
Vita, origine della
Vita, origine della
di Cyril Ponnamperuma
SOMMARIO: 1. Introduzione: a) le basi scientifiche; b) l'evoluzione chimica. □ 2. Concetti e teorie prima del nostro secolo: a) generazione spontanea; b) la chimica organica e la disfatta del vitalismo; c) le teorie scientifiche. □ 3. Le condizioni sulla Terra in epoca prebiotica: a) il materiale grezzo; b) la natura dell'atmosfera primitiva; c) il passaggio dall'atmosfera primitiva all'atmosfera contenente ossigeno; d) fonti di energia. □ 4. Esperimenti effettuati in laboratorio: a) amminoacidi; b) purine e pirimidine; c) monosaccaridi; d) nucleosidi e nucleotidi; e) idrocarburi e acidi grassi; f) porfirine; g) sintesi delle grandi molecole; h) polioeptidi; i) polinucleotidi. □ 5. Lo sviluppo delle nuove teorie scientifiche: a) organizzazione precellulare; b) interazione tra acidi nucleici e proteine. □ 6. La ricerca delle prime forme di vita: a) macropaleontologia; b) fossili molecolari. □ 7. La ricerca di forme di vita extraterrestri: a) analisi dei campioni lunari; b) condriti carboniose; c) la ricerca della vita su Marte; d) Giove, Saturno e Titano; e) molecole interstellari; f) esobiochimica. □ 8. L'acqua e l'origine della vita. □ 9. Attività ottica e origine della vita. □ 10. La ricerca di intelligenze extraterrestri. □ 11. Conclusioni. □ Bibliografia.
1. Introduzione.
L'uomo, fin dagli albori della sua esistenza, ha cominciato a porsi il fondamentale problema di come sia iniziata la vita. Insieme al problema dell'origine dell'Universo, l'origine della vita e quella dell'intelligenza possono essere considerati tra gli interrogativi essenziali di ogni scienza. Questo articolo descrive gli sforzi della scienza moderna per trovare una soluzione al problema dell'origine della vita. ‟Probabilmente, anche la sola formulazione di questo problema è al di là delle possibilità di ogni singolo scienziato, perché questi dovrebbe contemporaneamente essere un buon matematico, un fisico e un ottimo chimico organico, dovrebbe conoscere a fondo geologia, geofisica e geochimica, e per di più dovrebbe essere versato in tutte le discipline biologiche. Prima o poi questo compito dovrà venire affidato a gruppi di specialisti in tutte queste materie, che lavorino in stretto collegamento, sia nel campo teorico che in quello sperimentale". Questo era il parere espresso da J. D. Bernal (v., 1951; tr. it., pp. 6-7). Peraltro, oggi abbiamo forse ragione di essere più ottimisti: per la prima volta nella storia dell'uomo, infatti, scienze che erano nate come discipline separate si uniscono l'una all'altra, e la nostra generazione è stata testimone del sorgere di un lavoro interdisciplinare tra biofisici, biochimici, biologi molecolari e astrochimici.
a) Le basi scientifiche.
Tre fattori hanno permesso di affrontare in modo scientifico il problema di come è cominciata la vita, da un punto di vista non solo teorico ma anche sperimentale: le scoperte astronomiche del nostro secolo, i recenti progressi della biochimica e il trionfo della teoria dell'evoluzione di Darwin. Gli umanisti possono ritenere assurdo che un problema così complesso debba essere considerato di pertinenza del laboratorio, ma il ricercatore ha buone speranze che le sue indagini riusciranno un giorno a svelare questo profondo mistero.
Le scoperte astronomiche di questo secolo hanno relegato la Terra ai margini di un Universo costituito da miliardi di stelle. Lo studio dei cieli compiuto con i moderni telescopi ha rivelato la presenza di più di 1023 stelle. Al pari del nostro Sole, ognuna di queste stelle può fornire la base fotochimica necessaria alla vita vegetale e animale. Due fatti sono divenuti assolutamente chiari: che non c'è niente di unico nel nostro Sole, al quale peraltro dobbiamo la vita su questo pianeta, e che ci sono più di 1023 possibili fonti di vita nel nostro Universo. In vista della possibilità che esistano numerose condizioni limitanti, una valutazione restrittiva fatta dagli astronomi ha ridotto il aumero dei luoghi nell'Universo in cui la vita sarebbe possibile a circa l'1%; calcoli più ottimistici arrivano al 50% (v. Ponnamperuma e Cameron, 1974). In ogni caso, le possibilità di esistenza di vita nell'Universo sono almeno 1021. Esistono numerosissimi pianeti, e sulla base dei calcoli che abbiamo riportato la Terra sarebbe solamente uno dei luoghi in cui è stato compiuto un ‛esperimento' che ha avuto successo. Se le leggi della fisica e della chimica sono leggi universali, dovrebbe esser possibile considerarle valide anche in qualunque altro posto nell'Universo.
Se passiamo dalle stelle alla Terra, possiamo constatare come le recenti scoperte nel campo della biochimica abbiano messo in evidenza l'estrema omogeneità della materia vivente. In tutti gli organismi viventi, dal più piccolo microbo al più grande mammifero, esistono due gruppi di molecole fondamentali, gli acidi nucleici e le proteine. È dalla loro interazione che deriva quella straordinaria proprietà della materia che da un punto di vista biochimico è generalmente definita col termine ‛vita'. Mentre ognuna di queste molecole ha una struttura complessa, le unità che le compongono sono in numero limitato. Le molecole degli acidi nucleici sono costituite da nucleotidi uniti l'uno all'altro in serie. I nucleotidi, a loro volta, sono costituiti da una base purinica o pirimidinica, da una molecola di zucchero e da una di fosfato. Nelle molecole proteiche venti diversi amminoacidi sono legati l'uno all'altro per costituire una macromolecola; tra acidi nucleici e proteine, i componenti base utilizzati sono ventotto. È con queste lettere che pare sia scritto l'intero libro della vita. Pertanto, lo studio della composizione della materia vivente ci porta all'inevitabile conclusione che tutti gli organismi viventi devono avere avuto un'ascendenza chimica comune. In realtà si potrebbe andare ancora oltre e sostenere che non solo le molecole, ma addirittura i processi vitali abbiano natura universale. I biochimici sono portati a ritenere che quello che è vero per Escherichia coli può essere vero anche per l'elefante, il che ha particolare interesse nello studio del codice genetico, che è stato dimostrato avere validità universale per tutti gli organismi viventi (v. acidi nucleici e biologia).
Un altro fattore che ha permesso lo studio scientifico di questo problema è stato il trionfo della teoria evoluzionistica di Darwin, secondo la quale le più alte forme di vita si sono evolute dalle più basse nel corso di un lunghissimo periodo dell'esistenza del nostro pianeta. L'analisi dei fossili ha mostrato che le più antiche forme conosciute di sistemi viventi possono risalire a tre miliardi e mezzo di anni fa (v. Knoll e Barghoorn, 1977; v. Walter e altri, 1980; v. Orpen e Wilson, 1981). I dati geochimici ci dicono che la Terra ha 4,6 miliardi di anni (v. Tilton e Steiger, 1965); l'inizio della vita su questo pianeta, pertanto, deve essere fatto risalire a un periodo compreso tra i 4,6 miliardi di anni, che è l'età della Terra, e i 3,5 miliardi di anni, data alla quale è provata l'esistenza di un microbionte. Il primo problema che nasce riguarda la storia del nostro pianeta nel periodo compreso tra la sua nascita, 4,6 miliardi di anni fa, e l'origine della vita. Questo problema assillò la mente del fisico J. Tyndall, che nel 1871 così scrisse nel libro Fragments of science for unscientific people: ‟Darwin pose alla base della vita un germe primordiale dal quale egli riteneva potesse esser derivata la straordinaria ricchezza e varietà della vita che attualmente ritroviamo sulla superficie della Terra. Se questa ipotesi fosse vera, non sarebbe comunque definitiva. L'immaginazione umana sicuramente vorrebbe risalire al di là di questo germe, per quanto disperato sia questo tentativo, per cercare di capire la storia della sua genesi. Nasce spontaneo il desiderio di connettere la vita attuale del nostro pianeta col passato, di conoscere qualcosa dei nostri più remoti antenati. La vita è una proprietà intrinseca di ciò che noi indichiamo come materia, o è un principio indipendente che si è inserito nella materia in un periodo appropriato in cui le condizioni fisiche erano divenute tali da permettere lo sviluppo della vita stessa?" Queste riflessioni sull'evoluzione biologica ci portano a prendere in considerazione un'altra forma di evoluzione, e cioè l'evoluzione chimica. (V. anche evoluzione e paleontologia).
b) L'evoluzione chimica.
Si può ritenere che l'evoluzione chimica sia avvenuta in tre stadi: dalla chimica inorganica a quella organica, e da questa a quella biologica. Il primo stadio dell'evoluzione chimica è cominciato probabilmente con l'origine stessa della materia. Nella serie di reazioni cataclismatiche che hanno luogo con la nascita di una stella si devono essere formati gli elementi della tavola periodica. Studi sulla sintesi nucleare hanno dimostrato che l'origine degli elementi può essere ascritta in primo luogo al big bang, che segnò l'inizio della formazione del nostro Universo con la nascita delle stelle, quindi alle stelle di prima generazione, in cui si sarebbe verificata la fusione nucleare dell'idrogeno; il processo di fusione di elio, carbonio e silicio avrebbe dato infine origine agli elementi fino al ferro. In seguito, la seconda generazione di stelle derivanti dalle supernove avrebbe dato origine agli elementi più pesanti, situati dopo il ferro nella tavola periodica. Durante la formazione del sistema solare, per la quale è stato probabilmente utilizzato materiale reso disponibile dall'esplosione di una supernova - è certo che il nostro Sole è una stella di seconda generazione - nella nebula primordiale si formarono proprio quegli elementi che sono necessari alla vita (v. Shapley, 1958; v. origine degli elementi).
Nell'ambito di questa ipotesi, quindi, la vita può essere considerata come una proprietà speciale del nostro pianeta, derivante dallo sviluppo ordinato del pianeta stesso.
2. Concetti e teorie prima del nostro secolo.
a) Generazione spontanea.
Gli antichi credettero di aver trovato una facile soluzione a questo problema, in quanto ritenevano che la vita si generasse dal non vivente in un processo senza soluzione di continuità (v. Conant, 1959). Bastava accettare l'evidenza delle nostre percezioni sensoriali: i vermi emergevano dal fango, le larve delle mosche dalla carne putrescente, i topi dagli stracci vecchi, le lucciole dalla rugiada del mattino. Aristotele aveva sostenuto nella Metafisica la dottrina della generazione spontanea, e i suoi insegnamenti erano stati accettati per lungo tempo dai pensatori europei che lo avevano considerato l'autorità suprema in fatto di fisica e metafisica. Anche secondo le antiche scritture hindu il vivente si era evoluto dal non vivente: il Rgveda sosteneva che la vita aveva avuto inizio dagli elementi primi, mentre negli Atharvaveda si postulava che la culla di ogni forma vivente fossero gli oceani della Terra. La letteratura mondiale è piena di allusioni alla credenza popolare della generazione spontanea: Lucrezio nel De rerum natura considera la Terra come madre di ogni creatura vivente; Virgilio nelle Georgiche racconta di come uno sciame di api si fosse generato dalla carcassa di un vitello; il chimico e fisico belga J. B. van Helmont aveva addirittura una ‛ricetta' per fabbricare i topi. Credenze di questo tipo non potevano resistere a lungo al crescente rigore dei metodi scientifici. Francesco Redi, uno dei più famosi membri dell'Accademia del Cimento, provò che i vermi della carne in putrefazione erano larve di mosche, e le sue semplici dimostrazioni non lasciavano adito a dubbi; infatti, se si metteva un pezzo di carne sotto uno schermo in modo che le mosche non potessero deporvi le loro uova, non si sviluppava alcuna larva. Ma la dottrina della generazione spontanea ricomparve non appena A. von Leeuwenhoek inventò il microscopio, nella seconda metà del XVII secolo, in quanto alcuni ricercatori non riuscivano a spiegare l'origine dei molti organismi che il microscopio metteva in evidenza in soluzioni contenenti materia animale o vegetale. In questi organismi non riuscirono a scoprire niente che somigliasse al processo della generazione sessuata, e ciò li spinse a ritenere che la materia vivente mantenesse anche dopo la morte una speciale vitalità. All'incirca in questo periodo il sacerdote inglese J. Turberville Needham descrisse alcuni esperimenti che dovevano sostenere la teoria della generazione spontanea, ma fu contraddetto dall'abate Lazzaro Spallanzani, il quale dimostrò che un brodo di coltura mantenuto fuori del contatto dell'aria mentre bolliva, non generava mai dei microrganismi, e pertanto non andava mai a male. Needham obiettò che Spallanzani con la prolungata bollitura aveva reso il brodo e l'aria incompatibili con la vita, ma Spallanzani potè controbattere questa interpretazione sulla base di ulteriori esperimenti.
Verso la metà del secolo scorso, Louis Pasteur dimostrò con una serie di brillanti esperimenti che gli organismi viventi non potevano aver origine da materia non vivente, dando il colpo di grazia alla teoria della generazione spontanea, che era basata su osservazioni inadeguate e sul desiderio di accettare l'evidenza delle percezioni sensoriali. Purtroppo, l'opera di Pasteur fece nascere l'errata convinzione che il problema dell'origine della vita non potesse essere affrontato con metodi scientifici, e pertanto non fosse degno di attenzione da parte di uno scienziato serio. Ma è assolutamente chiaro che ciò che Pasteur aveva dimostrato era l'impossibilità di far sviluppare dei microrganismi da un materiale sterile. I suoi esperimenti, peraltro, non avevano niente a che vedere con la graduale formazione dei composti organici che portò alla comparsa di polimeri significativi da un punto di vista biologico.
b) La chimica organica e la disfatta del vitalismo.
Con lo sviluppo della chimica organica parve possibile affrontare in modo nuovo il problema dell'origine della vita. Berzelius pensava che i composti organici fossero prodotti in base a leggi diverse da quelle che regolano la formazione dei composti inorganici. Questo lo portò a ritenere che i composti organici si formassero sotto l'influenza di una ‛forza vitale', e che non potessero essere preparati artificialmente.
Nel 1828, Friedrich Wöhler trasformò il cianato d'ammonio in urea, una sostanza fino ad allora ottenuta solo da fonte animale. Questa sintesi indebolì la teoria che sosteneva la distinzione tra composti organici e inorganici. Tale distinzione scomparve definitivamente con la sintesi dell'acido acetico dagli elementi, operata da Kolbe nel 1845, e con quella del metano da un miscuglio di solfuro di carbonio e solfuro di idrogeno, compiuta da Marcelin Berthelot nel 1856. I vitalisti avevano obiettato che Wöhler aveva ottenuto il suo cianato d'ammonio da ammoniaca e acido cianico, che erano entrambi di origine animale. Ma già nel 1751 Joseph Priestley aveva preparato l'ammoniaca per riduzione dell'acido nitrico, che Henry Cavendish nel 1785 sintetizzò dai suoi elementi. Karl Wilhelm Scheele, riscaldando una miscela di carbonato di potassio e carbonio in presenza di azoto, aveva ottenuto anche il cianuro di potassio. Dato che il cianuro di potassio viene rapidamente trasformato in cianato di potassio, la sintesi operata da Wöhler può veramente considerarsi iniziata partendo da materiali inorganici. In conclusione, gli esperimenti di sintesi di questi primi chimici eliminarono gradualmente i confini tra il mondo inorganico e quello organico (v. Partington, 1957).
c) Le teorie scientifiche.
Uno dei primi a riflettere sulle condizioni necessarie per l'origine della vita fu Erasmus Darwin, il nonno di Charles Darwin. Nel suo Temple of nature (1803) aveva scritto: ‟Tutti i vegetali e gli animali adesso esistenti ebbero inizialmente origine dai più piccoli corrispondenti esseri microscopici formatisi per vitalità spontanea". Forse questa idea influenzò lo stesso pensiero di Charles Darwin, il quale, vari anni dopo, scrisse al suo amico Hooker di un ‟piccolo stagno caldo" pieno di ammoniaca, sali fosforici, luce, calore, elettricità, ecc. in cui egli postulava si fosse formato chimicamente un composto proteico, pronto ad andare incontro a cambiamenti ben più complessi (v. Darwin, 1959).
Circa nello stesso periodo, il fisico John Tyndall (v., 1871) sostenne che qualsiasi parte di un organismo vivente può essere ridotta a materia inorganica. Nel suo saggio sulla vitalità, del 1866, egli aveva suggerito che poteva essere concepito un cambiamento inverso, dalla materia inorganica a quella organica, e che la speciale disposizione degli elementi nei corpi viventi portava al fenomeno della vita. Nel 1868, Thomas Huxley tenne una conferenza a Edimburgo, nella quale fece notare che il protoplasma era sostanzialmente lo stesso nell'intero ambito degli esseri viventi. Per lui, l'esistenza della vita dipendeva da certe molecole come l'acido carbonico, l'acqua e i composti azotati: composti senza vita, ma che, una volta messi insieme, davano origine al protoplasma.
È al biochimico russo A. I. Oparin (v., 1924) che dobbiamo, più che a ogni altro, le nostre attuali idee su come affrontare scientificamente il problema dell'origine della vita. In termini chiari e scientificamente giustificabili, questo scienziato ha sostenuto che non c'è alcuna differenza fondamentale tra organismi viventi e materia bruta. Quella complessa combinazione di aspetti e di proprietà che è caratteristica della vita sarebbe scaturita dal processo di evoluzione della materia. Nel 1928, il biologo inglese Haldane ha espresso le sue idee sull'origine della vita; egli ha attribuito la sintesi dei composti organici all'azione della luce ultravioletta sull'atmosfera primitiva della Terra, e ha suggerito che i composti organici possano essersi accumulati fino a che gli oceani primitivi raggiunsero la consistenza di un ‛brodo' primordiale. Venti anni dopo la comparsa del saggio di Haldane, Bernal (v., 1951) espose la seguente teoria in una conferenza tenuta alla British Physical Society: ‟Condensazioni e deidrogenazioni portano necessariamente a sostanze sempre più insature e, alla fine, a strutture semplici e forse anche ad anelli condensati contenenti quasi certamente azoto, quali le pirimidine e le purine. La comparsa di queste molecole ha reso possibili sintesi ulteriori. Peraltro, la difficoltà fondamentale nel concepire che i processi chimici siano andati avanti fino a tal punto risiede nella estrema diluizione del sistema, se davvero essi hanno avuto luogo negli oceani. La concentrazione dei prodotti è il prerequisito necessario per ogni ulteriore evoluzione".
3. Le condizioni sulla Terra in epoca prebiotica.
a) Il materiale grezzo.
Il materiale grezzo da cui si sono formati i mattoni coi quali è costruita la vita è rappresentato dagli elementi della tavola periodica. L'esame della crosta terrestre, degli oceani e dell'atmosfera ci fornisce informazioni sull'abbondanza di questi elementi sulla Terra. Dati sulla composizione fondamentale della materia al di fuori della Terra sono stati forniti da diverse fonti. L'analisi spettroscopica della luce delle stelle ci ha rivelato la natura degli elementi che le compongono. Lo sviluppo della radioastronomia ci ha messo a disposizione la tecnica delle microonde, con la quale siamo in grado di mettere in evidenza vari elementi e specie eccitate nello spazio intergalattico. Le particelle dei raggi cosmici ci possono fornire campioni di materia extraterrestre. Le meteoriti e i campioni lunari ci hanno dato preziose cognizioni sulla composizione del nostro sistema solare. Possiamo così avere un quadro ragionevole e coerente della distribuzione degli elementi nell'Universo.
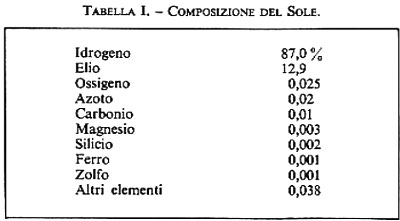
L'idrogeno è di gran lunga l'elemento più abbondante. Esso rappresenta il 93% del numero totale di atomi nell'Universo, e il 76% del suo peso. L'elio rappresenta quasi il 7% del numero totale di atomi e il 23% del peso. Se vengono poste in grafico le abbondanze atomiche in funzione dei pesi atomici, si osserva in genere una correlazione inversa, tranne una decisa interruzione nel caso degli elementi del gruppo del ferro. La relativamente alta abbondanza di questi elementi può essere collegata col fatto che le reazioni nucleari a cui prendono parte devono assorbire energia piuttosto che rilasciarla. Se si esclude l'elio, tutti gli altri elementi insieme contribuiscono per solo poco più dell'1% al peso dell'intero Universo. Le teorie di Gamow e Hoyle hanno tentato di delineare i processi mediante i quali furono formati, in modo graduale, gli elementi. Per spiegare i processi di sintesi nucleare sono stati invocati sia la cattura di neutroni che la fusione dell'idrogeno e dell'elio. C'è un parallelismo tra la graduale formazione degli elementi della tavola periodica da una parte, e lo sviluppo ed evoluzione delle stelle dall'altra. L'abbondanza atomica di alcuni degli elementi che si trovano nel Sole è riportata nella tab. I. La composizione del Sole può essere considerata rappresentativa della composizione media propria del sistema solare; l'idrogeno, l'elio, il carbonio, l'azoto e l'ossigeno sono gli elementi più abbondanti. A eccezione dell'elio, sono proprio questi elementi che costituiscono il 99% della materia vivente.
b) La natura dell'atmosfera primitiva.
L'effettiva comprensione della natura dell'atmosfera primitiva della Terra è il logico punto di partenza per qualsiasi discussione sul problema dell'origine chimica della vita. Mentre è difficile rispondere attraverso osservazioni dirette alle domande che emergono da tale problema, prove di vario tipo indicano che un'atmosfera riducente è stata gradualmente sostituita dall'odierna atmosfera ricca di ossigeno (v. Urey, 1952). Dati raccolti da osservazioni astronomiche, astrofisiche, chimiche, geologiche, biochimiche e dallo studio delle meteoriti possono essere utilizzati per chiarire questo problema. L'attuale rarità dei gas nobili nell'atmosfera terrestre in confronto alla loro distribuzione nell'Universo indica che l'atmosfera che è generalmente denominata atmosfera primitiva della Terra è, in realtà, successiva a una precedente di cui si è perduta traccia, probabilmente derivata da una fuoriuscita di gas dall'interno della Terra durante il periodo di accrezione dei pianeti. La successiva atmosfera deve essere stata molto simile alla prima, ed è a questa che ci riferiremo nella nostra discussione sull'atmosfera primordiale della Terra. La scoperta di H. N. Russell che l'idrogeno è l'elemento più abbondante nell'Universo indica che l'Universo stesso, considerato nel suo insieme, ha natura riducente. Uno dei problemi principali riguardo alla natura dell'atmosfera terrestre è pertanto quello di spiegare come da un'atmosfera con una così grande quantità di idrogeno può esserne derivata una contenente ossigeno libero.
I pianeti più piccoli per la maggior parte non sono riusciti a trattenere la loro atmosfera originale, mentre i pianeti maggiori, quali Giove e Saturno, contenevano grandi quantità di idrogeno, metano, ammoniaca e acqua. L'analisi di meteoriti a cui è stata attribuita un'età di 4,5 miliardi di anni ha fornito dei campioni della materia primordiale da cui si è formato il nostro sistema planetario. I metalli trovati in queste meteoriti sono generalmente nella loro forma ridotta, e lo studio degli equilibri chimici ci fornisce qualche indicazione circa la natura delle reazioni che devono aver avuto luogo nella primitiva atmosfera riducente. Le proprietà termodinamiche del carbonio, del diossido di carbonio, del metano, dell'ammoniaca e dell'acqua sono assai ben conosciute (v. tab. II). La composizione delle miscele all'equilibrio di queste sostanze è stata determinata in modo soddisfacente. Le costanti di equilibrio per la formazione del metano da carbonio e idrogeno, dell'acqua da ossigeno e idrogeno, e dell'ammoniaca da azoto e idrogeno sono tutte molto elevate, ed è pertanto ragionevole supporre che, in presenza di un grande eccesso di idrogeno, il carbonio, l'azoto e l'ossigeno esistessero nel loro stato ridotto.

c) Il passaggio dall'atmosfera primitiva all'atmosfera contenente ossigeno.
La presenza di ossigeno libero è una caratteristica peculiare del nostro sistema planetario. L'ossigeno che si trova nell'attuale atmosfera sembra si sia originato in due modi: per fotodissociazione dell'acqua da parte dei raggi ultravioletti a breve lunghezza d'onda, nella parte superiore dell'atmosfera, e per fotosintesi da parte delle piante. La fotosintesi probabilmente ebbe inizio quando si sviluppò, nella parte superiore dell'atmosfera, lo strato di ozono che ha protetto la Terra dai raggi ultravioletti emessi dal Sole, impedendo così l'ulteriore sintesi fotochimica di composti organici nell'ambiente primitivo. Gli organismi eterotrofi, che traevano il loro nutrimento da materiali prodotti fotochimicamente, furono sostituiti da organismi autotrofi, che erano in grado di fotosintetizzare i propri alimenti, dopo che si fu esaurito il materiale organico biogeno. Questo cambiamento può essere schematicamente raffigurato come una clessidra: solo gli organismi capaci di incorporare molecole quali le porfirine e di utilizzare lunghezze d'onda maggiori riuscirono a passare attraverso il collo della clessidra (v. fig. 1), mentre tutti gli altri si estinsero. Gli organismi che sopravvissero e andarono incontro a un ulteriore sviluppo riuscirono a evolversi nella grande varietà di forme di vita che vediamo sulla Terra oggi.
Nelle reazioni biochimiche che avvengono negli attuali organismi viventi, il carbonio deve essere ridotto prima di poter essere incorporato in amminoacidi, purine, pirimidine, ecc. Questo è un fenomeno universale nella biosfera. È merito della grande capacità intuitiva di Oparin (v., 1924) l'aver capito, anche prima che fosse stata scoperta la natura riducente dell'Universo, che queste dovevano esser state le condizioni prevalenti per almeno un miliardo di anni.
Altri studiosi non sono d'accordo su questo punto. Rubey (v., 1955) ha suggerito che l'atmosfera di metano e ammoniaca non sia durata più di 105-108 anni. Dato che la principale fonte di diossido di carbonio è rappresentata dai vulcani, oggi Rubey sostiene che, se l'atmosfera primitiva della Terra si formò per fuoriuscita di gas dall'interno della Terra stessa, il carbonio deve essere comparso sotto forma di diossido di carbonio. Con ragionamenti dello stesso tipo, Abelson (v., 1966) ha postulato che l'atmosfera primitiva della Terra consistesse prevalentemente di monossido e diossido di carbonio, azoto e una minore quantità di idrogeno; inoltre, egli ha dimostrato che l'ammoniaca deve essere stata dissociata dai raggi ultravioletti e che la sua permanenza nell'atmosfera non può essere durata più di 20.000 anni. Un'ulteriore argomentazione fornita da Abelson a favore della propria tesi è che le più antiche rocce avrebbero dovuto contenere una notevole quantità di materia organica. Studi più recenti, basati sulla nostra comprensione dei fattori evolutivi relativi al pianeta Marte, come pure le ricerche sul problema dell'accrezione planetaria sembrano indicare che il passaggio dall'atmosfera riducente a quella ricca di diossido di carbonio fu assai rapido; perciò è possibile che la materia organica si sia formata molto precocemente nella storia della Terra, e che gradualmente, via via che l'atmosfera si modificava, si sia sintetizzata materia organica dalla miscela di monossido e diossido di carbonio.
Holland (v., 1962) aveva diviso la storia dell'atmosfera della Terra in tre stadi: il primo, relativo al periodo precedente la formazione del nucleo della Terra, è quello durante il quale il ferro era probabilmente presente nella parte superiore del mantello. I gas vulcanici eruttati durante questo stadio devono aver contenuto una grande quantità di idrogeno, e l'atmosfera era fortemente riducente. Questo periodo può essere durato circa mezzo miliardo di anni. Il secondo stadio, secondo Holland, iniziò quando il ferro scomparve dalla parte più superficiale del mantello. L'atmosfera divenne allora meno riducente e il diossido di carbonio prese il posto del metano. La presenza di uraninite detritica nel Precambriano medio suggerisce che questo secondo stadio sia durato almeno fino a due miliardi di anni fa. Il terzo stadio iniziò quando la velocità di produzione di ossigeno per fotosintesi superò la velocità necessaria per ossidare i gas vulcanici ridotti. Berkner e Marshall hanno sviluppato una convincente teoria per spiegare come l'accumulazione di ossigeno sia stata un risultato della fotosintesi (v. Berkner e Marshall, 1964); essi sostengono che l'improvvisa proliferazione della vita durante il Cambriano sia stata causata dal raggiungimento del punto di Pasteur, cioè circa l'1% dell'attuale pressione di ossigeno.
Come abbiamo visto prima, Oparin aveva postulato che per la sintesi organica fosse necessario che il carbonio si trovasse nella sua forma ridotta. La maggior parte degli organismi possiedono un meccanismo che li rende capaci di ridurre l'azoto prima di incorporarlo nelle molecole viventi, e in tutti, senza eccezioni, è riscontrabile un procedimento grazie al quale l'azoto è eliminato sotto forma di ammoniaca o urea. Ciò è forse il risultato dell'evoluzione di vie biochimiche che si sono sviluppate nella primitiva atmosfera priva di ossigeno. Fintantoché la cellula non ebbe raggiunto uno stato di estrema organizzazione e protezione non era possibile che l'ossigeno si trovasse libero nell'atmosfera, perché la sua presenza avrebbe rapidamente ossidato quegli elementi che costituirono i precursori della vita, impedendo completamente la loro evoluzione e finanche la loro esistenza. Anche l'effetto deleterio esercitato dall'ossigeno sul nucleo della cellula, quale era in quel periodo, è una prova che le condizioni presenti nel periodo in cui la cellula cominciò a evolversi erano prevalentemente condizioni in cui l'ossigeno era assente. La presenza di organismi anaerobi obbligati e l'universalità delle vie glicolitiche anaerobie suggeriscono che la vita si originò in ambienti non ossidati. Via via che l'ambiente si modificava passando da una condizione riducente a una in cui l'ossigeno era presente nella sua forma libera, può essere stato selettivamente favorito lo sviluppo di meccanismi per l'eliminazione dei perossidi come difesa contro questi letali composti.
d) Fonti di energia.
Nelle condizioni presenti sulla Terra primitiva, l'energia disponibile per la sintesi dei composti organici era quella derivante dai raggi ultravioletti solari, dalle scariche elettriche, dalla radiazione ionizzante e dal calore (v. Miller e Urey, 1959). È chiaro che la principale fonte di energia è la luce del Sole. La radiazione solare è emessa in tutte le regioni dello spettro elettromagnetico, e al presente essa viene in gran parte schermata dallo strato di ozono che si trova nelle regioni superiori dell'atmosfera. Dal momento in cui ha raggiunto la sua collocazione nella sequenza principale, il Sole è rimasto molto stabile: la temperatura della sua superficie 4,5 miliardi di anni fa era praticamente la stessa di oggi. Tenendo conto di questo dato, è stato calcolato il flusso di energia solare a differenti lunghezze d'onda, e i risultati ottenuti illustrano chiaramente la predominanza che deve aver avuto la luce ultravioletta tra le primitive fonti di energia. La luce ultravioletta dovrebbe esser considerata di gran lunga la più importante fonte di energia tra quelle usate negli esperimenti di simulazione effettuati in laboratorio; tuttavia, dal momento che l'impiego di raggi ultravioletti di breve lunghezza d'onda pone delle difficoltà intrinseche, gli esperimenti effettuati con questo tipo di energia sono stati pochi. Le bande di assorbimento del metano arrivano a 1.450 Å, quelle dell'acqua a 1.650, quelle dell'ammoniaca a 2.200. La resa quantica per la fotolisi dell'ammoniaca è circa 0,5, per quella dell'acqua 0,3, per quella del metano circa 0,5. La dissociazione di queste molecole avviene nella regione delle brevi lunghezze d'onda, ma i prodotti della dissociazione possono assorbire a lunghezze d'onda maggiori.
La seconda fonte di energia in ordine di importanza è rappresentata dalle scariche elettriche, come i lampi e l'effetto corona causato da oggetti appuntiti, che avvengono vicino alla superficie terrestre e pertanto potrebbero aver efficacemente trasportato i prodotti della reazione agli oceani primitivi. Poiché il lampo può facilmente essere simulato in laboratorio, è stato effettuato un gran numero di esperimenti usando questa forma di energia.
Sulla Terra le principali fonti radioattive di radiazioni ionizzanti sono il potassio-40, l'uranio-238, l'uranio-235 e il torio-232. Tra questi sembra che attualmente il potassio-40 abbia maggiore importanza da un punto di vista quantitativo, ed è pertanto probabile che la sua importanza sia stata ancora maggiore in passato. L'energia del potassio-40 è sotto forma di raggi β e γ penetranti; invece, quasi il 90% dell'energia derivante dall'uranio-235 e dal torio-232 si rivela come particelle α, che possono non avere capacità di penetrazione tali da produrre un effetto significativo. In base ai calcoli effettuati, è risultato che il decadimento del potassio-40 sulla superficie terrestre produce attualmente 3 × 1019 calorie all'anno, che sarebbero state 12 × 1019 due miliardi di anni fa. Ciò rappresenta solo 1/30 dell'energia resa disponibile dalla luce ultravioletta a breve lunghezza d'onda.
La distribuzione dell'energia ha una notevole rilevanza. La maggior parte delle radiazioni prodotte da sorgenti radioattive viene assorbita dai solidi, e poiché la crosta terrestre ha uno spessore di circa 30 km, questo tipo di radiazione potrebbe non aver avuto alcun ruolo nella sintesi di materiale organico negli oceani primitivi. Tuttavia, è possibile che in passato siano esistite delle limitate regioni ad alta attività; potrebbero essere esistiti dei microambienti in cui l'azione catalitica esercitata dai metalli presenti avrebbe favorito le reazioni di condensazione delle prime molecole sintetizzate grazie alle radiazioni ionizzanti.
Un'altra forma di energia che può essere stata efficace è il calore prodotto dai vulcani. È ragionevole supporre che l'attività vulcanica sia stata maggiore sulla Terra primitiva che non oggi. Il dato riportato nella tab. III è stato calcolato presumendo che sia stato emesso un metro cubo di lava all'anno a una temperatura di 1.000 °C. Inoltre, anche le sorgenti calde intorno a pozze di fango bollente avrebbero prodotto calore.
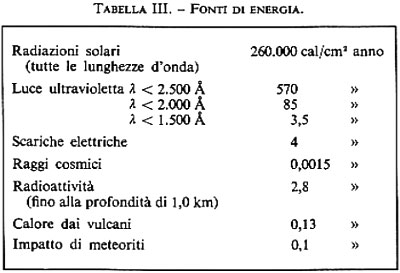
Anche la chemiosintesi dovuta all'impatto di meteoriti con l'atmosfera planetare è stata proposta come una delle possibili vie della primitiva sintesi di materiale organico (v. Hochstim, 1968). La reazione sarebbe il risultato dell'intenso calore generato momentaneamente in seguito all'onda d'urto prodotta dall'impatto.
4. Esperimenti effettuati in laboratorio.
Lo studioso di evoluzione chimica ha tentato di ricreare in laboratorio il ‟piccolo stagno caldo" di Darwin; in altre parole, la maggior parte delle varie fonti di energia che sono state considerate possibili è stata usata in laboratorio per vedere se la loro azione sull'atmosfera primitiva della Terra poteva determinare la formazione di molecole organiche.
a) Amminoacidi.
La maggior parte del lavoro effettuato nel campo della chimica prebiologica riguarda l'origine degli amminoacidi, probabilmente perché gli amminoacidi si formano con facilità. Un'altra ragione può risiedere nel fatto che solo in epoca relativamente recente l'attenzione si è focalizzata sulla chimica degli acidi nucleici; in precedenza, era la molecola proteica a occupare la posizione predominante in biochimica, e non deve perciò sorprendere che tutti gli sforzi fossero concentrati sulla ricerca dei suoi elementi costitutivi. Lo sviluppo delle tecniche cromatografiche ha reso possibile mettere in evidenza tracce anche minime di amminoacidi in miscele complesse.
È stato possibile ottenere la sintesi di amminoacidi, in condizioni che simulavano la Terra primitiva, mediante l'azione della luce ultravioletta, di scariche elettriche, calore e radiazioni ionizzanti (v. Lemmon, 1970). In alcuni casi, il materiale di partenza era rappresentato da composti intermedi capaci di reagire. E stato affermato che 14 dei 20 amminoacidi presenti nelle proteine sono stati ottenuti con sintesi non biologiche. In uno dei primi esperimenti di simulazione effettuati per riprodurre la sintesi di materia organica sulla Terra primitiva, Miller sottopose per circa una settimana una miscela di metano, ammoniaca, acqua e idrogeno a scariche elettriche, ottenute mediante l'impiego di un circuito di Tesla; si formarono diversi composti organici, tra cui gli amminoacidi glicina, alanina, β-alanina, acido aspartico e acido glutammico. Il 2,1% del carbonio introdotto inizialmente sotto forma di metano si era trasformato in glicina (v. Miller e Urey, 1959).
Abelson ha studiato l'effetto delle scariche elettriche su 20 diverse miscele di gas - idrogeno, metano, monossido di carbonio, diossido di carbonio, ammoniaca, azoto, acqua e ossigeno - e ha trovato che si erano formati parecchi amminoacidi, tra cui alanina, β-alanina, glicina e sarcosina. Gli amminoacidi si formavano esclusivamente quando le miscele non erano ossidanti. Questi esperimenti confermano indirettamente l'ipotesi che sulla Terra primitiva si producevano quantità significative di amminoacidi solo in condizioni non ossidanti. Più recentemente, Abelson ha studiato parecchie miscele diverse, contenenti monossido di carbonio, azoto e idrogeno, ed essendo riuscito in tutti i casi a identificare come prodotto intermedio il cianuro di idrogeno, ha suggerito che tale composto sia l'intermedio chiave nella sintesi di glicina, senna, alanina e acido aspartico (v. Abelson, Amino acids..., 1956).
Pavlovskaia e Pasynskii (v., 1960) hanno esposto a una scarica elettrica metano, ammoniaca, acqua e monossido di carbonio, e tra i prodotti così ottenuti sono riusciti a identificare glicina, alanina, β-alanina, acido α-amminobutirrico, acido glutammico, acido aspartico e lisina. Sembra che l'eliminazione dell'idrogeno dal sistema stimoli la sintesi degli amminoacidi, il che è comprensibile dato che i radicali metilici tendono a reagire con gli atomi di idrogeno riformando metano, mentre in un sistema in cui sia stato eliminato l'idrogeno viene favorita la reazione nella direzione che porta alla formazione di molecole più complesse. Sulla Terra primitiva l'idrogeno sfuggì nell'atmosfera, mentre in laboratorio viene mantenuto nell'apparecchio.
Reyns e altri (v., 1957) hanno lavorato con varie miscele di gas, riscontrando anch'essi che gli amminoacidi si formavano quando le condizioni erano non ossidanti. L'introduzione di solfuro di idrogeno dava luogo a tiocianato di ammonio, tiourea e tioacetammide, ma non ad amminoacidi contenenti zolfo. Negli esperimenti fatti usando metano, gli amminoacidi formatisi contenevano solo da due a quattro atomi di carbonio. Tuttavia, la sostituzione di parte del metano con etano permise a Oro di identificare prolina, valina e leucina, oltre a glicina, alanina, acido aspartico e asparagina, che erano già stati sintetizzati (v. Oro, Synthesis of.., 1963).
Grossenbacher e Knight (v., 1965) sottoposero a scarica elettrica per un periodo di tempo variabile da 12 a 48 ore un pallone da 12 litri contenente metano e ammoniaca, a una pressione di mezza atmosfera, su idrossido di ammonio; quindi analizzarono i prodotti che si erano così formati mediante un analizzatore di amminoacidi, trovando acido aspartico, treonina, senna, acido glutammico, glicina, alanina, isoleucina, leucina e lisina. Negli esperimenti eseguiti successivamente da Flores e Ponnamperuma (v., 1972), fu ottenuta la conversione di oltre il 95% del metano in composti organici esponendo per 24 ore una miscela di metano, ammoniaca e acqua a una scarica elettrica: il 18% del prodotto era rappresentato da cianuro di idrogeno. Molti furono gli amminoacidi identificati: glicina, alanina, treonina, acido aspartico, valina, acido glutammico, leucina, isoleucina e fenilalanina; l'identificazione fu effettuata dopo idrolisi del prodotto finale. Il tempo di ritenzione su una colonna a scambio ionico è stato confermato dall'analisi gascromatografica usando i derivati N-trifluoroacetilici degli amminoacidi.
Ponnamperuma e Woeller (v., 1967) hanno sottoposto delle miscele equimolari di metano e ammoniaca a una scarica elettrica ad arco. L'analisi per gascromatografia e spettrometria di massa dei prodotti volatili ha messo in evidenza la presenza di svariati nitrili alifatici: l'α-amminoacetonitrile, l'α-amminopropionitrile e i derivati C-metilico ed N-metilico dell'α-amminoacetonitrile.
Uno dei primi esperimenti fotochimici effettuati per determinare se l'azione della luce ultravioletta rendesse possibile la sintesi degli amminoacidi fu fatto da Abelson (v., Effects of.., 1956). Una soluzione di formiato d'ammonio in presenza di idrossido d'ammonio, cianuro di sodio e solfato ferroso venne irradiata a una lunghezza d'onda di 2.536 Å; si formò così glicinonitrile che, idrolizzato, dette luogo a glicina. Il 10% del formiato si convertì in glicina. Bahadur (v., 1954), dopo aver irradiato con luce visibile formaldeide e alcuni sali inorganici, quali nitrato di potassio e cloruro ferrico, riuscì a identificare acido aspartico, senna e lisina; questo esperimento, tuttavia, non può essere considerato molto significativo, in quanto l'uso del nitrato non è compatibile con le condizioni riducenti della Terra primitiva. Groth e von Weysenhoff (v., 1960) hanno esposto metano, etano, ammoniaca e acqua a radiazioni di 4.070 e 2.196 Å, riuscendo in tal modo a mettere in evidenza glicina, alanina e acido amminobutirrico. Al I Simposio internazionale sull'origine della vita, tenutosi a Mosca nel 1957, Terenin (v., 1959) riferì di aver riscontrato la presenza di alanina e di altri composti dopo aver irradiato ammoniaca, metano e acqua con radiazioni comprese nel la regione di Schumann.
Reid (v., 1960) usò radiazioni aventi lunghezza d'onda di 1.848 A su una miscela di formaldeide, ammoniaca e diossido di carbonio in presenza di alcuni sali inorganici, ottenendone fra l'altro i due amminoacidi glicina e alanina. Pavlovskaia e Pasynskii (v., 1960) sostennero di aver ottenuto senna, glicina, acido glutammico, alanina e valina irradiando formaldeide in presenza di nitrato d'ammonio e cloruro d'ammonio con una lampada a mercurio ad alta pressione. Più recentemente, Dodonova e Sidorova (v., 1961) usarono radiazioni comprese nella regione tra 1.450 e 1.800 Å su metano, ammoniaca, acqua e monossido di carbonio, riuscendo quindi a identificare diversi amminoacidi, tra cui glicina, alanina, valina e leucina.
Abelson aveva riferito che esponendo a una scarica elettrica una miscela contenente monossido di carbonio con azoto e idrogeno in proporzioni diverse aveva ottenuto cianuro di idrogeno. L'esposizione di una soluzione di cianuro di idrogeno a una lampada a mercurio dette luogo alla formazione di molti amminoacidi, tra cui glicina, senna, alanina e acido aspartico.
Riguardo al problema dell'origine della vita, sono stati compiuti numerosi studi per determinare l'effetto delle radiazioni ionizzanti su miscele primitive di gas (v. Ponnamperuma, 1967). Uno dei primi esperimenti fatti allo scopo di ricercare amminoacidi usando raggi X è stato quello di K. Dose e B. Rajewsky, i quali irradiarono una miscela di ammoniaca, acqua, metano, idrogeno, diossido di carbonio e azoto con raggi X e γ; si produssero in tal modo alcuni amminoacidi, messi in evidenza, come classe anche se non individualmente, dalla separazione elettroforetica.
Hasselstrom e altri ottennero glicina, acido aspartico e acido diamminosuccinico irradiando con particelle β una soluzione di acetato d'ammonio. Palm e M. Calvin esposero una miscela di metano, ammoniaca e acqua a elettroni di 5 MeV prodotti da un acceleratore lineare, e riuscirono a mettere in evidenza tra i prodotti finali alanina e glicina; usando un apparato identico a questo, e sostituendo in alcuni casi l'ammoniaca con l'azoto e il metano con etano, Oro ha riferito che si erano formati parecchi amminoacidi che non furono però identificati singolarmente. Dose e Ponnamperuma esponendo N-acetilglicina, usata come materiale di partenza, a raggi γ, riuscirono a identificare un'ampia gamma di amminoacidi.
In alcuni dei loro primi esperimenti S. W. Fox e altri riscaldarono fumarato e malato di ammonio e identificarono tra i prodotti finali della reazione acido aspartico e alanina. Successivamente, Fox e K. Harada usarono come sostanze di partenza glucosio, urea, acido α-idrossiglutanco e ammoniaca e, sotto l'azione del calore, ottennero glicina, acido glutammico, acido aspartico e alanina. In altri esperimenti, Harada e Fox hanno studiato l'effetto del calore su di una miscela di metano e ammoniaca: il metano è stato fatto gorgogliare in una soluzione acquosa concentrata di ammoniaca e fatto passare su sabbia di quarzo o allumina in un tubo di vetro riscaldato a 900 - 1.000 °C. I prodotti finali sono stati assorbiti in una soluzione acquosa di ammoniaca. I risultati ottenuti hanno portato Fox a sostenere che si potevano così sintetizzare 12 degli amminoacidi che si trovano normalmente nelle proteine: acido aspartico, treonina, senna, acido glutammico, prolina, glicina, alanina, valina, isoleucina, leucina, tirosina e fenilalanina (v. Rarada e Fox, 1964). Il rendimento in questi esperimenti era estremamente basso, a eccezione della glicina e dell'alanina.
Un esperimento simile è stato compiuto da J. Oro, che riuscì a identificare numerosi amminoacidi dopo aver fatto passare una miscela di metano, ammoniaca e acqua in una provetta riscaldata a circa 1.300 °K; tra gli amminoacidi ottenuti c'erano acido aspartico, treonina, senna, acido glutammico, glicina, alanina, alloisoleucina, isoleucina, leucina, tirosina, fenilalanina e β-alanina. Anche in questo esperimento gli amminoacidi prodotti in maggiore quantità furono glicina e alanina.
Lowe e altri studiarono l'azione del calore su di una miscela di cianuro d'idrogeno, idrossido d'ammonio e acqua, e identificarono tra i prodotti acido aspartico, treonina, serina, acido glutammico, glicina, alanina, isoleucina, leucina, β-alanina, acido α, β-diamminopropionico e acido α-amminobutirrico (v. Lowe e Rees, 1963).
In alcuni esperimenti compiuti da Oro e altri, che usarono una soluzione acquosa di formaldeide e di cloridrato di idrossilammina a temperatura ambiente in condizioni di leggera acidità, si formarono notevoli quantità di glicina e quantità minori di alanina, β-alanina, senna, treonina e acido aspartico. Facendo bollire a ricadere una soluzione acquosa di formaldeide e idrazina si formarono glicina, valina e lisina. In un altro esperimento, a temperatura ambiente, Oro e Kaurat ottennero la formazione spontanea di glicina, alanina e acido aspartico da cianuro di idrogeno, ammoniaca e acqua (v. Oro e Kaurat, 1961).
Nessuno di questi esperimenti ha mostrato che fosse possibile formare amminoacidi contenenti zolfo. Tuttavia, Choughuley e Lemmon (v., 1966) sono riusciti a mettere in evidenza acido cisteico, taurina e cisteina tra i prodotti finali ottenuti sottoponendo a irradiazione una miscela contenente metano, ammoniaca, solfuro d'idrogeno e acqua.
L'esser riusciti a identificare una così grande quantità di amminoacidi nel corso dei molti esperimenti che simulavano le condizioni primitive potrebbe far credere che il problema della loro origine sia ormai risolto, ma bisogna invece tener presente che molte di queste identificazioni necessitano ancora di analisi precise. Contrariamente a quanto accade quando si analizza un idrolisato di proteine di origine biologica, negli esperimenti di sintesi abiologica si forma un gran numero di composti molto simili gli uni agli altri. Per l'identificazione di un composto può non esser sufficiente la posizione che occupa su una carta cromatografica, o il tempo di ritenzione, o il tempo di eluizione su una colonna a scambio ionico; in alcuni casi si possono formare amminoacidi che non si trovano in natura e che possono avere gli stessi valori di Rf di quelli naturali; è quindi necessario eseguire altre analisi per chiarire la reale natura e struttura di questi composti. Tranne che in un caso in cui sia la cromatografia a scambio ionico, sia la gascromatografia e la spettrografia di massa hanno portato alla stessa identificazione, nella maggioranza degli altri casi l'identificazione si è basata o sulla sola cromatografia su carta o sulla sola cromatografia a scambio ionico.
Si sa poco sui meccanismi che intervengono in queste sintesi. Se si escludono gli esperimenti di Miller, per i quali è stata prospettata la possibilità di una sintesi di Strecker, non è stato fatto alcun tentativo per chiarire come si siano originati gli amminoacidi. Miller ha avanzato due ipotesi alternative: 1) dapprima si sono sintetizzati nella fase gassosa, per mezzo di una scarica elettrica, le aldeidi e il cianuro di idrogeno; queste aldeidi e il cianuro di idrogeno hanno poi reagito nella fase acquosa del sistema dando origine ad amminoe idrossinitrili che, a loro volta, si sono idrolizzati ad amminoe idrossiacidi; 2) gli ammino- e gli idrossiacidi si sono formati nella fase gassosa dagli ioni e dai radicali prodotti durante la scarica elettrica. Le ricerche di Miller hanno mostrato che la maggioranza degli amminoacidi si è formata secondo la maniera suggerita dalla prima ipotesi, dal momento che la velocità di produzione delle aldeidi e del cianuro d'idrogeno per scarica elettrica e la velocità di idrolisi degli amminonitrili erano sufficienti a spiegare la produzione totale di amminoacidi. Gli esperimenti di Miller sui meccanismi in gioco nella sintesi di amminoacidi dovuta a scarica elettrica indicano che l'insieme delle condizioni necessarie per la sintesi di tali composti non è molto particolare. Il processo o l'insieme di processi che hanno portato alla formazione sia dell'aldeide che del cianuro di idrogeno possono aver contribuito a formare la quantità di α-amminoacidi presente negli oceani della Terra primitiva secondo il meccanismo della sintesi di Strecker (v. Miller, 1957):
RCHO + NH3 + HCN ⇄ RCH(NH2)CN + H20
RCH(NH2)CN + 2H2O → RCH(NH2)COOH + NH3
RCHO + HCN ⇄ RCH(OH)CN
RCH(OH)CN + 2H2O → RCH(OH)COOH + NH3.
Gli esperimenti effettuati con metano e ammoniaca in assenza di acqua per formare nitrili hanno mostrato che può essere stato operante anche un meccanismo alternativo (v. Ponnamperuma e Woeller, 1967). Pertanto, se la reazione tra metano e ammoniaca dà luogo preferenzialmente alla sintesi di α-amminonitrili, e se questi nitrili reagiscono con l'acqua, è possibile che avvenga un processo di idrolisi dando origine ad amminoacidi. In questo caso gli amminoacidi si formano in assenza di aldeidi. Sanchez e altri (v., 1966) hanno avanzato l'ipotesi che la via sintetica che dà origine all'acido aspartico passi attraverso cianoacetilene.
b) Purine e pirimidine.
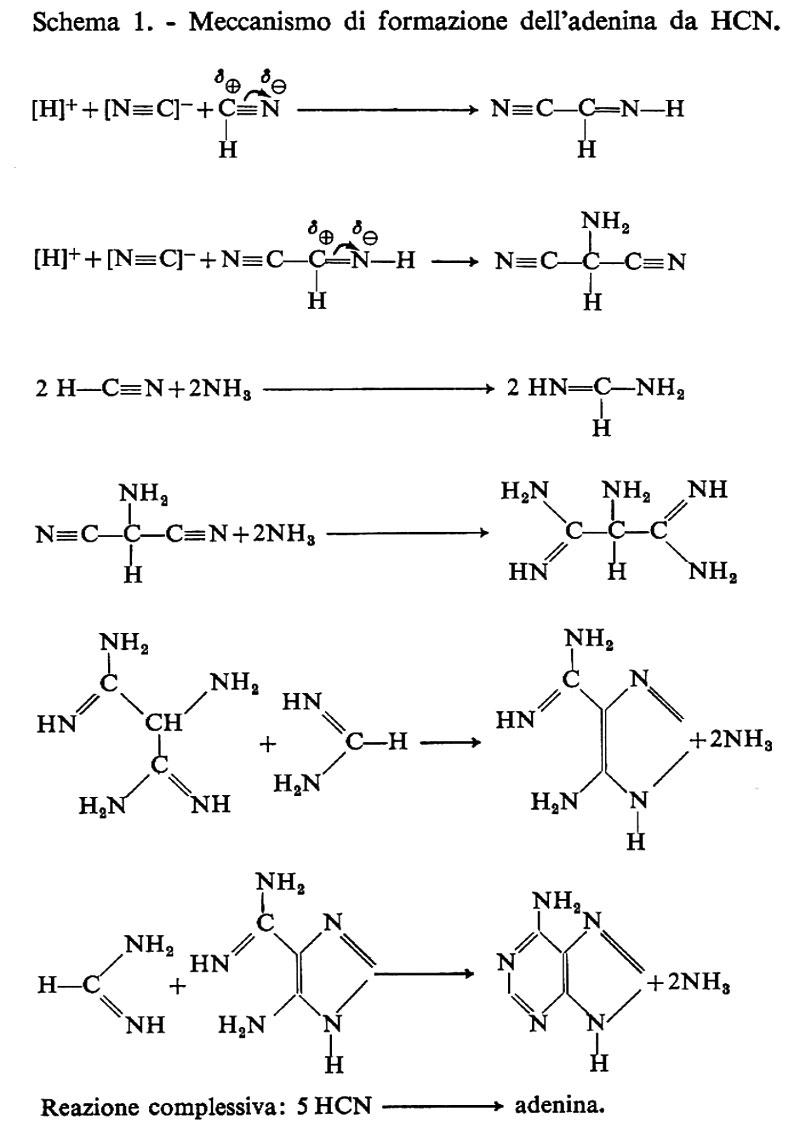
Le purine sono state sintetizzate per la prima volta in condizioni che simulavano quelle della Terra primitiva da Oro (v., Studies in..., 1963), il quale dimostrò che si poteva ottenere la sintesi di adenina da una soluzione concentrata di cianuro d'ammonio. Nelle sue linee essenziali la reazione può essere raffigurata come 5 molecole di cianuro di idrogeno che, in presenza di ammoniaca, danno luogo ad adenina. Questa sintesi è stata confermata da Lowe e collaboratori. Il particolareggiato studio di Oro ha mostrato che i probabili prodotti intermedi di questa reazione sono 4-amminoimidazolo-5-carbossimmide e formammidina (v. schema 1). In seguito, Oro riuscì a sintetizzare guanina e xantina portando a una temperatura di 100-140 °C una soluzione acquosa di amminoimidazolocarbossimmide. Il rendimento è stato dell'1,5% per ambedue le purine. È possibile che questo sia uno dei modi in cui è avvenuta la sintesi delle purine sulla Terra primitiva, ma le concentrazioni usate da Oro erano di gran lunga troppo alte per corrispondere a una situazione prebiotica. Se le condizioni sperimentali fossero state davvero simili a quelle prebiotiche, se, per esempio, si fossero usate concentrazioni più basse, allora queste reazioni sarebbero di grande aiuto alla comprensione dell'origine delle purine nelle condizioni presenti nella fase prebiotica della Terra.
Ponnamperuma e altri (v., Formation of.., 1963) hanno ottenuto anche la sintesi di adenina mediante irraggiamento con elettroni di metano, ammoniaca e acqua. Poiché in queste reazioni è stato possibile individuare la presenza di cianuro di idrogeno, è possibile che la via sintetica sia la stessa, che si tratti cioè di una reazione del cianuro di idrogeno catalizzata dalla base. Nell'irraggiamento di metano, ammoniaca e acqua con elettroni si ottenne un rendimento dello 0,01% del metano iniziale, il che non è poco e appare realistico nelle condizioni della Terra primitiva. È stata fatta l'interessante osservazione che la produzione di adenina era aumentata in assenza di idrogeno, il che non è sorprendente dato che il carbonio del metano deve essere ossidato per poter poi comparire nelle purine. In ogni caso, la concentrazione di materia organica divenne alta sulla Terra prebiotica probabilmente quando la maggior parte dell'idrogeno era sfuggito dalla sua atmosfera. Nonostante gli sforzi fatti, non è stato possibile identificare con sicurezza purine e pirimidine tra i prodotti finali in esperimenti che impiegavano scariche elettriche. Poiché si forma una gran quantità di cianuro di idrogeno, è difficile capire come le purine potrebbero essere assenti.
In altri esperimenti è stato mostrato come l'esposizione a luce ultravioletta di una soluzione diluita di cianuro di idrogeno desse luogo alla formazione sia di adenina che di guanina. Le quantità prodotte erano scarse, ma la quantità di adenina era maggiore di quella di guanina. La produzione di urea era abbondante (v. Ponnamperuma, 1965).
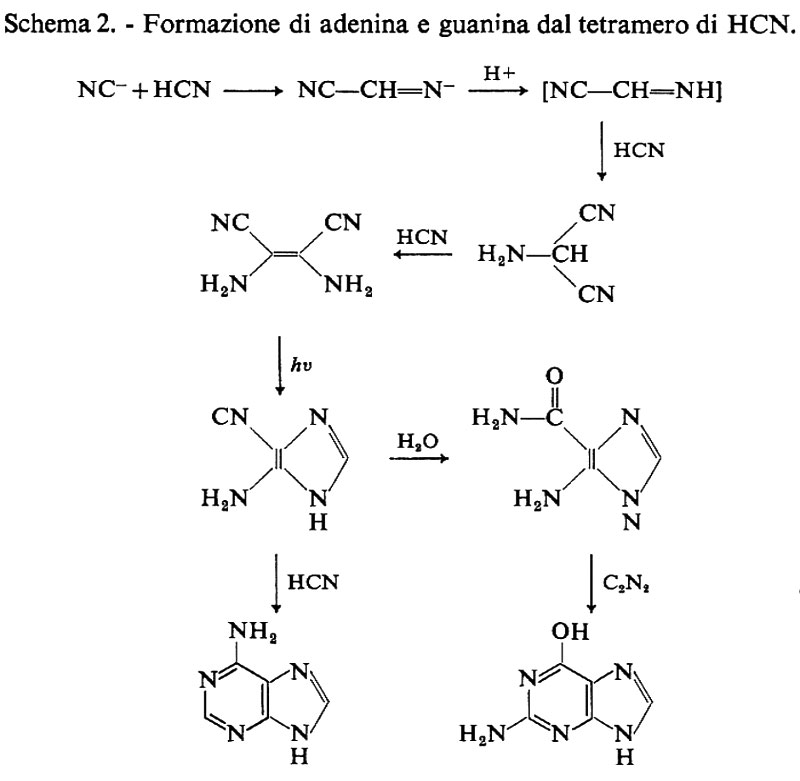
Le ricerche di Sanchez e altri (v., 1968) hanno mostrato che l'amminomalononitrile è un importante prodotto intermedio nella sintesi dell'adenina. Il monoamminomalononitrile si trasforma in 4-amminoimidazolo-5-carbossimmide o per reazione diretta con formammidina, o, cosa ancor più interessante per i nostri studi, per trasposizione fotochimica del tetramero del cianuro di idrogeno. Per idrolisi questo dà luogo a 4-amminoimidazolo-5-carbossimmide, che in condizioni più blande può, mediante una reazione di condensazione in soluzione acquosa con cianuro, formammidina o cianogeno, formare svariate purine, quali adenina, ipoxantina, diamminopurina e guanina (v. schema 2).
Ben poco è stato fatto nel campo della sintesi delle pirimidine. Fox e Harada (v., 1961) hanno dimostrato che uracile può essere ottenuto per riscaldamento di acido malico e urea. Mentre l'urea si forma facilmente negli esperimenti che simulano la Terra primitiva, non esiste alcuna indicazione della presenza di acido malico. Sanchez e altri hanno mostrato che uno dei principali prodotti azotati che si ottengono per azione di una scarica elettrica su di una miscela di metano e azoto è il cianoacetilene. Questo può essere il prodotto di partenza per la sintesi di svariate pirimidine. Attraverso la fusione di cianoacetilene con urea è stata ottenuta la citosina; in modo simile è stato preparato l'uracile dalla propiolammide (v. Sanchez e altri, 1966).
Riscaldando a 100 °C per un giorno una soluzione acquosa contenente cianato di potassio 1,0 M e cianoacetilene 1,0 M, è stata ottenuta citosina con un rendimento del 5%. È molto significativo il fatto che, lasciando la stessa soluzione a temperatura ambiente per sette giorni, fu ottenuta citosina con un rendimento dell'1%.
Oro aveva proposto la teoria che la formazione di pirimidine potesse aver luogo partendo da specie molecolari a 3 atomi di carbonio che si ritrovano nelle comete. Uno di questi derivati è la malonammidesemialdimmina, o il suo isomero β-amminoacrilimmide, che per condensazione con urea potrebbe formare uracile. Inoltre, questo autore riteneva che l'acrilonitrile, il β-amminopropionitrile e la fiamminopropionimmide fossero probabili prodotti intermedi, in quanto, facendo reagire ognuno di questi composti con urea in soluzioni acquose contenenti ammoniaca, egli aveva osservato costantemente la formazione di piccole quantità di uracile. In teoria, le tre pirimidine presenti negli acidi nucleici avrebbero potuto formarsi in sistemi acquosi, nelle condizioni esistenti sulla Terra primitiva, col meccanismo sopra descritto. Anche il sottossido di carbonio C3O2, che si ritrova nei corpi cosmici, potrebbe essere un adatto precursore nella sintesi delle pirimidine (v. Oro, Studies in..., 1963).
c) Monosaccaridi.
In contrasto con i progressi compiuti nel campo della chimica primordiale degli amminoacidi, l'origine di un altro gruppo di sostanze altrettanto importanti biologicamente, i monosaccaridi, è tuttora assai incerta.
Già nel 1861 A. Butlerow aveva mostrato che formaldeide sciolta in acqua va incontro a un processo di condensazione, in ambiente alcalino, dando luogo a una miscela di zuccheri (v. Butlerow, 1861). Poco dopo O. Loew riusci a isolare da questa miscela di condensazione un gruppo di esosi otticamente inattivi. In questa condensazione della formaldeide indotta dall'ambiente alcalino si formano, oltre a esosi, anche altri monosaccaridi di peso molecolare minore, quali la glicolaldeide (v. Butlerow, 1861), che pare sia il prodotto di condensazione di due molecole di formaldeide:
2CH2O → CH2OH−CHO
CH2OH−CHO + CH2O → CH2OH−CO−CH2OH
CH2OH−CHO + CH2OH−CO−CH2OH →
→ CH2OH−CHOH−CHOH−CO−CH2OH
2CH2OH−CHO → CH2OH−CHOH−CHOH−CHO
CH2OH−CO−CHOH−CH2OH
oppure
2C4 → [C8]? → C3 + C5.
È stata dimostrata l'esistenza di altri prodotti di condensazione, quali la gliceraldeide, il dudrossiacetone, l'entrosio e vari pentosi.
Gli esperimenti di Loew hanno mostrato che la condensazione alcalina della formaldeide procede nelle condizioni migliori quando sono presenti idrossidi alcalino-terrosi e gli idrossidi, debolmente basici, di piombo e stagno. La reazione di condensazione è caratterizzata anche da quello che è stato chiamato un ‛periodo di latenza', che peraltro può essere eliminato con l'aggiunta di glicolaldeide, diidrossiacetone o zuccheri più complessi. Inoltre, è stato osservato che la condensazione della formaldeide per dare una miscela di zuccheri non ha luogo in presenza di idrossido di tetrametilammonio, che è una base forte, a meno che non venga introdotto anche un sale di un metallo alcalino-terroso.
Dunque, la formazione di glicolaldeide per condensazione di due molecole di formaldeide pare essere un processo lento. Però, una volta che questo primo zucchero si è formato, si producono rapidamente zuccheri più complessi attraverso reazioni reversibili di condensazione aldolica e di enolizzazione. Dato che è noto che le condensazioni aldoliche sono catalizzate da basi e non richiedono la presenza di alcun catione, l'azione catalitica dei cationi dei metalli divalenti riguarda probabilmente la formazione di glicolaldeide.
L'ipotesi avanzata da S. L. Miller che, negli esperimenti in cui per azione di scariche elettriche si erano formati amminoacidi, questi potessero essersi originati secondo la sintesi di Strecker, richiamò l'attenzione sulla possibilità che la formaldeide potesse essere stata un precursore degli zuccheri ai tempi della chimica prebiologica. In effetti, la formaldeide è stata ritrovata tra i prodotti ottenuti sottoponendo miscele di metano, ammoniaca e acqua a scarica elettrica o irradiazione β o irradiazione γ. Nonostante che non siano ancora stati identificati zuccheri negli esperimenti in cui sono stati utilizzati metano, ammoniaca e acqua come sostanze di partenza, trattando la formaldeide con raggi ultravioletti si possono ottenere ribosio e desossiribosio, come pure altri zuccheri (v. Ponnamperuma, 1965). In uno studio sulla formazione di 2-desossiribosio da gliceraldeide e acetaldeide o da formaldeide e acetaldeide, Oro e Cox hanno trovato che il 2-desossiribosio e il suo isomero 2-desossixilosio si erano formati, con un rendimento di circa il 5%, da gliceraldeide e acetaldeide in sistemi acquosi. La reazione è catalizzata da ossidi di metalli divalenti o da ammoniaca. Il 2-desossiribosio fu identificato come prodotto di reazione anche quando la soluzione acquosa di partenza conteneva formaldeide, acetaldeide e idrossido di calcio.
Ci sono peraltro varie difficoltà nell'accettare l'ipotesi che la formaldeide possa essere stata utilizzata come precursore dei monosaccaridi. Horowitz e Miller hanno sottolineato che le elevate concentrazioni di formaldeide usate in alcuni esperimenti non riproducono in modo realistico le condizioni della Terra primitiva. Sono state sollevate obiezioni anche all'uso di soluzioni molto basiche. P. H. Abelson sostiene che la concentrazione di ammoniaca libera nei mari e nell'atmosfera non fu mai molto elevata, e che un oceano fortemente alcalino non è mai esistito. Inoltre, la formaldeide polimerizza assai rapidamente dando paraformaldeide, e può pertanto allontanarsi dall'ambiente.
Per tentare di superare queste obiezioni, Gabel e Ponnamperuma (v., 1967) hanno utilizzato come ambiente di reazione una sorgente idrotermale simulata. Soluzioni acquose di formaldeide a varie concentrazioni sono state fatte bollire a ricadere su caolinite e i prodotti cosi ottenuti sono stati separati in triosi, tetrosi, pentosi ed esosi. È stato osservato che a una concentrazione di formaldeide 0,5 M i prodotti della reazione comprendevano triosi, tetrosi e pentosi, mentre a una concentrazione 0,01 M si formavano anche esosi. Tra i prodotti finali è stato identificato il ribosio.
L'esistenza di sorgenti idrotermali sulla Terra primitiva, anche se limitata probabilmente a microambienti, eliminerebbe la necessità di postulare un oceano primitivo alcalino e avrebbe potuto fornire un'occasione per la demolizione della paraformaldeide. Resta comunque da analizzare con maggiore attenzione un'ulteriore via per la sintesi degli zuccheri, quella della sintesi fotochimica.
d) Nucleosidi e nucleotidi.
Il fatto di avere riprodotto in modo soddisfacente la sintesi delle basi e degli zuccheri degli acidi nucleici in condizioni simulanti quelle della Terra primitiva ha spinto alcuni ricercatori a compiere indagini sulla formazione abiogena di nucleosidi e nucleotidi.
Esponendo a luce ultravioletta soluzioni acquose diluite (10-3 M) di ribosio, adenina e fosfato, è stata ottenuta la formazione di adenosina (v. Ponnamperuma e altri, Synthesis of.., 1963) attraverso una reazione che pare sia catalizzata dal fosfato. Il meccanismo di questa reazione non è stato studiato, ma sembrerebbe che si formi uno zucchero-1-fosfato e che il fosfato sia poi sostituito dalla base purinica.
È stato suggerito che la primordiale atmosfera riducente della Terra fosse almeno un po' trasparente tra 2.400 Å e 2.900 Å, e che la fotoattivazione delle purine e pirimidine assorbenti luce ultravioletta abbia potuto rappresentare un passo nella formazione dei nucleosidi e nucleotidi (v. Sagan, 1957). Per verificare questa ipotesi sono stati compiuti quattro esperimenti utilizzando come sostanze di partenza rispettivamente adenina, adenosina, adenosinmonofosfato e adenosindifosfato (v. Ponnamperuma e altri, Synthesis of.., 1963). Fu così accertata la trasformazione di adenina in adenosina, di adenosina in adenosinmonofosfato, del monofosfato in difosfato e di quest'ultimo in trifosfato. L'adenosina si produceva in quantità misurabili solo in presenza di un composto fosforato. Mentre l'adenosina si formava in presenza sia di acido fosforico che di etilmetafosfato, il nucleoside fosfato è stato ottenuto solo in presenza di etilmetafosfato (v. Schramm, 1965). Nonostante sia improbabile che l'etilmetafosfato abbia potuto rappresentare una fonte di fosforo sulla Terra primitiva, il risultato di questo esperimento indica chiaramente che la sintesi di nucleotidi può avvenire abiologicamente, ed è comunque possibile che un altro composto fosforico più abbondante sia stato utilizzato in luogo dell'etilmetafosfato.
Miller e Parris hanno compiuto delle indagini con apatite, che è di gran lunga la forma di fosforo più abbondante sulla Terra. Usando idrossiapatite e cianati, questi autori hanno ottenuto pirofosfato. Dato che l'apatite e il pirofosfato di calcio sono entrambi insolubili, questa sintesi deve per forza avvenire sulla superficie dell'apatite. Il rendimento massimo di pirofosfato è stato ottenuto a pH 6,5, con una conversione del 27% del cianato aggiunto al sistema. W. F. Neuman e M. Neuman hanno anche sottolineato il possibile ruolo dell'apatite nelle reazioni di condensazione che portano alla formazione di nucleosidi e nucleotidi.
Per quello che riguarda la sintesi di nucleotidi, sono state esaminate anche le reazioni di varia natura che possono aver avuto luogo sul letto di un oceano prosciugato. Per simulare queste condizioni, una miscela intima di nucleosidi è stata riscaldata in presenza di un fosfato inorganico. Per questa reazione sono stati usati vari fosfati, tra cui il disodico, il trisodico, il monosodico-monoammonico, il monoammonico, il diammonico, il monocalcico e il tricalcico, nonché l'acido fosforico (v. Ponnamperuma e Mack, 1965). Riscaldando la miscela intima di nucleoside e fosfato, aveva luogo la fosforilazione: tra i prodotti finali sono stati identificati i mononucleotidi corrispondenti. Il rendimento più elevato è stato ottenuto a 160 °C, ma la reazione procedeva, sia pur con bassa efficienza, anche a temperature intorno a 50 °C. Negli esperimenti sopra descritti, l'ambiente simulato era assai povero di acqua, la cui presenza, peraltro, non è incompatibile con la reazione, a meno che essa non sia presente in grande eccesso. Le condizioni in cui questa reazione procede possono dunque essere considerate ipoidriche. Tra i monofosfati formatisi c'erano uguali quantità di monofosfati 2′, 3′, 5′ e ciclici. Non è da escludere che la presenza di certi amminoacidi possa dirigere la sintesi preferenzialmente verso la formazione di un particolare isomero.
Waenheldt e Fox hanno ottenuto la fosforilazione dei nucleosidi adenosina, citidina, guanosina, uridina, desossicitidina e timidina con acido polifosforico a temperature varianti fra 0 e 22 °C, con un rendimento variabile dal 25 al 45%. In queste condizioni la desossiadenosina e la desossiguanidina non vengono fosforilate. Il rendimento in questo tipo di esperimenti è eccezionalmente elevato, ma il fatto che sia stato utilizzato acido polifosforico concentrato non consente di ritenere che siano state simulate le autentiche condizioni di reazioni prebiologiche.
e) Idrocarburi e acidi grassi.
Benché siano stati prodotti idrocarburi in notevole quantità per azione di varie forme di energia sul metano, gli studi dettagliati compiuti in questo campo sono stati pochi. Risultati assai interessanti sono stati ottenuti in esperimenti effettuati per confrontare l'effetto di vari tipi di scarica sul metano (v. Ponnamperuma e Woeller, 1964). Con una scarica ad arco di grande intensità, il prodotto era un fluido chiaro, giallo, che dava cromatogrammi con picchi ben separati, mentre la scarica a semicorona dava luogo a un distillato incolore contenente sostanze mal separabili mediante gascromatografia. Con la scarica ad arco il prodotto più abbondante era il benzene, seguito immediatamente dal toluene, sostanze che erano virtualmente assenti con la scarica a semicorona. Mediante spettrometria di massa alcuni dei picchi cromatografici più importanti sono stati identificati come 2,2-dimetilbutano, 2-metilpentano, 3-metilpentano, 2,4-dimetilesano e 3,4-dimetilesano.
Quando sono stati esaminati i composti al di sopra di C9 è stato visto che, con la scarica a semicorona, la frazione alifatica era costituita da una miscela di composti non separabili, con una lunghezza massima intorno a C17. Le analisi di spettrometria di massa e di risonanza magnetica nucleare indicano che si tratta di derivati cicloesilici. Con l'uso di setacci molecolari si è dimostrato che in questo sistema non si formavano molecole normali o di tipo isoprenoide. È interessante notare che sono stati ritrovati in alcuni depositi idrocarburi simili a quelli ottenuti con scariche elettriche (v. Ponnamperuma e Pering, 1966). Un metodo per ottenere idrocarburi e acidi grassi a catena lineare è stato suggerito da Wilson, e comporta un ammassamento delle catene reattive degli acidi grassi su una superficie, in modo che solo le parti terminali siano disponibili per la reazione e si impedisca così la ramificazione.
Esponendo una miscela di metano e acqua a una scarica a semicorona e analizzando i prodotti mediante gascromatografia e spettrometria di massa, sono stati rinvenuti vari acidi monocarbossilici da C2 a C12; quelli al di sopra di C6 erano caratterizzati da catene di carbonio ramificate (v. Allen e Ponnamperuma, 1967). Un'area di notevole interesse nello studio dell'evoluzione chimica è rappresentata dalla formazione di acidi grassi normali contenenti 12 o più atomi di carbonio, che sono gli acidi grassi che prendono parte alla costituzione dei lipidi. Benché le condizioni di reazione usate sottoponendo a scariche a semicorona metano e acqua dovrebbero aver favorito la produzione di catene ramificate a causa della stabilità dei radicali di carbonio secondari, è sorprendente che si siano formati solamente lì dei possibili isomeri da C1 a C12; in contrasto con questo comportamento, sottoponendo a scariche a semicorona il solo metano, si ottengono miscele di idrocarburi ad alto contenuto di isomeri. Questi esperimenti indicano che certi acidi da C8 a C12 non eccessivamente ramificati potrebbero essersi accumulati per azione di scariche a semicorona passanti attraverso un'atmosfera di metano su una superficie acquosa.
f) Porfirine.
È opinione comune che gli organismi non avrebbero potuto sopravvivere al periodo di transizione dalla primitiva atmosfera riducente all'attuale atmosfera ossidante senza le porfirine (v. Gaffron, 1962). È probabile che i pirroli siano stati sintetizzati abiologicamente in uno stadio precoce nel corso dell'evoluzione chimica. A. E. Chichibabin ha dimostrato che facendo passare acetilene e ammoniaca attraverso un tubo riscaldato si formano pirroli. In precedenza V. Meyer aveva provato che i pirroli si possono ottenere da acetilene e cianuro di idrogeno. A. Szutka e collaboratori hanno fatto diversi tentativi di sintetizzare sostanze porfirino-simili in condizioni presumibilmente corrispondenti a quelle della Terra primitiva (v. Szutka, 1965). In alcuni esperimenti si è riscaldata benzaldeide insieme a pirroli per produrre materiale porfirino-simile; in altri si è riusciti a ottenere il derivato tetrafenilico della porfirina sottoponendo a raggi γ una miscela di benzaldeide e pirroli. Risultati simili sono stati ottenuti usando scariche elettriche. In esperimenti più recenti si è ottenuta la formazione di composti pirrolici per condensazione dell'acido δ-amminolevulinico.
Nel tentativo di ricreare in laboratorio le condizioni in cui potrebbero essersi formate le sostanze necessarie per la vita, l'obiettivo primario dello studioso di evoluzione chimica è stato quello di vedere se i componenti degli acidi nucleici e delle proteine avrebbero potuto formarsi nelle condizioni primordiali. Questa meta è stata in certo senso raggiunta, sia in modo diretto che per tappe successive. L'ipotesi più interessante sembra quella di un'atmosfera primitiva dalla quale avrebbero potuto generarsi cianuro di idrogeno e formaldeide; infatti il cianuro di idrogeno, o suoi sostituti, possono rappresentare la via sintetica per le basi, la formaldeide quella per gli zuccheri, mentre un'aldeide o il corrispondente nitrile potrebbero, attraverso la sintesi di Strecker, dare origine agli amminoacidi. Si tratta di uno schema certamente troppo semplificato, tuttavia esso raccoglie molti consensi per il fatto che i radioastronomi hanno riscontrato la presenza di formaldeide e cianuro di idrogeno nel mezzo interstellare.
g) Sintesi delle grandi molecole.
Per formare un dipeptide dall'unione di due amminoacidi, o un dinucleotide da due nucleotidi, è necessario un processo di disidratazione-condensazione. La disidratazione potrebbe essere stata il risultato dell'azione del calore, che potrebbe aver determinato l'evaporazione della soluzione primordiale di molecole organiche. Nella già citata conferenza tenuta alla British Physical Society e intitolata The physical basis of life, Bernal aveva avanzato l'ipotesi che i componenti organici contenuti nell'oceano potrebbero essere stati depositati sulla riva e adsorbiti dall'argilla, che avrebbe reso in tal modo possibile un processo di condensazione. L'ipotesi di Bernal è stata verificata in laboratorio e i risultati sono stati incoraggianti. Tuttavia dovrebbe essere analizzata anche la possibilità che questa reazione abbia avuto luogo nell'acqua. Poiché il nostro è un pianeta molto ricco di acqua, se fosse dimostrato che tali reazioni possono in realtà avvenire anche in acqua, si amplierebbero molto le circostanze favorevoli alla formazione di polimeri in condizioni prebiotiche. In effetti, negli organismi viventi tali reazioni hanno luogo, e per superare le barriere energetiche vengono usati gli enzimi. Ciò che sarebbe necessario nel sistema prebiotico sono i precursori di questi enzimi, o catalizzatori primitivi. Sono stati effettuati numerosi esperimenti allo scopo di saggiare entrambe le ipotesi menzionate e i risultati ottenuti sono stati oltremodo incoraggianti.
h) Polipeptidi.
Le sintesi simulate di polipeptidi primordiali sono state oggetto di maggiore attenzione rispetto alle sintesi di altri polimeri biologici, probabilmente per la grande importanza attribuita all'origine primordiale degli amminoacidi. Fox da molto tempo sostiene l'ipotesi dell'origine termica, anidra o ipoidrica, di tutti i composti organici primordiali (v. Fox e Dose, 1972). Fox e Harada hanno dimostrato che in presenza di una quantità relativamente elevata di acido glutammico o di acido aspartico si può polimerizzare mediante riscaldamento, a una temperatura di 180-200 °C, una miscela omogenea di tutti i diciotto amminoacidi normalmente presenti nelle proteine. A questi polimeri è stato dato il nome di proteinoidi; il loro peso molecolare aumentava da 3.600, in un proteinoide ottenuto a 160 °C, a 8.600 in uno ottenuto a 190 °C. Il più alto peso molecolare raggiunto è stato di 80.000. I polimeri danno la reazione del biureto positiva, possono essere nuovamente idrolizzati ad amminoacidi e vengono attaccati da svariati enzimi proteolitici.
È stato sostenuto che la quantità di energia resa disponibile dall'attività vulcanica è molto scarsa in confronto a quella resa disponibile da altre fonti energetiche. Tuttavia, non si può negare che in condizioni simulanti l'attività vulcanica avverrebbero reazioni organiche, inclusa la polimerizzazione di amminoacidi con formazione di legami peptidici. Un evidente punto debole insito in questo tipo di sequenza di reazioni per l'origine dei polipeptidi è la necessità di postulare la presenza di miscele omogenee, ad alta concentrazione, degli amminoacidi che si devono polimerizzare.
Oro e Guidry hanno rilevato un dato importante, e cioè che la glicinammide o la glicina possono essere polimerizzate termicamente in ammoniaca concentrata. Anche se i loro esperimenti dimostrano che non è necessaria una condizione anidra per la polimerizzazione degli amminoacidi mediante riscaldamento, tuttavia è dubbio che sulla Terra primitiva siano mai esistite concentrazioni così alte di ammoniaca. Questi esperimenti, però, potrebbero spiegare l'evoluzione chimica nei pianeti più grandi del sistema solare (v. Oro e Guidry, 1961).
È stato possibile ottenere la sintesi di peptidi direttamente da soluzioni diluite di amminoacidi, usando cianammide o diciandiammide come agenti condensanti. È noto che ambedue i composti si sono ottenuti mediante irradiazione ultravioletta di soluzioni acquose di cianuro di idrogeno o irradiando una miscela di metano, ammoniaca e acqua (v. Schimpl e altri, 1965). Ponnamperuma e Peterson hanno riportato che sottoponendo a irradiazione ultravioletta una soluzione acquosa diluita di glicina e leucina in presenza di cianammide a pH 5 si formavano glicilleucina e leucilglicina. Dal punto di vista del meccanismo di reazione, probabilmente la radiazione ultravioletta determinava un aumento della concentrazione di carbodummide, un tautomero della cianammide che può agire da agente condensante per gli amminoacidi:
H2N−CN→HṄ−C≡N ↔ HN=C=Ṅ→HN=C=NH.
L'esposizione di acidi carbossilici a radiazioni nel lontano ultravioletto, come è noto, promuove la scissione omolitica dei legami C−H sugli atomi di carbonio a-metilenici, seguita da dimerizzazione dei radicali risultanti ad acidi succinici sostituiti. L'irradiazione di ammoniaca produce radicali NH e NH2. Dal momento che nessuno di questi processi sembra fosse operante, e che i prodotti risultanti erano dipeptidi e tripeptidi, parrebbe che la luce ultravioletta non abbia agito direttamente sugli amminoacidi.
Le carbodiimmidi reagiscono con i gruppi carbossilici dando luogo a un addotto con un ottimo gruppo uscente sul carbonio carbossilico. Dato che le ammine sono assai più nucleofile dell'acqua, il gruppo amminico di un secondo amminoacido è a questo punto in grado di attaccare l'atomo di carbonio del summenzionato addotto formando un composto intermedio tetraedrico. La scissione del composto intermedio, che è instabile, produrrebbe allora il dipeptide e urea.
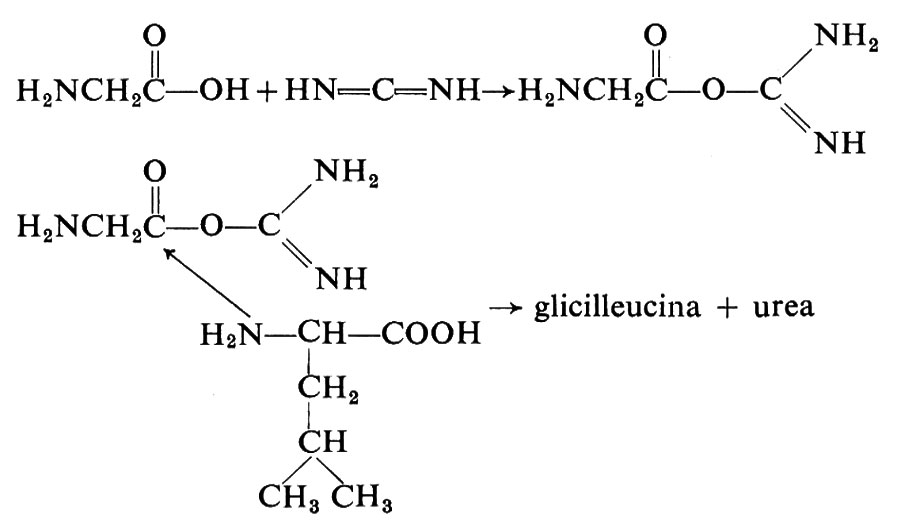
Steinman e collaboratori hanno riferito che in un esperimento analogo si formarono dipeptidi e tripeptidi dell'alanina quando una soluzione diluita dell'amminoacido era trattata con dicianammide (v. Steinman, 1967). Questa reazione, pur avvenendo preferenzialmente in ambiente acido, dà luogo anche a piccole quantità di peptidi a pH neutro. Questi autori hanno suggerito la possibilità che si formi un prodotto intermedio simile a quello descritto nell'esperimento precedente.
Rabinowitz (v., 1971) ha riferito che in soluzioni acquose contenenti amminoacidi e polifosfati (entrambi a concentrazioni tra 0,01 e 0,1 M) si formano peptidi a 70 °C o anche a temperatura ambiente, a vari valori di pH. Il pH ottimale è intorno alla neutralità (pH 7-8), e la reazione procede anche in ambiente alcalino ma non in un mezzo acido. La reazione di condensazione avviene, con un basso rendimento (0,4-0,5%), anche con pirofosfato e a temperatura ambiente. Quanto più lunga era la catena del polifosfato lineare, tanto maggiore era il rendimento (fino al 13,6%), come era stato originariamente suggerito da Gabel per le reazioni di polimerizzazione di sostanze organiche in soluzioni diluite. È possibile che polimerizzazioni apprezzabili avvengano a diluizioni molto maggiori se i reagenti sono adsorbiti su un'interfaccia o incorporati in una matrice solubilizzata o in sospensione. Lasciando per vari giorni della glicina marcata in una soluzione acquosa contenente polifosfati a basso peso molecolare (con una molarità in fosforo di 10-3) e sali inorganici in modo da rispecchiare la composizione dell'acqua di mare a pH 7,4, diventava non più dializzabile una quantità apprezzabile di materiale radioattivo.
La formazione di peptidi da una miscela di metano, ammoniaca e acqua è stata per la prima volta descritta da Ellenbogen (v., 1958). La miscela fu sottoposta a irradiazione ultravioletta in presenza di particelle sospese di minerali inorganici. Secondo Ellenbogen, una notevole quantità di materiale organico, comprendente anche polipeptidi, potrebbe essere stata sintetizzata durante il processo di accrezione che si ritiene si sia verificato durante la formazione della Terra primordiale. Si suppone che materiale ad alto peso molecolare possa essere stato presente anche prima della formazione della crosta terrestre.
Ponnamperuma e Flores hanno descritto la formazione di peptidi da miscele di metano, ammoniaca e acqua sottoposte a scarica elettrica; tali peptidi sono risultati composti di tre o quattro residui amminoacidici; dopo idrolisi sono stati identificati nove amminoacidi diversi (v. Flores e Ponnamperuma, 1972).
Kliss e Matthews (v., 1962) hanno avanzato un'ipotesi che renderebbe conto della formazione di polipeptidi anche senza passare attraverso l'iniziale formazione di amminoacidi. Questi autori hanno proposto che l'amminocianometilene H2N−???67???=C=Ṅ, un biradicale 1,3, si formi da cianuro di ammonio e cianuro di idrogeno attraverso un composto intermedio, il diamminoacetonitrile. Una polimerizzazione di tipo additivo porterebbe poi a precursori polimerici di un peptide contenenti una unità ripetentesi caratterizzata da gruppi ammidinici invece che ammidici:
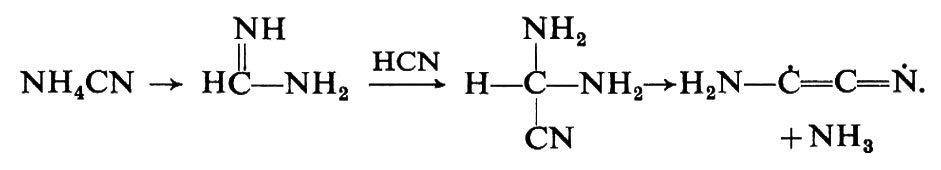
L'aggiunta di cianuro di idrogeno, una blanda idrolisi e la perdita di diossido di carbonio condurrebbero alla formazione di poliglicina:
Le catene laterali di altri amminoacidi potrebbero formarsi attraverso reazioni che avvengono a livello del carbonio metinico terziario prima o dopo idrolisi ed eliminazione del gruppo cianuro. A conferma di questa ipotesi è stato trovato che sottoponendo a scarica elettrica metano e ammoniaca, e riscaldando con acido diluito il residuo, si ottiene un materiale che appare essere un polipeptide (v. Matthews e Moser, 1966). La condizione limitante in questo schema di reazione è che in condizioni blandamente idrolitiche non si produce alcun effetto sul gruppo cianuro, ma si riuscirebbe solo a convertire i gruppi ammidinici in gruppi ammidici; il presupposto polipeptide, invece, dovrebbe avere un gruppo cianuro su ogni atomo α di carbonio. Inoltre, Huntress e collaboratori, dopo aver condotto uno studio approfondito per mezzo della risonanza ionociclotronica delle reazioni in fase gassosa del cianuro di idrogeno, hanno concluso che i processi di polimerizzazione non sono rilevanti (v. Huntress Jr. e altri, 1969). Akabori (v., 1955) ha avanzato un'ipotesi riguardo alla formazione di ciò che egli ha chiamato ‛proproteina' da amminoacetonitrile (v. Ponnamperuma e Woeller, 1967), dal quale si formerebbe una poliglicina. Anche in questo caso i gruppi ricorrenti in questa ‛proproteina' sarebbero gruppi ammidinici piuttosto che gruppi ammidici:
Hanafusa e Akabori hanno impiegato con successo questa reazione per produrre di- e triglicina. Akabori e altri hanno dimostrato che i gruppi metilenici della poliglicina possono condensarsi con aldeidi in condizioni debolmente basiche, formando in tal modo varie catene laterali.
i) Polinucleotidi.
Una volta Beadle ha osservato che i polinucleotidi primordiali avrebbero potuto esser costituiti da un polimero tipo adenina-timina, che in seguito si sarebbe sviluppato negli odierni polinucleotidi. La fosforilazione dei nucleotidi per riscaldamento con fosfati inorganici produce alcuni oligonucleotidi (v. Ponnamperuma e Mack, 1965); la temperatura necessaria per questa reazione è di circa 150 °C e il rendimento è molto scarso. Rabinowitz e altri hanno dimostrato che, proprio come quando si fa la sintesi di nucleotidi nelle stesse condizioni di reazione, il vero agente fosforilante è una miscela di polifosfati che si formano dai vari ortofosfati usati in questo esperimento.
I composti usati per queste reazioni di condensazione possono essere divisi in due classi: composti che contengono il gruppo CN e fosfati condensati. Alla prima classe appartengono i cianuri, le carbodiimmidi e il tetramero del cianuro di idrogeno, alla seconda i polifosfati lineari e quelli ciclici (v. tab. IV).
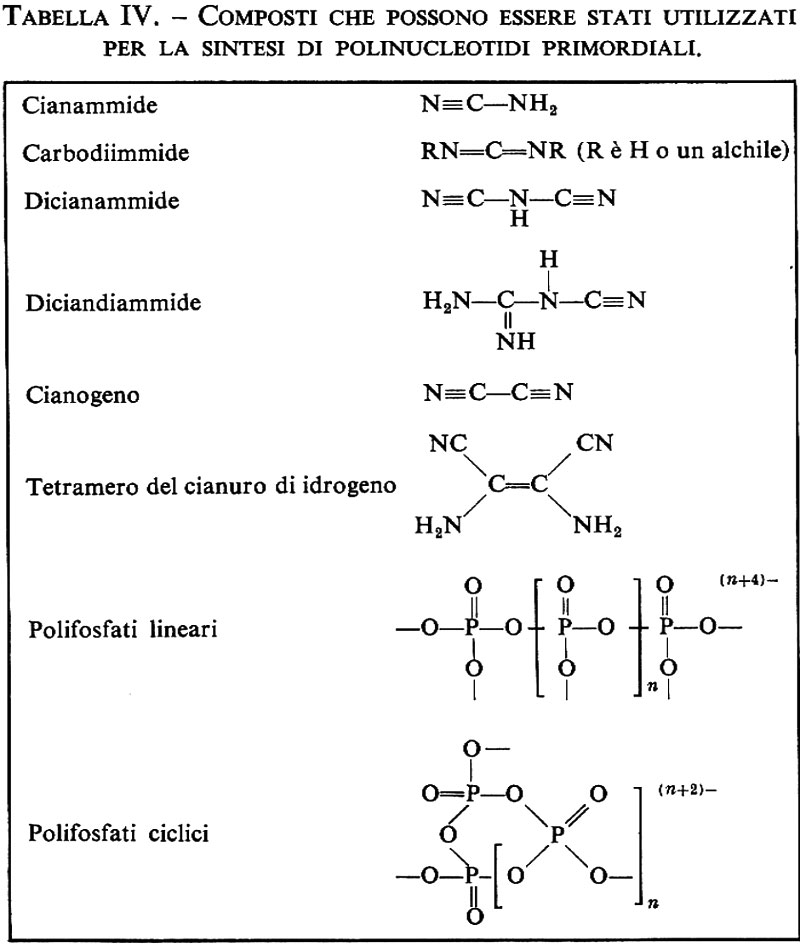
5. Lo sviluppo delle nuove teorie scientifiche.
I progressi della biochimica quantistica hanno gettato nuova luce sull'origine delle molecole biologiche. Il lavoro di B. e A. Pullman (v., 1962) ha chiarito alcune caratteristiche fondamentali: infatti, quando un chimico quanti stico prende per la prima volta in considerazione le molecole significative che si riscontrano negli organismi viventi, rimane colpito dal fatto che quasi tutte le biomolecole essenziali ai processi vitali sono sistemi coniugati, ricchi di elettroni π. Tutte e tre le unità fondamentali, gli acidi nucleici, le proteine e i fosfati ad alto contenuto energetico, mostrano questa caratteristica delocalizzazione elettronica.
I costituenti più significativi della molecola dell'acido nucleico sono le purine e le pirimidine, che sono eterocicli coniugati. Mentre a prima vista le proteine non appaiono essere coniugate, la loro struttura generale implica un certo grado di delocalizzazione. Nella struttura elicoidale delle proteine c'è una matrice di legami a idrogeno che rende possibile un certo grado di trasferimento di elettroni. Nel caso dei fosfati ad alto contenuto energetico, gli elettroni mobili di un gruppo fosforico interagiscono con gli elettroni di un altro gruppo fosforico o con gli elettroni π di un radicale organico. Tra le molecole coniugate importanti troviamo le porfirine, che sono costituite da quattro anelli pirrolici uniti a un atomo metallico centrale. La struttura esterna delle porfirine è rappresentata da una sequenza di doppi legami alternati, il che implica un alto grado di coniugazione e di delocalizzazione elettronica. Non si può quindi fare a meno di concludere, anche dopo un'osservazione così superficiale, che le manifestazioni basilari della vita sono intimamente connesse con l'esistenza di composti altamente coniugati. Per qualche ragione, la natura ha scelto questi composti quale mezzo per produrre la vita. La delocalizzazione elettronica è forse il più grande effetto quantistico riscontrabile nell'evoluzione biochimica.
Soffermiamoci per un momento a considerare come questa singolare caratteristica delle biomolecole possa spiegare il fatto che esse sono i principali mattoni con cui è stata costruita la materia vivente. Due dei fattori più importanti sembrano essere le caratteristiche di stabilità e funzionalità. Il principale risultato della delocalizzazione è l'aumento della stabilità, che da un punto di vista quantitativo è definito come energia di risonanza. Tale stabilizzazione termodinamica deve aver giocato un ruolo rilevante nella selezione delle molecole primitive. Si può supporre che ci sia stato un periodo durante il quale si è avuta una lotta per la sopravvivenza; c'è stata una selezione di biomolecole e alla fine vennero scelte le più stabili. Questo dato è confermato dalla straordinaria uniformità della biochimica: lo stesso limitato numero di composti esplica le stesse funzioni in tutto il regno animale e vegetale. Per fare un esempio, la clorofilla, che è usata dalle piante, è molto simile all'emoglobina, che è usata dagli animali: ambedue i composti vengono sintetizzati tramite la stessa sequenza di reazioni. La stabilizzazione attraverso la delocalizzazione di elettroni può aver avuto una parte anche nell'orientamento spaziale delle piccole molecole che danno luogo ai grandi polimeri, quali acidi nucleici e proteine.
In definitiva, si possono trarre le seguenti conclusioni sul possibile ruolo della delocalizzazione elettronica: la selezione evolutiva ha impiegato i composti più stabili che, grazie alla delocalizzazione di elettroni, sono risultati i più adatti agli scopi biologici; la vita non ha avuto inizio quando hanno fatto la loro comparsa i composti coniugati, ma le probabilità che si sviluppasse la vita quale noi la conosciamo sono aumentate in seguito alla loro apparizione.
a) Organizzazione precellulare.
La formazione delle membrane, la cui funzione è di isolare i microsistemi dall'ambiente circostante, si è probabilmente verificata a uno stadio precoce dell'evoluzione chimica. Gli esperimenti svolti in laboratorio hanno mostrato che si possono formare prodotti a struttura complessa da alcani che siano stati irradiati con luce ultravioletta in presenza di fosfato acquoso e ioni magnesio. Holland e collaboratori hanno suggerito che gli oceani primitivi avrebbero potuto essere coperti da uno strato di petrolio derivante dall'irradiazione del metano (v. Lasaga e altri, 1971). Se la radiazione ultravioletta avesse indotto la formazione di strutture membranose, le molecole presenti nell'ambiente avrebbero potuto essere incorporate in questi microsistemi.
Sono stati ipotizzati parecchi sistemi prebiologici in cui avrebbero potuto essere presenti delle membrane (v. Shah, 1972): tra questi ricordiamo le vescicole lipoproteiche di Goldacere, i coacervati di Oparin, le microsfere di Fox e le lipoproteine intorno a goccioline sotto forma di aerosol di Shaw, ma possiamo solo fare delle ipotesi su quale di questi sistemi sia stato il più probabile per l'evoluzione prebiologica. È ragionevole ritenere che in ambienti diversi siano esistiti sistemi diversi; secondo Goldacere la formazione di vescicole a due strati ha avuto luogo più probabilmente sulla superficie di contatto aria-acqua degli oceani primitivi, dei bacini di marea, di laghi e lagune. Se i lipidi sono presenti in fase acquosa è probabile che essi si trovino sulla interfaccia con l'acqua. La distribuzione di carica su questa sottile superficie è probabile che determini un certo grado di specificità nelle interazioni lipidi-polimeri a livello dell'interfaccia, dato che dei polimeri presenti in fase acquosa solo quelli aventi opposte distribuzioni di carica reagiranno con la sottile pellicola lipidica. Oltre alle interazioni ioniche, anche i polimeri tensioattivi tenderanno a favorire le associazioni lipidi-polimeri, e pertanto i polimeri dotati di molte catene laterali idrofobiche tenderanno ad adsorbire sullo strato lipidico la loro interfaccia con l'acqua. La fase acquosa che resta inclusa all'interno delle vescicole può contenere dei composti organici; le reazioni che avvengono all'interno di tali vescicole possono divenire selettive, dato che il trasporto dei diversi composti dall'esterno all'interno e quello del prodotto di una reazione dall'intemo all'esterno delle vescicole possono dipendere dalla carica superficiale e dall'estensione superficiale della membrana della vescicola.
La quantità di informazione contenuta nelle membrane di tali vescicole dipenderà dalla distanza intermolecolare esistente tra le molecole lipidiche, dalla distribuzione della carica sulla pellicola lipidica e sulle molecole polimeriche e da numero, caratteristiche e distribuzione dei gruppi idrofobici.
Il secondo sistema ipotizzato in relazione all'origine delle membrane prebiologiche è quello delle goccioline di coacervato. Il processo di coacervazione è uno dei metodi più efficaci per concentrare sostanze altamente polimerizzate da soluzioni molto diluite; per esempio, è possibile isolare una gocciolina di coacervato dalla soluzione in cui si trova, contenente gelatina a una diluizione dello 0,001%, ottenendo nella gocciolina una concentrazione di polimero che può arrivare al 10%. Anche se hanno la consistenza di un liquido, le gocce di coacervato sono nettamente segregate mediante uno strato limite dalla soluzione acquosa che le circonda. Questa separazione deve essere avvenuta nel ‛brodo primordiale' in cui ebbe luogo la polimerizzazione non specifica di una quantità di composti organici. Le gocce di coacervato rappresentano dei sistemi multimolecolari che possono passare con facilità da uno stato statico a uno dinamico interagendo con l'ambiente. Per formare dei coacervati, Oparin ha usato polimeri naturali e non naturali ad alto peso molecolare e svariate miscele di polimeri; ha incorporato degli enzimi nel sistema del coacervato dimostrando che l'aumento di attività nelle gocce di coacervato è pressappoco lo stesso che i composti presenti al loro interno avrebbero se si trovassero in soluzione. Oparin sostiene che il processo di coacervazione, che interessa i più semplici e importanti materiali biologici, potrebbe aver portato alla formazione della protocellula. A causa della loro struttura labile i coacervati possono dividersi o moltiplicarsi, determinando cosi la ricombinazione e la separazione di vari sistemi chimici. Questo processo può aver determinato la graduale evoluzione di goccioline di coacervato capaci di effettuare una serie di reazioni chimiche. La labilità della struttura dei coacervati è una delle loro caratteristiche più affascinanti, perché può portare a sistemi a reazioni multiple dovute alla ricombinazione di parecchie goccioline di coacervato.
Un terzo sistema di grande interesse per ciò che riguarda l'origine dell'organizzazione precellulare è quello delle microsfere, proposto da Fox e dai suoi collaboratori. Quando polipeptidi prodotti mediante riscaldamento vengono reidratati in acqua calda, il loro raffreddamento o la loro precipitazione darà luogo a una miriade di microsfere che, da un punto di vista morfologico, assomigliano moltissimo a dei micrococchi. Tali microsfere sono state chiamate ‛microsfere proteinoidi' e, a quanto è stato sostenuto, possono rappresentare una delle vie attraverso cui si sarebbero formate le prime protocellule. Esse hanno struttura sferica (2-7 μm di diametro), sopportano una centrifugazione a 3.000 giri al minuto e si mantengono stabili anche se manipolate o tagliate in sezioni sottili; in soluzioni ipotoniche si rigonfiano, e si restringono in soluzioni ipertoniche. Insomma, per molte delle loro caratteristiche esse sono state proposte come un modello plausibile della protocellula.
Secondo Shaw, la presenza di lipidi e proteine negli oceani primitivi può aver dato luogo alla formazione di una pellicola di lipoproteine adsorbita sulla loro superficie. Anche le goccioline sotto forma di aerosol che si sarebbero formate in conseguenza del frangimento delle onde avrebbero mantenuto sotto la superficie il sottile strato lipoproteico, con la porzione non polare delle molecole lipidiche rivolta verso l'esterno delle goccioline di acqua. Ricadendo sulla superficie dell'oceano, le goccioline si sarebbero nuovamente ricoperte con un sottile strato di lipoproteine. Tale processo avrebbe prodotto qualcosa di simile a delle strutture membranose. Le reazioni che sarebbero avvenute all'interno delle goccioline sarebbero state influenzate dalle caratteristiche della superficie e della permeabilità della membrana, che è strutturalmente simile a quella proposta da J. F. Danielli per i globuli rossi. Poiché sulla superficie dell'oceano avrebbe potuto formarsi una grandissima quantità di goccioline diverse, la natura avrebbe avuto la possibilità di mettere alla prova miliardi di strutture membranose saggiando le migliori per la futura evoluzione. Le goccioline sotto forma di aerosol coperte da membrana possono pertanto aver costituito una struttura autoformata e organizzata negli oceani primitivi.
b) Interazione tra acidi nucleici e proteine.
Forse il problema più importante che deve affrontare lo studioso di evoluzione chimica è quello di spiegare come si siano originati i sistemi viventi partendo dalla sintesi proteica guidata dagli acidi nucleici. Al momento non esistono prove sperimentali che indichino quali siano stati i primitivi precursori dell'attuale sistema di traduzione. Si può ipotizzare che le forme primitive di traduzione siano state abbastanza diverse da quelle odierne e che la traduzione sia stata imprecisa oppure ambigua, poiché l'accuratezza e la precisione dell'attuale sistema dipendono dalla precisa funzione di molte, se non di tutte le sue componenti altamente evolute e sofisticate, quali enzimi attivatori e proteine ribosomiali. Dato che l'accuratezza e la precisione di almeno alcune di queste proteine sono il risultato dell'evoluzione dell'intero sistema, si può presumere che abbia avuto minore importanza la precisione del ruolo dei precursori dell'attuale sistema di sintesi proteica e che abbia invece avuto importanza maggiore il ruolo dei corrispondenti acidi nucleici. Le ipotesi riguardo all'origine del codice genetico devono pertanto attribuire un ruolo importante alla funzione dell'acido nucleico nella determinazione dei compiti del codone e nella disposizione ordinata dell'attuale codice. Una delle possibilità è che la struttura del gruppo dei codoni e il processo di sintesi proteica riflettano in qualche modo il riconoscimento o una qualche forma di interazione catalitica tra nucleotidi e amminoacidi, magari solo durante uno stadio primitivo della storia dell'evoluzione cellulare. Per indagare questa possibilità, sono stati intrapresi degli studi sulle interazioni tra amminoacidi e oligonucleotidi, al fine di stabilire quali siano i possibili tipi di interazione tra queste due classi di composti, di imparare come utilizzare le nozioni derivanti da risultati di facile interpretazione per capire sistemi più complessi, e infine di determinare il grado di selettività ottenibile in sistemi di crescente complessità chimica.
Molti studi hanno mostrato che tra acidi nucleici e amminoacidi possono avvenire una quantità di reazioni diverse a seconda della composizione, conformazione, stato di polimerizzazione e ambiente in cui si trovano le specie chimiche reagenti. Si è così arrivati a concludere che un certo grado di specificità è innegabile, anche se non si è riusciti a capire a che cosa essa sia dovuta. Quando due specie reagenti sono semplici, non ci si può aspettare di osservare quello stesso tipo di ‛specificità' implicito nell'uso biologico di questo termine; ai livelli più semplici, si può solo cercare di mettere in evidenza un qualche tipo di selettività, cioè casi in cui, in determinate circostanze, un amminoacido forma con un nucleotide un legame la cui forza è in certo grado una funzione della composizione di tutte e due le specie reagenti. Anche se risultati di questo tipo ottenuti da semplici sistemi proteina-acido nucleico non potrebbero essere definiti spettacolari da un punto di vista biologico, tuttavia rappresentano un primo passo nello studio sistematico del ruolo delle interazioni amminoacidi-acidi nucleici durante l'evoluzione dei sistemi viventi. Gli studi su specie monomeriche in mezzi acquosi possono permettere di stabilire gli effetti di fattori individuali e fornire quelle informazioni di base che sono necessarie per interpretare le più complesse interazioni tra polinucleotidi e amminoacidi. Se queste interazioni esistono, di che natura sono? Sono selettive? Possono portare alla sintesi proteica guidata dagli acidi nucleici, o aiutare a capire il sistema? Possono essere usate per comprendere sistemi più complessi? Possono portare allo stabilirsi di condizioni limitanti il cui risultato sia il codice genetico in un definito sistema prebiotico?
L'uso di acidi nucleici immobilizzati su colonne a scambio ionico, gli studi con la risonanza magnetica nucleare per mettere in evidenza gli spostamenti dei protoni, e il possibile ruolo delle superfici complementari quali catalizzatori della polimerizzazione degli amminoacidi attivati hanno reso possibile avanzare alcune ipotesi sulla parte svolta dai fattori stereochimici nell'origine del codice genetico. Un altro possibile approccio sarebbe quello dello studio dei processi biologici, quali essi sono negli odierni organismi viventi, prendendoli in considerazione nella loro forma più semplice ed esaminandoli in modo da vedere se i componenti più primitivi degli acidi nucleici nell'attuale apparato di traduzione avrebbero potuto essere efficaci in un processo di traduzione e di replicazione organizzato in modo molto più semplice.
6. La ricerca delle prime forme di vita.
Gli studi effettuati hanno permesso di stabilire che le molecole necessarie per la vita possono essere sintetizzate in laboratorio. Un problema essenziale è quindi quello di stabilire se questo processo sia in realtà avvenuto, sulla Terra o in qualche altra parte dell'Universo. Risalendo indietro nel tempo ai primi periodi della Terra e del sistema solare, possiamo ottenere qualche indicazione della presenza di molecole organiche sintetizzate in un periodo prebiologico? Sono stati fatti molti tentativi per risolvere questo problema. Quando ebbe luogo il passaggio da sistemi chimici a sistemi biologici? La documentazione biologica, che si spinge sempre più indietro nel tempo, restringe il periodo in cui può essere avvenuta l'evoluzione chimica. L'età della Terra può essere rappresentata in un diagramma di 12 ore (v. fig. 2): in questo sistema l'inizio del Cambriano viene fatto risalire a 600 milioni di anni fa. Per un certo periodo si è ritenuto che prima del Cambriano la vita non esistesse: i paleontologi sostenevano che non esistevano scheletri e che quindi non poteva esservi stata vita. Invece, il lavoro dei micropaleontologi ha permesso di stabilire la presenza di vita in periodi ben anteriori ai 600 milioni di anni. Risalgono a un miliardo di anni fa le formazioni di Bitter Springs, in Australia, in cui è stata riscontrata la presenza di microstrutture, mentre la formazione dello Swaziland, in Sudafrica, risale addirittura a 3-3,5 miliardi di anni fa.
Ci sono solo tre luoghi sulla Terra che concordemente vengono fatti risalire a oltre 3,6 miliardi di anni fa: la regione di Isua nella Groenlandia occidentale, la regione di Saglefjord nella provincia di Nain nel Labrador settentrionale e il complesso granitico di Singhbhum nell'india orientale.
La prima prova che la vita risale a un periodo precedente al Cambriano è venuta nel 1954 dalla scoperta di microfossili, a opera di Barghoorn e Tyler (v., 1962), nelle sezioni sottili provenienti dalle selci di Gunflint. Da allora sono stati ritrovati parecchi depositi contenenti microfossili risalenti all'Algonchiano. La conservazione delle fini caratteristiche strutturali rende possibile un paragone con specie tuttora esistenti e con modelli ecologici e ha reso possibile accertare senza ombra di dubbio che molti microfossili proterozoici sono realmente resti di organismi precambriani.
Tra i più antichi microfossili rigorosamente accertati vi sono i semplici sferoidi ritrovati nelle selci di Swartkoppie, situate immediatamente sopra alla Onverwacht Superiore del Sudafrica, che hanno un'età di 3,4 miliardi di anni (v. Knoll e Barghoorn, 1977). Meno certa appare l'origine dei microfossili trovati recentemente nel Pilbara Block dell'Australia occidentale, che appaiono mal definiti e assomigliano ad alcune delle microstrutture delle selci di Onverwacht e che non sarebbero neppure stati considerati di origine biogena se non fossero stati associati con stromatoliti. Anche se è possibile, anzi probabile, che almeno alcune delle antiche microstrutture dell'Archeano siano davvero microfossili di organismi un tempo viventi, mancano prove inequivocabili in tal senso. Attualmente, tecniche quali quella della distribuzione delle dimensioni possono avere solo un'applicazione limitata, e la micropaleontologia non riesce a discriminare tra fossili provenienti da organismi viventi primitivi e fossili che possono aver avuto origine abiotica.
a) Macropaleontologia.
Le stromatoliti sono sedimenti macroscopici formati da detriti e minerali precipitati rimasti inglobati e tenuti insieme dal rivestimento calcareo di un banco di alghe. Questi banchi rappresentano ecosistemi completi. I sedimenti rimasti inglobati a opera delle Cianofite formano uno strato compatto e opaco che costringe la comunità a migrare verso l'alto; col progredire di tale processo, strati di detriti si depositano tra strati di resti organici e microrganismi inglobati. Mentre le stromatoliti attuali hanno dimensioni ridotte e sono limitate ad ambienti sfavorevoli che escludono la presenza di Metazoi, esse erano particolarmente fiorenti durante il Precambriano.
Benché anche alcuni processi di sedimentazione non biologica possano dar luogo a strutture stratiformi dello stesso tipo, le stromatoliti possono essere considerate la migliore indicazione di vita nell'Archeano. Gli esempi noti sono molti: le stromatoliti del Bulowaya sono state datate a circa 2,7 miliardi di anni fa (v. Schopf, 1972); quelle del supergruppo Pangola, in Sudafrica, sono un po' più vecchie; più spettacolari quelle dell'Australia occidentale che risalgono a 3,4 miliardi di anni fa (v. Walter e altri, 1980). Più recentemente sono state descritte stromatoliti simili nello Zimbabwe (v. Orpen e Wilson, 1981). Anche se appaiono metamorfosate nella faccia inferiore a scisti verdi, le caratteristiche di queste stromatoliti dell'Archeano inferiore sono incredibilmente simili a quelle più recenti e sicuramente biogeniche, e rappresentano la migliore prova che la vita è esistita almeno 3,4 miliardi di anni fa.
b) Fossili molecolari.
Con le analisi di geochimica organica, effettuate sui sedimenti del Precambriano, si è tentato di determinare la natura dei paleoecosistemi di origine sedimentaria attraverso la caratterizzazione dei costituenti molecolari del carbonio. Lo scopo principale della maggior parte di questi studi è quello di isolare e identificare dei marcatori biologici, cioè fossili chimici le cui configurazioni molecolari e abbondanze isotopiche possano essere indizio di attività biologica; infatti, specifici fossili chimici possono essere caratteristici di specifiche vie metaboliche. L'origine della vita, lo sviluppo di vie fotosintetiche da parte dei Batteri, la comparsa di un'organizzazione eucariotica e anche altri eventi dell'evoluzione biochimica potrebbero esser stati conservati nelle rocce non solo sotto forma di microfossili, ma anche sotto forma di fossili chimici di origine discreta. Tuttavia, quando si voglia arrivare alla rappresentazione di un paleoecosistema attraverso i fossili chimici del Precambriano, bisogna affrontare due problemi, e cioè l'alterazione dei composti organici primari e la contaminazione avvenuta in epoca successiva a quella in cui il deposito si è formato. Nei sedimenti recenti, i componenti estraibili sono simili ai composti biochimici sintetizzati dai microrganismi contemporanei; i composti organici primari, invece, vengono rapidamente alterati dalla degradazione batterica e dai processi diagenetici. I biopolimeri vengono raramente idrolizzati nei loro componenti. I gruppi funzionali sono facilmente sostituiti dall'idrogeno e le configurazioni molecolari vengono alterate da processi di racemizzazione e da cambiamenti di conformazione. L'aumento della temperatura, il cracking termico e la condensazione di composti organici possono dar luogo alla formazione di cherogene, una macromolecola insolubile senza struttura definita. Una gran parte del carbonio organico esistente sulla crosta terrestre era sotto forma di cherogene insolubile; temperature altamente metamorfiche determinarono ulteriori alterazioni del cherogene fino a ottenere composti come la grafite, in cui sono presenti solo legami carbonio-carbonio. Venne così meno qualsiasi informazione di tipo molecolare, e i dati riguardanti gli isotopi possono anch'essi risultare del tutto alterati. I composti organici avrebbero potuto inserirsi nei sedimenti precambriani in epoca successiva a quella di formazione dei depositi, e sembra che le selci permeabili siano abbastanza porose e permeabili alle acque sotterranee da aver potuto fornire quantità significative di contaminanti organici recenti. D'altra parte, il materiale carbonioso insolubile, che può costituire anche il 98% o più del carbonio totale di un sedimento precambriano, viene generalmente ritenuto singenetico, dato che ha dimensioni troppo grandi per esser considerato mobile.
Maggiori informazioni sono sicuramente ricavabili dalla frazione solubile che dal cherogene o dalla grafite; tuttavia, può essere assolutamente impossibile capire se le molecole osservate risalgano al tempo in cui si sono formati i depositi o se vi si siano introdotte successivamente. Perciò, volendo usare i fossili molecolari come indicazione della presenza di vita, bisogna essere estremamente cauti e tener presente la possibilità di contaminazioni e processi diagenetici. Nel lavoro effettuato sui sedimenti di Isua non è stata prestata alcuna attenzione ai materiali organici estraibili; tuttavia, la grafite presente è stata studiata attentamente e la maggior parte dei campioni sembra presentare un frazionamento isotopico che ricade nell'ambito biologico. Però, alcuni campioni possono dare risultati anomali che, fintantoché non vi siano prove del contrario, possono suggerire che la vita sulla Terra è vecchia quanto le più antiche rocce. Se ciò fosse davvero provato, risulterebbe forse impossibile trovare sulla Terra le prove di quei processi prebiologici che abbiamo tentato di verificare in laboratorio; tali prove dovranno allora essere cercate in altre parti del sistema solare, quali ad esempio la Luna, le meteoriti e la superficie dei pianeti.
7. La ricerca di forme di vita extraterrestri.
a) Analisi dei campioni lunari.
Le missioni Apollo hanno fornito un'occasione unica, quella di analizzare i campioni che sono stati riportati dalla Luna. I campioni prelevati dalle missioni 11-16 sono stati analizzati in laboratorio; tali studi hanno incluso analisi del contenuto totale di carbonio, della composizione isotopica del carbonio organico, dei microfossili, oltre a indagini mineralogiche. Il trattamento sequenziale del campione lunare con benzene, metanolo, acqua e acido cloridrico ha fornito degli estratti, che sono stati poi sottoposti a esame cromatografico e spettrometrico. Per minimizzare le possibilità di contaminazione, le analisi sono state eseguite nella massima pulizia, in laboratori la cui aria era stata filtrata, e tutta la sequenza delle estrazioni con solventi è stata effettuata nello stesso recipiente di vetro. Il contenuto di carbonio totale è stato determinato misurando il volume di CO2 che si sviluppa degassando un campione di 1 grammo a una temperatura di 150 °C e a una pressione inferiore a 10-3 mmHg e quindi bruciandolo a 1.050 °C. I valori variavano tra 140 e 200 μg/g; quelli più comunemente ottenuti erano compresi tra 140 e 160. La quantità di carbonio che si è riusciti a convertire in idrocarburi volatili è stata determinata sottoponendo a pirolisi circa 30 mg di polvere in un'atmosfera di idrogeno ed elio. I composti non volatili ottenuti in tal modo erano valutati con un rivelatore a ionizzazione di fiamma di idrogeno; i valori medi sono risultati di 40 μg/g. Le misurazioni degli isotopi su tutto il campione hanno dato un valore di δ 13C di +20 rispetto allo standard PDB, valore che è considerevolmente più alto di quelli ottenuti su meteoriti intatte (v. Ponnamperuma, 1970).
L'osservazione al microscopio ottico ed elettronico della polvere e della superficie delle microbrecce in sezioni sottili non ha messo in evidenza alcuna struttura biologica originaria. Per identificare composti organici estraibili, una parte della polvere lunare è stata trattata con una miscela di benzene e metanolo; gli spettri di fluorescenza, in eccitazione e in emissione, degli estratti così ottenuti possono suggerire la presenza di porfirine. Peraltro, risultati simili sono stati ottenuti per i prodotti di scarico dei motori per la discesa situati sui razzi lunari, ed è quindi possibile che queste porfirine si siano formate da alcuni componenti dello scarico dei razzi.
La cromatografia gas-liquido ha mostrato che nessuno degli alcani normali da C12 a C32 era presente a concentrazioni superiori a 2 × 10-5 μg/g. Trattando i campioni con HCl 6N è stato possibile identificare gli idrocarburi da C1 a C4, il che fa pensare che nei campioni fossero presenti dei carburi. Nonostante che la sensibilità delle tecniche impiegate fosse dell'ordine dei nanogrammi, non sono stati trovati alcani normali, isoprenoidi, idrocarburi aromatici, acidi grassi, amminoacidi, zuccheri e basi di acidi nucleici a queste concentrazioni. È chiaro che i campioni della superficie della Luna sono stati esposti a luce ultravioletta di grande intensità e a bombardamenti da parte di micrometeoriti, e pertanto in queste condizioni i legami carbonio-carbonio potrebbero essersi facilmente rotti.
b) Condriti carboniose.
Per oltre un secolo le meteoriti sono state esaminate per mettere in evidenza l'eventuale presenza di composti organici. La meteorite di Alais fu analizzata da Berzelius, quella di Kaba da Wöhler, quella di Orgueil da Berthelot. Ormai viene generalmente ammesso che le condriti carboniose contengono effettivamente materiale organico sotto forma di polimeri; tuttavia, i risultati ottenuti in passato sono dubbi per ciò che riguarda l'origine del materiale organico da esse estratto: era realmente extraterrestre o era dovuto a contaminazione di origine terrestre? L'analisi della meteorite di Murchison ha fornito la prima indubbia e decisiva prova della presenza di composti organici extraterrestri nelle meteoriti. La condrite carboniosa di tipo 2 Murchison cadde il 28 settembre 1969 vicino a Murchison, nello Stato australiano di Victoria. Molti frammenti vennero raccolti subito, altri durante i mesi di febbraio e marzo 1970. Le pietre scelte per essere analizzate furono quelle più grandi e che presentavano meno spaccature e il minor grado di contaminazione superficiale. I campioni analizzati contenevano una quantità di carbonio e di azoto pari al 2% e allo 0,16%, rispettivamente, del peso; in essi fu possibile identificare degli idrocarburi alifatici. I campioni avevano distribuzione simmetrica e il tempo massimo di ritenzione corrispondeva a C18. Gli idrocarburi mostravano una grande somiglianza con quelli prodotti tramite scarica elettrica in laboratorio: in entrambi i casi si trattava di alcani contenenti la stessa serie dominante di analoghi. Queste somiglianze fanno pensare che gli idrocarburi della meteorite di Murchison abbiano avuto un'origine abiogena. Anche gli studi sugli idrocarburi aromatici hanno dato risultati simili (v. Pering e Ponnamperuma, 1971). Gli estratti benzenici sono stati analizzati tramite gascromatografia e gascromatografia accoppiata a spettrometria di massa. L'origine termica degli idrocarburi aromatici della meteorite di Murchison è suggerita dalla loro somiglianza con quelli sintetizzati per via termochimica dal metano; in ambedue i gruppi di idrocarburi aromatici predominavano i composti polinucleari aventi un numero pari di atomi di carbonio. La relativa semplicità della composizione della frazione degli idrocarburi aromatici è interessante in vista della più complessa natura degli idrocarburi alifatici.
Per rivelare l'eventuale presenza di amminoacidi, le parti interne della meteorite sono state polverizzate, estratte con acqua bollente e analizzate mediante gascromatografia combinata a spettrometria di massa. Sono stati cosi individuati un gran numero di amminoacidi, quali glicina, alanina, valina, prolina, acido glutammico e acido aspartico; inoltre sono stati trovati vari amminoacidi non proteici, quali N-metilglicina, β-alanina, 2-metilalanina, acido α-amminobutirrico, acido β-amminobutirrico e acido γ-amminobutirrico. Degli amminoacidi con centri asimmetrici sono stati trovati sia gli enantiomeri D che quelli L. La presenza di amminoacidi non proteici e di amminoacidi con uguale quantità di enantiomeri D ed L fornisce un valido supporto all'ipotesi di una loro origine prebiotica. Questi sono i primi risultati decisivi riguardo agli amminoacidi di origine extraterrestre (v. Kvenvolden e altri, 1971).
Studi simili a quelli effettuati sulla meteorite di Murchison sono stati condotti sulla meteorite di Murray, che cadde nel Kentucky nel 1951, e su quella di Mighei, che cadde a Odessa nel 1866 (v. Ponnamperuma, 1978). Queste meteoriti sono state contaminate da una inadeguata protezione e dalle manipolazioni. Le tecniche della gascromatografia e della spettrometria di massa rendono possibile separare gli amminoacidi e gli idrocarburi originari da quelli dovuti alla contaminazione terrestre: fino a questo momento sono stati trovati tre campioni contenenti amminoacidi di origine extraterrestre.
Più recentemente, l'esplorazione dell'Antartide ha dato nuovo impulso agli studi delle meteoriti. Nel 1971, il geologo giapponese Kenzo Yanai, durante una spedizione sulle alture di Yamato nell'Antartide, ha scoperto otto frammenti meteoritici; quando i frammenti furono analizzati, egli scoprì che essi appartenevano a otto meteoriti diverse. L'anno successivo ne sono stati identificati 300, l'anno dopo ancora 600, e attualmente possediamo 4.000 frammenti riportati dall'Antartide nel corso di questi anni. Di questi, circa 40 appartengono alla categoria delle cosiddette condriti carboniose; due di questi frammenti, Alan Hills e Yamato, sono stati analizzati in laboratorio e confermano i risultati ottenuti sulle meteoriti di Murchison, di Murray e di Mighei (v. Kotra e altri, 1979).
Una sensazionale caratteristica delle meteoriti dell'Antartide è che esse non sono contaminate: l'esterno e l'interno contengono la stessa quantità di amminoacidi. Queste sono le migliori meteoriti che abbiamo a disposizione, se non vogliamo andare a prendercele nella fascia degli asteroidi!
La ricerca delle basi eterocicliche, puriniche e pirimidiniche, presenti negli acidi nucleici è stata condotta anche sulle meteoriti. In un primo tempo è stata riportata la presenza di adenina e guanina nella meteorite di Orgueil, ma successivamente la presenza di questi composti nelle condriti carboniose è stata da alcuni messa in discussione. Tuttavia, recentemente il lavoro di Schwartz e di altri ha dimostrato con certezza che le basi sono effettivamente presenti nelle meteoriti (v. Van der Velden e Schwaitz, 1977). In altre parole, nelle condriti carboniose è possibile riscontrare la presenza non solo dei componenti delle proteine, ma anche di quelli degli acidi nucleici. I processi prebiologici che sono stati postulati per la Terra e che sono stati simulati in laboratorio sembra che siano avvenuti anche in altre parti del sistema solare, come è possibile verificare grazie alle condriti carboniose.
c) La ricerca della vita su Marte.
Nella letteratura è stata spesso avanzata l'ipotesi che la vita sia possibile su Marte. Le strutture canaliformi di Marte e la stagionale oscurità che cala sul pianeta hanno portato a supporre che vi si possano essere evolute forme di vita. Quando sono state lasciate da parte le supposizioni e sono state prese in considerazione le reali condizioni esistenti su Marte, i parametri fisici riscontrati sono risultati tali da poter ancora far pensare che delle forme di vita inferiori, come i microrganismi, potessero sopravvivervi. Prima delle missioni Viking sapevamo che l'atmosfera di Marte è costituita in gran parte da diossido di carbonio e tracce di acqua. La superficie del pianeta è raggiunta da un flusso di intense radiazioni ultraviolette, da cui gli organismi marziani, se fossero realmente esistiti, avrebbero dovuto proteggersi scavando dei tunnel nella superficie.
Le calotte polari sembrava che fossero costituite principalmente da ghiaccio e diossido di carbonio solido. Le condizioni dell'atmosfera di Marte sono state per lungo tempo oggetto di controversie, finché le missioni Mariner mostrarono che essa ha uno spessore pari a circa 1/100 di quella della Terra. Anche se di per sé una pressione così bassa può non essere un fattore determinante per la sopravvivenza degli organismi, può aver avuto un effetto sulla disponibilità di acqua. La quantità di acqua presente, però, era circa 1/1.000 di quella riscontrabile nell'atmosfera terrestre. I parametri, peraltro molto scarni, che avevamo a disposizione su Marte rivelavano che le condizioni sul pianeta erano difficili rispetto a quelle sulla Terra, ma non tali da escludere definitivamente la possibilità che vi fossero dei microrganismi (v. Ponnamperuma e Klein, 1970).
Furono così programmate le missioni Viking 1 e 2 per esplorare la superficie di Marte e vedere se vi fosse del materiale organico o se vi fosse potuta esistere una qualche forma di vita microbica (v. Klein, 1977). Gli esperimenti più rilevanti ideati per le missioni Viking riguardavano la ricerca di molecole organiche (v. Biemann e altri, 1977). Due campioni di suolo marziano furono portati alla temperatura di 500 °C per sottoporre a pirolisi qualunque eventuale molecola organica; il materiale vaporizzato fu immesso in una colonna gascromatografica ad alta risoluzione, connessa con uno spettrometro di massa a scansione. I primi risultati indicavano che il materiale organico sulla superficie di Marte era meno di una parte per milione; in seguito, questo numero venne precisato meglio e portò al sorprendente risultato di 10 parti per miliardo. Alla luce di tali risultati, avrebbe potuto esser considerato superfluo continuare a cercare su Marte forme di vita basate sul carbonio. Tuttavia, i biologi avrebbero potuto prospettare una situazione nella quale il vento o la polvere portava le spore di microrganismi al deserto di Chryse dove avrebbero potuto divenire vitali grazie a un qualche stimolo esterno. Anche se la tecnica della gascromatografia associata alla spettrometria di massa ha una soglia di rivelabilità più bassa, gli esperimenti dei biologi possono individuare un singolo organismo amplificando la sua azione. La sonda Viking effettuò tre esperimenti di biologia, e cioè gli scambi gassosi (gas exchange, GEX), la liberazione marcata (labelled release, LR) e la liberazione pirolitica (pyrolytic release, PR). Si trattava di approcci diversi, effettuati senza l'aggiunta di elementi estranei al suolo - tranne forse il vapor acqueo all'acqua e alle sostanze organiche presenti, in modo da stimolare l'attività metabolica. Negli esperimenti di scambi gassosi, un campione di un centimetro cubico di suolo era trasferito in una camera di incubazione resa umida da una miscela di composti organici in presenza di atmosfera marziana. A vari intervalli di tempo, campioni di questa atmosfera vennero sottoposti a esame, in quanto cambiamenti nella loro composizione avrebbero indicato un'attività biologica. Gli esperimenti di liberazione marcata si svolgevano in una camera in cui un campione di suolo marziano era bagnato da una sostanza nutritiva contenente glicina, alanina, lattato e glicolato, un ‛brodo' considerato appetibile per i microrganismi terrestri; ogni atomo di carbonio era marcato radioattivamente, per cui la liberazione di un prodotto volatile radioattivo del carbonio avrebbe indicato la presenza di attività biologica. Negli esperimenti di liberazione pirolitica, furono ideati degli esami per mettere in evidenza l'attività fotosintetica degli organismi per mezzo della fissazione del diossido e del monossido di carbonio radioattivi in composti organici. Un campione del suolo venne incubato alla luce nell'atmosfera di Marte alla quale erano state aggiunte dosi traccianti di gas radioattivo; dopo l'incubazione il gas che non aveva reagito veniva rimosso dalla camera, il campione era sottoposto a pirolisi e i composti organici fissati su una colonna contenente ossido di rame. Riscaldando la colonna a 700 °C, il materiale organico viene ossidato e liberato sotto forma di CO2 radioattivo, mentre un rivelatore di 14C registra l'effluente. Nel caso che uno degli esami dia esito positivo, l'esperimento può essere ripetuto usando suolo marziano sterilizzato tramite riscaldamento a 160 °C per tre ore. Tutti gli esperimenti hanno dato esito positivo; esperimenti controllati hanno mostrato che il procedimento di sterilizzazione rendeva inattivo il suolo. Ma si sono incontrate molte difficoltà: per esempio, nel caso degli esperimenti di scambi gassosi sono stati osservati dei cambiamenti significativi solo nel livello di ossigeno e non in quello di altri gas; inoltre, il livello di ossigeno si innalzava improvvisamente e poi rimaneva uguale, situazione che difficilmente può essere riferita ad attività microbica. Negli esperimenti di liberazione marcata, la liberazione di radioattività aumentava improvvisamente per stabilizzarsi dopo poco. I risultati positivi ottenuti negli esperimenti di liberazione pirolitica, invece, sembrava che somigliassero all'attività biologica quale si riscontra sulla Terra. In assenza di carbonio organico, sembrava ragionevole ipotizzare che reazioni organiche di superficie potessero render conto di tutti questi risultati, e in realtà la presenza di superossidi o metalli può fornire una spiegazione plausibile dei risultati ottenuti. Esperimenti in laboratorio in cui veniva simulato il suolo marziano, basandosi sulle informazioni rese disponibili dalle analisi effettuate dal Viking (ossido di ferro, una traccia di umidità e argon), e questo era esposto a luce ultravioletta in ambiente umido, producevano liberazione di ossigeno; inoltre, aggiungendo a questa miscela la stessa miscela radioattiva che era stata mandata su Marte, si otteneva liberazione di CO2, il che suggerisce che vi sia stato un processo di decarbossilazione (v. Ponnamperuma e altri, 1977). In assenza di altri dati, bisogna concludere, anche se con riluttanza, che la missione Viking non ha fornito alcuna prova a favore della vita su Marte, anzi i dati più favorevoli a questa ipotesi sembrano dovuti a possibili reazioni di superficie.
d) Giove, Saturno e Titano.
Lo studio dei grandi pianeti basato sui dati ottenuti dagli osservatori terrestri ci ha fornito una buona conoscenza della composizione delle loro atmosfere, che appaiono fortemente ridotte e con un alto contenuto di idrogeno, elio e idrocarburi. La missione Voyager, grazie alla quale è stato possibile effettuare un gran numero di esperimenti, ha dato un'ulteriore possibilità di comprendere la composizione di questi grandi pianeti e dei loro satelliti, specialmente attraverso le osservazioni nell'infrarosso. È chiaro che se si riscontrasse la presenza di idrocarburi, quali metano o acetilene, e di ammoniaca e fosfato, potrebbero aver luogo molte reazioni organiche. Studi di simulazione effettuati in laboratorio hanno mostrato con chiarezza che in queste condizioni potrebbe venir sintetizzato un gran numero di molecole organiche che sono state ritenute prebiotiche (v. Ponnamperuma e Woeller, 1969); anzi, le immagini del loro materiale colorato (ad esempio la macchia rossa di Giove) potrebbero essere spiegate dalla formazione di materiale organico da metano e ammoniaca. La presenza di molecole organiche sui grandi pianeti, pertanto, è stata stabilita sia dalle osservazioni della navicella Voyager sia dagli esperimenti di simulazione effettuati in laboratorio.
Le eventuali reazioni che potrebbero avvenire su Titano, il satellite gigante di Saturno, sono per noi di grande interesse. Nelle fotografie che sono state inviate sulla Terra dalla missione Voyager, Titano appare come una sfocata palla arancione; i dati analitici indicano la presenza di un'atmosfera costituita principalmente da azoto con un contenuto di metano di circa il 2%, e per la prima volta è stata individuata la presenza di cianuro di idrogeno nell'atmosfera di un pianeta. Quando i dati della missione Voyager hanno indicato un'atmosfera composta da azoto per il 98% e da metano per il 2% sono stati effettuati molti esperimenti esponendo una tale miscela a varie fonti di energia; è stata in tal modo sintetizzata una grande varietà di molecole organiche (v. Gupta e altri, 1981), che in gran parte corrispondevano a quelle che si erano formate o erano state identificate nel corso della missione Voyager. I dati a nostra disposizione hanno chiaramente indicato la presenza di molecole organiche nei grandi pianeti e su Titano, il satellite di Saturno.
Per quanto riguarda la vita, le condizioni presenti su questi pianeti potrebbero essere tali da determinare forze di convezione che trasportano le molecole sintetizzate al centro del pianeta stesso, ad altissime temperature, e ciò potrebbe causare la loro demolizione. Le possibilità che in tali condizioni abbia origine la vita sono minime. Nel caso di Titano, nonostante che esso abbia un'atmosfera, la temperatura è probabilmente così bassa da ridurre le probabilità che le molecole si incontrino; tuttavia, il processo di evoluzione prebiotica può avvenire anche in queste condizioni.
e) Molecole interstellari.
La presenza di carbonio insieme a idrogeno, azoto e ossigeno porta inevitabilmente alla formazione di composti del carbonio. Le osservazioni astronomiche confermano la presenza nella Galassia di ammoniaca e vapor acqueo. Dal 1968 a oggi sono state identificate nel mezzo interstellare più di 50 molecole (v. Gammon, 1978), molte delle quali, come cianuro di idrogeno, formaldeide e cianoacetilene, sono considerate molecole chiave dell'evoluzione biochimica; quelle che hanno importanza particolare per l'evoluzione biochimica sono formaldeide, cianuro di idrogeno, cianoacetilene, metanolo, acido formico, monossido di carbonio, monosolfuro di carbonio, acetonitrile, metilacetilene, formammide, acetaldeide, acido isocianico, diformaldeide, metilenimmina, etere dimetilico, metirammina, etanolo, cianodiacetilene, acetilene, acrilonitrile, acetonitrile e solfuro di carbonio. È stato anche suggerito che il materiale altamente polimerico eventualmente presente potrebbe essere costituito da idrocarburi aromatici policiclici; altri pensano, invece, che potrebbe trattarsi di polisaccaridi. È difficile stabilire se queste molecole, singolarmente, abbiano contribuito in modo diretto a dare origine alla vita in qualche luogo. Tuttavia, da un punto di vista intellettuale, è di grande soddisfazione constatare che la sequenza dagli atomi di idrogeno, carbonio, azoto, ossigeno, fino alle piccole molecole quali cianuro di idrogeno e formaldeide, sembra essere la stessa in tutto il cosmo. Inoltre, la verifica nel mezzo interstellare del lavoro svolto in laboratorio, nel quale molecole come cianuro di idrogeno e formaldeide sembrano essere gli intermedi della sintesi prebiologica, aggiunge una nuova dimensione alla nostra comprensione del processo dell'evoluzione cosmica.
f) Esobiochimica.
Ci si chiede spesso se un elemento diverso dal carbonio o un solvente diverso dall'acqua avrebbe potuto avere un ruolo predominante in biologia. Sarebbe possibile scoprire su un altro pianeta una diversa forma di vita, basata magari sul silicio? La risposta a questa domanda ci riporta ancora una volta a considerare l'importanza del carbonio nella biologia terrestre. La maggioranza delle biomolecole è costituita da carbonio, idrogeno, azoto e ossigeno, con piccole quantità di fosforo e zolfo. I più importanti ioni monoatomici sono sodio, potassio, magnesio, calcio e cloro; gli elementi ferro, manganese, cobalto, rame e zinco compaiono solo in tracce, ed essendo elementi di transizione sono adatti a fungere da leganti in composti organometallici o entrano a far parte di enzimi di ossidoriduzione. Tutti questi sono elementi leggeri, compresi tra i primi trenta nella tavola periodica; degli altri, solo due - iodio e molibdeno - hanno una funzione come elementi in dosi traccia. Una delle risposte pertanto ci viene dal fatto che gli elementi più leggeri sono i più abbondanti. Tuttavia, al di là del semplice fatto di essere disponibili, devono esistere anche altri fattori, che potremmo genericamente raggruppare nel termine ‛idoneità', cioè una particolare combinazione di proprietà che rende questi elementi adatti per certe funzioni necessarie agli organismi. Sulla base della forza dei legami e della configurazione elettronica, il carbonio sembra più adatto del silicio a ricoprire il ruolo centrale nelle molecole che hanno importanza biologica.
8. L'acqua e l'origine della vita.
L'acqua ha un ruolo primario in quei processi che danno origine alla vita e la conservano. È stata spesso discussa la possibilità che si siano originate forme di vita in solventi che non siano l'acqua (v. Miller, 1964). Tra i molti solventi presi in considerazione ci sono ammoniaca liquida, cianuro di idrogeno, fluoruro di idrogeno, cloruro di idrogeno, alcoli, idrocarburi e anche alcuni sali; alcuni di questi possono essere scartati subito sulla base delle abbondanze atomiche, della rapida decomposizione, dell'instabilità dei composti organici disciolti in essi. Uno che merita una maggiore attenzione è l'ammoniaca liquida; data la grande abbondanza di acqua nel cosmo, essa contamina l'ammoniaca liquida dando luogo a tre miscele eutettiche contenenti rispettivamente il 34%, il 58% e l'82% di ammoniaca. Neppure questo sistema sarebbe adatto per gli organismi viventi, poiché la composizione del solvente cambierebbe considerevolmente a causa della diversa volatilità dell'ammoniaca e dell'acqua. Ciò inibirebbe la crescita di organismi, dato che i sistemi viventi hanno bisogno di un ambiente costante. Inoltre, i solidi hanno densità maggiore dei liquidi, per cui oceani di questo tipo gelerebbero a partire dal fondo e l'ambiente ne risulterebbe sicuramente danneggiato. (V. anche acqua e vita).
9. Attività ottica e origine della vita.
L'attività ottica è stata spesso indicata come una delle caratteristiche distintive delle molecole presenti nei sistemi viventi (v. Bonner, 1972). Negli organismi viventi qualunque tipo di sintesi e di degradazione coinvolge un solo enantiomero; mentre è logico che si sarebbe comunque perpetuato l'enantiomero di partenza, qualsiasi esso fosse, è difficile capire come sia stata fatta la scelta iniziale. Forze fisiche quali la luce polarizzata circolarmente, la superficie di cristalli asimmetrici o la cristallizzazione spontanea non spiegano la tendenza, assolutamente predominante, a produrre una forma piuttosto che l'altra (v. stereochimica).
La maggior parte del lavoro sperimentale fatto fino a ora ha fornito solo risultati marginali riguardo a questi processi di sintesi. Una ragionevole spiegazione può risiedere nel fatto che le necessità strutturali delle grandi molecole richiedono l'utilizzazione di una forma sola piuttosto che di forme miste, il che conferisce sicuramente stabilità ai polimeri; tuttavia, questo ancora non fornisce una risposta alla domanda su come venne fatta la scelta iniziale. Considerazioni di ordine matematico porterebbero a concludere che le fluttuazioni farebbero perpetuare una forma piuttosto che l'altra. I lavori di Frank e le considerazioni di I. Prigogine sembrano indicare che questo è ciò che è realmente accaduto (v. Walker, 1979).
G. Wald ha raccontato che una volta egli discusse questo argomento con Einstein che gli rispose: ‟Vede, un tempo ero solito chiedermi perché mai l'elettrone avesse carica negativa. Negativo e positivo sono perfettamente simmetrici in fisica. Non c'è assolutamente nessuna ragione di preferire l'uno piuttosto che l'altro. E allora, perché l'elettrone è negativo? Ci ho pensato a lungo, e alla fine l'unica conclusione che ho potuto trarre è questa: è risultato vincente". Lo stesso deve essere successo nel caso delle molecole destrogire o levogire. Se fossimo in grado di analizzare la vita nell'intero Universo, dovremmo trovare un ugual numero di amminoacidi destrogiri e di amminoacidi levogiri.
10. La ricerca di intelligenze extraterrestri.
I dati ottenuti dallo studio delle meteoriti e delle molecole interstellari e dagli esperimenti effettuati in laboratorio indicano chiaramente che l'evoluzione biochimica deve avere natura cosmica. Se le cose stanno effettivamente cosi, potrebbe esistere un altro luogo nell'Universo nel quale i processi evolutivi abbiano raggiunto un punto simile a quello riscontrabile sulla Terra. Viene naturale chiedersi se siamo soli nell'Universo (v. Ponnamperuma e Cameron, 1974). L'unica maniera di ottenere risposta a questa domanda sembra possa venire dall'ascolto di eventuali segnali provenienti dallo spazio. F. Drake ha cominciato una ricerca di questo tipo già nel 1959, e benché i primi esperimenti non abbiano dato risultati certi, hanno suscitato un notevole interesse. Un piano, per ora sulla carta, chiamato ‛progetto Ciclope', prevede l'installazione di antenne di circa 10 m di diametro nella parte sud-occidentale degli Stati Uniti, orientate verso l'altra faccia della Luna per poter ascoltare in modo continuativo i segnali provenienti dallo spazio. Alla fine si potrebbe riuscire a sentire un segnale; una singola parola avrebbe probabilmente maggior valore di tutti gli esperimenti fatti in laboratorio. Attualmente sono in corso svariate ricerche, ma per venire a capo di qualcosa è necessario uno sforzo coordinato da parte di tutte le nazioni. I risultati di una ricerca di questo tipo potrebbero essere grandiosi. La risposta alla domanda se siamo soli nell'Universo è oggi a portata di mano.
11. Conclusioni.
Gli studi intrapresi in laboratorio col ciclotrone di Berkeley o gli esperimenti con le scariche elettriche di Urey e Miller ci hanno portato attraverso il sistema solare nello spazio. Nutriamo buone speranze che lo schema dell'evoluzione biochimica possa essere delineato in laboratorio. Le conoscenze biochimiche che si sono accumulate nel corso degli ultimi anni ci hanno permesso di capire alcuni dei più segreti processi della natura. Grazie a queste conoscenze, il tempo necessario per risolvere il nostro problema può non essere molto lungo. Non possiamo negare l'immensa importanza di questa prospettiva per qualsiasi posizione filosofica, né sottrarci a questa ricerca per la difficoltà del compito.
Bibliografia.
Abelson, P. H., Amino acids formed in primitive atmospheres, in ‟Science", 1956, CXXIV, p. 935.
Abelson, P. H., Effects of ultraviolet light on the primitive environment, in Carnegie Institution of Washington yearbook, Washington 1956, pp. 156-157.
Abelson, P. H., Chemical events on the primitive earth, in ‟Proceedings of the National Academy of Sciences", 1966, LV, pp. 1365-1372.
Akabori, S., Asymmetric synthesis of amino acids and formation of fore-proteins, in ‟Kagaku", 1955, XXV, pp. 54-58.
Allen, W. V., Ponnamperuma, C., A possible prebiotic synthesis of monocarboxylic acids, in ‟Currents in modern biology", 1967, I, pp. 24-28.
Bahadur, K., Photosynthesis of amino acids from paraformaldehyde and potassium nitrate, in ‟Nature", 1954, CLXXIII, p. 1141.
Barghoorn, E. S., Tyler, S. A., Microfossils form the Middle Pre-Cambrian of Canada, in First international conference on palynology, Tucson 1962.
Berkner, L.V., Marshall, L.C., The history of the growth of oxygen in the earth's atmosphere, in The origin and evolution of atmospheres and oceans (a cura di P. J. Brancazio e A. G. W. Cameron), New York 1964, pp. 86-101.
Bernal, J. D., The physical basis of life, London 1951 (tr. it. in: L'origine della vita sulla Terra, a cura di A. I. Opearin, Torino 19612).
Biemann, K., Oro, J., Toulmin, P. III, Orgel, L.E., Nier, A. O., Anderson, D. M., Simmonds, P. G., Flory, D., Diaz, A. V., Rushneck, D. R., Biller, J. E., Lafleur, A. L., The search of organic substances and inorganic volatile compounds in the surface of Mars, in ‟Journal of geophysical research", 1977, LXXXII, 28, pp. 4641-4658.
Bonner, W. A., Origins of molecular chirality, in Exobiology (a cura di C. Ponnamperuma), Amsterdam 1972, pp. 170-234.
Butlerow, A., Bildung einer zuckerartigen Substanz durch Synthese, in ‟Justus Liebigs Annalen der Chemie", 1861, CXX, pp. 295-298.
Calvin, M., Chemical evolution, Oxford 1969.
Choughuley, A. S., Lemmon, R. M., Production of cysteic acid, taurine and cystamine under primitive earth conditions, in ‟Nature", 1966, CCX, pp. 628-629.
Conant, J. B., Pasteur and Tyndall's study of spontaneous generation, Cambridge, Mass., 1959.
Darwin, C., Life and letters; including an autobiographical chapter (a cura di F. Darwin), 2 voll., New York 1959.
Darwin, E., The temple of nature, London 1803.
Dodonova, N. Y., Sidorova, A. L., Photochemical synthesis of amino acids from a mixture of simple gases under the influence of vacuum ultraviolet radiation, in ‟Biofizika", 1961, VI, pp. 14-20.
Ellenbogen, E., Abiogenic photosynthesis of polypeptides, in 134th National meeting of the American Chemical Society, Chicago 1958, p. 47c.
Flores, J. J., Ponnamperuma, C., Polymerization of amino acids under primitive earth conditions, in ‟Journal of molecular evolution", 1972, II, pp. 1-9.
Fox, S. W., Dose, K., Molecular evolution and the origin of life, San Francisco 1972.
Fox, S. W., Harada, K., Synthesis of uracil under conditions of a thermal model of prebiological chemistry, in ‟Science", 1961, CXXXIII, pp. 1923-1924.
Gabel, N. W., Ponnamperuma, C., Model for the origin of monosaccharides, in ‟Nature", 1967, CCXVI, pp. 453-455.
Gaffron, H., On dating stages in photochemical evolution, in Horizons in biochemistry (a cura di M. Kasha e B. Pullman), New York 1962, pp. 59-89.
Gammon, R. H., Chemistry of interstellar space, in ‟Chemical and engineering news", 1978, LVI, 40, pp. 21-23.
Grossenbacher, K. A., Knight, S. C., Amino acids, peptides and spherules obtained from primitive earth gases in a sparking system, in The origins of prebiological systems and of their molecular matrices (a cura di S. W. Fox), New York 1965, pp. 173-186.
Groth, W., Weysenhoff, H. von, Photochemical formation of organic compounds from mixtures of simple gases, in ‟Planetary science", 1960, II, pp. 79-85.
Gupta, S., Ochiai, E., Ponnamperuma, C., Organic synthesis in the atmosphere of Titan, in ‟Nature", 1981, CCXCIII, pp. 725-727.
Haldane, J. B. S., The origin of life, in ‟Rationalist annual", 1928, CXLVIII, pp. 3-10.
Harada, K., Fox, S. W., The thermal synthesis of natural amino acids from a postulated primitive terrestrial atmosphere, in ‟Nature", 1964, CCI, pp. 335-336.
Heyns, K., Walter, W., Meyer, E., Modelluntersuchungen zur Bildung organischer Verbindungen in Atmosphären einfacher Gase durch elektrische Entladungen, in ‟Naturwissenschaften", 1957, XLIV, pp. 385-389.
Hochstim, A. R., Hypersonic chemosynthesis and possible formation of organic compounds from impact of meteorites on water, in ‟Proceedings of the National Academy of Sciences", 1968, LIX, pp. 22-28.
Holland, H. D., Model for the evolution of the earth's atmosphere. Petrological studies. A volume to honor A. F. Buddington, Boulder 1962, pp. 447-477.
Huntress, W. T. Jr., Baldeschwieler, J. D., Ponnamperuma, C., Ion-molecule reactions in hydrogen cyanide, in ‟Nature", 1969, CCXXIII, pp. 468-471.
Huxley, T. H., On the physical basis of life, in Collected essays, vol. I, London 1868.
Klein, H. P., The Viking biological investigation: general aspects, in ‟Journal of geophysical research", 1977, LXXXII, 28, pp. 4677-4680.
Kliss, R. M., Matthews, C. N., HCN dimer and chemical evolution, in ‟Proceedings of the National Academy of Sciences", 1962, XLVIII, pp. 1300-1306.
Knoll, A. H., Barghoorn, E. S., Archean microfossils showing cell division from the Swaziland system of South Africa, in ‟Science", 1977, CXCVIII, pp. 396-398.
Kotra, R. K., Shimoyama, A., Ponnamperuma, C., Hare, P. E., Amino acids in a carbonaceous chondrite from Antarctica, in ‟Journal of molecular evolution", 1979, XIII, pp. 179-183.
Kvenvolden, K. A., Lawless, J. G., Ponnamperuma, C., Nonprotein amino acids in the Murchison meteorite, in ‟Proceedings of the National Academy of Sciences", 1971, LXVIII, pp. 486-490.
Lasaga, A. C., Holland, H. D., Dwyer, M. J., Primordial oil slick, in ‟Science", 1971, CLXXIV, pp. 53-55.
Lemmon, R. M., Chemical evolution, in ‟Chemical reviews", 1970, LXX, pp. 95-109.
Lowe, C. U., Rees, M. W., NH4CN and primitive earth synthesis, in ‟Federation proceedings", 1963, XXII, p. 479.
Matthews, C. N., Moser, R. E., Prebiological protein synthesis, in ‟Proceedings of the National Academy of Sciences", 1966, LVI, pp. 1087-1094.
Miller, S. L., The mechanism of synthesis of amino acids by electric discharges, in ‟Biochimica et biophysica acta", 1957, XXIII, pp. 480-489.
Miller, S. L., Liquid water and the origin of life, in Isotopic and cosmic chemistry (a cura di H. Craig, S. L. Miller e G. J. Wasserburg), Amsterdam 1964, pp. 103-113.
Miller, S. L., Urey, H. C., Organic compound synthesis on the primitive earth, in ‟Science", 1959, CXXX, pp. 245-251.
Oparin, A. I., Proischûodenie ûizni, Moskva 1924 (tr. ingl.: Origin of life, New York 1938).
Oparin, A. I., Vozniknovenie ûizni na zemle, Moskva 19412 (tr. it.: L'origine della vita sulla Terra, Torino 1956, 19612).
Oro, J., Studies in experimental organic cosmochemistry, in ‟Annals of the New York Academy of Sciences", 1963, CVIII, pp. 464-481.
Oro, J., Synthesis of organic compounds by electric discharges, in ‟Nature", 1963, CXCVII, pp. 862-867.
Oro, J., Guidry, C. L., Direct synthesis of polypeptides. I. Polycondensation of glycine in aqueous ammonia, in ‟Archivs of biochemistry and biophysics", 1961, XCIII, pp. 166-177.
Oro, J., Kaurat, S. S., Amino acid synthesis from hydrogen cyanide under possible primitive earth conditions, in ‟Nature", 1961, CXC, pp. 442-443.
Orpen, J. L., Wilson, J. F., Stromatolites at ~3.500 Myr and greenstone-granite unconformity in the Zimbabwean Archean, in ‟Nature", 1981, CCXCI, pp. 218-220.
Partington, J. R., A short history of chemistry, London 1957.
Pavlovskaia, T. E., Pasynskii, A. G., The original formation of amino acids under the action of ultraviolet rays and electric discharges, in Aspects of the origin of life on earth (a cura di M. Florkin), New York 1960, pp. 98-104.
Pering, K. L., Ponnamperuma, C., Aromtic hydrocarbons in the Murchison meteorite, in ‟Science", 1971, CLXXIII, pp. 237-239.
Ponnamperuma, C., Abiological synthesis of some nucleic acid constituents, in The origins of prebiological systems and of their molecular matrices (a cura di S. W. Fox), New York 1965, pp. 221-242.
Ponnamperuma, C., Role of radiation in primordial organic synthesis, in Radiation research. Proceedings of the 3rd international congress ( cura di G. Silini), Amsterdam 1967, pp. 700-713.
Ponnamperuma, C., A la recherche de la vie sur la Lune, in ‟Atomes", 1970, XXV, 274, pp. 169-174.
Ponnamperuma, C., Cosmochemistry and the origin of life, in Proceedings of the Robert A. Welch Foundation conferences on chemical research, vol. XXI, Cosmochemistry (a cura di W. O. Milligan), Houston 1978, pp. 137-197.
Ponnamperuma, C., Cameron, A. G. W., Interstellar communication: scientific perspectives, Boston 1972.
Ponnamperuma, C., Cameron, A. G. W. (a cura di), Interstellar communication: scientific perspectives, Boston 1974.
Ponnamperuma, C., Klein, H. P., The coming search for life on Mars, in ‟Quarterly review of biology", 1970, XLV, pp. 235-258.
Ponnamperuma, C., Lemmon, R. M., Mariner, R., Calvin, M., Formation of adenine by electron irradiation of methane, ammonia and water, in ‟Proceedings of the National Academy of Sciences", 1963, XLIX, pp. 737-740.
Ponnamperuma, C., Mack, R., Nucleotide synthesis under possible primitive earth conditions, in ‟Science", 1965, CXLVIII, pp. 1221-1223.
Ponnamperuma, C., Pering, K. L., Possible abiogenic origin of some naturally occurring hydrocarbons, in ‟Nature", 1966, CCIX, pp. 979-982.
Ponnamperuma, C., Sagan, C., Mariner, R., Synthesis of adenosine triphosphate under possible primitive earth conditions, in ‟Nature", 1963, CIC, pp. 222-226.
Ponnamperuma, C., Shimoyama, A., Yamada, M., Hobo, T., Pal, R., Possible surface reactions on Mars: implications for Viking biology results, in ‟Science", 1977, CXCVII, 4302, pp. 455-457.
Ponnamperuma, C., Woeller, F., Differences in the character of C6 to C9 hydrocarbons from gaseous methane in low frequency electric discharges, in ‟Nature", 1964, CCIII, pp. 272-274.
Ponnamperuma, C., Woeller, F., Alpha-aminonitriles formed by an electric discharge through a mixture of anhydrous methane and ammonia, in ‟Currents in modern biology", 1967, I, pp. 156-158.
Ponnamperuma, C., Woeller, F., Organic synthesis in a simulated Jovian atmosphere, in ‟Icarus", 1969, X, pp. 386-392.
Pullman, B., Pullman, A., Electronic delocalization and biochemical evolution, in ‟Nature", 1962, CXCVI, pp. 1137-1142.
Rabinowitz, J., Note on the role of cyanides and polyphosphates in the formation of peptides in aqueous solutions of amino acids at room temperature as a possible prebiotic process, in ‟Helvetica chimica acta", 1971, LIV, 5, pp. 1483-1485.
Reid, C., The relation between primitive and present day photobiological process, in Aspects of the origin of life (a cura di M. Florkin), New York 1960, pp. 189-195.
Rubey, W. W., Development of the hydrosphere and atmosphere with special reference to the probable composition of the early atmosphere, in Crust of the earth (a cura di A. Poldervaart), New York 1955, pp. 631-650.
Sagan, C., Radiation and the origin of the gene, in ‟Evolution", 1957, XI, pp. 40-55.
Sanchez, R. A., Ferris, J. P., Orgel, L.E., Cyanoacetylene in prebiotic synthesis, in ‟Science", 1966, CLIV, pp. 784-785.
Sanchez, R. A., Ferris, J. P., Orgel, L.E., Studies in prebiotic synthesis. IV. Conversion of 4-aminoimidazole-5-carbonitrile derivatives to purines, in ‟Journal of molecular evolution", 1968, XXXVIII, pp. 121-128.
Schimpl, A., Lemmon, R. M., Calvin, M., Cyanamide formation under primitive earth conditions, in ‟Science", 1965, CXLVII, pp. 149-150.
Schopf, J. W., Precambrian paleobiology, in Exobiology (a cura di C. Ponnamperuma), Amsterdam 1972, pp. 16-61.
Schramm, G., Synthesis of nucleotides and polynucleotides with metaphosphate esters, in The origin of prebiological systems and of their molecular matrices (a cura di S. W. Fox), New York 1965, pp. 299-315.
Shah, D. O., The origin of membranes and related surface phenomena, in Exobiology (a cura di C. Ponnamperuma), Amsterdam 1972, pp. 235-265.
Shapley, H., Of stars and men, Boston 1958.
Steinman, G. D., Sequence generation in prebiological peptide synthesis, in ‟Archives of biochemistry and biophysics", 1967, CXXI, pp. 533-539.
Szutka, A., Probable synthesis of porphine-like substances during chemical evolution, in The origins of prebiological systems and of their molecular matrices (a cura di S. W. Fox), New York 1965, pp. 243-254.
Terenin, A. N., Photosynthesis in the shortes ultraviolet, in The origin of life on earth (a cura di A. I. Oparin), Oxford 1959, pp. 136-139.
Tilton, G. R., Steiger, R. H., Lead isotopes and the age of the earth, in ‟Science", 1965, CL, pp. 1805-1808.
Tyndall, J., Fragments of science for unscientific people, London 1871.
Urey, H. C., The planets, their origin and development, New Haven 1952.
Van der Velden, W., Shwartz, A. W., Search for purines and pyrimidines in the Murchison meteorite, in ‟Geochimica et cosmochimica acta", 1977, XLI, 7, pp. 961-968.
Walker, D. C., Origins of optical activity in nature, Amsterdam 1979.
Walter, M. R., Buick, R., Dunlop, J. S. R., Stromatolites 3,400-3,500 Myr old from the North Pole area, Western Australia, in ‟Nature", 1980, CCLXXXIV, pp. 443-445.