
Oceanografia
Oceanografia
Oceanografia biologica, di Emer Steemann Nielsen
Oceanografia fisica, di Henry Charnock
Oceanografia biologica
SOMMARIO: 1. Introduzione. □ 2. L'oceanografia biologica all'inizio del secolo: a) ricerche quantitative sul plancton; b) ricerche sui processi di base della produzione oceanica. □ 3. Opinioni attuali sulla produzione oceanica: a) introduzione; b) riserva costante di alghe planc toniche e produzione primaria; c) i fattori che regolano la produzione primaria; d) alcuni tipi di produzione oceanica primaria; e) produzione globale di vegetali planctonici e bentonici. □ 4. Riserva costante di zooplancton e catene alimentari nell'oceano. □ 5. Gli animali del fondo marino. □ 6. Regioni faunistiche e masse d'acqua: a) plancton; b) gli animali dei fondi oceanici. □ 7. Batteriologia oceanica. □ 8. Conclusione. □ Bibliografia.
1. Introduzione.
Il termine ‛oceanografia biologica' è recente: in quasi tutti i paesi dell'Europa occidentale è entrato nell'uso comune soltanto negli ultimi quindici anni. Secondo alcuni scienziati l'oceanografia biologica studia tutti gli aspetti della biologia degli organismi marini, ma questa accezione così ampia non è pratica. È opportuno invece distinguere tra oceanografia biologica e biologia marina: anche se non è possibile stabilire confini netti tra le due discipline, si può dire che nella prima si studia in particolare l'oceano, mentre nella seconda si tiene conto degli organismi in tutta la loro varietà. Una distinzione analoga si fa tra limnologia e biologia delle acque dolci.
L'oceanografia biologica opera in stretta collaborazione sia con l'oceanografia fisica, sia con l'oceanografia chimica, con la quale si è sviluppata di pari passo.
Una breve rassegna dello sviluppo dell'oceanografia biologica in questo secolo non può evitare un certo grado di soggettività. Non è possibile menzionare tutti i fatti importanti né tutti gli autori che hanno contribuito a far progredire questa scienza. In questo articolo è stato necessario limitarsi all'oceano vero e proprio e non si è potuto dare notizia dei problemi particolari relativi alle zone costiere, come estuari, ecc.
2. L'oceanografia biologica all'inizio del secolo.
a) Ricerche quantitative sul plancton.
Durante la prima parte del sec. XIX gli zoologi intrapresero intense ricerche sugli animali costieri, mentre le ricerche sugli oceani aperti incominciarono negli ultimi decenni, quando si costruirono molti strumenti di campionamento per catturare gli animali e i vegetali sia pelagici sia di fondo, e furono fatte numerose e importanti scoperte fondamentali per la conoscenza degli organismi oceanici.
Nello stesso periodo nacque la scienza che oggi chiamiamo oceanografia biologica, di cui è stato pioniere lo scienziato tedesco V. Hensen. Egli comprese chiaramente che i vegetali marini, sia di mare aperto sia costieri, sono i soli produttori di sostanza organica e che il punto di partenza per rendersi conto della produzione di tutti gli organismi negli oceani era la misurazione della quantità dei vegetali. Poiché i soli vegetali autotrofi dell'oceano aperto sono praticamente le alghe planctoniche, ossia le alghe microscopiche galleggianti negli strati d'acqua superficiali, il primo compito di Hensen fu quello di costruire reti adatte a raccogliere le alghe e a misurarne la distribuzione a determinati livelli di profondità.
Per ottenere informazioni significative con questo metodo è necessario che si realizzino due condizioni: a) che siano catturati tutti gli organismi; b) che le alghe planctoniche siano equamente distribuite negli oceani, così che ogni cattura possa essere considerata rappresentativa per una data area. Come vedremo, la prima condizione non fu affatto realizzata da Hensen, mentre della seconda si ebbero, nella maggior parte dei casi, prove positive anche se in proposito furono avanzati dubbi da alcuni scienziati nei primi decenni del secolo. Oggi la critica relativa all'uniformità della distribuzione quantitativa delle alghe planctoniche negli oceani non è ritenuta più valida, in linea di massima, ma quando si scende nei particolari si possono trovare distribuzioni ineguali dovute al caso. Il fatto che la distribuzione sia per lo più molto regolare è ancora un enigma.
L'apparecchio costruito da Hensen per misurare il numero delle alghe planctoniche a una determinata profondità era un retino verticale che si calava nel mare aperto fino alla profondità di 200 m e si riportava poi lentamente alla superficie. La rete era costituita dalla più fine seta da buratto, con maglie di circa 0,05 mm di diametro. Per garantire la massima efficienza filtrante del retino, la bocca doveva essere piccola in relazione all'area filtrante del tessuto di seta. Ciò nonostante Hensen si accorse che non si poteva ottenere che attraverso il retino passasse tant'acqua quanta era teoricamente prevedibile. Egli riuscì a stabilire empiricamente dei coefficienti di filtrazione per le varie dimensioni dei retini. Attualmente, nei paesi in cui la scienza oceanografica è più avanzata, i retini per le misure quantitative del fitoplancton non si usano più, a causa della loro inefficienza.
Nuove tecniche sono oggi a disposizione. Nei primi anni del sec. XX, un altro scienziato tedesco, H. Lohmann, descrisse una serie di tecniche nuove, in particolare quella della centrifugazione (v. Lohmann, 1908). Egli poté dimostrare che le alghe planctoniche quantitativamente più importanti non erano catturate dal retino di seta. L'uso della centrifugazione per misurare la quantità di alghe planctoniche in un determinato volume di acqua di mare - per es. 100 ml - si basa sul fatto che tali alghe hanno praticamente tutte un peso specifico più elevato di quello dell'acqua. In seguito sono stati elaborati metodi più perfezionati e perciò la tecnica della centrifugazione si usa di rado; essa però è stata per molti anni uno dei metodi più importanti per studiare gli aspetti quantitativi della biologia degli oceani.
Nel 1920 il Lohmann recò un contributo della massima importanza presentando i risultati della Deutschland Expedition, che nel corso di una crociera dall'Europa all'America meridionale (1911) aveva esplorato alcune zone rappresentative dell'Atlantico. Furono osservati fatti molto rilevanti, per esempio la scarsità di alghe nel Mare dei Sargassi rispetto alla loro abbondanza nelle aree attorno all'equatore.
Le ricerche del Lohmann furono continuate su scala più vasta dalla spedizione della nave Meteor (1925-1927), che esplorò l'Atlantico meridionale. I dati raccolti, numerosissimi, furono pubblicati da Hentschel (v., 1933-1936). Da allora siamo in possesso di un materiale molto importante relativo alla riserva costante di fitoplancton e zooplancton di piccole dimensioni in tutte le stagioni dell'anno, nella maggior parte delle zone sudatlantiche.
Fino a dopo la seconda guerra mondiale, gli scienziati britannici continuarono a usare con molta frequenza la tecnica del retino, nonostante la sua scarsa attendibilità, mentre altri, in particolare i biologi scandinavi e statunitensi, già nel periodo fra le due guerre mondiali avevano proseguito col metodo di Lohmann le ricerche sulla riserva costante di fitoplancton nella parte più settentrionale dell'Atlantico del Nord. A poco a poco si introdusse una nuova tecnica, basata sulla sedimentazione in cilindri di piccole dimensioni, muniti sul fondo di un sottilissimo vetrino che permette di contare le alghe sedimentate usando un microscopio invertito, cioè un microscopio con l'obiettivo rivolto verso l'alto: questa tecnica fu elaborata da H. Utermöhl (v., 1932).
b) Ricerche sui processi di base della produzione oceanica.
Verso la fine del secolo scorso erano stati raccolti molti dati sulla distribuzione quantitativa del plancton e dei pesci negli oceani: da essi si pervenne a una generalizzazione, accettata dalla maggioranza degli oceanografi di quel tempo, secondo cui le parti fredde degli oceani sono ricche di organismi, mentre le acque calde sono più povere. Come vedremo più avanti, la moderna oceanografia ha dimostrato che questa generalizzazione non è sempre vera, in quanto regioni ricche di organismi e regioni povere si trovano sia nei mari caldi sia nei mari freddi; essa, tuttavia, contiene qualcosa di vero.
Verso la fine del secolo, lo zoologo tedesco K. Brandt (v., 1899), tentò di dimostrare che l'azoto è un fattore determinante per la produzione oceanica. Egli si riferì alla legge del minimo formulata da J. von Liebig, secondo la quale il fattore presente in quantità minima è, tra i fattori di crescita, quello che determina il ritmo di accrescimento. Brandt sostenne che le sostanze presenti in quantità minima negli oceani sono i nitrati, sebbene, secondo le conoscenze del tempo, grandi quantità di questi sali si sarebbero trasferite dalla terra nel mare, nel corso di lunghi periodi geologici. Secondo Brandt le basse concentrazioni presenti negli oceani si dovevano attribuire all'attività dei batteri denitrificanti: poiché il metabolismo di questi batteri - come quello di tutti i Batteri - aumenta con l'aumentare della temperatura, secondo la teoria di Brandt nelle zone più calde degli oceani si dovrebbe riscontrare una minor quantità di nitrati. Molti scienziati, però, misero in dubbio l'esattezza della teoria, primo e più importante fra tutti un altro autore tedesco, A. Nathansohn, che cercò invano di reperire batteri denitrificanti nelle acque del Mediterraneo. Dato poi che i batteri denitrificanti utilizzano l'ossigeno dei nitrati per il loro metabolismo (ossidazione nel processo respiratorio), si doveva ritenere improbabile che essi si potessero trovare nelle condizioni di aerobiosi degli oceani.
Purtroppo il Nathansohn sostenne anche che la legge del minimo non era adeguata a spiegare la crescita delle alghe planctoniche marine, e una delle ragioni da lui addotte per negare l'importanza dei nitrati come fattore limitante per lo sviluppo delle alghe planctoniche fu il fatto che secondo le analisi imperfette di quel tempo né le concentrazioni dei nitrati, né quelle dei fosfati risultavano particolarmente basse negli strati superficiali del mare. Il Nathansohn propose quindi una teoria secondo cui una produzione considerevole di alghe planctoniche può aver luogo soltanto laddove gli strati d'acqua sottostanti sono portati alla superficie. In tal modo era possibile rendersi ragione della quantità quasi sempre abbondante di fitoplancton sia nell'Artico - per i movimenti verticali (convezione) dell'acqua durante l'inverno - sia in quelle parti dei tropici dove si verifica un moto ascensionale costante dell'acqua situata sotto la zona superficiale e dove proprio in quegli anni erano state osservate forti concentrazioni di plancton.
Il Nathansohn (v., 1906) ipotizzò dapprima un effetto di avvelenamento determinato dai prodotti del metabolismo accumulati nelle masse d'acqua dove per lunghi periodi si era verificata una produzione di alghe planctoniche. Un fenomeno analogo era stato già osservato durante lo sviluppo di colonie batteriche. L'autore tedesco suppose che l'effetto dell'avvelenamento fosse controbilanciato dall'innalzamento delle acque subsuperficiali, nelle quali non vi era stato sviluppo di alghe planctoniche. Le successive critiche all'ipotesi del Nathansohn sembra non abbiano tenuto conto di una sua ulteriore supposizione (v. Nathansohn, 1908), secondo cui gli organismi che si nutrono di fitoplancton si accumulerebbero nelle vecchie acque di superficie, mantenendo, così, bassa la concentrazione delle alghe. Come vedremo più avanti, la scienza oceanografica attuale considera che l'attività alimentare dei consumatori sia uno dei fattori più importanti nella regolazione della produzione di fitoplancton.
Brandt e Nathansohn non trovarono un accordo tra le loro teorie. Come diremo più avanti, lo sviluppo di tecniche specifiche atte a determinare con precisione la concentrazione dei nitrati e dei fosfati nel mare permette agli oceanografi, chimici e biologi, di dimostrare come entrambi i fattori, le concentrazioni di questi sali nutritivi (secondo l'ipotesi di Brandt) e la dinamica del rinnovamento delle acque superficiali (secondo l'ipotesi di Nathansohn), possano, congiuntamente, fornire la spiegazione principale - ma non la sola - dei processi di base della produzione oceanica.
3. Opinioni attuali sulla produzione oceanica.
a) Introduzione.
Dopo aver passato brevemente in rassegna lo sviluppo dell'oceanografia biologica durante i due primi decenni del secolo, non esporremo più i fatti e le teorie secondo un rigoroso ordine storico, ma passeremo a discutere lo stato attuale delle conoscenze. Sarebbe errato sostenere che dopo la prima guerra mondiale gli orientamenti siano cambiati rapidamente, fino a guadagnare ben presto le posizioni attuali; al contrario, il cammino è stato lungo ma in continuo progresso. Oggi siamo nel vivo di uno sviluppo assai rapido per molti aspetti dell'oceanografia biologica.
È facilmente comprensibile come questa scienza abbia avuto il massimo sviluppo in quei settori della biologia che si occupano dell'inizio e della fine delle catene alimentari, cioè dei produttori primari e dei Pesci. Nel capitolo precedente nulla si è detto sullo sviluppo della biologia della pesca; non certo per mancanza di informazione, ché anzi la biologia della pesca nei primi decenni del secolo e negli anni successivi si è sviluppata costantemente fino a raggiungere l'elevato livello delle conoscenze attuali. Ritengo tuttavia opportuno lasciare la trattazione di questo aspetto dell'oceanografia biologica ad autori più competenti e limitarmi qui ai produttori primari. In un altro capitolo cercherò di descrivere lo stato attuale delle conoscenze relative ai due anelli successivi della catena alimentare, cioè allo zooplancton erbivoro e carnivoro, e mi limiterò a dare brevi cenni sui Pesci.
Nell'oceanografia sia chimica sia biologica si devono considerare i due aspetti, statico e dinamico. Ambedue le discipline hanno necessariamente avuto inizio con lo studio dell'aspetto statico. Hensen e Lohmann hanno studiato le concentrazioni degli organismi planctonici nel mare (la riserva costante). I chimici del sec. XX hanno anche studiato le concentrazioni dei sali nutritivi, cioè nitrati e fosfati. Per alcuni problemi la concentrazione è particolarmente importante, ma per molti altri si deve considerare anche l'aspetto dinamico, cioè a dire per le sostanze chimiche la velocità di rifornimento, e per gli organismi la velocità di riproduzione.
Oggi, grazie a metodi tecnici adeguati, è possibile studiare direttamente la velocità della produzione primaria, cioè la produzione autotrofica di sostanza organica da parte delle alghe planctoniche. Produzione primaria, infatti, se non se ne dà una definizione diversa, significa produzione di sostanza organica da parte delle alghe planctoniche, che sono i soli organismi di mare aperto ad avere importanza per questo processo. Invece la velocità di produzione e il trasferimento delle sostanze organiche attraverso i vari anelli della catena alimentare si possono in pratica studiare soltanto con metodi indiretti, e perciò i risultati sono assai meno sicuri. Spesso ci si deve accontentare di conoscere l'entità della riserva costante. Anche il rifornimento di un sale nutritivo è, in molti casi, difficile da studiare; spesso è più facile calcolarne indirettamente la velocità in base a quella della produzione primaria.
b) Riserva costante di alghe planctoniche e produzione primaria.
Non è possibile concentrare il fitoplancton partendo da una grande quantità d'acqua, perciò per valutare il materiale organico con metodi chimici si devono usare tecniche molto sensibili. Von Brand (v., 1935) ideò alcuni metodi per determinare l'azoto e il carbonio in piccole quantità di plancton, ma si tratta di metodi non molto utili, perchè normalmente, nel materiale organico particolato, i detriti morti sono presenti in quantità molto superiore a quella del plancton vivo. Krey (v., 1951) descrisse un metodo per la determinazione delle proteine; attraverso tale determinazione, però, è impossibile distinguere il fitoplancton dagli organismi di tipo diverso. Recentemente Hamilton e altri (v., 1968) hanno usato per lo stesso scopo misurazioni di ATP.
I pigmenti fotosintetici sono i soli componenti specifici per tutte le alghe planctoniche fotoautotrofe e la loro determinazione è relativamente facile. L'analisi di questi pigmenti ebbe inizio intorno al 1930, su campioni presi con retini. Krey (v., 1939) introdusse una tecnica per filtrare in modo adeguato l'acqua di mare. Negli anni più recenti si è fatto largo uso delle analisi dei pigmenti, in particolare della determinazione della clorofilla; Richards e Thompson (v., 1952) introdussero una tecnica spettrofotometrica. Le analisi del pigmento presentano varie fonti di errore, e tuttavia sono da considerarsi il metodo migliore di cui si disponga attualmente per misurare la riserva costante di fitoplancton. Una fonte di errore sta nel fatto che la percentuale di clorofilla sulla quantità totale di sostanza organica nelle alghe planctoniche varia almeno di un fattore 10. Tale variazione però non è casuale e si può eliminare in parte introducendo un fattore di correzione per le varie aree e per le diverse profondità degli oceani. Alcuni autori hanno utilizzato il materiale in sospensione come indice della quantità di fitoplancton nel mare aperto; Jerlov (v., 1953) misurò questo materiale con mezzi ottici, per mezzo dell'effetto Tyndall. Hentschel (v., 1933-1936) osservò una stretta correlazione fra il colore del mare e la quantità di plancton; però nelle acque costiere, dove si possono trovare grandi quantità di detriti e di particelle inorganiche, il materiale in sospensione e il colore non forniscono indicazioni molto utili per valutare la quantità di plancton.
Il conteggio delle cellule delle alghe planctoniche è una tecnica che ha trovato largo impiego per calcolare la riserva costante di fitoplancton, e che presenta anche il grande vantaggio di consentire di identificare, contemporaneamente all'operazione di conteggio, i vari tipi di organismi. Il solo numero delle cellule non è un elemento sufficiente a misurare la riserva costante: infatti, alcune specie sono grandi, altre piccole. Questo problema fu risolto dal Lohmann (v., 1908), il quale, oltre al numero delle cellule delle varie specie, determinò anche il volume del plasma; ma spesso anche il volume totale delle cellule è di scarso significato, perché può in gran parte dipendere dalla quantità di succo cellulare. La tecnica del Lohmann è estremamente laboriosa e perciò non ha trovato molto favore presso i biologi marini.
Molti studiosi del plancton preferiscono usare la già ricordata tecnica di Utermöhl (del microscopio invertito) per calcolare il numero delle cellule. L'unico difetto di questa tecnica sta nel fatto che essa richiede la conservazione delle alghe, secondo un processo che distrugge molti piccoli flagellati privi di membrana, riducendo così quello che è talvolta il costituente più importante della biomassa. Come abbiamo già detto, le ricerche basate sul conteggio delle cellule per mezzo della centrifugazione o della tecnica di Utermöhl hanno reso eccellenti servigi all'oceanografia nella prima metà di questo secolo. La ricerca sulla riserva costante di fitoplancton ha recato un notevole contributo alla comprensione della produttività primaria degli oceani. Tuttavia soltanto le tecniche moderne che hanno reso possibile studiare gli aspetti dinamici, cioè la velocità della produzione primaria nel mare, hanno consentito di comprendere appieno questo processo.
Il primo tentativo di misurare l'effettiva produzione organica del fitoplancton in una determinata area marina fu realizzato da Atkins (v., 1922) nella Manica: il metodo da questi usato consiste nella determinazione della perdita di diossido di carbonio in 1 m2 di acqua superficiale, dalla fine dell'inverno al culmine dell'estate. Poiché la perdita di CO2 è uguale alla quantità utilizzata nella fotosintesi, si può calcolare il valore minimo della produzione organica di sei mesi. Non fu possibile calcolare lo scambio di CO2 con l'atmosfera, ossia la sua rigenerazione, per la presenza dei prodotti della respirazione di tutti gli organismi, comprese le stesse alghe planctoniche. Atkins (v., 1923) calcolò nello stesso modo la diminuzione dei fosfati. Tali metodi sono stati però poco usati, perché presentano notevoli difficoltà.
Nel caso della vegetazione terrestre, la produzione di sostanza organica è concentrata per lo più in unità di dimensioni piuttosto vaste, tali da rendere le misurazioni relativamente facili. Diverso è il comportamento del fitoplancton marino. Nell'oceano aperto le alghe planctoniche di piccole dimensioni sono distribuite in uno strato che ha uno spessore compreso tra i 50 e i 100 metri, perciò la quantità di alghe che si raccolgono in 1 litro di acqua prelevata dalla zona fotica è uguale a quella contenuta in un cilindro di solo 0,1-0,2 cm2 di base e di altezza uguale a quella di tutta la zona fotica. Poiché la velocità della produzione giornaliera per m2 è di regola uguale a circa 150 mg di C, la produzione media per litro nella zona fotica è soltanto di circa 2 μg di C al giorno, corrispondenti a circa 6 μg di O2 Ciò significa che la media giornaliera del diossido di carbonio assimilato equivale a 1 decimillesimo della quantità totale del CO2 presente nell'acqua di mare, ciò che corrisponde a 1 millesimo dell'ossigeno. Non è possibile misurare differenze tanto piccole con metodi chimici.
Se lo stesso plancton è concentrato in una zona fotica di scarsa profondità - per es. 10 metri - come quello che si trova in molte acque costiere, si può calcolare una velocità di produzione giornaliera di 150 mg di C per m2 con il metodo della determinazione dell'ossigeno. Se fossero disponibili soltanto metodi chimici, i metodi sperimentali per misurare la produzione primaria si potrebbero applicare soltanto alle zone costiere più produttive.
Gaarder e Gran (v., 1927) usarono per primi, in un fiordo della Norvegia, una tecnica basata sulla produzione dell'ossigeno in bottiglie da esperimento sospese in acqua di mare. Campioni d'acqua di mare furono raccolti a differenti profondità entro la zona fotica e aspirati in bottiglie con tappi di vetro. Alcune bottiglie furono usate per determinare la concentrazione dell'ossigeno prima dell'inizio dell'esperimento, applicando la tecnica di Winkler; le altre, delle quali alcune trasparenti e alcune oscurate con vernice nera, furono immerse di nuovo alla profondità da cui erano stati tratti i campioni e qui lasciate per 24 ore, sospese a un filo legato a un gavitello ancorato. La quantità di ossigeno contenuta in una bottiglia oscura, meno quella di una bottiglia campione, dà la misura della velocità di respirazione di tutti gli organismi presenti: alghe planctoniche, piccoli animali e batteri. La quantità di ossigeno contenuta in una bottiglia trasparente, meno quella di una bottiglia oscura, dà la misura della velocità di fotosintesi delle alghe (cioè la produzione primaria lorda).
La tecnica dell'ossigeno è stata usata con successo in varie zone costiere ad alta produttività. Invece nelle acque oceaniche limpide essa è di scarsa utilità per la misurazione della produzione organica, perché la velocità del processo fotosintetico è troppo bassa per permettere esperimenti di breve durata. Negli esperimenti più lunghi insorgono serie complicazioni, dovute al fatto che nell'acqua contenuta nelle bottiglie si verifica un imponente sviluppo di batteri.
Steemann Nielsen (v., 1952) escogitò una nuova tecnica per misurare la produzione organica negli oceani, basata sulla marcatura con carbonio radioattivo 14C. Questa tecnica fu usata per la prima volta nel corso della spedizione danese della Galathea, che nel 1950-1952 percorse tutti gli oceani (v. Steemann Nielsen e Jensen, 1957). L'in- corporazione del tracciante ne!la sostanza organica delle alghe planctoniche viene usata come misura della produzione : prima dell'esperimento si aggiunge all'acqua di mare una quantità determinata di una soluzione di NaH14CO3, quindi il contenuto di 14C rilevato nel plancton dopo l'e- sperimento si moltiplica per un fattore corrispondente al rapporto tra CO2 (totale) e 14CO2 esistente nell'acqua all'inizio dell'esperimento.
La quantità di 14C assimilato si determina misurando la radiazione β del plancton trattenuto da una membrana filtrante.
Per la determinazione della produzione di sostanza al disotto di 1 m2 di superficie oceanica, si possono impiegare diverse variazioni del metodo. La tecnica in situ, simile a quella dell'ossigeno usata da Gaarder e Gran, generalmente non è utilizzabile nelle spedizioni oceaniche effettuate da grandi navi, perché richiede troppo tempo ed è eccessivamente costosa. Sebbene siano state messe in evidenza varie complicazioni inerenti a questa tecnica, gli errori introdotti nelle misurazioni della produzione oceanica primaria hanno scarsa importanza. La velocità della pro- duzione, che può variare secondo una distribuzione casuale delle alghe nell'acqua marina, non richiede di essere determinata con un'altissima precisione.
Attualmente lo studio della produzione di sostanza or- ganica da parte del fitoplancton è uno dei problemi centrali della scienza del mare. È necessario sottolineare che non ci si deve limitare alla misurazione della produzione di so- stanza organica, ma si deve anche mettere in evidenza quali sono le particolari alghe che la producono.
Non vi sono ancora prove assolute, ma è lecito pensare che nel destino della sostanza organica abbia molta importanza il tipo degli organismi che la producono, in quanto a esso sono legati gli organismi che se ne nutrono. Gli organismi planctonici erbivori non si cibano di qualunque tipo di alghe, ma soltanto di quelle che hanno certe dimensioni e altre proprietà caratteristiche.
c) I fattori che regolano la produzione primaria.
Fin dall'inizio del secolo sono stati compiuti numerosi tentativi per individuare i fattori che regolano la produzione primaria. Esponendo l'insieme dei vari fattori faremo menzione dello sviluppo storico attraverso il quale si è giunti alle concezioni attuali sulle fonti della produzione oceanica primaria.
È ormai stabilito che i fattori principali della produzione del fitoplancton nel mare sono i seguenti: 1) rifornimento di sali nutritivi; 2) luce; 3) animali che si cibano di fitoplancton. Altri fattori, fra cui la temperatura, hanno un'importanza relativa.
Se si considerano tutti i mari nel loro complesso, si può affermare che, di solito, il rifornimento di sali nutritivi negli strati produttivi è il fattore essenziale che determina l'intensità della produzione organica annuale. Tuttavia in alcuni periodi dell'anno e in molte zone del mare vi sono altri fattori - in primo luogo la luce - che possono assumere momentaneamente maggiore importanza.
Normalmente la natura ha stabilito un equilibrio tra tutti i fattori che regolano la produzione di fitoplancton; solo di quando in quando è possibile indicare uno dei fattori come l'unico limitante. Di solito un cambiamento di uno dei fattori influenza gli altri e instaura un nuovo equilibrio: tutti i fattori sopra ricordati devono essere considerati inscindibili. In pratica, però, si possono fare solamente tentativi in questo senso. Sembra, in generale, che le dimensioni di una popolazione di fitoplancton siano mantenute direttamente da un equilibrio fra la sua crescita e il suo consumo da parte dello zooplancton che se ne nutre. Tuttavia l'equilibrio dipende dalla disponibilità dei sali nutritivi e quindi, in ultima analisi, è da questa governato.
1. Sali nutritivi. - I principali elementi che possono diventare limitanti nell'oceano sono l'azoto (proveniente soprattutto dai nitrati) e il fosforo (dai fosfati inorganici). Altri elementi, come il ferro, possono, in date circostanze, diventare troppo scarsi per l'accrescimento del fitoplancton.
Nelle masse d'acqua situate a profondità inferiori ai 500-1.000 m, la concentrazione dei fosfati e dei nitrati inorganici è piuttosto alta, tranne poche eccezioni: per esempio quella del fosforo da fosfati è di circa 1-3 m-grammoatomi per m3. Nell'Atlantico settentrionale le concentrazioni, di norma, sono più basse che negli altri oceani.
La concentrazione di fosfati e di nitrati nelle acque profonde è dovuta in primo luogo alla rigenerazione della sostanza organica che originariamente viene prodotta in prossimità della superficie, ma che, per effetto di vari meccanismi, viene poi trascinata in basso. Quanto più l'acqua di profondità è ‛vecchia', tanto maggiore è la quantità di materiale organico che è stato trasportato in basso. L'acqua profonda del Pacifico è più vecchia di quella dell'Atlantico; infatti una parte essenziale della prima deriva da quella del secondo. Si comprende perciò facilmente perché la concentrazione dei nitrati e dei fosfati sia notevolmente più alta nelle acque profonde del Pacifico che in quelle dell'Atlantico.
Ciò che determina la produttività di una zona marina non è la concentrazione che si osserva in un dato momento, ma la velocità di rifornimento dei sali nutritivi nello strato fotico. Questo fatto importante fu notato per la prima volta da Ketchum (v., 1947). Il rifornimento è determinato da due processi: la decomposizione e la circolazione dell'acqua.
La rigenerazione dei sali nutritivi dalla sostanza organica si può suddividere in una rigenerazione diretta, e rapida, dovuta alle escrezioni da parte dello zooplancton che si nutre di fitoplancton (v. Ketchum, 1962), e in una rigenerazione, prodotta da batteri, di composti organici provenienti da piante e da animali parzialmente digeriti.
La rigenerazione batterica ha luogo nelle masse d'acqua libere e negli strati superficiali dei sedimenti. Generalmente, la rigenerazione microbiologica nelle masse d'acqua libere appare lenta. È molto probabile che nelle zone poco profonde quella che si svolge nei sedimenti costituisca la parte più importante della rigenerazione totale.
Mentre la temperatura sembra avere un'importanza piuttosto limitata nella velocità dei processi biologici del mare, tutto sembra dimostrare l'importanza considerevole di questo fattore nella velocità di rigenerazione dei sali nutritivi. Nelle acque costiere, dove la rigenerazione dei sali nutritivi negli strati più superficiali dei sedimenti del fondo deve essere considerata come il fattore essenziale che determina la velocità della produzione organica per la maggior parte dell'anno, si può trovare una stretta correlazione tra la velocità della produzione primaria e la temperatura dell'acqua.
In altre aree, e in particolare nell'oceano aperto, affinché si verifichi un abbondante rifornimento dei sali nutritivi nella zona fotica è necessaria la circolazione dell'acqua. Questa può essere orizzontale, e in tal modo acqua ricca di sali nutritivi è rifornita dalle zone adiacenti. Più importante è, tuttavia, la circolazione verticale: per questo le masse d'acqua immediatamente sottostanti la zona fotica sono in genere piuttosto ricche di sali nutritivi.
Si deve al chimico francese G. Denigès un metodo per la misurazione quantitativa di concentrazioni minime di fosfati nell'acqua. Per l'acqua di mare la tecnica fu usata per la prima volta da Atkins (v., 1923). Alcuni anni dopo Harvey (v., 1926) introdusse una tecnica piuttosto buona, derivata da un metodo elaborato originariamente dal Denigès, per misurare i nitrati nell'acqua di mare. Si avevano ormai a disposizione i mezzi adatti per dimostrare che, a conferma dell'ipotesi di Brandt, i nitrati e i fosfati sono realmente i fattori limitanti della produzione primaria. La spedizione tedesca della Meteor nell'Atlantico meridionale (1925-1927) eseguì ricerche su larga scala sulla distribuzione orizzontale e verticale dei sali nutritivi in tutto l'oceano. Essendosi contemporaneamente realizzata su vasta scala la determinazione quantitativa della riserva costante delle alghe planctoniche, Hentschel e Wattenberg (v., 1930) poterono stabilire la correlazione tra sali nutritivi e biomassa di fitoplancton. Quest'operazione rappresenta una pietra miliare nella storia dell'oceanografia biologica. I due autori dimostrarono che è possibile mettere in evidenza una stretta correlazione tra la concentrazione dei fosfati inorganici e la riserva costante di plancton, sia vegetale sia animale, quando se ne consideri la media nello strato compreso tra la superficie e i 50 m di profondità (v. fig. 1). Se invece essi avessero utilizzato la concentrazione superficiale dei fosfati, la maggior parte delle zone sarebbero risultate povere, pur essendovi una considerevole variazione della riserva costante. Ciò conferma quanto abbiamo detto prima, cioè che si deve considerare il rifornimento di un sale nutritivo e non la sua concentrazione. Una forte concentrazione di sali nutritivi nello strato più superficiale e una considerevole riserva costante furono trovate soltanto nelle zone in cui ha luogo un movimento ascensionale delle acque sottostanti la superficie. Tutto ciò conferma l'ipotesi del Nathansohn (1910) sul rinnovo delle acque contenenti alghe planctoniche, anche se le cause non sono esattamente le stesse indicate da questo autore.
Si è trovato che le zone in cui vi è un rifornimento mi- nimo di sali nutritivi sono quelle in cui l'apporto di acqua nello strato fotico proviene esclusivamente da un flusso laterale di acque superficiali relativamente ‛vecchie' provenienti dalle aree confinanti e dove si verifica una tendenza al movimento discendente dell'acqua di superficie. Tipici in tali condizioni sono i centri di grandi vortici anticiclonici, come quelli che si trovano nell'Atlantico settentrionale e meridionale. In queste zone si riscontrano le punte più basse della produzione primaria.
2. La luce. - Fornisce alle alghe planctoniche l'energia necessaria per la trasformazione delle sostanze inorganiche (diossido di carbonio, acqua, nitrati e fosfati) in sostanza organica. Per queste trasformazioni viene utilizzata soltanto la luce assorbita dai pigmenti attivi nella fotosintesi; in generale tali pigmenti assorbono solo una minima parte della luce sottomarina. Nella maggior parte delle aree marine la quantità di luce disponibile è sufficiente a fornire l'energia necessaria a una produzione di sostanza organica equivalente alle quantità di fosforo e di azoto annualmente disponibili nello strato eufotico.
In molte aree, in certi periodi dell'anno, l'energia luminosa è insufficiente, per esempio d'inverno alle alte latitudini; però il fenomeno non si può generalizzare a tutto l'anno. Una delle rare eccezioni è quella del bacino del polo Nord, perché qui la coltre di ghiaccio che assorbe la luce è praticamente permanente in tutti i periodi dell'anno.
Le condizioni di luce, insieme con le condizioni idrografiche generali, determinano la profondità della zona fotica. Per lo zooplancton che si nutre del plancton vegetale è importante che una determinata velocità di produzione primaria per unità di superficie si verifichi in una zona eufotica bassa o profonda, come si dirà in seguito.
Nel mare l'utilizzazione per la fotosintesi dell'energia luminosa incidente nella parte visibile dello spettro varia tra lo 0,02 e il 2% circa. Si considera che l'utilizzazione media abbia un valore di circa 0,1%. Una forte utilizzazione della luce incidente è possibile soltanto se le alghe planctoniche sono concentrate in uno strato poco profondo in cui si compie la fotosintesi: in tal caso l'assorbimento di luce da parte dell'acqua è ridotto al minimo. In conformità con le previsioni teoriche, la velocità della produzione organica per unità di superficie è inversamente proporzionale alla profondità dello strato fotico (v. fig. 2).
Le alghe planctoniche sono trasportate passivamente dalle masse d'acqua in movimento e quindi subiscono anche eventuali spostamenti verticali di notevole ampiezza, che incidono sull'utilizzazione della luce. Nel mare aperto la zona fotica per lo più ha un'estensione superiore a quella dello strato (profondo circa 25 m) in cui le masse d'acqua si mescolano giornalmente in senso verticale.
Alle latitudini più alte, durante l'inverno, si può formare uno strato d'acqua ben mescolata, profondo varie centinaia di metri; ciò è dovuto al raffreddamento dell'acqua di superficie, ed è quanto avviene nell'Atlantico settentrionale. In queste condizioni una singola cellula di alga non può assorbire l'energia luminosa sufficiente al processo fotosintetico nel corso del giorno, anche se durante l'ultima fase dell'inverno l'illuminazione presso la superficie può essere ottima. Poiché il tasso di respirazione è uniforme in tutto lo strato mescolato, ne risulta un deficit nel metabolismo delle alghe. In queste aree la produzione di fitoplancton può avere inizio solamente quando, per effetto del riscaldamento estivo, si è stabilito presso la superficie uno strato fotico relativamente basso, e di conseguenza viene inibito il trasporto delle alghe a maggiori profondità.
Braarud e Kleem (v., 1931) segnalarono per primi l'importanza determinante del mescolamento verticale per l'utilizzazione della luce da parte del fitoplancton. Sverdrup (v., 1953) elaborò una trattazione matematica del problema, considerando in particolare l'avvio della produzione durante la primavera, alle alte latitudini.
3. Organismi fitoplanctofagi. - Le alghe planctoniche allevate in laboratorio crescono in modo del tutto diverso da quelle che vivono in condizioni naturali. In laboratorio la scarsità di sali nutritivi - per es. quelli contenenti azoto e fosforo - determina ben presto l'assoluta deficienza di queste sostanze, finché le alghe raggiungono uno stadio in cui la crescita si arresta totalmente. Se l'accrescimento delle alghe nel mare non fosse controllato dagli organismi fitoplanctofagi, nelle acque prive di un costante rifornimento di sali nutritivi, per movimenti verticali o altre cause, dovremmo aspettarci di trovare un'alternanza di periodi con biomasse relativamente ricche di fitoplancton, e di lunghi periodi privi di alghe in accrescimento. Però in natura tale situazione non si verifica. Nelle aree tropicali, in cui le condizioni idrografiche non presentano variazioni notevoli nel corso dell'anno, la quantità delle alghe rimane estremamente stabile sia nello spazio sia nel tempo. Perciò deve quivi esistere normalmente un equilibrio fra l'accrescimento delle alghe e la loro scomparsa, dovuta al consumo da parte dei fitofagi e al movimento verso il fondo. Come si è già detto fu il Nathansohn (1910) che per primo richiamò l'attenzione sull'importanza del consumo delle alghe da parte dei fitofagi per la produzione del fitoplancton.
d) Alcuni tipi di produzione oceanica primaria.
Passiamo ora a illustrare alcuni schemi fondamentali di produzione primaria per certi tipi di aree. Come abbiamo visto, i fattori che regolano la produzione primaria si combinano in molti modi, ciascuno dei quali determina particolari modelli di produzione.
Le zone tropicali e subtropicali del mare aperto sono state studiate in modo piuttosto esauriente dal punto di vista della produzione primaria. Una caratteristica comune, tanto nel conteggio del plancton quanto nelle misurazioni della produzione organica, è la scarsa variabilità dei valori riscontrati nelle aree uniformi dal punto di vista idrografico.
Tutti gli oceani sono caratterizzati da vasti sistemi di correnti. Nell'Atlantico meridionale, nel Pacifico meridionale e nell'Oceano Indiano le correnti circolano in senso antiorario. In tutte le stagioni, nelle acque dell'emisfero meridionale esiste una tipica corrente sudequatoriale che parte dal continente e procede in direzione ovest; essa deriva da una corrente che si muove in direzione nord lungo il continente. La corrente sudequatoriale si muove trasversalmente all'oceano a sud dell'equatore. Quando sta per raggiungere il continente a ovest, la maggior parte dell'acqua volge verso sud e segue le coste continentali; verso il 45° di latitudine sud la corrente volge verso est. Nel centro di queste correnti si forma un vortice anticiclonico.
Contrariamente a quanto avviene negli oceani dell'emisfero meridionale, le correnti dell'Atlantico settentrionale e del Pacifico settentrionale circolano in senso orario. In tutti gli oceani si trova una controcorrente che si muove verso est fra la corrente nordequatoriale e quella sudequatoriale, che hanno entrambe un movimento verso ovest. Nell'Oceano Indiano, per la sua forma particolare, i sistemi di correnti della zona settentrionale assomigliano solo in parte a quelli dell'Atlantico e del Pacifico settentrionali. La corrente nordequatoriale diretta verso ovest si trova soltanto durante parte dell'anno, infatti essa è neutralizzata o sostituita da una corrente verso est quando soffia il monsone di sud-ovest.
In teoria là dove ha luogo il movimento verticale si dovrebbero trovare regioni ad alta produttività. Ciò si verifica lungo le coste dei continenti nelle parti orientali degli oceani, alle latitudini in cui soffiano gli alisei. Dovremmo aspettarci di trovare una produttività relativamente alta anche nei luoghi dove si osservano correnti superficiali divergenti, come avviene nelle zone di confine tra la controcorrente equatoriale e le correnti equatoriali settentrionale o meridionale. D'altronde, nella parte centrale dei vortici anticiclonici ci si dovrebbe aspettare una bassa produttività. In queste zone l'acqua di superficie si sposta lentamente verso le zone più profonde; un rinnovo della massa d'acqua si realizza per l'azione laterale di acqua di superficie ‛vecchia', povera di sali nutritivi.
L'Oceano Indiano, che in passato fu il meno studiato di tutti gli oceani, oggi è forse il meglio conosciuto, per merito di una recente spedizione internazionale (v. fig. 3). Con lo sforzo congiunto di molte nazioni è stata raccolta un'importante massa di notizie relative al tasso della produzione primaria. Questa varia secondo le stagioni, in relazione al regime monsonico, che nell'Oceano Indiano è particolarmente importante.
Nelle regioni oceaniche alle alte latitudini non si può trascurare l'influenza delle stagioni: questo principio vale soprattutto per l'Oceano Artico e per l'Antartico, ma si applica anche alle zone temperate dell'oceano.
Il mescolamento verticale dell'acqua che ha luogo durante l'inverno, a causa del raffreddamento della superficie, può avere, sulla produzione primaria, tanto un'influenza positiva (per un più elevato apporto di sali nutritivi allo strato fotico) quanto un'influenza negativa (dovuta al trasporto di alghe in strati sottostanti, dove l'illuminazione è scarsa).
Nelle aree in cui l'inverno non provoca alcun mescola- mento verticale, sembra prevalere, immediatamente, l'influenza benefica. Questo è quanto si osserva, per esempio, nell'Oceano Atlantico vicino alle isole Bermude, immediatamente a sud del Mar dei Sargassi, dove la produzione primaria presenta un tipico massimo invernale, mentre il suo valore è basso in tutte le altre stagioni. In generale, più alta è la latitudine, più è pronunciata l'influenza negativa durante l'inverno; l'influenza positiva del mescolamento verticale dell'inverno si riscontra di solito durante la primavera, quando un termoclino si è ben stabilizzato.
Nel bacino del polo Nord si trova una produzione di fitoplancton estremamente bassa. Il pack polare permanente, dello spessore medio di 2-4 metri, è ricoperto di neve durante la maggior parte dell'anno. I canali aperti fra i ghiacci non rappresentano più dell'1% della superficie. La produzione annua di materiale organico è inferiore a 1 g C/m2 (v. Ryther, 1963). Recenti ricerche (v., per es., El-Sayed, 1970) hanno dimostrato nelle acque antartiche valori molto variabili della produzione ; è probabile che i valori molto alti che si calcolavano per queste zone fino a pochi anni orsono non siano esatti.
Ryther (v., 1963) ha pubblicato una vasta rassegna sulla variazione della produttività oceanica.
e) Produzione globale di vegetali planctonici e bentonici.
Evidentemente non è ancora possibile fare una stima esatta del totale della produzione marina primaria. Dopo alcune controversie sorte intorno agli anni cinquanta, i vari autori (v. Ryther, 1960; v. Steemann Nielsen, 1960) si trovano ora d'accordo nel calcolare che la produzione totale netta di plancton per anno sia di circa 2•1010 tonnellate di carbonio.
Non si può non tener conto anche dei vegetali bentonici, nonostante che essi abbiano un'importanza praticamente nulla ai fini della produzione marina totale nell'oceano aperto. Le ricerche sulla velocità di produzione del macro e del microbentos hanno rivelato un alto valore di produzione nelle regioni costiere (v., per es., Odum, 1957; v. Grøntved, 1960). Bogorov (v., 1969) ha calcolato che la produzione annua dei vegetali bentonici in tutto il globo rappresenta lo 0,05% della produzione totale di fitoplancton oceanico (v. tabella). Ma probabilmente questo valore è un po' più basso del reale.
È probabile che le alghe bentoniche abbiano un ruolo molto importante anche se indiretto - per gli animali delle regioni costiere. La maggior parte dei vegetali bentonici non è mangiata direttamente dagli animali: sono piuttosto i Batteri che decompongono il materiale organico in essi contenuto e costituiscono così un'importante via di trasmissione dell'energia agli animali.
Keys e altri (v., 1935) dimostrarono che l'acqua degli oceani, a tutte le profondità, contiene una quantità assai notevole di sostanze organiche in soluzione e costituisce la più vasta riserva di sostanza organica di tutto il globo. Zobell e Anderson (v., 1936) osservarono che di norma questa sostanza organica viene attaccata solo lentamente dai Batteri. Se si pone l'acqua in bottiglie, una parte della sostanza organica disciolta viene metabolizzata rapidamente, forse per la presenza di superfici attive. Sebbene da molto tempo sia noto il contenuto relativo degli elementi che la compongono, ad esempio del carbonio e dell'azoto, la costituzione chimica della sostanza organica disciolta non è ancora nota. Servendosi della datazione con il carbonio 14 Williams e collaboratori (v., 1969) hanno dimostrato che l'età della sostanza organica contenuta nelle profondità del Pacifico è, in media, di circa 3.400 anni. Partendo dalla produzione primaria annua dell'oceano è stato calcolato (v. Strickland, 1970) che circa lo 0,5% di tale produzione viene immesso nell'acqua sotto forma di sostanza organica disciolta. Tale valore è ovviamente approssimato per difetto, poiché non comprende la sostanza organica, rapidamente metabolizzabile, che si trova spesso nelle acque superficiali.
4. Riserva costante di zooplancton e catene alimentari nell'oceano.
Oltre a escogitare una tecnica adatta a misurare la riserva costante di fitoplancton, Lohmann (v., 1908) mise a punto una tecnica per la misurazione delle unità di zooplancton presenti nel mare, che consiste nel pompare l'acqua da varie profondità e filtrarla attraverso tessuti di seta a maglie minute. Così si raccoglie la maggior parte dello zooplancton microscopico e nel sedimento dei campioni centrifugati si trovano e si possono contare con il fitoplancton anche organismi più piccoli, come gli Infusori. Purtroppo nei decenni successivi pochi zoologi compresero quale fosse l'importanza dello zooplancton di più piccole dimensioni. Hentschel, che rappresentò una delle poche eccezioni, contò, durante la spedizione della Meteor (1925-1927), tutti gli organismi zooplanctonici contenuti nei campioni del sedimento, ottenuti prelevando di volta in volta 3 litri d'acqua a varie profondità. Questo volume, decisamente troppo piccolo per un campionamento dello zooplancton contenuto nell'acqua marina, tuttavia fu sufficiente a dare un'idea generale della sua distribuzione, per esempio a dimostrare la stretta correlazione che esiste tra la riserva costante di zoo- e di fitoplancton (v. fig. 1).
Negli anni seguenti furono descritte, e usate, varie tecniche basate sulla filtrazione, attraverso tessuti di seta a maglie sottilissime, di determinati volumi d'acqua prelevati a diverse profondità. Strickland (v., 1970) ha presentato una rassegna delle ricerche eseguite presso lo Institute of Marine Resources dell'Università di California sulla rete delle catene alimentari del plancton marino, in cui il microzooplancton ha un'importanza predominante. Sebbene attualmente sia ancora assai difficile calcolare, a partire dalla riserva costante, la quantità del microzooplancton consumato, si è potuto stabilire, per le acque della California, che esso rappresenta in media il 30% della produzione giornaliera di fitoplancton. I soli Ciliati possono consumare il 50% o della produzione giornaliera del fitoplancton di piccolissime dimensioni (inferiori a 10 μ), che generalmente rappresenta il 25% del totale della biomassa vegetale.
Steemann Nielsen (v., 1962) richiamò l'attenzione sul fatto che gli erbivori, per assumere il cibo necessario, devono utilizzare più energia nelle acque oligotrofiche che in quelle eutrofiche. La maggior parte del cibo che viene assimilato nelle acque oligotrofiche è utilizzata per l'attività respiratoria e soltanto una piccola parte resta a disposizione dell'accrescimento e della riproduzione. Anche l'estensione verticale della zona fotica ha importanza: se in una zona fotica profonda e in una bassa si ha la stessa intensità di produzione primaria, la concentrazione delle alghe planctoniche sarà maggiore nella zona bassa e in questa quindi sarà più facile per gli erbivori procurarsi il cibo. Ciò spiega perché le acque costiere con zona fotica relativamente bassa siano generalmente più ricche di animali di tutti i tipi - Pesci compresi - rispetto alle regioni oceaniche in cui le zone fotiche sono più profonde.
Fin dall'inizio del secolo è stato fatto molto lavoro per determinare la riserva costante dello zooplancton marino di maggiori dimensioni. Si sono incontrate però difficoltà tecniche considerevoli, innanzi tutto perché gli animali dotati di rapido movimento non si lasciano catturare con il retino. Sebbene sia quasi impossibile costruire una rete o altri apparecchi con cui raccogliere tutto lo zooplancton di maggiori dimensioni, le ricerche oceaniche su vasta scala hanno dato informazioni importanti perfino quando si sono usate tecniche piuttosto primitive. Jespersen (v., 1935) ha pubblicato i risultati quantitativi di catture eseguite con reti a imboccatura orizzontale, nel corso della spedizione danese del Dana; tali tipi di cattura danno termini di confronto molto utili.
Gli studiosi russi, in particolare, hanno prodotto una gran quantità di lavoro sulla distribuzione quantitativa dello zooplancton a tutte le profondità e su scala mondiale (v. Bogorov, 1967) usando reti verticali a chiusura. Con queste tecniche le misurazioni assolute della biomassa sono difficili da realizzare, ma in senso relativo i dati che se ne ricavano sono molto importanti (v. Hansen e Andersen, 1962). Le aree con alta intensità di produzione primaria sono in generale identiche a quelle che hanno valori elevati della riserva costante di zooplancton, nonostante che nelle aree con acque subsuperficiali dotate di movimenti verticali la massa dello zooplancton - specialmente dei carnivori - sia alquanto diversa da quella del fitoplancton. Ciò dipende dal fatto che lo zooplancton erbivoro e soprattutto quello carnivoro si sviluppano più lentamente del fitoplancton, in modo tale che l'acqua viene trasportata lontano dai centri di moto ascensionale quando la concentrazione è massima (v. Steemann Nielsen, 1937; v. Vi- nogradov e Voronina, 1962).
Come abbiamo già detto, attualmente l'Oceano Indiano può considerarsi il meglio studiato, grazie alla spedizione internazionale che lo ha esplorato. La produzione organica primaria netta, per anno, è di 3,9•109 t di C per un'area di 51.106 km2, il che costituisce circa un quinto della produzione oceanica mondiale.
La distribuzione della biomassa di zooplancton nell'Oceano Indiano è rappresentata nella fig. 4, secondo Prasad e altri (v., 1970). L'emisfero meridionale, specialmente a sud del 10° di latitudine, presenta valori molto bassi della biomassa zooplanctonica, mentre le regioni a sud dell'arcipelago indonesiano e l'area prossima all'Australia hanno una maggior densità di plancton. Le aree con alta produttività si trovano nella parte nordoccidentale dell'oceano, lungo le coste della Somalia e dell'Arabia e in parte in prossimità delle coste meridionali della penisola indiana. Le zone a bassa produttività occupano la parte centrale del Mare Arabico e del Golfo del Bengala.
Le ricerche eseguite in tutti gli oceani nelle stazioni delle zone profonde, da navi russe e di altri paesi, hanno dimostrato che la quantità di plancton nei vari strati di tali zone è sempre correlata con il plancton dello strato superficiale: più è ricco il plancton degli strati superiori, più è ricco quello degli strati più profondi, e viceversa. Esiste a questo proposito una rassegna del Vinogradov (v., 1962). La quantità di biomassa planctonica diminuisce rapidamente con l'aumentare della profondità, e la diminuzione di solito è più forte tra i 100 e i 200 m e tra i 500 e i 700. La diminuzione totale del plancton negli strati profondi è molto forte: per esempio, lo strato compreso tra 6.000 e 6.500 m di profondità nella fossa tra le Curili e la penisola di Camciatca contiene per ogni m3 una quantità di plancton mille volte inferiore a quella che si trova a 50 m di profondità.
Si dispone ormai di dati sufficienti per poter sostenere che le profondità oceaniche sono rifornite di cibo soprattutto attraverso il trasporto attivo di materiale organico proveniente dalle zone superficiali, operato dalla migrazione degli animali. Gli animali delle zone intermedie si spostano di notte verso la superficie per nutrirsi di fitoplancton. Altri animali si nutrono nella zona di superficie durante certi periodi del loro ciclo vitale, per discendere poi alle profondità di 1.000-2.000 m, dove possono essere predati dagli animali di queste zone. Questi ultimi, discendendo a profondità maggiori, diventano a loro volta preda degli animali che vivono colà. La maggior parte degli animali pelagici di profondità sono carnivori.
Gli ittiologi marini del passato videro giustamente la relazione tra plancton e Pesci, soprattutto pesci pelagici. Si consiglia il lettore di consultare l'abbondante bibliografia relativa a questo argomento.
Nelle conclusioni sul lavoro eseguito in passato dall'Institute of Marine Resources dell'Università della California, Strickland (v., 1970, p. 111) scrive: ‟Nel decennio scorso ero fortemente prevenuto contro l'uso dei modelli matematici per prevedere e spiegare la dinamica del plancton. Secondo la matematica formale, alcuni modelli furono elaborati correttamente, altri in modo profondamente errato. Comunque, mi sembra che elaborare tali modelli sia una pura illusione, perché le nostre conoscenze sulla fisiologia degli animali e delle piante sono molto scarse. Neppure sappiamo quali siano i parametri critici, nè come impostare matematicamente le correlazioni tra luce, sali nutritivi e foto sintesi, nè quelle tra densità di preda e predatori. Ancor più difficile appare il problema quando si considerino gli innumerevoli sistemi di retroazione che insorgono per effetto di vari fattori, come per es. la rigenerazione dei sali nutritivi da parte degli organismi che si nutrono di plancton e le complesse situazioni idrodinamiche delle zone in cui si hanno movimenti verticali". Strickland aggiunge: ‟L'elaborazione matematica dei risultati di esperimenti eseguiti in laboratorio usando un ambiente controllato è, evidentemente, cosa del tutto diversa; se non si tiene conto di questo fatto, ogni ricerca significativa è inattuabile". Quando si discute delle catene alimentari si devono tener presenti queste affermazioni.
5. Gli animali del fondo marino.
Nel primo decennio del secolo lo zoologo danese C. G. J. Petersen intraprese un lavoro sperimentale usando un apparecchio di cattura da lui inventato, con cui si poteva calcolare il numero e il peso degli animali del fondo marino. Scopo principale delle ricerche del Petersen era quello di calcolare la quantità di cibo dei Pesci appartenente alla macrofauna (v. Petersen, 1918). In seguito fu misurata anche la quantità degli animali di minor mole, la cosiddetta meiofauna. Questi animali sono molto più numerosi, ma relativamente al peso costituiscono una piccola percentuale rispetto alla macrofauna. D'altra parte la velocità di accrescimento degli animali più piccoli è molto più alta e in un anno si producono parecchie generazioni, mentre di norma la durata di una generazione per gli animali della macrofauna è di alcuni anni. Da questi fattori deriva che la velocità di produzione della macro- e della meiofauna è quasi uguale nella maggior parte delle aree. Nelle zone poco profonde sottoposte alle maree il peso umido degli animali del fondo può essere molto elevato, fino a parecchi kg/m2. Invece sul fondo del Mare del Nord si trova un peso molto inferiore.
Fino al periodo successivo alla seconda guerra mondiale, con questo tipo di ricerche quantitative si sono sondate profondità di poche centinaia di metri. Durante la spedizione danese della Galathea (1950-1952) furono fatti campionamenti a profondità superiori, usando l'apparecchio di Petersen (v. Spärck, 1951) e ne risultò una media di circa 10 animali per m2 e un peso umido totale di circa 1 grammo. Ciò rivela una densità sorprendente e indica che forse la situazione alimentare negli abissi non è così povera come si pensava. Tuttavia nulla sappiamo sulla velocità di accrescimento, che probabilmente è molto bassa. A differenza degli animali pelagici di profondità, che sono carnivori, la maggior parte degli animali che vivono sul fondo marino si nutrono di sospensioni o detriti (v. Knudsen, 1970).
6. Regioni faunistiche e masse d'acqua.
a) Plancton.
In un articolo sull'oceanografia biologica non si può fare a meno di ricordare sia pur brevemente gli elementi faunistici e floristici che caratterizzano le varie masse d'acqua oceaniche. Queste sono solitamente classificate dagli oceanografi fisici in base al rapporto temperatura-salinità, stabilito dalle curve T-S. Vi sono però altri fattori importanti, per esempio la durata dei massimi e dei minimi di temperatura e le competizioni interspecifiche.
Lo studio della distribuzione geografica di varie specie, sia animali sia vegetali, ebbe inizio molti anni orsono, ma la situazione si fece più chiara dopo che gli oceanografi fisici ebbero sviluppato le tecniche adeguate per caratterizzare le masse d'acqua. Il pioniere tra i biologi fu Russell (v., 1939), il quale dimostrò che alcune specie preferiscono le acque dell'Oceano Atlantico, altre invece quelle del Mare del Nord. Come si prevedeva, gli organismi planctonici si sono dimostrati i migliori indicatori del tipo di acqua.
Johnson e Brinton (v., 1963) hanno pubblicato un'eccellente rassegna sullo zooplancton, sottolineando il fatto che parecchie specie con ampia distribuzione di vari caratteri sono state suddivise in sottospecie proprio in base alla loro presenza in masse d'acqua diverse.
Esula dallo scopo del nostro articolo entrare in particolari, ma è necessario ricordare che nelle acque del Pacifico e dell'Atlantico vicine alla superficie, al largo delle coste, si possono facilmente distinguere tre zone, servendosi di due elementi: le masse d'acqua e le specie presenti. Le tre zone sono designate con i seguenti nomi: subartica, centrale ed equatoriale. Una quarta zona faunistica si trova nelle acque temperate, al limite settentrionale della regione centrale. Si tratta di una zona di transizione tra la fauna centrale e quella subartica, ma parecchie specie a vasto areale hanno qui la loro più alta concentrazione e vi sono anche diverse specie endemiche caratteristiche di questa zona. Nell'Oceano Antartico s'individuano varie zone, i cui limiti sono segnati dall'Antartide, dal confine del park, dalla convergenza antartica e da quella subtropicale.
Molte specie planctoniche in grado di riprodursi solamente all'interno di determinate masse d'acqua possono essere trasportate dalle correnti in altre masse, in cui possono sopravvivere per qualche tempo. Esse sono buoni indicatori del trasporto dell'acqua, come è stato riconosciuto anche dagli oceanografi fisici. È opportuno ricordare che il botanico svedese P. T. Cleve, al volgere del secolo passato, si era già servito della distribuzione geografica delle specie di alghe planctoniche per le ricerche sul movimento dell'acqua nei mari dell'Europa settentrionale: tuttavia si è visto in seguito che egli aveva dato eccessivo peso all'influenza delle correnti sulla distribuzione delle specie.
b) Gli animali dei fondi oceanici.
Gli animali che vivono sul fondo a maggiore o minore profondità, presso le coste, sono stati studiati intensamente per più di cento anni, ma sono esclusi dalla presente rassegna, che si prefigge altri scopi. Lo studio degli animali che vivono a grande profondità è stato intrapreso soltanto dopo la seconda guerra mondiale. Nel tracciare il profilo che segue ci siamo attenuti alle idee di Knudsen (v., 1970).
È ormai generalmente accettato che la fauna dei piani abissali sia distinta da quella delle fosse, la cosiddetta fauna adale (v. Bruun, 1955). Mentre la fauna abissale sembra essere più o meno uniforme nell'ambito di vaste estensioni, la maggior parte delle specie adali sono diverse da una fossa all'altra; sembra inoltre che le specie presenti nelle parti più profonde della zona abissale abbiano, con qualche eccezione, una distribuzione più limitata delle specie che vivono nelle parti superiori di tale zona. Non sembra che il fenomeno della bipolarità ricorra nella distribuzione degli Invertebrati bentonici abissali. Per alcune specie il Pacifico centrale costituisce una barriera alla diffusione, forse a causa della povertà, per esse, del nutrimento ivi presente.
Dato che le acque artiche poco profonde non albergano alcuna specie abissale, è evidente che deve esistere un'efficace barriera ecologica, diversa dalla temperatura, che separa la fauna abissale dalla fauna artica di acque poco profonde che vive nelle stesse condizioni di temperatura; può darsi che la pressione idrostatica sia un fattore determinante.
7. Batteriologia oceanica.
Molti esperti batteriologi si sono dedicati nel nostro secolo ai problemi dell'oceano. La sistematica è già a un punto relativamente avanzato e sono stati chiariti molti problemi particolari, per esempio gli effetti della pressione. Però, se consideriamo gli aspetti dinamici - i più importanti per l'oceanografia biologica - dobbiamo ammettere che le conoscenze sono ancora a un livello piuttosto basso: non si è neppure stabilito con certezza quale sia il numero delle cellule batteriche presenti nell'oceano alle varie profondità, e si mettono in dubbio le valutazioni finora eseguite. Un serio inconveniente sta nel fatto che i batteri in bottiglia si comportano molto diversamente da quelli liberi nelle masse d'acqua, così che gli esperimenti di laboratorio hanno scarsa validità. Vari indizi lasciano supporre, comunque, che una nuova strada stia per aprirsi nella batteriologia oceanica. Perciò mi sembra opportuno tralasciare tutti i particolari e le tappe storiche riguardanti questo ramo specialistico, ma molto importante, dell'oceanografia.
8. Conclusione.
La tabella che riproduciamo rappresenta in sintesi le informazioni raccolte da Bogorov (v., 1969).
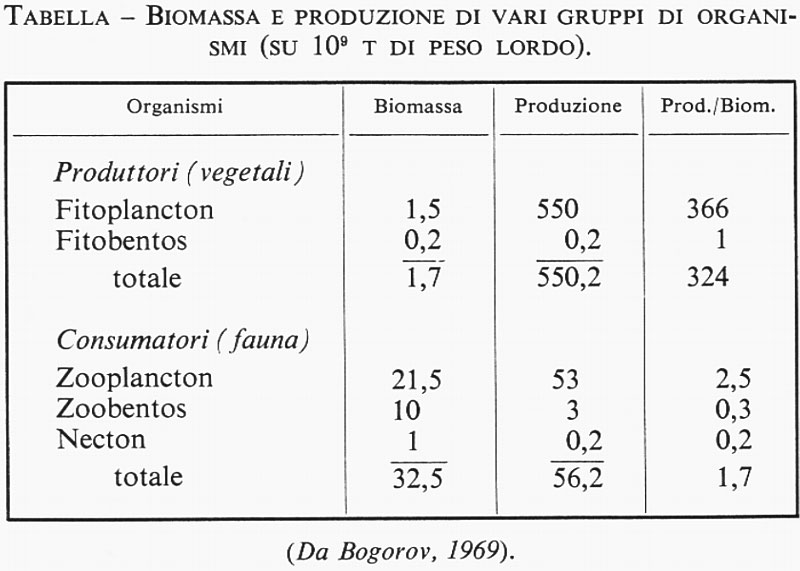
A differenza dei continenti, in cui la massa vegetale supera di oltre 1.000 volte la massa animale, negli oceani la biomassa animale è di circa 20 volte superiore a quella vegetale. Ciò dipende dal fatto che le alghe planctoniche producono giornalmente una quantità di sostanza organica che eguaglia la loro biomassa. Il rapporto fra la produzione di alghe marine e quella animale è dell'ordine di 10:1.
L'oceanografia biologica si è sviluppata rapidamente durante il sec. XX e dobbiamo considerare molto importante il fatto che si possa oggi compilare una tabella come la precedente. Anche se nei particolari ci potrà essere qualche inesattezza, il principio generale sarà difficilmente criticabile da parte degli esperti contemporanei.
Bibliografia.
Atkins, W. R. G., The hydrogen ion concentration of sea water in its biological relations, in ‟Journal of the Marine Biological Association of the United Kingdom", 1922, XII, pp. 717-771.
Atkins, W.R.G., The phosphate content of fresh and salt waters in its relationship to the growth of algal plankton, in ‟Journal of the Marine Biological Association of the United Kingdom", 1923, XIII, pp. 119-150.
Bogorov, V. G. (a cura di), Biology of the Pacific Ocean, vol. I, Plankton, Moskva 1967.
Bogorov, V. G., Productivity of the Ocean; primary production and its utilization in food chains, in Morning review lectures of the Second International Oceanographic Congress, Moskva 1966, UNESCO 1969 pp. 117-122.
Braarud, T., Kleem, A., Hydrographical and chemical investigations in the coastal waters off Möre and in the Romsdalfjord, in ‟Hvalrådets skrifter", 1931, I, pp. 1-88.
Brand, T. von, Methods for the determination of nitrogen and carbon in small amounts of plankton, in ‟Biological bulletin", 1935, LXIX, pp. 221-232.
Brandt, K., Über den Stoffwechsel im Meere, in ‟Wissenschaftliche Meeresuntersuchungen der Kommission zur wissenschaftlichen Untersuchung der deutschen Meere", Abt. Kiel, 1899, IV, pp. 213-230.
Buun, A. F., The ecological zonation of the deep sea, in Proceedings of the UNESCO symposium on physical oceanography, 1955, Tokyo 1955, pp. 160-168.
El-Sayed, S. N., On the productivity of the southern ocean. Symposium on Antartic biology held at Cambridge University in August 1968, Cambridge 1970, pp. 119-134.
Fairbridge, R. W. (a cura di), The encyclopedia of oceanography, in Encyclopedia of earth sciences, vol. I, New Yok 1963.
Gaarder, T., Gran, H. H., Investigations of the production of plankton in the Oslo fjord, in ‟Rapport et procès-verbaux des réunions, Conseil permanent international pour l'exploration de la mer", 1927, XLII, pp. 3-31.
Grontved, J., On the productivity of microbenthos and phytoplankton in some danish fjords, in ‟Meddelser fra Danmarks Fiskeri-og Havundergogelser (N.S.)", 1960, III, pp. 55-92.
Hamilton, R. D., Holm-Hansen, O., Strickland, J. D. H., Notes on the occurrence of living microscopic organisms in deep water, in ‟Deep sea research", 1968, XV, pp. 651-656.
Hansen, V. K., Andersen, K. P., Sampling the smaller zooplankton, in ‟Rapport et procès-vebaux des réunions. Conseil permanent international pour l'exploration de la mer", 1962, CLIII, pp. 39-47.
Harvey, H. W., Nitrate in the sea, in ‟Journal of the Marine Biological Association of the United Kingdom", 1926, XIV, pp. 71-88.
Hentschel, E., Allgemeine Biologie des Südatlantischen Ozeans, in ‟Wissenschaftliche Ergebnisse der deutschen Atlantischen Expedition auf dem Vermessungs- und Forschungsschiff ‛Meteor', 1925-1927", 1933-1936, IX, pp. 1-343.
Hentschel, E., Wattenberg, H., Plankton und Phosphat in der Oberflächenschicht des Südatlantischen Ozeans, in ‟Annalen der Hydrographie und maritimen Meteorologie", 1930, LVIII, pp. 273-277.
Jerlov, N. G., Particle distribution in the ocean, in ‟Report of the Swedish deep sea expedition, 1947-1948", 1953, III, pp. 73-125.
Jespersen, P., Quantitative investigations on the distribution of macroplankton in different oceanic regions, in ‟Dana reports", 1935, VII, pp. 1-44.
Johnson, M. W., Brinton, E., Biological species, watermasses and currents, New York-London 1963.
Ketchum, B. H., Biochemical relations between marine organisms and their environments, in ‟Ecological monographs", 1947, XVII, pp. 309-315.
Ketchum, B. H., The regeneration of utrients, in ‟Rapport et procès-verbaux des réunions. Conseil permanent international pour l'exploration de la mer", 1962, CLIII, pp. 142-147.
Keys, A., Christensen, E. H., Krogh, A., Organic metabolism of sea-water, in ‟Journal of the Marine Biological Association of the United Kingdom", 1935, XX, pp. 181-196.
Knudsen, J., The systematics and biology of abyssal and hadal Bivalvia, in ‟Galathea report", 1970, XI, pp. 7-238.
Krey, J., Bestimmung des Chlorophylls in Meerwasserschopfproben, in ‟Journal du conseil. Conseil permanent international pour l'exploration de la mer", 1939, XIV, pp. 201-209.
Krey, J., Quantitative Bestimmung von Eiweiss im Plankton mittels der Biuretreaktion, in ‟Kieler Meeresforschungen", 1951, VIII, pp. 16-29.
Lohmann, H., Untersuchungen zur Festsellung des vollständigen Gehaltes des Meeres an Plankton, in ‟Wissenschaftliche Meeresuntersuchungen der Kommission zur wissenschaftlichen Untersuchung der deutschen Meere", Abt. Kiel, 1908, X, pp. 129-370.
Lohmann, H., Die Bevölkerung des Ozeans mit Plankton nach den Zentrifugenfänge während der Ausreise der Deutschland' 1911. Zugleich ein Beitrag zur Biologie des Atlantischen Ozeans, in ‟Archiv für Biontologie", 1920, IV, pp. 1-617.
Nathansohn, A., Über die Bedeutung vertikaler Wasserbedingungen für die Produktion des Planktons im Meere, in ‟Abhandlungen der mathematisch-physikalischen Klasse der Königlichen Sächsischen Gesellschaft der Wissenschaften", 1906, XXXIX.
Nathansohn, A., Über die allgemeinen Produktionsbedingungen im Meere, in ‟Revue internationale d'hydrobiologie et d'hydrographie", 1908, I, pp. 37-72.
Odum, H. T., Primary production in eleven Florida springs and a marine turtlegrass community, in ‟Limnology and oceanography", 1957, II, pp. 85-97.
Petersen, C. G. J., The sea bottom and its production of fish-food. (A survey of the work done in connection with valuation of the Danish waters from 1883-1917), in ‟Report of the Danish biological station to the board of agriculture", 1918, XXV, pp. 1-62.
Prasad, R. R., Banerji, S. K., Nair, P. V. R., A quantitative assessment of the potential fishery resources of the Indian Ocean and adjoing seas, in ‟Indian journal of animal sciences", 1970, XL, pp. 73-98.
Raymont, J. E. G., Plankton and productivity in the oceans, in ‟International series monograph of pure and applied biology. Division of zoology", 1963, XVIII, p. 660.
Richards, F. A., Thompson, T. G., The estimation and characterization of plankton populations by pigment analyses. II. A spectrophotometric method for the estimation of plankton pigments, in ‟Journal of marine research", 1952, XI, pp. 156-172.
Russell, F. S., Hydrographical and biological conditions in the North Sea as indicated by plankton organisms, in ‟Journal du conseil. Conseil permanent international pour l'exploration de la mer", 1939, XIV, pp. 171-192.
Ryther, J. H., Organic production by plankton Algae and its environmental control, in The ecology of Algae, Pittsburgh 1960, pp. 72-83.
Ryther, J. H., Geographic variations in productivity, in The sea. Ideas and observations on progress in the study of the sea (a cura di M.N. Hill), vol. II, The composition of sea water, comparative descriptive oceanography, New York 1963, pp. 347-380.
Schaefer, M. B., Beverton, J. H., Fishery dynamics. Their analysis and interpretation, in The sea. Ideas and observations on progress in the study of the sea (a cura di M.N. Hill), vol. II, The composition of sea water, comparative descriptive oceanography, New York 1963, pp. 464-483.
Spärck, ., Density of bottom animals on the ocean floor, in ‟Nature", 1951, CLXVIII, p. 112.
Steemann Nielsen, E., On the relation between the quantities of phytoplankton and zooplankton in the sea, in ‟Journal du conseil. Conseil permanent international pour l'exploration de la mer", 1937, XII, pp. 147-153.
Steemann Nielsen, E., Use of radio-active carbon (C14) for measuring organic production in the sea, in ‟Journal du conseil. Conseil permanent international pour l'exploration de la mer", 1952, XVIII, pp. 117-140.
Steemann Nielsen, E., Productivity of the oceans, in ‟Annual review of plant physiology", 1960, XI, pp. 31-362.
Steemann Nielsen, E., The relationship between phytoplankton and zooplankton in the sea, in ‟Rapport et procès-verbaux des réunions. Conseil permanent international pour l'exploration de la mer", 1962, CLIII, pp. 178-182.
Steemann Nielsen, E., Productivity, definition and measurement, in The sea. Ideas and observations on progress in the study of the sea (a cura di M.N. Hill), vol. II, The composition of sea water, comparative descriptive oceanography, New York 1963, pp. 129-164.
Steemann Nielsen, E., Jensen, E. A., Primary oceanic production. The autotrophic production of organic mattert in the oceans, in ‟Galathea report", 1957, I, pp. 49-135.
Strickland, J. D. H., Reserach on the marine food chain. Part IV. Research on the marine planktonic food web at the Institute of Maine Resources. A review of the past six years work, University of California Institute of Marine Resources, IMR Report, 1970, LXX, 5, pp. 1-135 (manoscritto non pubblicato).
Sverdrup, H.N., On conditions for the vernal blooming of phytoplankton, in ‟Journal du conseil. Conseil pemanent international pour l'exploration de la mer", 1953, XVIII, pp. 287-295.
Sverdrup, H.N., Johnson, M. W., Fleming, R. H., The oceans, their physics chemistry and general biology, Englewood Cliffs, N. J., 1942.
Utermöhl, H., Neue Wege in der quantitativen Erfassung des Planktons, in ‟Internationale Vereinigung für theoretische und angewandte Limnologie, Verhandlungen", 1932, V, pp. 567-595.
Vinogradov, M. E., The feeding of deep-sea zooplankton, in ‟Rapport et procès-verbaux des réunions. Conseil pemanent international pour l'exploration de la mer", 1962, CLIII, pp. 114-120.
Vinogradov, M. E., Voronina, N., The distribution of different groups of plankton in accordance with their trophic level in the Indian Equatorial Current area, in ‟Rapport et procès-verbaux des réunions. Conseil permanent international pour l'exploration de la me", 1962, CLIII, pp. 200-204.
Williams, P. M., Oeschger, H., Kinney, P., The natural radiocarbon activity of the dissolved organic carbon in the northeast Pacific Ocean, in ‟Nature", 1969, CCXXIV, pp. 256-258.
Zobell, C. E., Anderson, D. Q., Observations on the multiplication of Bacteria in different volumes of stored sea water, in ‟Biological bulletin. Marine biological laboratory, Woods Hole, Mass.", 1936, LXXI, pp. 324-342.
Oceanografia fisica
SOMMARIO: 1. Introduzione. □ 2. I bacini oceanici. □ 3. Proprietà fisiche e chimiche dell'acqua di mare. □ 4. Distribuzione della temperatura, della salinità e della densità nell'oceano: a) caratteristiche di superficie; b) caratteristiche delle profondità oceaniche. □ 5. Correnti oceaniche. □ 6. Studi teorici. □ 7. Interazione aria-mare. □ 8. Onde. □ 9. Maree. □ 10. Come modificare l'oceano. □ Bibliografia.
1. Introduzione.
Dopo la grande spedizione del vascello Challenger nel 1872-1876, fu compiuta qualche altra crociera dello stesso genere, ma l'interesse del pubblico per l'oceano era ormai venuto meno ed era molto più attratto dalle esplorazioni polari; nel 1900, in realtà, l'oceanografia destava scarso interesse e i pochi scienziati che studiavano il mare non avevano sufficienti mezzi.
Ci fu bensì un ritorno d'interessi dopo l'affondamento del Titanic, che però la guerra del 1914-1918 stroncò, cosicché furono pochi i paesi, tra le due guerre, che continuarono a esplorare gli oceani. A questo periodo, terminato solo nel 1939, appartengono la spedizione della Meteor e le ricerche della Discovery.
La seconda guerra mondiale, con la minaccia dei sottomarini, gli sbarchi dei mezzi anfibi e l'aumentata dipendenza dalla scienza, cambiò rapidamente l'atteggiamento ufficiale nei confronti dell'oceanografia. Da allora quasi tutti gli Stati hanno mostrato un certo interesse per la scienza del mare, in parte per ragioni militari, in parte per l'importanza pratica che il mare ha come fonte di cibo, deposito di rifiuti, via di comunicazione e regolatore del clima.
Col passare del tempo, via via che si impara di più sui processi oceanici e sulla loro interazione con l'atmosfera, diviene evidente che esiste la possibilità di servirsi con intelligenza dell'oceano. Si sta lavorando su molti problemi, per esempio su quelli suscitati dalla crisi energetica. Esiste un futuro per l'energia ricavata dal moto ondoso? Sarà necessario eliminare scorie fortemente radioattive? È in grado l'oceano di assorbire l'anidride carbonica prodotta dal combustibile fossile bruciato senza posa dall'uomo? Cambierà rapidamente il clima?
Tutti questi problemi, e molti altri ancora, richiedono una più approfondita conoscenza dell'oceano e dei suoi complicati processi.
2. I bacini oceanici.
Il primo e più ovvio fatto da notare è che il mare ha un'estensione quasi doppia della terraferma (circa il 71% della Terra è coperto dai mari). L'Oceano Pacifico da solo ricopre il 36% del nostro pianeta, l'Oceano Atlantico il 21% e l'Oceano Indiano il 14%. È inoltre noto che il rapporto acque-terraferma è molto maggiore nell'emisfero meridionale che non in quello settentrionale.
I moderni geofisici si sono serviti della teoria della tettonica a zolle per studiare lo sviluppo dei bacini oceanici e altri caratteri topografici primari della Terra. In seguito all'intensa ricerca geofisica degli ultimi tempi essi sono in grado di datare i vari stadi della formazione dell'Atlantico e degli altri oceani e hanno acquisito una notevole conoscenza dei processi implicati. Le configurazioni maggiori sono da certi punti di vista comprese più facilmente di quelle minori: corpi d'acqua come il Mediterraneo, il Mar dei Caraibi, il Mare di Bering, ben delimitati dalla terraferma o da catene di isole, presentano una problematica ancora aperta per quanto riguarda la loro origine, oltre a rivestire particolare interesse oceanografico. La grande estensione in senso orizzontale dell'oceano balza subito agli occhi, mentre le conoscenze sulla sua estensione verticale sono state raccolte lentamente. I primi oceanografi, come per esempio quelli della Challenger, effettuavano faticosi sondaggi per mezzo di funi zavorrate. Questo metodo richiedeva molto tempo ed era spesso impreciso, poiché le correnti oceaniche allontanavano le funi dalla verticale e a profondità che non fossero modeste era spesso impossibile capire quando il peso aveva toccato il fondo. Lo sviluppo di sistemi di cavi sottomarini per l'invio di messaggi condusse al miglioramento delle tecniche di sondaggio con l'adozione di cavi metallici di diametro ridotto al posto delle funi: si poté così rilevare che la profondità media dell'oceano è di circa 4.000 metri. L'oceano è quindi poco profondo rispetto alla sua estensione orizzontale, e va considerato alla stregua di una sottile pelle sulla superficie della Terra.
Gli affascinanti dettagli della topografia sottomarina non poterono essere studiati fino all'invenzione dell'ecoscandaglio acustico (brevettato quasi immediatamente dopo l'affondamento del Titanic durante il suo viaggio inaugurale nel 1913). A seguito degli sviluppi tecnologici avvenuti nel corso degli anni, soprattutto durante e dopo la seconda guerra mondiale, si è messo a punto un ecoscandaglio di precisione in grado di misurare il tempo impiegato da un impulso sonoro per andare da una nave al fondale marino e ritornare indietro, con una precisione di circa un millisecondo, che corrisponde a una precisione, nella misura della profondità, di un metro o poco più.
L'immagine globale che emerge da un gran numero di sondaggi oceanici è che a partire dalla linea costiera verso il largo c'è una regione piatta e poco profonda nota come piattaforma continentale. Questa regione ha una profondità media di poco più di 100 metri e un ampiezza variabile, ma in media inferiore a 100 chilometri. Generalmente sabbiosa, la piattaforma continentale presenta lievi variazioni di profondità e occasionalmente dà luogo a secche o isole al largo. È una regione di grande utilità pratica e commerciale: vi si trovano i porti e la maggior parte dei punti di pesca, e costituisce anche la zona di scarico per i liquami e per gli altri rifiuti dei paesi industrializzati. In data relativamente recente si è incominciato anche a estrarre da essa il petrolio sottomarino. Non è sorprendente quindi che questa zona sia oggetto di interessi internazionali, né che la Conferenza delle Nazioni Unite sulle leggi del mare stia compiendo un particolare sforzo per regolare la cosiddetta Zona Economica Esclusiva, che in pratica è la piattaforma continentale stessa.
Al largo della piattaforma continentale il fondale marino scende rapidamente, con pendenze dell'ordine di 1:10, fino al raggiungimento dei fondali oceanici a 4.000 metri circa. Il materiale che forma la scarpata è generalmente fango, ma con affioramenti rocciosi e grande varietà di forme, taltalvolta impressionanti, come nel caso dei canyons sottomarini. Si tratta di ripide vallate distribuite dappertutto, ma che sono per lo più associate alle coste in cui sboccano fiumi.
I fondali oceanici sono la regione più estesa del nostro pianeta, dato che in più di tre quarti dell'oceano si trovano profondità tra i 3.000 e i 6.000 m. Solamente l'1% del fondo oceanico ha profondità maggiori di 6.000 m, generalmente nella regione delle fosse, e le massime profondità (circa 11.000 m) sono paragonabili alle massime altezze sulla terraferma (circa 9.000 m). Il fondale oceanico mostra tuttavia una topografia assai varia: ci sono monti, valli e pianure. Il rilievo più esteso è la grande dorsale che si estende dal sud della Groenlandia lungo il centro dell'Oceano Atlantico da nord a sud, e poi attraverso l'Oceano Indiano e l'Oceano Pacifico. La dorsale oceanica è una configurazione di fondamentale importanza nella teoria della tettonica a zolle. Le configurazioni opposte sono le fosse, che sono le regioni più profonde dell'oceano. Esse si trovano soprattutto nell'Oceano Pacifico (fosse delle Aleutine, delle Curili, delle Filippine e delle Marianne), ma ne esistono anche nell'Oceano Atlantico (fosse di Portorico e delle Sandwich Australi) e nell'Oceano Indiano (fossa della Sonda).
Esistono anche grandi aree completamente pianeggianti, note come pianure abissali, e montagne isolate, veri monti marini, alcune delle quali si ergono al di sopra della superficie del mare a formare le isole oceaniche.
Il materiale dei fondali oceanici è spesso costituito da particelle molto fini, in gran parte formatesi nel mare stesso, piuttosto che dal materiale più grossolano portato dai fiumi o dai venti dalla terra alla scarpata e alla piattaforma continentale. Pertanto la velocità media di sedimentazione sui fondali è bassa e una carota di materiale raccolto negli abissi può fornire informazioni sulla sedimentazione e sul clima di milioni di anni fa.
3. Proprietà fisiche e chimiche dell'acqua di mare.
I bacini oceanici contengono una complessa soluzione acquosa di sali; vale la pena di dedicare un po' di tempo a considerare le proprietà del solvente. Noi diamo comunemente per scontate le proprietà dell'acqua, ma l'acqua è una sostanza eccezionale: essa è uno dei pochi liquidi inorganici esistenti in natura e per di più è una delle poche sostanze che si riscontrino in natura in tutte e tre le fasi, cioè allo stato gassoso, liquido e solido (v. acqua; v. tab. I per le proprietà più caratteristiche). Forse le proprietà più importanti sono il calore specifico e il calore latente, entrambi assai elevati. Essi fanno sì che occorrano grandi quantità di calore per far aumentare la temperatura dell'acqua e per farla fondere o evaporare. Il risultato è che l'acqua è un fluido eccellente per il condizionamento dell'aria grazie all'elevata capacità di accumulare calore. L'abbassamento della temperatura corporea superficiale in seguito alla sudorazione è un effetto delle caratteristiche fisiche dell'acqua; le stesse proprietà, attraverso un meccanismo più complesso, controllano la temperatura della Terra. Tutti sanno cosa sono i climi continentali, con calde estati e inverni rigidi, tipici della Siberia, e i climi oceanici delle isole e delle regioni costiere, dove l'accumulo di calore da parte dell'oceano modera l'escursione termica e mantiene entro limiti ristretti la variazione della temperatura nel corso di tutto l'anno.
Il ghiaccio è più leggero dell'acqua, vi galleggia sopra e la isola da ulteriori perdite di calore. Nonostante le variazioni stagionali, si mantiene così nel mare un ambiente a temperatura quasi costante, nel quale gli esseri viventi possono sopravvivere più facilmente.
Ci sono molte altre proprietà dell'acqua - la capacità di solvente, la costante dielettrica, la tensione superficiale - che fanno sì che tutte le reazioni essenziali per la vita avvengano rapidamente; la maggior parte di esse non è modificata di molto dai sali disciolti che distinguono l'acqua di mare dall'acqua dolce.
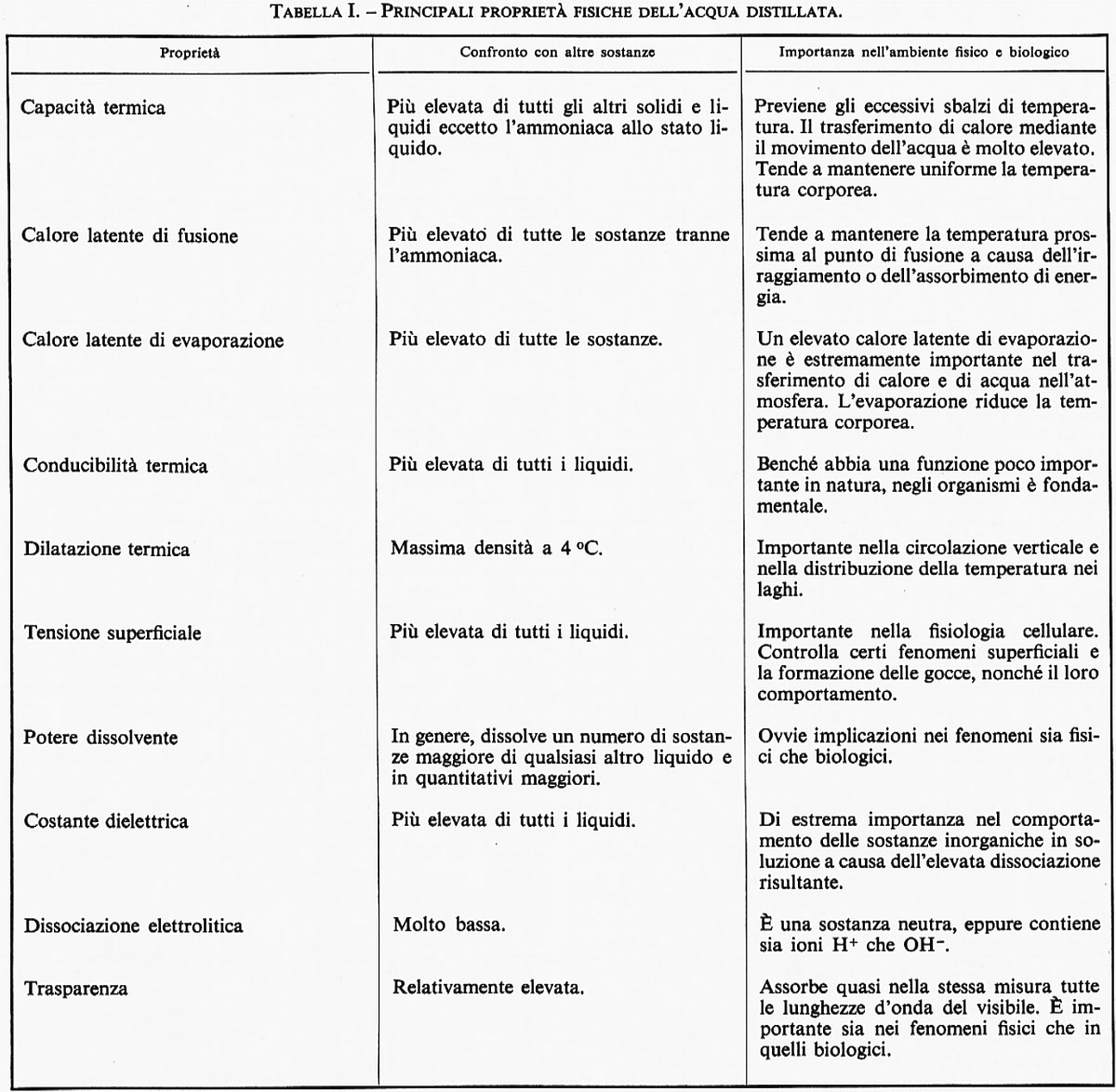
L'acqua di mare è una soluzione complessa che probabilmente contiene tutti gli elementi; le attuali tecniche analitiche ne hanno identificato circa la metà. Alcuni dei più abbondanti sono elencati nella tab. II. Gli altri sono presenti in concentrazione di circa una parte su un milione, o anche meno. Si è trovato che campioni di acqua di mare prelevati da quasi ogni parte dell'oceano aperto contengono i principali costituenti riportati nella tab. II in proporzioni pressoché costanti. Nel corso delle ere geologiche i diversi componenti chimici sono stati molto ben mescolati tra di loro e non esiste alcun processo efficace che li possa separare l'uno dall'altro. Ne consegue che, così come possiamo pensare alla miscela uniforme dei gas atmosferici come ‛aria', allo stesso modo possiamo considerare l'acqua di mare come una miscela di acqua dolce con un cosiddetto ‛sale marino' consistente di un miscuglio uniforme degli elementi riportati nella tab. II. La quantità totale di materiale disciolto nell'acqua di mare è detta salinità: essa viene misurata in grammi per chilogrammo di acqua di mare e generalmente indicata con S‰. Poiché molte delle più importanti proprietà dell'acqua di mare dipendono dalla salinità, è necessario poterla misurare con precisione. Dato che la composizione è pressoché costante, si può misurare l'abbondanza di uno qualsiasi dei componenti principali e dedurne la salinità. In passato ci si serviva comunemente dello ione cloruro (‛clorurazione') e la salinità era generalmente stimata mediante titolazione con nitrato d'argento. Al giorno d'oggi di solito si deduce la salinità misurando la conducibilità di un campione di acqua di mare a temperatura nota e usando una relazione nota (empirica) che lega salinità e conducibilità. Con questo metodo si riesce a stimare la salinità con una precisione fino a +0,003‰, il che equivale a una precisione di circa una parte su diecimila per quanto riguarda il sale.
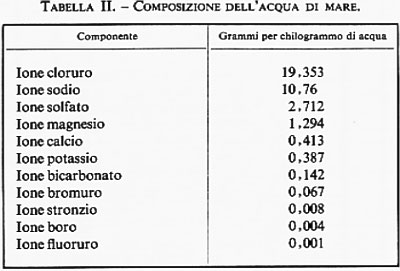
Le proprietà dell'acqua dolce dipendono dalla temperatura e dalla pressione; quelle dell'acqua di mare non dipendono solamente dalla temperatura e dalla pressione, ma anche dalla salinità. Forse la più importante proprietà dell'acqua di mare è la densità: questo perchè l'acqua dell'oceano, come qualsiasi altro fluido, tende a muoversi in maniera che la parte più densa si trovi in fondo e quella più leggera in superficie. La relazione tra densità e pressione, temperatura e salinità è chiamata ‛equazione di stato'. Si tratta di una relazione complessa che non è ancora stata dedotta teoricamente, ma che è stata ricavata dal- l'osservazione. L'effetto della pressione, o compressibilità, non è tanto elevato quanto si potrebbe supporre. L'acqua di mare è quasi incomprimibile e la sua densità aumenta solo di una minima percentuale a pressioni corrispondenti alle massime profondità dell'oceano.
Questa quasi incompressibilità dell'oceano fa sì che in genere un corpo solido o galleggi in superficie o cali sul fondo. È possibile, mediante una progettazione molto accurata, costruire un corpo che galleggi a una profondità intermedia.
Per molte applicazioni pratiche in oceanografia si può prescindere dall'effetto della pressione e confrontare le densità rapportandole alla stessa pressione, generalmente quella atmosferica che si ha alla superficie del mare. La relazione empirica che lega temperatura, salinità e densità (a 1 atmosfera) è riprodotta nella fig. 3. Prendendo una particolare linea di densità, si può vedere che essa cambia pendenza quando si passa da temperature più alte a temperature più basse. Questo significa che alle basse temperature (ai poli) è il cambiamento di salinità e non quello di temperatura che fa cambiare la densità. Alle alte temperature (ai tropici) la situazione è rovesciata.
L'acqua dolce ha la massima densità a 4 °C; col crescere della salinità la temperatura di massima densità si abbassa e altrettanto fa il punto di solidificazione, ma, come si vede nella fig. 4, il risultato è che l'acqua la cui salinità è maggiore di 24,69‰ continua a diventare più densa finché non si solidifica. L'acqua di mare ha mediamente una salinità di circa 35‰ e quindi deve raffreddarsi fino a quasi −2 °C prima di divenire solida.
Un'altra proprietà assai importante dell'acqua di mare è la capacità di assorbire le radiazioni elettromagnetiche. La componente più penetrante è la luce blu-verde, di lunghezza d'onda di circa 0,5 μm. Essa è fortemente assorbita anche in acqua pura: la sua energia è ridotta di metà dopo circa 35 m di percorso. Nell'acqua dell'oceano la sua penetrazione è molto minore, giacché l'energia viene pressoché dimezzata dopo 5 m in acqua limpida e dopo soli 0,5 m nelle acque costiere, generalmente più torbide. La riduzione della penetrazione è dovuta tanto alle particelle in sospensione quanto al materiale disciolto.
In acque trasparenti e pulite, come si è già detto, la luce blu-verde è quella che penetra a profondità maggiore. Le lunghezze d'onda più corte (verso l'ultravioletto) e quelle più lunghe (rosso e infrarosso) sono più fortemente assorbite.
Questo può essere facilmente osservato dai sommozzatori e illustrato dalle fotografie subacquee a colori, prese senza l'uso di illuminazione artificiale. Gli oggetti rossi o gialli appaiono sempre più scuri man mano che si scende, mentre i blu e i verdi mantengono il loro colore. Sia i pesci abissali che gli esseri umani (i cui antenati provenivano dal mare) hanno la massima sensibilità visiva alle lunghezze d'onda prossime a 0,5 μm, che penetrano più in profondità di tutte le altre nell'acqua marina limpida.
Siccome la maggior parte dell'energia solare raggiunge la superficie terrestre sotto forma di radiazione visibile (il 99% ha lunghezze d'onda inferiori a 4 μm), essa non penetra molto profondamente nell'acqua. Anche nelle acque più limpide meno dell'10/0 della luce solare raggiunge i 100 m di profondità; nell'acqua torbida in prossimità delle coste, poi, è quasi completamente assorbita nei primi 10 m. Il Sole agisce quindi solamente su un sottile strato in superficie.
Vedremo più avanti come ciò abbia importanti effetti sulla temperatura e sulle correnti oceaniche, ma qui dobbiamo sottolineare il fatto ovvio, importante ma spesso dimenticato, che l'oceano è buio. Fatta eccezione per i lampi, energeticamente insignificanti, prodotti dagli organismi bioluminescenti, non c'è alcuna luce ambientale. Questo è noto da molti anni, fin da quando gli oceanografi scoprirono che è possibile esporre per molte ore a mezz'acqua delle lastre fotografiche senza che si impressionino minimamente.
L' opacità dell'oceano alle radiazioni elettromagnetiche non si limita alla sola luce visibile. Lunghezze d'onda minori (raggi X e ultravioletti) e maggiori (infrarossi, radar e microonde) sono tutte anche più fortemente assorbite. Le onde radio molto lunghe possono penetrare per una certa profondità (pari a circa mezza lunghezza d'onda), ma il sistema più efficace di trasmissione nel mare è quello che si avvale di onde sonore.
Per esse infatti la situazione è diversa: possono essere trasmesse nell'oceano con perdite molto minori che nell'atmosfera e per questo motivo l'uomo e le creature marine si servono dei suoni per ottenere informazioni. Mediante l'ecoscandaglio la profondità del mare è calcolata dal tempo che occorre perché un impulso sonoro raggiunga il fondo e sia riflesso indietro. Un principio analogo è sfruttato nell'ASDIC e nel SONAR, ma in questi il fascio non è necessariamente verticale e la distanza non è misurata fino al fondale ma fino a un sottomarino, a un banco di pesci o a qualche altro oggetto che rifletta il suono. Nei primi esperimenti ci si serviva dell'udito per percepire il suono di ritorno, ma questo non è un procedimento efficace, dato che il suono perde molta energia nell'attraversare l'interfaccia aria-acqua. Oggi si usano microfoni e idrofoni per convertire l'energia sonora in un segnale elettrico che può essere amplificato e visualizzato in un grafico. Le condizioni del mare non sono però così uniformi da permettere una riproduzione nitida delle immagini degli oggetti riflettenti. Questo è dovuto in parte al fatto che le sorgenti sonore sono approssimativamente dello stesso ordine di grandezze della lunghezza d'onda (un suono di 10 KHz ha una lunghezza d'onda di circa 15 cm), ma è anche dovuto alla variazione della velocità del suono con la temperatura, la salinità e la pressione. Il percorso del fascio è alterato da lievi disomogeneità presenti nell'oceano e il fascio stesso risulta confuso, per cui non è possibile formare un'immagine nitida. Fluttuazioni analoghe, prodotte sui raggi luminosi dalle disomogeneità presenti nell'atmosfera, causano il luccichio delle stelle.
Un altro effetto è dovuto alle variazioni della velocità del suono, causate soprattutto dalla distribuzione della temperatura, che possono rifrangere il percorso delle onde sonore secondo linee curve invece che rette. L'equivalente atmosferico in questo caso è il miraggio. È ovvio che la distorsione dovuta ai diversi valori assunti nei pressi della superficie dalla velocità del suono (essa stessa strettamente legata alla temperatura) è di grande importanza, nella guerra sottomarina, per la localizzazione e per il computo delle distanze delle unità nemiche. Per questo motivo gran parte dell'attività degli oceanografi militari consiste nel cercare di prevedere la distribuzione della temperatura negli strati superiori dell'oceano.
Sono numerosi gli studi dedicati alle proprietà fisiche dei liquidi e dei gas, in particolare a quelle dell'aria e dell'acqua. Qui ci siamo limitati a esaminare brevemente quelle proprietà dell'acqua di mare che condizionano maggiormente la struttura e il comportamento dell'oceano.
L'aspetto della superficie dell'oceano è ben noto. Dal- l'aria (o dallo spazio) si distinguono la terraferma, il ghiaccio e l'enorme superficie dell'oceano coperta da quelle ondulazioni in movimento che chiamiamo onde. Un osservatore che studi attentamente le linee costiere potrebbe distinguere i movimenti, giornalieri e semigiornalieri, di interi bacini oceanici, che costituiscono le maree ; su questi fenomeni di moto ritorneremo in seguito.
Fotografato in bianco e nero dallo spazio, l'oceano appare nero mentre le nuvole appaiono bianche. Questo perché le nuvole riflettono gran parte della radiazione solare visibile, mentre il mare assorbe la maggior parte dei raggi luminosi e ne riflette solo una piccola frazione. Ripreso a colori e da minori altezze, il mare presenta una gamma di colori ristretta, che va dal blu scuro, al verde, al giallo- verde.
Anche se è difficile misurare ed esprimere in termini quantitativi i colori, e quindi non esistono diagrammi che lo confermino, in genere il colore blu scuro o indaco è tipico dei mari tropicali e di quelle regioni, come il Mar dei Caraibi o il Mediterraneo, in cui la produzione biologica è scarsa (il blu è il colore del mare deserto). A latitudini più elevate il colore è per lo più verde bluastro, e più particolarmente color acquamarina. Le acque polari e costiere sono generalmente verdastre.
La spiegazione dei diversi colori assunti dal mare non è semplice. La ragione principale della colorazione blu risiede nella dispersione selettiva della luce solare ad opera delle molecole: il mare sembra blu per lo stesso motivo per cui il cielo sembra azzurro. Quando l'acqua contiene particelle, soprattutto fitoplancton, c'è meno trasmissione e più dispersione della luce e probabilmente un certo assorbimento da parte delle sostanze organiche solubili prodotte dal plancton. Oltre al colore del mare stesso, è importante anche considerare la luce riflessa. I colori tendono a cambiare in continuazione, a causa del luccichio del sole sulle onde che si infrangono e sulle increspature, a causa del gioco delle ombre e dei riflessi delle nuvole e a causa delle lievi differenze fra regioni increspate e regioni calme della superficie. Se, per esempio, la superficie è cosparsa di una sottile pellicola oleosa, prodotta dagli scarichi di una petroliera o da qualche fonte planctonica, le piccole increspature della superficie risultano smorzate. Le aree lisce sembrano più chiare e riflettono il cielo quasi perfettamente. Quando non esiste una pellicola superficiale la superficie appare più scura, in parte perché la regione frontale di ogni increspatura riflette un punto più alto (e quindi di un blu più scuro) del cielo, in parte perché la riflessione è meno completa. Dato che la superficie del mare è generalmente increspata, c'è quasi sempre un contrasto di colore tra il mare e il cielo all'orizzonte.
Per quanto belli, i colori del mare, si è detto, sono difficili da misurare e di essi si è fatto uno scarso uso scientifico, benché in alcune interessanti ricerche si siano sfruttate le decolorazioni dovute al deflusso dei fiumi, a fioriture di plancton e a veli oleosi, come indicative di movimenti dell'acqua. Tali osservazioni sono ovviamente limitate agli strati superficiali; per poter effettuare ricerche sugli strati più profondi occorrono strumenti che rendano possibile effettuare misurazioni su varie lunghezze d'onda a qualsiasi profondità.
4. Distribuzione della temperatura, della salinità e della densità nell'oceano.
a) Caratteristiche di superficie.
La temperatura dell'oceano in vicinanza della superficie fu una delle prime caratteristiche fisiche a essere misurata. Appena furono disponibili i termometri a mercurio, i naviganti iniziarono a raccogliere campioni di acqua di mare usando semplici secchi e a misurarne la temperatura. Simili rilevamenti per molti anni furono effettuati dalle navi mercantili; i dati ricavati sono stati utilizzati dagli istituti meteorologici per costruire atlanti climatologici. La temperatura superficiale dell'oceano scende da valori prossimi a 30 °C nelle regioni equatoriali a circa −2 °C nelle regioni ghiacciate delle alte latitudini. L'effetto della temperatura sulla densità fa sì che le densità maggiori si riscontrino alle alte latitudini e le densità minori presso l'equatore.
La corrispondente distribuzione superficiale della salinità dà un quadro diverso, legato più alla differenza tra piovosità ed evaporazione che all'insolazione. La salinità cresce andando dalle alte latitudini alle basse, raggiunge un massimo intorno ai 25° di latitudine nord e sud, e poi giunge a un minimo all'equatore. Vi è una precisa indicazione che la salinità è legata al bilancio delle quantità di acqua che si trovano vicine alla superficie dell'oceano, mentre la temperatura è legata al bilancio termico. Il minimo di salinità all'equatore va collegato con le foreste pluviali sulla terraferma, mentre i massimi subtropicali si trovano alle stesse latitudini dei deserti. La distribuzione della salinità è una manifestazione della grande cella atmosferica che produce gli alisei.
b) Caratteristiche delle profondità oceaniche.
Misurare le caratteristiche dell'acqua di mare a una certa profondità nell'oceano è un'impresa assai difficile. Finché ci si occupa della salinità o della concentrazione di altre sostanze chimiche occorrono dei campioni. Questi si ottengono generalmente immergendo bottiglie o sonde appese a un cavo collegato con l'argano di una nave. Una tipica calata può raggiungere qualche migliaio di metri, con dieci bottiglie su un cavo di 4 mm di diametro. Le bottiglie sono calate aperte, per evitare che vengano schiacciate dall'aumento della pressione idrostatica, e poi chiuse in profondità mediante un comando azionato da un peso anulare fatto scendere dalla superficie lungo il cavo; i campioni vengono quindi recuperati per essere analizzati direttamente a bordo della nave oppure a terra.
È difficile misurare la salinità con la necessaria precisione. Nei primi anni di questo secolo il Consiglio internazionale per l'esplorazione del mare ha portato a un'unificazione dei metodi, il più importante dei quali consiste nella valutazione della salinità in base alla determinazione della concentrazione dello ione cloro. La concentrazione di questo ione è rappresentata dalla quantità di cloro contenuta in un dato campione d'acqua, quantità che viene determinata per titolazione del campione di acqua di mare con nitrato d'argento. Per ottenere una precisione accettabile è necessario usare speciali pipette e provette graduate, tarandole per mezzo di ‛acqua di mare campione'. Questa è stata per anni fornita in fiale da un servizio internazionale; si tratta di acqua di mare naturale, filtrata, sterilizzata e diluita in modo da avere una salinità molto prossima al 35‰. La sua concentrazione di ione cloro viene accuratamente determinata e il valore specificato per ogni fiala.
Il metodo funzionava bene perché il rapporto della quantità di ione cloro rispetto alla quantità totale di sostanze disciolte è quasi costante, ma la titolazione era relativamente lenta e difficile da fare sulle navi. Oggigiorno la maggior parte delle misure di salinità sono fatte determinando la conducibilità elettrica del campione di acqua di mare e quindi servendosi della relazione nota tra conducibilità e salinità. Questo metodo è molto più conveniente, benché si debba tener conto della temperatura dell'acqua, che modifica anch'essa la conducibilità. Anche questa apparecchiatura è tarata mediante acqua di mare campione, sebbene si stia lavorando per fissare un campione assoluto di conducibilità dell'acqua di mare.
Il metodo della conducibilità, usato accuratamente, può dare valori della salinità precisi entro lo 0,003‰, mentre il metodo di titolazione (concentrazione dello ione cloro) solitamente dava valori precisi entro lo 0,02‰.
Siccome generalmente la valutazione della densità è importante, è opportuno misurare la temperatura alla stessa profondità a cui si misura la salinità. Quindi di solito alle bottiglie campionatrici sono attaccati dei termometri a rovesciamento d'alto mare. Si tratta di termometri a mercurio costruiti in modo che la colonna di mercurio si spezzi allorché il termometro viene capovolto. La rottura avviene nel tubo capillare, che presenta un corto bracciolo laterale, di modo che dopo che il termometro è stato capovolto esso diviene quasi insensibile agli ulteriori cambiamenti della temperatura. Ciò significa che il termometro può essere recuperato da grandi profondità e la temperatura in situ a quella profondità può essere letta a bordo della nave anche se su questa la temperatura non ha niente a che vedere (di solito è molto più alta) con la temperatura dell'acqua alle profondità oceaniche.
Se il termometro a rovesciamento ha una robusta custodia esterna di vetro che lo protegge dalla pressione idrostatica, la precisione della misura della temperatura, dopo alcune piccole correzioni, è circa di 0,02 °C. Se il termometro non fosse protetto contro la pressione, la temperatura indicata sarebbe in parte dovuta alla temperatura in situ e in parte alla pressione in situ alla profondità del rovesciamento. Pertanto si può usare una coppia di termometri, uno protetto contro la pressione e uno non protetto, per determinare sia la temperatura che la pressione (o la profondità) alla quale è stata effettuata la misura.
Le nostre attuali conoscenze sulla distribuzione della temperatura, della salinità e della densità delle acque oceaniche sono state in gran parte ottenute grazie a un'accurata serie di stazioni di rilevamento (la stazione di rilevamento è il punto in cui una nave misura la distribuzione verticale della temperatura e della salinità, per mezzo di bottiglie campionatrici e di termometri a rovesciamento). Questo metodo fornisce i valori della temperatura e della salinità solamente a quelle particolari profondità alle quali sono stati presi i campioni e rovesciati i termometri. Recentemente sono stati realizzati degli strumenti in grado di registrare con continuità la temperatura e la salinità al variare della profondità, via via che lo strumento è calato nell'acqua. Generalmente questi strumenti consistono di un termometro a resistenza al platino, un misuratore di salinità (o di conducibilità) e un manometro. Tutti e tre controllano la frequenza di un oscillatore e le tre frequenze risultanti, mescolate tra di loro, sono trasmesse mediante un solo cavo conduttore alla nave, dove i segnali sono demodulati e visualizzati.
Questi strumenti, detti STD (Salinity, Temperature, Depth = salinità, temperatura, profondità) o CTD (Conductivity, Temperature, Depth = conducibilità, temperatura, profondità), hanno dato interessanti risultati nello studio delle variazioni su scala ristretta in funzione della profondità; sono comodi da usare e la loro precisione relativa può essere resa sufficientemente elevata mediante una taratura accurata. Per una precisione assoluta occorre ancora tararli sui termometri a rovesciamento e su acqua di mare campione.
Una determinazione rapida e conveniente, benché meno accurata, della distribuzione della temperatura nei primi 500 m a partire dalla superficie si può ottenere usando un batitermografo a perdere (Expendable Bathy Thermograph, XBT). L'XBT, che rappresenta un perfezionamento di un precedente strumento meccanico che poteva essere calato in mare da un argano posto su una nave in movimento, consiste di un termistore a risposta rapida montato entro un corpo affusolato che affonda liberamente lungo un cavo sottile da una nave in movimento. La temperatura è registrata in funzione del tempo, presumendo che il corpo scenda verso il fondo con velocità uniforme e nota. L'XBT è utile per rilevamenti rapidi; ne esiste una versione più costosa che può essere sganciata anche da un aereo.
Il numero delle osservazioni in superficie è maggiore di quello delle osservazioni in profondità, che sono più difficili e più costose. Cionondimeno oggi abbiamo una buona conoscenza della distribuzione media della temperatura, della salinità e del contenuto di ossigeno, mentre le conoscenze sugli altri costituenti sono molto meno complete.
Ciò che conosciamo meglio è la distribuzione della temperatura: essa varia tra −2 °C e 30 °C (più o meno l'intervallo di temperatura entro cui può vivere l'uomo), ma c'è molta più acqua fredda che calda: la temperatura media è di 3,5 °C. Tuttavia l'acqua a temperatura superiore a 5 °C è confinata in una fascia relativamente stretta tra 50° nord e 50° sud.
Tra queste latitudini la struttura tipica prevede uno strato di acqua quasi isotermica in superficie, uno strato più profondo in cui la temperatura varia con relativa rapidità e uno spesso strato freddo che arriva fino al fondo. Quanto sopra è illustrato nella fig. 7; la regione in cui la temperatura varia più rapidamente con la profondità e nota come ‛termoclino principale dell'oceano'.
Il modo in cui la distribuzione tipica della temperatura cambia col variare della latitudine è mostrato nella fig. 8. A latitudini maggiori di 50° circa la temperatura cambia poco al variare della profondità. Man mano che si va verso l'equatore aumenta la temperatura superficiale e diminuisce la profondità del termoclino principale che diviene un cambiamento di temperatura molto brusco e vicino alla superficie negli oceani tropicali ed equatoriali.
La fig. 9 mostra una tipica sezione nord-sud della distribuzione della temperatura: tutta l'acqua a temperatura superiore a 5 °C è confinata nello strato superiore, il resto dell'oceano è freddo, persino ai tropici e all'equatore.
Questa struttura, almeno fino a un certo punto, è spiegabile in base alle summenzionate proprietà dell'acqua di mare. La luce solare non penetra in profondità e tutti i processi che tendono a modificare la temperatura agiscono entro pochi metri dalla superficie. Dopo che una massa d'acqua ha lasciato la superficie, la sua temperatura può cambiare solo per mescolamento e questo processo non è molto efficace. Inoltre, come abbiamo già visto, più l'acqua è fredda più, in genere, è pesante. Ci si deve quindi attendere che l'acqua più pesante (la più fredda) generalmente affondi per riempire i bacini abissali. L'acqua più fredda si trova alla superficie nelle regioni polari, d'inverno, dopo che il suo calore è stato irradiato nella lunga notte polare.
La distribuzione della salinità nell'oceano ha un andamento più complesso di quello della temperatura, per cui non è altrettanto facile descriverla in poche parole. La salinità, infatti, agisce meno marcatamente sulla densità, cosicché non è detto che essa aumenti con l'aumentare della profondità. Però sia i processi che tendono a modificare la temperatura sia quelli che tendono a modificare la salinità (scambio termico, precipitazione ed evaporazione) avvengono in superficie: allorché l'acqua lascia la superficie la sua temperatura e la sua salinità possono cambiare solo per mescolamento. Siccome i processi oceanici in genere tendono a mescolare calore e sale nello stesso modo, un buon sistema per rappresentarli si ottiene mediante un ‛diagramina caratteristico', o diagramma T-S, nel quale la temperatura (più precisamente la temperatura potenziale) è tracciata in funzione della salinità. Qualsiasi campione di acqua di mare può essere rappresentato da un punto nel diagramma e una massa d'acqua di una certa estensione che possa essere identificata con un punto del genere è detta ‛tipo' d'acqua. Più spesso una massa estesa d'acqua può essere rappresentata da una linea del diagramma, dato che i valori della temperatura e della salinità variano con la profondità secondo una regola fissa. È facile vedere che se si mescolano due masse d'acqua, entrambe rappresentate da un punto del diagramma T-S, il punto che rappresenta la miscela risultante giacerà sulla linea che congiunge i primi due punti, a una distanza determinata dalle proporzioni delle masse che costituiscono la miscela finale.
I processi climatici alla superficie del mare tendono a formare ‛tipi' di acque; le masse d'acqua risultano dalla miscela di due o più tipi. La forma del diagramma è spesso caratteristica dell'acqua proveniente da un particolare punto dell'oceano e dà preziose informazioni sull'origine dei tipi d'acqua che la compongono. Per molti punti dell'oceano la relazione T-S è sorprendentemente simile, ovverossia i punti che rappresentano singoli valori giacciono presso una linea, con poca dispersione. Non è ancora stata avanzata una soddisfacente spiegazione dell'esatto meccanismo responsabile dell'unicità di questa relazione, tuttavia esso rende il diagramma utile nel controllo delle osservazioni: se un punto giace lontano dalla curva T-S usuale per quella regione, probabilmente l'osservazione è errata. Il diagramma T-S può servire anche per indicare la quantità d'acqua avente una particolare temperatura e una particolare salinità: basta suddividere il diagramma in righe (di stanziate da particolari intervalli di temperatura) e colonne (distanziate da particolari intervalli di salinità) e contrassegnare ogni casella della griglia così ottenuta con il volume (o la massa) dell'acqua avente quelle particolari caratteristiche.
La distribuzione verticale della temperatura - strato superficiale, termoclino principale, strato profondo - si mantiene, alle basse come alle medie latitudini, per tutta la durata dell'anno. Nelle acque polari invece non esiste un termoclino permanente. Questo semplice quadro è in parte complicato dagli effetti stagionali che sono evidenti soprattutto alle medie e alte latitudini. La temperatura degli strati superficiali dell'oceano, fino a una profondità variabile tra 25 e 200 m è generalmente la stessa che in superficie, a causa del mescolamento dovuto al vento; questo strato più alto, così rimescolato, è generalmente chiamato ‛strato di mescolamento'. D'inverno la temperatura superficiale è bassa e lo ‛strato di mescolamento' si estende fino al termoclino principale. In primavera la temperatura superficiale aumenta a causa dell'incremento della radiazione solare e lo strato di mescolamento si estende generalmente per circa 30 m dalla superficie, seguito immediatamente da una regione di rapido calo di temperatura. In autunno il mare comincia a perdere calore e una combinazione del mescolamento ad opera dei venti e della convezione gravitazionale fa affondare e infine distrugge la struttura termica della regione superficiale.
In pieno oceano le variazioni termiche diurne sono piccole, ma in condizioni di bonaccia il riscaldamento dovuto alla radiazione solare si fa sentire vicino alla superficie, producendovi uno strato tiepido. Una moderata azione del vento può rendere omogeneo uno strato di uno o due metri, originando così uno strato di mescolamento molto poco profondo, con uno strato stabile al di sotto, per il quale è perfettamente appropriato, per analogia, il termine ‛termoclino diurno'. L'aumento dei venti o il raffreddamento notturno generalmente distruggono una struttura del genere nel giro di uno o due giorni. Si tratta di un fenomeno variabile, difficile da studiare sia perché è sporadico sia perché le variazioni si estendono solo a profondità paragonabili al pescaggio di una nave.
L'importanza della stratificazione della temperatura sta nel fatto che nella maggior parte delle regioni dell'oceano la temperatura determina la densità e, se l'acqua diventa più fredda con l'aumento della profondità, diventa anche più pesante; in una simile situazione stabile la stratificazione può essere eliminata solo mediante mescolamento o qualche processo analogo, che richiede energia. I vari termoclini sono fortemente stabili, dato che la temperatura cala (e pertanto la densità aumenta) rapidamente con la profondità. Quasi ovunque mancano moti verticali che producano mescolamento e il termoclino agisce alla stregua di una barriera: è difficile che qualcosa che si trovi al di sotto del termoclino passi nell'acqua soprastante, e che qualcosa che si trovi al di sopra passi al di sotto. Le zone di pesca si trovano dove esistono processi che riescono a destabilizzare i termoclini, così che le sostanze nutritive che generalmente si trovano al di sotto di questi possano giungere negli strati superiori, illuminati dal sole.
Nei pressi della superficie il vento può fornire l'energia necessaria per mescolare l'acqua e quindi le sue varie proprietà, ma non si è ancora compreso appieno il meccanismo che provoca gli stessi effetti a profondità maggiori. La stabilità di base tende a smorzare le fluttuazioni turbolente che caratterizzano i fluidi omogenei presenti entro grandi volumi, tuttavia i coefficienti molecolari di diffusione sono così bassi (per il calore k ≃ 10-3 cm2 s-1, per il sale D ≃ 10-5 cm2 s-1) che il tempo necessario per il trasferimento di quantità apprezzabili di calore o di sale risulta molto lungo. È in questo contesto che si sono dimostrate di grande interesse le recenti osservazioni sulla microstruttura dell'oceano. Immediatamente dopo lo sviluppo dei registratori continui di salinità, temperatura e profondità, si trovò che né la temperatura né la salinità variano in modo continuo con la profondità. Talvolta cambiano in maniera piuttosto discontinua, secondo una serie di piccoli gradini. È stato provato che alcune di queste discontinuità presenti su scala assai ristretta sono dovute a ‛fontane di sale', cioè a un'instabilità che si produce, allorquando acqua relativamente calda e salata si trova al di sopra di acqua relativamente fredda e dolce, a causa della maggiore rapidità con cui avviene il trasferimento molecolare del calore rispetto a quello del sale. Altre sembrano dovute all'intersecarsi e al diffondersi di masse d'acqua di densità quasi costante, ma di diversa temperatura e salinità. Tali masse d'acqua dalle dimensioni di metri potrebbero forse essere prodotte da un processo noto come ‛instabilità di Kelvin-Helmholtz', che spiega perché il movimento con una velocità di scorrimento in un fluido stabilmente stratificato può produrre vortici che causano moti di avvolgimento e infine rendono localmente omogeneo il fluido.
5. Correnti oceaniche.
Il movimento dell'oceano è più difficile da misurare della sua temperatura o della sua salinità, soprattutto perché è necessario un sistema di riferimento fisso, il che difficilmente si può ottenere a bordo di una nave lontana dalla terraferma. Le correnti oceaniche nei pressi della superficie in alcune località sono abbastanza forti da agire sulle navi, e soprattutto sui velieri; come tali sono note ai marinai da sempre.
Ancor oggi la maggior parte delle nostre conoscenze in proposito sono basate sulla raccolta dei dati relativi alla deriva delle navi, riportati dai naviganti. Una nave che segua una rotta costante a una velocità data non viaggia su quella rotta e con quella velocità rispetto alla terraferma, ma ne è allontanata dalle correnti oceaniche. Nel caso di una navigazione tranquilla, un semplice calcolo fornisce una stima della velocità e della direzione effettive della corrente prossima alla superficie. Per molti anni le autorità marittime dei diversi paesi hanno raccolto tali dati, ne hanno elaborato le medie e pubblicato i risultati sotto forma di carte. Abbiamo così una buona conoscenza della circolazione superficiale dell'oceano, benché la maggior parte delle osservazioni riguardino le rotte mercantili e siano scarse quelle relative alle parti più remote del mondo.
Le caratteristiche principali delle correnti sono illustrate nella fig. 15; benché gli oceani Pacifico, Atlantico e Indiano abbiano forme piuttosto differenti, essi presentano una disposizione delle correnti, o circolazione generale, abbastanza simile: vi è una circolazione assai estesa in senso orario (o rotazione) nelle parti settentrionali e una in senso antiorario in quelle meridionali. Le due circolazioni non sono simmetriche, dato che le correnti sono molto più forti in una regione ristretta nei pressi dei margini occidentali del Pacifico settentrionale e dell'Atlantico settentrionale (nell'Oceano Indiano la situazione è complicata dalle variazioni stagionali dovute ai monsoni). Queste forti correnti costiere (la Corrente del Golfo nell'Atlantico e la Curo Scivo nel Pacifico) sono le correnti meglio conosciute dell'oceano.
Nei pressi dell'equatore in tutti e tre gli oceani esistono due correnti equatoriali che scorrono verso ovest. La corrente sud-equatoriale si trova in corrispondenza dell'equatore o a sud di esso, la corrente nord-equatoriale a nord. Negli oceani Pacifico e Indiano, e in parte dell'Atlantico, le due correnti equatoriali dirette verso ovest sono separate da una controcorrente equatoriale diretta verso est.
Nell'oceano meridionale, che circonda l'Antartide, non esistono barriere continentali (benché lo Stretto di Drake, di larghezza relativamente limitata, possa avere un effetto analogo) e la corrente superficiale principale scorre intorno alla terraferma sotto forma di un flusso diretto verso est, noto come corrente circumpolare o come deriva del vento dell'ovest.
Bisogna sottolineare che le carte delle correnti oceaniche sono carte climatologiche, cioè basate su medie di osservazioni eseguite nel corso di lunghi periodi di tempo. In casi particolari una nave può incontrare una corrente molto differente dalla corrente indicata sulla carta nautica. Questo fenomeno è notevole specialmente nelle regioni attraversate da una corrente costiera veloce, diretta a ovest, come la Corrente del Golfo che serpeggia e cambia la posizione del proprio asse in maniera imprevedibile.
In un caso del genere la carta climatologica può indurre in errore, dal momento che le osservazioni in un particolare punto sono mediate su lunghi periodi senza riguardo alla presenza o meno della corrente. Una corrente stretta e forte, che cambia di posizione, è rappresentata su una carta climatologica come una corrente più larga ma più debole. In questo modo quella che si può definire ‛corrente del golfo climatologica' (cioè quella rappresentata su una carta climatologica costruita in base ai valori medi nel tempo) è forse dieci volte più larga, e decisamente più debole, della Corrente del Golfo.
Le carte nautiche danno tuttavia una rappresentazione adeguata e utile delle correnti nei primi 10 m dalla superficie dell'oceano. A profondità maggiori la loro distribuzione è conosciuta molto meno bene, soprattutto a causa delle difficoltà inerenti alla loro osservazione.
Nel corso dei primi tentativi fatti per misurare le correnti oceaniche in profondità ci si serviva di correntometri muniti di un rotore che misurava la velocità della corrente e di un timone che ne confrontava la direzione rispetto a una bussola magnetica. Questi strumenti venivano calati da navi ancorate, ma i risultati indicati erano ambigui. Le correnti erano deboli e potevano essere seriamente influenzate dal moto della nave. Non è esagerato dire che fino al 1955 circa non si avevano ancora osservazioni dirette attendibili delle correnti più profonde dell'oceano. Poi J. C. Swallow, un oceanografo inglese, sviluppò un metodo lagrangiano. Se si usa un rotore e una banderuola e si misura il flusso dell'acqua in un dato punto, il metodo è detto euleriano, perché fu Eulero che propose di specificare il flusso dei liquidi in tale maniera. Un metodo lagrangiano, invece, misura il percorso di una data particella che si muove col fluido.
Precedenti tentativi di osservazioni lagrangiane delle correnti oceaniche si erano basati sull'uso di maniche a vento, simili a paracadute, sospese mediante sottili cavi a piccoli galleggianti di superficie. Si sperava che la forza trainante agente sulla manica a vento avrebbe fatto muovere l'intero sistema con la corrente al suo livello: il percorso della manica a vento veniva tracciato seguendo il galleggiante di superficie. Il metodo ottenne qualche successo, ma era difficile garantirne l'accuratezza, tanto questa dipendeva dalla corrente alle diverse profondità comprese tra la superficie e la profondità a cui operava la manica a vento. Swallow eliminò il galleggiante di superficie e i cavi e costruì un oggetto che rimaneva in sospensione a una data profondità e che andava alla deriva con l'acqua a quella profondità. Egli trovò che le proprietà meccaniche di alcune leghe di alluminio erano particolarmente adatte per la costruzione di capsule meno compressibili dell'acqua di mare. Adeguatamente zavorrata, una capsula del genere sarebbe affondata finché la pressione differenziale non avesse reso uguali la sua densità e quella dell'acqua circostante; da quel momento si sarebbe mossa con la corrente a quella profondità.
La rotta di questi galleggianti veniva seguita acustica- mente: essi contenevano un circuito elettronico, completo di batterie, che emetteva impulsi sonori di frequenza in- torno a 10 KHz a intervalli di circa un secondo. Per mezzo di due idrofoni, uno a prua e uno a poppa di una nave da ricerca, si poteva eseguire il rilevamento del galleggiante; una successione di tali rilevamenti localizzava il galleggiante rispetto alla nave. La nave, dal canto suo, veniva localizzata rispetto alla terraferma mediante rilevamenti radar di una boa ormeggiata.
Da allora si è sempre usato questo metodo. Sono stati apportati svariati miglioramenti per comodità pratica: i moderni galleggianti Swallow contengono dei ripetitori, cosicché possono essere localizzati su domanda piuttosto che seguendone l'emissione; coi moderni sistemi di navi- gazione, inoltre, si può fare il punto senza bisogno di or- meggiare una boa, e cosi via. Si sono persino potute costruire grandi capsule in sospensione dotate di una sorgente sonora abbastanza potente da essere captata dalle stazioni costiere d'ascolto, ovviando alla necessità di una costosa nave. Però tali galleggianti possono essere usati solo a certe profondità (a non molta distanza dal canale sonoro) e solo in quelle regioni dell'oceano dove esistono appropriate stazioni costiere d'ascolto.
Anche i metodi euleriani si sono sviluppati nel corso degli ultimi due decenni. Dato che gran parte delle incertezze proveniva dal tentativo di fare le osservazioni da bordo di una nave, si è fatto un considerevole sforzo per lo sviluppo e la costruzione di un correntometro-registratore, nel quale i giri di un rotore, che misura la velocità, e l'orientamento di una banderuola, che valuta la direzione, sono registrati su un nastro magnetico contenuto all'interno dello strumento stesso.
Si potevano sospendere a una boa in superficie file di questi strumenti ancorati al fondo, e quindi lasciarli registrare anche per un mese prima di recuperarli e tradurre i dati per l'analisi. Le prime esperienze non ebbero tutte il medesimo successo, ma gradualmente il metodo migliorò finché i dati acquisiti non compensarono decisamente gli sforzi. Ci si accorse presto che l'uso di un galleggiante di superficie non era conveniente, in primo luogo perché spesso le navi di passaggio operavano il ‛salvataggio' dell'apparecchiatura; in secondo luogo, e cosa più importante, perché il movimento del galleggiante di superficie, causato dalle onde, veniva trasmesso al cavo di ormeggio sottostante falsando così i risultati. Il metodo moderno consiste nel sospendere i correntometri a un oggetto più leggero dell'acqua che è mantenuto a 100 o più metri sotto la superficie per mezzo di zavorre poggiate sul fondo. Alla fine del periodo di registrazione un segnale acustico proveniente dalla nave aziona uno sgancio, liberando il cavo di ormeggio dal peso che lo ancora e permettendo alla boa sommersa di risalire alla superficie insieme con gli strumenti. Gli strumenti elettrici, elettronici e acustici necessari hanno raggiunto attualmente un considerevole grado di perfezionamento, cosicché si dispone dei mezzi adatti per misurare le correnti profonde dell'oceano, benché il loro impiego richieda un considerevole grado di abilità ed essi siano, come ci si può aspettare, costosi.
Nonostante la disponibilità di tecniche di osservazione, la nostra conoscenza delle correnti profonde dell'oceano è ancora frammentaria: poche di esse sono permanenti, nel senso che sono sempre presenti nei pressi di una particolare località e a una certa profondità, quantunque con variazioni di velocità e di direzione; l'impressione generale è di grande variabilità, con fluttuazioni molto più ampie del valore medio.
Forse le più interessanti correnti sottomarine ‛permanenti' sono le sottocorrenti equatoriali. Esse si trovano nel Pacifico e nell'Atlantico (e sporadicamente nell'Oceano Indiano) sotto forma di una corrente veloce e stretta, che scorre da ovest a est a una profondità di circa 100 m esattamente lungo l'equatore.
La sottocorrente equatoriale dell'Atlantico fu ipotizzata da Buchanan più di ottanta anni fa, ma fu quella del Pacifico ad essere riscoperta (nel 1952) e studiata accuratamente (1958) dagli oceanografi statunitensi.
Il lavoro recente è iniziato con l'osservazione, fatta da Cromwell, che lunghe reti da pesca calate presso l'equatore per la pesca di diverse specie di tonni tendevano a derivare verso est, in direzione contraria al flusso superficiale. Ciò implicava l'esistenza di una corrente sottomarina, che fu chiamata sottocorrente equatoriale. Essa fu studiata esaurientemente da J. A. Knauss (v. figg. 16 e 17): si trattava di una corrente stretta (300 km), sottile (0,2 km), veloce (fino a 150 cm/s), che si estendeva almeno da 150° ovest a 92° ovest (per 6.500 km). L'asse centrale della corrente giace lungo l'equatore a una profondità che diminuisce lentamente da 100 m, a ovest, fino a circa 40 m, nei pressi delle isole Galapagos. La corrente trasporta circa 40 milioni di tonnellate di acqua al secondo - più o meno la stessa quantità trasportata dalla corrente della Florida, che è la parte più veloce della Corrente del Golfo.
L'esistenza di una corrente analoga nell'Atlantico fu confermata intorno al 1960; le ricerche fatte nel tentativo di individuarla anche nell'Oceano Indiano indicano che talvolta è presente, in primavera, ma non tutti gli anni e non in maniera così marcata come nell'Atlantico e nel Pacifico. La differenza è dovuta alla variabilità dei monsoni sopra l'Oceano Indiano.
Sono note altre correnti sottomarine che prendono la forma di correnti di travaso. Queste correnti si riscontrano ogniqualvolta in un bacino con un'imboccatura stretta e alta si forma acqua di densità elevata: l'acqua densa tra- bocca dall'imboccatura sotto forma di corrente nel bacino oceanico esterno. Esempi tipici sono il flusso di acqua profonda e pesante dal Mediterraneo all'Atlantico in corrispondenza dello stretto di Gibilterra (v. fig. 18) e il flusso di acqua densa dall'Artico all'Atlantico attraverso varie spaccature della dorsale che unisce la Groenlandia, l'Islanda e la Scozia. La conoscenza di questi travasi è ancora inadeguata, benché essi rivestano un ruolo importante nella determinazione della struttura delle profondità oceaniche.
A parte tali flussi semipermanenti, anche la conoscenza delle correnti sottomarine è scarsa. Esistono lunghe serie di osservazioni per alcune località particolari, come il punto D a 39° nord e 70° ovest, sul lato rivolto verso la costa, della Corrente del Golfo nell'Atlantico settentrionale. In questa località scienziati dell'Istituto oceanografico di Woods Hole hanno effettuato ripetuti stazionamenti per rilevare la variazione della velocità e della direzione delle correnti. Parte dell'energia delle correnti compete ai moti a lungo periodo, ma la maggior parte alla cosiddetta frequenza inerziale data da 2π/sen ϕ, in cui ϕ rappresenta la latitudine. Inoltre esiste energia alle varie frequenze di marea. Risultati analoghi sono stati ottenuti in stazionamenti ripetuti presso la piattaforma continentale a ovest del Portogallo e in serie più brevi di osservazioni in mare aperto.
Particolare interesse rivestono le correnti profonde dell'oceano nelle regioni a est delle correnti costiere occidentali. È stata effettuata una lunga serie di osservazioni in una regione di acque profonde a sud-est delle Bermude (41° nord 14° ovest), nel 1959-1960, ad opera di una nave di modeste dimensioni, la Aries. Le misure avrebbero dovuto confermare l'ipotesi teorica che a tutte le profondità l'acqua si muove lentamente verso nord, a velocità non superiore a circa 1 mm/s (100 m al giorno).
Si trovò invece che le correnti erano più forti (circa 10 cm/s) e che non seguivano direzioni preferenziali. Si trovò anche che durante un periodo di osservazione di una settimana o due le correnti a tutte le profondità andavano più o meno nella stessa direzione e alla stessa velocità, ma che la loro direzione generale, e in qualche misura anche la loro velocità, erano differenti se le osservazioni venivano ripetute a distanza di un mese o due. Se ne ricavò la chiara impressione che nell'oceano ci fossero vasti vortici, con periodi dell'ordine di 100 giorni e con un'estensione spaziale compresa tra 100 e 200 km.
Non sembrava che tali vortici avessero alcuna relazione con i vortici e gli anelli di maggiori dimensioni, associati ai meandri e alle biforcazioni della Corrente del Golfo: il loro studio richiedeva metodi che permettessero misure continue per alcuni mesi, effettuate su un'area di qualche centinaio di chilometri di diametro. Durante il decennio successivo furono messi a punto congegni ormeggiati e metodi lagrangiani più adatti, cosicché gradualmente divenne possibile dare il via a una serie di programmi di misurazione su larga scala.
Contemporaneamente anche in URSS era aumentato l'interesse per le correnti sottomarine variabili. Una serie di correnti oceaniche della durata di un mese venne registrata nel Mar Caspio nel 1935. Questo esperimento fu seguito da osservazioni eseguite nell'Atlantico settentrionale (1958), nell'Oceano Indiano nordoccidentale (1967) e nella zona tropicale dell'Atlantico settentrionale (1970), mediante file di correntometri fissi, dislocati in aree sempre più vaste per tempi di osservazione sempre più lunghi. In tutte queste serie di osservazioni la maggior parte dei correntometri si trovava entro i primi 1.200 m dalla superficie dell'oceano, sebbene alcuni fossero a profondità superiori. In tutte le serie furono registrate fluttuazioni della corrente su una grande varietà di scale. Nel 1970 una serie di 17 ormeggi che copriva una superficie di 200 km di diametro fu mantenuta in funzione per sei mesi, durante i quali un vortice ellittico con moto orario (con assi di 200 km e 90 km) si spostò attraverso lo schieramento in direzione ovest.
Nel 1971 scienziati di 15 enti di ricerca degli Stati Uniti e del Regno Unito decisero che la comprensione del problema e lo sviluppo della tecnologia erano sufficientemente avanzati da giustificare l'iniziativa di un esperimento sulla dinamica del mare aperto (Mid-Ocean Dynamics Experiment, MODE I). Il lavoro sperimentale fu nuovamente compiuto in una regione a sud-ovest delle Bermude: era incentrato su una serie di osservazioni intensive protratte per un periodo di 5 mesi, cui parteciparono sei navi, e sulla misurazione simultanea di molte particolarità della distribuzione delle correnti, della temperatura e della salinità mediante l'uso di numerosi e differenti tipi di strumenti. I metodi principali comprendevano la registrazione delle correnti a diverse profondità, mediante boe ancorate, galleggianti a profondità fissa e derive ad affondamento lento, e la misurazione della temperatura e della salinità per mezzo di strumenti CTD, STD e XBT. Di particolare interesse erano i grandi galleggianti a profondità fissa seguiti dalle stazioni costiere. Erano zavorrati in modo da porsi in galleggiamento intorno a 1.500 m di profondità (la profondità del cosiddetto canale sonoro, entro cui il suono percorre grandi distanze nell'oceano). I risultati del MODE I accrebbero di molto la nostra conoscenza sui fenomeni variabili dell'oceano. Fu confermato che esiste un campo di vortici dotati di energia sufficiente da influire significativamente sulla circolazione dell'oceano. Queste conoscenze hanno in certo modo concluso una rivoluzione nelle nostre concezioni della circolazione oceanica: l'idea che i vortici siano un'importante componente della dinamica dell'oceano è ora accettata in tutti gli studi di chimica e biologia oceanica e in quei settori dell'oceanografia che trattano fenomeni in cui si pensa che giochino una parte importante i processi del trasporto orizzontale.
Una volta accettato il concetto generale dell'esistenza di vortici nell'oceano, furono presto trovate prove della loro ubiquità; sembra che esistano movimenti su scala simile in molte altre parti dell'oceano. Sono stati trovati vortici nelle vicinanze della maggior parte delle correnti oceaniche principali; per lo più hanno un diametro compreso tra 10 e 1.000 km e generalmente sono più forti nei pressi della superficie. Alcuni sono stati rilevati mediante osservazioni oceanografiche dirette, altri a distanza, dai satelliti, che hanno registrato anomalie termiche superficiali e particolari conformazioni di nubi. Alcune delle prove più convincenti dell'esistenza dei vortici d'alto mare sono venute da osservazioni fatte per scopi diversi e non analizzate pensando alla possibilità di una struttura a vortici.
Sembrano esserci validi motivi per credere che i vortici sottomarini più forti siano associati alla distribuzione della densità che accompagna le forti correnti costiere occidentali, come la Corrente del Golfo, dove è probabile che si formino.
Per la prossima fase di ricerca, che avrà l'obiettivo di capire come tali vortici oceanici agiscano sulla circolazione generale dell'oceano, saranno necessari anni di osservazioni in molte parti del mondo e il contributo degli oceanografi di molti paesi. Già adesso gli Stati Uniti, l'URSS, il Regno Unito, il Canada, la Francia e la Germania Occidentale stanno collaborando a un progetto di primaria importanza, il POLYMODE, una combinazione, come suggerisce il nome, del sovietico POLYGON e dell'occidentale MODE I.
Resta ancora molto da fare, non solo a proposito dei vortici oceanici e della loro dinamica, ma anche per quel che riguarda le configurazioni di correnti su scale diverse, da quella dei bacini oceanici a quella della più piccola increspatura, prima di poter dire che si sia compreso quale parte l'oceano gioca nel bilanciamento termico globale e nella regolazione del clima terrestre.
6. Studi teorici.
Abbiamo sin qui delineato le moderne tecniche di osservazione delle più importanti caratteristiche dell'oceano che ne costituiscono l'‛anatomia': la distribuzione della temperatura, della salinità e delle correnti. Sono questi i dati che devono essere spiegati dalla teoria.
Lo studio della distribuzione nel tempo e nello spazio delle tre componenti della velocità della corrente, della pressione, della densità, della temperatura e della salinità dell'oceano richiede la definizione di un certo numero di funzioni, precisamente di u, v, w, p, ρ, T, S, nella forma di f(x, y, z, t)
Le equazioni di cui disponiamo sono sette: tre equazioni del moto, quelle di Navier e Stokes ricavate dalle leggi di Newton; un'equazione empirica, ricavata in laboratorio, che lega tra di loro la pressione, la densità, la temperatura e la salinità; tre equazioni di continuità, che esprimono l'equilibrio di massa, calore e salinità.
Alcune di queste equazioni non sono lineari e le loro condizioni al contorno sono complicate: in esse interviene non solo la complessa geometria dei bacini oceanici, ma anche l'assorbimento dei raggi solari, l'evaporazione e lo scambio di calore alla superficie del mare, nonché l'effetto del vento. L'oceano, come l'atmosfera, è in moto turbolento su scale molto diverse; la distribuzione delle variabili non è a lenta variazione e queste quindi non possono essere completamente rappresentate analiticamente o mediante un elaboratore elettronico: esistono sempre moti su scala minore che non possono essere risolti e rappresentati esplicitamente, per cui è chiaro che si può progredire solamente facendo grossolane semplificazioni, scegliendo nell'oceano particolari fenomeni nei quali è probabile che siano dominanti certi processi specifici ed eseguendo un continuo controllo sperimentale delle previsioni teoriche.
La profondità dell'oceano è piccola rispetto alla sua estensione e la componente verticale della velocità è generalmente piccola rispetto a quelle orizzontali. L'equazione del moto lungo la verticale può essere ridotta con buona approssimazione quasi dappertutto (tranne nelle immediate vicinanze della superficie oceanica) all'equazione dell'equilibrio idrostatico:

Per convenzione prendiamo l'asse x in direzione est, l'asse y in direzione nord e l'asse z verticale. Indichiamo le corrispondenti componenti della velocità con u, v, w; p rappresenta la pressione, ρ la densità, g l'accelerazione di gravità.
Le equazioni del moto orizzontale contengono termini di tipo diverso, quali le variazioni nel tempo (come ∂u/∂t), termini inerziali (come u∂u/∂x), termini che implicano il gradiente di pressione (come ρ-1 ∂p/∂x), termini che implicano le forze di Coriolis (come fν, con f = 2ω sen ϕ, in cui ω è la velocità angolare della Terra e ϕ la latitudine) e termini che implicano l'attrito (come il coefficiente ν ∂2u/∂z2). Se trascuriamo tutti questi termini tranne il gradiente di pressione e quello legato alle forze di Coriolis, si ottiene:
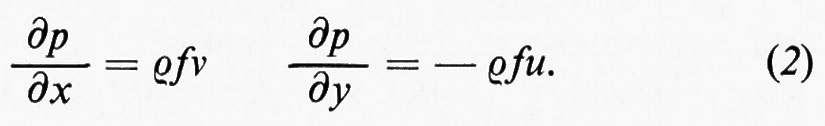
Queste sono le cosiddette equazioni geostrofiche, note ai meteorologi fin dal XIX secolo. Nell'atmosfera possono essere utilizzate per fare una stima della velocità e della direzione del vento: i venti soffiano lungo le isobare a velocità inversamente proporzionale alle distanze relative tra queste e al seno della latitudine. Le equazioni geostrofiche esprimono anche l'equilibrio primario delle forze nell'oceano: le correnti circolano intorno ai vortici oceanici nello stesso modo in cui i venti circolano intorno alle depressioni atmosferiche.
Sfortunatamente non esiste ancora il modo di misurare il gradiente della pressione in un punto dell'oceano con precisione sufficiente per usare direttamente l'equazione geostrofica, per cui gli oceanografi si servono contemporaneamente delle equazioni geostrofiche e dell'equazione idrostatica per calcolare lo shear, ovvero le variazioni secondo la verticale, delle correnti geostrofisiche sulla base di osservazioni della distribuzione della densità. Questo è il motivo per cui occorre una grande precisione nella determinazione della temperatura e della salinità: esse servono infatti per calcolare il campo di densità che determina la distribuzione secondo la profondità della corrente geostrofica.
Le correnti delle profondità oceaniche sono quasi in equilibrio geostrofico ma, siccome l'oceano è termodinamicamente meno attivo dell'atmosfera, il suo moto è in gran parte guidato dai venti. Se, per esempio, consideriamo una regione nella quale l'equilibrio principale è dovuto alle forze di Coriolis e di attrito, le equazioni si semplificano in
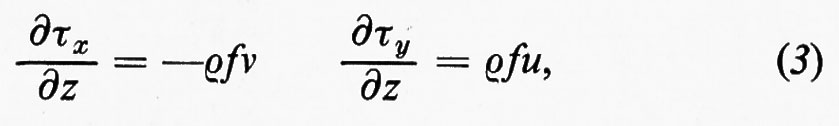
in cui τx e τy sono le componenti verso est e verso nord dello sforzo di taglio orizzontale. Gli effetti principali del trascinamento ventoso sono ristretti a uno strato relativamente sottile vicino alla superficie per cui è conveniente integrare la (3) dalla superficie a una profondità d, oltre la quale le velocità risultanti sono nulle.
Allora:
fù = τy (0) fã = −tx(0), (4)
in cui ù = ∫²0 ρudz, ã = ∫d0 ρνdz e τx (0) e ty(0) sono le componenti in direzione est e nord della forza di trascina- mento ventoso del mare. L'equazione (4) mostra che lo spostamento di masse dovuto all'attrito, cioè la deriva di Ekman, è perpendicolare allo sforzo di taglio in superficie, e diretto verso destra nell'emisfero settentrionale. Questo risultato è dovuto a V. W. Ekman, che studiò il problema dietro suggerimento di Fr. Nansen, il quale aveva notato che gli icebergs non andavano alla deriva esattamente nella direzione del vento. Più importante è l'applicazione al fenomeno noto come upwelling: se il vento soffia parallelamente alla costa, con la risultante deriva di Ekman diretta verso il largo, allora l'acqua che si trova immediatamente al di sotto della superficie risale per prendere il posto dell'acqua superficiale trascinata dalla deriva. Quest'acqua che risale è più fredda e più ricca di sostanze nutritive e porta quindi un aumento della produzione biologica. Alcuni dei punti di pesca più importanti del mondo dipendono dall'upwelling, in particolare quelli al largo del Perù, dove le variazioni del fenomeno hanno effetti economici importantissimi.
Fu solo nel 1947 che H. U. Sverdrup collegò l'equilibrio geostrofico con quello di Ekman e trovò, eliminando la pressione mediante differenziazione incrociata, che
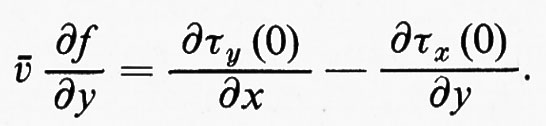
Questa è un'equazione di vorticosità, più spesso scritta sotto la forma βῡ = rot γ (0), che mostra come il trasporto di masse verso sud e il rotore dell'azione del vento siano legati dalla variazione della rotazione terrestre con la latitudine.
Alla maggior parte delle medie latitudini il rotore dell'azione del vento è negativo, dato che la componente occidentale del vento aumenta andando dagli alisei ai venti occidentali. Così in genere le correnti oceaniche trasportano acqua da nord verso sud. Una difficoltà si incontra nel soddisfare l'equazione di continuità per la massa nel caso di una sottile corrente costiera occidentale, per la quale la semplice approssimazione di Sverdrup non è valida. Stommel indicò che la soluzione di Sverdrup aggiunta a una corrente costiera occidentale di ordine maggiore può spiegare l'asimmetria delle correnti superficiali degli oceani settentrionali, nei quali il trasporto verso nord è concentrato in una corrente veloce e stretta localizzata presso la costa occidentale.
Si sono fatti diversi tentativi per ampliare questa semplice trattazione della circolazione dovuta ai venti, ma i risultati dipendono molto: dalle approssimazioni fatte nelle equazioni complete del moto per tener conto di strutture su scala minore, come i vortici oceanici.
Questo assume un'importanza ancora maggiore allorché si considerano le correnti oceaniche che sono causate da differenze di densità invece che dai venti. È noto che in tutti i bacini oceanici profondi l'acqua abissale è fredda e di origine polare. Le sorgenti di acqua fredda e pesante, che si incanala sul fondo, sono aree relativamente piccole dei mari di Irminger e di Weddell. Nel resto dell'oceano esiste probabilmente un lento flusso di compensazione verso l'alto alle medie profondità, cui corrisponde in generale una componente verso il polo del flusso orizzontale e una corrente costiera occidentale diretta a sud. È stata osservata una corrente più profonda della Corrente del Golfo, diretta verso sud, ma uno studio completo richiederà una migliore comprensione del flusso termico verticale (microstruttura) e del flusso termico orizzontale (vortici oceanici) dell'oceano.
7. Interazione aria-mare.
È chiaro che la distribuzione della temperatura e della salinità nell'oceano e i complessi sistemi di correnti sono in gran parte determinati dagli scambi energetici alla superficie del mare. È perciò che su questi scambi si è concentrata un'attenzione sempre crescente durante gli ultimi decenni.
La più importante fonte di energia è costituita dalla radiazione solare, che fornisce energia a tutti i tipi di moti, sia nell'oceano sia nell'atmosfera, fatta eccezione per le maree. L'energia della radiazione solare si trova concentrata intorno a una lunghezza d'onda di circa 0,5 μm, corrispondente alla temperatura di un corpo nero a circa 6.000 °K. Soltanto una piccola frazione dell'energia irradiata dal Sole raggiunge l'atmosfera terrestre: essa ammonta a circa 2 cal/cm2 al minuto.
La quantità di energia relativa alle radiazioni a breve lunghezza d'onda che penetra nel mare dipende da quattro fattori: la durata del giorno, l'assorbimento dell'atmosfera (che dipende notevolmente dall'altezza del Sole sull'orizzonte), l'assorbimento e la riflessione dovuti alle nubi, la riflessione da parte della superficie del mare. La ripartizione di questa energia può essere convenientemente espressa mediante l'equazione del bilancio termico per una colonna di oceano al di sotto di una superficie di area unitaria:
Qs = Qb + Qh + Qe + Qv + QT,
in cui ogni termine rappresenta un flusso di calore: Qs è il flusso di energia solare che attraversa nell'unità di tempo l'unità di area della superficie del mare; Qb è il flusso di calore netto perduto dall'oceano sotto forma di radiazioni di grande lunghezza d'onda nell'atmosfera e nello spazio; Qh è il flusso di calore trasferito nell'atmosfera per conduzione; Qe è il flusso di calore dissipato nei processi di evaporazione; Qv è la quantità di calore rimosso dalla colonna nell'unità di tempo ad opera delle correnti oceaniche per unità di tempo e QT è il calore speso nei processi di riscaldamento della colonna nell'unità di tempo. Altre fonti e altre perdite di calore, come per esempio il calore proveniente dall'interno della Terra, possono essere trascurate.
Tutti questi flussi termici possono essere misurati con speciali strumenti, tranne Qv, che richiede la misura del flusso termico dovuto alle correnti oceaniche a tutte le profondità. Qv deve essere trovata per differenza, quindi le stime che se ne possono fare hanno una precisione relativamente scarsa.
Le misure di tutte le componenti del bilancio termico dell'oceano sono rare: ciò che serve non sono tanto i valori in un punto e a un istante particolari, ma carte che mostrino i flussi in relazione allo sviluppo sinottico o climatico. Per questo fine è opportuno che tali misure siano collegate ad altre osservazioni, come quelle meteorologiche di routine effettuate da navi di superficie.
I termini di radiazione Qs e Qb sono piuttosto difficili da valutare, per la variabilità sia delle grandezze stesse, sia delle caratteristiche radiative delle nuvole; tuttavia dai valori medi sono state ricavate formule empiriche abbastanza buone.
Per valutare Qh di solito si correla il flusso termico con la differenza di temperatura tra il mare e l'aria:
Qh = Cρa(Tm − Ta) U,
dove C è una costante dell'ordine di 10-3, ρa è la densità dell'aria, Tm è la temperatura alla superficie del mare, Ta e U sono la temperatura dell'aria e la velocità del vento a un'altezza di circa 10 metri sopra la superficie del mare.
Un'espressione corrispondente per il flusso termico relativo all'evaporazione è data da
Qe = Cρa (qm − qa) U L,
in cui L è il calore latente di evaporazione, qm è l'umidità specifica alla superficie del mare e qa quella a un'altezza di circa 10 metri al di sopra. L'aria sulla superficie del mare può essere considerata virtualmente (98%) satura alla temperatura della superficie stessa.
La derivazione di queste formule è complicata, dato che dipende dalle condizioni di turbolenza del flusso orizzontale. Se il moto dell'atmosfera fosse laminare, il flusso sarebbe proporzionale a k∂T/∂z con k coefficiente molecolare di conduzione termica. In un flusso turbolento, invece, il coefficiente non è costante, ma aumenta con l'altezza secondo una funzione grosso modo lineare, poiché le effettive dimensioni dei vortici aumentano. Se le condizioni sono costanti, il prodotto k ∂T/∂z è costante, quindi ∂T/∂z è inversamente proporzionale all'altezza e quindi la differenza di temperatura tra aria e mare diminuisce col logaritmo dell'altezza. I profili verticali della temperatura, della umidità e del vento hanno andamenti più o meno logaritmici e quindi la maggior parte delle differenze tra aria e mare risultano concentrate nei primissimi metri di altezza.
I grafici di Qh e di Qe ricavati dalla media climatologica delle osservazioni effettuate da navi mostrano che gran parte dell'energia è trasferita dal mare all'aria nelle aree a est delle masse continentali, alle medie latitudini. Queste sono aree nelle quali aria fredda (per esempio da Terranova) scorre su un tratto di oceano caldo (la parte settentrionale della Corrente del Golfo). Risulta anche che in queste aree hanno origine molte depressioni atmosferiche di media latitudine e che quindi esiste una stretta connessione tra l'interazione aria-mare e i fenomeni meteorologici.
Se applichiamo l'equazione del bilancio termico agli oceani come a un tutto unico ed eseguiamo la media su alcuni anni, allora QT e Qv si annullano e i termini rimanenti si riducono a
Qs = Qb + Qh + Qe,
per i quali i valori medi sono: 320 = 130 + 20 + 170 calorie/cm2 giorno, il che mostra come il surplus della radiazione vada in gran parte speso nell'evaporazione.
L'accumulo di calore nell'oceano, rappresentato da QT, è ben noto: abbiamo parlato in precedenza del termoclino stagionale. È il calore accumulato nell'oceano durante l'estate che fornisce energia a gran parte delle perturbazioni invernali. Il valore di Qv misura l'effetto principale dell'oceano sul clima e sulle variazioni climatiche, una materia di interesse crescente, dato che si è capito che ogni cambiamento climatico sarà probabilmente in peggio. Dal punto di vista di un climatologo, l'atmosfera e l'oceano sono componenti di un esteso sistema concatenato. I moti nell'aria e nel mare insorgono perché la maggior parte della radiazione solare arriva alle basse latitudini e mantiene una differenza di temperatura tra i tropici e i poli.
L'atmosfera tropicale riceve calore, oltre che dal Sole, anche dall'evaporazione e dal trasporto di calore e il bilancio resta attivo anche dopo aver tenuto conto della restituzione delle radiazioni a grande lunghezza d'onda (v. fig. 24). L'atmosfera polare, al contrario, è in deficit: essa, infatti, mentre emette circa la stessa quantità di radiazioni delle regioni tropicali, ne assorbe di meno. Se l'atmosfera e l'oceano fossero immobili, i tropici diverrebbero sempre più caldi e le regioni polari sempre più fredde; essi, invece, si mantengono alle temperature attuali solo perché c'è un passaggio di calore dalle basse alle alte latitudini, dovuto ai venti e, in minor misura, alle correnti oceaniche.
In tal senso l'atmosfera può esser vista come un gigantesco motore termico molto poco efficiente, che assorbe calore nella calda fascia equatoriale e lo dissipa presso i poli. Siccome gran parte del passaggio di calore è dovuta all'evaporazione e alla condensazione di vapor acqueo, si può pensare all'intero meccanismo come a un motore a vapore e usare termini espressivi come ‛caldaia equatoriale' considerando le gigantesche torri dei cumulonembi alla stregua di cilindri nei quali il vapore è trasformato in gocce d'acqua. Il motore ha un rendimento molto basso, ma indubbiamente è molto grande; il calore che viene trasferito dall'equatore al polo attraversa un determinato parallelo alla media di circa 1015 W, un valore assai alto, che rappresenta quasi un milione di volte la potenza consumata da tutta la popolazione del mondo. Come fa l'atmosfera ad operare questo enorme scambio di calore dai tropici alle regioni polari? Esistono due meccanismi principali, che operano uno tra l'equatore e le regioni poste a circa 30° nord o sud e l'altro a latitudini più elevate.
Nel meccanismo che opera alle latitudini inferiori, la caratteristica fondamentale è che l'aria si innalza all'equatore termico e poi si muove verso i poli. La maggior parte poi di questa massa d'aria si abbassa di nuovo a 30° nord o sud e ritorna verso l'equatore, acquistando una componente verso est dovuta alla rotazione terrestre e formando gli alisei. Questi trasportano meno energia dei venti d'alta quota diretti verso i poli così che si ha un trasferimento netto di energia dalle basse alle alte latitudini.
I fattori essenziali di questa circolazione sono due linee chiuse di venti a nord e a sud dell'equatore; esse furono descritte da O. Hadley fin dal 1735 e sono note come celle di Hadley. La loro esistenza dà una spiegazione razionale delle cinture di pioggia equatoriali (dovute al moto ascendente dell'aria) e degli anticicloni subtropicali intorno ai 30° nord o sud. Questi sono connessi coi moti discendenti dell'aria, con scarsità di pioggia, e quindi con aree desertiche sulla terraferma e acqua blu scuro a elevata salinità nel mare. I venti orizzontali corrispondenti a questi moti sono gli alisei, alla superficie del mare, e la corrente a getto subtropicale ad alta quota. La corrente a getto subtropicale è una fascia di forti venti occidentali alle latitudini comprese tra 30 e 40°, a un'altezza di circa 12 km. Essa insorge perché l'aria che si è innalzata all'equatore tende a mantenere la sua velocità angolare mentre si muove verso i poli. Quando raggiunge circa 30° di latitudine si muove verso est molto più velocemente della terra sottostante (v. atmosfera).
Le celle di Hadley offrono una buona spiegazione di molti fenomeni osservati e il meccanismo della circolazione atmosferica generale alle basse latitudini è relativamente ben compreso. In entrambi gli emisferi oltre i 30° nord o sud, lo scambio di calore non avviene per effetto di moti analoghi alle celle di Hadley, ma di perturbazioni su scala minore. Qui i venti predominanti sono occidentali a tutti i livelli, al contrario di quanto avviene nelle regioni tropicali, dove gli alisei in superficie hanno una componente orientale.
Incuneate tra i venti occidentali delle medie latitudini si trovano le depressioni in movimento che causano le instabilità del tempo. In queste depressioni l'aria più calda della media tende ad andare verso i poli e quella più fredda verso l'equatore. La covarianza tra il moto verso i poli e le anomalie della temperatura e dell'umidità determina il segno e l'entità dello scambio di calore verso i poli.
Attualmente nell'emisfero settentrionale c'è un numero sufficiente di stazioni meteorologiche di osservazione da permettere valutazioni quasi dirette dello scambio termico atmosferico mediante la covarianza tra il vento diretto verso i poli e le anomalie di temperatura e di umidità.
Ciò ha condotto ad alcuni nuovi e interessanti risultati relativi allo scambio termico verso i poli dovuto all'oceano. L'uso dell'equazione del bilancio termico dell'oceano lascia da calcolare Qv come differenza e la precisione non è sufficiente per effettuare stime affidabili dello scambio termico medio attraverso i paralleli. Tuttavia l'avvento della tecnologia dei satelliti ha fornito buone stime del surplus (e del deficit) di radiazioni del sistema combinato atmosfera-oceano.
In questo modo ora è possibile calcolare il contributo dell'oceano, dato dal surplus totale di radiazioni meno la quantità trasferita dall'atmosfera. Si ricava che l'oceano contribuisce largamente, in ragione di circa il 40% del totale tra 0° e 70° nord. Ad alcune latitudini, come per esempio 20° nord, pare che l'oceano trasferisca il 75% del flusso termico verso i poli: l'impressione generale è che lo scambio termico alle basse latitudini sia dovuto principalmente all'oceano e sia invece dovuto all'atmosfera alle alte latitudini.
Sembra così sufficientemente chiaro che l'oceano ha realmente un ruolo importantissimo nella determinazione del clima della Terra. Per alcuni anni a venire gli oceanografi cercheranno di stabilire attraverso quali processi avvenga lo scambio termico, se mediante correnti su larga scala o vortici nelle profondità oceaniche o per l'affondamento invernale dell'acqua fredda o tramite fenomeni superficiali causati dal vento. Una volta che si sia giunti alla comprensione dei processi, sarà possibile simulare con l'ausilio di un elaboratore i possibili sviluppi climatici.
8. Onde.
Le onde sono forse la caratteristica più tipica dell'oceano, e sono state studiate a lungo; ancora oggi tuttavia la loro natura presenta aspetti inspiegati e non sappiamo quale sia il loro effetto sulla meccanica globale dell'atmosfera e dell'oceano. Esse ricevono energia dal vento e la dissipano nell'acqua o contro le coste, ma il quantitativo in gioco è piccolo rispetto alla quantità di energia proveniente dal Sole. Forse il loro principale effetto consiste nella determinazione dell'aerodinamica della superficie marina e quindi dell'attrito fra aria e mare. Non sussistono dubbi sull'importanza di questo fatto - l'attrito superficiale è importantissimo nella generazione delle correnti oceaniche - ma lavori recenti tendono a confermare l'ipotesi che l'azione di trascinamento del vento sul mare non dipenda tanto dalle onde più grandi quanto dalle increspature di piccole dimensioni. Forse qualche moto circolatorio causato dalle onde nello strato superficiale misto dell'oceano è un meccanismo di trasferimento più importante?
Che siano significative da un punto di vista geofisico o no, le onde hanno certamente un'importanza pratica, almeno per quel che riguarda la progettazione dei porti e per la difesa dal mare. Inoltre è sempre crescente l'interesse per la possibilità di utilizzare l'energia del moto ondoso e tale ipotesi è sempre più attraente man mano che le riserve di petrolio diminuiscono e i pericoli della radioattività vengono meglio riconosciuti.
I matematici del XIX secolo studiarono le onde nei liquidi; i migliori risultati cui pervennero sono forse quelli che riguardano le onde di piccola ampiezza su acque profonde, per le quali si trovò che la superficie dell'acqua si disponeva secondo una sinusoide e che le onde erano dispersive, con
ω2 = g k,
in cui ω è la frequenza e k è il numero d'onda.
La regolarità sinusoidale delle onde trovata teoricamente era in contrasto con l'aspetto generalmente disordinato della superficie marina. Si sapeva che le dimensioni delle onde sono controllate dalla forza del vento, dal tempo in cui soffia e dall'apertura, cioè dal fronte d'acqua su cui agisce. Si distinse anche tra onde vive, generate da un vento locale, e onde morte, termine usato per indicare un'onda che ha lasciato la zona di generazione e non subisce più l'influenza del vento.
C'erano tuttavia pochi dati registrati sulle onde marine, almeno fino agli anni quaranta, allorché la prospettiva di una guerra nel Pacifico, combattuta da isola a isola, indusse i militari a studiare la possibilità di prevedere il moto ondoso. Si svilupparono rapidamente numerosi metodi di registrazione e divenne necessario metter ordine nelle risultanti serie di tempi apparentemente distribuiti a caso. Presto si scoprì che una buona rappresentazione era fornita da uno spettro di frequenza, ovvero da un grafico in funzione della frequenza dell'energia delle onde in un dato intervallo di frequenza. Un grafico di questo genere è ancor oggi uno strumento di primaria importanza nell'analisi delle onde.
Uno dei primi risultati fu che si potevano separare le onde morte in una complessa mescolanza di onde vive e morte. Siccome le onde lunghe viaggiano più velocemente di quelle corte, sono le onde più lunghe (di frequenza più bassa) che raggiungono per prime una stazione di registrazione da una perturbazione distante. La rapidità con la quale cambia la frequenza registrata delle onde permette di fare una valutazione della distanza della perturbazione: i primi ricercatori rimasero piacevolmente sorpresi nel constatare che avevano registrato, sulle coste sudoccidentali dell'Inghilterra, le onde provenienti da una tempesta al largo del Capo Horn, a migliaia di chilometri di distanza. Si capì presto che la rappresentazione spettrale era particolarmente interessante perché le onde di diversa frequenza si muovono quasi indipendentemente, senza interferenze reciproche: rappresentarle come somma di sinusoidi era fisicamente appropriato oltreché matematicamente conveniente.
Non abbiamo ancora a disposizione una teoria generalmente accettata per quanto riguarda la generazione delle onde a opera del vento, ma osservazioni abilmente eseguite hanno fatto molto per chiarire il problema. Si è trovato, per esempio, che per un vento costante che soffi dalla costa l'energia totale delle onde cresce col quadrato della velocità del vento e con l'area della superficie sulla quale questo agisce (E ∝ U2S/g).
In tali condizioni è stata osservata anche la forma dello spettro, che alle alte frequenze risulta indipendente dal vento e dalla superficie (E(ω) ∝ g2ω-5), mentre tutta la variabilità è associata alle onde più lunghe, di bassa frequenza. La forma specifica dello spettro è dovuta a numerosi fattori, quali l'avvezione dell'energia del moto ondoso, l'azione atmosferica e la piccola ma cionondimeno significativa interazione fra le onde stesse.
In base a numerose osservazioni fatte risulta chiaro che l'energia connessa con le onde dell'oceano può contribuire in larga misura al fabbisogno energetico dell'uomo. È anche abbastanza chiaro d'altra parte che ci sono molti pro- blemi pratici da risolvere. Un risultato, per esempio, riguarda la forza d'ormeggio agente su un generatore di energia ricavata dalle onde. Tale forza sarebbe notevole in caso di onde alte; infatti l'energia ricavabile dalle onde cresce all'incirca come la quinta potenza della velocità del vento e la forza d'ormeggio circa come la quarta. Così nelle tempeste entrambe sono decisamente elevate. Attualmente stanno progredendo ampi programmi di ricerca, perché l'idea di ricavare energia ‛pulita' dalle onde è innegabilmente attraente.
Le onde di superficie di cui ci siamo finora occupati sono onde corte, nel senso che la loro lunghezza non è maggiore della profondità dell'acqua. Con un'espressione più pittoresca, non sentono il fondo.
Le onde lunghe (di lunghezza d'onda considerevolmente maggiore della profondità dell'acqua) si comportano in maniera diversa: la loro velocità non dipende più dalla lunghezza d'onda o dal periodo, ma solo dalla profondità dell'acqua (esse viaggiano a una velocità data dalla radice quadrata del prodotto della profondità dell'acqua per l'accelerazione di gravità). Un buon esempio di tali onde è dato dagli tsunami, che sono onde lunghe generate dai terremoti e che viaggiano a grandi velocità (generalmente intorno a 200 m/s) attraverso l'oceano. Un esempio terribile fu lo tsunami del 1° di aprile del 1946, generato da un terremoto nei pressi delle Aleutine. Esso raggiunse le Hawaii (a 3.700 km di distanza) dopo aver viaggiato per 4 ore e 40 minuti, causando gravi danni e 150 vittime. La sua lunghezza d'onda era di circa 200 km e la sua altezza in mare aperto fu valutata in poco più di 50 cm, ma aumentò fino a più di 10 metri quando raggiunse le acque basse. Attualmente funziona un servizio di allarme per gli tsunami, che fornisce il maggior numero possibile di informazioni alle isole che potrebbero esserne colpite.
9. Maree.
Le maree sono onde oceaniche lunghe, che differiscono da tutti i moti sin qui menzionati in quanto la loro energia proviene dall'energia rotatoria dei sistemi Terra-Luna e Terra-Sole, invece che dall'energia termica del Sole.
Le basi della scienza delle maree risalgono a Newton; Laplace la considerò una branca dell'oceanografia dinamica. Le ricerche sui fondamenti della dinamica delle maree sono tuttora in atto. La maggior parte della gente ha una certa conoscenza dei meccanismi più elementari delle forze di marea e pensa alle maree come a un fenomeno conosciuto abbastanza a fondo, che può essere facilmente previsto con una notevole precisione. In effetti le maree sono tra le pochissime variabili geofisiche prevedibili con notevole precisione senza che si sappia alcunché del processo fisico implicato. Tutto ciò che occorre è un grafico di registrazione delle maree in una data località per alcuni cicli lunari, forse per un anno, dal quale risultino le ampiezze e le fasi di un certo numero di termini armonici a frequenze astronomiche note. La serie può quindi essere estrapolata con adattamenti di scarsa entità per decenni. La previsione delle maree rimane un procedimento essenzialmente empirico, ma secondo Cartwright (v., 1977) le maree come sistema meccanico non saranno comprese finché non si potrà definire spazialmente il grande complesso delle onde di marea che oscillano sugli oceani, ove sono generate. Le maree vanno definite in funzione dello spazio, piuttosto che del tempo, ma finora non è ancora stata fatta una ‛mappa cotidale' soddisfacente dell'oceano. A tal fine occorre combinare difficili misure effettuate in mare aperto con la soluzione di una serie di equazioni differenziali parziali, con complesse condizioni al contorno, che soddisfino la meccanica delle onde. Il problema comincia solo ora a sembrare risolvibile, con lo sviluppo dei metodi oceanografici e di simulazione computerizzata. Ora che si dispone di elaboratori elettronici sembra possibile trovare soluzioni realistiche per le equazioni di marea di Laplace per tutti gli oceani e i mari che abbiano una geometria non troppo complicata.
Gli scienziati hanno ottenuto risultati differenti, per lo più dovuti alle diverse maniere di includere l'attrito, ma anche al fatto di aver trascurato altri fattori fisici di una certa importanza, in particolare l'elasticità della crosta terrestre sotto il carico delle maree.
I problemi energetici delle maree terrestri sono anch'essi non del tutto risolti. Si pensa che la maggior parte della dissipazione delle maree abbia luogo nei mari poco profondi e che assommi a 1,7 × 1012 watt per la componente M2: la potenza assorbita (proveniente dalla Luna, dal Sole e dalle maree terrestri) è valutata in 3,5 × 1012 watt. Le correnti che circolano intorno agli atolli e agli altri gruppi insulari, o nelle profondità oceaniche, si ritiene che siano di poca importanza. Cartwright ritiene improbabile che l'energia delle maree possa essere dissipata sotto forma di onde o all'interno della Terra, ma ritiene più probabile che la dissipazione avvenga nella conversione delle normali maree oceaniche in maree interne agli strati di densità dell'oceano.
È noto che negli strati di diversa densità presenti nell'oceano esistono onde con una vasta gamma di frequenze. Molte di queste onde hanno una frequenza relativamente alta, dell'ordine di 10 cicli all'ora, e generalmente un'ampiezza di pochi metri senza nessuna direzione privilegiata; restano da chiarire i dettagli della loro generazione, della loro propagazione e del loro decadimento. Però la frequenza di alcune di queste onde interne coincide con quella delle maree, dando origine a correnti più forti delle maree che esisterebbero in un oceano di densità uniforme. Si ritiene che queste maree interne ricevano energia da quelle superficiali attraverso il moto del fluido stratificato sulle asperità del fondo: infatti esse hanno la frequenza delle maree ma la loro lunghezza d'onda è determinata dalla topografia del fondale e viaggiano molto più lentamente.
Vi è qualche speranza che si progredisca nello sviluppo di un programma di misurazione delle maree in tutto il mondo. Esistono al giorno d'oggi dei registratori di marea adatti per le profondità del mare, ma è logisticamente difficile eseguire le osservazioni nei luoghi, spesso remoti, dove sarebbero più necessarie. È possibile che si riesca a misurare l'altezza della superficie marina per mezzo di altimetri a radar montati su satelliti, la cui orbita si può conoscere con estrema precisione per mezzo di misure effettuate col laser. Il problema tecnico è appena al di sopra delle possibilità attuali e l'interpretazione dei dati sarà ardua, ma, come dice Cartwright, la prospettiva di una copertura quasi globale senza le spese e i problemi logistici connessi con le navi-laboratorio fa riporre nei satelliti la principale speranza per la compilazione di una carta cotidale del mondo. Per lo studio delle maree interne e del possibile ruolo che svolgono nella dissipazione delle maree probabilmente si renderanno necessarie misurazioni oceanografiche effettuate a mezzo di navi.
10. Come modificare l'oceano.
Data la nostra attuale conoscenza dell'ambiente che ci circonda e dei meccanismi che ne conservano le caratteristiche, viene spontaneo chiedersi se sia possibile modificare su larga scala l'ambiente, per il bene dell'umanità. Certo non e un'idea nuova, ma viene proposta sempre più spesso ora che si sono resi disponibili grandi quantitativi di energia ricavata dalle reazioni nucleari controllate. Nei capitoli precedenti abbiamo visto come l'atmosfera e l'oceano insieme costituiscano un motore termico molto grande ma molto poco efficiente, il quale riceve l'energia necessaria per il suo funzionamento dalla radiazione del Sole. Modificare un reattore termonucleare di tali dimensioni e a tale distanza va certamente al di là delle nostre capacità tecniche attuali. Non potremo quindi alterare la quantità di radiazione solare che raggiunge la Terra, ma potremo modificare la sua distribuzione.
Dato che una frazione notevole (circa un terzo) viene riflessa dalla Terra senza essere utilizzata, si potrebbe tentare di cambiare la riflettività della Terra stessa. È stato suggerito di annerire le nevi e i ghiacci delle alte latitudini mediante qualche sostanza idonea, come la fuliggine, o di coprire il Sahara con una pellicola di politene nero. L'oceano assorbe la maggior parte della radiazione solare su di esso incidente, ma potrebbe essere reso più riflettente ricoprendolo con una qualche schiuma bianca adatta, dall'alto potere riflettente. Questo potrebbe avere l'effetto di ridurre il tasso di evaporazione, cui si deve la dissipazione di cerca metà dell'energia proveniente dal Sole.
Si è sperimentato l'uso di sottili pellicole di esadecanolo (una sostanza grassa ricavata dalle teste dei capodogli). Su masse d'acqua di dimensioni ristrette si sono ottenuti sensibili cali dell'evaporazione, e questa tecnica si rivela certamente utile nel tentativo di conservare l'acqua potabile. Però ci sono problemi tecnici per mantenere intatta sotto l'effetto di forti venti una simile pellicola su grandi masse d'acqua.
Questo metodo, come pure tutti i metodi che implicano il deposito di pellicole su grandi superfici, comporta costi molto elevati. Per esempio, immaginiamo di dover depositare una pellicola spessa solo 0,001 mm su una superficie quadrata di 1.000 km di lato. La quantità di sostanza necessaria potrebbe facilmente essere un milione di tonnellate e costare qualcosa come due miliardi di dollari.
Lo stesso genere di calcolo indica il costo proibitivo per formare nubi artificiali nella parte alta dell'atmosfera, al fine di ridurre la radiazione incidente. Abbiamo un'idea dei possibili effetti di una simile operazione, perché immani esplosioni vulcaniche proiettano talvolta grandi quantità di pulviscolo negli strati superiori dell'atmosfera e il clima che ne risulta sembra differire molto poco da quello precedente, tranne per il fatto che i tramonti diventano più pittoreschi.
Cambiare la nuvolosità è un'idea che è stata popolare per molto tempo. I moderni ‛fabbricanti di pioggia' seminano le nuvole con ioduro d'argento, nella speranza che ciò cambi i processi microfisici che portano alla formazione delle gocce di pioggia. I loro sforzi non hanno per ora cambiato molto le precipitazioni piovose e non esistono progetti per un'utilizzazione su scala mondiale di questa tecnica.
Il motore termico aria-mare è poco efficiente e converte in moto solamente una piccola parte dell'energia solare. Tuttavia molta di questa energia viene dissipata per attrito e la si potrebbe diminuire solo modificando il profilo della crosta terrestre. Si potrebbero, per esempio, eliminare delle montagne scelte all'uopo, ma questo è un processo per il quale occorrerebbero o molta fede o alcune bombe da un milione di megatoni! È stato anche proposto di aumentare l'attrito tra aria e terra erigendo nelle praterie e nelle steppe un gran numero di cartelloni pubblicitari, a spese degli inserzionisti.
Proposte più sofisticate coinvolgono l'oceano, e ne sono state avanzate parecchie. La chiusura dello Stretto di Gibilterra avrebbe sicuramente conseguenze tanto oceanografiche e meteorologiche quanto politiche. Il clima dei paesi mediterranei ne risulterebbe profondamente modificato e la circolazione in tutti i mari e nell'Atlantico settentrionale in particolare sarebbe probabilmente alterata dall'assenza dell'acqua calda e salata proveniente dai bacini del Mediterraneo.
Isolare l'Artico bloccando lo Stretto di Bering è un'altra possibilità che è stata considerata. In questo caso si parte dall'idea che la fusione e la solidificazione dell'Oceano Artico sono associate ai mutamenti climatici maggiori, quali le glaciazioni.
Come mai tutti questi suggerimenti su come modificare l'ambiente sembrano fantascientifici? Non perché implicano quantità tanto elevate di energia, o perché verrebbero a costare tanto, ma perché non siamo in grado, con la nostra attuale conoscenza del funzionamento dell'atmosfera e dell'oceano, di prevedere le conseguenze della nostra forza e della nostra ignoranza. Gran parte dei danni fatti dalla scienza derivano dal tentativo di fare del bene, soprattutto in situazioni complesse nelle quali non è stato possibile prevedere le conseguenze di un'azione particolare.
Bibliografia.
Bogdanov, K. T., Magarik, V. A., Numerical solution of the problem of semidiural tides (M2 and Sa) in the ocean, in ‟Dokladȳ kademii nauk SSSR", 1967, CLXXII, pp. 1315-1317.
Cartwright, D. H., Oceanic tides, in ‟Reports on progress in physics", 1977, XL, pp. 665-708.
Groen, P., The waters of the sea, London 1967.
Hasselmann, K. e altri, Measurements of wind-wave growth and swell decay during the Joint North Sea Wave Project (JONSWAP), in ‟Deutsche hydrographische Zeitschrift" (Reihe A 8), 1973, pp. 1-95.
Herring, P. J., Clarke, M. R., Deep oceans, London 1971.
Houghton, H. G., On the annual heat balance of the northern hemisphere, in ‟Journal of meteorology", 1954, XI, pp. 1-9.
Knauss, J. A., Measurement of the Cromwell current, in ‟Deep sea research", 1960, VI, pp. 265-286.
Lafond, E. C., Internal waves, in The sea (a cura di M.N. Hill), vol. I, New York-London 1962, pp. 731-751.
McLellan, H. J., Elements of physical oceanography, London 1965.
Monin, A. S., Kamenkovich, V M., Kort, V. G., Variability of the oceans, New York 1977.
Montgomery, R. B., Water characteristics of Atlantic Ocean and of world ocean, in ‟Deeps sea research", 1958, V, pp. 134-148.
Munk, W. H., On the wind-driven ocean circulation, in ‟Journal of meteorology", 1950, VII, pp. 79-93.
Pickard, G. L., Descriptive physical oceanography, London 1964.
Rossby, T., Voorhis, A. D., Webb, D., A quasi-Lagrangian study of mid-ocean variabilities using long-range SOFAR floats, in ‟Journal of marine research", 1975, XXXIII, pp. 355-382.
Simmons, W., Barnes, N., Schematic representation of variability in the Western North Atlantic, in ‟Oceanus", 1976, XIX, p. 3.
Sverdrup, H. U., Oceanography, in The eath as a planet (a cura di C. F. Kuiper), Chicago 1954, pp. 215-257.
Sverdrup, H. U., Johnson, M. W., Fleming, R. H., The oceans. Their physics, chemistry and biology, Englewood Cliffs, N. J., 1942.
Swallow, J. C., Variable currents in mid-ocean, in ‟Oceanus", 1976, XIX, pp. 18-25.
Swallow, M., Measuring deep currents in mid ocean, in ‟New scientist", 1961, IX, pp. 740-743.
Wüst, G., Oberflachensaltzgehalt, Verdungstung und Niederschlag auf den Weltmeeren, in Fetschrift für Norbert Krebs (Länderkundliche Forschung), Stuttgart 1928, pp. 347-359.