
Ormoni
Ormoni
Ormoni negli Invertebrati, di Vincent B. Wigglesworth
Ormoni nei Vertebrati, di Gerald Litwack
Ormoni locali, di Heinz Otto Schild
Ormoni vegetali, di Erasmo Marrè
Ormoni negli Invertebrati
SOMMARIO: 1. Introduzione. □ 2. Gli ormoni negli Insetti: a) ormoni e muta; b) chimica dell'ormone della muta; c) ormoni e metamorfosi; d) chimica dell'ormone giovanile; e) ormoni e poli- morfismo; f) ormoni e riproduzione; g) diapausa riproduttiva; h) diapausa nell'uovo; i) altri effetti ormonali negli Insetti; l) neuroumori; m) feromoni. □ 3. Gli ormoni nei Crostacei. □ 4. Gli ormoni in altri Artropodi. □ 5. Gli ormoni nei Molluschi. □ 6. Gli ormoni negli Anellidi. □ 7. Gli ormoni nei Celenterati e negli Echinodermi. □ 8. Conclusione. □ Bibliografia.
1. Introduzione.
Gli ormoni sono ‛messaggeri chimici' che costituiscono una delle principali modalità di comunicazione tra una parte e l'altra del corpo, accanto al maggior sistema di comunicazione, quello nervoso. Questi due sistemi si integrano, sovrapponendosi tra loro. Il sistema nervoso opera attraverso la rapida trasmissione di perturbazioni elettriche, o ‛stimoli nervosi', lungo le fibre nervose. Ma il passaggio di tali stimoli da un neurone all'altro, attraverso le giunture interneuronali, o ‛sinapsi', avviene comunemente mediante un processo chimico, ovvero mediante la liberazione, all'estremità della fibra nervosa, di un ‛neuroumore', o ‛neurotrasmettitore', che si diffonde attraverso gli interstizi sinaptici, per stimolare il neurone adiacente e stabilire, così, un nuovo impulso nervoso. Allo stesso modo, dall'estremità di un nervo che termina in un muscolo o in una ghiandola può liberarsi un ‛neuroumore', ed è proprio questo a stimolare la cellula ghiandolare o la fibra muscolare. I neuroumori hanno per lo più un effetto puramente locale, ma talvolta possono essere immessi in circolo, esplicando allora un'azione più complessa.
Nervi e ormoni si integrano anche per altra via: tutte le cellule nervose elaborano ininterrottamente una sostanza, che defluisce lentamente lungo la fibra nervosa, o assone; esistono tuttavia alcune particolari cellule nervose, i cui assoni possono trasmettere stimoli elettrici come quelli di qualsiasi altra cellula nervosa, che sono caratterizzate dalla produzione di minuscoli granuli sferici, di natura proteica o polipeptidica, di diametro compreso tra i .000 e 3.000 Å, comunemente, ma non sempre, opachi agli elettroni del microscopio elettronico e dotati di proprietà specifiche di colorazione (v. fig. 1). Questi granuli scorrono lungo gli assoni e il loro contenuto viene scaricato alle estremità dei filamenti nervosi: le cellule nervose che li elaborano vengono dette ‛neurosecretrici'.
Il prodotto delle cellule neurosecretrici può avere un effetto farmacologico locale sulle cellule in prossimità delle quali viene rilasciato: per esempio, in alcuni invertebrati, può raggiungere il muscolo cardiaco, stimolandone il battito; oppure, tale secrezione può essere immessa nella circolazione sanguigna e, attraverso questa, essere distribuita a tutti gli organi e i tessuti. Spesso i corpi delle cellule neurosecretrici risiedono nei gangli del sistema nervoso centrale; i loro assoni possono terminare in estremità bulbose, raggruppate nei cosiddetti ‛organi neuroemali', che fungono da sistemi di regolazione per l'immissione della neurosecrezione proveniente dagli assoni nella circolazione sanguigna.
La sostanza attiva prodotta dalle cellule neurosecretrici, per solito di natura polipeptidica, può esercitare la propria azione direttamente sui tessuti, oppure può agire su particolari ‛ghiandole endocrine', o organi a secrezione interna, inibendo o stimolando la loro attività secretrice. Tali ghiandole cominciano a svilupparsi precocemente, per lo più in prossimità dell'apparato boccale, per un processo di gemmazione dell'ectoderma; successivamente possono spostarsi all'interno del corpo, dando luogo a organi ben definiti, lontani dalla loro zona di origine.
2. Gli ormoni negli Insetti.
Conviene prendere in esame, innanzitutto, gli ormoni degli Insetti, giacché sono stati oggetto di studi più dettagliati. Questi ormoni intervengono nel controllo della crescita e della muta, nella regolazione della metamorfosi e nella riproduzione; influiscono su alcuni cambiamenti metabolici, sulla diuresi e sull'arresto di diuresi, sulla trascolorazione e, infine, sulla frequenza del battito cardiaco. I neuroumori sono probabilmente implicati nella trasmissione nervosa, secondo le linee che ne abbiamo tracciato.
a) Ormoni e muta.
In tutti i gangli del sistema nervoso s'incontrano cellule neurosecretrici; nella parte dorsale del cervello (pars intercerebralis) si trovano quelle che partecipano al processo di crescita, provviste di assoni che si estendono fino al corpus cardiacum, tipico organo neuroemale posto proprio sotto il cervello, dove l'ormone (probabilmente un polipeptide) viene liberato nel sangue. Sembra che, in genere, l'ormone del cervello (detto anche ‛ormone di attivazione' ovvero ‛ormone protoracotropo') non agisca direttamente sui tessuti; esso innanzitutto stimolerebbe una ghiandola endocrina, situata nella testa o nel torace, a secernere l'ormone vero e proprio, responsabile dell'inizio della crescita e della muta. Negli insetti a struttura più primitiva, questa ghiandola, detta ‛ghiandola ventrale', si trova nella testa; invece in quelli a struttura più evoluta essa si è spostata in zone più interne del corpo, tanto da essere denominata ‛ghiandola protoracica'. Sotto l'azione dell'ormone cerebrale, le cellule della ghiandola protoracica si sviluppano enormemente e incominciano a secernere l'ormone della muta' (v. figg. 2 e 3).
L'ormone della muta agisce su tutti i tessuti del corpo e in particolare sull'epidermide, che ha la maggiore responsabilità nei processi di crescita e di muta. Infatti, stimolate da questo ormone, le cellule epidermiche sintetizzano acidi nucleici e proteine, e, aumentando di dimensioni, si staccano dalla vecchia cuticola, si dividono e si moltiplicano, disponendosi in maniera opportuna a formare adeguatamente nuova cuticola. Gli strati più interni della vecchia cuticola vengono dissolti dagli enzimi prodotti dalle cellule epidermiche e, infine, quel che resta della vecchia cuticola cade. Contemporaneamente, come è ovvio, analoghi processi di crescita si verificano a carico dei muscoli e degli altri organi interni: l'ormone della muta è indispensabile per sostenere l'intero fenomeno della crescita.
Alcuni insetti, come le cimici ematofaghe, consumano un solo pasto per ogni fase di muta, e dopo la muta non mostrano ulteriore crescita, fino al pasto successivo. E facile dimostrare il ruolo svolto dagli ormoni nella crescita di simili insetti: valga l'esempio di Rhodnius, grossa cimice ematofaga, la cui dilatazione addominale, provocata da una cospicua ingestione di sangue, fornisce un impulso nervoso al cervello, stimolandolo a secernere l'ormone di attivazione. Qualora, subito dopo il pasto, l'insetto venga decapitato, la sua crescita si arresta all'istante, pur restando esso in vita ancora per mesi; se però la regione del cervello dove si trovano le cellule neurosecretrici attivate viene trapiantata nell'addome dell'insetto decapitato, la ghiandola protoracica viene riattivata e si riavvieranno anche i processi di crescita e di muta (v. fig. 4). D'altra parte, se l'addome viene separato dal torace, il trapianto di cellule neurosecretrici non sarà sufficiente a ripristinare la crescita, mentre quello della ghiandola protoracica attivata provocherà la muta persino nell'addome isolato. È chiaro quindi che è l'ormone della ghiandola protoracica ad agire direttamente sull'epidermide.
Tali osservazioni sono di notevole importanza per la conoscenza dell'arresto naturale della crescita, o ‛diapausa', in cui incorrono parecchi insetti, durante le avverse stagioni, a causa del freddo invernale o del caldo e della siccità estivi. Quando la diapausa si verifica durante le fasi postembrionali degli Insetti, come nella larva matura o nella crisalide, è conseguente a un arresto della secrezione di ormoni talvolta da parte della ghiandola protoracica, ma, più spesso e primitivamente, da parte del cervello. Per esempio, nella crisalide ibernante del baco da seta gigante Hyalophora cecropia, le cellule neurosecretrici del cervello diventano inattive, ma l'attività riprende se l'esposizione a bassa temperatura viene prolungata: allora, il trapianto di un cervello cosi riattivato darà luogo, in una crisalide in diapausa, a una ripresa della crescita.
b) Chimica dell'ormone della muta.
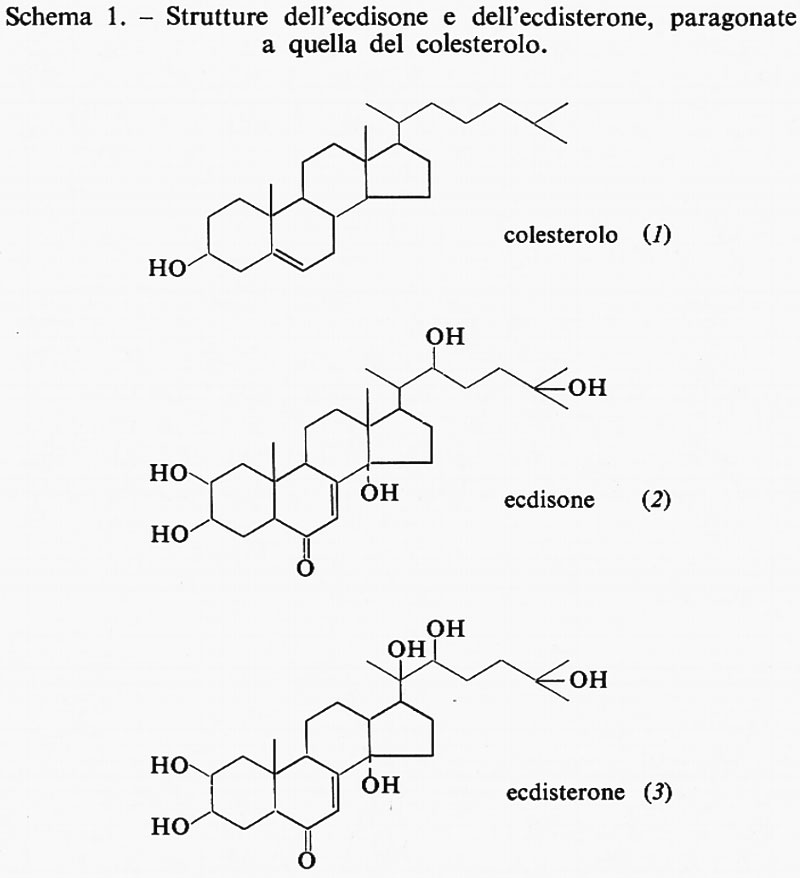
L'ormone della muta è stato isolato da numerosi insetti, ed è stato dimostrato che si tratta di uno steroide, chimicamente correlato con il colesterolo (1), ma molto più solubile nell'acqua della maggior parte degli steroidi, in quanto contiene un maggior numero di gruppi idrossilici. In effetti esistono due forme dell'ormone suddetto: la prima a essere riconosciuta venne detta ‛ecdisone' (con la formula di struttura 2), la seconda, che contiene un gruppo idrossilico in più, fu indicata con il termine ‛ecdisterone' (3). Tali sostanze sono state sintetizzate e, iniettate nell'insetto, riproducono tutti gli effetti dell'ormone naturale, come l'indurimento cutaneo nelle larve della mosca per la formazione della crisalide, o come i cambiamenti di colore, che precedono, in alcuni bruchi, la formazione della crisalide, o come infine l'intero fenomeno della muta: una dose di circa 10 μg per grammo è necessaria per provocare una muta completa nella maggior parte degli insetti. Anche negli insetti adulti un iniezione di ecdisone può determinare un nuovo processo di muta.
c) Ormoni e metamorfosi.
È caratteristica degli Insetti subire un notevole cambiamento di forma durante la muta finale, nel diventare adulti: generalmente spuntano le ali, si formano complessi organi genitali esterni, e l'intero aspetto può esserne radicalmente modificato. E noto infatti che si riscontra una ‛metamorfosi completa' negli insetti di struttura più evoluta, come i Lepidotteri (farfalle e falene), i Coleotteri (scarabei), gli Imenotteri (formiche, api, ecc.) e i Ditteri (mosche): durante lo stadio dell'accrescimento, essi sono larve a forma di bruco; durante quello della riproduzione, quando l'adulto è alato, assumono una forma completamente diversa, e spesso cambiano radicalmente condizioni di vita; infine fra i due stadi attraversano una forma intermedia, quella della ‛pupa'.
La natura ormonale del controllo della metamorfosi fu dimostrata per la prima volta nel corso di esperimenti sulla cimice ematofaga Rhodnius, quando venne osservato che, se la testa veniva rimossa dal corpo di giovani larve durante un periodo critico, alcuni giorni dopo il pasto di sangue, esse continuavano regolarmente a trasformarsi, secondo il loro processo di muta. Ma, una volta completato questo processo e rimossa la vecchia cute, si rilevò che le larve avevano subito una metamorfosi precoce, sviluppandosi in adulti di taglia minuscola: era chiaro che la testa della giovane larva produceva un ormone atto a impedire la metamorfosi.
Tale ormone, detto generalmente ‛ormone giovanile', o ‛neotenina', si rivelò prodotto dal corpus allatum, piccola ghiandola a secrezione interna, strettamente associata con il corpus cardiacum, posta proprio dietro il cervello. Il meccanismo di controllo esplicato da questo ormone è presente in tutti gli Insetti: l'asportazione della coppia di corpora allata, compiuta in giovani filugelli (v. fig. 5), provocò il loro sviluppo in piccole crisalidi, dalle quali uscirono minuscole farfalle (v. fig. 6) capaci di deporre uova; i corpora allata, asportati negli stadi giovanili di sviluppo e trapiantati nell'ultima fase larvale, provocarono nuovamente una muta in larva. In alcuni insetti, come in Tenebrio molitor, possono aversi molti stadi larvali supplementari, con conseguente nascita di adulti giganti. Negli insetti a metamorfosi completa, sembra che l'ormone giovanile determini la persistenza della forma larvale, se presente ad alto tenore, e la comparsa della forma crisalidica, se presente a tenore assai basso; mentre sembra che la sua completa assenza provochi lo sviluppo dei caratteri adulti.
L'ormone giovanile agisce localmente sulle cellule epidermiche: talvolta, quando viene trapiantata una ghiandola si forma soltanto una piccola superficie di cuticola larvale che ricopre la zona di impianto della ghiandola stessa: il resto dell'insetto è un normale adulto (v. fig. 7). Si suppone che l'ormone agisca, direttamente o indirettamente, sul sistema genetico cellulare, provocando le attività geni- che responsabili dei caratteri larvali e sopprimendo quelle responsabili dei caratteri adulti. Se un insetto adulto è indotto sperimentalmente alla muta, per mezzo di un'iniezione di ecdisone in presenza dell'ormone giovanile, alcune superfici del tegumento riassumeranno in parte i loro caratteri larvali; non è stato tuttavia possibile provocare questa reversione nell'insetto intero. Nello sviluppo normale dell'insetto, perché possa avvenire la metamorfosi, la secrezione dell'ormone giovanile s'interrompe al momento opportuno: l'epoca in cui avviene questa interruzione sembra essere determinata dal cervello, ma non si conoscono i particolari di tale processo.
Normalmente l'insetto, una volta diventato adulto, non affronterà una seconda muta, dato che le ghiandole protoraciche degenerano e scompaiono nel corso della metamorfosi. Ma se nell'ultima fase larvale di un emittero, o nella fase crisalidica di un lepidottero, si somministra sperimentalmente l'ormone giovanile, l'insetto conserva le caratteristiche larvali o crisalidiche quando subisce la muta, e la persistenza della ghiandola protoracica lo mette in grado di affrontare nuove trasformazioni.
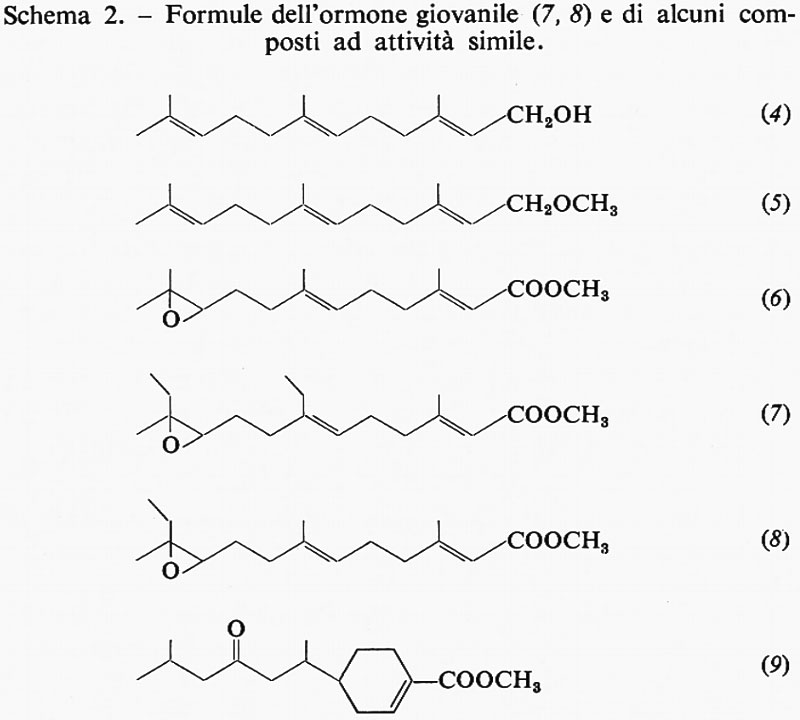
d) Chimica dell'ormone giovanile.
Sono riportate nello schema 2 le formule di alcuni composti con attività di ormone giovanile. Si tratta di derivati di un alcool terpenoide, il farnesolo (4). Il primo e più noto ormone giovanile (7) fu isolato dall'estratto lipidico della farfalla adulta del baco da seta gigante Hyalophora cecropia (che, per una qualche ragione, possiede tale sostanza in quantità considerevole). Può essere considerato come l'estere metilico dell'acido farnesenico, con un anello epossidico in posizione 10 e con due catene laterali etiliche in luogo dei gruppi metilici che si trovano nella maggior parte dei composti isoprenoidi. L'isomero naturale (il più attivo) ha la configurazione trans, trans ai due doppi legami, mentre il gruppo epossidico è in cis. Se questo sia l'ormone attivo per tutti gli Insetti, rimane ancora da accertare. E interessante il fatto che dalla farfalla cecropia sia stato isolato un secondo ormone, ma non altrettanto attivo, che differisce dal primo per avere una catena laterale metilica al posto di una etilica (8). È stato sperimentato che una vasta gamma di composti, quasi tutti chimicamente collegati con il farnesolo (4) - salvo alcuni, di struttura piuttosto diversa - provoca gli effetti dell'ormone giovanile. Per la verità, alcuni di questi composti sintetici sono molto più attivi dell'ormone isolato dalla cecropia. Un altro derivato è l'estere metilico dell'acido todomatuico (9), fattore presente nella carta fatta con polpa di abete del balsamo del Canada (Abies balsamea), che è specificamente attivo negli Emitteri della famiglia dei Pirrocoridi.
e) Ormoni e polimorfismo.
Alcuni insetti si presentano morfologicamente sotto forme diverse, ciascuna con la propria costituzione genetica. In altri insetti, invece, la forma varia anche tra individui che abbiano la stessa struttura genetica; queste variazioni sono prodotte ugualmente dall'azione di geni. Tale operazione ‛scatta', ovvero geni latenti sono indotti ad agire, per l'improvviso insorgere di un qualche cambiamento ambientale. Queste trasformazioni sono paragonabili a quelle della metamorfosi, giacché, pur restando invariata la costituzione genetica, vengono messe in funzione nuove componenti del sistema. Citiamo alcuni esempi di questo tipo di polimorfismo: 1) l'adattamento del tono di colore dei bruchi al fondo sul quale sono allevati; 2) il cambiamento delle locuste dalla forma migratoria di gregario a quella non migratoria di individuo solitario; 3) le forme stagionali delle farfalle: in Araschnia levana una diapausa crisalidica è provocata dall'esposizione delle larve al sole, durante le giornate corte, come si deduce dalla loro varietà primaverile (forma levana), mentre le crisalidi esposte al sole durante le giornate lunghe, non avendo diapausa, danno vita alla varietà estiva molto diversa (forma prorsa) (v. flg. 8); 4) le alterazioni, per cui alcune afidi femmine partenogenetiche, sotto l'influsso di giornate di diversa lunghezza, danno vita o a una discendenza che diventa vivipara, o a femmine partenogenetiche, o a forme bisessuate (v. figg. 9 e 10); 5) le diverse caste (forme sessuate, operai, guerrieri, ecc.) nelle formiche e in altri insetti a organizzazione sociale. La natura precisa del controllo di tutte queste trasformazioni non è chiaramente identificata, ma è evidente che vi sono implicati gli ormoni circolanti e, in alcuni casi, sembra che ne sia responsabile la quantità di ormone giovanile secreto.
L'esempio più comune di polimorfismo è l'esistenza di forme maschili e di forme femminili: la distinzione tra gli individui dei due sessi è il risultato di fattori genetici, negli Insetti come nei Vertebrati. In molti vertebrati, tuttavia, l'effetto dell'apparato genetico può essere neutralizzato dall'azione degli ormoni sessuali. Sembra che questo non accada nella maggior parte degli insetti, ma vi è un esempio che proverebbe il contrario, quello della lucciola Lampyris, nella quale il sesso è chiaramente sotto il controllo ormonale, in quanto il testicolo della larva maschile secerne un ‛ormone androgeno', capace di trasformare una femmina in via di sviluppo in un maschio con funzioni normali.
f) Ormoni e riproduzione.
Quando la cimice Rhodnius diventa adulta, il corpus allatum dà segni di rinnovata attività, e la decapitazione, così come impedisce la muta negli individui in fase giovanile, impedisce la formazione di uova mature in quelli adulti. Ecco poi quanto avviene in seguito all'asportazione del corpus allatum: il trapianto dei corpora allata nell'addome della femmina decapitata ristabilisce lo sviluppo delle uova. Il corpus allatum è necessario al maschio non per la produzione dello sperma, ma per la piena attività delle ghiandole accessorie, che forniscono lo ‛spermatoforo', cioè la capsula in cui sono chiusi gli spermatozoi prima di essere trasmessi alla femmina: l'ormone implicato in questa operazione è lo stesso ormone giovanile, anzi un iniezione di ormone giovanile puro nella femmina decapitata ristabilirà lo sviluppo dell'uovo.
Naturalmente, per la produzione dell'uovo è necessaria una nutrizione adeguata, ma talvolta il primo effetto della nutrizione sembra essere quello di ristabilire la secrezione dell'ormone giovanile da parte del corpus allatum. Nella cimice della cicerbita Oncopeltus e nello scarafaggio Leucophaea, se un insetto denutrito accoglie per trapianto il corpus allatum prelevato da un insetto ben nutrito, può essere indotto a deporre un piccolo numero di uova mature. L'ormone giovanile sembra determinare essenzialmente la deposizione del vitello all'interno dell'uovo. Femmine di Rhodnius o della locusta Schistocerca, in stato di inanizione oppure decapitate, daranno vita a una serie di cellule uovo capaci di svilupparsi normalmente solo fino al momento in cui dovrebbe cominciare la normale formazione del vitello; in quel momento, a meno che non venga loro fornito ormone giovanile, le cellule uovo muoiono e vengono riassorbite.
I Tisanuri, insetti dalla struttura più primitiva, che comprendono il pesciolino d'argento Lepisma e Thermobia domestica, hanno la caratteristica singolare di cominciare la muta e la riproduzione prima di essere completamente sviluppati. Essi serbano per tutta la vita le ghiandole ventrali che producono gli ormoni della muta e i loro cicli di muta si alternano con quelli di riproduzione. I corpora allata svolgono un ruolo modesto per ciò che riguarda la forma del corpo, poiché non esiste metamorfosi, ma la loro secrezione è necessaria alla formazione del vitello dell'uovo durante i cicli riproduttivi.
g) Diapausa riproduttiva.
Esattamente come l'arresto della secrezione nelle cellule neurosecretrici o nelle ghiandole protoraciche fa cessare la crescita durante le stagioni inclementi dell'anno, così quello della secrezione dell'ormone giovanile del corpus allatum può essere causa immediata della diapausa riproduttiva nell'adulto. Ciò si può osservare agevolmente nel parassita della patata Leptinotarsa, nel quale tale arresto ha effetti di vasta portata: questo insetto, dopo aver scavato nel suolo, vi si stabilisce; i muscoli delle ali degenerano, rendendolo incapace di volare; cessa nelle sue uova la formazione di vitello e gli stessi ovai entrano in uno stato d'inerzia. Un trapianto di corpora allata attivi inverte tutte queste modificazioni e ripristina rapidamente la riproduzione.
La riproduzione tuttavia non dipende esclusivamente dai corpora allata, la cui importanza, in molti insetti, sembra essere superata da quella delle cellule neurosecretrici. È troppo presto per affermarlo con sicurezza, eppure si direbbe che in molti insetti le cellule neurosecretrici cerebrali siano indispensabili per la riproduzione, giacché forniscono, influenzando la sintesi proteica (il che spesso significa sintesi di enzimi in generale) quelle materie prime essenziali, il cui passaggio agli ovai è regolato dall'ormone giovanile. Per concludere, in alcuni insetti (come Rhodnius) il corpus allatum sembra avere il controllo quasi esclusivo della riproduzione, mentre in altri, come i Ditteri (Calliphora, ecc.), le cellule neurosecretrici sembrano essere ancora più determinanti. Alle stesse conclusioni si perviene per quanto riguarda la diapausa: in Leptinotarsa il corpus allatum svolge il ruolo più importante, che spetta invece alla neurosecrezione nelle zanzare ibernanti.
h) Diapausa nell'uovo.
La diapausa può verificarsi anche nell'uovo, dopo che è stato deposto, e anche in questo caso possono essere implicati gli ormoni. L'esempio più noto è quello del baco da seta Bombyx mori, le cui uova, deposte in autunno, entrano in una stabile diapausa, che ba termine solo dopo una prolungata esposizione ai rigori invernali. Le uova nell'ovaio sono soggette all'azione di un ‛ormone della diapausa', secreto da due grosse cellule neurosecretrici, situate nel ganglio subesofageo (v. fig. 11), che ha la capacità di provocarne la diapausa. Nella farfalla è il cervello che stabilisce se l'ormone debba essere liberato (provocando la diapausa) o trattenuto nelle cellule neurosecretrici (permettendo così lo sviluppo delle uova deposte). L'azione del cervello viene determinata dall'esperienza fatta dalla madre in uno stadio precoce del suo sviluppo: se essa era stata esposta ad alta temperatura (25 °C) e a una giornata lunga - di 14 o più ore di luce - il suo cervello libererà l'ormone, onde essa deporrà uova in diapausa; se esposta a bassa temperatura (15 °C) e a una giornata corta, l'effetto sarà opposto.
i) Altri effetti ormonali negli Insetti.
Cellule neurosecretrici si trovano non soltanto nel cervello, ma anche nel corpo cardiaco, nel ganglio subesofageo e lungo tutti i gangli della catena ganglionare ventrale. In base al poco che si conosce a proposito di queste cellule, sembrerebbe che abbiano le funzioni più disparate, di cui tenteremo un elenco approssimativo. Alcuni insetti hanno delle cellule nel cervello (altri le hanno altrove, nei gangli), che producono un ormone capace di avviare un processo d'indurimento e inscurimento della cuticola dopo la muta. Alcune cellule del cervello controllano invece la trascolorazione negli insetti Ortotteri (Phasmida), provocando una migrazione, attraverso tutto il corpo, dei granuli di pigmento verso le cellule epidermiche. Le cellule del corpo cardiaco liberano ormoni implicati nel metabolismo dei carboidrati in grassi, mentre quelle neurosecretrici dei gangli toracici di Rhodnius producono un ormone diuretico che, liberato lungo il decorso dei nervi addominali, fa fluire rapidamente l'urina, dopo un pasto di sangue. In altri insetti, poi, anche l'equilibrio idrico è probabilmente sotto il controllo ormonale. Negli scarafaggi, gli ormoni del cervello o del ganglio subesofageo controllano il ritmo dell'attività locomotrice. Ma questa lista è ben lungi dall'essere completa, giacché in quasi tutti gli insetti esiste un sistema di cellule neurosecretrici, e di organi neuroemali associati, lungo l'intera catena ganglionare ventrale, la cui funzione è praticamente sconosciuta.
l) Neuroumori.
Dal sistema nervoso degli Insetti sono state estratte molte sostanze dotate di proprietà farmacologiche, ma si ignora quante di esse partecipino a normali processi fisiologici. É concordemente accettato che l'acetilcolina sia implicata nella trasmissione sinaptica del sistema nervoso centrale, sebbene non abbia alcun ruolo (come invece nel caso dei Mammiferi) nella trasmissione neuromuscolare, e sembra che non sia neppure immessa nel sangue circolante. Del pari, s'incontrano, negli Insetti, delle catecolammine (dopammina, adrenalina, ecc.), ma non si conosce il ruolo fisiologico che esse svolgono in questi animali. Il glutammato sembra sia la sostanza che regola la trasmissione dell'impulso tra nervo e muscolo e l'acido γ-ammino-butirrico (GABA) sembra funzionare da trasmettitore chimico durante l'inibizione nervosa, proprio come nei Vertebrati.
m) Feromoni.
Gli Insetti, come altri animali, comunicano con gli altri membri della loro specie per mezzo di sostanze odorose, le quali, per analogia tra questo tipo di comunicazioni e il sistema di comunicazione degli ormoni nell'interno di un corpo, furono denominate ‛ormoni sociali'. Per lo più tali sostanze chimiche agiscono soltanto sugli organi sensoriali, conducendo a modifiche di comportamento, come l'associazione o la dispersione di individui, l'emettere odori o seguire una scia odorosa, il senso di solidarietà nella specie, i segnali di avvertimento e di allarme, i richiami di accoppiamento (sia gli attraenti sessuali, prodotti dalla femmina, che gli eccitanti sessuali, prodotti dagli ‛androconi' del maschio), ecc. Un esempio ben noto di ormoni sociali è la ‛sostanza reale', o acido 9-idrossidecenoico, che l'ape regina emette dalle ghiandole mandibolari, sia per attirare il fuco durante l'accoppiamento, sia per rassicurare le api operaie della sua continua presenza tra loro. Le operaie, infatti, prelevano la secrezione, leccandola, dalla bocca della regina e se la trasmettono l'un l'altra durante gli scambi di cibo. Appena vedono scarseggiare la sostanza, si danno a costruire nuove celle per allevare più regine.
Il termine ‛feromone' è stato adottato per indicare tutte le sostanze chimiche che servono per comunicazioni nell'ambito della specie. Esse non possono considerarsi generalmente come ormoni. In un caso, però, i feromoni esercitano un ruolo più importante, in quanto controllano la formazione delle varie caste: nelle termiti. Per esempio, nella termite europea che si annida nel legno secco, Calotermes, la colonia consiste di una coppia di forme riproduttive primarie (ovvero la regina e il suo maschio), di una categoria di soldati e di una di larve (che fungono da operaie). Le forme riproduttive primarie secernono continuamente feromoni, atti a impedire lo sviluppo delle larve operaie in forme sessuate (‛forme riproduttive supplementari'), giacché la sostanza prodotta dal maschio inibisce la formazione di forme supplementari maschili e quella prodotta dalla femmina sopprime ogni possibilità di sviluppo per forme supplementari femminili. Probabilmente le termiti soldati esercitano azione analoga nell'impedire la comparsa di altri soldati. Si possono produrre sperimentalmente soldati somministrando a larve di operaie quantità elevate di ormone giovanile. Può essere dunque che i feromoni controllino tra l'altro la secrezione dell'ormone giovanile, ed esistono dati che provano come l'ormone giovanile sia effettivamente contenuto nei feromoni prodotti dalle forme riproduttive.
3. Gli ormoni nei Crostacei.
La classe di Invertebrati più studiata, dopo gli Insetti, è quella dei Crostacei, che possiedono un sistema neurosecretore ed endocrino in più punti simile al sistema degli Insetti. Come questi hanno ghiandole ventrali, o protoraciche, così i Crostacei hanno nel cefalotorace una coppia di ghiandole endocrine, gli ‛organi Y'. I gangli contengono: cellule neurosecretrici, molte delle quali scaricano la loro secrezione attraverso organi neuroemali (per es., attraverso la ‛ghiandola sinusale', situata nel peduncolo dell'occhio o, se l'occhio è sessile, nella testa); ‛organi X', anch'essi nel peduncolo dell'occhio; ‛organi postcommissurali' e ‛organi pericardici', nel cefalotorace. Vi è poi generalmente nel maschio dei Crostacei una ‛ghiandola androgena', associata ai condotti spermatici, paragonabile alla ghiandola testicolare nel maschio della lucciola (v. fig. 12). I neuroumori dei Crostacei sembrano dunque strettamente simili a quelli degli Insetti.
La muta nei Crostacei è continua per tutta la vita di un adulto, proprio come nei Tisanuri, insetti dalla struttura più primitiva. A provocarla è l'ormone ecdisterone, identico al ‛-ecdisone, o ecdisterone degli Insetti, il quale è prodotto dall'organo Y; ma la regolazione della muta nei Crostacei avviene per un cammino inverso rispetto alla muta negli Insetti. Mentre in questi ultimi l'ormone del cervello rende attiva la ghiandola della muta, nei Crostacei gli organi Y sembrano sempre pronti a secernere l'ormone della muta, ma sono tenuti sotto controllo da un ‛ormone inibitore', prodotto dal sistema neurosecretore del peduncolo oculare. Infatti, se vengono recisi i peduncoli oculari, la muta risulta accelerata.
Il processo della muta studiato nei Crostacei è quello dell'adulto, mentre la metamorfosi, che avviene in uno dei primissimi stadi dello sviluppo, non è mai stata oggetto di troppa attenzione. Non è stato scoperto nei Crostacei alcun ormone capace d'inibire lo svolgimento della metamorfosi, come l'ormone giovanile degli Insetti, e tuttavia un ormone che induce la metamorfosi potrebbe anche esistere, dato che nei granchi il peduncolo oculare sembra produrre un ormone capace di accelerare la metamorfosi della larva zoea.
Nei Crostacei, come nei Tisanuri tra gli Insetti, cicli riproduttivi si alternano con cicli di muta. Il loro controllo ormonale è complicato, in quanto la risposta del sistema varia durante tutto il ciclo; ma è chiaro che, durante la fase quiesciente della riproduzione, è il sistema organi X-ghiandola sinusale a produrre un ormone inibitore per la formazione del vitello: durante questo periodo, infatti, l'asportazione dei peduncoli oculari provoca immediatamente la vitellogenesi. Tale processo è il contrario di quello che si riscontra negli Insetti; in questi ultimi, infatti, è l'assenza dell'ormone giovanile a impedire la formazione di uova mature.
Come la riproduzione, così il sesso dei Crostacei è controllato dagli ormoni. I Malacostraci offrono un classico esempio di trasformazione ormonale del sesso negli Invertebrati. Nell'animale giovane non è possibile riscontrare differenziamento del sesso, benché questo sia già genetica- mente determinato. Ma più tardi la cosiddetta ‛ghiandola androgena', mentre cessa di svilupparsi nella femmina, continua a farlo nel corpo del maschio, in cui provoca, mediante la secrezione, l'apparire dei testicoli, organi sessuali primari, nonché degli altri caratteri sessuali secondari. In mancanza delle ghiandole androgene, le cellule germinali primordiali si sviluppano spontaneamente in oociti. Se le ghiandole androgene vengono trapiantate da un giovane maschio in una femmina, gli ovai si trasformano in testicoli funzionanti e tutto l'animale assume carattere maschile. I caratteri sessuali secondari maschili in tutte le parti del corpo sono dovuti all'azione che l'ormone androgeno esplica direttamente sui tessuti, sopraffacendone la determinazione genetica: ovvero, viene risvegliata la capacità latente nel sistema genetico di produrre caratteri maschili.
Molti altri ormoni sono stati identificati nei Crostacei. Un ormone regolatore dell'equilibrio idrico, prodotto dalle ghiandole sinusali, provoca l'assorbimento di acqua durante l'ecdisi, mentre ormoni secreti dal peduncolo oculare regolano la quantità di zucchero circolante nel sangue e forse la sintesi proteica in generale, nonché, in particolare, la formazione di enzimi nell'epatopancreas. L'organo pericardico libera un ormone neurosecretorio di natura peptidica, che incide sull'ampiezza e sulla frequenza del battito cardiaco. E ormoni, infine, determinano gli spettacolari trascoloramenti dei Crostacei. Infatti, quando Koller nel 1928 scopri nel sangue dei gamberetti delle specie Crangon e Palaemonetes sostanze capaci di causare questi trascolaramenti, si ebbe la prima prova certa dell'esistenza di un ormone in un invertebrato. Nei Crostacei, le cellule cromatofore, portatrici del pigmento, contengono granuli di pigmento di colore nero, bianco, rosso, azzurro e giallo; reagendo agli ormoni circolanti, tali cellule possono espandersi o contrarsi, producendo i più vari effetti di colore. In tale processo sono implicati in certo numero gli ormoni, diversi a seconda della specie, che derivano dal sistema neurosecretore costituito dagli organi X e dalla ghiandola sinusale del peduncolo oculare, nonché dalle cellule secretrici del cervello e del torace, che si scaricano attraverso organi postcommissurali. Alcuni crostacei, come il granchio violinista Uca, trascolorano secondo un ritmo circadiano, essendo scuri di giorno e chiari di notte: se i peduncoli oculari vengono escissi, il pigmento si concentra nei cromatofori neri e l'animale impallidisce definitivamente. In effetti, diversi ormoni partecipano al normale ciclo di mutamento: anche le ghiandole sinusali producono una coppia di ormoni antagonisti, capaci di indurre il pigmento retinico degli occhi composti del gamberetto Palaemonetes a spostarsi, secondo una posizione di adattamento, ora al buio, ora alla luce. Tali ormoni sono ben distinti da quelli che controllano le cellule cromatofore nell'integumento.
4. Gli ormoni in altri Artropodi.
In tutti gli altri gruppi di Artropodi sono state trovate cellule neurosecretrici, nonché svariati tipi di organi neuroemali a esse associati; sembra che siano presenti anche ghiandole endocrine paragonabili a quelle protoraciche degli Insetti. Un ‛iniezione di ecdisone induce certamente a una muta il ragno Araneus, il merostoma Limulus, il gambero Procambarus e l'isopodo oniscide Armadillidium. Quindi sembra probabile che un analogo sistema di ormoni possa trovarsi nell'intero phylum.
5. Gli ormoni nei Molluschi.
Si hanno molti dati sulla distribuzione di quelle che sembrano le cellule neurosecretrici dei Molluschi, ma vi sono ben poche prove sperimentali sulla loro funzione. In Octopus vi è chiaramente una ghiandola a secrezione interna, detta ‛ghiandola ottica' (pari), che sembra essere implicata nella maturazione sessuale. Nell'animale immaturo, l'attività di queste ghiandole è soppressa da nervi inibitori di origine cerebrale, tagliando i quali le ghiandole s'ingrandiscono e sia gli ovai sia i testicoli maturano, secondo un sistema che ha qualche analogia con quello corpus allatum-ormone giovanile degli Insetti.
6. Gli ormoni negli Anellidi.
Nessuna ghiandola endocrina epiteliale è stata rintracciata negli Anellidi, presso i quali però abbondano le cellule neurosecretrici, che producono un ormone necessario per mantenere il processo di crescita e per rigenerare i segmenti perduti in seguito a traumi. Alcuni tipi di Nereidi, nel diventare sessualmente maturi, subiscono una sorta di metamorfosi, detta ‛epitochia', che comporta lo sviluppo di parapodi ingranditi con chele più appiattite e muscoli più forti, sì da poter abbandonare il fondo marino e al tempo stesso nuotare nel plancton. Che questo cambiamento si verifichi anzitempo è impedito da un secondo ormone, anch'esso derivato dalle cellule neurosecretrici del cervello e talvolta chiamato ‛ormone giovanile' per analogia con la metamorfosi degli Insetti, benché chimicamente del tutto diverso da quello degli Insetti: solo se diminuisce la concentrazione di questo ormone può avvenire la metamorfosi, o epitochia. Sperimentalmente, infatti, si può impedire l'inizio della metamorfosi epitochiale e quello della maturazione sessuale mediante un trapianto di cervelli prelevati da vermi più giovani.
7. Gli ormoni nei Celenterati e negli Echinodermi.
In questi gruppi possono trovarsi cellule neurosecretrici, anzi vi è qualche prova a favore di una loro funzione endocrina. In Hydra, proprio vicino all'ipostoma, vi sono numerose cellule che sembrano essere neurosecretrici: sostanze estratte da questa regione, applicate ad altra regione del tronco, stimolano la crescita, che nel caso specifico è determinata dalla comparsa di gemme generatrici di teste supplementari. Altrettanto non avviene applicando sostanze estratte da altre parti del corpo. Sembra perciò che le cellule neurosecretrici localizzate dietro l'ipostoma siano indispensabili per la crescita, e che questa cessi in loro assenza. Ma allora le cellule interstiziali si differenziano in gonadi e può aver luogo la riproduzione sessuale.
Nella stella di mare le gonadi mature sono stimolate a scaricare i loro gameti nel mare da un ormone neurosecretorio, che può essere estratto dai nervi radiali: tale ormone ‛diffusore' è un polipeptide di basso peso molecolare (circa 2.600). Liberato apparentemente dai nervi di una stella marina nel mare, può essere quivi captato dai pedicelli ambulacrali di altre stelle marine, provocando la deposizione di uova da parte di queste. Pertanto tale sostanza si comporta sia da ormone, sia da feromone che influenza altri membri della specie.
8. Conclusione.
Sostanze chimiche di ogni genere intervengono negli animali a regolare i cambiamenti legati alla crescita. Per esempio, prodotti di lesioni cellulari, chiamati talvolta ‛ormoni da ferita', danno inizio al processo di cicatrizzazione. La forma del corpo di un animale è il risultato di un differenziamento, durante il quale diversi gruppi di cellule vengono ‛determinati' a produrre strutture particolari. Sembra molto verosimile che questi cambiamenti siano determinati da ‛regolatori', o ‛sostanze induttrici', che agiscono analogamente all'ormone giovanile e producono differenze strutturali risvegliando potenzialità specifiche presenti allo stato latente nel sistema genetico.
È stato anche dimostrato che queste sostanze induttrici possono essere distribuite in ‛gradienti' e che la natura della trasformazione genetica indotta è dovuta al grado di concentrazione di una singola sostanza - proprio come l'ormone giovanile degli Insetti può determinare la formazione di larve, crisalidi o adulti, a seconda della sua concentrazione. Talvolta, particolari geni possono far produrre specifiche sostanze chimiche responsabili di cambiamenti in tutto il corpo, riguardanti vari fenomeni, quale la pigmentazione o altri.
Tutti questi processi, che riscontriamo in tutti gli animali multicellulari e che, sia pure in forma modificata, devono aver luogo perfino nei Protozoi unicellulari con la loro struttura variata, hanno qualcosa in comune con gli ormoni propriamente detti, che circolano nel sangue. A mano a mano che gli Invertebrati sono divenuti più complessi e hanno acquisito un sistema nervoso più complicato, hanno raggiunto una forma sempre più elaborata di coordinazione chimica per mezzo degli ormoni circolanti.
Bibliografia.
Butler, C. G., Insect pheromones, in ‟Biological reviews", 1967, XLII, pp. 42-87.
Cottrell, C. B., Insect ecdysis with particular emphasis on cuticular hardening and darkening, in ‟Advances in Insect physiology", 1964, II, pp. 175-218.
Danilevskij, A. S., Photoperiodism and seasonal development of Insects, Edinburgh 1965.
Engelmann, F. A., Endocrine control of reproduction in Insects, in ‟Annual review of entomology", 1968, XIII, pp. 1-26.
Engelmann, F. A., The physiology of Insect reproduction, Oxford-New York 1970.
Gilbert, L. I., Hormones regulating Insect growth, in The hormones: physiology, chemistry and applications (a cura di G. Pincus e K. V. Thimann), vol. IV, New York 1964, pp. 67-134.
Herman, W. S., The ecdysial glands of Arthropods, in ‟International review of cytology", 1967, XXII, pp. 269-347.
Highnam, K.C., Hill, L., The comparative endocrinology of the Invertebrates, London 1969.
Jenkin, P. M., Animal hormones: a comparative survey, vol. II, Control of growth and metamorphosis, Oxford-New York 1970.
Kennedy, J. S. (a cura di), Insect polymorphisms. Symposium of the Royal Entomological Society of London, London 1961.
Krause, G., Sander, K., Ooplasmic reaction systems in Insect embryogenesis, in ‟Advances in morphogenesis", 1962, II, pp. 259-303.
Lees, A. D., The physiology of diapause in Arthropods, London 1955.
Lees, A. D., The control of polymorphism in aphids, in ‟Advances in Insect physiology", 1966, III, pp. 207-277.
Locke, M., The development of patterns in the integument of Insects, in ‟Advances in morphogenesis", 1967, VI, pp. 33-88.
Smallman, B. N., Mansingh, A., The cholinergic system in Insect development, in ‟Annual review of entomology", 1969, XIV, pp. 387-408.
Treherne, J. E., The neurochemistry of Arthropods, London 1966.
Weaver, N., Physiology of caste determination, in ‟Annual review of entomology", 1966, XI, pp. 79-102.
Wigglesworth, V. B., The hormonal regulation of growth and reproduction in Insects, in ‟Advances in Insect physiology", 1964, II, pp. 247-336.
Wigglesworth, V. B., Insect hormones, Edinburgh 1970.
Wilde, J. de, Photoperiodism in Insects and Mites, in ‟Annual review of entomology", 1962, VII, pp. 1-26.
Ormoni nei Vertebrati
SOMMARIO: 1. Introduzione. □ 2. Ormoni simili ad amminoacidi o derivati da essi: a) ormoni catecolamminici; b) ormone tiroideo. □ 3. Ormoni polipeptidici: a) ormoni dell'ipotalamo; b) ormoni adenoipofisari; c) ormoni neuroipofisari; d) insulina e glucagone; e) ormone paratiroideo e calcitonina; f) ormoni gastrointestinali. □ 4. Ormoni steroidi: a) ormoni della corticale del surrene; b) ormoni sessuali. □ 5. Prostaglandine. □ 6. Considerazioni conclusive. □ Bibliografia.
1. Introduzione.
Gli ormoni sono composti chimici che portano nella loro struttura un messaggio che usualmente induce una cellula a cambiare il proprio metabolismo. Questa definizione indica subito che devono esistere due tipi di cellule, uno che elabora il messaggio e l'altro che lo riceve (v. endocrinologia). A tutt'oggi noi non comprendiamo la relazione tra struttura chimica e messaggio metabolico specifico che essa contiene. Alcune ipotesi sul modo in cui può essersi sviluppato il ‛codice metabolico' sono presentate qui di seguito.
Gli ormoni che si trovano nei Vertebrati rappresentano uno sviluppo che può essere considerato una combinazione di adattamento, mutazione ed evoluzione ortogenetica, ben diverso dai meno complessi sistemi di regolazione degli organismi unicellulari. Gli ormoni dei Vertebrati, dal punto di vista strutturale, possono essere classificati in tre categorie generali: 1) ormoni correlati ad amminoacidi, come, per esempio, la tiroxina e l'adrenalina (o epinefrina); 2) ormoni a struttura polipeptidica, come: gli ormoni tropici dell'ipofisi anteriore; gli ormoni della parte endocrina del pancreas, insulina e glucagone; gli ormoni del tratto gastrointestinale e i peptidi a basso peso molecolare, caratterizzati di recente; i fattori liberatori ipotalamici (releasing factor); 3) ormoni a struttura steroide, come gli ormoni sessuali e gli ormoni corticosurrenali.
Origine degli ormoni. - Dal punto di vista delle nostre attuali conoscenze sull'origine delle cellule primitive dalle condizioni prebiotiche esistenti un tempo sulla Terra - campo scientifico, questo, relativamente nuovo - sembra probabile che gli amminoacidi e i polipeptidi possano essersi formati più facilmente delle strutture steroidi. Da ciò si potrebbe concludere, in via del tutto ipotetica, che gli ormoni a struttura peptidica siano apparsi sulla Terra prima degli ormoni a struttura steroide. Ancora, sembra probabile che le prime sostanze nutritive che hanno permesso la crescita e la riproduzione delle cellule primordiali negli oceani primitivi siano state di natura proteica o peptidica. Ciò se fosse vero, spiegherebbe la specificità strutturale degli ormoni peptidici nel processo evoluzionistico attraverso le mutazioni. In altri termini, le cellule primordiali si sarebbero nutrite dei peptidi più abbondanti negli oceani, peptidi che potrebbero avere avuto una struttura abbastanza uniforme. Sono stati fatti esperimenti a sostegno di questa ipotesi, particolarmente da S. Fox e collaboratori e da altri: si è dimostrato che in condizioni simulanti lo stato prebiotico la serie di polipeptidi ottenuti è abbastanza costante per dimensioni e composizione di amminoacidi, pur essendo stata fatta variare alquanto la concentrazione dei singoli amminoacidi usati come precursori.
Quindi, i polipeptidi abbondantemente presenti negli oceani prebiotici potrebbero aver costituito gli alimenti primari delle cellule primitive; quando, aumentando le cellule, questi polipeptidi divennero scarsi e la loro utilizzazione come cibo risultò insufficiente, la prima grande mutazione potrebbe essere stata l'adattamento delle cellule a un diverso, abbondante materiale, utilizzabile come nutrimento. E possibile che la mutazione sia stata accompagnata dalla formazione di una ‛memoria' (acidi nucleici) della struttura del polipeptide usato in precedenza come cibo e che questo polipeptide abbia svolto in seguito una funzione regolatrice. Esperimenti di laboratorio simulanti le condizioni prebiotiche idonee alla formazione di ‛proteidi' o proteine primigenie hanno dimostrato che nella miscela formatasi vi è, per esempio, un'attività stimolante i melanociti (cioè un ormone).
Per quanto riguarda la successione temporale in cui sono comparsi, nel corso del processo evolutivo, i vari tipi di ormoni, è probabile che gli ormoni steroidi, i quali svolgono un importante ruolo nello sviluppo e nell'attività sessuale come pure nella funzione della corteccia surrenale, abbiano fatto la loro apparizione in un tempo successivo rispetto agli ormoni peptidici. Fino a oggi l'organismo più primitivo in cui si sia stabilita la presenza di uno steroide con funzione ormonale è un fungo, la muffa d'acqua; lo steroide isolato da questo organismo facilita il processo di riproduzione, agendo quindi come ormone sessuale.
Gli ormoni corticosurrenali appaiono successivamente nell'evoluzione, ma le cellule che li sintetizzano possono essere derivate dallo stesso tipo di cellule che elaborano gli ormoni sessuali. E plausibile che la necessità di nutrirsi abbia preceduto lo sviluppo della sessualità, nel corso della evoluzione. Inoltre, la struttura polipeptidica contiene una informazione derivata dalla traduzione dell'RNA, che l'ha, a sua volta, ottenuta dal gene, mentre la struttura steroide non implica un'informazione simile; possiamo forse considerare le attività ormonali delle strutture steroidi come un tardo perfezionamento del sistema di regolazione degli organismi complessi multicellulari.
Derivazione degli ormoni dal tessuto nervoso. - Negli animali gli ormoni e il sistema nervoso centrale agiscono in tandem, ed è interessante notare che il sistema nervoso centrale innerva tutti gli organi del corpo; alcuni neurotrasmettitori (come la noradrenalina) sono considerati ormoni. È essenziale correlare l'attività ormonale dei Vertebrati con l'insieme del sistema nervoso centrale: dalla biologia comparata risulta che l'evoluzione dei sistemi regolatori potrebbe essersi svolta così: da cellule nervose (neuroumori o neurotrasmettitori) a cellule neurosecretrici (neurormoni) e infine a ghiandole endocrine (ormoni).
Nei Vertebrati inferiori il sistema nervoso centrale è l'agente principale per l'integrazione delle funzioni; in alcuni Invertebrati superiori cominciano ad apparire cellule neurosecretrici, che sono sempre presenti nei Vertebrati. Le ghiandole senza dotto (o endocrine), caratteristiche degli organismi superiori, non sono presenti negli Invertebrati inferiori, in cui le funzioni principali, come riproduzione e metabolismo, sono controllate da neurosecrezioni: è quindi probabile che i sistemi neurosecretori si siano sviluppati prima delle ghiandole endocrine. Queste, comparse più tardi nell'evoluzione, sono controllate dalle cellule neurosecretrici per coordinare le risposte dei due sistemi a stimoli interni ed esterni. L'interazione tra i due sistemi nei Vertebrati superiori apparirà più evidente quando, nel corso di questo articolo, si vedrà come stimoli nervosi sono correlati all'elaborazione di ormoni: nei Mammiferi i centri neurosecretori che rispondono alla stimolazione nervosa iniziando il processo di elaborazione degli ormoni sono collocati principalmente in un'area del diencefalo ventrale, l'ipotalamo.
Differenziazione dei tessuti e ormoni. - Negli organismi superiori la distribuzione dei compiti tra cellule di tessuti differenti ha raggiunto un alto grado di efficienza: per esempio alcune cellule sono destinate a sintetizzare ormoni e a riversarli nel torrente circolatorio, dopo aver ricevuto un opportuno segnale. L'ormone prodotto arriva agli organi-bersaglio, che possono essere indotti, tra l'altro, a sintetizzare specifici enzimi. La divisione dei compiti, che affida la produzione degli ormoni a particolari cellule, permette alle cellule-bersaglio di funzionare senza possedere la notevole serie di enzimi necessari alla sintesi dell'ormone; a loro volta le cellule che producono ormoni dipendono, per la loro normale attività cellulare, da altre cellule specializzate; l'alto grado d'interdipendenza consente a un organismo di raggiungere una complessità tale da compiere funzioni sofisticate. Malauguratamente, più complesso diviene un organismo, più grande è la possibilità di disfunzioni, in quanto vi sono più funzioni da controllare; queste disfunzioni si manifestano come quadri patologici diversi, caratterizzati da iperfunzioni o da ipofunzioni dei tessuti che producono ormoni. Meno chiari sono a tutt'oggi l'ipo- e l'iperfunzione dei vari sistemi che trasportano gli ormoni agli organi-bersaglio o la presenza di livelli insolitamente bassi o alti di proteine ‛recettrici' all'interno delle cellule-bersaglio.
Ormoni e sviluppo. - È noto che gli ormoni sono importanti per la formazione di enzimi nei diversi tessuti durante la gestazione e il periodo neonatale; da interferenze con queste funzioni ormonali derivano gravi alterazioni. Ciò che non è chiaro è il potenziale ruolo degli ormoni nell'iniziale sviluppo degli organismi; molte ricerche sono state compiute sulle cosiddette ‛sostanze induttrici' nell'embriogenesi; tuttavia, dato il genere di esperimenti necessari per confermare quest'ipotesi, non è stato possibile chiarire definitivamente se in effetti queste sostanze induttrici siano responsabili della differenziazione organizzata delle varie cellule per formare i diversi tessuti. Se così non fosse, tutte le informazioni necessarie per il futuro sviluppo potrebbero essere contenute già nell'uovo fecondato. Ciò nondimeno è interessante osservare che alcuni tipi di funzioni differenziate nelle primitive cellule totipotenti possono essere influenzati da sostanze semplici, come la fenilalanina, che è un amminoacido precursore degli ormoni tiroidei e delle catecolammine previa trasformazione in tirosina. Non verranno trattati quindi, nel presente articolo, gli ormoni degli animali superiori sotto l'aspetto di sostanze induttrici, soprattutto perché l'esistenza di ‛induttori' embrionali è ancora in discussione.
Come agiscono gli ormoni a livello cellulare. - Gli ormoni, negli animali superiori, possono esercitare i loro effetti sia ‛all'interno delle cellule' (ormoni steroidi) sia, nel caso degli ormoni polipeptidici, ‛sulle membrane cellulari', stimolando per esempio uno specifico enzima, la adenilatociclasi, a produrre AMP ciclico all'interno della cellula; quest'ultima sostanza è stata considerata come ‛secondo messaggero' all'interno della cellula. L'AMP ciclico, come vedremo, può stimolare determinati enzimi a compiere determinate funzioni, la maggior parte delle quali riguarda la biosintesi di macromolecole, come il glicogeno, di steroidi ecc. Gli ormoni possono influenzare l'informazione genetica all'interno della cellula causando la derepressione di zone del gene e di conseguenza la produzione di maggiori quantità dell'RNA messaggero codificato dal gene specifico; così la traduzione di questo RNA informazionale produce maggiori quantità di una proteina fenotipica. Infine, gli ormoni possono in alcuni casi influenzare vari processi nel citoplasma, particolarmente quelli riguardanti la traduzione, a livello citoplasmatico, dell'informazione genetica mediante l'RNA messaggero. Bisogna comunque tener presente che nessuna di queste funzioni è stata definitivamente accertata : rimane infatti ancora da chiarire la specifica interazione molecolare per cui un dato ormone causa una serie correlata di eventi culminante nel suo specifico effetto biologico ; sono tuttavia in corso numerose ricerche che prima o poi chiariranno l'effetto degli ormoni anche a livello molecolare. Il problema del meccanismo con cui gli ormoni agiscono a livello sub- cellulare è molto complesso e richiede molte più informa- zioni sui processi cellulari fondamentali di quelle che si possiedono oggi. Inoltre, poiché gli ormoni fungono da regolatori di cellule differenziate, la conoscenza dei loro meccanismi d'azione può chiarire aspetti dello stesso processo di differenziamento.
Probabilmente l'aspetto non chiarito e più imbarazzante degli ormoni negli organismi superiori è il rapporto tra la struttura specifica di un dato ormone e il genere di risposte da esso provocate : in quest'area quasi inesplorata potrebbe essere collocato il problema del ‛codice metabolico'. Come vedremo in seguito, una parte di questo codice può essere compresa nella struttura dei recettori delle cellule-bersaglio a livello della membrana cellulare o all'interno della cellula. Comunque, il problema della struttura ormonale specifica e della sua evoluzione resta, in massima parte, un problema per il futuro.
2. Ormoni simili ad amminoacidi o derivati da essi.
I più importanti ormoni simili ad amminoacidi o derivati da essi sono le catecolammine e l'ormone tiroideo.
a) Ormoni catecolamminici.
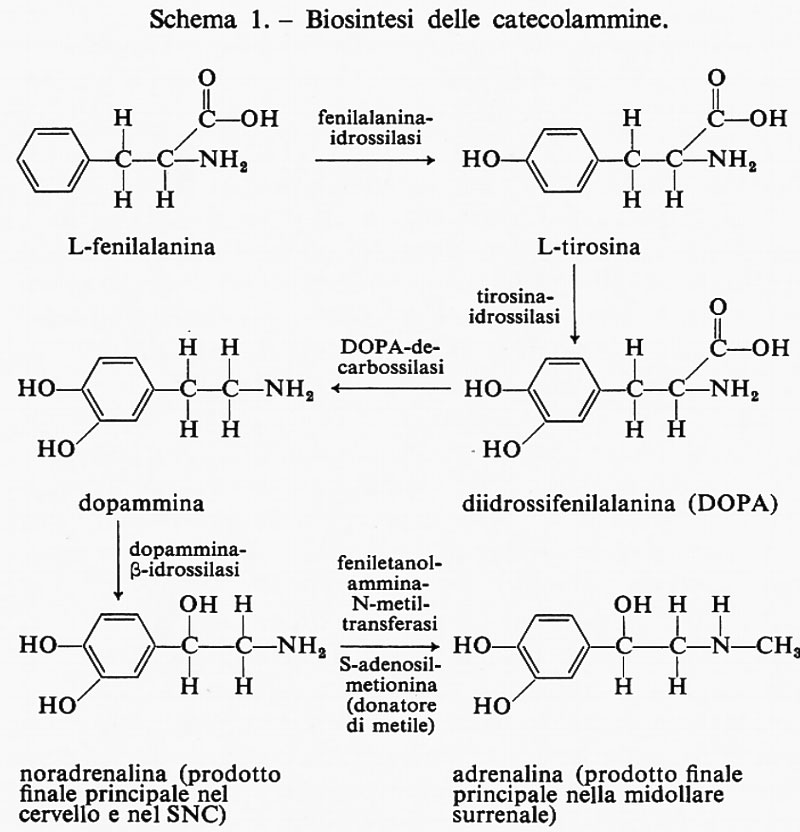
Tra le catecolammine l'adrenalina (o epinefrina) è il principale prodotto della midollare del surrene, mentre la noradrenalina è un neurotrasmettitore del sistema nervoso centrale. Entrambi gli ormoni sono derivati dalla tirosina, o dalla fenilalanina per idrossilazione di quest'ultima a tirosina, mediante reazioni enzimatiche simili; l'adrenalina deriva dalla noradrenalina, particolarmente nella midollare del surrene, per mezzo di un enzima specifico di questo tessuto, la feniletanolammina-N-metil-translerasi (PNMT) (v. schema 1); l'enzima sembra essere controllato dagli ormoni della corticale del surrene.
L'idrossilazione specifica della tirosina da parte della tirosinaidrossilasi è il passaggio limitante nella sintesi delle catecolammine: si tratta di un enzima complicato, ad alto peso molecolare, che pare sia sotto controllo allosterico. Non si è ancora avuta una prova definitiva di ciò, in quanto la purificazione completa dell'enzima è tuttora in corso; tuttavia, sulla base di studi preliminari cinetici e di inibizione si può congetturare che, per esempio, nel cervello possa esistere un sistema a feedback negativo il cui scopo sarebbe di controllare la quantità totale di noradrenalina sintetizzata nelle terminazioni nervose. Quando, attraverso le reazioni suddette, si sono formate quantità relativamente alte del prodotto finale, la noradrenalina, la concentrazione di questa raggiungerà valori tali da superare la costante di associazione per il sito allosterico, e la noradrenalina stessa si potrebbe legare a questo sito causando un cambiamento di conformazione della tirosinaidrossilasi polimerica e, di conseguenza, una modificazione dell'architettura del sito attivo che renderebbe l'enzima meno reattivo verso il substrato, la tirosina. Si produrrebbe così un'interruzione nella sintesi di noradrenalina, finché questa non venga consumata per la trasmissione nervosa; l'utilizzazione a tale scopo della noradrenalina potrebbe determinarne la diminuzione della concentrazione a livello delle terminazioni nervose fino a un valore forse inferiore a quello della sua costante di associazione e provocarne pertanto il distacco dal sito allosterico. Conseguentemente l'enzima, riacquistata la sua conformazione cataliticamente più attiva, potrebbe nuovamente legare la tirosina e formare altra noradrenalina finché la concentrazione di questo prodotto sia di nuovo più alta della costante di affinità per il sito allosterico della tirosinaidrossilasi.
Con questo presunto meccanismo ciclico la sintesi della noradrenalina potrebbe essere controllata semplicemente dalla quantità formatasene in vicinanza dell'enzima limitante, la tirosinaidrossilasi.
Effetti delle catecolammine. - Prese nel complesso, le catecolammine sono ormoni di ‛emergenza', che preparano l'organismo ‛alla fuga o alla lotta' e influenzano la pressione del sangue e le vie metaboliche che controllano la quantità di carboidrati da bruciare per impreviste necessita energetiche.
Di recente, grande interesse ha suscitato l'uso clinico di un intermedio della via biosintetica della noradrenalina, dianzi illustrata, cioè la DOPA. Questo composto viene usato con qualche successo nel trattamento del morbo di Parkinson, che è un'affezione del sistema nervoso centrale caratterizzata tra l'altro da tremore: poiché è noto che le catecolammine possono abolire tremori e brividi, potrebbe darsi che in questa malattia vi sia un'insufficiente produzione di neurotrasmettitori in determinate parti del cervello.
La tirosinaidrossilasi può essere inibita da un analogo della tirosina, l'α-metiltirosina, che impedisce la sintesi di adrenalina. L'uso di questo composto in clinica costituisce un tentativo sperimentale per ridurre la pressione sanguigna.
Struttura e funzione. - L'attività biologica delle catecolammine sembra risiedere nei gruppi idrossilici presenti, uniti ai C-3 e C-4 dell'anello; anche l'anello benzenico è essenziale per gli effetti stimolatori del sistema nervoso centrale. Sostituzioni nel carbonio a prolungano l'effetto di questi ormoni riducendo l'azione della monoamminossidasi che inattiva le catecolammine.
Metabolismo delle catecolammine. - Le reazioni di degradazione delle catecolammine, che portano alla loro inattivazione e ai prodotti di escrezione, sono riportate nello schema 2, in cui non sono indicate molte delle altre possibili trasformazioni: per esempio è possibile, per azione della catecolammina-O-metiltransferasi (COMT), un accumulo di dimetossifenetilammina che, somministrata ad animali, produce reazioni allucinogene.
Una delle ipotesi attuali a proposito della base molecolare delle malattie mentali è che si verifichi un'alterazione che impedisca la normale escrezione di composti di questo genere, oppure che vi sia una loro accelerata sintesi cosicché si avrebbero sindromi psichiche causate da concentrazioni relativamente elevate di agenti allucinogeni endogeni. Tuttavia non è stata ancora raggiunta la certezza di questo fatto.
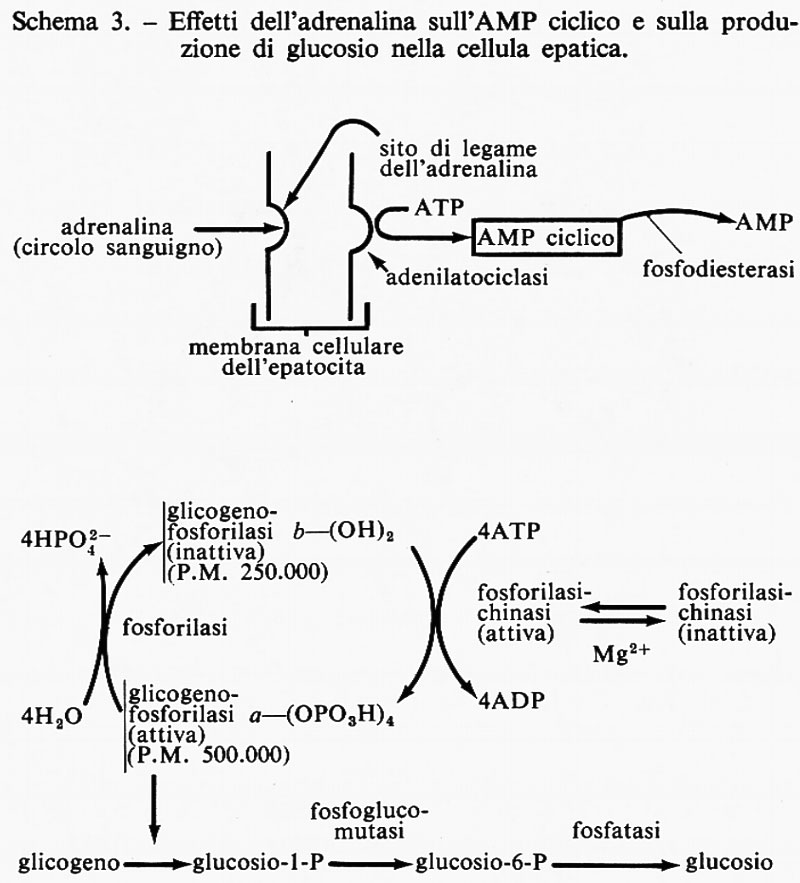
Effetti sul metabolismo. - Come gli altri ormoni, le catecolammine producono numerosi effetti nell'organismo; molti di questi sono riportati nella tab. I. I maggiori effetti sul metabolismo sono dovuti all'adrenalina: dopo somministrazione di questo ormone vi è un rapido aumento del glucosio e dell'acido lattico nel sangue, a spese del glicogeno epatico e, in seguito, anche di quello muscolare. Si pensa che questi effetti siano dovuti all'attivazione, da parte dell'adrenalina, del sistema fosforilasico e, contemporaneamente, all'inibizione della secrezione di insulina: entrambi questi eventi provocano un aumento della glicemia (effetto iperglicemizzante). Anche gli acidi grassi liberi del sangue sono aumentati, in quanto l'ormone provoca la scissione dei grassi nel tessuto adiposo. A causa dell'aumentata concentrazione di composti rapidamente utilizzabili per produrre energia, si ha un aumento del consumo di ossigeno (v. tab. I). Vi è anche, in risposta a un aumento dell'adrenalina, una secrezione, da parte dell'ipofisi anteriore, di ormone adrenocorticotropo (ACTH), il cui effetto è di indurre la sintesi e l'immissione nel torrente circolatorio di corticosteroidi da parte della ghiandola surrenale. L'ACTH stimola anche la sintesi di zuccheri: si verifica così una serie di eventi a cascata che prepara l'animale all'utilizzazione improvvisa di quantità di energia relativamente grandi.
Adrenalina e AMP ciclico. - L'effetto dell'adrenalina sul sistema fosforilasico con la conseguente produzione di glucosio è mediato dalla produzione di AMP ciclico, la cui struttura è:
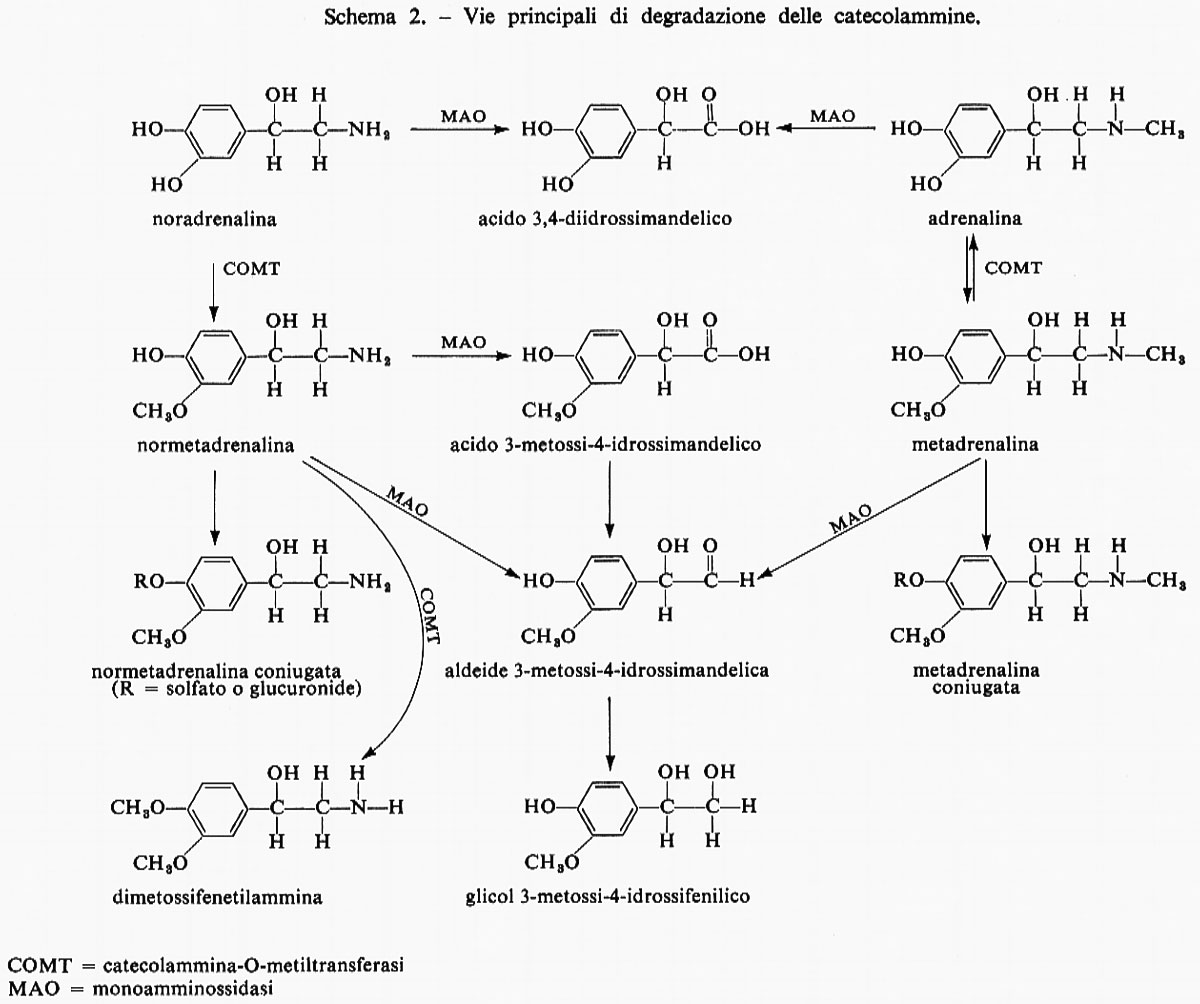
e il cui modello è riportato nella fig. 1. Questo importante composto intracellulare è stato chiamato ‛secondo messaggero' da E. W. Sutherland e collaboratori. Alcune conseguenze dell'azione dell'adrenalina sono illustrate nello schema 3; la stessa serie di eventi si verifica nelle cellule muscolari fino alla produzione di glucosio-6-fosfato: attraverso il metabolismo glicolitico questo composto è trasformato in acido lattico.
b) Ormone tiroideo.
Un altro ormone correlato con l'amminoacido tirosina, o da esso derivato, è la tiroxina o tetraiodotironina della ghiandola tiroide. Questo ormone è caratterizzato dal contenere iodio e un legame etere tra due strutture cicliche; è del tutto differente dall'ormone peptidico anch'esso prodotto in questa ghiandola e conosciuto come calcitonina, di cui si parlerà in seguito.

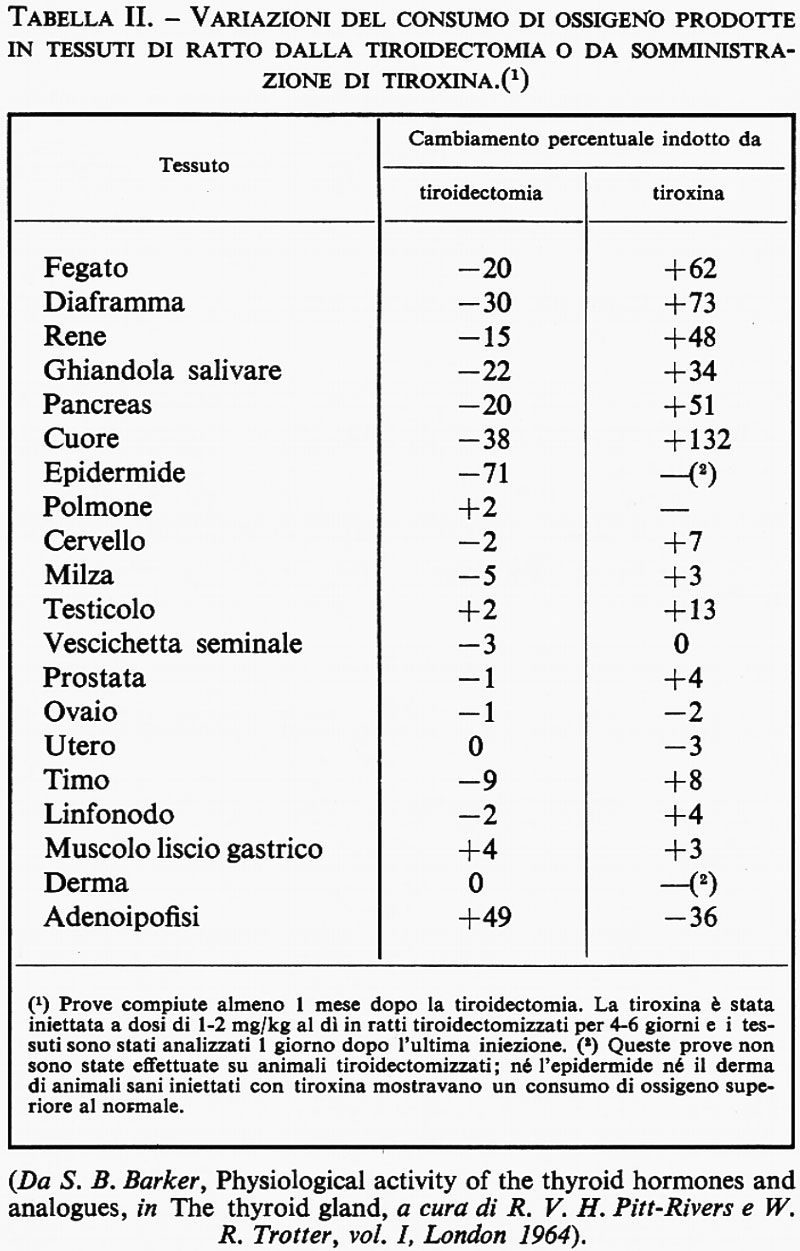
Ormone tiroideo e metabolismo. - L'ormone tiroideo regola il metabolismo della maggior parte dei principali tessuti adulti; i suoi effetti sono riassunti nella tab. II.
L'aumento del metabolismo si riflette in un aumento della frequenza del polso: la relazione tra la velocità di utilizzazione periferica di tiroxina e il rapporto tra frequenza del polso e peso corporeo è illustrata nella fig. 2. Di conseguenza, questo ormone è importante nello sviluppo e nella maturazione di molti tessuti (v. tab. II) e può agire, durante lo sviluppo, anche sul processo morfogenetico di tessuti che nel prosieguo della vita divengono insensibili o impermeabili all'ormone: un esempio di questo genere è costituito dal tessuto cerebrale. A causa dei suoi considerevoli effetti sul metabolismo, la tiroide è stata spesso indicata come ‛ghiandola maestra'.
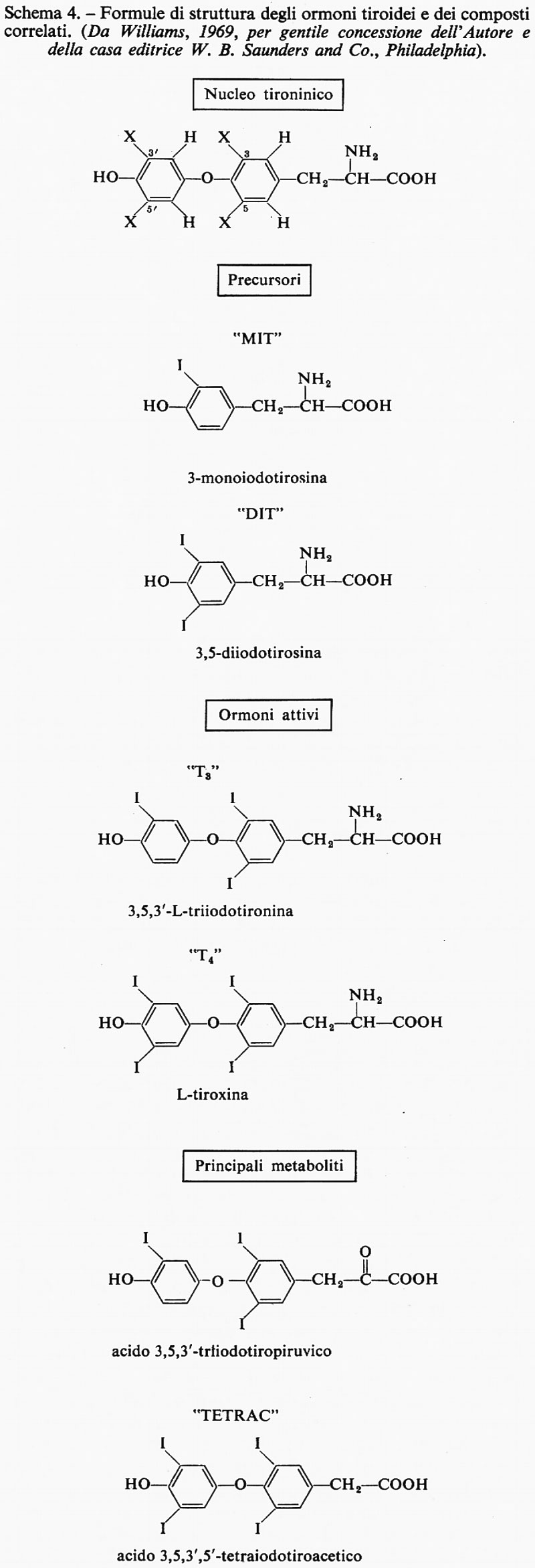
Struttura e funzione dell'ormone tiroideo. - La struttura degli ormoni tiroidei è presentata nello schema 4 e nella fig. 3; gli ormoni attivi sono la L-tiroxina o 3,5,3′,5′-L-tetraiodotironina (T4) e la 3,5,3′-L-triiodotironina (T3), che differiscono solo per la presenza o l'assenza di un atomo di iodio nella posizione 5′ dell'anello più esterno. La ghiandola tiroide (circa 20-30 grammi nell'adulto) secerne l'ormone per soddisfare la richiesta dei tessuti periferici. La sintesi di ormone tiroideo nella ghiandola dipende dall'apporto alimentare di iodio, che varia ampiamente a seconda della località e della disponibilità dei diversi cibi che contengono questo elemento. Nella tiroide si trova il 90% circa dello iodio dell'organismo e circa il 10% di esso è presente come ioduro. Il tasso di ricambio dello iodio tiroideo nel circolo periferico è circa del 10% al dì. La prima tappa della biosintesi dell'ormone tiroideo è il trasporto attivo dello ioduro nella ghiandola tiroide, seguito dall'ossidazione dello ioduro e dalla iodurazione, da parte della forma ossidata, di residui di tirosina contenuti nella tireoglobulina (la forma di deposito dell'ormone tiroideo nella ghiandola tiroide) con produzione di tirosine iodate inattive dal punto di vista ormonale, e infine dall'accoppiamento delle iodotirosine a formare le iodotironine ormonicamente attive. I derivati tironinici attivi sono legati per mezzo di legami peptidici alla tireoglobulina, forma macromolecolare di deposito dell'ormone.
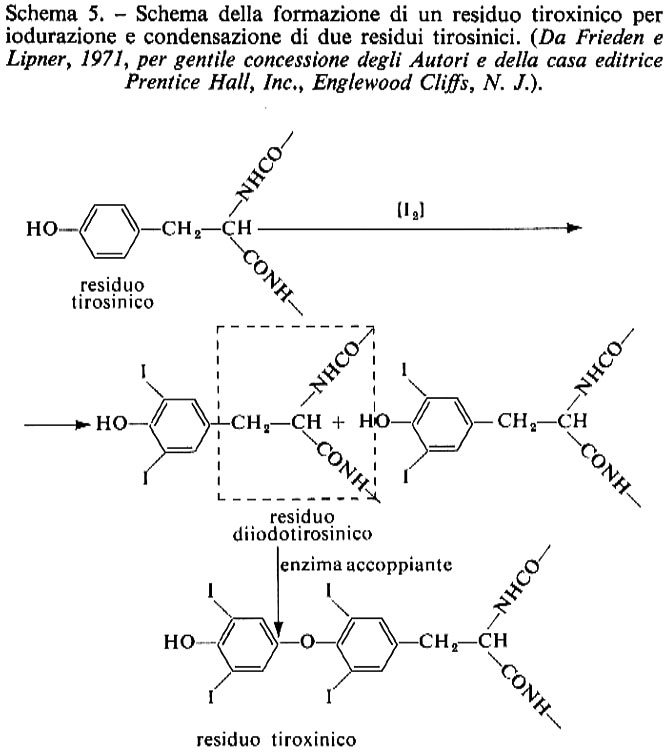
La sintesi del composto T4 potrebbe avvenire come mostrato nella fig. 4 e nello schema 5. Per idrolisi della tireoglobulina, effettuata da enzimi proteolitici e da peptidasi all'interno della tiroide, vengono messe in circolo le iodotironine e le iodotirosine (MIT e DIT); queste ultime vengono poi private dello iodio.
Secrezione della ghiandola tiroide ed effetti molecolari nelle cellule-bersaglio. - Gli ormoni messi in circolo per proteolisi parziale della tireoglobulina avvenuta nella ghiandola, vengono legati a tre proteine seriche che sono i loro agenti di trasporto e di accumulo nel sangue: in particolare il T4 si lega a una globulina specifica (TBG) oltre che a una prealbumina (che deve questo nome al comportamento elettroforetico) e all'albumina. Le prime due proteine sembrano essere molto più specifiche dell'ultima nel legare l'ormone. E probabile che l'ormone sia ceduto ai vari tessuti dal sangue circolante per dissociazione del complesso proteina-ormone e l'ormone libero sia captato dalle cellule di un determinato tessuto, secondo lo schema:
Benché sembri che venga prodotta, e messa in circolo, più T4 che T3, quest'ultima forma ormonale ha, per cause non completamente chiarite, un maggiore effetto biologico. Comunque, recenti ricerche hanno dimostrato che all'interno della cellula vi potrebbero essere recettori proteici specifici: studi condotti su cellule epatiche inducono a ritenere che il T4 e il T8 vengano legati a proteine diverse. Poiché la concentrazione intracellulare di T4 e di T5 nei tessuti dove agiscono è dell'ordine di 10-8M, il legame tra ormoni e recettori dev'essere molto specifico. Il complesso formato da recettore proteico e ormone tiroideo potrebbe dare inizio a una catena di eventi intracellulari, ciascuno causato dal precedente, che porta a un aumento del metabolismo di un dato tessuto, ma questi meccanismi sono oggi ancora poco noti.
Inattivazione e rimozione dell'ormone tiroideo. - Per essere allontanato dall'organismo, l'ormone può essere coniugato come glucuronide nel fegato ed escreto attraverso il sistema epatobiliare; si può verificare un certo riassorbimento dall'intestino previa idrolisi del gruppo coniugato. Il rene, particolarmente la frazione mitocondriale, è capace di attaccare la catena laterale di T4 e di T3 e alcuni prodotti di degradazione possono essere escreti dal rene stesso.
3. Ormoni polipeptidici.
Come ricordato in precedenza, questi ormoni possono essere arbitrariamente divisi in due classi quelli ad alto peso molecolare e i peptidi a basso peso molecolare, in particolare quelli di recente individuati nell'ipotalamo.
a) Ormoni dell'ipotalamo.
L'ipotalamo, che sembra controlli le secrezioni dell'ipofisi (v. sotto), è a sua volta innervato dal sistema nervoso centrale, costituendo così un'importante connessione tra stimolazione nervosa (che è una reazione a fattori ambientali) ed elaborazione di ormoni. La fig. 5 illustra le connessioni anatomiche tra ipotalamo e ipofisi: il peduncolo ipofisario, che connette l'ipofisi anteriore con l'eminenza mediana del tuber cinereum, contiene un piccolissimo sistema portale di vasi sanguigni; gli stimoli nervosi vengono trasmessi all'ipotalamo, dove probabilmente provocano la produzione, da parte di varie zone nervose (cellule neurosecretrici), di sostanze umorali che, immesse nel circolo portale suddetto, portano questi segnali nell'ipofisi anteriore per causare a loro volta la produzione degli specifici ormoni tropici. Una volta immessi nel circolo generale, questi ultimi sono captati a distanza dai recettori delle membrane cellulari dei tessuti-bersaglio. Alcuni aspetti di questo fenomeno a cascata sono illustrati nella fig. 6. La specificità della successiva secrezione di un particolare ormone tropico da parte dell'ipofisi anteriore sembra essere controllata dalla speciale struttura del fattore liberatore (releasing factor) ipotalamico. Recentemente vi è stato un grande interesse riguardo all'identificazione dei fattori liberatori ipotalamici, alla determinazione della loro struttura e alla sintesi chimica di ormoni attivi. Le loro dimensioni relativamente ridotte e la loro specificità fisiologica li rendono particolarmente idonei per il trattamento clinico di vari disordini endocrini. Fino a oggi sono stati individuati almeno sette diversi neurormoni ipotalamici; per lo più sono polipeptidi formati da 3 o più amminoacidi. La fig. 7 mostra la zona dell'ipotalamo e la direzione del flusso dei segnali che provocano la secrezione degli ormoni tropici da parte dell'ipofisi anteriore.
Fattore liberatore della tirotropina. - Uno di questi ormoni, il fattore liberatore della tirotropina (TRF, Thyrotropin Releasing Factor, o TRH, Thyrotropin Releasing Hormone) è un tripeptide: piroglutammil-istidil-prolinammide.
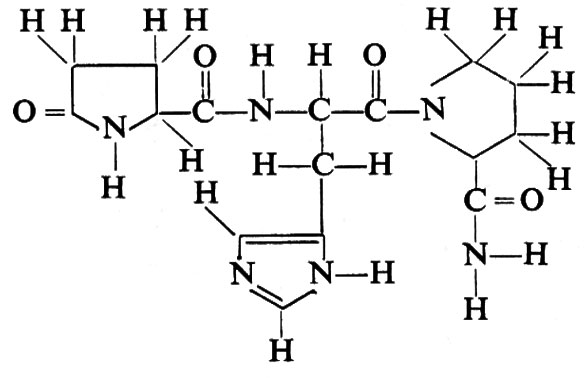
La struttura tridimensionale di questa molecola è illustrata nella fig. 8. Le particolari cellule dell'ipotalamo da cui derivano i fattori liberatori non sono state ancora identificate; inoltre niente si sa della biosintesi di questi piccoli peptidi. Esperimenti recenti hanno dimostrato che la produzione di ormone luteinizzante (LH) e di ormone follicolo-stimolante (FSH) viene stimolata da un unico fattore, per il quale è stata proposta la struttura: piroglutammil-istidil-triptofanil-seril-tirosil-glicil-leucil-arginil-prolil-glicinammide; chimicamente è stato sintetizzato un polipeptide più semplice, con attività simile nel causare la secrezione di questi due ormoni tropici, la cui struttura è: piroglutammiltirosil-arginil-triptofanilammide.
Ormone inibitore della secrezione dell'ormone stimolante i melanociti. - Un altro ormone dell'ipotalamo bovino è quello che inibisce la produzione dell'ormone stimolante i melanociti (MRIH, Melanocyte stimulating hormone Release-Inhibiting Hormone); la sua struttura, di recente chiarita, è: L-prolil-Leucil-glicinammide. Il modello molecolare di
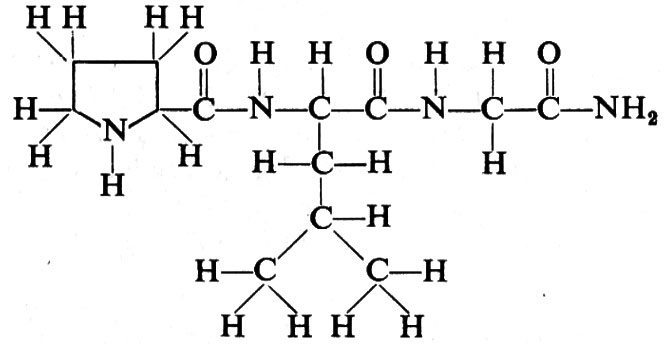
questo polipeptide è riportato nella fig. 9, e può essere comparato con quello del TSH-RH (o TRF) della fig. 8. Oltre all'MRIH (o MIH) e al fattore liberatore dell'ormone stimolante i melanociti (MRF o MRH), è stato possibile individuare altri ormoni ipotalamici, quali il fattore liberatore della corticotropina (CRF o CRH), i fattori liberatore e inibitore della secrezione dell'ormone somatotropo (GRF o OHRF, Growth Hormone Releasing Factor, e GHRIF o GHRIH o somatostatina) e i fattori liberatore e inibitore della secrezione dell prolattina (PRF o PRH e PIF o PIH o PRIH). La struttura del CRH non è completamente chiarita: sembra in parte correlata a quella dell'ormone stimolante i melanociti (MSH). Dall'α-MSH è stata separata una sostanza, detta -CRF o βCRH, la cui possibile struttura parziale è acetilseril-tirosil-cisteinil-fenilalanil-istidil (asparaginil, glutammil)-cisteinil-(prolil, valil)-lisil-glicinammide. Alcuni frammenti dell'α-MSH hanno in vitro attività CRH; tra gli altri ricorderemo i due composti seguenti: H-metionil-glutamminil-istidil-fenilalanil-arginiltriptofanil-glicina-OH e H-seril-tirosil-seril-metionil-glutamminil-istidil-fenilalanil-arginil-triptofanil-glicina-OH.
Il GRF o GRH è conosciuto meno bene chimicamente: pare sia un polipeptide acido dal peso molecolare intorno a 2.500. La somatostatina è un tetradecapeptide ciclico. PRH e PIH non sono ancora caratterizzati dal punto di vista chimico.
Questi ormoni sono generalmente attivi in quantità dell'ordine dei nanogrammi, cioè di gran lunga inferiori a quelle degli ormoni fin qui considerati; si comprende quindi come l'isolamento e la caratterizzazione di queste sostanze abbiano richiesto uno sforzo considerevole, data la necessità di ottenere quantità sufficienti di materiale. Tutti questi fattori possono essere inquadrati, dal punto di vista chimico, in un gruppo di piccoli polipeptidi simili tra loro, che presentano, per quanto oggi se ne sappia, il gruppo -NH2 terminale protetto e il gruppo carbossilico terminale sotto forma di ammide.
Il TRH è rapidamente inattivato nel plasma da un enzima. Ogni fattore liberatore è specifico nello stimolare la secrezione di un particolare ormone ipofisario; per la maggior parte dei fattori studiati non vi è, nei Mammiferi, specificità di specie. Sarebbe di grande interesse determinare quali sono gli esseri che per primi nella scala filogenetica posseggono tali peptidi; è possibile che questi siano presenti anche in specie prive di ipotalamo: in tal caso sarebbe interessante determinarne la specifica funzione. Sarebbe anche interessante vedere se tali peptidi si formino nelle miscele di proteoidi ottenute in condizioni che simulano lo stato di prebiosi sulla Terra. Finora non vi sono prove che i fattori liberatori ipotalamici, oltre a stimolare la secrezione degli ormoni dell'ipofisi anteriore, partecipino in qualche modo alla loro biosintesi. Per svolgere la loro azione essi richiedono la presenza di Ca2+ e di Na+; la loro attività non è ostacolata da inibitori della trascrizione o della traduzione, come l'actinomicina D, la puromicina o la cicloesimmide.
b) Ormoni adenoipofisari.
L'ipofisi è costituita da due parti, l'anteriore (adenoipofisi) e la posteriore (neuroipofisi), come si vede nella fig. 7. L'ipofisi anteriore secerne da 7 a 10 ormoni, che a loro volta, direttamente o indirettamente, regolano molti altri organi (organi-bersaglio), come è indicato parzialmente nella fig. 6. L'ipofisi anteriore deriva da un'estroflessione del tetto della primitiva cavità orale; il foglietto di cellule estroflesso costituisce la parte epiteliale destinata a formare la ghiandola, mentre un'estroflessione dal precursore del pavimento del terzo ventricolo dà origine alla parte nervosa destinata a formare l'ipofisi posteriore. I sette ormoni secreti dall'ipofisi anteriore derivano dalla pars tuberalis e dalla pars distalis della ghiandola; l'ormone stimolante i melanociti (MSH) viene prodotto dalla pars intermedia, che in alcune specie rappresenta una porzione distinta della ghiandola. Nella fig. 5 sono posti in risalto i numerosi capillari che irrorano le cellule secretrici dell'ipofisi anteriore.
Nel lobo anteriore della ghiandola pituitaria sono presenti diversi tipi di cellule: forse ve ne sono 6, ciascuno responsabile di una secrezione diversa; il diametro dei granuli di secrezione nelle diverse cellule varia da 150 a 600 nm.
Come già ricordato, l'ipotalamo regola l'attività dell'ipofisi anteriore mediante i fattori liberatori: questi ormoni sono trasportati alle cellule dell'ipofisi anteriore (che costituiscono il bersaglio) per mezzo di uno speciale sistema vascolare, mentre sembra che le connessioni nervose esistenti tra ipotalamo e ipofisi non abbiano importanza. Ogni tipo di cellula dell'ipofisi anteriore riceve istruzioni da almeno un regolatore chimico, di cui si è parlato a proposito degli ormoni dell'ipotalamo.
Gli ormoni elaborati dall'ipofisi anteriore sono: ormone della crescita (GH); ormone tireostimolante (TSH); la tireostimulina ad azione ritardata (LATS); la sostanza esoftalmizzante (EPS); l'adrenocorticotropina (ACTH); la prolattina; le gonadotropine: follicolo-stimulina (FSH), luteostimulina (LH) od ormone stimolante le cellule interstiziali (ICSH); le lipotropine (LPH); l'ormone stimolante i melanociti (MSH).
Ormone della crescita o somatotropo. - L'ormone della crescita (OH, o somatotropo, STH) è un ormone polipeptidico dal peso molecolare di circa 22.000. Nella fig. 10 è riportata la sequenza in amminoacidi del GH umano: sembra che vi siano considerevoli rassomiglianze tra ormoni somatotropi di specie diverse, dal momento che un anticorpo preparato contro uno di essi reagisce in modo cr0- ciato con numerosi altri. Ciò nonostante ormoni di differenti specie presentano pesi molecolari notevolmente diversi, per lo più intorno a 22.000 o a 46.000: una spiegazione di questa disparità può essere trovata in fenomeni di polimerizzazione di una struttura protomerica, ma questa interpretazione attende una conferma.
Secrezione dell'ormone della crescita. - La regolazione della secrezione dell'ormone della crescita è complessa ma, come abbiamo già visto, è effettuata in parte dal fattore liberatore dell'ormone somatotropo (GRH).
Si può tracciare lo schema di questa regolazione nel modo seguente:

L'effetto feedback dell'insulina in circolo sull'ipotalamo può stimolare la produzione di ormone della crescita da parte dell'ipofisi anteriore, mentre la somministrazione di glucosio blocca, secondo le previsioni, la produzione dell'ormone. Lesioni ipotalamiche impediscono questo effetto dell'insulina, che probabilmente è mediato dalla produzione di catecolammine, come la noradrenalina o la L-dopammina (v. sopra, cap. 2, È a). Farmaci capaci di svuotare i depositi di catecolammine nel sistema nervoso centrale, come per esempio la reserpina, interferiscono con la capacità dell'insulina di causare la secrezione di ormone somatotropo.
Effetti dell'ormone della crescita. - Questo ormone è capace d'indurre la crescita di tutti i principali organi viscerali; a questo effetto finale concorrono una positivizzazione del bilancio dell'azoto, una riduzione dell'azoto amminico circolante, un aumento del trasporto di amminoacidi attraverso le membrane cellulari e un aumento della sintesi proteica.
L'ormone probabilmente agisce a livello subcellulare, nei processi di trascrizione e di traduzione che precedono la sintesi proteica: l'RNA-polimerasi del fegato, enzima responsabile della trascrizione dell'RNA a partire dal gene, viene stimolata nelle 24 ore successive alla somministrazione in animali trattati con ormone somatotropo. Tra i diversi tipi di RNA, sia quello solubile (tRNA) sia il messaggero (mRNA) vengono prodotti in maggiore quantità per azione dell'ormone.
L'ormone della crescita è capace di ristabilire parzialmente la capacità dei ribosomi d'incorporare amminoacidi, quando tale capacità sia stata perduta in seguito a ipofisectomia.
Alcune delle principali funzioni dell'ormone somatotropo nello stimolare processi metabolici sono riportate qui di seguito:
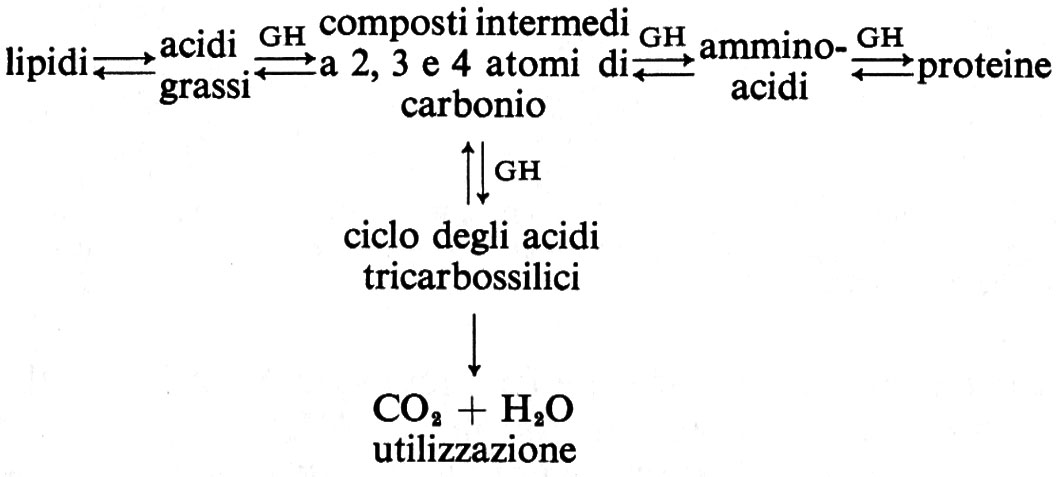
Altri effetti generali di questo ormone sono l'aumento del riassorbimento renale di ioni fosfato e solfato, l'aumento del trasporto di acqua nei mitocondri, l'aumento della incorporazione di amminoacidi, già menzionato, nei muscoli scheletrici, nel cuore, nel fegato, oltre che nel diaframma e nel tessuto adiposo, in cui l'ormone provoca anche un incremento del contenuto glucidico. Non è chiaro se e fino a qual punto l'AMP ciclico funga da mediatore dell'ormone della crescita.
Ormone tireostimolante (TSH). - Questo ormone è una glicoproteina con peso molecolare di circa 28.000 dalton; la frazione glucidica legata covalentemente alla proteina è formata da un singolo oligosaccaride contenente mannosio, glucosammina, galattosammina e fucosio. L'ormone tireostimolante è probabilmente assai simile all'ormone luteinizzante, in quanto è difficile separarli; ne consegue che il TSH non è facilmente ottenibile in forma pura. La composizione in amminoacidi è diversa nelle diverse specie in quanto, per esempio, anticorpi anti-TSH bovino ottenuti da coniglio danno una debole reazione crociata con l'ormone umano.
Secrezione del TSH. - La secrezione di ormone tireostimolante è regolata a due livelli: in primo luogo l'ormone tiroideo prodotto dalla tiroide interagisce con l'ipofisi anteriore; inoltre, intervengono fattori ambientali come l'alimentazione, gli effetti sul centro della termoregolazione e l'azione di feedback sull'ipotalamo dell'ormone tiroideo circolante, che è responsabile della produzione del fattore liberatore dell'ormone tireostimolante. Questi rapporti possono essere schematizzati cosi:

Gli effetti a feedback dell'ormone tiroideo sull'ipotalamo non sono del tutto accertati, ma la loro esistenza è probabile, per analogia con altri sistemi.
Effetti del TSH. - Gli effetti dell'ormone tireotropo sulla ghiandola tiroide sono i seguenti: l'ormone aumenta il flusso sanguigno nella tiroide e quindi induce una maggiore velocità di demolizione dei depositi tireoglobulinici di ormone tiroideo, con aumentata secrezione di ormone tiroideo nel sangue; contemporaneamente alla degradazione della tireoglobulina con immissione in circolo di ormone tiroideo vi è un'aumentata produzione di quest'ultimo. L'ormone ha anche importanti effetti in vivo sul trasporto di vari metaboliti in alcuni organi: in particolare provoca un aumento dell'incorporazione di ioduro nella tiroide, di amminoacidi in fettine di pelle, di muscolo, di fegato e di tiroide e infine di zuccheri in fettine di tiroide.
TSH e AMP ciclico. - L'azione dell'ormone tireotropo, come quella di altri ormoni peptidici, sembra essere mediata dall'AMP ciclico, particolarmente per ciò che riguarda i suoi principali effetti sulla ghiandola tiroide: lo stimolo della produzione dell'ormone tiroideo e della sua secrezione nel sangue. Ciò potrebbe avvenire con questo meccanismo sequenziale: TSH (in circolo) → recettore di membrana della ghiandola tiroide → TSH legato alla membrana → attivazione della adenilciclasi → conversione dell'ATP in AMP ciclico → attacco dell'AMP ciclico a una subunità regolatrice della fosfochinasi → distacco da parte dell'AMP ciclico della subunità regolatrice della chinasi → la chinasi attivata catalizza la fosforilazione di una proteina che può provocare un aumento della biosintesi e/o della demolizione della tireoglobulina.
Un'ipotesi alternativa è che la subunità regolatrice della fosfochinasi proteica, una volta legata all'AMP ciclico, possa agire, dopo il distacco dall'enzima, da derepressore sul genoma, aumentando la velocità di sintesi dell'ormone tiroideo; tuttavia quest'ultima ipotesi non è ancora suffragata da dati sperimentali.
Stimolatore della tiroide ad azione ritardata. - Il plasma di pazienti ipertiroidei contiene una sostanza che provoca il distacco, dalla tiroide di ratto, di iodio organico marcato. In confronto all'azione del TSH, che richiede dalle 2 alle 3 ore, il siero di pazienti ipertiroidei agisce in 12-16 ore: da qui il nome della sostanza, ‛stimolatore ad azione ritardata'. Il principio attivo è una proteina a coefficiente di sedimentazione 7S, appartenente alla frazione α-globulinica del sangue, attiva di per sé più che in unione al TSH. Tale proteina è formata da due catene polipeptidiche, una delle quali è attiva. Non è presente nell'ipofisi. È stato ipotizzato che il LATS (Long Acting Thyroid Stimulator) sia un autoanticorpo diretto contro qualche componente della tiroide, in quanto esso perde attività dopo incubazione con fettine di ghiandola tiroide.
Agente esoftalmizzante. - Quando in pazienti affetti da morbo di Graves si asporta chirurgicamente la tiroide, spesso si osserva come conseguenza una protrusione dei bulbi oculari (esoftalmo); lo stesso effetto si produce nelle cavie dopo tiroidectomia. Preparazioni molto pure di TSH non producono questo effetto; lo producono invece preparazioni non purificate. È probabile quindi che la sostanza che produce l'esoftalmo (EPS, Exosphthalmos Producing Substance) sia presente nell'ipofisi anteriore e sia in qualche modo simile al TSH, ma probabilmente distinta da esso. Questo effetto può essere spiegato come l'instaurarsi di una condizione anomala per la cessazione di un feedback negativo tiroideo: ipofisi anteriore TSH ghiandola ormone tiroideo
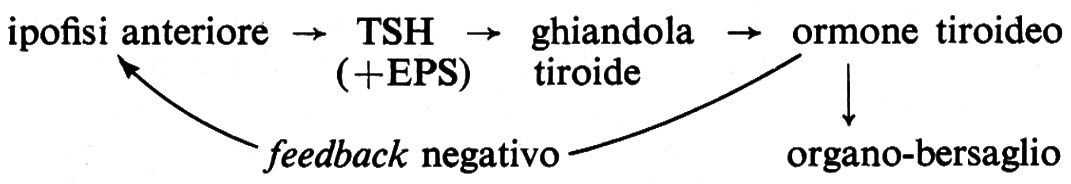
Ancora si conosce ben poco di questo fattore.
Adrenocorticotropina (ACTH). - L'adrenocorticotropina è formata da una singola catena polipeptidica dal peso molecolare di circa 4.500 dalton; la sua molecola è costituita da 39 residui di amminoacidi. L'isolamento e lo studio della struttura di questo ormone hanno richiesto un notevole sforzo. La struttura dell'ormone è riportata nella fig. 11: è chiaro che vi sono notevoli analogie tra le diverse specie; le differenze sono infatti contenute tra i residui amminoacidici 25-33, che rappresentano una parte della molecola non importante per l'attività biologica.
Secrezione dell'ACTH e suo controllo a feedback. - L'ACTH ha come bersaglio la ghiandola surrenale, nella quale stimola la produzione degli ormoni corticosteroidi. Il meccanismo di secrezione e di controllo a feedback dell'ACTH non è dissimile dagli altri già descritti: stimoli ambientali nocivi → mesencefalo → impulsi nervosi → stimoli ('stressori') → ipotalamo → azione degli stimoli nervosi → diminuita concentrazione dell'ormone liberatore della corticotropina (CRH) → passaggio del CRH nell'ipofisi anteriore accumulo di CRH nell'ipofisi anteriore → secrezione di ACTH nel sangue → inibizione a feedback dell'ACTH sull'ipotalamo per interrompere la produzione di CRH l→ 'ACTH passa dal sangue alla corteccia surrenale (bersaglio) → accumulo di ACTH nella corteccia surrenale stimolazione della sintesi e della secrezione di glucocorticoidi surrenalici passaggio in circolo di ormoni

Sembra che l'azione dell'ACTH sulle cellule della corteccia surrenale si esplichi con la mediazione dell'AMP ciclico: ACTH ematico sito di attacco per il peptide sulla cellula surrenale (membrana) → complesso → stimolazione dell'adenilciclasi → conversione dell'ATP in AMP ciclico → attivazione di una fosfoproteinchinasi fosforilazione di una proteina specifica aumento della biosintesi degli steroidi.
L'effetto specifico finale potrebbe ragionevolmente aversi o per un aumento della sintesi di enzimi-chiave per la conversione del colesterolo in pregnenolone o per un effetto diretto sulla reazione limitante. La maggior parte di questo meccanismo è ipotetica, tuttavia è chiara la mediazione dell'azione dell'ACTH da parte dell'AMP ciclico.
Lo schema di regolazione della secrezione di ACTH è confermato da prove sperimentali: per esempio, steroidi impiantati nel mesencefalo causano una diminuzione della secrezione di glucocorticoidi da parte della corteccia surrenale; questo effetto può essere mediato, in tutto o in parte, dall'inibizione della produzione di ormone liberatore della corticotropina da parte dell'ipotalamo, impedendo così al segnale di arrivare nell'ipofisi anteriore e di stimolare la produzione di ACTH. I glucocorticoidi possono anche agire direttamente sulla secrezione di ACTH da parte dell'ipofisi anteriore, dal momento che diminuiscono il consumo di ossigeno in cellule pituitarie in coltura. Tuttavia questo feedback degli steroidi sull'ipofisi sembra di importanza secondaria rispetto a quello, più importante, sull'ipotalamo. Questi meccanismi a feedback richiedono comunque uno studio più sistematico.
Effetti dell'ACTH. - L'azione principale dell'ormone adrenocorticotropo si esplica, come indicato dal nome, a livello della corteccia surrenale, in particolare a carico dei due strati più interni di cellule, la zona fascicolare e la zona reticolare (ben identificabili nella fig. 12), ove avviene la sintesi e la secrezione dei glucocorticoidi. Lo strato più esterno, la zona glomerulare, non è molto influenzato dall'ormone adrenocorticotropo: in questo strato di cellule viene sintetizzato e secreto l'aldosterone, steroide che regola l'equilibrio salino. L'azione principale dell'ACTH consiste nello stimolare la trasformazione del colesterolo in glucocorticoidi e la secrezione di questi ultimi nel torrente circolatorio. Si può supporre che per azione dell'ACTH sulla cellula surrenale si abbia la seguente serie di eventi: ACTH circolante → sito di legame della membrana cellulare → formazione del complesso ACTH-membrana → stimolazione dell'adenilciclasi → conversione dell'ATP in AMP ciclico → legame dell'AMP ciclico alla subunità regolatrice di una proteinfosfochinasi → fosforilazione di una proteina specifica che determina un aumento nella velocità di sintesi dei glucocorticoidi, oppure, a seconda del livello di produzione dell'AMP ciclico: ATP → AMP ciclico → si lega a una proteina citoplasmatica → va nel nucleo → si lega al DNA → induce la sintesi di enzimi limitanti specifici per la sintesi dei glucocorticoidi.
Tutte queste azioni sono attualmente oggetto di ipotesi, ma devono essere considerate come possibili meccanismi d'azione dell'ACTH.
Prolattina. - La prolattina, altro ormone polipeptidico dell'ipofisi anteriore, ha un peso molecolare di 23.000 dalton. È molto simile all'ormone della crescita: nell'uomo, i due ormoni non possono essere separati, mentre ciò è possibile per gli ormoni estratti dall'ipofisi anteriore di altre specie.
Secrezione della prolattina. - Sembra che l'ipotalamo inibisca la secrezione di prolattina da parte dell'ipofisi anteriore elaborando una sostanza detta ormone inibitore della secrezione di prolattina (PIH). La secrezione di prolattina è controllata da altri ormoni, cosa del resto prevedibile data la dipendenza multiormonale del processo della lattazione: in particolare estrogeni e progesterone sembrano agire per feedback sull'ipotalamo per aumentare la produzione di PIH. Lo stimolo dato dal succhiamento, invece, causa una diminuzione del livello di PIH nell'ipotalamo, cosi che più frequente è il succhiamento, minore è il PIH presente nell'ipotalamo. Questo ormone, la cui formazione dipende pertanto da vari parametri, arriva dall'ipotalamo all'ipofisi anteriore, dove il suo accumulo produce una diminuzione della prolattina. D'altra parte gli estrogeni secreti dall'ovaio hanno un effetto bifasico sull'ipofisi anteriore: all'aumentare della concentrazione di estrogeni nella ghiandola, vi è un aumento del contenuto di prolattina fino a un massimo; per concentrazioni ancora più elevate di estrogeno nell'ipofisi anteriore, la quantità di prolattina diminuisce. È probabile che, in gran parte, la diminuzione di prolattina sia dovuta alla sua immissione in circolo; dal torrente circolatorio questo ormone viene concentrato negli organi-bersaglio, la ghiandola mammaria e anche il corpo luteo (nel ratto), e, all'aumentare della sua concentrazione nella ghiandola mammaria, aumenta la produzione di latte. La ghiandola mammaria funzionante è un sistema molto complesso, sottoposto al controllo di altri ormoni, come l'ACTH, il OH, la tiroxina, l'insulina, il paratormone e gli estrogeni. Recenti ricerche condotte su organocolture di ghiandola mammaria hanno permesso di determinare accuratamente il ruolo dei diversi ormoni: infatti la presenza d'insulina, cortisolo e prolattina nel mezzo d'incubazione induce i nidi irregolari di cellule dell'epitelio mammario a disporsi ordinatamente intorno a lumi alveolari allargati che si riempiono di secreto e provoca un aumento numerico delle cellule funzionalmente differenziate svelato dall'aumentata produzione di proteine proprie del latte, quali la caseina e l'enzima lattosiosintetasi. L'insulina da sola è in grado di mantenere il grado iniziale di produzione di caseina e di provocare un leggero aumento di attività lattosio-sintetasica; quando oltre all'insulina viene aggiunta prolattina, si riscontra un moderato aumento dei due componenti proteici della lattosiosintetasi; solo quando tutti e tre gli ormoni - insulina, cortisolo e prolattina - sono contemporaneamente presenti si ha il massimo incremento quantitativo delle proteine proprie del latte e della lattosiosintetasi. Queste proteine costituiscono senza dubbio un indice accurato del differenziamento cellulare, in quanto nello sviluppo in vivo della ghiandola mammaria la loro velocità di formazione aumenta durante la gravidanza e l'allattamento. L'azione dell'insulina, del cortisolo e della prolattina sulle cellule mammarie che si differenziano in coltura è schematizzata nella fig. 13.
Azione della prolattina. - Sembra che inizialmente la prolattina possa legarsi a un recettore della membrana cellulare, poichi l'ormone legato covalentemente a microsfere di sefarosio (che non possono entrare nella cellula) interagisce con la membrana esterna delle cellule mammarie in coltura e stimola la sintesi di RNA nucleare. Non è del tutto chiaro quali siano gli eventi molecolari che culminano in un'aumentata sintesi di proteine del latte, benché si possano indicare, come nella fig. 14, alcune ipotesi generiche.
La prolattina esplica altre attività in Vertebrati non Mammiferi: così nel piccione il gozzo secerne un ‛latte del gozzo' in risposta all'ormone; nel girino l'ormone inibisce il riassorbimento della coda, normale stadio di transizione della metamorfosi, e inibisce anche l'escrezione di urea; in una forma terrestre di tritone la prolattina stimola una attrazione verso l'acqua e in alcuni pesci ossei causa melanogenesi.
Gonadotropine. - Tra gli ormoni gonadotropi, che hanno peso molecolare di circa 30.000 dalton, si possono distinguere cinque sostanze diverse: ormone luteinizzante (LH), ormone follicolostimolante (FSH), gonadotropina cononica umana (HCG), gonadotropina serica di cavalla gravida (PMSO) e gonadotropina umana della menopausa (HMG). Sono chimicamente simili all'ormone tireostimolante (TSH), già trattato (v. sopra, È b).
L'ormone follicolostimolante e l'ormone luteinizzante, come l'ormone tireotropo, vengono estratti dal tessuto ipofisario; gli altri vengono ottenuti dalle urine (HCG e HMG) o dal siero (PMSG). L'ormone luteinizzante è formato da due subunità non identiche, ciascuna con peso molecolare di circa 15.000 dalton e con diversa composizione in amminoacidi; la molecola nel suo insieme presenta una struttura non elicoidale, stabilizzata da ponti disolfuro.
L'ormone follicolostimolante viene inattivato dalla neuraminidasi; ciò indica che esso contiene un oligosaccaride, probabilmente l'acido sialico, e che questo componente glucidico è essenziale per la sua attività ormonale. La gonadotropina umana della menopausa (HMG) è costituita da una molecola provvista di una duplice azione: ciò è suggerito dal fatto che essa lungo tutto il processo di purificazione presenta uguale attività luteostimolante e follicolostimolante.
La gonadotropina corionica umana (HCG) deriva dalla placenta e la gonadotropina serica di cavalla gravida (PMSG) dall'endometrio uterino: entrambe sono ricche di componenti glucidici, fino al 30% 50% del peso secco; tuttavia può anche darsi che questi carboidrati siano contaminazioni residuate nel corso della purificazione.
Funzioni dell'FSH e dell'LH. - Gli ormoni FSH e LH agiscono in modo complementare: la spermatogenesi è possibile solo se entrambi gli ormoni sono presenti e secreti contemporaneamente, poiché il primo (FSH) è necessario negli stadi intermedi della spermatogenesi e il secondo (LH, detto anche, nel maschio, ormone stimolatore delle cellule interstiziali, ICSH) provoca la secrezione di androgeni, che esplicano un ruolo importante nel processo di maturazione dei gameti maschili. Nella femmina l'ovulazione avviene solo se FSH e LH sono presenti sequenzialmente.
Secrezione dell'LH nel maschio. - Il controllo della secrezione di LH (ICSH), come quello degli altri ormoni dell'ipofisi anteriore, implica un meccanismo abbastanza complesso: stimoli nervosi provenienti dal sistema nervoso centrale giungono all'ipotalamo; questo produce il relativo fattore liberatore dell'ormone luteinizzante, fattore che si accumula nelle specifiche cellule dell'ipofisi anteriore, contenenti LH. All'aumentare della concentrazione di fattore liberatore, anche la quantità di LH prodotto e immesso in circolo aumenta ; dal circolo l'ormone luteinizzante raggiunge le proprie cellule-bersaglio. Le cellule-bersaglio dell'ICSH sono, nel maschio, le cellule interstiziali del testicolo e i tubuli seminiferi, mentre la follicolostimolina viene captata principalmente dagli spermatociti. All'aumentare di LH nelle cellule interstiziali del testicolo, aumenta il livello di testosterone, che viene versato in circolo ; questo ormone raggiunge le cellule a cui è destinato, come si vedrà in seguito, e può esercitare un'inibizione a feedback sull'ipotalamo, bloccando la produzione di fattore liberatore dell'ormone luteinizzante.
Il testosterone agisce congiuntamente all'FSH sugli spermatociti, aumentando il numero di spermatozoi prodotti da queste cellule. I tubuli seminiferi possono anche sintetizzare estrogeni, che aumentano in risposta a un incremento della concentrazione di ormone luteinizzante nei tubuli seminiferi. Questi estrogeni sono immessi nel torrente circolatorio sia dall'organo maschile sia da quello femminile e possono agire per feedback sull'ipotalamo in modo del tutto analogo a quello del testosterone, inibendo la liberazione di fattore liberatore dell'FSH dall'ipotalamo stesso.
Secrezione dell'LH e dell'FSH nella femmina. - I meccanismi di controllo e di regolazione della secrezione di gonadotropine nella femmina sono anch'essi molto complicati.
Impulsi nervosi arrivano all'ipotalamo causando la liberazione di FRH, fattore liberatore dell'ormone follicolostimolante, la cui quantità aumenta localmente in proporzione all'incremento degli stimoli nervosi. L'FRH liberato dall'ipotalamo è captato dalle cellule specifiche dell'ipofisi anteriore, dove avviene la sintesi e l'accumulo dell'ormone follicolostimolante. All'aumentare di FRH una quantità crescente di FSH si accumula nelle cellule dell'ipofisi anteriore e viene poi riversata in circolo.
Sia gli impulsi nervosi, sia gli ormoni estrogeni e progestinici circolanti (fra cui il 17α-idrossiprogesterone) possono agire per feedback sull'ipotalamo provocando la produzione del fattore liberatore di LH. Tale fattore, come abbiamo già detto, si accumula nell'ipofisi anteriore, che risponde con la produzione di LH da parte delle cellule a ciò deputate e con la successiva immissione dell'ormone nel torrente circolatorio. Come conseguenza di questa azione i livelli di FSH e LH circolanti vengono aumentati dagli impulsi nervosi, dagli estrogeni, dal progesterone e dal 17α-idrossiprogesterone.
Una volta entrati in circolo, entrambi gli ormoni ipofi- san, FSH e LH, si concentrano nell'ovaio. In tal modo l'aumento della concentrazione dei due ormoni gonadotropi determina un aumento della secrezione di estrogeni e l'accrescimento cellulare. Gli estrogeni immessi nel torrente sanguigno agiscono per feedback sull'ipotalamo, nel modo che abbiamo sopra descritto.
L'ormone LH si concentra anche nel corpo luteo e ivi stimola la secrezione del progesterone, che a sua volta viene immesso in circolo e agisce con meccanismo di feedback sull'ipotalamo. Queste interrelazioni completano i meccanismi di equilibrio fra ipotalamo e ovaio, grazie all'azione di feedback mediata dall'ipofisi anteriore, che è responsabile della secrezione dei due importanti ormoni FSH e LH. In alcuni casi, a seconda della concentrazione di 17-α-idrossiprogesterone o dello stesso progesterone, l'azione sull'ipotalamo può essere di inibizione o di stimolo. Tuttavia non si discuteranno in questo articolo i particolari di tali fenomeni, come non si presenteranno i dati, del resto largamente prevedibili, che permettono di ritenere che gli ormoni steroidi possano agire direttamente sull'ipofisi anteriore.
Effetti dell'LH e dell'FSH. - Il ruolo dell'FSH nel maschio non è molto chiaro, in quanto tale ormone, da solo, non è in grado, in ratti ipofisectomizzati, di mantenere integro l'epitelio germinativo, né di conservare invariato il peso dei testicoli. Quando però LH viene somministrato insieme a FSH, nel rapporto di 1 a 400, i tessuti suddetti restano vitali e, in base a questo dato, si pensa che sia essenziale l'azione combinata dei due ormoni, che comporterebbe la stimolazione della produzione di androgeni, necessari per le ultime fasi della spermatogenesi.
Nelle femmine l'ormone follicolostimolante agisce sui follicoli in accrescimento stimolando la loro maturazione.
L'ormone stimolatore delle cellule interstiziali (ICSH, cioè l'equivalente nel maschio dell'ormone LH) provoca la secrezione di testosterone da parte delle cellule interstiziali.
Nella femmina l'ovulazione può avvenire spontaneamente, nel corso di un ciclo, o in risposta a uno stimolo specifico. Molto probabilmente l'induzione della sintesi di steroidi a opera dell'LH o ICSH comporta la mediazione dell'AMP ciclico, analogamente a quanto abbiamo visto in precedenza, per esempio nel caso dell'adrenalina.
Una più completa esposizione dell'azione degli ormoni sessuali si troverà nel paragrafo a essi dedicato (v. sotto, cap. 4, È b).
Lipotropine. - Sembra che l'ipofisi anteriore contenga sostanze in grado di mobilizzare i grassi. Sono polipeptidi, due dei quali sono stati isolati: il primo, detto β-lipotropina, ha peso molecolare di circa 9.500 dalton ed è formato da 90 amminoacidi; il secondo, o γ-lipotropina, ha peso molecolare di 5.800 dalton ed è formato da circa 58 amminoacidi. La γ-lipotropina sembra essere un frammento della β-lipotropina; entrambi questi polipeptidi contengono un eptapeptide basico centrale, presente anche nell'ACTH e nell'MSH. Questo eptapeptide mostra attività melanocitastimolanti; l'attività MSH-simile della β-lipotropina è circa il doppio di quella della γ-lipotropina.
Ben poco si sa della regolazione della secrezione di questi ormoni e del ruolo specifico che essi hanno nei Vertebrati.
Ormone stimolante i melanociti (MSH) . - Questo ormone è presente nell'ipofisi anteriore, o nel lobo intermedio in quei Vertebrati che possiedono questa struttura. Esso è presente sotto due forme, indicate come α- e β-MSH: il primo è un tridecapeptide, ha attività biologica maggiore del β-MSH e si trova in tutti i Mammiferi; il β-MSH presenta una catena di lunghezza variabile e di variabile sequenza di amminoacidi. Come detto in precedenza a proposito delle lipotropine, tutte le molecole di MSH presentano una parte centrale comune, costituita da 7 amminoacidi, che si riscontra anche nell'ACTH e nelle lipotropine.
Effetti e controllo dell'MSH. - L'ormone stimolante i melanociti provoca la dispersione del pigmento nella pelle di rana, mentre la concentrazione del pigmento può essere controllata da un altro ormone, la melatonina, che è prodotto dalla ghiandola pineale. Il controllo della secrezione dell'ormone stimolante i melanociti può essere schematizzato nel modo seguente. La melatonina prodotta dalla ghiandola pineale va in circolo e si concentra nella regione sopraottica. La concentrazione dell'ormone inibente la melatonina cade quando la concentrazione di melatonina aumenta nella regione sopraottica, mentre l'ormone mibente la liberazione di MSH, l'MIH, si concentra nel lobo intermedio dell'ipofisi. L'MSH viene accumulato dai melanociti e se la sua concentrazione aumenta si verifica un conseguente aumento percentuale della dispersione del pigmento; al contrario, la percentuale di dispersione del pigmento nella pelle di rana diminuisce quando nei melanociti si accumula la melatonina formata nella ghiandola pineale. Così viene regolata la dispersione del pigmento, in quanto nell'epifisi la concentrazione di melatonina è inversamente proporzionale all'intensità della luce: tanto minore è questa, tanto maggiore è la quantità di melatonina presente nella ghiandola e quindi secreta nel torrente circolatorio per essere captata dai melanociti.
Le cellule contenenti melanina, nei Vertebrati a sangue freddo, sono usualmente di due tipi. La melanina è conte- nuta nei melanofori e il colore della pelle è determinato dalla quantità di pigmento distribuito nel citoplasma. L'α-MSH aumenta la sintesi di melanina sia nei melanofori sia nei melanociti e la dispersione della melanina nei melanofori. Il ruolo dell'α- e del β-MSH nei Mammiferi non è chiaro ; invece nei Vertebrati a sangue freddo, come i Pesci, gli Anfibi e i Rettili, l'α-MSH ha una funzione protettiva, in quanto questi animali hanno la capacità di diventare più chiari su uno sfondo chiaro o più scuri su uno sfondo scuro.
Anche l'ACTH agendo come l'α-MSH può provocare un imbrunimento della pelle, come per esempio nel morbo di Addison ; ciò non sorprende considerando che questi ormoni hanno un eptapeptide centrale comune.
Per quanto negli organismi superiori la funzione dell'ormone stimolante i melanociti non sia del tutto chiara come nel caso degli Anfibi, i dati fin qui acquisiti fanno pensare che la somministrazione di questo ormone abbia effetto sul sistema nervoso centrale, in particolare sul comportamento e sulla capacità di apprendimento.
c) Ormoni neuroipofisari.
Ossitocina e vasopressina. - La struttura della neuroipofisi è presentata nella fig. 7. Mentre le cellule secretrici dell'adenoipofisi sono largamente irrorate da capillari, la neuroipofisi contiene fibre nervose e pituiciti: questi ultimi sono cellule connettivali del tessuto nervoso generalmente indicate col termine di neuroglia. L'ipotalamo invia alla neuroipofisi filamenti nervosi che costituiscono la via ipotalamo-ipofisaria e che terminano in prossimità dei capillari della neuroipofisi.
I principali ormoni che nei Mammiferi superiori vengono prodotti nella neuroipofisi sono l'ossitocina e la vasopressina (arginina-vasopressina e lisina-vasopressina).
Questi ormoni vengono inviati, lungo gli assoni dei neuroni che compongono i nuclei ipotalamici, in prossimità dei capillari della neuroipofisi, ove nella parte terminale degli assoni si accumulano in granuli da cui devono essere staccati per passare in circolo. I granuli, detti neurofisine, possono partecipare al trasporto e all'immagazzinamento degli ormoni; sono state isolate tre diverse proteine capaci di legare gli ormoni: le neurofisine 1, 2 e 3.
Ciascun ormone viene secreto indipendentemente dal proprio centro ipotalamico: il nucleo sopraottico viene indicato come luogo di produzione della vasopressina, mentre il nucleo paraventricolare sarebbe deputato all'elaborazione dell'ossitocina.
Aspetti strutturali. - Come accennato in precedenza, l'ossitocina e la vasopressina sono presenti nell'ipofisi posteriore di tutti i Mammiferi (più precisamente, questi animali possiedono l'arginina-vasopressina, salvo i suini che hanno la lisina-vasopressina); queste sostanze sono presenti anche nell'ipotalamo e possono quindi essere considerate ormoni dell'ipofisi posteriore o dell'ipotalamo. Nei Pesci cartilaginei od ossei e negli Anfibi sono presenti altri ormoni, come la mesotocina e la vasotocina. Tutte queste sostanze ormonali contengono un anello pentapeptidico chiuso da un ponte disolfuro tra due mezze molecole di cistina. Nella fig. 15 è riportata la struttura dell'ossitocina; l'arginina-vasopressina ha una struttura simile, ma, rispetto all'ossitocina, in posizione 4 invece di una isoleucina contiene una fenilalanina e in posizione 9 invece di una leucina presenta un'arginina. In quest'ultima posizione, nella lisina-vasopressina si riscontra una lisina invece che un'arginina.
Le strutture degli altri ormoni presenti negli organismi inferiori, come la vasotocina, l'ittiotocina, la mesotocina e la glumitocina, sono del tutto simili a quella dell'ossitocina, eccetto che per sostituzioni nelle posizioni 5 e 9.
Effetti degli ormoni dell'ipofisi posteriore. - Questi ormoni hanno principalmente un'azione antidiuretica e, a dosi maggiori, provocano la contrazione dei muscoli lisci, in particolare quelli delle pareti vasali. L'ossitocina è importante solo nella femmina gravida e nutrice, per la contrazione della muscolatura uterina e la eiezione del latte. La vasopressina ha un effetto considerevole sulla permeabilità all'acqua e all'urea dei nefroni renali degli Anfibi inferiori; nei Mammiferi superiori la mancanza di questo ormone provoca il diabete insipido, malattia che comporta l'eliminazione con le urine, e quindi l'assunzione, di grandi quantità d'acqua.
Questi ormoni peptidici, similmente ad altri ormoni della stessa natura chimica, possono influire sull'attività della adenilciclasi a livello delle membrane delle cellule-bersaglio, provocando l'attivazione di una fosfoproteinchinasi responsabile dell'effetto ormonico generale.
La regolazione della secrezione della vasopressina è effettuata probabilmente da numerosi fattori che agiscono sull'ipotalamo. Uno di essi, forse il piu importante, è costituito dall'alterazione della pressione osmotica plasmatica; tuttavia anche variazioni del volume o della pressione del sangue potrebbero essere fattori che intervengono in questo processo di regolazione.
In quanto all'ossitocina, come si è detto, questo ormone non ha un ruolo ben definito nel maschio, mentre nella femmina la sua azione è ben nota: in animali gravidi l'ossitocina induce l'espletamento del parto: il tasso plasmatico di ossitocina aumenta significativamente nel primo stadio del travaglio per ritornare ai livelli normali una volta espletato il parto.
L'ossitocina è molto probabilmente un fattore importante anche per l'allattamento, dal momento che in sua assenza non si ha eiezione di latte da parte della ghiandola mammaria.
Vasopressina e controllo del riassorbimento idrico. - Accenneremo brevemente al processo di regolazione degli effetti della vasopressina per ciò che riguarda il controllo del riassorbimento dell'acqua. Quando stimoli dolorifici o emozionali giungono alle aree superiori del cervello, le scariche nervose aumentano di numero interessando direttamente l'ipotalamo, cosicché la produzione di ormone antidiuretico aumenta. Attraverso il tratto ipotalamo-ipofisario la vasopressina giunge alla neuroipofisi, ove la sua concentrazione si accresce proporzionalmente all'intensità dello stimolo; passa quindi in circolo e si accumula nel rene, ove la quantità di acqua riassorbita è in certo qual modo proporzionale alla quantità di ormone antidiuretico accumulato ; il riassorbimento dell'acqua modifica la pressione osmotica del sangue e ciò influenza per feedback l'ipotalamo. Bisogna ricordare però che anche l'intestino può intervenire nel regolare la pressione osmotica del plasma sanguigno eliminando acqua o assorbendo cloruro di sodio.
Nel caso dell'ossitocina, l'ipotalamo è influenzato sia dal progesterone, che diminuisce la quantità di ossitocina, sia dagli stimoli prodotti dalla suzione o dalla distensione della cervice uterina. Questi fattori provocano un aumento dell'ossitocina accumulata attraverso la via ipotalamo-ipofisaria nella neuroipofisi: tale aumento è proporzionale all'intensità della scarica di impulsi nervosi e alla quantità di ormone immesso in circolo. L'ossitocina viene accumulata dall'utero e dalla ghiandola mammaria: l'attività contrattile dell'utero e la quantità di latte emessa dalla ghiandola mammaria sono proporzionali alla quantità di ossitocina accumulata.
d) Insulina e glucagone.
Il pancreas secerne, nell'uomo e in altri Mammiferi superiori, due importanti ormoni responsabili della regolazione del tasso di glucosio circolante: l'insulina e il glucagone. Entrambi questi ormoni sono polipeptidi; la loro struttura è riportata nella fig. 16.
L'insulina viene prodotta nelle cellule β del pancreas, mentre le cellule α elaborano il glucagone. La secrezione di insulina, come si vedrà in seguito, è determinata dal livello di glucosio circolante e l'azione dell'ormone consiste nell'abbassare questo livello; sembra che l'elevarsi del tasso plasmatico di glucosio possa provocare la liberazione di ‛glucagone intestinale', ormone peptidico capace d'interagire con un recettore sito sulla membrana delle cellule β, scatenando una successione di eventi che culminano con la immissione di insulina in circolo.
Per contro, il glucagone è un fattore iperglicemizzante la cui secrezione risponde a una diminuzione del tasso glicemico: la sua azione consiste nell'aumentare la glicemia ed è quindi antagonista dell'insulina.
La struttura dell'insulina è ben nota: questo ormone ha un peso molecolare di circa 5.700 dalton ed è formato da due catene polipeptidiche, A e B, costituite rispettivamente da 21 e da 30 amminoacidi. Le due catene sono unite da ponti disolfuro, com'è indicato nella fig. 16 B. L'insulina deriva da un precursore, la proinsulina, priva di attività ormonale, la cui struttura è anch'essa indicata nella fig. 16 B. La proinsulina viene trasformata in insulina attiva mediante l'idrolisi del peptide di interconnessione tra le due catene. Ancora non si è trovata una precisa funzione fisiologica per questo peptide una volta staccato dall'insulina.
La composizione in amminoacidi del segmento di insulina compreso tra i due ponti disolfuro sembra non essere di fondamentale importanza per l'attività fisiologica. Vi sono notevoli variazioni, da specie a specie, nella sequenza di amminoacidi dell'insulina e ciò deve essere tenuto presente nella terapia con questo ormone. Lo studio dei tumori originati dalle cellule β del pancreas umano ha rivelato che la proinsulina si forma con meccanismi di biosintesi analoghi a quelli delle altre proteine e che l'attivazione della molecola insulinica è correlata all'attività di alcune peptidasi capaci di idrolizzare la catena della proinsulina nei punti indicati nella fig. 16 B.
Il glucagone, la cui struttura è riportata nella fig. 16 A, è formato da 29 amminoacidi, per un peso molecolare di 3.483 dalton. Questo ormone si può ottenere perfettamente separato dall'insulina.
Il diabete mellito è una delle più importanti e frequenti manifestazioni cliniche di disfunzioni ormonali; questa malattia è dovuta a un'alterazione delle cellule β delle insulae di Langerhans e i sintomi che la caratterizzano possono essere eliminati con l'iniezione intramuscolare di un'insulina purificata, la cui struttura sia abbastanza simile a quella dell'ormone umano, in modo da non causare gravi disturbi immunologici.
Secrezione dell'insulina. - Il fattore principale che stimola la secrezione d'insulina da parte delle cellule β è la concentrazione di zucchero nel sangue: l'iperglicemia causa un'aumentata secrezione di insulina; il glucosio è più efficace nel suscitare la risposta rispetto ad altri zuccheri come mannosio, fruttosio o galattosio. L'ingestione di cibo aumenta transitoriamente la glicemia dando il via alla secrezione di insulina; il meccanismo dell'immissione di insulina nel torrente circolatorio in risposta a un aumento della glicemia è schematizzato nella fig. 17. In termini estremamente semplificati, la secrezione di insulina è regolata nel modo seguente: l'aumento della glicemia induce la secrezione di ‛glucagone intestinale', un ormone peptidico del tratto intestinale, che, interagendo con le cellule fi, causa la liberazione di insulina nello spazio extracellulare e in circolo. Alcuni studiosi di questi problemi ritengono che il glucosio all'interno delle cellule β sia metabolizzato a sorbitolo e che il sorbitolo sia la sostanza scatenante la secrezione di insulina. Ovviamente queste indagini sono molto complicate e i loro risultati richiedono ulteriori conferme.
Gli effetti dell'insulina sulle cellule-bersaglio sono responsabili della riduzione del tasso di glucosio circolante, facendo cessare lo stimolo per la secrezione di insulina.
Effetti dell'insulina. - Per comprendere gli effetti dell'insulina nel processo di regolazione della glicemia bisogna tener presenti, almeno in parte, gli effetti che questo ormone esplica sul metabolismo, particolarmente su quello dei carboidrati, degli acidi grassi e degli amminoacidi.
Gli effetti dell'insulina sono certamente mediati da un recettore di membrana sia a livello delle cellule adipose, sia a livello delle cellule epatiche. Nel fegato l'insulina può attivare l'enzima fosfodiesterasi, per effetto del quale la concentrazione di AMP ciclico potrebbe diminuire. I cambiamenti che si determinano nel metabolismo dopo l'iniziale associazione dell'insulina con il recettore di membrana provocano altre modificazioni metaboliche, che alla fine influenzano il livello del glucosio ematico circolante. L'aggancio iniziale dell'ormone alla membrana cellulare, soprattutto a livello epatico, porta alla conversione del glucosio nei suoi metaboliti: attraverso la reazione esochinasica si ha la formazione di glucosio-6-fosfato, che successivamente viene metabolizzato sino ad acido piruvico. La riduzione del tasso di glucosio circolante avviene per stimolazione della sequenza primaria di eventi metabolici che portano all'utilizzazione del glucosio a scopi energetici, diminuendo così le scorte di glucosio ematico per soddisfare ulteriormente questa esigenza. Sembra che l'insulina intervenga anche nel processo di sintesi del glicogeno e ciò rappresenta una via alternativa per l'utilizzazione del glucosio. L'insulina, infine, esplica notevoli effetti sul metabolismo degli acidi grassi e delle proteine; i dati a questo riguardo sono talmente numerosi che non possono purtroppo essere trattati nella presente sommaria esposizione.
Secrezione e interazioni del glucagone. - Come abbiamo detto in precedenza, la secrezione di glucagone viene stimolata dalla diminuzione del livello ematico del glucosio. La regolazione del tasso glicemico a opera dell'insulina e del glucagone, che agiscono a livello epatico, può essere riassunta nel modo seguente: mentre il glucosio ematico stimola la secrezione di insulina da parte delle cellule βi del pancreas, le catecolammine si oppongono a tale effetto (v. al cap. 2, È a, la discussione sulle catecolammine). Le catecolammine, elaborate dalla midollare del surrene in risposta a impulsi nervosi, una volta in circolo, raggiungono le cellule fi del pancreas, ove inibiscono la secrezione d'insulina; effetto opposto è prodotto dal glucosio, da ormoni del tratto gastrointestinale, dal glucagone e dall'arginina in circolo che si accumulano nelle cellule fi o inviano a queste il segnale di produrre e secernere una maggior quantità di insulina. L'insulina così elaborata si accumula sui recettori specifici di membrana delle cellule epatiche, stimolando la sintesi del glicogeno. L'azione del glucagone o delle catecolammine sulla cellula epatica provoca invece la scissione del glicogeno in glucosio che, fuoriuscito dal fegato, stimola, come già detto, le cellule β a produrre insulina. Se il livello ematico di glucosio è basso, viene prodotto glucagone dalle cellule a del pancreas, mentre se la glicemia è elevata le cellule β secernono insulina e contemporaneamente risulta inibita la produzione di glucagone. Anche la pancreozimina, secreta dal tratto gastrointestinale in risposta all'assunzione di cibo, o altri fattori come l'arginina, la gastrina, la secretina o lo stesso glucagone possono inibire la produzione di glucagone da parte delle cellule α e stimolare l'elaborazione di insulina nelle cellule β. Il glucagone prodotto dalle cellule a del pancreas agisce a livello epatico, presumibilmente legandosi anch'esso a un recettore sito sulla membrana cellulare, e fa diminuire la concentrazione di glicogeno nel fegato stesso; infatti è necessaria la glicogenolisi per aumentare il glucosio nel sangue.
Questo breve riepilogo illustra il controllo dell'equilibrio del glucosio ematico, processo che ruota intorno agli effetti primari del glucagone e dell'insulina sulla sintesi e la scissione del glicogeno e anche sull'utilizzazione del glucosio nelle cellule epatiche.
Effetti molecolari del glucagone. - L'effetto specifico del glucagone richiede probabilmente l'attacco dell'ormone alla membrana degli epatociti e ciò determina la stimola- zione dell'adenilciclasi a produrre AMP ciclico all'interno della cellula. Questo AMP ciclico induce la conversione della forma inattiva della fosforilasichinasi nella forma attiva; ciò causa la conversione della glicogenofosforilasi dalla forma B, inattiva, nella forma A, attiva, e quindi permette la scissione del glicogeno in glucosio-l-fosfato, che passa poi a glucosio libero. Questa quindi sarebbe, dopo la nascita, l'azione specifica del glucagone (e anche dell'adrenalina) sull'utilizzazione, nella cellula epatica, del glicogeno per produrre glucosio libero.
e) Ormone paratiroideo e calcitonina.
Le paratiroidi sono due coppie di ghiandole, localizzate sulla superficie della ghiandola tiroide, costituite da due tipi di cellule, le cellule principali e le cellule ossifile. L'ormone paratiroideo (PTH), ormone ipercalcemizzante, viene elaborato dalle cellule principali: questo ormone, isolato dalle paratiroidi di bovino, risulta formato da una sola catena polipeptidica del peso molecolare di 9.500 dalton, contenente 83 residui di amminoacidi, tra i quali manca la cisteina. Una sua probabile struttura è riportata nella fig. 18; gli ultimi quattro amminoacidi del segmento carbossiterminale della catena polipeptidica possono essere rimossi, per azione della carbossipeptidasi, senza che si abbia perdita di attività, mentre l'ulteriore distacco di amminoacidi adiacenti comporta marcata diminuzione dell'attività biologica.
La calcitonina, detta anche tirocalcitonina a causa della sua presenza nella ghiandola tiroide, ha un peso molecolare di 3.600 dalton circa e contiene 32 amminoacidi, tra cui due cisteine e una metionina.
Questo ormone è stato anche sintetizzato completamente in laboratorio; la struttura della calcitonina di maiale è riportata nella fig. 19.
Effetti dell'ormone paratiroideo e della calcitonina. - L'ormone paratiroideo provoca la solubilizzazione della sostanza minerale delle ossa, agendo direttamente sugli osteoclasti. Non è ancora ben chiarito l'esatto meccanismo con cui il calcio viene attivamente solubilizzato e allontanato dalle ossa: le ipotesi formulate in proposito postulano o la distruzione delle fibre collagene a opera della collagenasi con conseguente mobilizzazione dei minerali ossei; oppure la solubilizzazione dei sali di calcio per formazione di un eccesso di sostanze chelanti, come l'acido citrico o l'acido lattico; o, infine, un cambiamento nelle caratteristiche di permeabilità delle cellule in modo che il trasporto di calcio avvenga con maggiore efficienza dall'osso ai liquidi interstiziali. È stato provato che si verifica un aumentato riassorbimento di fibre osteocollagene con conseguente aumentata escrezione di idrossiprolina: questo dato sperimentale può fornire una base per alcune delle ipotesi esposte in precedenza.
Nel rene, l'ormone paratiroideo diminuisce la quantità di ioni calcio presenti nelle urine.
L'azione dell'ormone a livello intestinale non è chiara, tuttavia sembra che l'assorbimento di calcio aumenti in seguito a somministrazione di ormone. L'ipercalcemia e la fosfaturia indotte dal paratormone sono inibite nel ratto dalla somministrazione di actinomicina D: ciò fa pensare che l'azione di questo ormone sull'osso coinvolga direttamente la biosintesi di macromolecole.
L'azione della calcitonina consiste nell'indurre rapida, ma transitoria, ipocalcemia e ipofosfatemia; apparentemente la breve durata di questo effetto è legata alla contemporanea stimolazione della secrezione di paratormone: infatti nel ratto paratiroidectomizzato l'azione della calcitonina si prolunga per diverse ore. La calcitonina incrementa la captazione di calcio da parte degli osteoblasti; inoltre blocca l'effetto del paratormone sugli osteoblasti, consistente nell'indurre tali cellule a trasformarsi in osteoclasti, com'è dimostrato dal fatto che l'aggiunta di calcitonina al mezzo d'incubazione di frammenti ossei inibisce la solubilizzazione del calcio, indotta dal paratormone.
La calcitonina ha poco o nessun effetto sull'escrezione renale di fosfato. Probabilmente l'azione ipocalcemizzante della calcitonina non si esplica attraverso un processo di biosintesi proteica, dal momento che l'actinomicina D non influisce su questo effetto.
Controllo della secrezione. - Pur non essendo conosciuto a fondo il meccanismo di controllo della secrezione di paratormone e di calcitonina, sembra che la calcemia agisca per feedbaek negativo, regolando la secrezione di ormone paratiroideo: concentrazioni plasmatiche elevate di ioni calcio bloccherebbero la secrezione di paratormone. Per- ché si abbia secrezione d'ormone paratiroideo, la calce- mia deve diminuire di 0,4-1 mg per 100 ml; d'altra parte la secrezione di calcitonina è stimolata da un aumento del tasso plasmatico di calcio di 0,15-0,5 mg per 100 ml: il risultato finale è una diminuzione di ioni calcio nel plasma. La calcitonina agisce molto rapidamente e per breve tempo; il paratormone esplica un'azione più lenta, ma più protratta: l'attività contemporanea di entrambi questi ormoni mantiene le concentrazioni di calcio e fosfato plasmatici entro limiti di variabilità assai ristretti. L'osso è in equilibrio con gli ioni calcio e fosfato del plasma, di modo che i suoi sali sono rapidamente mobilizzabili; sulle cellule ossee agiscono gli ormoni che regolano calcemia e fosfatemia, ma questo effetto è relativamente lento rispetto a quello esercitato sul rene. La regolazione della secrezione del paratormone e della calcitonina può essere inquadrata nel seguente sistema semplificato: il pool intestinale di calcio risente della concentrazione di vitamina D o meglio della sua forma attiva, nel senso che all'aumentare di essa aumenta la quantità di ioni calcio assorbiti. Gli ioni calcio assorbiti in maggior misura passano in circolo e si accumulano nelle paratiroidi e nella tiroide; all'aumentare del calcio nelle paratiroidi, diminuisce proporzionalmente il livello di ormone paratiroideo e, nello stesso tempo, l'aumento di ioni calcio nella tiroide provoca un aumento di calcitonina. Questi ormoni, la cui secrezione è legata ai valori della calcemia, vengono immessi nel sangue: a livello osseo, l'ormone paratiroideo aumenta la mobilizzazione di ioni calcio, mentre la calcitonina determina l'effetto opposto; a livello del nefrone l'ormone paratiroideo aumenta considerevolmente la quantità di ioni calcio riassorbita. In tal modo gli ioni calcio mobilizzati dall'osso o riassorbiti dal rene possono essere messi in circolo e regolare per feedback i valori della calcemia dovuti all'entità del pool intestinale.
Altri fattori che influenzano il metabolismo del calcio. - Numerose altre sostanze che influenzano il metabolismo del calcio meritano di essere ricordate sia pur brevemente. Come abbiamo già detto, i due principali fattori di regolazione della calcemia sono la calcitonina o tirocalcitonina e l'ormone paratiroideo: il primo incrementa la deposizione di calcio nell'osso e perciò provoca una diminuzione della concentrazione degli ioni calcio e magnesio nel siero e della idrossiprolina e dei fosfati nelle urine; l'ormone paratiroideo attiva gli osteoblasti favorendone la trasformazione in osteoclasti, causando così il riassorbimento dell'osso, il conseguente aumento del tasso di ioni calcio e magnesio nel sangue e l'aumento di fosfati e idrossiprolina nell'urina. L'ormone paratiroideo agisce anche sul rene, dove aumenta il riassorbimento di calcio, e sull'intestino, dove esplica un'azione sinergica a quella della vitamina D.
La vitamina D assunta con l'alimentazione agisce sulla mucosa intestinale, dove aumenta la quantità di proteina calciofissatrice necessaria per l'assorbimento di calcio dal tubo gastroenterico (v. vitamine).
Altri tre ormoni intervengono nel metabolismo del calcio: l'ormone somatotropo, che stimola gli osteoblasti aumentando la loro utilizzazione di ioni calcio durante l'accrescimento; l'estradiolo, che agisce anch'esso attivando gli osteoblasti, causando la chiusura delle epifisi e impedendo il riassorbimento di ioni calcio; l'ormone tiroideo, infine, che negli animali in accrescimento facilita l'effetto dell'ormone della crescita, necessario per la maturazione delle epifisi ossee e che negli adulti impedisce il riassorbimento degli ioni calcio.
f) Ormoni gastrointestinali.
Numerose sostanze elaborate dal tratto gastroenterico influenzano l'attività secretrice o quella motoria di altre parti dell'intestino: ciò può essere verificato iniettando questi preparati ormonali isolati e opportunamente purificati. La gastrina e la secretina sono le due più importanti sostanze ormonali di questo tipo; sono di natura polipeptidica e la loro sequenza amminoacidica primaria è presentata nella fig. 20. Gli effetti della gastrina possono essere indotti anche dall'istamina, e poiché sembra che la gastrina possa provocare liberazione di istamina, non è del tutto chiarita la sua azione specifica. Sappiamo invece qualcosa di più sulla secretina, polipeptide composto da 27 amminoacidi, i primi 14 dei quali, a partire dall'estremità ammino-terminale, presentano una sequenza simile, anche se non identica, a quella del glucagone; ciò è di particolare interesse in quanto, poiché il glucagone viene secreto dal pancreas, sviluppatosi come un'estroflessione dell'intestino primitivo, la somiglianza dei due polipeptidi indica una possibile origine primitiva comune di entrambi gli ormoni.
La secretina provoca la secrezione di un succo pancreatico particolarmente ricco in bicarbonato, ma non contenente una quantità notevole di enzimi pancreatici. Altri ormoni elaborati dalla mucosa intestinale sono la colecistochinina, che causa lo svuotamento della cistifellea, e la pancreozimina, che stimola il pancreas a secernere nel lume intestinale i suoi enzimi. Oltre a quelli già detti esistono numerosi altri ormoni gastrointestinali, la cui natura chi- mica non è bene accertata; ricorderemo la enterocrinina, che controlla la secrezione di succo intestinale; la villichinina, che controlla il movimento dei villi, e la duocrinina, che controlla la secrezione delle ghiandole di Brunner, situate nella sottomucosa della parte superiore del duodeno; l'enterogastrone, infine, che inibisce la motilità gastrica (v. ormoni locali).
4. Ormoni steroidi.
a) Ormoni della corticale del surrene.
Gli ormoni corticosurrenali sono prodotti dalla ghiandola surrenale, nella cui corticale (v. corteccia surrenale) si distinguono tre strati di cellule (v. fig. 12). Come abbiamo già detto, la sintesi e la secrezione degli ormoni corticosurrenali vengono indotte dall'ipofisi anteriore tramite l'ACTH; questo, a sua volta, viene stimolato dall'ipotalamo per mezzo del fattore liberatore della corticotropina, la cui elaborazione è scatenata dall'arrivo di stimoli stressanti ricevuti dal sistema nervoso centrale.
Nell'uomo il principale corticosteroide è il cortisolo; invece in altri animali, come il ratto, molto impiegato nella ricerca sperimentale, lo steroide più importante elaborato dal surrene è il coiticosterone, che differisce dal cortisolo solamente per la mancanza del gruppo α-idrossilico in posizione 17. Gli ormoni corticosurrenali, la cui funzione consiste nel permettere all'organismo di sopravvivere agli stress causati da improvvisi cambiamenti ambientali, vengono elaborati dalla zona fascicolare e dalla zona reticolare, i due strati più interni della corticale, mentre la zona glomerulare, la più esterna, è il sito di produzione dell'aldosterone, un ormone steroide simile ai precedenti, la cui funzione primaria è di influenzare la ritenzione salma a livello del rene.
La funzione principale dei corticosteroidi è di regolare i processi metabolici a livello del fegato e di altri organi- bersaglio, come la ghiandola timo: infatti, inducendo variazioni metaboliche in biochimismi fondamentali quali la glicogenosintesi e la glicogenolisi, e inducendo in tal modo l'aumento del tasso glicemico, esplicano un'attività estremamente importante per l'organismo animale, facilitandone l'adattamento a improvvise variazioni ambientali.
L'azione e i meccanismi di controllo dell'aldosterone, l'ormone che influenza la ritenzione salma, e del cortisolo, il principale ormone corticosteroide nell'uomo, sono riportati nelle figg. 21 e 22.
b) Ormoni sessuali.
Gli effetti e i meccanismi di controllo degli ormoni sessuali sono riportati nelle flgg. 23 e 24.
Gli ormoni sessuali vengono per convenienza divisi in tre classi: androgeni (ormoni sessuali maschili), estrogeni (ormoni sessuali femminili) e progestinici (ormoni che preparano e proteggono la gestazione).
Senza entrare nel merito dei loro ruoli nello sviluppo embrionale dei caratteri sessuali, centriamo la nostra attenzione sulle azioni di questi ormoni a livello delle cellule differenziate che costituirono il loro bersaglio.
Effetti degli androgeni. - Gli androgeni stimolano la sintesi proteica e rendono positivo il bilancio dell'azoto negli organi recettori come il pene, la prostata ventrale, le vescichette seminali e i muscoli (v. fig. 23). Questi tessuti fungono da bersaglio in virtù della presenza in essi di un recettore proteico per il dudrotestosterone. Questo ormone promuove la sintesi proteica, provocando, presumibilmente per un'azione sul genoma cellulare, la trascrizione di alcuni geni i quali, in assenza di questo ormone, non darebbero luogo a nuovi RNA messaggeri (v. acidi nucleici).
Dopo il trasferimento su di un poliribosoma funzionale, nel citoplasma, questi RNA messaggeri sono tradotti in nuove proteine, il che giustifica l'aumentata sintesi proteica totale ricordata in precedenza. Questo schema concorda con i fatti finora noti, benché non siano disponibili tutte le informazioni concernenti il preciso succedersi degli eventi dopo l'interazione del complesso ormone-recettore con il genoma.
I caratteri sessuali secondari maschili compaiono per la presenza del recettore ormonale nei tessuti specifici: a ciò è dovuto il manifestarsi di fenotipi particolari, caratteristici di questi tessuti, come lo sviluppo di peli, la muscolatura e gli organi sessuali.
Effetti degli estrogeni. - Gli estrogeni determinano la comparsa nell'organismo femminile di una risposta analoga a quella che gli androgeni provocano nel maschio: infatti, come si può desumere dalla fig. 24, stimolano la sintesi proteica negli organi-bersaglio, come l'utero. È presumibile che ciò avvenga perché questi organi contengono un recettore proteico per l'estradiolo che può interagire con il genoma nello stesso modo visto prima per gli androgeni. Sino a oggi, anche se un maggior numero di ricerche sono state condotte con il recettore degli estrogeni piuttosto che con quello degli androgeni, sembra che queste molecole siano simili per struttura e per attività caratteristica: ovviamente differiscono sostanzialmente per la specificità nei riguardi dello steroide da legare. Il meccanismo d'azione a livello molecolare implica la captazione dell'estrogeno non legato che, proveniente dal sangue, diffonde liberamente sino a penetrare in una cellula-bersaglio, per esempio una cellula uterina. Una volta all'interno della cellula, l'estrogeno, nel citoplasma, si lega specificamente a una proteina recettrice; il complesso steroide-recettore viene traslocato nel nucleo, dove si lega a un accettore che potrebbe essere caratteristico per ciascun particolare tessuto-bersaglio e che è probabile sia una proteina legata al DNA genetico o un sito posto sullo stesso DNA genetico. Questo processo avviene solo a temperature fisiologiche; esso non si verifica sperimentalmente in sistemi acellulari mantenuti a bassa temperatura, mentre può manifestarsi, nelle stesse condizioni, per temperature elevate. Può darsi che il trasferimento intranucleare, che dipende dalla temperatura, consista in un cambiamento di conformazione della proteina recettrice tale da renderla capace di entrare nel nucleo: questo cambiamento conformazionale non avverrebbe a basse temperature.
Gli effetti del progesterone possono essere mediati da simili eventi molecolari specifici; tuttavia non è chiaro se in questo caso esista un cambiamento conformazionale della proteina accettrice dei progestinici, necessario per il trasferimento nel nucleo delle cellule sensibili.
5. Prostaglandine.
È questo un nuovo gruppo di sostanze ormonali molto attive, isolate inizialmente dalla prostata, ma ampiamente diffuse in diversi tessuti (v. ormoni locali).
Nella fig. 25 è riportata la struttura della prostaglandina E1 (PGE1), una delle più importanti molecole di questo tipo. E possibile che le prostaglandine modulino l'attività ormonale interagendo specificamente, in qualche modo, con la membrana cellulare: alcune di esse stimolano la contrazione dei muscoli lisci, abbassano la pressione del sangue e ostacolano l'azione di ormoni come la vasopressina. Come si vede dalla struttura della fig. 25, queste sostanze poco comuni si formano da acidi grassi polinsaturi per chiusura di un anello ciclopentanico o ciclopentemco nel mezzo di una catena di un acido grasso. In colture cellulari, talora, le prostaglandine simulano alcuni effetti dell'AMP ciclico.
6. Considerazioni conclusive.
Esiste un'enorme varietà di ormoni e di composti chimici provvisti di attività ormonale, sostanze per le quali si comincia solo vagamente a comprendere il rapporto tra struttura chimica e funzioni specifiche. Per esempio, le ghiandole derivate dall'intestino primitivo secernono tutte ormoni polipeptidici o proteici e alcuni di questi possono avere una sequenza di amminoacidi simile. Gli ormoni steroidi derivano dal mesoderma e probabilmente sono comparsi con l'introduzione della riproduzione sessuale durante l'evoluzione, rappresentando la ghiandola surrenale un ulteriore sviluppo, avente ancora la capacità di produrre ormoni sessuali.
Vi è ancora molto cammino da fare per comprendere come gli ormoni funzionino negli organismi superiori, problema che è alla base del differenziamento cellulare stesso.
Bibliografia.
Burgus, R., Guillemin, R., Hypothalamic releasing factors, in ‟Annual review of biochemistry", 1970, XXXIX, pp. 499-526.
Fox, S. W., Harada, K., Krampitz, G., Müller, G., Chemical origins of cells, in ‟Chemical and enginering news", 1970, XLVIII, 26, pp. 80-94.
Frieden, E., Lipner, H., Biochemical endocrinology of the Vertebrates, Englewood Cliffs, N. J., 1971.
Litwack, G. (a cura di), Biochemical actions of hormones, voll. I-V, New York 1970 ss.
Litwack, G., Kritchevsky, D. (a cura di), Actions of hormones on molecular processes, New York 1964.
Robison, G. A., Butcher, R. W., Sutherland, E. W., Cyclic AMP, New York 1971.
Turne, C.D., Bagnara, J. T., General endocrinology, Philadelphia 19715.
Williams, R. H. (a cura di), Textbook of endocrinology, Philadelphia 1969.
Omoni locali
SOMMARIO: 1. Introduzione. □ 2. Acetilcolina: a) teoria della trasmissione neuroumorale; b) scoperta dell'acetilcolina; azioni muscariniche e nicotiniche; c) liberazione dell'acetilcolina dalle terminazioni nervose; d) effetti dell'acetilcolina al livello della giunzione neuromuscolare e sul cuore; e) recettori dell'acetilcolina; f) le colinesterasi; g) azioni dell'acetilcolina nel sistema nervoso centrale. □ 3. Catecolammine: a) cenni storici; b) biochimica; c) effetti delle catecolammine; d) inattivazione delle catecolammine; e) le catecolammine nel sistema nervoso centrale; f) applicazioni terapeutiche. □ 4. Istamina: a) origine, chimica, formazione e distruzione; b) azioni farmacologiche e fisiologiche; c) farmaci antistaminici; recettori istaminici; d) metodi di estrazione e di dosaggio; e) liberazione di istamina nell'anafilassi e a opera di liberatori chimici dell'istamina. □ 5. 5-Idrossitriptammina (serotonina, enterammina): a) la 5-HT come mediatore dell'impulso nervoso; b) implicazioni cliniche; c) derivati indolici similari. □ 6. Chinine: a) callicreina; b) formazione e distruzione delle chinine; c) azioni farmacologiche e possibili ruoli fisiologici e patologici delle chinine; d) polipeptidi similari. □ 7. Prostaglandine: effetti farmacologici e possibili funzioni. □ 8. Conclusione. □ Bibliografia.
1. Introduzione.
Quando W. M. Bayliss ed E. H. Starling introdussero il termine ‛ormone', intesero applicano a un principio attivo formato in un organo e portato dal sangue ad altri organi in cui esso produceva i suoi effetti specifici. Fu sir H. H. Dale a proporre, per chiarezza, che l'uso del termine ormone dovesse essere limitato al significato originariamente attribuitogli da Bayliss e Starling. Dale tuttavia andò oltre, affermando che in aggiunta ai veri ormoni esistevano nell'organismo sostanze ad attività fisiologica ugualmente intensa, l'azione delle quali era circoscritta alle immediate vicinanze del sito della loro liberazione. Egli chiamò queste sostanze ‛stimolanti chimici ad azione locale'; altri le hanno chiamate ‛ormoni locali' o ‛ormoni tessutali' o, semplicemente, ‛sostanze farmacologicamente attive dei tessuti'. Nella trattazione di questo gruppo inomogeneo ci si imbatte nella difficoltà che, mentre per alcuni di questi agenti è stato assodato il possesso di funzioni chiaramente definite, per altri, che pure sono farmacologicamente attivi quando siano saggiati in varie condizioni sperimentali, non è stata stabilita una funzione nell'organismo.
Noi ci proponiamo di discutere in questa rassegna varie sostanze trovate nei tessuti, particolarmente di Mammifero, che sono farmacologicamente attive ma non possono essere classificate fra i veri ormoni nel senso di Bayliss e Starling. Per ciascun ormone locale non solo saranno descritti gli effetti, ma sarà posta la domanda del significato funzionale che a esso è stato o può essere attribuito. Non c'è dubbio che in molti casi si scopriranno nuove funzioni degli ormoni locali e pertanto questo nostro modo di parlarne sarà alquanto speculativo, cosa giustificabile in un'opera rivolta al futuro, quale è questa.
2. Acetilcolina.
a) Teoria della trasmissione neuroumorale.
L'acetilcolina (ACh) ha avuto una parte centrale nello sviluppo della teoria della trasmissione neuroumorale, che ha profondamente mutato le nostre vedute sulla propagazione dell'impulso nervoso e sul funzionamento del sistema nervoso. Allo scopo di comprendere pienamente le acquisizioni di O. Loewi e H. H. Dale, che diedero inizio alla teoria della trasmissione neuroumorale, è necessario richiamarsi a precedenti concetti sul modo d'azione dei farmaci in rapporto ai nervi, che risalgono a Cl. Bernard. Questi usò i farmaci per studiare le funzioni fisiologiche, definendole come reagenti della vita, les réagents de la vie. Una delle sostanze chiave da lui studiate fu il veleno delle frecce usato in Sudamerica, il curaro, che egli dimostrò agire in corrispondenza della giunzione fra nervo motore e muscolo. In un famoso esperimento, mostrato nella fig. 1, egli impiegò due preparati neuromuscolari di rana immersi in una soluzione di curaro. Quando il muscolo era immerso nella soluzione, esso rispondeva con una brusca contrazione alla stimolazione elettrica diretta, ma non alla stimolazione indiretta attraverso il nervo; quando invece era il tronco nervoso a essere immerso nella soluzione di curaro, allora il muscolo rispondeva a entrambi i tipi di stimolazione. Questo esperimento escludeva sia il tronco nervoso motore sia lo stesso muscolo come siti della paralisi e Cl. Bernard concludeva - come ora sappiamo, erroneamente - che il curaro paralizzava le fini terminazioni nervose del nervo motore nel punto in cui esse si congiungono al muscolo.
Le concezioni di Cl. Bernard influenzarono il pensiero dei fisiologi per i successivi sessant'anni. Così O. Schmiedeberg, studiando l'azione del veleno fungino muscarina e trovando che esso produceva effetti simili a quelli ottenibili con la stimolazione vagale, concludeva che la muscarina stimolava presumibilmente le terminazioni nervose del vago. Egli studiò anche l'atropina, scoprendo che era capace di antagonizzare gli effetti sia della muscarina, sia della stimolazione vagale e concluse che l'atropina agiva sulle terminazioni vagali più perifericamente della muscarina, producendo un blocco. Era così costruito un convincente sistema d'azione dei farmaci, basato sulla stimolazione e paralisi delle terminazioni nervose.
Questo sistema concettuale venne rivoluzionato, negli anni venti, dagli esperimenti del Loewi sul cuore isolato di rana, nei quali egli dimostrò la liberazione di Vagusstoff (più tardi identificato come ACh) dal nervo vago. Il Loewi condusse a termine una serie di esperimenti di grande semplicità ed economia di mezzi, i quali hanno decisamente influenzato il nostro modo di pensare non solo sulla trasmissione dell'impulso nervoso, ma anche sulla maniera d'interferire in tale trasmissione. I reperti fondamentali del Loewi relativi all'ACh sono illustrati nella fig. 2 e possono essere riassunti come segue.
1. La stimolazione del vago causa la comparsa, nel perfusato di cuore di rana, di una sostanza capace di produrre, in un secondo cuore di rana, un effetto rassomigliante alla stimolazione vagale. Loewi concluse che doveva essersi liberata una sostanza atta a trasmettere normalmente la stimolazione vagale, che venne chiamata Vagusstoff (sostanza vagale).
2. L'atropina (alcaloide della Atropa belladonna) previene gli effetti della stimolazione vagale sul cuore donatore, ma non impedisce la liberazione della sostanza vagale. Infatti, se il perfusato raccolto durante la stimolazione vagale di un cuore atropinizzato viene trasferito a un secondo cuore, esso provoca l'inibizione di questo. L'atropina pertanto previene l'effetto del trasmettitore, ma non la sua liberazione.
3. Loewi intuì l'esistenza della colinesterasi, dimostrando che la sostanza vagale viene inattivata per incubazione con muscolo cardiaco triturato. L'alcaloide eserina (dalla fava del Calabar), che potenzia la stimolazione vagale, previene la distruzione della sostanza vagale a opera del muscolo cardiaco. Era così raggiunta la prova che il potenziamento eserinico era dovuto a inibizione dell'enzima inattivante il trasmettitore.
In tal modo gli esperimenti del Loewi dimostrarono che la trasmissione neuroumorale in corrispondenza delle terminazioni nervose del vago avviene a opera dell'ACh e chiarirono il meccanismo col quale i farmaci possono bloccare o potenziare la trasmissione.
Un altro tipo di trasmissione neuroumorale da ACh venne dimostrato circa dieci anni dopo gli esperimenti del Loewi: precisamente, la trasmissione acetilcolinica in corrispondenza della sinapsi gangliare. L'esperimento cruciale fu compiuto da A. W. Kibjakov, che perfuse il ganglio cervicale superiore di gatto, organo del peso di circa 20 mg. Egli dimostrò che la stimolazione preganghare liberava nel perfusato un fattore che, riperfuso attraverso il ganglio, stimolava la fibra postgangliare. Esperimenti successivi di W. Feldberg, J. H. Gaddum e altri provarono che il fattore non era altro che acetilcolina la quale, perfusa attraverso il ganglio, stimolava le cellule gangliari a scaricare impulsi lungo le fibre postgangliari. Durante la stimolazione nervosa pregangliare l'ACh appare nell'effluente venoso dal ganglio (a condizione che il liquido di perfusione contenga eserina) e la sua presenza può essere dimostrata con vari test biologici. L'ACh è liberata in seguito a stimolazione nervosa preganghare anche se la trasmissione gangliare è paralizzata da nicotina o esametonio, ma non è liberata da stimolazione antidromica del tronco postgangliare. La degenerazione della fibra pregangliare determina la scomparsa del contenuto di ACh del ganglio e anche la scomparsa degli enzimi colinacetilasi, che sintetizza l'ACh, e colinesterasi, che la distrugge, ma le cellule gangliari rimangono sensibili alla ACh iniettata.
Oltre che agire come trasmettitore in corrispondenza delle terminazioni nervose postgangliari parasimpatiche e delle sinapsi gangliari, l'ACh funge da trasmettitore in altri siti periferici. Essa funge da mediatore dell'impulso nervoso dal nervo splancnico alle cellule della midollare surrenale che liberano adrenalina, cellule che embriologicamente sono affini ai neuroni postgangliari ; inoltre trasmette l'impulso dal nervo motore al muscolo striato, come è provato dalla liberazione di ACh quando un nervo motore è stimolato elettricamente (v. Dale e altri, 1936). Introdotta per iniezione intrarteriosa vicina, l'ACh produce un breve tetano asincrono. ACh infine é liberata, atipicamente, da certe terminazioni nervose postgangliari simpatiche, per es. da quelle delle ghiandole sudoripare. Dale ha chiamato ‛colinergiche' le fibre nervose che producono i loro effetti attraverso una liberazione di ACh.
b) Scoperta dell'acetilcolina; azioni muscariniche e nicotiniche.
L'Ach è un estere della colina e dell'acido acetico che è altamente instabile e facilmente si scompone nei suoi due componenti:
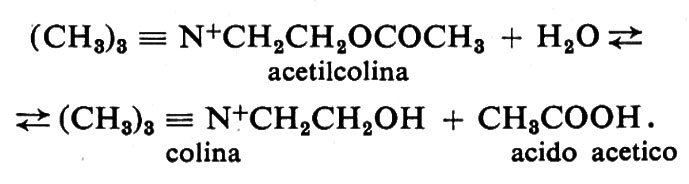
Essa fu sintetizzata nel 1894 da G. Nothnagel. Nel 1906 R. Hunt e M. Taveau scoprirono che esplicava un potente effetto ipotensivo quando era iniettata nel circolo dei Mammiferi e fin da allora essi prospettarono la possibilità che l'ACh potesse essere un costituente naturale del corpo. Questo punto tuttavia non fu provato che molto più tardi, quando Dale e H. W. Dudley estrassero ACh dalla milza di cavallo.
L'interesse di Dale per l'ACh, come per l'istamina, nacque dall'averla ritrovata in estratti di segale cornuta. Nel 1914 egli condusse una ricerca sistematica sull'ACh, che rivelò un impressionante parallelismo fra le sue azioni e gli effetti della stimolazione nervosa parasimpatica. Dale trovò che le azioni vasodilatatrici e parasimpatiche dell'ACh (v. sistema nervoso autonomo) erano facilmente antagonizzate dall'atropina ma che, dopo atropina, l'iniezione di elevate dosi di ACh produceva una stimolazione nicotinosimile delle cellule gangliari e della midollare surrenale. Questo reperto lo indusse a suddividere le azioni farmacologiche dell'ACh in ‛muscariniche' e ‛nicotiniche'. Tale classificazione venne pienamente giustificata dalla teoria della trasmissione neuroumorale, la quale dimostrò che le azioni dell'ACh liberata nell'organismo erano classificabili in base a questo schema. Nella terminologia moderna, recettori per l'ACh esistono in corrispondenza delle sinapsi muscariniche e a livello di due tipi di sinapsi nicotiniche. Ciascun tipo di recettore è interessato da antagonisti specifici dell'ACh, che sono l'atropina per le giunzioni muscariniche, la tubocurarina per la giunzione nicotinica della placca motrice e l'esametonio per la giunzione nicotinica gangliare.
c) Liberazione dell'acetilcolina dalle terminazioni nervose.
L'ACh dei tessuti dei Vertebrati è sintetizzata dall'enzima colinacetilasi (D. Nachmansohn e A. L. Machado, 1943), presente in tutti i neuroni colinergici e capace di trasferire l'acetilcoenzima A alla colina. Pertanto un'efficiente sintesi di ACh richiede la presenza di adeguate quantità di colinacetilasi, colina libera, coenzima A e acetato (per la formazione di acetilcoenzima A). L'energia chimica necessaria per l'acetilazione del coenzima A è resa disponibile in forma di ATP.
Colinacetilasi e ACh sono presenti lungo tutto l'assone dei nervi colinergici, come pure a livello delle loro terminazioni. La colinacetilasi è probabilmente elaborata nel corpo cellulare e portata alla periferia da movimenti dell'assoplasma, ma questo non è stato provato in modo così preciso come nel caso dei nervi adrenergici. La colina è un costituente normale dei liquidi extracellulari, che non penetra facilmente nell'interno della cellula. Eppure, quando il ricambio dell'ACh è vivace, come durante l'attività nervosa, le scorte di colina nell'interno del nervo devono continuamente essere ricostituite, e ciò si ottiene per l'esistenza di un meccanismo di trasporto specifico. Infatti J. F. C.MacIntosh e collaboratori hanno provato che una base emicolinica, che deprime fortemente la sintesi di ACh da parte dei neuroni colinergici, esplica la sua azione competendo con il trasportatore specifico a livello della membrana cellulare.
Nei nervi, e più specificamente nelle terminazioni nervose colinergiche, l'ACh sembra essere presente in due forme, libera nell'assoplasma e localizzata in granuli intracellulari dai quali è liberata in corrispondenza delle terminazioni nervose. Il processo di liberazione dell'ACh dalle terminazioni nervose è stato ampiamente chiarito dal lavoro di B. Katz e collaboratori, i quali hanno dimostrato che la liberazione dell'ACh dalle terminazioni nervose costituisce una forma di secrezione controllata elettricamente (v. Katz, 1969). I loro esperimenti sono stati condotti con microelettrodi. Se si penetra nella fibra muscolare, il potenziale interno è negativo, di circa 90 mV, rispetto al bagno esterno. Questo vale per l'intera fibra muscolare, ma se l'oscilloscopio è regolato a elevata amplificazione, la regione della placca motrice terminale sembra essere in uno stato di continua attività elettrica subliminale anche quando non è stato applicato nessuno stimolo. Si ha una serie di piccole scariche elettriche intermittenti, ciascuna corrispondente a una breve depolarizzazione di circa 0,5 mV (v. fig. 3), che s'innalza rapidamente e declina più lentamente per una durata di circa 20 ms. Katz chiamò queste depolarizzazioni ‛potenziali miniatura della placca terminale' (miniature end plate potentials). Queste modificazioni di potenziale sono simili ai potenziali della placca terminale indotti dalla stimolazione del nervo motore, da cui si differenziano soprattutto per la loro ampiezza molto più piccola (circa 1/100) e per la loro spontaneità. Quando il potenziale d'azione invade la terminazione nervosa, i potenziali miniatura della placca terminale divengono molto più frequenti, tanto frequenti che non possono più essere contati singolarmente. In queste condizioni i potenziali miniatura della placca terminale sommano le loro ampiezze, per quanto non in proporzione lineare al loro numero, e producono una forte depolarizzazione della membrana (v. fig. 3).
Si ritiene che i potenziali miniatura della placca terminale siano dovuti alla liberazione di pacchetti unitari di ACh, ciascuno contenuto in una vescicola sinaptica nella terminazione nervosa. La sostanza trasmettitrice, l'ACh, è impacchettata nell'interno di un sacco vescicolare ed è separata dall'organo-bersaglio da due barriere, la membrana vescicolare e la membrana della terminazione dell'assone. La frequenza delle collisioni fra le membrane vescicolare e assonale è probabilmente elevata, ma normalmente la grande maggioranza di queste collisioni è improduttiva e non porta a liberazione del trasmettitore. La depolarizzazione indotta dall'impulso nervoso può produrre modificazioni nella membrana assonale in modo che questa presenta temporaneamente un numero molto maggiore di siti attivi alle membrane che entrano in collisione, col risultato che la probabilità statistica della liberazione del trasmettitore è accresciuta. Che il verificarsi dei potenziali miniatura della placca terminale sia un processo statistico ‛randomizzato' può essere provato matematicamente dall'applicazione a esso della legge di distribuzione statistica di Poisson, applicabile a eventi le cui probabilità di successo sono molto basse. La comparsa di potenziali miniatura della placca terminale segue la legge di Poisson.
La presenza di calcio nel liquido extracellulare è essenziale per la liberazione del trasmettitore. Se il livello del calcio si abbassa si ha una notevole diminuzione della quantità di ACh liberata. Anche il magnesio ha una rimarchevole azione nella liberazione del trasmettitore, antagonizzando gli effetti giunzionali del calcio. Quando il calcio è eliminato dall'ambiente esterno, l'impulso nervoso raggiunge ancora le terminazioni nervose, ma è incapace di liberare ACh. Katz e R. Miledi hanno dimostrato che, se in queste condizioni si applica alla terminazione nervosa una micropipetta riempita di una soluzione concentrata di CaC2 e se ne lascia fluire una piccola quantità, in quel punto si ha liberazione di ACh con produzione di un potenziale di placca terminale localizzato. Poiché il calcio è necessario nel processo per cui la depolarizzazione produce liberazione di ACh, gli autori suddetti suggerirono l'ipotesi che la depolarizzazione aprisse una porta agli ioni calcio permettendogli di penetrare nella membrana assonale. È possibile che il calcio sia essenziale nel processo che causa quella fusione transitoria delle membrane vescicolare e assonale che conduce alla fuoriuscita del pacchetto di trasmettitore.
d) Effetti dell'acetilcolia al livello della giunzione neuromuscolare e sul cuore.
La placca motrice terminale del muscolo striato è sensibilissima all'ACh per la presenza sulla sua superficie di speciali molecole recettrici capaci di reagire con l'ACh liberata dalle terminazioni nervose. L'azione dell'ACh sui recettori provoca un aumento della permeabilità della membrana al sodio e al potassio, come pure ad altri anioni e cationi liberi dall'una e dall'altra parte della membrana. In questo modo il potenziale attraverso la placca terminale motrice diviene cortocircuitato e ne risulta una depolarizzazione locale, il potenziale di placca terminale. Il potenziale di placca terminale è una risposta graduata che, quando raggiunge un livello soglia, dà origine a correnti locali di intensità sufficiente per aumentare la permeabilità al sodio delle porzioni circonvicine della membrana e per dare inizio a un potenziale d'azione muscolare diffuso. L'ACh liberata dalla terminazione nervosa è distrutta in pochi millisecondi dalla colinesterasi nella regione giunzionale, permettendo alla placca terminale di ripolarizzarsi prima che sia passato il periodo refrattario del muscolo.
Un blocco neuromuscolare può essere prodotto in modi diversi. Una possibilità teorica è l'interferenza nel meccanismo liberatore di ACh, per es. compromettendo il trasporto della colina a mezzo di emicolina e diminuendo la liberazione di ACh con magnesio. Molto più importante in pratica è l'interferenza con l'apparato recettore dell'ACh in corrispondenza della placca motrice terminale, che può essere ottenuta per due vie diverse, esemplificate da due rilassanti muscolari d'importanza clinica. La tubocurarina antagonizza lo sviluppo del potenziale della placca terminale, competendo con l'ACh per i recettori. Il sussametonio (succinilcolina) produce una depolarizzazione acetilcolinosimile dei recettori, seguita da blocco neuromuscolare. Si ritiene che questo tipo di blocco sia dovuto in parte a desensibilizzazione del recettore e in parte a ineccitabilità locale della membrana cellulare conseguente a depolarizzazione prolungata; viene così ostacolata la diffusione del potenziale d'azione.
Per quanto l'ACh inibisca il cuore, il meccanismo che è alla base dell'effetto inibitore è essenzialmente simile a quello che sostiene l'effetto eccitatorio dell'ACh in corrispondenza della giunzione neuromuscolare: entrambi sono dovuti ad accresciuta permeabilità della membrana agli ioni. La differenza sta nella specie di ioni interessata. A livello della giunzione neuromuscolare l'ACh provoca un aumento non selettivo della permeabilità con conseguente depolarizzazione, mentre in corrispondenza del pacemaker cardiaco essa provoca un aumento selettivo nella permeabilità agli ioni potassio e forse cloruro. Questo tende a ridurre la velocità di accrescimento del potenziale del pacemaker e a rallentare il ritmo cardiaco. Elevate dosi di ACh o un'intensa stimolazione vagale sopprimono i potenziali del pacemaker e arrestano il cuore.
e) Recettori dell'acetilcolina.
Come è stato detto, esistono almeno due tipi di recettori dell'ACh, quelli muscarinici e quelli nicotinici, distinti per le diverse affinità per gli antagonisti. Varie ragioni sono state suggerite per spiegare perché la molecola dell'ACh è capace di reagire con entrambi i tipi di recettore. Una spiegazione è basata sul fatto che la molecola dell'ACh è flessibile, così che differenti isomeri conformazionali potrebbero interagire con differenti recettori: per esempio un isomero potrebbe presentarsi nella forma distesa e l'altro in una forma quasi anulare. Un'altra spiegazione è quella che diversi gruppi reattivi nella molecola dell'ACh possano reagire con recettori differenti.
L'efficacia dell'ACh (e di altri farmaci) nell'attivare i recettori dipende da due variabili: affinità per il recettore e capacità d'indurre una risposta (attività intrinseca o efficacia). Non è noto quale parte della molecola dell'ACh contribuisca all'affinità e quale all'efficacia o per quale meccanismo l'ACh legata ai suoi siti recettivi specifici induca una risposta fisiologica, quale un aumento nella permeabilità di membrana. Una proposta (T.R. Podleski e J. P. Changeux, 1970) è che vi possa essere un'interazione indiretta o ‛allosterica' fra il sito di agganciamento e il sito producente la risposta, simile all'interazione cooperativa postulata per certi enzimi regolatori (v. catalisi enzimatica).
La localizzazione dei recettori dell'ACh nel muscolo striato può essere esplorata a mezzo della ionoforesi. Se l'ACh è applicata per ionoforesi attraverso una sottile micropipetta in un punto vicino alla giunzione neuromuscolare, una piccola dose di essa produce una depolarizzazione della membrana della fibra muscolare, che può essere registrata a mezzo di un elettrodo intracellulare collocato nelle vicinanze. I recettori possono in tal modo essere localizzati nell'area giunzionale, ma se la pipetta è spostata di alcuni millimetri da quest'area, l'ACh cessa di produrre depolarizzazione, presumibilmente perché il sito non contiene recettori. Se un muscolo viene denervato e il nervo viene lasciato degenerare, si hanno notevoli cambiamenti nella distribuzione dei recettori. Si trova allora che la sensibilità di regioni lontane dalla giunzione neuromuscolare può salire da un livello inapprezzabile a un livello che si avvicina a quello della placca terminale. Si può concludere che dopo denervazione si formano nuovi recettori per l'ACh in regioni che prima non ne contenevano.
Un tentativo di individuare siti recettori dell'ACh a mezzo di tecniche autoradiografiche è stato fatto da P.G. Waser (1970), il quale trovò che, se della tubocurarina marcata con 14C viene applicata a un diaframma di topo, le immagini autoradiografiche mostrano che essa è specificamente localizzata nella regione della placca terminale, probabilmente perché si aggancia ai recettori dell'ACh.
f) Le colinesterasi.
Sono stati distinti due tipi di colinesterasi, la colinesterasi vera o acetilcolinesterasi, presente in corrispondenza dei siti effettori dei nervi colinergici, e la pseudocolinesterasi o colinesterasi, che è ampiamente presente nel plasma. Gli aspetti caratteristici dell'acetilcolinesterasi sono stati chiariti particolarmente dal lavoro dii. B. Wilson e sono illustrati nella fig. 4, la quale mostra che il sito attivo dell'enzima consiste di due subsiti, un sito anionico, che lega e orienta ioni ammonici sostituiti, e un sito esterasico contenente un gruppo essenziale acido e basico che lega funzioni estere e reagisce nel processo idrolitico.
Esistono due tipi di inibitori dell'acetilcolinesterasi. Un tipo contiene ioni ammonici sostituiti, con affinità per il sito anionico dell'enzima. Queste anticolinesterasi, per es. eserina e neostigmina, competono con l'acetilcolina per il sito attivo. L'altro tipo di inibitori, gli inibitori ‛irreversibili', reagisce col sito esterasico trasferendo su di esso un gruppo derivato da un acido. I composti organofosforici sono tipici rappresentanti di questa classe. La forma fosforilata dell'acetilcolinesterasi si riattiva lentamente nell'acqua, ma questo processo può essere grandemente accelerato dall'idrossilammina e particolarmente da certe ossime (pralidossima) che riattivano l'enzima rimuovendo il gruppo fosforico e possono salvare la vita in caso di intossicazione da composti organofosforici (v. pesticidi).
g) Azioni dell'acetilcolina nel sistema nervoso centrale.
L'acetilcolina è ampiamente distribuita nel sistema nervoso centrale insieme con gli enzimi responsabili della sua sintesi e della sua distruzione. Le concentrazioni di ACh trovate in varie parti del cervello non sono uniformi. La prova più recente sulla localizzazione cellulare dell'ACh e degli enzimi a essa relativi nel cervello è venuta da studi di frazionamento subcellulare con i quali è possibile ottenere preparati di terminazioni nervose isolate. Si è trovato che le terminazioni nervose separate contengono vescicole sinaptiche piene di ACh. La colinacetilasi è localizzata principalmente nel citoplasma solubile e la colinesterasi nella frazione microsomiale. La separazione dell'ACh dagli enzimi che la interessano e il suo presentarsi in vescicole sinaptiche corroborano l'opinione di una sua funzione di trasmettitore nel sistema nervoso centrale.
Nonostante i numerosi studi effettuati, è stato difficile identificare con certezza neuroni colinergici nel sistema nervoso centrale. Il miglior esempio documentato di una trasmissione sinaptica centrale mediata dall'ACh è quello dei collaterali di nervi motori nel midollo spinale innervanti le cellule di Renshaw. Questa sinapsi colinergica appare nicotinica, come ci si doveva aspettare, poiché essa corrisponde alle terminazioni del nervo motore che si sfioccano su recettori nicotinici.
Ci sono molte prove accessorie della trasmissione colinergica nel cervello. Non sono disponibili metodi istochimici per la dimostrazione dell'ACh, ma abbiamo metodi per la colorazione della colinesterasi che dimostrano che certe vie sono ricche in enzima. Può essere raccolta ACh dalla corteccia cerebrale in seguito ad applicazione di farmaci anticolinesterasici e la quantità liberata aumenta con l'attività elettrica del cervello, ma questo metodo non permette l'identificazione di singole vie colinergiche.
Un'importante tecnica per l'individuazione di neuroni colinergici consiste nell'applicazione microionoforetica di ACh e nella registrazione simultanea dell'attività elettrica. Solo alcune cellule rispondono all'applicazione ionoforetica di ACh; esse sono localizzate in particolare nel tronco encefalico, nel nucleo caudato e nell'ippocampo. Ci sono prove che le proiezioni diffuse del sistema reticolare attivatore nella corteccia cerebrale sono colinergiche. Alcune cellule nel nucleo caudato e nell'ippocampo e le fibre raggiungenti le cellule di Purkinje del cervelletto sono probabilmente colinergiche. Le risposte all'ACh nel cervello sono antagonizzate dall'atropina e appaiono essere largamente muscariniche.
Il tronco encefalico contiene parecchi neuroni reattivi all'ACh, dalla quale alcuni sono eccitati e altri inibiti. In questo caso le risposte eccitatorie appaiono miste, nicotiniche e muscariniche, e le risposte inibitorie principalmente muscariniche. Ci si può attendere che informazioni molto più precise sulle funzioni dell'ACh nella trasmissione sinaptica saranno disponibili nei prossimi anni.
3. Catecolammine.
Le catecolammine costituiscono un gruppo di ammine contenenti un nucleo catecolico portante una catena laterale etilamminica. Le tre catecolammine, dopammina, noradrenalina (norepinefrina) e adrenalina (epinefrina), sono costituenti naturali dell'organismo di Mammifero e ciascuna di esse esplica importanti attività fisiologiche. Dopammina e noradrenalina sono presenti anche negli Invertebrati. Queste sostanze consentono di comprendere le difficoltà che s'incontrano volendo distinguere ormoni veri e ormoni tessutali. L'adrenalina e, in misura minore, la noradrenalina possono infatti essere considerate ormoni veri in quanto esse sono riversate dalle cellule della midollare surrenale nella corrente sanguigna ed esercitano le loro azioni su effettori a distanza, quali i vasi sanguigni. D'altra parte la noradrenalina e, in misura minore, l'adrenalina, possono anche essere considerate ormoni locali, poiché esse sono liberate dalle terminazioni nervose simpatiche postgangliari per produrre effetti vasocostrittori locali. La principale funzione nota della dopammina è quella di trasmettitore chimico del sistema nervoso centrale. Nel seguito dell'articolo porremo in particolare risalto le principali azioni che le catecolammine esplicano a livello cellulare; il loro ruolo generale nella trasmissione neuroumorale e nell'organizzazione particolareggiata della branca simpatica del sistema nervoso autonomo viene discusso altrove (v. sistema nervoso autonomo).
a) Cenni storici.
L'effetto ipertensivo degli estratti di surrenale fu scoperto da O. Oliver ed E. A. Schäfar nel 1895. Il principio attivo, l'adrenalina, venne isolato da J. Takamine (1901) e sintetizzato, insieme con la noradrenalina, da F. Stolz nel 1904. T. R. Elliott (1905) suggerì che la somiglianza fra gli effetti della stimolazione simpatica e quelli dell'adrenalina potesse essere dovuta al fatto che quest'ultima veniva liberata in corrispondenza delle terminazioni nervose simpatiche, ma dati probanti a favore di questa ipotesi si fecero attendere fino ai classici esperimenti compiuti da Loewi a partire dal 1924 (v. fig. 5). Il significato della noradrenalina nell'organismo venne scoperto nel 1946 quando U. S. von Euler ne dimostrò la presenza nelle terminazione nervose, e P. W. J. Holtz nelle ghiandole surrenali. Il gruppo delle sostanze adrenalinosimili fu studiato in un lavoro classico da O. Barger e Dale (1910), che le denominarono ammine simpaticomimetiche.
b) Biochimica.
È molto importante comprendere bene il meccanismo di formazione delle catecolammine, poiché ogni interferenza nei loro passaggi metabolici può causare alterazioni funzionali. Fonte primaria dell'adrenalina nell'organismo è l'amminoacido L-tirosina, e i più probabili passaggi nella formazione dell'adrenalina, per la prima volta suggeriti da H. Blaschko, sono mostrati nello schema 1. Si vede che la dopammina e la noradrenalina sono entrambe formate nei corso della biosintesi dell'adrenalina. Dei vari passaggi enzimatici implicati nella biosintesi dell'adrenalina, il passaggio catalizzato dalla tirosinaidrossilasi è quello limitante la velocità, cioè quello che effettivamente determina l'accelerazione o il ritardo della formazione del prodotto finale. Nel sistema nervoso simpatico la noradrenalina controlla la propria velocità di formazione attraverso un meccanismo di feedback.
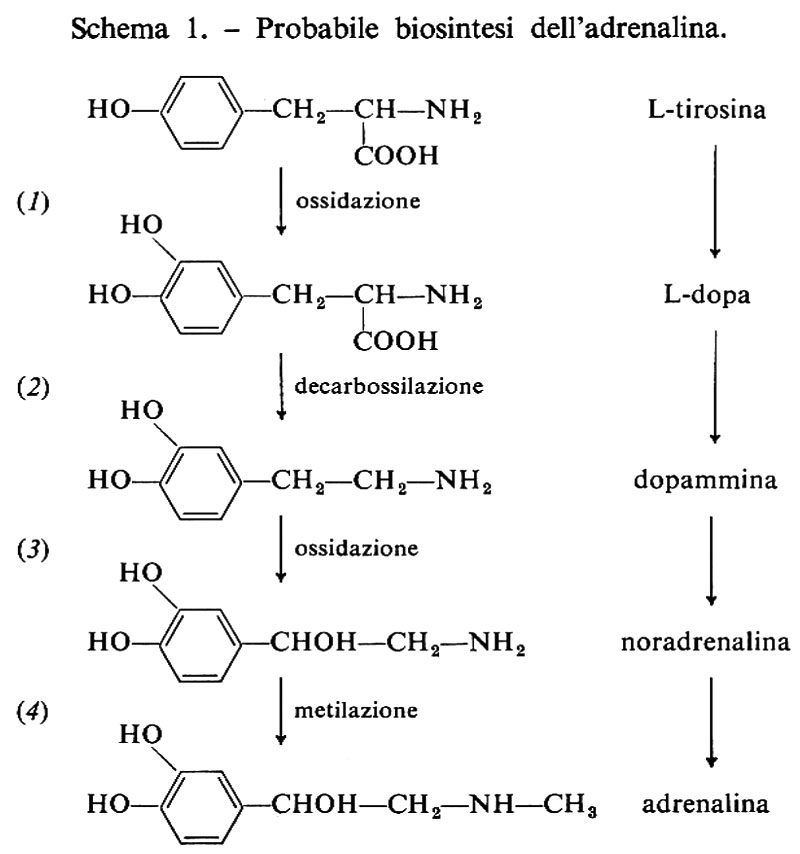
L'adrenalina nelle ghiandole surrenali e la noradrenalina nelle terminazioni nervose simpatiche sono contenute in granuli di deposito identificabili col microscopio elettronico come vescicole a centro denso. Il passaggio finale del processo di formazione della noradrenalina nel sistema nervoso simpatico avviene nei granuli di deposito che contengono l'enzima idoneo alla trasformazione della dopammina in noradrenalina. I granuli di deposito si formano nei corpi cellulari dei neuroni del sistema nervoso simpatico dai quali scendono, migrando lungo i cilindrassi, alle terminazioni nervose. Le prove di questo meccanismo di trasporto sono basate principalmente sul metodo di visualizzazione delle catecolammine a mezzo della loro fluorescenza. Come è mostrato nella fig. 6, quando il tronco nervoso viene legato si ha diminuzione della fluorescenza al di sotto della legatura e intensificazione al di sopra, presumibilmente per accumulo di granuli a monte della legatura. Il passaggio dei granuli lungo il nervo può implicare la presenza di microtubuli nell'interno della fibra nervosa.
La scarica del mediatore chimico dalle terminazioni nervose simpatiche è di tipo quantico, cioè avviene in forma di pacchetti, ciascuno dei quali contiene migliaia di molecole di noradrenalina. Una prova della liberazione sotto forma di quanti è fornita dal presentarsi di potenziali miniatura alle terminazioni nervose simpatiche, simili a quelli visibili per l'acetilcolina a livello della placca terminale motrice (v. sopra). Ogni pacchetto di molecole nasce dallo svuotarsi di una vescicola sinaptica, e questo processo è grandemente accelerato quando un impulso nervoso arriva alla periferia. Le vescicole nella ghiandola surrenale si scaricano per ‛esocitosi', cioè esse svuotano completamente il loro contenuto di catecolammine e di ATP (in proporzione di circa 1:4), nonché di una proteina specifica, la cromogranina. Le vescicole delle terminazioni nervose simpatiche scaricano il trasmettitore, ma molto del loro contenuto residuo è trattenuto e può essere riutilizzato. La scarica del contenuto è preceduta da fusione della vescicola con la membrana cellulare. È noto che il processo di liberazione delle catecolammine dai granuli richiede calcio come fattore essenziale.
c) Effetti delle catecolammine.
Dopo la sua liberazione dalle terminazioni nervose simpatiche postgangliari, il mediatore simpatico (principalmente noradrenalina) raggiunge la cellula effettrice attraverso una via di diffusione relativamente lunga. Esso poi si combina con recettori specifici che fungono da mediatori delle sue azioni fisiologiche, siano esse rappresentate da contrazione o rilasciamento del muscolo liscio, da stimolazione del cuore, da glicogenolisi o lipolisi. Sono stati caratterizzati due principali tipi di recettori adrenergici, gli alfa- e i beta-recettori. Gli alfa-recettori sono i mediatori della contrazione del muscolo liscio, i beta-recettori del rilasciamento del muscolo liscio e della stimolazione cardiaca. I recettori metabolici possono appartenere a entrambe le classi. I due tipi di recettori sono distinguibili a mezzo dei loro agenti di blocco specifici. Le due classi di recettori sono anche distinguibili per i loro specifici agenti attivanti: la noradrenalina attiva soprattutto gli alfa-recettori e l'isoprenalina (isoproterenolo), una catecolammina sintetica, i beta-recettori. L'adrenalina attiva entrambi i tipi di recettori. Sono state postulate ulteriori sottoclassi di beta-recettori, una interessante soprattutto il cuore, l'altra la muscolatura liscia.
Mentre le interazioni farmaco-recettore-antagonista sono state analizzate a fondo (esse tendono a seguire la semplice cinetica di Michaelis-Menten), molto meno è noto sull'intimo modo d'azione delle catecolammine. Due importanti meccanismi interessati sono i seguenti: I) il sistema ciclasico: E. W. Sutherland jr. e collaboratori hanno dimostrato che le catecolammine promuovono la formazione di adenosina-3′,5′-fosfato, cioè di AMP ciclico:
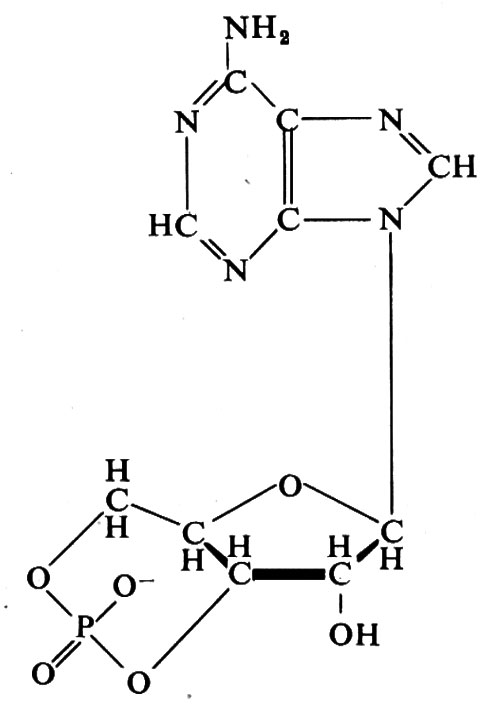
2) il calcio: le catecolammine producono contrazione o rilasciamento del muscolo liscio anche in completa assenza del potenziale di membrana, ma in assenza di calcio non riescono a produrre né contrazione né rilasciamento, facendo supporre che modificazioni nei livelli intracellulari del calcio siano d'importanza fondamentale.
d) Inattivazione delle catecolammine.
Due enzimi sono implicati nell'inattivazione delle catecolammine: l'amminossidasi, enzima intracellulare contenuto principalmente nei mitocondri, che provoca la deamminazione ossidativa della catena laterale etilamminica, e la catecol-O-metiltransferasi, che causa la metilazione di un idrossile dell'anello. Il primo funziona soprattutto durante la liberazione delle catecolammine, il secondo dopo la liberazione. Si ritiene ora che il più importante meccanismo di inattivazione della noradrenalina dopo la sua liberazione non sia di natura enzimatica, ma si attui attraverso una ricattura dell'ammina da parte delle terminazioni nervose, che ne provoca una reale inattivazione. La ricattura può essere prevenuta da certi farmaci, quali la cocaina e l'antidepressivo imipramina, i cui effetti farmacologici potrebbero essere ascritti a questa attività. Il fenomeno della ricattura è stato studiato con adrenalina triziata e si è dimostrato che il riassorbimento avviene in parte nel liquido cellulare e in parte nelle vescicole sinaptiche. La penetrazione in queste dell'ammina è impedita dalla reserpina, farmaco antipsicotico e antipertensivo. Gli inibitori della monoamminossidasi (anti-MAO), usati anche come antidepressivi, ostacolano la distruzione della noradrenalina ad opera dell'amminossidasi e potenziano gli effetti dell'ammina (v. psicofarmacologia).
e) Le catecolammine nel sistema nervoso centrale.
A differenza dell'acetilcolina, le catecolammine sono distribuite nel sistema nervoso centrale in modo non uniforme, essendo esse presenti in maggior abbondanza nel tronco cerebrale e nell'ipotalamo. La fig. 7 mostra la distribuzione della noradrenalina nel cervello di cane, con la più elevata concentrazione nell'ipotalamo. La dopammina è distribuita in modo differente dalla noradrenalina, essendo presente soprattutto nel corpus striatum. E opinione ora ampiamente accettata, soprattutto in base alle convincenti prove fornite dalla microscopia a fluorescenza, che nel sistema nervoso centrale esistano fibre noradrenergiche, dopamminergiche e probabilmente 5-idrossitriptamminergiche.
Quando adrenalina e noradrenalina sono somministrate per via endovenosa, esse non penetrano attraverso la barriera ematoencefalica che in minima quantità. Quando, invece, le stesse sostanze vengono infuse nei ventricoli cerebrali, esse producono effetti rimarchevoli, come è stato dimostrato da Feldberg e collaboratori. Per es., quando l'adrenalina è somministrata per questa via nel gatto provoca il sonno. Le catecolammine e la 5-idrossitriptammi- na soministrate per via intraventricolare modificano la temperatura del corpo; è stato perciò suggerito che esse possano essere responsabili della regolazione centrale della temperatura.
Quantunque le catecolammine non riescano a penetrare nel cervello come tali, il loro amminoacido precursore vi penetra facilmente e diviene farmacologicamente attivo dopo trasformazione biologica. Per es. A. Carlsson e collaboratori (1957) hanno dimostrato che in topi tranquillizzati con reserpina la somministrazione dell'amminoacido L-dopa ha effetto stimolante, presumibilmente a motivo della sua metabolizzazione a dopammina.
f) Applicazioni terapeutiche.
Sono stati fatti tentativi d'interferire, per scopi terapeutici, nella biosintesi delle catecolammine nel cervello e alla periferia, mediante la somministrazione di amminoacidi precursori. Uno di questi tentativi è stato l'introduzione, nella medicina clinica, dell'α-metildopa con l'obiettivo d'inibire la dopadecarbossilasi, l'enzima responsabile della formazione di dopammina dalla L-dopa. Risultò che la metildopa era molto attiva nell'ipertensione umana, ma una successiva analisi della sua azione ha dimostrato che responsabile della sua efficacia non è un'inibizione enzimatica. La sua attività terapeutica è dovuta al fatto che essa è un substrato della dopadecarbossilasi, che la trasforma in ametildopammina. Questa a sua volta è un substrato della dopammina-β-idrossilasi, che la converte in α-metilnoradrenalina. Quest'ultima viene depositata nei siti che normalmente trattengono la noradrenalina ed è liberata in seguito a stimolazione nervosa al posto della noradrenalina, l'autentico trasmettitore. Il ‛falso trasmettitore' è meno efficace del vero nel mantenere il tono simpatico.
Gli studi sulla dopa nel corpus striatum e nella substantia nigra hanno portato a un importante progresso terapeutico. L'effetto biochimico preminente associato con la malattia parkinsoniana è un'accentuata diminuzione della concentrazione di dopammina nelle aree sopra nominate del cervello. Questo ha portato all'impiego del precursore L-dopa nel trattamento del parkinsonismo, con vantaggio di numerosi pazienti refrattari ad altre misure terapeutiche.
Nel complesso, le ricerche nel campo delle catecolammine hanno portato ad alcuni dei più importanti progressi terapeutici moderni. Per esempio, noi possediamo ora un gruppo di farmaci, bloccanti i neuroni adrenergici, che interferiscono nella liberazione di noradrenalina dalle terminazioni nervose simpatiche e sono usati con successo nel trattamento a lungo termine dell'ipertensione essenziale.
4. Istamina.
a) Origine, chimica, formazione e distruzione.
L'istamina venne sintetizzata da A. Windaus e W. Vogt (1907) prima che fossero conosciute le sue azioni fisiologiche. Dale e collaboratori la isolarono dalla segale cornuta nel 1910 e ne studiarono gli effetti farmacologici. La presenza di istamina nella mucosa intestinale venne dimostrata subito dopo, ma venne considerata come dovuta a decarbossilazione batterica dell'istidina; solo attorno al 1920 l'istamina venne riconosciuta come costituente effettivo dell'organismo.
Molta ma non tutta l'istamina dei tessuti è contenuta nei mastociti, cellule che si colorano metacromaticamente, e nei leucociti basofili del sangue, elementi simili ai precedenti. I mastociti possono essere considerati organi endocrini unicellulari capaci di produrre eparina, acido ialuronico, istamina e 5-idrossitriptammina. Certi tessuti patologici, per es. quelli dell'orticaria pigmentosa, sono estremamente ricchi in istamina e mastociti. Nei mastociti l'istamina è presente in combinazione con l'eparina.
La struttura chimica dell'istamina è mostrata qui sotto.
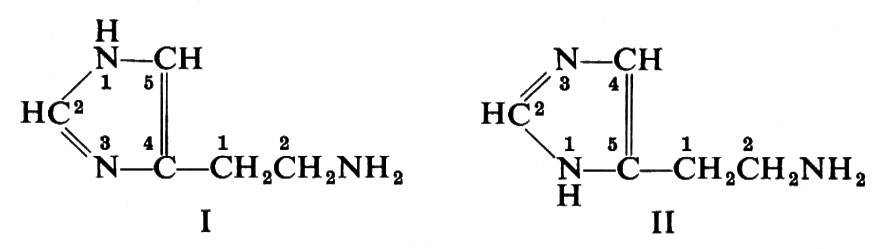
L'istamina dei tessuti di Mammifero ha origine dalla decarbossilazione intracellulare dell'istidina ad opera dell'istidindecarbossilasi. Per la sua attività questo enzima richiede il piridossalfosfato; l'enzima è inibito dalla semicarbazide e dalla α-metilistidina. La capacità dei tessuti di formare istamina è stata intensamente studiata da due gruppi di ricercatori. R. W. Schayer e collaboratori considerano l'istamina ‛indotta' quale regolatore intrinseco della microcircolazione in vari tipi di stress. G. Kahlson e collaboratori hanno studiato l'istamina ‛nascente', vale a dire la neoformazione d'istamina in tessuti in fase di rapido accrescimento. Questi autori hanno trovato che nel ratto l'istamina urinaria aumenta fortemente nella femmina gravida, come conseguenza dell'accresciuta capacità del fegato fetale di formare istamina. Un'aumentata capacità di formare istamina si ha anche durante la guarigione delle ferite nel ratto e durante l'accrescimento di alcune neoplasie. Un forte incremento di tale capacità si manifesta infine quando leucociti umani sensibilizzati vengono messi a contatto in vitro con l'antigene.
L'istamina è distrutta nel corpo soprattutto a opera di due sistemi enzimatici, l'istaminasi e la imidazolo-N-metiltransferasi. L'istaminasi è strettamente affine alla diamminossidasi, ma può essere distinta da questa (R. Kapeller-Adler, 1970). L'istaminasi degrada l'istamina ad aldeide imidazolacetica in una reazione richiedente la presenza di rame, piridossalfosfato e flavinadenindinucleotide (FAD). L'enzima è inibito da reagenti carbonilici, quali l'amminoguanidina. L'istaminasi è ampiamente distribuita nel corpo; elevate concentrazioni si ritrovano nella placenta umana, che è la fonte degli elevati livelli ematici di istaminasi riscontrabili nella gravidanza.
b) Azioni farmacologiche e fisiologiche.
L'istamina è un potente stimolante di certi tipi di muscoli lisci, fra cui in particolare quelli bronchiali, ed è un dilatatore dei piccoli vasi sanguigni, particolarmente dei capillari, che essa rende permeabili alle proteine plasmatiche, causando edema. Un effetto vascolare caratteristico dell'istamina è la ‛triplice risposta' del Lewis dopo sua iniezione nella pelle, consistente in una macchia rossa localizzata, un ponfo e un alone rosso vivo circostante. Questa reazione, che rassomiglia, anche per la sensazione di prurito, a un'orticaria localizzata, si ha anche per offese cutanee ed è allora probabilmente causata da istamina liberata dai mastociti. Dale ha suggerito l'ipotesi che la liberazione di istamina, agente sui capillari posti nelle immediate vicinanze, possa fornire un perfetto meccanismo per la regolazione del flusso sanguigno locale. L'istamina può pertanto fungere da ‛ormone locale'. Altre funzioni proposte per l'istamina sono più dubbie. È stato spesso ipotizzato che istamina liberata nel sangue possa essere la causa della iperemia reattiva o della vasodilatazione da esercizio, ma queste ipotesi non sono state sufficientemente comprovate. Similmente non è stata provata l'idea che una liberazione di istamina possa essere la causa dello shock traumatico, quantunque sia agevole dimostrare che l'iniezione endovenosa di un'elevata dose di istamina provoca dilatazione capillare, perdita di liquidi e collasso circolatorio.
L'istamina è un potente stimolatore della secrezione gastrica acida, per azione diretta sulle cellule parietali. Fin dalla prima dimostrazione di tale attività stimolatrice dell'istamina, a opera di L. Popielski (1920), si è molto discusso se l'ammina svolga un ruolo fisiologico nella secrezione gastrica. Attualmente molti ricercatori interessati al problema tendono a dare una risposta affermativa alla domanda. L'istamina è stata trovata nella mucosa gastrica di tutte le specie studiate e la sua concentrazione è massima nell'area in cui sono più numerose le cellule parietali. La capacità della mucosa gastrica di formare istamina aumenta considerevolmente dopo un pasto o dopo iniezione dell'ormone gastrico, la gastrina. È pertanto possibile che la gastrina possa agire attraverso una liberazione di istamina nella mucosa gastrica.
Non è noto se l'istamina abbia una funzione nella trasmissione neuroumorale. Elevate concentrazioni di istamina vengono ritrovate nei nervi simpatici postgangliari ma, a differenza di quanto avviene per la noradrenalina, il contenuto in istamina di questi nervi non diminuisce palesemente dopo sezione nervosa. È possibile che l'istamina sia contenuta nei mastociti della guaina connettivale piuttosto che nel neurone e che essa non abbia alcuna azione mediatrice diretta. L'istamina è presente anche in certe parti del sistema nervoso centrale, soprattutto nell'ipotalamo (H. M. Adam, 1960), in questo caso non in mastociti ma in sinaptosomi; ciò suggerisce una possibile funzione trasmettitrice dell'ammina nel sistema nervoso centrale.
c) Farmaci antistaminici; recettori istaminici.
Gli effetti dell'istamina su certe strutture a fibrocellule muscolari lisce (intestino di cavia, bronchi di cavia e di uomo) sono facilmente antagonizzati da un gruppo di sostanze chiamate antistaminici. Gli antistaminici, inizialmente scoperti da D. Bovet e dai suoi collaboratori, sono antagonisti competitivi dell'istamina a livello dei recettori. Le affinità degli antistaminici per i recettori sono in questi tessuti strettamente simili, suggerendo l'interessamento di un recettore uniforme, il recettore H1 (A. S. F. Ash e H. O. Schild, 1966). Altri effetti farmacologici dell'istamina, per es. il suo effetto stimolante sulla secrezione gastrica, non sono antagonizzati dagli antistaminici convenzionali. Ciò ha indotto a supporre e successivamente a dimostrare l'esistenza di un tipo diverso di recettore istaminico, l'H2, del quale si conosce attualmente un antagonista, la cimetidina.
Un'importante scoperta è stata la caratterizzazione, di Black e collaboratori, dei recettori H2 dell'istamina. Il lavoro di questi autori rappresenta un'applicazione pratica della teoria dei farmacorecettori e dell'antagonismo competitivo. Poiché gli effetti dell'istamina sulla secrezione gastrica non sono antagonizzati dalle antistamine classiche (H1), dovevano presumibilmente agire mediante recettori diversi. Questi autori hanno dimostrato per mezzo di una indagine sistematica che certi composti di nuova sintesi (in un primo tempo la burimammide e in seguito altri), strutturalmente simili all'istamina, antagonizzavano l'azione dell'istamina sulla secrezione gastrica, senza peraltro neutralizzare i suoi effetti H1 sulla muscolatura liscia. Si ritenne quindi che questi composti agissero sui recettori H2 dell'istamina. È interessante rilevare che anche le antistamine H2 erano antagoniste molto efficaci della secrezione dovuta all'ormone gastrico naturale, la gastrina; si poteva pertanto ritenere che l'istamina fosse un normale mediatore della secrezione gastrica. Gli antagonisti H2 sono da allora divenuti farmaci utili in clinica, particolarmente efficaci in alcuni tipi di ulcere peptiche.
d) Metodi di estrazione e di dosaggio.
L'istamina può essere estratta dai tessuti sfruttando la sua stabilità in soluzione acida: anche una prolungata ebollizione in acido non inattiva l'istamina, mentre distrugge altri agenti farmacologici che possono interferire nel suo dosaggio biologico. Tali metodi sono stati perfezionati con l'impiego di apparecchi di dosaggio automatici e di schemi programmati statisticamente. Sono disponibili anche metodi chimici di dosaggio dell'istamina; fra i quali un metodo fluorimetrico. Essi hanno all'incirca lo stesso grado di sensibilità dei metodi biologici.
e) Liberazione di istamina nell'anafilassi e a opera di liberatori chimici dell'istamina.
La liberazione di istamina nell'anafilassi e nell'allergia è uno degli esempi meglio accertati in cui un normale costituente delle cellule produce gravi effetti patologici quando venga liberato. La liberazione di istamina può essere dimostrata nel polmone isolato perfuso della cavia sensibilizzata. Quando si aggiunge l'antigene specifico al liquido di perfusione si ha, insieme con broncocostrizione e vasocostrizione, liberazione di istamina e sua comparsa nell'effluente venoso. La liberazione di istamina a opera dell'antigene può essere dimostrata anche per diffusione da tessuti isolati, per es. dal polmone sminuzzato.
Un preparato utile per l'identificazione di reagine umane è il polmone umano sminuzzato, sensibilizzato passivamente per incubazione con siero contenente reagine, e poi addizionato di antigene che provoca la liberazione di istamina. Un altro strumento utile nella diagnosi dell'allergia umana sono i leucociti di pazienti allergici, che liberano istamina quando siano esposti all'antigene (L. M. Lichtenstein e A. G. Osler, 1964). In questi casi l'istamina proviene dai mastociti tessutali o dai basofili ematici, loro equivalenti. Il processo di liberazione implica l'attivazione intracellulare di una reazione enzimatica che richiede calcio e la partecipazione di un ingrediente termolabile che viene inattivato per esposizione del tessuto a una temperatura lievemente superiore alla norma (v. fig. 8).
Molte sostanze, fra cui vari inibitori metabolici, interferiscono con la reazione di liberazione d'istamina nell'anafilassi. Un'interferenza interessante, poiché riguarda un ormone naturale, è quella da parte dell'adrenalina (o del suo congenere isoprenalina). Le catecolammine prevengono la liberazione di istamina nell'anafilassi a concentrazioni assai basse (E. S. K. Assem e Schild, 1969). Anche parecchi antipiretici interferiscono nel meccanismo dell'anafilassi.
È stato dimostrato che la reazione anafilattica cellulare si svolge in due momenti: il primo implica un meccanismo di bypass, il secondo porta alla liberazione di istamina. Se un inibitore viene applicato durante il primo stadio e successivamente rimosso, non si ha alcuna liberazione d'istamina. È così possibile effettuare una desensibilizzazione senza liberazione d'istamina. Sono disponibili inibitori che possono raggiungere questo risultato (cromoglicato disodico) e che possono essere utilizzati nel trattamento dell'asma bronchiale.
Varie ammine agiscono come liberatori d'istamina quando vengano messe a contatto con tessuti non sensibilizzati. Esse possono agire per un semplice meccanismo di scambio ionico, spostando l'istamina dal suo legame con l'eparina. Altri liberatori d'istamina agiscono in modo più complesso, inducendo reazioni che sono affini alla reazione anafilattica e possono essere usate come sistemi modello per lo studio dell'anafilassi.
5. 5-Idrossitriptammina (serotonina, enterammina).
In questa trattazione degli ormoni tessutali procediamo da quelli meglio noti a quelli meno noti. Nel caso dell'acetilcolina e delle catecolammine è già disponibile una vasta massa di conoscenze sulle loro azioni fisiologiche, sebbene rimanga ancora molto da scoprire. Nel caso dell'istamina numerose funzioni sono state proposte, ma nessuna di esse è stata provata in modo inequivocabile. Questo è valido ancor più per il campo della 5-idrossitriptammina (5-HT), in cui persiste ancora molta incertezza che forse contribuisce al suo fascino.
La storia recente della 5-HT inizia col lavoro di M.M. Rapport nel 1949, che identificò la sostanza nel siero. Apparve subito evidente che la serotonina di Rapport era identica all'enterammina, scoperta molti anni prima da V. Erspamer nei tessuti di Vertebrati e Invertebrati. I primi lavori sulla 5-HT si basavano soprattutto sul saggio biologico. I tessuti venivano estratti con acetone, che elimina i polipeptidi e altre sostanze interferenti, e l'attività 5-idrossitriptamminica veniva determinata su preparati quali l'utero di ratto o la sensibile striscia di stomaco di ratto. Ora sono stati svilupppati per la titolazione della 5-HT metodi fluorimetrici che, per quanto meno sensibili dei metodi biologici, trovano impiego crescente.
La 5-HT è stata ritrovata in parecchie specie di Invertebrati. Nei Mammiferi essa è principalmente concentrata nel tratto gastrointestinale, ove è contenuta in cellule appartenenti al sistema enterocromaffine. La 5-HT del sangue è contenuta nelle piastrine. 5-HT è presente anche nel sistema nervoso centrale, soprattutto nell'ipotalamo, mesencefalo e midollo spinale. Le vie metaboliche di trasformazione e biosintesi della 5-HT nell'organismo di Mammifero sono riportate nello schema 2.
La 5-HT esplica varie azioni farmacologiche, per esempio è un potente broncocostrittore. Essa produce sia vasocostrizione, sia vasodilatazione; nel ratto è fornita di forte azione antidiuretica, probabilmente per vasocostrizione renale (Erspamer). La droga allucinogena dietilammide dell'acido lisergico (LSD) è un potente antagonista periferico della 5-HT, per es. della sua azione stimolante sul muscolo liscio dell'utero di ratto. La 5-HT stimola anche l'intestino di cavia, ma questa azione è ritenuta indiretta, in quanto dovuta a liberazione di acetilcolina dalle terminazioni nervose.
La reserpina, un farmaco con attività antipsicotica e antipertensiva, ha il rimarchevole effetto di svuotare i tessuti, compreso il cervello, e le piastrine del sangue del loro contenuto in 5-HT. Le piastrine normalmente captano avidamente la 5-HT contro un gradiente di concentrazione, ma perdono questa loro capacità per effetto della reserpina, la quale sembra distruggere il loro speciale meccanismo di captazione entro granuli intracellulari ‛osmiofili'.
a) La 5-HT come mediatore dell'impulso nervoso.
Ci sono notevoli prove, per quanto non del tutto conclusive, che la 5-HT sia un trasmettitore neuroumorale. Le prove si basano sul fatto che la 5-HT è contenuta in certi neuroni insieme con gli enzimi necessari per la sua sintesi e la sua inattivazione; che essa è depositata in tipiche vescicole sinaptiche; che nel sistema nervoso centrale essa ha una velocità di ricambio molto elevata; che essa produce stimolazione neuronica quando venga applicata per microelettroforesi; infine che essa viene, almeno in certi casi, liberata per stimolazione nervosa. La sua presenza nel sistema nervoso centrale può essere dimostrata con metodi di fluorescenza istochimica. Grossi neuroni contenenti 5-HT sono stati osservati soprattutto negli Invertebrati. Una liberazione di 5-HT dal midollo spinale, dopo stimolazione elettrica, è stata dimostrata nella rana e nel topo (N. E. Anden e altri, 1964). E. Bülbring e altri (1968) hanno trovato che la stimolazione elettrica dello stomaco di topo provoca una liberazione di 5-HT, impedita dalla tetrodotossina, e perciò suggeriscono che la 5-HT possa essere un trasmettitore in corrispondenza della sinapsi gangliare inibitrice del vago. Ci sono considerevoli prove che la 5-HT intervenga nel processo di regolazione centrale della temperatura: così Feldberg e collaboratori trovarono che l'introduzione intratecale di 5-HT nel gatto innalzava la temperatura corporea, mentre la noradrenalina l'abbassava.
La sintesi della 5-HT nell'organismo può essere quasi completamente bloccata dalla p-clorofenilalanina, che inibisce l'azione dell'enzima triptofano-5-idrossilasi, ma gli effetti dell'inibizione sono sorprendentemente piccoli e piuttosto diffusi, e consistono in un abbassamento della soglia convulsiva e in iperalgesia. Forse i processi di trasmissione triptamminergica hanno un fattore di sicurezza assai elevato, che permette loro di operare in condizioni di liberazione minima. E stato ipotizzato che le vie 5-idrossitriptamminiche possano differire da quelle incaricate del trasferimento rapido e preciso d'informazioni. Esse hanno spesso basse velocità di conduzione e forse funzionano da modulatrici dell'attività neuronica in parecchie regioni del sistema nervoso centrale, particolarmente nelle porzioni filogeneticamente più antiche. Concentrazioni particolarmente elevate di 5-HT si hanno nel sistema del rafe del midollo allungato e del mesencefalo. M. Jouvet (1968) ha dimostrato che questo sistema ha una funzione importante nel sonno e che la sua lesione meccanica o chimica, quest'ultima ottenuta attraverso l'eliminazione della 5-HT a mezzo della p-clorofenilalanina, porta, nel gatto, a insonnia permanente che può essere alleviata con la somministrazione di 5-idrossitriptofano. Questi dati indicano che il sonno può essere un processo attivo dipendente da neuroni triptamminergici.
b) Implicazioni cliniche.
Uno dei primi reperti concernenti la 5-HT in condizioni morbose (F. Lembeck, 1953) fu quello dell'elevato contenuto di 5-HT nei tumori carcinoidi. I pazienti con carcinoidi eliminano nelle urine 5-HT e acido 5-idrossundolacetico (5-HIAA); la titolazione di quest'ultimo fornisce un mezzo per la conferma della diagnosi di carcinoide. I pazienti affetti da carcinoide presentano episodi di colica intestinale, di broncocostrizione e di arrossamento cutaneo (flushing), tutti fenomeni attribuiti alla 5-HT, per quanto ora si ritenga che l'arrossamento possa in parte dipendere dalla formazione di chinine. E stata anche suggerita l'ipotesi che la 5-HT possa essere implicata nella tossiemia gravidica e nell'emicrania: alcuni pazienti con emicrania eliminano con le urine un'aumentata quantità di acido 5-idrossundolacetico. È possibile che la 5-HT abbia una parte nella patogenesi di taluni difetti mentali congeniti: nella fenilchetonuria, in cui non avviene l'ossidazione della fenilalanina a tirosina, c'è una diminuzione del tasso plasmatico di 5-HT e ciò giustifica l'ipotesi che nella malattia sia compromessa tanto l'ossidazione della fenilalanina quanto quella del triptofano.
c) Derivati indolici similari.
Un importante derivato indolico sintetizzato nell'organismo è la melatonina, trovata nella ghiandola pineale. La melatonina è la N-acetil-5-metossitriptammina. Nei Mammiferi la ghiandola pineale è l'unico organo contenente l'enzima che catalizza l'ultimo passaggio nella sintesi della melatonina. La melatonina produce vari effetti, per lo più inibitori; fra l'altro essa deprime lo sviluppo dell'ovaio e la secrezione dell'ormone tiroideo. La sintesi della melatonina nella pineale varia diurnalmente; l'esposizione alla luce la deprime, l'oscurità ha l'effetto opposto. J. Axelrod ha suggerito l'ipotesi che le fluttuazioni ritmiche nella secrezione della melatonina possano fornire all'organismo un ‛orologio biologico' che sincronizza i ritmi in altri organi.
Molti derivati indolici esplicano effetti farmacologici. La psilocibina, un derivato naturale della 4-idrossitriptammina, è un antagonista di questa. Essa possiede nell'uomo effetti allucinogeni simili a quelli dell'LSD, che pure è un derivato indolico sostituito in posizione 4 (v. droga). La metisergide, composto sintetico, è strettamente affine all'LSD: essa è un antagonista della 5-HT ed è stata usata nel trattamento di pazienti affetti da carcinoide e nell'emicrania.
6. Chinine.
L'organismo utilizza un elaborato sistema di meccanismi di difesa in cui gli enzimi proteolitici esplicano un ruolo importante. Questi meccanismi comprendono la coagulazione del sangue, la fibrinolisi, i sistemi del complemento e delle chinine; il loro effetto precipuo è quello di produrre emostasi, rimozione dei coaguli di sangue, attivazione delle reazioni anticorpali e vasodilatazione in seguito a lesioni dei tessuti. È probabilmente in quest'ultimo tipo di attività che interviene particolarmente la formazione di chinine. Gli enzimi proteolitici implicati in queste varie reazioni di difesa sono correlati fra loro, ma nella nostra trattazione ci limiteremo agli enzimi che hanno direttamente a che fare col sistema formatore di chinine. Un sistema proteolitico affine di grande importanza, il sistema renina-angiotensina, viene discusso altrove (v. ipertensione arteriosa).
Ricercatori tedeschi (E. K. Frey, H. Kraut, E. Werle) descrissero, negli anni trenta, una sostanza trovata nell'urina umana, la callicreina, che produceva una caduta della pressione del sangue se iniettata nel cane. La callicreina era generalmente inattiva su preparati isolati, ma dopo incubazione con siero essa dava origine, per azione enzimatica, a una sostanza capace di contrarre l'intestino (Darmkontrahierende Subrtanz, DKS, callidina).
Indipendentemente, M. Rocha e Silva e collaboratori trovarono che incubando con certi veleni di serpente le globuline plasmatiche, queste davano origine a un prodotto farmacologicamente attivo, che essi chiamarono bradichinina.
a) Callicreina.
La callicreina interviene nella produzione delle chinine come indicato nello schema 3. Si vede che la callicreina è normalmente presente sotto forma di precursore inattivo richiedente attivazione; l'enzima attivato agisce poi su una o più frazioni proteiche del sangue dando origine alle chinine farmacologicamente attive. È nota l'esistenza di almeno due forme di callicreina: la callicreina plasmatica e la callicreina ghiandolare, trovata nel pancreas, nelle ghiandole salivari e nell'urina. Si ritiene che la callicreina plasmatica liberi bradichinina, quella ghiandolare callidina. Alcuni ricercatori distinguono due diverse forme di callicreina plasmatica. Un attivatore fisiologico del callicreinogeno è il fattore di Hageman (fattore XII della coagulazione), che circola sotto forma inattiva e che dopo attivazione innesca non solo il sistema chimico, ma anche la coagulazione del sangue e la fibrinolisi. Il callicreinogeno può essere attivato anche da un basso pH, cosa che può essere importante nelle lesioni tessutali. La callicreina è una idrolasi proteolitica con specifiche esigenze di substrato; essa scinde certe ammidi e certi esteri di amminoacidi, particolarmente quelli dell'arginina. Un buon substrato artificiale per la callicreina è il tosil-arginina-metilestere (TAME). Il sito attivo della callicreina può essere inibito dal dii sopropilfiuorofosfato (DFP). L'inibitore tripsinico dei semi di soia inibisce la callicreina del siero, ma non quella pancreatica.
La callicreina serica è inattivata dal siero umano. Si ritiene che questa inattivazione, che è irreversibile, sia dovuta alla presenza nel siero di un inibitore avente alta affinità per il sito attivo della callicreina. L'inibitore della callicreina è strettamente affine all'inibitore della tripsina di Kunitz ed è disponibile in commercio (Trasylol). Mentre il callicreinogeno è di per sé fisiologicamente inattivo, esso viene attivato dopo iniezione endovenosa, nel cane, producendo una caduta della pressione del sangue dovuta a formazione di chinina. La caduta della pressione ematica può essere utilizzata per il saggio biologico della callicreina. Un altro metodo di dosaggio consiste nell'incubare la callicreina con un idoneo substrato di proteine plasmatiche e nel misurare poi la formazione di chiama col saggio biologico su un muscolo liscio isolato.
b) Formazione e distruzione delle chinine.
Parecchie chinine sono state identificate chimicamente e sintetizzate. Fra le più importanti sono: 1) l'enneapeptide bradichinina: Arg-Pro-ProGly-Phe-Ser-Pro-Phe-Arg; 2) il decapeptide Lys-bradichinina o callidina: Lys-Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg; 3) l'endecapeptide Met-Lysbradichinina.
I chininogeni del siero, da cui prendono origine le chinine, sono glicoproteine acide di peso molecolare all'incirca 50.000. Il plasma nativo possiede un'attività amminopeptidasica capace di trasformare la callidina disponibile in bradichinina. Il plasma e il siero contengono anche delle chininasi, che distruggono le chinine. Le chininasi possono essere inattivate da inibitori specifici, quali la o-fenantrolina; questi inibitori debbono essere aggiunti alla miscela di reazione ogni qual volta si misuri la formazione di chinine a opera della callicreina.
c) Azioni farmacologiche e possibili ruoli fisiolo gici e patologici delle chinine.
Le chinine sono dotate di elevata attività farmacologica. La bradichinina stimola la contrazione di alcuni preparati isolati di fibrocellule muscolari lisce, tra i quali quelli di utero di ratto, di ileo di cavia, di duodeno di coniglio e di gatto, mentre determina il rilasciamento di altre strutture muscolari lisce, quali il duodeno e il colon di ratto. Per identificare le chinine possono essere impiegate varie reazioni farmacologiche specifiche, facendo uso del principio dei saggi paralleli quantitativi rispetto a un campione comune (J. H. Gaddum). Finora non sono stati individuati antagonisti specifici delle chinine, ma il potente effetto broncocostrittore della bradichinina nella cavia può essere antagonizzato dall'aspirina.
La bradichinina è una delle più potenti sostanze vasodilatatrici conosciute: essa abbassa la pressione del sangue in tutte le specie esaminate. Nell'animale integro la sostanza accresce la gettata cardiaca e produce dilatazione delle coronarie. Un'interessante proprietà delle chinine è quella di produrre dolore quando siano iniettate per via endoarteriosa o applicate sulla base di una vescichetta aperta (D. Armstrong e C. A. Keele). Per quanto le terminazioni nervose si desensibilizzino rapidamente all'effetto algogeno delle chinine, la formazione di queste è verosimilmente un importante fattore nell'insorgenza del dolore da lesione tessutale. Le chinine esplicano effetti sistemici e locali sul circolo. Esse possono liberare adrenalina dalla midollare surrenale, ed è probabile che svolgano un ruolo importante nell'infiammazione; ovunque un tessuto sia leso, il sistema formatore di chinine è facilmente attivato. Le chinine possono produrre vasodilatazione nelle ghiandole sottomascellari e in altre ghiandole, durante la loro attività, e molto probabilmente intervengono anche nelle reazioni allergiche.
d) Polipeptidi similari.
La sostanza P (Euler e Gaddum, 1931) è un polipeptide scoperto in estratti d'intestino e di cervello di cavallo. La sua azione è simile a quella della bradichinina, ma distinguibile da essa. Parecchi nuovi peptidi sono stati scoperti da Erspamer e collaboratori in Vertebrati inferiori e in Invertebrati.
Fra essi l'eledoisina ottenuta dalle ghiandole salivari di Eledone, la fisalemina ottenuta dalla pelle di Physalaemus e la ceruleina trovata nella pelle di Hyla caerulea. Tutti questi peptidi sono ora caratterizzati chimicamente. La pelle degli Anfibi contiene anche bradichinina preformata.
7. Prostaglandine: effetti farmacologici e possibili funzioni.
La presenza nel liquido seminale umano di un'attività ipotensiva e stimolatrice per la fibrocellula muscolare liscia fu scoperta indipendentemente da M. W. Goldblatt e U. S. von Euler nel 1933-1934. L'attività era dovuta a un materiale acido liposolubile che fu chiamato prostaglandina (PG). In seguito si vide che molti tessuti differenti contengono varie prostaglandine.
L'isolamento e la caratterizzazione chimica delle prostagì andine furono condotti a termine da ricercatori svedesi (S. Bergström e collaboratori) con l'uso di modemi metodi di gascromatografia, spettrometria di massa e cristallografia a raggi X. Chimicamente tutte le prostaglandine possono essere considerate come derivati dell'acido prostanoico, acido grasso a 20 atomi di carbonio contenente un anello a cinque termini. Viene riportata qui di seguito la formula di una delle prostaglandine (PGE1).
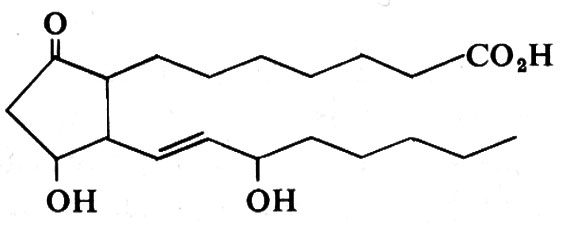
Sono state isolate almeno 14 diverse prostaglandine, divise in quattro serie designate con le lettere A, B, E, F. Sebbene tutte le prostaglandine siano strutturalmente simili, esiste una grande varietà nelle loro proprietà biologiche.
Azione sull'utero. - Le prostaglandine hanno un effetto prevalentemente stimolante sull'utero e poiché lo sperma è una fonte concentrata di prostaglandine si è pensato che esse possano avere una funzione nel trasporto dello sperma, attraverso l'attivazione dei movimenti uterini. Le prostaglandine sono assorbite dalla vagina e possono pertanto interessare il tratto genitale femminile sia per la loro azione locale sia dopo assorbimento sistemico. La PGF2~, in particolare, è un potente stimolatore uterino ed è stata usata per provocare le contrazioni dell'utero a termine di gravidanza o per indurre l'aborto.
Azione sul sistema nervoso centrale. - Le prostaglandine possono avere funzioni di mediatori nel sistema nervoso centrale e sono state identificate come costituenti naturali del cervello in svariate specie. A differenza di quanto avviene per altri ipotetici trasmettitori, tuttavia, la loro distribuzione non è localizzata chiaramente nel cervello. Dette sostanze sono costituenti normali del liquido cerebrospinale e sono liberate dal cervello a riposo e dal midollo spinale dopo stimolazione afferente.
Le prostaglandine hanno potenti effetti sul sistema nervoso centrale. La PGE1 iniettata nei ventricoli cerebrali del gatto non anestetizzato produce torpore e catatonia, che iniziano 20-30 minuti dopo l'iniezione e durano parecchie ore (E. W. Horton). L'applicazione microionoforetica di PGE1 e PGF2~ a cellule del tronco encefalico di gatti decerebrati produce una scarica in un'aliquota di questi neuroni.
Le prostaglandine possono fungere da trasmettitori a livello delle terminazioni nervose periferiche, come risulta dai seguenti esperimenti. La stimolazione antidromica del nervo trigemino causa una miosi di lunga durata, per la prima volta osservata da Cl. Bernard. N. Ambache e collaboratori studiarono questo fenomeno attraverso l'esame dell'attività miotica di estratti di tessuto irideo e scoprirono una sostanza provvista di attività stimolante sulla muscolatura liscia, che essi chiamarono irina. L'irina è probabiimente una miscela di prostaglandine è stato trovato che le prostaglandine iniettate nella camera anteriore dell'occhio producono miosi.
Nel tessuto adiposo ha luogo un'interessante interazione fta nervi adrenergici e prostaglandine. La stimolazione adrenergica causa un aumento della lipolisi, probabilmente attraverso un'azione sull'AMP ciclico. Le prostaglandine antagonizzano questo effetto e diminuiscono la lipolisi. È stato dimostrato che prostaglandine vengono liberate dal tessuto adiposo in risposta a stimolazione nervosa adrenergica e ad apporto di catecolammine. Questo può rappresentare un meccanismo di feedback negativo col quale le prostaglandine modulano effetti dipendenti da un sistema con AMP ciclico. Meccanismi similari possono operare in altri sistemi attivanti l'AMP ciclico.
8. Conclusione.
La lista delle attività ormonali prese in esame in questo articolo non è per nulla completa. Fra quelle omesse sono da ricordare gli effetti cardiaci dei composti adenosinici, gli effetti sulla permeabilità capillare di polipeptidi a elevato peso molecolare e infine gli effetti neuroumorali di certi amminoacidi. L'intero argomento è di sua propria natura inesauribile e continuerà a essere fonte fruttuosa di nuove ricerche e di problemi interessanti.
Bibliografia.
Achesom, G. A. (a cura di), Second symposium on catecholamines, in ‟Pharmacological review", 1966, XVIII, pp. 1-803.
Bergström, S., Samuelsson, B. (a cura di), Prostaglandins. 2. Nobel symposium, New York 1967.
Dale, H. H., Feldberg, W., Vogt, M., Release of acetylcholine at voluntary motor nerve endings, in ‟Journal of physiology", 1936, LXXXVI, pp. 353-380.
Erdös, E. G. (a cura di), Bradykinin, kallidin and kallikrein, in Handbuch der experimentellen Pharmakologie, vol. XXV, Berlin 1970.
Ersparmer, V. (a cura di), 5-Hydroxytryptamine and related indolealkylamines, in Handbuch der experimentellen Pharmakologie, vol. XIX, Berlin 1966.
Euler, U. S. von, Noradrenaline: chemistry, physiology, pharmacology and clinical aspects, Springfield, Ill., 1956.
Garattini, S., Shore, P. A. (a cura di), Biological role of indolealkylamine derivatives, in ‟Advances in pharmacology", 1968, VI, part B.
Horton, E. W., Hypothesis on physiological roles of prostaglandins, in ‟Physiological reviews", 1969, XLIX, pp. 121-161.
Kahlson, G., Rosengren, E., Histamine, in ‟Annual review of pharmacology", 1965, V, pp. 305-320.
Katz, B., The release of neural transmitter substances, Liverpool 1969.
Koelle, G. B. (a cura di), Symposium on neurohumoral transmission, in ‟Pharmacological review", 1954, VI, pp. 3-123.
Koelle, G. B. (a cura di), Cholinesterases and anticholinesterase agents, in Handbuch der experimentellen Pharmakologie, vol. XV, Berlin 1963.
Loewi, O., Über humorale Übertragbarkeit der Heznervenwirkung, in ‟Pflügers Archiv", 1921, CLXXXIX, pp. 239-242.
Mongar, J. L., Schild, H. O., Cellula mechanisms in anaphylaxis, in ‟Physiological reviews", 1962, XLII, pp. 226-270.
Pickles, V. R., The prostaglandins, in ‟Biological reviews", 1967, XLII, pp. 614-652.
Rocha e Silva, M. (a cura di), Histamine. Its chemistry, metabolism and physiological and pharmacological actions, in Handbuch der experimentellen Pharmakologie, vol. XVIII, t. 1, Berlin 1966.
Rocha e Silva, M. (a cura di), Kinin hormones: with special reference to bradykinin and related hormones, Springfield, Ill., 1970.
Schümann, H. J., Kroneberg, G., New aspects of stoage and release mechanisms of catecholamines, New York 1970.
Vane, J. R. (a cura di), Adrenergic mechanisms, London 1960.
Ormoni vegetali
SOMMARIO: 1. Generalità. □ 2. Fenomeni di correlazione ormonale e identificazione degli ormoni nelle piante. □ 3. Ormoni vegetali e organizzazione della pianta. □ 4. Auxine: a) definizione, aspetti biochimici, biosintesi; b) effetti fisiologici. □ 5. Etilene: a) aspetti biochimici, biosintesi; b) effetti fisiologici. □ 6. Gibberelline: a) definizione, aspetti biochimici, biosintesi; b) effetti fisiologici. □ 7. Citochinine: a) definizione, aspetti biochimici, biosintesi; b) effetti fisiologici. □ 8. Acido abscissico e abscissine o dormine: a) definizione, aspetti biochimici, biosintesi; b) effetti fisiologici. □ 9. Gli ormoni vegetali come sistema regolatore unitario. □ 10. Meccanismo d'azione degli ormoni vegetali. □ Bibliografia.
1. Generalità.
Ormoni, dal greco ὁρμάω, stimolo, sono definite quelle sostanze che, prodotte in parti specializzate di un organismo, da queste possono muovere ad altre parti esercitandovi, a concentrazioni di solito molto basse, effetti fisiologici caratteristici. Questa definizione implica tre idee centrali: 1) la capacità di gruppi di cellule di un organismo di specializzarsi per quanto riguarda taluni aspetti del metabolismo, in modo che determinate sostanze (come appunto gli ormoni) sono sintetizzate solo, o in modo prevalente, in alcune regioni e non in altre; 2) la capacità di sostanze attive, prodotte solo da alcuni tessuti od organi, di muovere verso altri; 3) la capacità dei tessuti od organi di reagire con risposte caratteristiche a fattori chimici provenienti da altri territori. Gli ormoni vengono così a rappresentare dei messaggeri chimici che collegano tra loro parti differenziate di un organismo, integrandone in modo unitario il comportamento. L'insieme delle correlazioni ormonali costituisce una funzione, in quanto assicura la coordinazione tra le diverse attività dell'organismo.
La funzione delle correlazioni ormonali si presta a essere considerata sotto due aspetti principali: a) del suo significato generale ai fini della sopravvivenza dell'organismo o della specie; b) del suo meccanismo. Entrambi questi aspetti possono essere considerati a livelli diversi. I tipi chimici di mediatori ormonali sono molteplici, differendo per caratteristiche di origine metabolica, di natura chimica e di attività fisiologica; inoltre, i diversi ormoni possono interagire, talora potenziandosi, talora neutralizzandosi reciprocamente.
Ogni cellula, in un organismo pluricellulare, è quindi soggetta all'azione integrata di una ‛costellazione' di ormoni che ne controlla il comportamento.
Un altro aspetto importante della definizione di ormone è che essa si restringe a quelle sostanze che agiscono sul comportamento cellulare a concentrazioni estremamente ridotte. Ciò esclude dalla definizione di ormone tutti i fattori di tipo nutritizio, che pure possono migrare dal sito di origine (per es. dalle foglie, per gli zuccheri ivi fotosintetizzati) a quelli di utilizzazione. Ma un aspetto più importante di questa restrizione è nel fatto che essa sottolinea la capacità di intensa reazione del comportamento della cellula a stimoli chimici anche quantitativamente esilissimi (esistono ormoni che agiscono a concentrazioni estremamente piccole, inferiori a un microgrammo per chilogrammo di tessuto di accettore). Ciò implica la presenza di meccanismi tali da assicurare un'enorme amplificazione degli effetti primari delle poche molecole che, giungendo alla cellula dall'esterno, vengono a interferire con i suoi sistemi intrinseci di regolazione.
Il progresso delle conoscenze nei campi della chimica biologica e della biologia molecolare dà oggi esauriente documentazione dell'esistenza di questi sistemi di amplificazione e, nello stesso tempo, permette la loro classificazione in categorie diverse, a seconda della natura dei sistemi di regolazione sui quali i diversi ormoni possono intervenire.
2. Fenomeni di correlazione ormonale e identificazione degli ormoni nelle piante.
La possibilità di rilevamento di un ormone si basa sull'esistenza, nello stesso organismo, di parti che producono ed esportano una determinata sostanza attiva, accanto ad altre che non producendola (o eventualmente producendola in quantità insufficiente) la possono assorbire, reagendo a essa in modo manifesto e caratteristico. La scoperta della maggior parte degli ormoni sia animali sia vegetali segue una linea relativamente monotona che può così schematizzarsi: primo, si osserva che l'asportazione o l'inattivazione patologica di un organo induce determinate alterazioni del comportamento di altre parti dell'organismo; secondo, l'innesto dell'organo isolato, o la somministrazione di suoi estratti o di quelle sostanze che da esso diffondono, determinano la soppressione o la riduzione, nell'organismo amputato o in parti isolate di esso, delle alterazioni conseguenti all' amputazione; terzo, l'individuazione della natura chimica del fattore attivo presente negli estratti apre la via allo studio della sua sintesi e del suo meccanismo d'azione. Così, per esempio, sono state scoperte negli animali l'insulina, prodotta nel pancreas e ricevuta dagli organi che in vario modo regolano il livello del glucosio nel sangue; gli ormoni sessuali, prodotti dalle gonadi e responsabili dello sviluppo dei caratteri sessuali secondari; la tiroxina, prodotta dalla tiroide e attiva sul metabolismo ossidativo in genere; e, nelle piante, le auxine, prodotte dagli apici meristematici e dalle foglie, e attive sulla distensione della parete cellulare dei tessuti prossimi agli apici stessi, oltre che su tutta una serie di processi più avanti dettagliatamente descritti. Questo schema generale di rilevamento della presenza e dell'attività degli ormoni comporta d'altra parte tutta una serie di varianti a esso sostanzialmente riconducibili: per es., altre due classi di ormoni vegetali sono state individuate osservando l'effetto su piante, o su parti isolate di esse, di fattori presenti in preparati non vegetali (come nel caso delle citochinine, presenti nell'idrolisato di acidi nucleici di sperma di salmone) oppure prodotti da parassiti (caso delle gibberelline, elaborate da un fungo parassita delle Graminacee). La classificazione delle citochinine e delle gibberelline come ormoni vegetali ha dovuto tuttavia attendere la dimostrazione che esse sono prodotte dalle piante superiori ove sono capaci di muoversi da tessuto a tessuto, nonché la loro purificazione e individuazione chimica e l'analisi dei loro effetti quando somministrate pure e a dosi fisiologiche. Un aspetto che emerge in modo costante nel rilevamento di ogni ormone è quello della specificità di azione, intesa in due sensi: 1) specificità di organo (‛bersaglio'), in quanto uno stesso ormone tende a influenzare, in un organismo, solo determinati organi o tessuti; 2) specificità di risposta, in quanto ogni ormone, pur essendo in genere capace di controllare processi fisiologici anche molto diversi, e in modo diverso a seconda della diversa natura o condizione degli organi-bersaglio, restringe la propria azione a un campo relativamente ben definito di effetti fisiologici. La specificità di organo e la specificità di azione dei singoli ormoni sono alla base della possibilità di allestire test biologici per il loro rilevamento: per esempio, - e limitandoci a tre tipi di ormoni vegetali, le auxine, le gibberelline e le citochinine - la quantità di auxina presente in un estratto, anche se accompagnata da altri ormoni, può essere determinata con precisione misurando l'allungamento che essa induce su di un segmento lungo 3-10 mm di coleottile (guaina delle prime foglioline delle Graminacee) di avena prelevato in fase di accrescimento; la quantità di gibberellina, misurando il suo effetto nello stimolare la produzione di un enzima, l'α-amilasi, prodotta dagli strati esterni (aleurone) dell'endosperma di semi d'orzo cui sia stato asportato l'embrione; la quantità di citochinina, dall'effetto nel promuovere l'ingrandimento delle cellule di giovani cotiledoni di semi di ravanello, ecc. In tutti questi test, ognuno degli ormoni sopracitati induce infatti una risposta quantitativamente ben definita e notevolmente specifica.
Un'ultima osservazione generale a proposito della caratterizzazione dell'azione degli ormoni e dell'individuazione degli organi (o tessuti) produttori e degli organi-bersaglio, è che non sempre, o non necessariamente, la capacità dell'organo-bersaglio di reagire alla somministrazione di ormone richiede che esso sia incapace di produrre l'ormone; in certi casi, il basso, o comunque non ottimale, livello dell'ormone nel bersaglio può essere determinato dal prevalere in esso di condizioni favorevoli a una sua rapida distruzione. È necessario tener presente che la concentrazione di un ormone nella cellula dipende ovviamente dalla differenza tra la somma di fattori che tendono ad aumentarne il livello e la somma di quei fattori che tendono a diminuirlo; i primi sono evidentemente l'assorbimento netto (assunzione meno fuoriuscita) dall'esterno e la capacità di sintesi all'interno della cellula, mentre i secondi sono rappresentati essenzialmente dalla velocità di demolizione o inattivazione metabolica. Lo stesso carattere degli ormoni quali regolatori del comportamento cellulare impone che essi possano essere prontamente inattivati o distrutti, in modo che si eviti il pericolo di un loro accumulo non controllato nelle diverse regioni dell'organismo. L'integrarsi del duplice meccanismo di sintesi in siti specializzati e di inattivazione fisiologicamente controllata negli organi-bersaglio permette il mantenimento nell'organismo di ben definiti gradienti delle concentrazioni dei diversi ormoni; gradienti sul valore dei quali si basa, in ultima analisi, il coordinamento delle varie attività delle singole parti dell'individuo in un quadro funzionale unitario. Il significato del controllo del livello di ogni ormone, in ogni momento e in ogni regione dell'organismo, risulta chiaro dall'analisi della relazione tra concentrazione ed effetto fisiologico. Solo in pochi casi, infatti, l'intensità di un determinato effetto ormonale è funzione semplice della concentrazione - o dose - in ormone; molto più spesso variazioni di concentrazione dell'ormone determinano variazioni complesse, anche qualitative, dell'attività fisiologica. Appare tipico, a questo proposito, il comportamento delle auxine che, in molti tessuti, a bassa concentrazione stimolano, ad alta concentrazione inibiscono una serie di processi collegati con l'accrescimento. Questo fenomeno, descritto in maggior dettaglio più avanti, è oggi soddisfacentemente interpretato come conseguenza della capacità delle auxine di indurre, quando presenti a concentrazioni superiori a una determinata soglia, la sintesi di un altro ormone, l'etilene, ad azione prevalentemente inibente e quindi in larga misura antagonista di quella tipica delle auxine.
La descrizione della natura e della funzione di ogni ormone si risolve nell'esposizione di quanto si conosce a proposito dei seguenti punti : a) scoperta e isolamento del fattore chimico responsabile di un dato fenomeno ormonale; b) delucidazione della sua struttura chimica; c) individuazione dei siti di origine e della via biochimica della sua sintesi e della sua demolizione nell'organismo; d) fisiologia del suo trasporto; e) suoi effetti fisiologici e morfologici negli organi-bersaglio; f) suo meccanismo d'azione a livello molecolare; g) sue interazioni con altri stimoli ormonali e, in generale, con i diversi fattori che influenzano il comportamento degli organi-bersaglio.
3. Ormoni vegetali e organizzazione della pianta.
Gli ormoni vegetali presentano, rispetto a quelli animali, una serie di divergenze corrispondenti a diversità nei piani organizzativi caratteristici dei due tipi di organismi. Nell'animale l'accrescimento tende a esaurirsi in una fase relativamente precoce, e i diversi organi e tessuti (prodotti da un processo molto spinto di differenziamento) mantengono dimensioni e caratteri morfologici immutati per un lungo periodo, durante il quale essi svolgono attività fisiologiche qualitativamente e quantitativamente diverse nei diversi momenti. Negli animali, sia i processi di crescita e di differenziamento, sia l'attività degli organi già differenziati sono quindi soggetti a fenomeni di regolazione ormonale. Nella pianta, al contrario, i processi di sviluppo sono più protratti nel tempo, continuando per molti organi fino alla morte; il differenziamento è molto più blando, e investe un numero ristretto di componenti cellulari (tra i fattori morfologicamente più appariscenti, essenzialmente le pareti cellulari, i vacuoli e i plastidi); pochi organi, come la foglia, conservano per un tempo relativamente lungo una struttura definitiva capace di sostenere variazioni fisiologiche reversibili e sottoposte al controllo di stimoli esterni o interni. Inoltre la pianta, a differenza dell'animale, tende a risolvere il problema della propria autonomia non tanto isolandosi dall'ambiente o modificandolo, come fa l'animale (si pensi a omeotermia, locomozione, attività connesse allo sviluppo del sistema nervoso), bensi, piuttosto, rimanendo aperta all'ambiente e ricevendone gli stimoli in modo da poter a esso rispondere in modo adeguato (arresto di sviluppo ed entrata in dormienza quando le condizioni esteme sono sfavorevoli; sensibilità al variare del ritmo stagionale giorno-notte per quanto riguarda la riproduzione; regolazione della crescita in funzione della disponibilità di sostanze nutritizie, ecc.). Non stupisce quindi che le manifestazioni più appariscenti dell'azione degli ormoni nelle piante investano soprattutto crescita e differenziamento, nonché aspetti metabolici a essi strettamente collegati, come sintesi e demolizione delle riserve.
Nelle piante, d'altra parte, crescita e differenziamento risentono della presenza di due strutture caratteristiche, assenti negli animali, e assolutamente determinanti ai fini della morfologia e della funzione della cellula vegetale: il vacuolo e la parete cellulare. Il vacuolo, metabolicamente quasi inerte ma sede di concentrazioni elevate di soluti, tende, assorbendo acqua dall'ambiente, a espandersi, spingendo verso l'esterno il citoplasma; questa spinta trova un limite nella resistenza della parete cellulare, involucro relativamente rigido, disposto subito all'esterno della membrana cellulare lipoproteica, o plasmalemma, comune a tutte le cellule animali o vegetali. La crescita degli organismi vegetali è affidata, oltre che all'aumento del numero delle cellule per moltiplicazione cellulare, a un processo caratteristico definito ‛crescita per distensione', nel quale la parete si lascia distendere in modo fisiologicamente controllato dalla forza dovuta alla differenza tra valori di pressione osmotica all'interno e all'esterno della membrana cellulare. Inoltre, per tutto il periodo giovanile della cellula, il metabolismo della parete (composta da cellulosa e altri polisaccaridi, nonché vari altri polimeri e proteine) presenta caratteristiche diverse sia nel tempo, sia da tessuto a tessuto. Queste diverse caratteristiche incidono in modo determinante su forma, struttura e funzione finale della cellula (pareti cutinizzate dell'epidermide, suberificate del sughero, lignificate dei vasi legnosi e delle fibre, perforate dei tubi cribrosi del floema; pareti con forma e dimensioni caratteristiche per ognuno dei diversi tessuti parenchimatici). In conseguenza di questa preminenza morfologico-funzionale della parete, l'azione degli ormoni nelle piante, pur investendo aspetti intimi del metabolismo cellulare analoghi a quelli primariamente toccati negli animali, si esplica molto spesso nel modo più evidente a livello dell'accrescimento e del differenziamento della parete cellulare.
Le classi di ormoni vegetali meglio note che, a quanto risulta, sono presenti in tutte le piante superiori, sono, tra gli ormoni a effetto prevalentemente (anche se non esclusivamente) stimolante, le auxine, le gibberelline e le citochinine; tra gli ormoni prevalentemente inibenti, l'etilene e l'acido abscissico. Altri fattori di regolazione che probabilmente rientrano nella definizione di ormone sono stati descritti in specie diverse, ma sembrano rivestire importanza molto meno generale di quelli sopra elencati. Vi sono inoltre buone ragioni per credere nell'esistenza di altri tipi di ormoni ad azione generale, per esempio quelli coinvolti nel controllo della fioritura (ormoni florigeni) o in altri processi specifici. Per questi ipotetici ormoni, tuttavia, manca l'individuazione della struttura chimica e quindi molto spesso si è portati a pensare che gli effetti a loro attribuiti possano dipendere dall'interazione tra gli ormoni già noti.
4. Auxine.
a) Definizione, aspetti biochimici, biosintesi.
La dimostrazione che una sostanza chimica specifica (auxina) prodotta nel meristema apicale del giovane coleottile di Graminacee diffonde nei tessuti sottostanti stimolandovi il processo di crescita per distensione della parete (distensione cellulare) è generalmente attribuita a P. Boysen Jensen e risale al 1910 (v. fig. 1). L'ormone naturale è identificabile nell'acido indol-3-acetico: questa sostanza è stata inizialmente scoperta e studiata chimicamente nell'urina dell'uomo, ove rappresenta un prodotto della flora batterica intestinale, e nei liquidi di colture di funghi e di batteri, e solo successivamente (1934-1966) nelle piante superiori. L'acido indol-3-acetico è oggi considerato come la principale - forse l'unica - auxina naturale diffusa in tutte le piante superiori, anche se diversi suoi derivati o precursori metabolicamente vicini possono esser presenti nelle piante e rivelarsi capaci di azione di tipo auxinico (probabilmente previa trasformazione in acido indolacetico). D'altra parte, intense ricerche circa il rapporto tra struttura e azione dell'auxina hanno portato alla scoperta di una grande varietà di composti sintetici dotati dell'azione caratteristica dell'auxina naturale. Questi, pur essendo apparentemente molto diversi per struttura, presentano alcuni aspetti in comune, sufficientemente ben definiti per permettere di prevedere un'azione ‛auxinica' in base al semplice esame della formula. Le formule di alcune auxine naturali e sintetiche sono riportate nella pagina seguente.
I principali caratteri comuni sono: 1) una struttura ad anello con una catena laterale inserita in prossimità di un doppio legame; 2) la presenza di una carica negativa, portata da un carbossile, al termine della catena laterale, con distanza ottimale di due atomi di carbonio dalla parte ciclica della molecola; 3) una configurazione spaziale tale da assicurare una distanza di circa 5,5 Å tra la carica negativa del gruppo carbossilico e una carica positiva della parte ciclica della molecola.
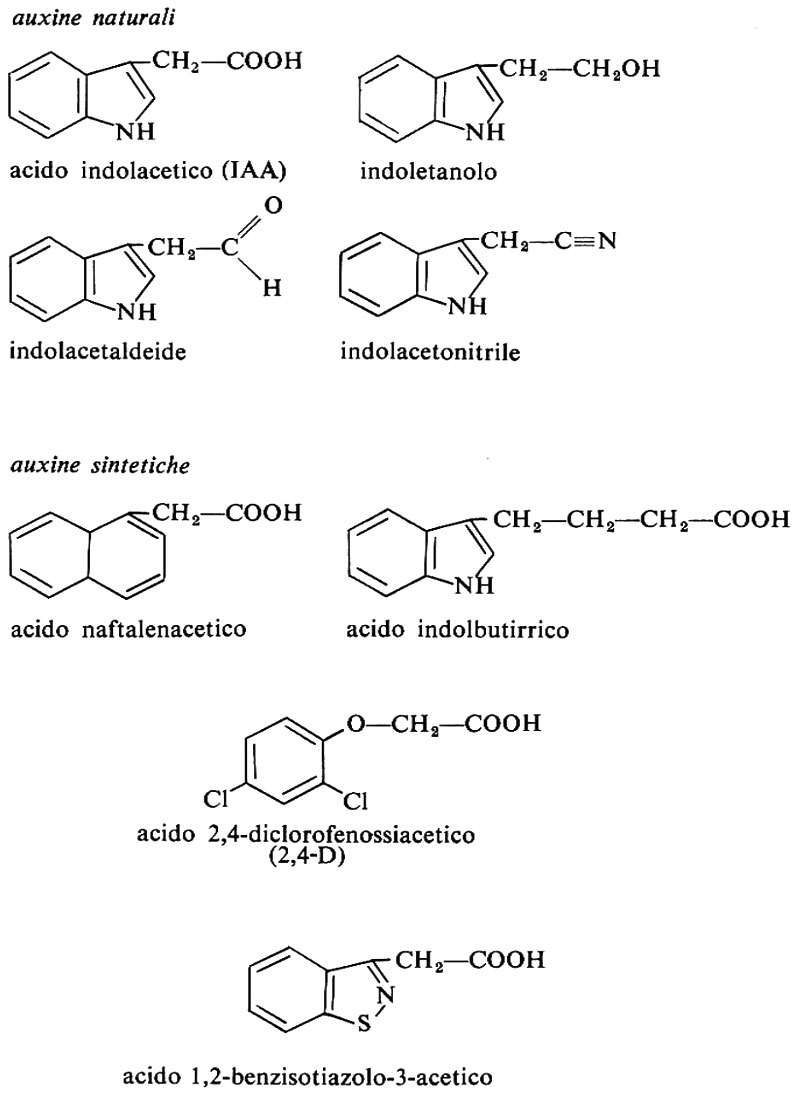
L'auxina è prodotta, nella pianta, in regioni ben definite, identificabili soprattutto nei meristemi in attività: sia in quelli apicali del fusto e (in misura minore) della radice, sia in quelli cambiali e in quelli del fiore, del seme e del frutto in stadio giovanile. Una notevole sintesi di auxine si svolge anche tra i tessuti adulti, soprattutto in quelli della lamina fogliare. La biosintesi dell'acido indolacetico, pur sottoposta a intensa indagine, non è ancora esaurientemente chiarita, anche se la maggioranza degli autori accetta l'ipotesi che essa passi attraverso il triptofano, avendo come composti intermedi alternativi l'acido indolpiruvico (per deamminazione ossidativa) o la triptammina (per decarbossilazione) e, come precursore immediato, l'indolacetaldeide (v. schema 1).
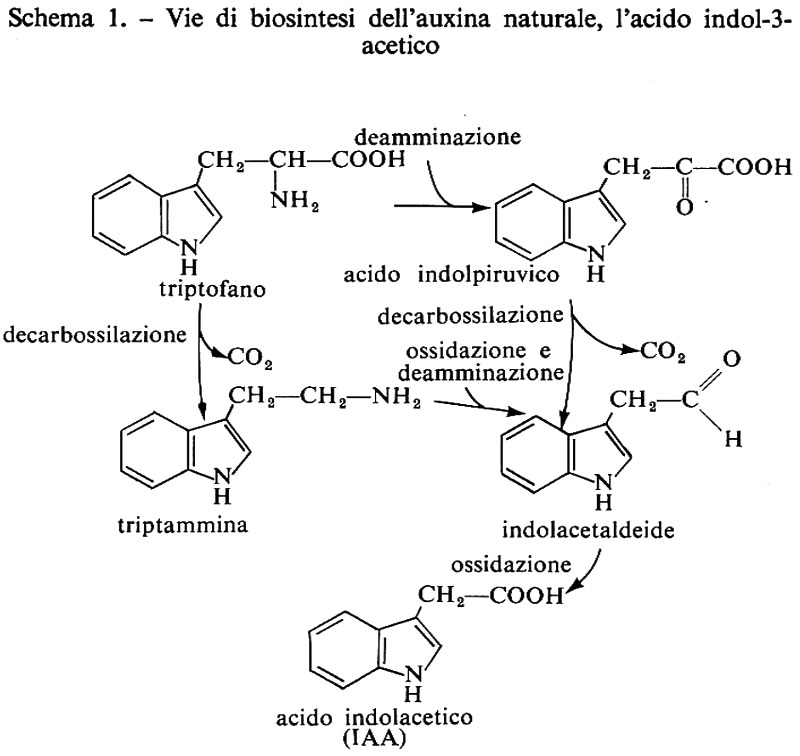
L'inattivazione fisiologica reversibile dell'auxina può avvenire per coniugazione con diversi composti, in particolare amminoacidi come l'acido aspartico (con formazione di acido indolacetilaspartico, presumibilmente inattivo).
La principale via di distruzione dell'acido indolacetico nei tessuti risulta quella ossidativa, mediata da un enzima specifico del tipo delle perossidasi (IAA-perossidasi), con 3-metilen-ossindolo e indol-3-acetaldeide come primi prodotti.
Diverse sostanze fotodinamicamente attive presenti nelle piante possono intervenire nella fotossidazione dell'auxina, mediante meccanismi diversi; l'importanza fisiologica di queste vie di demolizione rimane peraltro incerta.
Il trasporto delle auxine nella pianta è tipicamente polare, risultando diretto, nel fusto, nel senso apice-base, con una velocità dell'ordine di 10 mm/ora a 25 °C; nelle radici esso è rivolto verso la regione apicale, almeno nelle parti più adulte, mentre sembra accertato che, negli apici radicali, l'auxina formata nei meristemi apicali tenda a muoversi per un tratto relativamente breve verso la retrostante regione in crescita per distensione, fino alla zona pilifera. D'altra parte, dati recenti indicano che nella pianta intera il senso del flusso dell'auxina (come di altri ormoni) in una data regione non è costante, bensì controllato dalla situazione fisiologica delle diverse parti dell'organismo.
b) Effetti fisiologici.
Gli effetti fisiologici dell'auxina sono straordinariamente vari, e investono crescita per moltiplicazione, crescita per distensione, differenziamento della parete cellulare, tropismi, trasporto e assorbimento dei soluti, abscissione e invecchiamento. Questi effetti mostrano una specificità di organo, nel senso che in un dato momento dello sviluppo solo determinati organi danno una determinata risposta all'auxina. Il senso dell'azione, se essa risulti cioè nello stimolo oppure nell'inibizione di un dato fenomeno, può essere diverso per organi-bersaglio diversi, e anche diverso per uno stesso organo, a seconda della concentrazione dell'auxina. Ci limitiamo qui a riferire pochi esempi dell'estrema varietà dell'azione di questo ormone.
Crescita meristematica. - Comprende la crescita di frutti e meristemi cambiali, e la formazione delle radici laterali. In piante intere, organi come l'ovaio non fecondato possono essere stimolati dall'auxina alla moltiplicazione cellulare fino a dare sviluppo di frutti privi di semi (partenocarpia; v. fig. 2); pare inoltre accertato che nel caso più normale dello sviluppo del frutto dopo la fecondazione, l'auxina portata dal polline sia responsabile dello stimolo primario dei tessuti dell'ovaio alla moltiplicazione cellulare. Anche la moltiplicazione cellulare dei meristemi secondari è stimolata da concentrazioni relativamente basse di auxina. Infine, tessuti isolati dai più diversi organi e portati in condizioni nutritive favorevoli (colture in vitro) esigono di regola per entrare in moltiplicazione cellulare l'aggiunta di auxina al mezzo (v. fig. 3); solo in condizioni particolari (tessuti ‛abituati' o tessuti cancerogeni) le cellule diventano capaci di produrre auxina esse stesse e cessano di esigerne la somministrazione dall'esterno. E da notare che in molti tessuti lo stimolo alla moltiplicazione richiede anche altri ormoni, in particolare citochinine.
Altro effetto caratteristico dell'auxina, legato allo stimolo della moltiplicazione cellulare, è quello dell'induzione della formazione di radici avventizie (v. fig. 4); il trattamento della parete basale di segmenti di fusto (talee) con dosi basse di auxine per stimolare la radicazione è largamente applicato in agricoltura.
Crescita per distensione. - Nelle piante superiori la crescita per distensione cellulare, anche se di regola accompagnata a più o meno intensa sintesi di proteine, consiste essenzialmente in una distensione plastica della parete sotto la spinta della differenza tra pressione osmotica interna ed esterna; in altri termini, in un'assunzione netta di acqua condizionata da un particolare stato chimico-fisico della parete. Le auxine sono implicate in modo molto appariscente nel controllo delle proprietà della parete, e quindi del fenomeno di distensione. Parti giovani (ancora in crescita per distensione) di fusto di molte piante (pisello, fagiolo, ecc.), se isolate e incubate in acqua, pura o addizionata di semplici sostanze nutritizie, cessano di crescere; ma la crescita riprende attivamente per aggiunta di concentrazioni anche molto basse di auxine. Lo stesso avviene per sezioni di coleottile delle Monocotiledoni. Anche nelle radici la crescita per distensione della zona tra apice meristematico e zona dei peli assorbenti è spesso stimolata da concentrazioni estremamente basse (dell'ordine di 10-9 M) di auxina. Nelle radici, tuttavia, concentrazioni appena più alte mibiscono energicamente il processo di distensione. In realtà questa differenza tra fusto e radice è piuttosto quantitativa che qualitativa, in quanto dosi relativamente molto alte di auxina inibiscono la distensione anche dei tessuti del fusto; solamente, la concentrazione limite tra azione stimolante e azione inibente è spostata, per il fusto, a valori di 102-104 volte più alti (v. fig. 5). Su questa tipica capacità inibente dell'auxina ad alte concentrazioni si tornerà più avanti, parlando dell'etilene.
L'effetto dell'auxina sulla distensione cellulare è pure ben evidente in tessuti coltivati in vitro, e anche in cellule isolate in coltura, esprimendosi, anche qui, come aumento del volume cellulare per assunzione d'acqua nel vacuolo e per distensione della parete. Lo stimolo alla distensione è in questi casi accompagnato di regola da quello, sopra menzionato, alla crescita per moltiplicazione cellulare. In alcuni casi i due effetti possono essere scissi agendo sulle condizioni sperimentali: per esempio, nel midollo di tabacco isolato, l'auxina, se somministrata da sola, induce distensione senza moltiplicazione, mentre l'aggiunta di auxina insieme a citochinina (v. sotto) scatena una rapida moltiplicazione.
Differenziamento. - Sia la moltiplicazione sia la distensione cellulare portano, in quanto diversamente stimolate dall'auxina in parti diverse della pianta, a un differenziamento morfologico dell'organismo vegetale. Interventi più specifici delle auxine nel differenziamento macroscopico si riconoscono nelle curvature tipicamente indotte nelle piante da fattori fisici come la luce (fototropismi) e la gravità (geotropismi). Queste curvature risultano infatti largamente dipendenti dalla capacità di tali stimoli fisici di determinare un'ineguale distribuzione trasversale dell'auxina, e quindi un'ineguale crescita, nei due lati dell'organo diversamente colpiti dallo stimolo luminoso o gravitazionale (v. fig. 6). È inoltre ben dimostrato l'intervento delle auxine in processi di differenziamento a livello cellulare, in particolare per quanto riguarda la struttura chimica e la morfologia della parete: in cellule isolate in coltura la lignificazione della parete richiede auxina (in rapporto preciso con altri ormoni, come citochinine e gibberelline). Inoltre, in tessuti in coltura proliferanti (calli) la somministrazione di auxina a dosi relativamente alte (dell'ordine di 10-5 M), soprattutto se eseguita in modo da creare gradienti di concentrazione in determinate zone, induce il differenziamento di elementi lignificati del tipo delle tracheidi, e talora di veri abbozzi di fasci vascolari nel territorio sottostante. Questi dati suggeriscono che le auxine prodotte dalle gemme in via di sviluppo e dalle giovani foglie siano molto importanti nell'indurre il differenziamento, nel fusto, dei cordoni vascolari che assi- cureranno gli scambi tra organi come le foglie, i fiori, i frutti e il resto della pianta (cordoni vascolari).
Sviluppo delle gemme laterali e dominanza apicale. - La forma delle piante (per es. l'aspetto generale a cono o a colonna di molte piante legnose) è largamente determinata dal fenomeno della cosiddetta dominanza apicale, che può riassumersi dicendo che la gemma apicale in via di sviluppo inibisce, per un tratto sottostante variamente esteso, lo sviluppo delle gemme laterali. Fenomeno molto simile a questo è l'inibizione della crescita della gemma sita all'ascella della foglia stessa da parte della foglia funzionante. In entrambi i casi l'arresto dello sviluppo delle gemme laterali o ascellari sembra essenzialmente determinato dalla produzione dell'auxina nella gemma apicale (in crescita) o nel lembo fogliare, e risulta riproducibile sostituendo questi organi con preparati auxinici. Altri ormoni, in particolare le citochinine, agiscono antagonisticamente all'auxina sullo sviluppo delle gemme, esempio tipico di processo dipendente da un complesso rapporto tra le concentrazioni di ormoni diversi (v. fig. 7).
Trasporto e accumulo di soluti. - Qualunque effetto di stimolo sulla crescita presume una buona capacità della cellula di assumere sostanze nutritizie dall'esterno, capacità chiaramente legata a processi metabolici sottoposti a regolazione fisiologica. Non stupisce quindi che l'auxina intervenga, oltre che sulla crescita, anche sul trasporto dei soluti nella pianta e sulla capacità di assorbirli da parte delle cellule stimolate a crescere. Organi e tessuti in fase di rapida crescita, come i meristemi apicali e quelli del giovane boccio fiorale, del seme e del frutto in via di sviluppo, risultano caratteristicamente capaci di richiamare a sé e nei tessuti a essi prossimi sostanze nutritizie come glucidi, fosfati e altri sali.
In questa loro azione di richiamo di soluti i centri in via di sviluppo possono essere completamente sostituiti, dopo essere stati rimossi, dall'applicazione di preparati auxinici. È interessante osservare che quest'azione dell'auxina sembra comprendere due opposte componenti: una di richiamo (aumento di capacità di assunzione) nei territori prossimi al punto di applicazione dell'ormone, e una di mobilitazione (caduta della capacità di trattenere soluti) nei territori distali. Entrambe queste componenti sono evidentemente essenziali nel determinare il senso e la velocità del trasporto nella pianta di sostanze nutritizie e, presumibilmente, anche di fattori ormonali; entrambe, inoltre, devono essere l'espressione di un gioco complesso di fattori diversi di regolazione, di cui uno solo, anche se molto importante, è rappresentato dall'auxina (v. fig. 8).
Invecchiamento. - L'invecchiamento - complesso irreversibile di fenomeni degradativi che culmina nella morte - rappresenta, a livello di organi e tessuti, un processo normale fisiologicamente utilizzato dalla pianta durante il suo ciclo di sviluppo. Così certi tessuti vascolari, dopo un periodo di funzionalità, muoiono e vengono espulsi nella corteccia; le foglie e gli organi vessilliferi del fiore, dopo il periodo di tempo richiesto per l'esaurimento della loro funzione, determinato eventualmente dalle condizioni fisiologiche dell'intera pianta, vengono funzionalmente inattivati e scartati, dopo essere stati ‛costretti' a restituire al resto dell'organismo quanto possibile di materiali utili. Che il processo d'invecchiamento in queste parti della pianta sia chiaramente determinato da stimoli di tipo correlativo (in particolare, ormonale) piuttosto che da un inevitabile determinismo intrinseco, tale da porre un termine temporale alla sopravvivenza dell'organo, è chiaramente indicato dal fatto che: a) la soppressione dei centri in attivo sviluppo (meristemi, organi attivi dei giovani fiori e frutti) prolunga talora moltissimo la sopravvivenza degli organi sottoposti a invecchiamento; b) in organi isolati (come foglie intere o anche suddivise in frammenti) la velocità d'invecchiamento può essere grandemente accelerata o ritardata dalla presenza di ormoni nel mezzo di incubazione. È facile dimostrare che sia l'auxina sia, come si dirà più avanti, altri ormoni (soprattutto la cinetina) influenzano l'invecchiamento: in particolare l'auxina applicata in un dato territorio della pianta (per es. in sostituzione della gemma apicale) può accelerare l'invecchiamento di regioni od organi (per es. foglie) situati distalmente. D'altro lato, in tessuti posti in contatto diretto con l'auxina (per es. parti di foglia isolate) l'invecchiamento in genere non è accelerato, e in qualche materiale è addirittura ritardato.
Effetti sull'abscissione. - L'abscissione è il processo per cui in determinate condizioni fisiologiche alcuni organi come foglie, fiori o frutti possono indebolire le proprie connessioni meccaniche col resto della pianta, fino a distaccarsi in seguito a una minima trazione. La scissione avviene a livello di una zona ben definita, detta zona di abscissione (v. fig. 9). Zone di abscissione sono spesso ben riconoscibili alla base dei peduncoli dei fiori e dei frutti, e alla base o, talora, nel tratto intermedio dei piccioli delle foglie.
Il fenomeno dell'abscissione è imputabile alla lisi dei polisaccaridi (e altri polimeri) che tengono unite le cellule, come costituenti delle pareti cellulari, ed è spesso preceduto dall'attivazione della moltiplicazione cellulare in uno o pochi strati di cellule disposti trasversalmente rispetto all'asse dell'organo che abscide, in corrispondenza della zona di abscissione (formazione del cosiddetto meristema di abscissione). Sia la usi delle pareti cellulari sia l'attivazione del meristema di abscissione sono sotto stretto controllo di diversi ormoni vegetali. Questo è facilmente comprensibile, in quanto l'abscissione di un organo rappresenta l'aspetto terminale del suo invecchiamento e deve avvenire in un momento ben preciso del suo ciclo di sviluppo (per es.: per il frutto, quando la maturazione è compiuta ; per i fiori non fecondati e per le foglie, quando hanno esaurita la loro attività funzionale e hanno restituito alla pianta quanto possibile dei prodotti utili in essi contenuti).
L'auxina risulta, di regola, un efficace agente inibitore dell'abscissione. I peduncoli di foglie il cui lembo - in cui è forte la produzione di auxina - sia stato reciso, o anche semplicemente impedito nella sua capacità fotosintetica (per es. fasciandolo con carta nera) abscidono entro un breve penodo di tempo; l'applicazione di auxina all'estremo distale del picciolo ne impedisce efficacemente l'abscissione. Non mancano tuttavia casi in cui la somministrazione di auxina può accelerare, anziché inibire, l'abscissione; tanto che la somministrazione di auxine sintetiche è comunemente usata in agricoltura per ridurre il numero dei fiori o dei giovani frutti quando risulta eccessivo. In questi casi l'azione di stimolo dell'auxina, peraltro, è probabilmente indiretta, e può dipendere sia dalla capacità di questo ormone, quando presente ad alte concentrazioni, d'indurre la biosintesi di etilene (agente stimolante l'abscissione, v. sotto, cap. 5), sia dall'attivazione, da parte dell'ormone spruzzato indiscriminatamente sull'intera pianta, di altre regioni, diverse dall'organo suscettibile di abscissione.
5. Etilene.
a) Aspetti biochimici, biosintesi.
Il trattare l'etilene subito dopo le auxine è giustificato dal fatto che la sintesi di questo composto è fortemente stimolata dalla presenza dell'auxina stessa, il che fa sì che i due ormoni siano molto spesso simultaneamente presenti negli stessi tessuti. Inoltre, poiché gli effetti dell'etilene sono spesso parzialmente antagonisti a quelli dell'auxina, i due ormoni vengono a costituire un unico complesso sistema di regolazione ormonale.
L'etilene è oggi generalmente incluso tra gli ormoni vegetali, malgrado qualche perplessità dovuta soprattutto al fatto che, alle temperature fisiologiche, esso si trova allo stato gassoso. In realtà, nelle cellule l'etilene è presente in soluzione e in tale stato può muoversi nella pianta, per diffusione o trasportato con i liquidi, per distanze brevi ma tuttavia apprezzabili, spostandosi così dai tessuti dove è prodotto (come gli apici meristematici) verso i tessuti vicini.
La capacità delle piante di produrre etilene e anche l'attività dell'etilene nel determinare curvature anormali dei giovani fusti e dei piccioli fogliari, come pure di accelerare la fase finale della maturazione in molti frutti, sono conosciute da oltre cinquant'anni. Tuttavia, solo dopo il 1962 studi di diversi autori, soprattutto di S. P. Burg e di F. B. Abeles, hanno messo in luce la grande importanza di questo composto nel regolare tutta una serie di processi fisiologici nella pianta.
L'etilene è il più semplice idrocarburo insaturo, con formula CH2=CH2. Altri idrocarburi insaturi (propilene, ecc.) risultano dotati di effetti fisiologici simili a quelli dell'etilene, anche se più lievi, e decrescenti con l'aumentare della lunghezza della catena carboniosa. Anche l'ossido di carbonio a bassa concentrazione può determinare effetti che ricordano quelli dell'etilene: fatto che, essendo riconducibile ad analogie strutturali tra i due composti, renderebbe ipotizzabile un meccanismo d'azione basato sull'interazione con enzimi metalloproteici. Gli effetti fisiologici dell'etilene sono impediti, o ridotti, dalla presenza di CO2 ad alte concentrazioni, il che suggerisce una competizione tra etilene e CO2 a livello del recettore primario.
Nelle piante l'etilene è prodotto da tessuti molto diversi, come i meristemi degli apici vegetativi, l'embrione dei semi germinanti, i tessuti adulti, soprattutto se sottoposti a lesione, e diversi organi al termine del loro ciclo di sviluppo, quali, in particolare, i frutti maturi e le foglie invecchiate nel periodo che precede l'abscissione.
La via biochimica di sintesi dell'etilene non è nota nei suoi dettagli, anche se sembra accertato che essa parta dalla metionina e passi poi attraverso il metionale e la demolizione ossidativa di questo da parte di enzimi del tipo delle perossidasi (v. schema 2). Un aspetto molto interessante, anche se soprattutto agli effetti fisiologici, della biosintesi dell'etilene è che essa risulta strettamente controllata dal livello in auxina, nel senso che un aumento di questa oltre un certo valore (diverso per i diversi tessuti) porta con sé un forte aumento della produzione di etilene (v. fig. 10). Anche altri ormoni capaci di stimolare la crescita, come le citochinine e le gibberelline, sembrano capaci di stimolare la sintesi di etilene, pur se in misura molto minore rispetto all'auxina.
b) Effetti fisiologici.
L'etilene influenza la crescita per moltiplicazione cellulare, la crescita per distensione, le curvature dei fusti, lo sviluppo delle gemme, la germinazione dei semi, l'abscissione, l'invecchiamento dei tessuti fogliari, la formazione di radici laterali, la maturazione dei frutti.
In un tentativo di discussione sintetica di questi effetti, occorre tener conto del fatto che molto spesso l'etilene agisce in senso opposto, o quanto meno diverso, da quello in cui agisce l'auxina, con la quale normalmente coesiste, anche in conseguenza della sopracitata relazione tra presenza di auxina e sintesi di etilene. Così, nei tessuti in cui la somministrazione di auxina stimola la crescita per moltiplicazione cellulare (caso dei tessuti in coltura, della formazione di radici avventizie nelle talee, dello sviluppo a frutto partenocarpico di ovai non fecondati) oppure la crescita per distensione (segmenti isolati di giovani fusti o coleottili), l'etilene agisce in senso antagonista all'auxina, inibendo più o meno fortemente l'uno e l'altro tipo di crescita. Effetti antagonistici a quelli dell'auxina si hanno anche per l'abscissione e il risveglio delle gemme dormienti laterali o ascellari (dominanza apicale, v. sopra), fenomeni fortemente inibiti dall'auxina e invece stimolati dall'etilene. Questo antagonismo tra effetti dell'auxina e dell'etilene, insieme al fatto che l'auxina al di sopra di una certa concentrazione stimola la sintesi di etilene, suggerisce l'ipotesi che molti, forse tutti quegli effetti di tipo inibente esplicati dall'auxina a concentrazioni relativamente elevate siano, almeno in parte, imputabili all'induzione da auxina di un aumento della produzione di etilene, ipotesi che spiegherebbe tra l'altro, per esempio, l'inibizione della crescita per distensione in radici trattate con auxina. Il sistema auxina-etilene sembra quindi presentare un interessante carattere di autoregolabilità, in quanto ogni aumento dell'ormone stimolante trascinerebbe con sé la sintesi dell'antagonista inibente.
È interessante ricordare che l'interazione auxina-etilene sembra ricomparire anche a livello applicativo, in quanto può in parte spiegare alcuni degli effetti di quella classe di erbicidi di tipo cosiddetto auxinico, come il 2,4-D (acido 2,4-diclorofenossiacetico). Questi, infatti, somministrati a dosi alte, inducono sintesi di etilene a concentrazioni sufficienti per esplicare un'azione fortemente inibente.
L'antagonismo tra auxine ed etilene è del resto soltanto parziale e in molti casi non sembra sufficiente a spiegare l'azione dell'etilene. Possiamo ricordare, in proposito, l'intensa azione dell'etilene nell'indurre o nell'accelerare la maturazione del frutto (espressa dall'aumento della respirazione, dalla scomparsa di clorofilla, dalla comparsa di pigmenti colorati e dall'aumento di zuccheri solubili); lo stimolo alla germinazione di molti semi; l'induzione alla fioritura in alcune specie come l'ananas; lo stimolo della crescita per distensione in senso trasversale (anziché longitudinale, come indotto dall'auxina).
Dati recenti dimostrano, inoltre, che l'etilene interagisce fisiologicamente non solo, come si è detto, con l'auxina, ma anche con tutti gli altri ormoni vegetali noti. Tali interazioni coinvolgono sia gli effetti finali di questi ormoni, sia il loro stesso trasporto, e probabilmente anche le relative biosintesi. In particolare, la capacità dell'etilene di inibire il trasporto (soprattutto in senso trasversale) dell'auxina sembra una componente importante del meccanismo mediante il quale il sistema etilene-auxina controlla i fenomeni di orientamento fisiologico della pianta nello spazio (tropismi) oppure, quando gli ormoni sono somministrati dall'esterno, induce curvature anormali. E infine da ricordare che sia la sintesi di etilene che la sensibilità dei diversi tessuti a questo ormone sono fortemente influenzati dalla luce a quelle stesse lunghezze d'onda (rosso e rosso porpora) responsabili del controllo fotoperiodico della riproduzione nelle piante (fotoperiodismo). Molte piante dicotiledoni crescono, subito dopo germinato il seme, con l'estremità ripiegata a uncino, (proteggendo, così, il delicato apice vegetativo dagli attriti con il terreno) e questo uncino si raddrizza appena la piantina emerge dal suolo e riceve la luce; questo comportamento viene spiegato attribuendo la primitiva curvatura della parte apicale all'etilene (abbondantemente prodotto al buio) e il suo raddrizzarsi all'azione della componente rossa della luce, che riduce sia la sintesi dell'etilene che la sensibilità dei tessuti all'ormone.
6. Gibberelline.
a) Definizione, aspetti biochimici, biosintesi.
Malgrado siano anch'esse definibili come ormoni di crescita, le gibberelline rappresentano una famiglia ben distinta da quella delle auxine. Sia auxine che gibberelline possono stimolare la crescita per moltiplicazione cellulare e quella per distensione, anche se in genere tessuti diversi rispondono in modo diverso all'uno e all'altro tipo di ormoni. Le gibberelline non mostrano, o mostrano in grado molto ridotto, la caratteristica azione inibente tipica delle auxine quando siano presenti a concentrazioni relativamente elevate: tale azione, come si è detto sopra, è, almeno in larga misura, riconducibile allo stimolo della sintesi di etilene, mentre questo effetto non è indotto, o è indotto in misura molto modesta, dalle gibberelline. Infine le gibberelline costituiscono, dal punto di vista della struttura chimica, un gruppo omogeneo e ben definito, nettamente diverso da quello che comprende l'auxina naturale e le au- xine sintetiche.
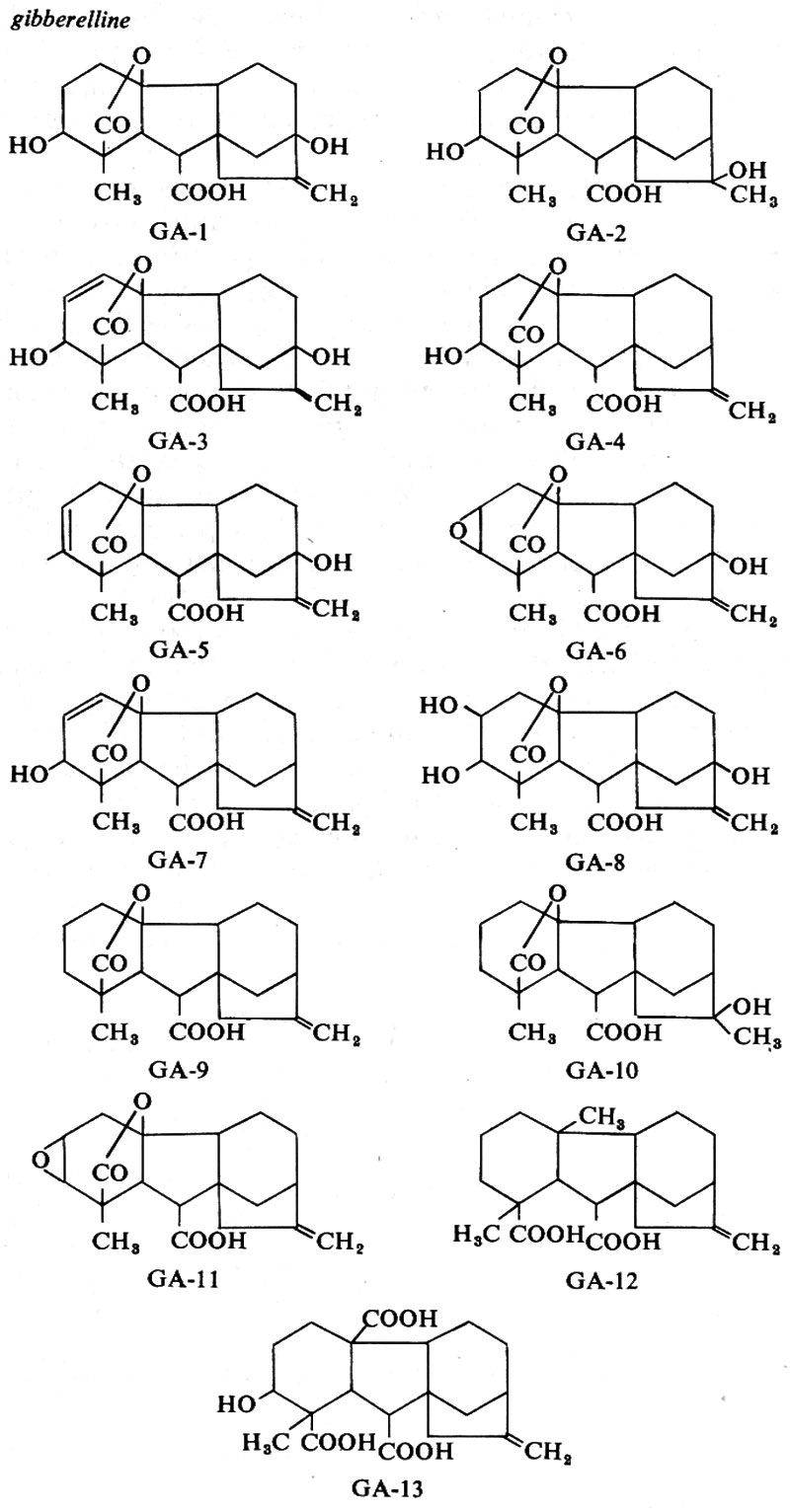
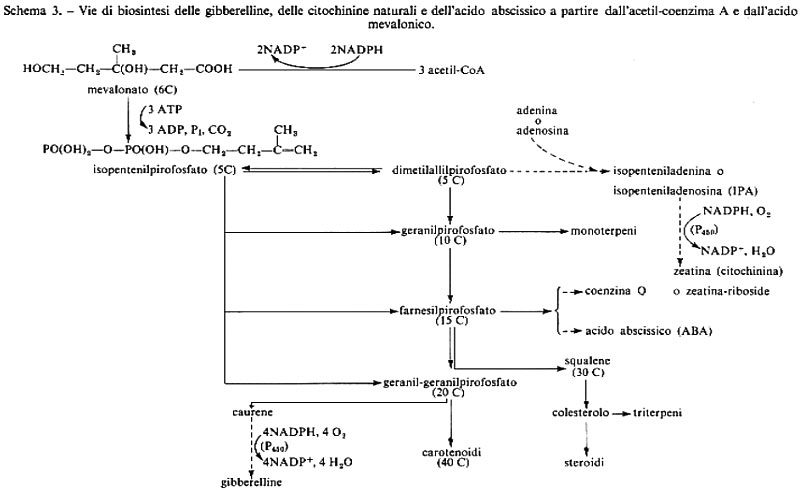
Le gibberelline sono state scoperte nel 1930 da T. Yabuta e F. Hayashi, che individuarono in un composto di questa classe il principio attivo prodotto da un fungo parassita, la Gibberella fujikuroi, capace di determinare nelle piante di riso infetto un allungamento abnorme dello stelo fruttifero. Successive pazienti ricerche portarono alla conclusione che composti dello stesso tipo erano prodotti da tutte le piante superiori e vi esplicavano un'importantissima azione di regolazione di numerosi processi fisiologici, oltre che della crescita.
Delle molte (oltre 30) gibberelline note, diverse tra loro per particolari strutturali di modesto rilievo e probabilmente metabolicamente interconvertibili, la meglio nota è la gibberellina A3 o acido gibberellico o GA-3 (v. le formule riportate nella pagina precedente.
Delle altre gibberelline, alcune sono da considerarsi precursori o derivati inattivi dell'acido gibberellico, mentre per altre resta aperto il problema di una possibile diversa specificità di azione.
La via di biosintesi dell'acido gibberellico (e delle altre gibberelline) parte dall'acetato attivo e passa per il mevalonato, il farnesilpirofosfato e il caurene (v. biosintesi), e comprende una serie di ossidazioni miste catalizzate da enzimi apparentati alla classe dei citocromi (v. schema 3). Nella pianta, organi molto diversi risultano capaci di sintetizzare gibberelline: gli apici del fusto e delle radici, gli organi riproduttivi fiorali, l'embrione nel seme sia in via di sviluppo che in fase di germinazione. Il trasporto delle gibberelline nella pianta non è prevalentemente polare, in senso apice-base, come quello delle auxine, e mostra piuttosto una tendenza inversa, dalle radici verso le parti superiori del fusto e verso fiori e frutti. La sintesi di gibberelline è fortemente influenzata dalla luce, in particolare da quella attiva sui fenomeni fotoperiodici; l'azione della luce rossa sulla germinazione dei semi, sull'allungamento degli internodi e, in qualche caso, sulla stessa fioritura sembra almeno in parte riconducibile a uno stimolo della sintesi o del trasporto di gibberelline, anche se la natura di questo rapporto è tuttora oscura.
b) Effetti fisiologici.
L'acido gibberellico (in genere meglio se associato a citochinina e auxina) stimola la moltiplicazione cellulare sia nei tessuti in coltura (per es. nel parenchima midollare di tabacco) sia nella parte giovane del fusto di piante intere: effetto, questo, particolarmente evidente nei mutanti nani, geneticamente ben identificati, del pisello e del mais (v. fig. 11).
L'effetto di stimolo alla moltiplicazione è in genere accompagnato, e spesso preceduto, da un notevole stimolo alla distensione cellulare. In alcuni casi (parenchina midollare di Helianthus tuberosus incubato in presenza di auxina e citochinine) l'acido gibberellico inibisce la moltiplicazione e stimola la distensione cellulare. Nelle piante a rosetta, che sviluppano un unico lungo stelo fiorifero solo quando entrano in riproduzione, la somministrazione di gibberelline induce la formazione dello stelo anche in condizioni di fotoperiodo sfavorevoli alla fioritura. È evidente che in questi casi l'ormone ha agito stimolando, tra l'altro, la crescita per divisione del meristema da cui si sviluppa lo stelo fiorale (v. fig. 12).
Resta d'altra parte del tutto aperto il problema di quanto, in fenomeni di attivazione della crescita così complessi, sia possibile discriminare l'azione specifica dei diversi ormoni. Infatti, un risveglio, a opera della gibberellina, della crescita meristematica del tessuto, e quindi del numero delle cellule in divisione, potrebbe molto verosimilmente portare all'aumento della produzione o del richiamo di auxina e altri ormoni, con tutte le conseguenze che ne derivano.
Strettamente collegata all'induzione da gibberellina dello sviluppo dello scapo fiorale nelle piante a rosetta sembra essere la capacità dell'ormone d'indurre la fioritura in queste come in altre piante (per es. Conifere dei generi Taxus e Cupressus), sempre in condizioni fotoperiodiche sfavorevoli. È stata avanzata l'ipotesi che in questi casi il fattore limitante lo sviluppo del sistema riproduttivo consista nell'inerzia del relativo meristema (o abbozzo embrionale) e che la somministrazione di gibberellina agisca rompendo questa inerzia e avviando il processo.
La gibberellina induce pure la partenocarpia in certe specie (come nella vite, dove peraltro la partenocarpia è indotta anche da auxina e dalle citochinine) e stimola la distensione cellulare in foglie giovani (mais). Infine, una delle azioni caratteristiche e più specifiche delle gibberelline è la loro capacità di rompere la dormienza di molti semi, che altrimenti germinerebbero solo dopo un lungo periodo di riposo, oppure se sottoposti a determinati stimoli fisici (irradiazione con luce rossa). In questi casi, uno dei primi effetti osservabili è l'attivazione della sintesi di enzimi necessari per la trasformazione delle riserve immagazzinate nel seme in composti facilmente utilizzabili per la crescita della nuova piantina. Particolarmente ben studiato è l'effetto di stimolo delle gibberelline sulla sintesi dell'α-amilasi (enzima che idrolizza l'amido) nell'aleurone dei semi di Graminacee, effetto così preciso e sensibile da poter essere usato come metodo di dosaggio dell'ormone.
Insieme a quella dell'amilasi, peraltro, risulta attivata dalle gibberelline la sintesi di altri enzimi caratteristici della prima fase della germinazione, tra cui quelli interessati alla demolizione delle proteine (proteasi) e alla trasformazione dei lipidi di riserva in zuccheri (isocitratoliasi).
Il trattamento con freddo (5 °C seguito da 20 °C) in alcuni casi (per es. nei semi di nocciolo) aumenta il livello di gibberelline, il che può spiegare un aspetto dell'efficacia della vernalizzazione (trattamento dei semi dormienti con basse temperature, che spesso accelera l'entrata nella condizione di germinabilità; v. fotoperiodismo).
Anche processi come l'invecchiamento di parti isolate di foglie e l'abscissione possono, almeno in certi casi, essere influenzati dalle gibberelline, in modo tuttavia meno imponente che da auxina, citochinine ed etilene.
7. Citochinine.
a) Definizione, aspetti biochimici, biosintesi.
Le citochinine costituiscono una classe di ormoni vegetali ben caratterizzata per fisionomia sia chimica sia fisiologica, pur avendo in comune con auxina e gibberelline la capacità di stimolare la crescita per moltiplicazione cellulare e, talora, anche quella per distensione, nonché d'intervenire in molti fenomeni di differenziamento e nell'invecchiamento.
I primi studi sull'azione di sostanze di questo gruppo risalgono alle esperienze di F. Skoog e collaboratori (1954) sull'effetto delle basi azotate, e in particolare dell'adenina, sulla crescita e il differenziamento di tessuti vegetali (soprattutto midollo di tabacco) coltivati in vitro. Lo sviluppo di questi studi portò a riconoscere la spiccata attività (simile ma molto maggiore di quella dell'adenina) della quale è dotato un componente estratto da materiali animali (sperma di pesce), la furfuriladenina, tuttora usata in fisiologia vegetale e comunemente chiamata chinetina; successivamente, consentì l'isolamento da materiali diversi, tra cui l'estratto di lievito, di altri composti dotati della stessa azione della chinetina. Le formule di alcuni di questi composti sono riportate nella pagina seguente.
Studi successivi dimostrano che i composti ad azione chinetino-simile, o citochinine, avevano soprattutto origine dall'idrolisi dell'acido ribonucleico, specialmente di quello adibito alla funzione di trasporto degli amminoacidi (RNA di trasferimento o transfer RNA, tRNA). Particolarmente attivo come citochinina risultò un nucleotide anomalo, l'isopenteniladenosina, che si trova immediatamente adiacente ai tre nucleotidi dell'anticodone nei tRNA del lievito specifici per la tirosina e la senna (v. acidi nucleici).
Soltanto molto recentemente si è giunti all'individuazione di una citochinina, la zeatina, presente allo stato libero (cioè non inserita in un RNA) nel mais, e alla dimostrazione che questa sostanza, e altre a essa chimicamente molto simili, sono generalmente diffuse in tutte le piante superiori.
La formula della zeatina (la più attiva tra le citochinine note) è molto simile a quella della isopenteniladenosina ottenibile per idrolisi dai tRNA; il fatto che l'isopenteniladenosina è appena meno attiva della zeatina suggerisce l'ipotesi che nei tessuti la prima possa essere trasformata nella seconda (v. schema 3).
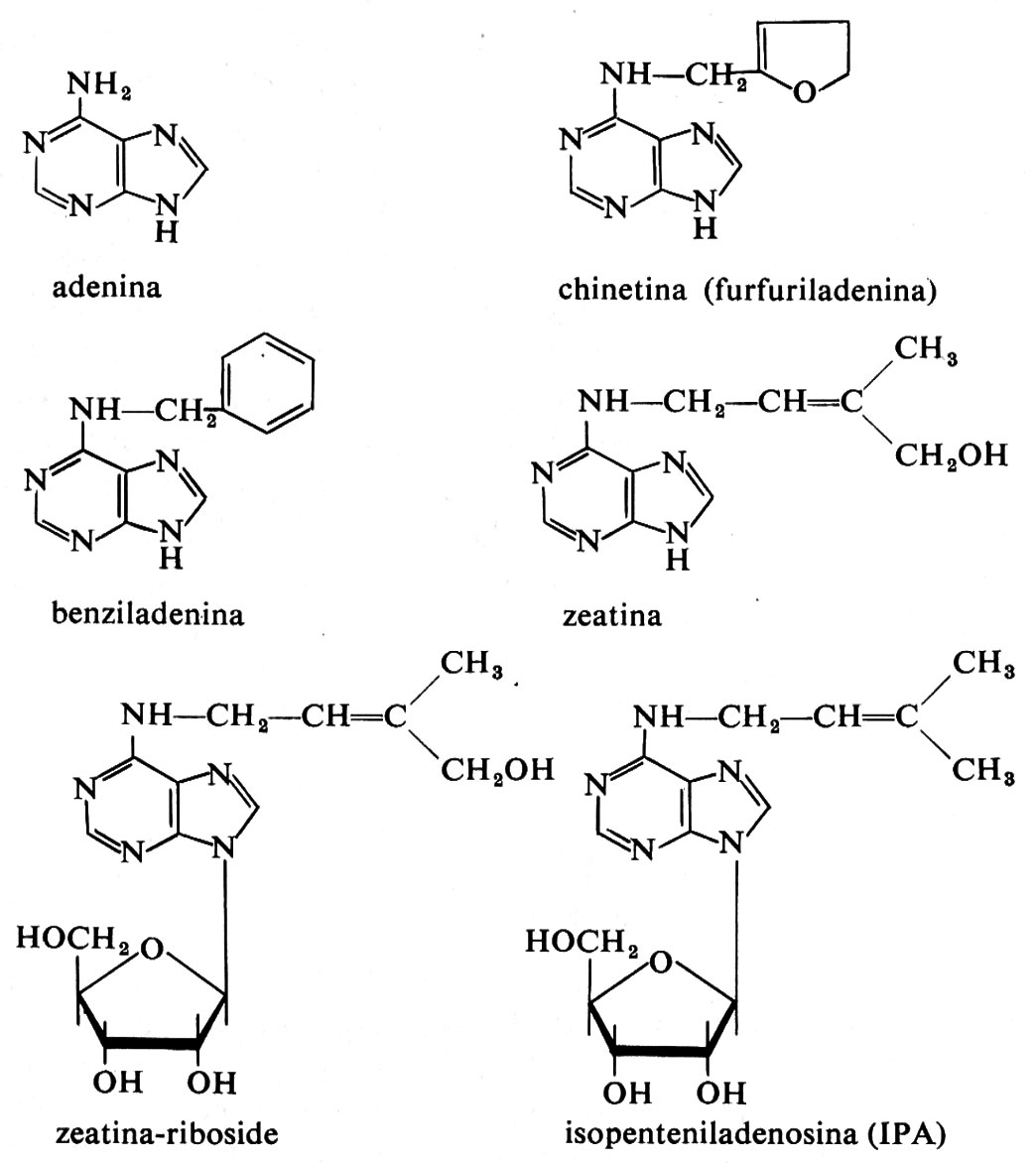
Dal punto di vista della struttura chimica le citochinine rappresentano un gruppo relativamente omogeneo, caratterizzato da una base eterociclica (in genere purinica) con una catena lipofila legata a un gruppo amminico. Le citochinine più attive sono derivati dell'adenina, o del suo nucleoside adenosina, o del suo nucleotide acido adenilico, in cui il gruppo amminico in 6 è legato a una catena carboniosa lipofila. L'attività, minima per l'adenina, aumenta coll'aumento del numero di atomi di carbonio della catena lipofila fino a quello caratteristico della isopenteniladenosina e della zeatina.
Una citochinina sintetica molto attiva è la benziladenina. Anche derivati di sostituzione dell'urea, come la difenilurea, rivelano una certa attività citochininica. Il fatto che l'isopenteniladenina e altre citochinine siano ottenibili per idrolisi di tRNA ha suggerito l'ipotesi che l'azione fisiologica di questi composti dipenda dalla loro capacità di essere incorporati nelle catene polinucleotidiche di questa classe di acidi nucleici, e quindi di regolare, attraverso il controllo del livello di alcuni tRNA, la capacità di sintesi proteica nelle cellule. Questa ipotesi è peraltro contraddetta da esperienze che dimostrano che alcuni tipi di citochinine, pure fisiologicamente molto attive, non vengono certamente incorporate nei tRNA.
Territori preferenziali di sintesi delle citochinine sembrano essere le radici, anche se la capacità di produrre questi ormoni appare largamente diffusa nella pianta. Il trasporto risulta preferenzialmente acropeto (almeno nella pianta intera e in base a dati relativamente scarsi) e affidato soprattutto al floema. La biosintesi delle citochinine naturali (zeatina e derivati) non è nota nei dettagli, anche se deve comprendere, secondo ogni evidenza, l'unione dell'isopentenilpirofosfato (derivato dal mevalonato) con adenina, o adenosina, o acido adenilico (v. schema 3). La demolizione nella pianta è presumibilmente affidata all'azione di enzimi del tipo delle adenosina-amminoidrolasi presenti nei tessuti animali.
b) Effetti fisiologici.
Gli effetti più caratteristici delle citochinine sono quelli di stimolo della moltiplicazione cellulare, ben riconoscibili sia in tessuti in coltura (parenchima midollare di tabacco e di Helianthus tuberosus, ‛calli' ottenuti da cotiledoni di molti semi) sia in sistemi rappresentati da parti isolate della pianta (parte immediatamente subapicale dei coleottili di Graminacee, fettine di tuberi di dalia, patata, begonia). In questi casi spesso l'azione delle citochinine sulla moltiplicazione cellulare è sinergica con quella dell'auxina e delle gibberelline. Pure riconducibile allo stimolo della moltiplicazione cellulare è la capacità delle citochinine di indurre sia la crescita delle gemme dormienti (v. fig. 7) sia lo sviluppo partenocarpico degli ovai non fecondati di diverse specie, e di stimolare, almeno in qualche caso (sezioni di radice di pisello), la crescita delle radici laterali.
Molto più modesti e più rari sono gli effetti delle citochinine sulla crescita per distensione; uno stimolo sensibile di questo tipo di crescita si osserva tuttavia sui cotiledoni di certi semi, come quelli di ravanello, e sulle sezioni dei giovani coleottili isolati delle Monocotiledoni.
Anche la germinazione di semi dormienti, o la cui germinazione sia stata inibita da trattamento con acido abscissico, può essere, in determinate condizioni (per es. in sinergismo con dosi anche molto basse di gibberelline), efficacemente stimolata dalle citochinine.
Il rapporto tra livelli in citochinine, auxina e altri ormoni è molto importante per quanto riguarda i fenomeni di differenziamento. Così, per es., in sezioni di radici di pisello un alto rapporto citochinine/auxina stimola la lignificazione delle cellule corticali (v. fig. 13), mentre nei ‛calli' in coltura la stessa condizione è favorevole al differenziamento di gemme vegetative e un eccesso di auxina tende a stimolare lo sviluppo delle radici (v. fig. 3). Anche nei protonemi dei muschi la citochinina induce la formazione di gemme.
Anche i fenomeni d'invecchiamento sono fortemente influenzati dalle citochinine. In un sistema classico per questo tipo di ricerche, costituito da parti isolate di lembo fogliare incubate in condizioni standard, l'azione di questi ormoni nell'impedire, o nel ritardare, la degradazione della clorofilla e la lisi degli acidi nucleici e delle proteine è così efficace da poter rappresentare un test della loro presenza: test peraltro non specifico, in quanto anche auxine e gibberelline possono esplicare effetti simili, pur se di solito molto minori, e limitatamente ad alcune specie (v. fig. 14). Un altro fenomeno riconducibile all'invecchiamento, l'abscissione dei peduncoli fogliari privati del lembo, tipicamente inibito dall'auxina, risulta pure inibito, almeno in alcuni materiali, dalle citochinine.
8. Acido abscissico e abscissine o dormine.
a) Definizione, aspetti biochimici, biosintesi.
La presenza di sostanze inibenti processi come la crescita e la germinazione dei semi negli estratti di tessuti vegetali è nota da tempo. Alcune di queste sostanze, come gli acidi caffeico e ferulico e diversi fenoli, sono state ben caratterizzate. Lo studio della loro azione non ne ha tuttavia dimostrato nè l'importanza generale nei fenomeni di inibizione, nè caratteristiche tali da poterle classificare nella categoria degli ormoni.
Rientra invece chiaramente in questa categoria l'acido abscissico, che è probabilmente presente in tutte le piante superiori, e che può considerarsi come il principale rappresentante di una classe di ormoni ad azione prevalentemente inibente i processi di accrescimento, e quindi in qualche modo opposta a quella delle auxine, gibberelline e citochinine. La scoperta dell'acido abscissico è dovuta soprattutto alle ricerche condotte indipendentemente dai gruppi di F. T. Addicott, interessato all'isolamento dei fattori responsabili dell'abscissione dei giovani frutti di cotone, e di P. F. Wareing, impegnato nell'analisi sistematica dei fattori stimolanti o inibenti rivelati dalla cromatografia di estratti di tessuti vegetali e, in particolare, di gemme dormienti. Nel periodo tra il 1973 e il 1976 sia Addicott che Wareing riuscirono a purificare, il primo dai peduncoli dei frutti in procinto di abscidere, il secondo da gemme dormienti di acero, uno stesso composto corrispondente al cosiddetto ‛inibitore β' rivelato dall'analisi cromatografica degli estratti di molti organi vegetali (v. fig. 15). La successiva analisi chimica dimostrò per questo composto la formula riportata qui di seguito.

Il nome ‛acido abscissico' dato al composto purificato e giustificato solo in parte dalla sua capacità di accelerare l'abscissione di piccioli di foglie private del lembo, in quanto in altri casi l'effetto sull'abscissione risulta scarso o addirittura nullo. Preferibile sarebbe il nome di ‛dormina' (e di ‛dormine' per altri eventuali ormoni della stessa classe) in quanto l'azione più caratteristica dell'acido abscissico è quella di mantenere la dormienza dei semi e delle gemme, e ‛dormine' venivano definiti, anche prima del loro isolamento e identificazione chimica, i fattori ritenuti responsabili di questo tipo di fenomeni.
L'acido abscissico è stato trovato in pressoché tutte le piante superiori sottoposte ad analisi accurata per svelarne la presenza, e risulta particolarmente abbondante, fino a concentrazioni massime dell'ordine del mg/kg di peso fresco, nei semi e nelle gemme dormienti. Luoghi preferenziali di sintesi (o per lo meno di accumulo) sembrano essere i tessuti in fase di passaggio dall'attiva crescita a uno stato di relativo riposo fisiologico, come i frutti e i semi in via di maturazione.
Anche le foglie pare siano sede di attiva produzione di questo ormone. Trattamenti che accelerano l'invecchiamento (per es., con etilene) o che in qualche modo ledono o diminuiscono l'attività dei tessuti (come lo stato di sofferenza per deficit idrico) risultano indurre, almeno in molti casi, un aumento del livello di acido abscissico.
L'esame della formula dell'acido abscissico mostra come esso sia riconducibile all'unione di tre unità isoprenoidi, derivate dal mevalonato; e infatti esperienze di somministrazione in vivo di precursori marcati dimostrano che la biosintesi dell'ormone parte dall'acido mevalonico e comprende come prodotti intermedi il farnesilpirofosfato (v. schema 3). È interessante a questo proposito sottolineare il fatto che ben tre dei cinque principali tipi di ormoni vegetali noti, e cioè le gibberelline, l'acido abscissico e (per la catena legata sul gruppo amminico dell'adenina) le citochinine naturali, abbiano origine dalla via biosintetica che dal mevalonato porta agli steroidi; via, come noto, utilizzata anche negli animali per la biosintesi di molti e importanti fattori ormonali.
Il trasporto dell'acido abscissico nella pianta intera sembra avvenire soprattutto nel floema. Nel picciolo delle foglie l'ormone si muove soprattutto in senso basipeto, il che è probabilmente da porre in relazione con la sua sintesi nel lembo e la sua azione nel mantenimento da parte dell'ormone dello stato di dormienza della gemma ascellare.
b) Effetti fisiologici.
Un'azione inibente dell'acido abscissico sulla crescita sia per moltiplicazione sia per distensione si rivela (con forti variazioni per i diversi materiali) anche nei sistemi più semplici quali le colture di tessuti, per es. nel callo di Ipomoea batatas coltivato in vitro.
In queste condizioni, come del resto anche sulla pianta integra, l'azione inibente risulta chiara quando nei tessuti la crescita è stimolata dalla presenza degli ormoni di tipo stimolante (auxina, gibberelline e citochinine). In condizioni fisiologiche, l'acido abscissico risulta inibire la crescita (probabilmente anche qui tanto per moltiplicazione che per distensione) negli apici di giovani coleottili di Graminacee e, in modo vistoso e dettagliatamente studiato, nei semi posti in condizioni adatte alla germinazione. Questo effetto inibente sulla germinazione è probabilmente di grande rilevanza fisiologica, in quanto induce a ritenere che l'alto contenuto in abscissine nei semi sia responsabile della loro dormienza, e la germinazione avvenga solo quando la sintesi di ormoni stimolanti, soprattutto gibberelline e cinetine, raggiunge livelli tali da spostare il rapporto tra fattori stimolanti e fattori inibenti a favore dei primi. È interessante osservare che nei semi non solo la crescita per moltiplicazione e distensione, ma anche processi metabolici che la precedono (come l'attivazione della sintesi degli enzimi che mobilizzano le riserve, quali l'α-amilasi, e anche altri) sono inibiti dall'acido abscissico, il cui effetto inibente può essere efficacemente rovesciato dall'aggiunta di gibberellina o citochinine.
Per esempio, la germinazione dei semi dormienti di lattuga, indotta da basse dosi di gibberellina, è di nuovo inibita da un successivo trattamento con acido abscissico; ma l'aggiunta, a questo punto, di citochinine provoca, ancora una volta, la ripresa della germinazione. In questo caso, come in molti altri, risulta evidente che la germinazione, o più generalmente la crescita, è sotto controllo simultaneo dei diversi ormoni delle due classi stimolanti e inibenti. L'andamento degli effetti sia sinergici sia antagonistici sembra peraltro dimostrare che i diversi ormoni interessano diverse aree metaboliche attraverso tipi diversi di recettori, anche quando le loro azioni confluiscono in manifestazioni finali in larga parte comuni.
Anche il mantenimento delle gemme nello stato dormiente, altro processo fisiologico caratterizzato dall'inibizione della crescita, è chiaramente influenzato dall'acido abscissico (v. quanto detto sopra circa la scoperta e l'isolamento di questo ormone).
Che l'acido abscissico influenzi i processi di differenziamento è facilmente intuibile in base alla sua azione sulla moltiplicazione e la distensione cellulare. Manifestazioni vistose di questa capacità di agire sul differenziamento sono l'induzione con acido abscissico della fioritura in alcune piante brevidiurne, come Pharbitis e Ribes, e l'inibizione dello stesso processo in longidiurne, come Lolium e Spinacia oleracea.
L'acido abscissico - come già detto - esplica un'azione di stimolo sull'abscissione, soprattutto chiara in test quali l'abscissione di piccioli di foglie il cui lembo sia stato asportato. Tale azione, peraltro, è spesso molto variabile, il che è comprensibile in considerazione della forte capacità di altri ormoni, soprattutto le auxine e l'etilene, d'influenzare lo stesso fenomeno.
Anche la senescenza, specie in certi suoi aspetti come la degradazione della clorofilla e la proteolisi in foglie recise, può, almeno in alcuni materiali, essere accelerata dall'acido abscissico. Il contenuto relativamente elevato dell'ormone nei frutti in via di maturazione suggerisce inoltre un suo intervento nelle fasi finali di questo processo, che può considerarsi come uno dei tanti casi di invecchiamento fisiologicamente controllato.
9. Gli ormoni vegetali come sistema regolatore unitario.
La rapida rassegna degli effetti fisiologici dei cinque principali tipi di ormoni vegetali noti, fatta nei paragrafi precedenti, porta ad alcune considerazioni di carattere generale, che possiamo così elencare: a) sia la crescita sia il differenziamento, nei loro principali effetti, sono fortemente influenzati da tutti e cinque i tipi di ormoni; b) per ogni effetto fisiologico appaiono costantemente fenomeni d'interazione, di tipo talora sinergico, talora antagonista, fra i diversi ormoni; c) mentre per un particolare organo o tessuto in una data situazione sperimentale la risposta fisiologica a un dato ormone o combinazione di ormoni risulta ben definita e riproducibile, in organi diversi la stessa risposta può essere provocata da ormoni diversi; inoltre un dato ormone spesso induce risposte diverse (talora opposte) in organi diversi, o addirittura nello stesso organo, se somministrato a concentrazioni diverse o in diverse condizioni sperimentali.
Il fatto che ogni ormone (o gruppo di ormoni) sia ben definito riguardo ai caratteri strutturali chimici e risulti capace d'indurre risposte fisiologiche caratteristiche in determinati tessuti e test, fa pensare che investa nella sua azione primaria un'area specifica del metabolismo cellulare, interagendo con uno o più recettori strettamente specifici. D'altra parte, queste diverse aree primarie di azione devono tenere sotto controllo, partendo da direzioni diverse, gli stessi fenomeni fisiologici generali che risultano, nelle diverse situazioni, influenzati, di volta in volta, dall'uno o dall'altro ormone. I cinque tipi di ormoni (o, per lo meno, la maggior parte di essi) sembrano essere, almeno di regola, simultaneamente presenti nei vari organi della pianta intera. Ciò è molto probabile per due considerazioni: 1) la localizzazione sfumata dei siti di sintesi degli ormoni nella pianta (in così stridente contrasto con la localizzazione precisa nelle cosiddette ghiandole a secrezione interna degli animali); 2) il fatto che non si sia finora riusciti a ottenere mutanti vitali incapaci di sintetizzare anche uno solo dei cinque tipi di ormoni vegetali noti, il che fa pensare che ognuno di essi (e in particolare dei tre ormoni stimolanti) sia in realtà necessario per il normale svolgimento di processi di base condizionanti sia crescita sia differenziamento. Le differenti risposte dei diversi organi o tessuti ai vari ormoni sarebbero quindi determinate dal fatto che il livello dell'uno o dell'altro ormone può scendere, in conseguenza di una particolare situazione, al di sotto di un certo valore e quindi diventare limitante per un dato processo fisiologico; mentre, d'altra parte, condizioni particolari di un tessuto (stato di differenziamento, particolarità dell'ambiente interno o esterno) possono modificare fortemente il grado di sensibilità di un processo metabolico a una data concentrazione di un determinato ormone.
Se queste considerazioni sono valide, lo svolgimento di fenomeni come moltiplicazione, distensione e differenziamento cellulare, dormienza o risveglio di organi dormienti, abscissione e invecchiamento fisiologico, trasporto e assorbimento attivo di soluti e sostanze nutritive, regolazione dell'acidità, della composizione ionica e della pressione osmotica dell'ambiente intra- e intercellulare (tutti toccati dai diversi ormoni) deve considerarsi come controllato e regolato non solo dalle concentrazioni dei singoli ormoni, ma anche, e talora soprattutto, dai loro rapporti relativi di concentrazione; infatti a diversi rapporti relativi corrispondono manifestazioni diverse delle potenzialità fisiologiche globali delle singole cellule e dei singoli tessuti e organi. In altri termini, lo stimolo e l'inibizione delle attività sopra menzionate, in ogni loro aspetto, esprimerebbero la risultante di un particolare stato di un unico sistema ormonale sostanzialmente unitario.
Una volta accettata questa visione globale del fenomeno delle correlazioni ormonali nella pianta, il problema principale che si pone, per una migliore conoscenza della funzione specifica di ogni ormone nell'ambito del sistema unitario cui esso partecipa, è quello della ricostruzione del suo meccanismo d'azione, a livello biochimico; è, come si suol dire, un problema di biologia molecolare. L'analisi a livello fisiologico è infatti impotente a risolvere il nodo del sottile gioco d'interazioni per cui i diversi stimoli ormonali, partendo da aree presumibilmente diverse, confluiscono, reciprocamente condizionandosi, in manifestazioni finali caratterizzate da enorme molteplicità di elementi determinanti e, insieme, da un altissimo grado di coordinamento.
10. Meccanismo d'azione degli ormoni vegetali.
L'analisi a livello molecolare del meccanismo d'azione di un ormone (come di qualunque composto attivo) presume la ricostruzione delle singole tappe d'azione, dall'interazione primaria tra ormone e recettore, giù giù lungo la catena di ripercussioni secondarie, fino all'ultimo anello della catena, a livello della manifestazione finale. Nel caso degli ormoni vegetali quest'analisi, intrapresa ormai da diversi decenni, ha portato finora a risultati estremamente modesti e incerti. In particolare: a) per nessun ormone è stata individuata la natura del recettore primario; b) in nessun caso è stata definita la natura di quella componente metabolica limitante a livello del processo finale che deve essere toccata da un ormone quando esso promuove (o inibisce) questo processo; c) anche la conoscenza delle tappe biochimiche intermedie del meccanismo d'azione dei singoli ormoni rimane incerta e in genere di difficile interpretazione.
Ciò nonostante, alcune tra le informazioni finora ottenute sono di notevole importanza soprattutto ai fini di una più precisa impostazione del problema. Le più importanti di queste informazioni possono essere riassunte come segue.
1. L'azione dei cinque gruppi di ormoni investe profondamente i meccanismi di regolazione della sintesi degli acidi nucleici e delle proteine. È ovvio che nei casi in cui gli ormoni stimolano (o inibiscono) la moltiplicazione cellulare, devono essere stimolate o inibite sia l'attività di gran parte del genoma nel promuovere la sintesi di RNA e proteine, sia la stessa replicazione del corredo genetico. Anche molti effetti degli ormoni sul differenziamento (per es. la lignificazione delle pareti cellulari indotta dall'auxina e dalle citochinine, la lisi delle pareti delle cellule nella zona di abscissione e molti tra i fenomeni caratteristici dell'invecchiamento) risultano riconducibili alla sintesi preferenziale di questo o quel tipo di proteine enzimatiche. Anche effetti ormonali che si svolgono, almeno in certe condizioni, senza variazioni apparenti della velocità di sintesi proteica potrebbero in realtà dipendere dallo stimolo della sintesi di alcuni enzimi chiave. Si è avanzata l'ipotesi, per es., che l'effetto dell'auxina sulla distensione cellulare sia dovuto all'induzione della sintesi di enzimi capaci di aumentare la plasticità della parete, diminuendone la resistenza alla spinta derivante dalla differenza tra pressione osmotica interna ed esterna.
Il fatto che l'azione degli ormoni sia in molti casi accompagnata da variazioni del livello in acidi nucleici, proteine ed enzimi specifici e, nella generalità dei casi, fortemente inibita dalla presenza di inibitori specifici della sintesi di queste macromolecole ha indotto molti autori a formulare l'ipotesi che l'azione primaria degli ormoni consista in un'interazione (attivante o inibente) con proteine regolatrici specificamente adibite al controllo dell'attività dei singoli geni. Tuttavia, dati recenti si oppongono a una simile generalizzazione e dimostrano che in alcuni casi l'azione degli ormoni rimane apprezzabile anche in condizioni di blocco totale della sintesi di proteine (per es. nel caso dell'effetto dell'auxina sulla distensione). Rimane quindi, al momento, del tutto aperto il problema del meccanismo d'azione dei diversi ormoni sulla sintesi proteica (sia generale che specifica), meccanismo circa il quale, d'altra parte, le moderne conoscenze di biologia molecolare forniscono un numero elevato di modelli possibili (due di questi sono schematizzati nella fig. 16, A e B).
È d'altra parte importante tener presente che, anche indipendentemente da un'azione sulla sintesi di determinati enzimi o altre proteine, gli ormoni potrebbero esplicare alcuni dei loro effetti agendo sull'attività di proteine già esistenti. Questo meccanismo appare molto probabile nei casi in cui l'attività fisiologica degli ormoni risulta accompagnata da rapide modificazioni di parametri metabolici (per es., l'azione dell'auxina sulla crescita per distensione è in genere accompagnata da simultaneo aumento della respirazione). Effetti degli ormoni sull'attività di singoli enzimi potrebbero, d'altra parte, portare a modificazioni di livelli endocellulari di metaboliti tali da coinvolgere secondariamente anche la regolazione della sintesi proteica (v. fig. 16 C).
Negli ultimi anni molti ricercatori hanno cercato di di- mostrare in sistemi cellulari isolati l'esistenza di un'azione diretta dei diversi ormoni (soprattutto auxina e gibberelline) sia sulla sintesi di acidi nucleici e proteine, sia sull'attività di enzimi isolati. Purtroppo, i risultati di questi studi appaiono finora incerti, o contraddittori, o scarsamente riproducibili.
2. Anche l'analisi degli aspetti terminali del meccanismo d'azione degli ormoni appare appena iniziata. Il caso più intensamente studiato è, ancora una volta, quello dell'effetto dell'auxina sulla distensione. La maggior parte degli studiosi ritiene oggi che la crescita per distensione sia riconducibile a un aumento della distensibilità plastica della parete cellulare, tale da far si che la tendenza della cellula a richiamare acqua (dovuta a un gradiente osmotico) non sia più bilanciata dalla capacità di resistenza della parete. Di fatto, nei tessuti stimolati da auxina un aumento della distensibilità plastica della parete risulta ben evidente e strettamente correlato con l'effetto sulla distensione.
Rimane tuttora oscuro non solo il tipo di modificazione della struttura della parete da cui dipende la variazione di distensibilità, ma anche il meccanismo attraverso il quale l'auxina induce questa variazione; anche se risulta chiaro che l'ormone non agisce direttamente sulla parete, bensì a livello del citoplasma, interessandovi processi di trasporto e di utilizzazione di energia (tutti i veleni della respirazione e del trasporto di energia e, in genere, ogni lesione dell'integrità cellulare inibiscono l'azione dell'ormone sulla distensione). Recentissimamente, alcune osservazioni prospettano, come suggestiva ipotesi di lavoro, la possibilità che l'auxina agisca attivando il trasporto di idrogenioni attraverso la membrana cellulare e che la conseguente diminuzione del pH nella parete sia responsabile dell'aumento di plasticità. Tale ipotesi è sostenuta da due ordini di considerazioni: a) la capacità (aspecifica) di soluzioni acide di indurre sulla distensione di cellule vive, o sulla distensibilità della parete di cellule paralizzate da inibitori o ‛uccise' da congelamento e scongelamento, effetti del tutto simili a quelli indotti dall'auxina in condizioni fisiologiche; b) il reperto che in quelle condizioni sperimentali in cui non solo l'auxina ma anche altri ormoni, come le citochinine, stimolano la distensione, è di regola altresì ben dimostrabile un proporzionale effetto di aumento dell'acidità del mezzo esterno.
Questo tentativo di ricostruzione del meccanismo della distensione introduce, quale nuova dimensione nell'analisi del modo di agire degli ormoni, l'ipotesi di un controllo del trasporto attivo a livello delle membrane. Un'ipotesi quanto mai interessante, in quanto da un lato prospetta un collegamento tra effetti degli ormoni sul metabolismo ossidativo e sulla distensione (ogni fenomeno di trasporto attivo, infatti, deve dipendere dall'energia resa disponibile dai processi respiratori), dall'altro apre il problema della regolazione nelle piante dello stato chimico-fisico del mezzo intercellulare, rappresentato essenzialmente dallo spazio libero di cui le pareti pectocellulosiche sono parte. È pertinente ricordare, in proposito, che argomenti di ordine generale rendono estremamente probabile che non solo il pH, ma anche la concentrazione dei singoli ioni e, in genere, dei soluti nella parete cellulare siano accuratamente regolati su valori nettamente diversi da quelli del citoplasma. In particolare, sembra necessaria una regolazione del valore della pressione osmotica nella parete, in quanto proprio dalla differenza tra pressione osmotica all'interno e all'esterno della membrana cellulare dipende la possibilità di distensione delle cellule. Dati recenti fanno quindi ritenere che anche nelle piante, come negli animali superiori, osmoregolazione e controllo del pH e della composizione del mezzo intercellulare rappresentino un momento comune di fondamentale importanza nel quadro dell'integrazione correlativa delle diverse attività funzionali.
Bibliografia.
Abeles, F. B., Biosynthesis and mechanism of action of ethylene, in ‟Annual review of plant physiology", 1972, XXIII, pp. 259-292.
Addicott, F. T., Lyon, J. L., Physiology of abscisic acid and related substances, in ‟Annual review of plant physiology", 1969, XX, pp. 139-164.
Carr, D. J. (a cura di), Plant growth substances 1970, Berlin 1972.
Cleland, R., Cell wall extension, in ‟Annual review of plant physiology", 1971, XXII, pp. 197-222.
Galston, A. W., Davies, P. J., Control mechanisms in plant development, Englewood Cliffs, N. J., 1970.
Key, J. L., Hormones and nucleic acid metabolism, in ‟Annual review of plant physiology", 1969, XX, pp. 449-474.
Lang, A., Gibberellins: structure and metabolism, in ‟Annual review of plant physiology", 1970, XXI, pp. 537-570.
Marrè, E., Ciferri, O., Regulation of cell membrane activities in plants, Amsterdam 1976.
Milborrow, B. V., The occurence and function of abscisic acid in plants, in ‟Science progress", 1969, LVII, pp. 533-558.
Salisburg, F. B., Ross, C., Plant physiology, Belmont 1969.
Skoog, F., Armstrong, D. J., Cytokinins, in ‟Annual review of plant physiology", 1970, XXI, pp. 359-384.
Steward, F. C., Plant physiology, vol. VI B, New York 1972.
Wareing, P. F., Saunders, P. F., Hormones and dormancy, in ‟Annual review of plant physiology", 1971, XXII, pp. 261-288.
Wightman, F., Setterfield, G., Biochemistry and physiology of plant growth substances, Ottawa 1968.
Wilkins, M. B. (a cura di), Physiology of plant growth and development, New York 1969.