
Origine della vita
Origine della vita
La vita è caratterizzata da un aspetto duplice: (a) l'esistenza individuale, ontogenetica, sincronica e di sviluppo basata sulle proprietà autopoietiche (cioè di autocostruzione) delle cellule e sulle loro conseguenze ecologiche (ossia biogeochimiche); (b) il processo evolutivo storico-collettivo, diacronico, popolazionale. Metabolismo e genetica sono quindi i due pilastri centrali che sostengono la vita. Ogni essere vivente utilizza risorse esterne di energia e di materia per lottare contro disordine, equilibrio e morte. Le membrane e i meccanismi molecolari situati al confine tra l'interno e l'esterno controllano e gestiscono i flussi di energia e di materia a beneficio della cellula stessa. Nelle forme di vita odierne queste attività dipendono dai codici genetici digitali. Le modifiche, le acquisizioni e la perdita di frammenti delle molecole informative sono alla base dei meccanismi di evoluzione, cioè della capacità differenziale di adattamento, sopravvivenza e riproduzione. La persistenza storica dei codici genetici è interamente dipendente dalle capacità metaboliche ed ecologiche degli organismi. È quindi fondamentale sapere in che modo le funzioni metaboliche e genetiche, così inestricabilmente legate nelle forme di vita terrestri, siano diventate una parte integrante della storia del pianeta. Le proprietà autopoietiche della materia vivente sono precedenti a quelle ereditarie?
Nonostante la difficoltà del compito, di recente abbiamo proposto una definizione di vita che cerca di riunire in una frase sintetica entrambi gli aspetti: un essere vivente è un qualsiasi sistema autonomo con capacità evolutive di tipo aperto. Il termine 'autonomia' si riferisce al rapporto tra l'essere vivente e il suo ambiente e alle loro reciproche modificazioni, oltre che alla capacità dell'essere vivente di utilizzare energia e materia per sintetizzare i propri componenti, cioè per autocostruirsi, quindi per crearsi un'identità separata da quella dell'ambiente. Nel caso più semplice, un essere vivente è una cellula. L'espressione 'evoluzione aperta' si riferisce alla capacità di un essere vivente di esplorare nuove funzioni e relazioni con l'ambiente (inclusi altri esseri viventi) e di adattarsi alle diverse situazioni in modi quasi illimitati. La gamma di condizioni fisiche compatibili con 'l'infezione planetaria' di microrganismi è impressionante: laddove siano disponibili una fonte di energia utile (per gli esseri viventi ciò significa fotoni di luce visibile, alcune reazioni chimiche inorganiche, oppure materia organica) e acqua allo stato liquido, si potrà trovare una qualche forma di attività biologica. A livello molecolare, la biodiversità mostra un'unità biochimica altrettanto impressionante: la stessa organizzazione (cellulare) di base, l'uso ricorrente del DNA come materiale ereditario, il codice genetico universale e le variazioni sugli stessi meccanismi bioenergetici (i processi di produzione di energia chimica e chemiosmotica sono simili tra loro). La spiegazione più sintetica di queste osservazioni ormai ben documentate è quella evoluzionista, un'ipotesi che già Charles Darwin aveva avanzato, anche se con meno prove a disposizione: tutte le forme odierne di vita terrestre condividono un progenitore ancestrale comune, il cenancestor universale. Tuttavia, l'individuazione dei diversi stadi del processo evolutivo è oggetto di numerose controversie, e lo stesso punto di partenza (una molecola autoreplicante, o un gene primordiale dotato di un sistema metabolico autocatalitico?) è ancora al centro di vivaci dibattiti.
I primi passi
In che modo possiamo accostarci ai passi primordiali dell'evoluzione della vita? Nell'approccio 'dall'alto', tutti gli organismi conosciuti vengono messi a confronto per poter ricostruire la composizione genetica e metabolica del progenitore comune. Quindi, nell'albero filogenetico universale basato sulla struttura ribosomica, questo significa essenzialmente che vengono paragonati i due domini procariotici, batteri e archei, visto che il dominio degli eucarioti è ritenuto il probabile risultato della simbiosi tra organismi procariotici. I metodi cladistici di ricostruzione di possibili progenitori comuni tra due gruppi di organismi sono tra gli strumenti più potenti a disposizione per fare questi confronti. Tuttavia, le possibilità di risalire ancora più indietro con questo tipo di studio retrospettivo e di azzardare alcune prudenti ipotesi sugli stadi precedenti al progenitore universale sono limitate.
Nella direzione opposta, l'approccio 'dal basso' prende le mosse da risorse cosmochimiche, planetologiche, geologiche, e da qualsiasi altro elemento utile a fornire prove che ricostruiscano l'ambiente, l'inventario chimico e i processi coinvolti nell'origine della vita. D'altro canto, è anche possibile considerare le condizioni geochimiche primordiali in astratto, per poi esaminare i principî teorici che potrebbero essere applicati ai processi più antichi dell'organizzazione della materia. I modelli matematici e le simulazioni al computer possono essere d'aiuto nella definizione di alcune proprietà generali del sistema in questione. Naturalmente, molti passi, stadi e transizioni dall'evoluzione chimica a quella biochimica sono, e saranno per molto tempo ancora, al centro di accese discussioni scientifiche.
Dagli storici contributi di Aleksandr I. Oparin, negli anni Venti del Novecento, la sfida intellettuale sull'enigma dell'origine della vita si è sviluppata a partire dalla premessa che la vita sia nata sulla Terra attraverso processi fisico-chimici che possono essere ipotizzati, compresi e simulati; in altre parole, non vi sarebbero stati né miracoli né generazioni spontanee. I meriti scientifici della proposta di Oparin, dunque, non consistono tanto nei contenuti, sempre soggetti a modifiche con il perfezionarsi della conoscenza scientifica, bensì nella possibilità di testare sperimentalmente la plausibilità di ipotesi storiche e, addirittura, di implementare forme molto semplici di vita artificiale.
Per esempio, nel 1953 l'esperimento di Stanley L. Urey e Harold C. Miller (i quali in un contenitore chiuso riempito da una miscela ricca di metano, ammoniaca, idrogeno e vapore acqueo, con numerose scintille elettriche ottennero degli amminoacidi) diede inizio alla ricerca di chimica prebiotica. Da allora sono state proposte molte reazioni diverse ‒ in particolare, la sintesi di quasi tutti gli amminoacidi e delle basi di acido nucleico ‒ per spiegare l'origine abiotica della materia organica presente nello spazio interstellare, nelle meteoriti, nelle comete e sul nostro pianeta.
Anche se alcune importanti sintesi abiotiche sono più facili da descrivere che da verificare, incluse le sintesi di alcune sostanze molto labili come gli zuccheri (per es., il ribosio), o altri complessi molecolari più sofisticati come i nucleotidi, la partecipazione di catalizzatori minerali (per es., argille come promotori di polimerizzazione e il borato come stabilizzante di zuccheri) facilita alcune delle sintesi più complesse. Ciononostante, non solo è necessario avere uno schema plausibile di sintesi abiotica per una biomolecola, ma anche spiegare come questa si accumulerebbe (se cioè durerebbe abbastanza a lungo da essere utilizzabile) sul pianeta primitivo. Nello scenario che fu delineato da Oparin, le molecole organiche prebiotiche, sia di origine terrestre che extraterrestre, si sono accumulate negli oceani, promuovendo poi l'organizzazione della materia in sistemi primitivi capaci di crescere anaerobicamente dal materiale organico ambientale. Secondo questa prospettiva, la vita sarebbe iniziata da cellule primordiali molto semplici, anossigeniche ed eterotrofe.
Le difficoltà concernenti le reazioni chimiche prebiotiche nell'atmosfera (dal momento che ancora non disponiamo di solide prove sulla sua esatta composizione chimica e sulle sue condizioni fisiche) e le incertezze riguardo all'accumulo di materiale organico extraterrestre hanno spostato l'attenzione sulla ricerca di luoghi alternativi per i processi di vita primordiale. Da qui nasce l'interesse per lo studio dei camini idrotermali sottomarini come culle chimiche della vita. Infatti, alcune simulazioni recenti indicherebbero che la materia organica può avere origine in questi ambienti estremi.
La proposta, da parte di Günter Wächtershäuser, di un'origine della vita termofila e autotrofa ha indotto a concentrare l'attenzione sui camini sottomarini. In questa prospettiva, la sintesi senza ossigeno della pirite sarebbe la fonte dell'energia e degli elettroni necessari a fissare il diossido di carbonio e alla genesi di ogni componente organico della nascente materia vivente. In altre parole, questa ipotesi sostituisce al brodo prebiotico un metabolismo primitivo autotrofo e bidimensionale sulla superficie di pirite: si tratta quindi di un metabolismo acellulare, non istruito geneticamente e favorito dalle alte temperature, che utilizza i gas solforosi ridotti che abbondano nelle sorgenti idrotermali sottomarine. Esiste del materiale sperimentale a supporto di alcuni dei postulati chimici, ma per certi aspetti ancora mancano dati a sostegno di un'origine autotrofa della vita, incluso il carattere ancestrale del ciclo di Arnon. Un metabolismo primordiale chemiolitoautotrofo basato sulla sintesi finale di acetilcoenzima A è stato proposto come collegamento tra una geochimica idrotermale e la biochimica primordiale.
La complessiva diversità dei processi e degli scenari ‒ la generazione abiotica della complessità chimica nella Terra primordiale, sia endogena (atmosfera, superficie marina, camini sottomarini) che esogena (meteoriti, comete, particelle di polvere interstellare), il ruolo dei minerali (catalisi e stabilità) e delle diverse fonti di energia (elettromagnetica, chimica) ‒ ci fa approdare a una visione eclettica del problema dell'origine della vita, in cui l'eterogeneità dei meccanismi e delle situazioni ambientali è essenziale per ricomporre una storia coerente.
Punti di vista a confronto

Le numerose controversie sulle origini della vita hanno profonde radici filosofiche e storiche. In questo senso il dibattito scientifico non differisce da altri conflitti intellettuali che interessano la biologia. La fig. 3 riassume i temi principali in discussione.
Origini eterotrofe versus origini autotrofe
O la vita ha avuto inizio come un sistema semplice, sfruttando un ambiente chimicamente complesso, come hanno suggerito ‒ tra gli altri ‒ Oparin e Haldane, oppure la nascita di sistemi autosufficienti si è basata sull'utilizzo delle molecole più semplici disponibili, secondo l'ipotesi di Wächtershäuser. Se consideriamo che una mappa metabolica eterotrofa è più semplice di una autotrofa (un postulato che è tuttavia ancora da dimostrare), allora la prima opzione è quella più economica, come ammette la maggior parte degli studiosi.
I replicatori precedono i metabolismi?
Si tratta di un vecchio dibattito, che mette a confronto i sostenitori di un'origine precoce dei geni (ancor prima che la loro natura chimica fosse definita, come ipotizzò Hermann J. Muller negli anni Venti del Novecento) con quelli favorevoli a una comparsa precedente delle proteine (per es., Leonard Troland negli anni Dieci del Novecento). Molti scienziati (fra cui Manfred Eigen e Richard Dawkins) ritengono che l'origine della vita equivalga all'origine dei primi replicanti, cioè le prime molecole autoreplicanti. Le intuizioni teoriche e sperimentali (per es., il classico scritto di Leslie E. Orgel sui modelli di replicazione senza enzimi) e la scoperta dell'RNA catalitico (la descrizione dei primi ribozimi proposta da Tom Cech e Sydney Altman nei primi anni Ottanta del Novecento) hanno rinnovato l'interesse per l'idea di una comparsa precoce e spontanea dei polimeri genetici.
Questo è uno dei postulati fondamentali dell'ipotesi del 'mondo a RNA', la presunta soluzione alla versione genetica del quesito se sia nato prima l'uovo o la gallina'. Tuttavia, altri scienziati la pensano diversamente: come sarebbe stata possibile l'origine dell'ordine (per es., i polimeri sintetizzati da monomeri puri in senso chirale ed energetico) senza una modalità plausibile ed efficace per canalizzare l'energia nel sistema? Diversi autori, come Christian de Duve, David Deamer, Tibor Gánti, Harold Morowitz, Stuart Kauffman, Robert Shapiro e Wächtershäuser, sostengono che la comparsa di primitivi sistemi metabolici autocatalitici sia stata il primo passo verso la sintesi dei replicatori. Questi processi prebiotici autorganizzati avrebbero fornito una sorta di struttura ossea per la successiva comparsa della genetica. Tuttavia, i fautori della tesi basata sulla precedenza della replicazione hanno confutato le proposte relative alla precedenza del metabolismo.
Cellularizzazione: prima o dopo?
È tuttora oggetto di discussione se la formazione di compartimenti sia stato un fenomeno tardo o precoce nell'origine della vita, e si tratta di un dibattito strettamente legato alla controversia sull'origine precoce dei metabolismi versus la genetica. Molti sostenitori di una comparsa precoce di replicatori considerano la cellula come un mero compartimento fisico per la separazione dei polimeri dalle diverse capacità replicanti, anche se alcuni autori hanno adottato una posizione intermedia. D'altra parte, esiste un'ampia evidenza biochimica a dimostrazione del fatto che la cellula non è un semplice spazio chiuso da una membrana semipermeabile. Al contrario, il suo interno, sia in senso fisico che chimico, è diverso dall'ambiente esterno per via della presenza funzionalmente attiva di membrane fosfolipidiche e delle loro macchine molecolari (le proteine), ovvero i protagonisti della transduzione e del controllo bioenergetici delle connessioni tra esterno e interno. Per costruire i propri componenti, la materia vivente deve poter accedere alle risorse esterne attraverso le sue biomembrane. L'essenza della bioenergetica consiste nella disposizione asimmetrica delle macchine molecolari sulla membrana cellulare e nei corrispondenti flussi di messaggeri chimici che permettono un efficace accoppiamento tra le fonti primarie di energia (luce visibile, composti chimici) e i sistemi metabolici.
Alcune proteine (ATPasi, catene di trasporto di elettroni) sintetizzano unità di scambio bioenergetiche (chemiosmotiche o chimiche), mentre altre utilizzano tali unità per eseguire un'ampia gamma di compiti cellulari: motilità, biosintesi di architetture complesse, mantenimento dell'identità a fronte delle fluttuazioni ambientali, eredità, ecc. È difficile immaginare un altro modo (per non parlare dei sistemi acellulari) di costruire e mantenere strutture complesse, dai comportamenti altrettanto complessi, utilizzando energia solare o reazioni chimiche a base di energia proveniente dall'esterno e semplici particelle elementari.
Dunque la vita sarebbe stata ab initio cellulare, anche se permangono molti aspetti controversi, fra i quali la natura chimica delle membrane e il tipo di trasduttori molecolari primitivi di fonti primarie di energia e di trasportatori della membrana, necessari per prevenire una crisi osmotica prima dell'esistenza di macchine proteiche complesse. Quest'argomento ha pervaso perfino le strategie dal basso per la sintesi artificiale (chimica) della vita, coinvolgendo gli scienziati interessati al concetto di ribozimi autoreplicanti o all'implementazione chimica dell'autopoiesi attraverso vescicole lipidiche autorganizzate e autoriproduttive. Altre ricerche hanno tentato di trovare un terreno comune nei sistemi chimici, come le vescicole contenenti sia ribozimi autoreplicanti che ribozimi coinvolti nella sintesi delle componenti anfipatiche della vescicola. La nostra comprensione della natura e dell'origine della vita migliorerà quando riusciremo a colmare, con ipotesi fondate, la distanza tra questi approcci dal basso, sia teorici che sperimentali, e le strategie dall'alto finalizzate a immaginare le forme più semplici di vita.
Un'ipotesi di percorso per il passaggio dalla materia inerte alla materia vivente
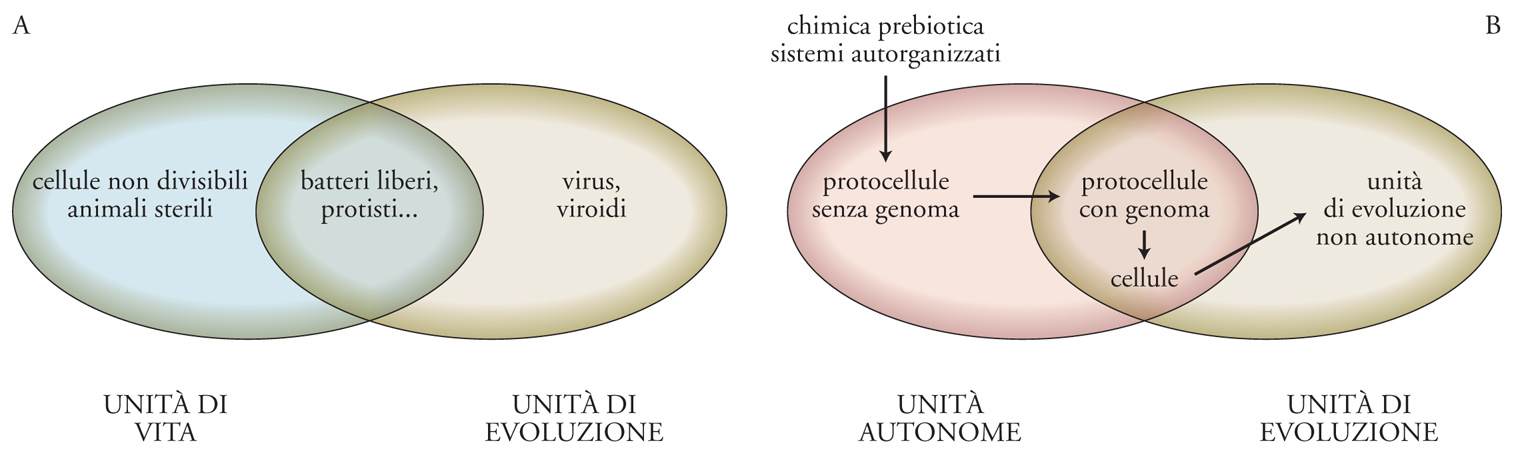
Eörs Szathmáry ha proposto una distinzione tra unità di vita (entità che mostrano autosostentamento) e unità di evoluzione (entità le cui istruzioni sono replicabili, mutabili ed ereditabili). Alcuni esempi tipici di esseri viventi (per es., i batteri liberi) sono sia unità di vita che unità di evoluzione, quindi si collocano sulla linea d'intersezione tra le due categorie. Tuttavia, si possono immaginare anche casi limite: unità di vita incapaci di evolvere (animali sterili), oppure unità di evoluzione strettamente dipendenti da unità di vita (parassiti obbligati, come virus e viroidi, fig. 4A).
La prospettiva genealogica sulla vita può essere combinata con lo schema di Szathmáry, ribattezzando le unità di vita come unità autonome. In questo caso, il primo passo nell'autorganizzazione della materia nella Terra primitiva sarebbe stata la comparsa di unità autonome prive della capacità di evoluzione in senso darwiniano. In altre parole, l'evoluzione chimica avrebbe dato luogo a sistemi chimici primitivi capaci di autosostentamento, dotati di meccanismi rudimentali di trasduzione energetica, ma privi di codici digitali (replicanti e mutabili). Questi sistemi 'infrabiologici', che comportano un'associazione primordiale tra metabolismo e membrana, sono denominati protocellule (fig. 4B). Una volta acquisiti i codici genetici e superato il metabolismo non istruito da uno geneticamente istruito, ha avuto inizio uno stadio evolutivo (fig. 4B).
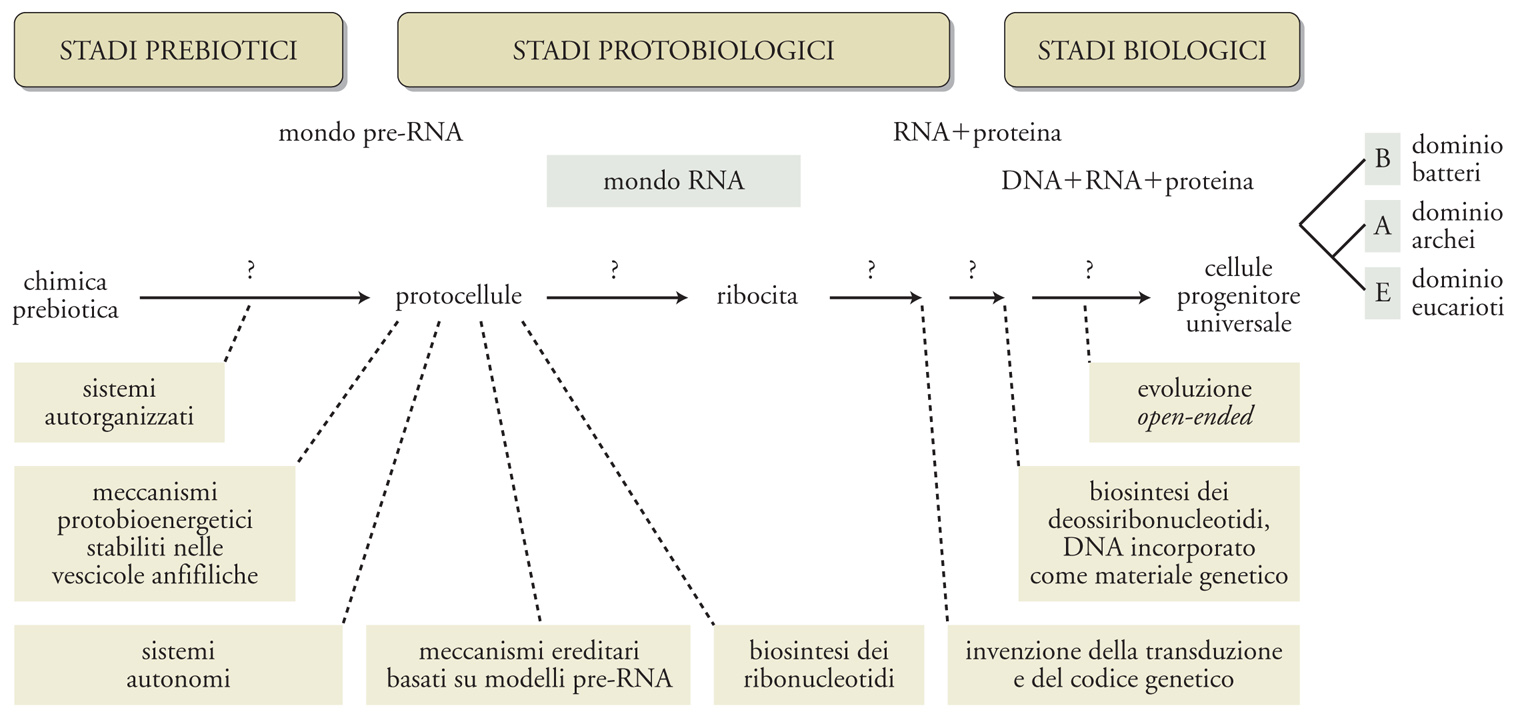
Una protocellula dipendente da un singolo polimero potrebbe essere la versione primordiale di questi sistemi, un mondo a RNA cellulare. È opportuno segnalare che gran parte di coloro che propongono questo scenario con un polimero singolo fanno riferimento a un ipotetico mondo a RNA acellulare, che tuttavia non sembra molto plausibile in una prospettiva prebiotica. L'invenzione della sintesi proteica catalizzata dall'RNA e la determinazione di un codice hanno introdotto un secondo passaggio cruciale: da un metabolismo primitivo catalizzato dal ribozima al controllo dinamico dei sistemi metabolici da parte delle proteine. Questo fondamentale passaggio evolutivo ha permesso lo sdoppiamento tra fenotipo (organizzazione) e genotipo (struttura), così peculiare della vita terrestre. Un ultimo, importante passaggio è stato la sostituzione del DNA all'RNA come materiale genetico, che ha segnato l'inizio del processo di evoluzione aperta, avendo fra le sue conseguenze la formazione delle cellule del progenitore universale. Alla fine, alcune unità autonome con capacità evolutive aperte hanno completamente perso la loro autonomia (cioè le loro capacità metaboliche) per diventare dipendenti obbligati di altre unità autonome (per es., virus e viroidi, fig. 4B). La fig. 5 descrive più in dettaglio i processi coinvolti nell'origine della vita secondo la nostra proposta, dalla chimica prebiotica alle protocellule, alle cellule. Se partiamo dal presupposto che l'origine della vita è stata necessariamente l'inizio dell'evoluzione organica, allora tutti gli stadi precedenti alla fase dell'evoluzione aperta dovrebbero essere considerati protobiologici.
Per concludere, durante la vita individuale la comunicazione con l'ambiente è permanente, mentre durante la storia evolutiva vi sono alcuni casi di comunicazione (unidirezionale) tra popolazioni del passato e del presente. All'origine della vita, sono emersi per primi l'organizzazione e i meccanismi che hanno permesso a un sistema chimico di collegarsi materialmente e in senso energetico (ecologico) all'ambiente, dotandolo della capacità di autocostruirsi. Anche ai primordi i sistemi ecologici sono stati la culla dell'evoluzione. Ciò ha preceduto la comparsa di strutture chimiche complesse, che hanno fornito al sistema un legame temporale attraverso le generazioni successive. In futuro, le linee di ricerca sulle origini della vita dovranno stabilire una maggiore interazione tra chi propone che l'origine della vita sia stata l'origine della vera eredità ‒ si potrebbe addirittura dire l'origine del primo gene ‒ e chi ritiene che nessun modello biopoietico meriti di essere preso in seria considerazione, a meno che non fornisca una fonte energetica evidente e sostenibile, oltre a modalità plausibili di accoppiamento tra l'energia e il flusso di materia all'interno del sistema emergente.
Bibliografia
Bada, Lazcano 2002: Bada, Jeffrey L. - Lazcano, Antonio, The origin of life: some like it hot, but not the first biomolecules, "Science", 296, 2002, pp. 1982-1983.
De Duve 2005: de Duve, Christian, The onset of selection, "Nature", 433, 2005, pp. 581-582.
Dworkin 2002: Dworkin, Jason P. - Lazcano, Antonio - Miller, Stanley L., The roads to and from the RNA world, "Journal of theoretical biology", 222, 2002, pp. 127-134.
Ehrenfreund 2002: Ehrenfreund, Pascale e altri, Astrophysical and astrochemical insights into the origin of life, "Reports on progress in physics", 65, 2002, pp. 1427-1487.
Gánti 2003: Gánti, Tibor, The principles of life. With a commentary by J. Griesemer and E. Szathmáry, Oxford, Oxford University Press, 2003, p. 159.
Luisi 2004: Luisi, Pier Luigi e altri, A possible route to prebiotic vesicle reproduction, "Artificial life", 10, 2004, pp. 297-308.
Orgel 2004: Orgel, Leslie E., Prebiotic chemistry and the origin of the RNA world, "Critical reviews in biochemistry and molecular biology", 39, 2004, pp. 99-123.
Ruiz-Mirazo 2004: Ruiz-Mirazo, Kepa - Peretó, Juli - Moreno, Álvaro, A universal definition of life: autonomy and open-ended evolution, "Origins of life and evolution of the biosphere", 34, 2004, pp. 323-346.
Szathmáry 2005: Szathmáry, Eörs, Life: in search of the simplest cell, "Nature", 433, 2005, pp. 469-470.
Wächtershäuser 1988: Wächtershäuser, Günter, Before enzymes and templates: theory of surface metabolism, "Microbiological reviews", 52, 1988, pp. 452-484.