toccava un oggetto o lo afferrava. Gli oggetti potevano essere afferrati in due maniere diverse e il modo in cui ciò avveniva permetteva di capire perché l’agente afferrava l’oggetto. Era chiesto quindi ai bambini di spiegare cosa vedevano (toccare o afferrare) e nel caso dell’afferrare il perché dell’atto motorio. Per es., nel caso di una tazzina dovevano prima dire se la mano la toccava o la afferrava e poi, a seconda dell’atto motorio (prensione per il manico o prensione dall’alto), specificare l’intenzione dell’agente (per bere o per spostare, rispettivamente). Sia i bambini a sviluppo tipico sia quelli con autismo distinguevano virtualmente senza errori il toccare dall’afferrare (l’atto motorio). Tuttavia, solo i bambini a sviluppo tipico spiegavano correttamente il perché dell’azione. I bambini con autismo davano anche loro una spiegazione, ma questa poggiava non sull’atto motorio osservato, ma sul riconoscimento della funzione d’uso più comune dell’oggetto di volta in volta in questione. Per riprendere il nostro esempio, la tazzina era per loro sempre ‘presa per bere’, indipendentemente dal tipo di prensione. L’azione degli altri non era codificata in base all’atto motorio osservato, ma in base all’uso comune dell’oggetto osservato. Quello che i bambini con autismo, quindi, non risultano in grado di fare è interpretare le azioni degli altri attivando il proprio pattern motorio corrispondente: il loro mondo, infatti, è fatto di cose e non di esseri che agiscono come loro.
Tornando all’esperimento elettromiografico, vi è un importante dato che occorre sottolineare: la difficoltà di reclutamento del sistema motorio nei bambini durante l’esecuzione delle azioni. Questa difficoltà cambia la prospettiva di come interpretare la causa fisiologica primaria del deficit autistico. Questa sembra risiedere, prima ancora che nell’incapacità di ‘rispecchiare’ le azioni degli altri, nell’incapacità di organizzare il proprio comportamento intenzionale. Mancando una normale organizzazione delle catene motorie, anche la comprensione delle intenzioni degli altri basata sull’attivazione di queste catene (comprensione esperienziale) diventa deficitaria.
Conclusioni
Il sistema dei neuroni specchio è stato scoperto alla fine del 20° secolo. Oggi sappiamo molto sulla sua organizzazione anatomica e su come esso funziona. Esistono tuttavia varie lacune, la più importante delle quali riguarda il suo sviluppo. I sistemi specchio sono innati? E se sì, entro che limiti? E ancora, è innato tanto il sistema specchio responsabile della comprensione delle emozioni quanto quello alla base della comprensione delle azioni ‘fredde’? Infine, nell’ambito di queste ultime, il sistema specchio ha la stessa origine per i movimenti della mano, per i quali è facile postulare l’apprendimento, e per i movimenti della bocca e della faccia, per i quali invece un simile assunto appare molto più difficile?
Si sa poco, inoltre, sull’esistenza di un eventuale sistema specchio sottocorticale che potrebbe mediare certi fenomeni di imitazione immediata già presenti alla nascita, quando la corteccia cerebrale è ancora immatura, come, per es., l’imitazione della protrusione della lingua. Un meccanismo specchio sottocorticale potrebbe essere alla base anche di veri e propri fenomeni di contagio, come lo sbadiglio.
Un altro interessante campo d’indagine che certamente si svilupperà nei prossimi anni è quello della possibile presenza del meccanismo specchio in specie animali diverse dai primati. Di recente la presenza del meccanismo specchio è stata dimostrata negli uccelli e, più precisamente, nel passero di palude. Nel telencefalo di questa specie esistono infatti neuroni che rispondono in modo temporalmente preciso alla presentazione uditiva di certe sequenze di suoni e quando l’uccello emette la medesima sequenza. Se si interrompe il feedback uditivo il pattern di attività non cambia, indicandone la loro natura essenzialmente motoria.
Importante sarebbe anche dimostrare l’esistenza del meccanismo specchio in specie, quali, per es., i roditori, comunemente usate nella sperimentazione animale. La dimostrazione dell’esistenza del meccanismo specchio in queste specie avrebbe un interesse non solo concettuale ma anche medico-applicativo, permettendo di affrontare il problema della specificità istologica e chimica del sistema specchio. A loro volta queste conoscenze potrebbero creare la possibilità di avere un modello animale per studiare a livello cellulare e molecolare i disturbi causati da un deficit di funzionamento del sistema specchio.
Bibliografia
G. Rizzolatti, G. Luppino, The cortical motor system, «Neuron», 2001, 31, 6, pp. 889-901.
M.C. Corballis, From hand to mouth. The origins of language, Princeton (N.J.) 2002 (trad. it. Milano 2008).
A.R. Damasio, Looking for Spinoza, Orlando (Flo.)-London 2003.
V. Gallese, Ch. Keysers, G. Rizzolatti, A unifying view of the basis of social cognition, «Trends in cognitive sciences», 2004, 8, 9, pp. 396-403.
G. Rizzolatti, L. Craighero, The mirror-neuron system, «Annual review of neuroscience», 2004, 27, pp. 169-92.
T. Singer, B. Seymour, J. O’Doherty et al., Empathy for pain involves the affective but not sensory components of pain, «Science», 2004, 303, 5661, pp. 1157-62.
L. Fogassi, P.F. Ferrari, B. Gesierich et al., Parietal lobe. From action organization to intention understanding, «Science», 2005, 308, 5722, pp. 662-67.
M. Iacoboni, M. Dapretto, The mirror neuron system and the consequences of its dysfunction, «Nature reviews. Neuroscience», 2006, 7, pp. 942-51.
V.S. Ramachandran, L.M. Oberman, Broken mirrors: a theory of autism, «Scientific American», 2006, 295, pp. 62-69.
G. Rizzolatti, C. Sinigaglia, So quel che fai. Il cervello che agisce e i neuroni specchio, Milano 2006.
L. Cattaneo, M. Fabbri-Destro, S. Boria et al., Impairment of actions chains in autism and its possible role in intention understanding, «PNAS. Proceedings of the National academy of sciences of the USA», 2007, 104, 45, pp. 17.825-30.
F. Filimon, J.D. Nelson, D.J. Hagler, M.I. Sereno, Human cortical representations for reaching: mirror neurons for execution, observation, and imagery, «NeuroImage», 2007, 37, 4, pp. 1315-28.
D. Tkach, J. Reimer, N.G. Hatsopoulos, Congruent activity during action and action observation in motor cortex, «The journal of neuroscience», 2007, 27, 48, pp. 13.241-50.
M. Fabbri-Destro, G. Rizzolatti, The mirror system in monkeys and humans, «Physiology», 2008, 23, pp. 171-79.
A.F. de C. Hamilton, S.T. Grafton, Action outcomes are represented in human inferior frontoparietal cortex, «Cerebral cortex», 2008, 18, 5, pp. 1160-68.