
Orologi biologici
Orologi biologici
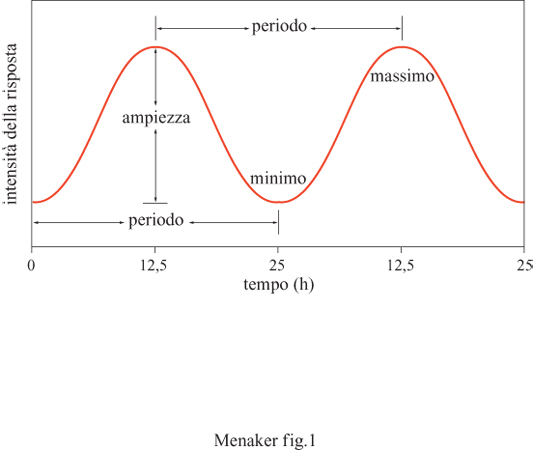
Variazioni giornaliere nel comportamento di molti organismi sono state osservate sino dai tempi più antichi, ma solo a partire dalla seconda metà del 20° secolo i biologi hanno iniziato a interessarsi in modo scientifico a questi fenomeni. Per molti anni si è creduto che questi ritmi fossero una semplice risposta alla variazione quotidiana dell'ambiente fisico. II risveglio d'interesse per questo tipo di studi seguì la dimostrazione che tali variazioni nel comportamento o nella fisiologia sono controllate da accurati sistemi interni di misurazione: gli orologi biologici. Molti studi hanno dimostrato che la maggior parte degli organismi viventi, sia vegetali che animali, possiede un oscillatore endogeno, dotato di un periodo di oscillazione simile alla durata del giorno astronomico. Inoltre molti organismi possiedono orologi che oscillano con un periodo simile a quello di altri cicli geofisici che rivestono un ruolo importante nella loro vita. Introduzione Lo studio degli orologi biologici e dei ritmi da essi generati è chiamato cronobiologia. Un ritmo viene definito come un evento che si ripete regolarmente dopo un certo intervallo di tempo, ed è caratterizzato da tre parametri: il periodo, l'ampiezza e la fase (fig. 1).
Il periodo è il tempo necessario al ritmo per completare un intero ciclo (cioè il tempo che intercorre fra due picchi dell'oscillazione). L'ampiezza è la differenza, in valore assoluto, fra il valore minimo e il valore massimo dell'oscillazione. La fase rappresenta invece la relazione temporale fra un punto identificabile del ciclo e un punto su un ciclo di riferimento. Il periodo dei ritmi biologici varia da pochi millisecondi a molte ore, giorni e persino anni (tab. I). Alcuni di questi ritmi non mostrano una relazione diretta con i cicli presenti nell'ambiente fisico; essi si sono evoluti in risposta alle richieste fisiologiche dell'organismo, quindi la loro periodicità dipende dalle dinamiche di tali richieste (per esempio, il battito del cuore e la ritmicità spontanea della scarica di molti neuroni). Altri ritmi, invece, si sono evoluti in risposta ai cicli ambientali e mantengono una chiara relazione di fase con essi. Questi ritmi biologici possono avere un periodo simile a quello del giorno astronomico (ritmi circadiani), a quello delle maree (ritmi circatidali), delle fasi lunari (ritmi circalunari) e dell'alternanza delle stagioni (circannuali). Tali ritmi persistono quando gli organismi vengono isolati dal ciclo ambientale con cui sono in relazione. Ciò dimostra in modo inequivocabile che essi sono generati in maniera endogena e autonomamente dagli stimoli esterni. In assenza di segnali ambientali i ritmi circadiani hanno un periodo di circa 24 ore, mentre quelli circatidali, circalunari e circannuali hanno periodi rispettivamente di circa 12,4 ore, 29,5 ore e 12 mesi (v. tabella l).
Ritmi biologici senza relazione con i cicli geofisici
Poiché la maggior parte dei processi biologici è regolata da meccanismi di controllo coinvolgenti una retro azione che genera sempre oscillazioni, in ogni organismo sono presenti migliaia di ritmi; tuttavia, solo una minima parte di questi è sotto il controllo di un orologio biologico. l cronobiologi hanno deciso di raggruppare i ritmi in due categorie: i ritmi con periodicità più corta di un giorno (cioè di durata inferiore alle 24 ore) e quelli con periodicità più lunga di un giorno (cioè di durata maggiore alle 24 ore).
Ritmi ultradiani
l ritmi ultradiani sono ritmi endogeni con una periodicità che può variare da alcuni millisecondi ad alcune ore, senza una correlazione con i cicli ambientali a noi noti. Esempi specifici di tale ritmicità sono la frequenza di scarica dei recettori del freddo nel nervo linguale, che ha una periodicità di circa 100 ms, e l'attività elettrica del ganglio parietale della lumaca (Helix pomatia), che ha una periodicità di circa 5 s. l ritmi ultradiani sono anche presenti a livello comportamentale. Per esempio, il verme Arenicola marina, che vive nella zona sabbiosa intertidale emerge dalla propria tana a intervalli regolari della durata di 6÷8 min. In maniera simile, l'arvicola Microtus pennsylvanicus, che è attiva di giorno, ha uno spiccato ritmo ultradiano caratterizzato da periodi di attività e riposo che si susseguono con un periodo di circa 2 ore. Ritmi ultradiani simili sono presenti in altri piccoli mammiferi. Anche la presenza nel sangue di alcuni ormoni, quali l'ormone luteinizzante, l'ormone della crescita e il corticosterone, è regolata da ritmi ultradiani.
Ritmi infradiani
l ritmi con un periodo più lungo di 24 ore e che non mostrano nessuna relazione con i cicli geofisici sono chiamati ritmi infradiani. l più comuni e i più studiati sono i ritmi infradiani associati con il ciclo estrale delle femmine dei mammiferi. Per esempio, le femmine dei criceti hanno un ciclo estrale di circa 4 giorni, mentre nelle femmine dei ratti la durata è di circa 4÷5 giorni. Pecore e cavie domestiche hanno, invece, un ciclo di circa 16 giorni. Ci sono dubbi circa l'appartenenza del ciclo mestruale delle donne a questa categoria, poiché esistono buoni motivi per credere che esso si sia evoluto per essere sincronizzato dal ciclo lunare.
Ritmi biologici con periodicità simile a quella dei cicli geofisici
Passiamo ora a descrivere i ritmi biologici generati dagli orologi biologici interni e sincronizzati con i cicli geofisici in modo tale da consentire l'adattamento degli organismi all'ambiente esterno.
Ritmi circatidali
La caratteristica principale dell'ambiente costiero è la variazione delle condizioni fisiche dovuta alle maree. Le maree sono il risultato dell'attrazione che la forza gravitazionale della Luna (e del Sole) esercita sulla Terra ogni 12,4 ore. Molti organismi che vivono nelle zone intertidali hanno evoluto ritmi nel comportamento e nella fisiologia con un periodo simile a quello delle maree. Le ostriche, per esempio, aprono le loro valve per nutrirsi durante l'alta marea, quando si trovano immerse nell'acqua, e le chiudono quando l'acqua si ritira durante la bassa marea. l granchi violino emergono dalle loro tane con la bassa marea, per farvi ritorno solo all'arrivo dell'alta marea. Molte ricerche hanno chiaramente dimostrato che questi ritmi persistono quando gli animali sono tenuti in ambienti artificiali costanti, in assenza di maree. Tale risultato dimostra in modo inequivocabile che gli organismi intertidali possiedono un orologio biologico tidale o, più precisamente, un orologio intertidale.
Ritmi circadiani
Molti parametri biochimici, fisiologici e comportamentali che caratterizzano gli organismi animali e vegetali mostrano cospicue fluttuazioni giornaliere, che nella maggioranza dei casi persistono in condizioni d'isolamento dall'ambiente esterno; ciò dimostra che sono indotte da un oscillatore endogeno. l ritmi circadiani sono stati descritti in quasi tutti i gruppi di organismi, da quelli unicellulari (persino qualche batterio) ai mammiferi. Attualmente siamo in possesso di molte informazioni sulla fisiologia, l'organizzazione, la genetica e i meccanismi molecolari che sono alla base di tale oscillazione. Al contrario, la nostra conoscenza dei meccanismi, o anche della semplice localizzazione anatomica delle strutture che generano l'oscillazione dei ritmi circatidali, circalunari e circannuali è molto limitata. Per questo motivo nel presente saggio ci occuperemo principalmente dei ritmi circadiani. Vogliamo però ricordare al lettore che molte delle caratteristiche generali dei ritmi circadiani sono applicabili anche a tutti gli altri tipi di ritmicità, al di là del periodo caratteristico della loro oscillazione.
Ritmi circalunari
L'intensità dell'illuminazione notturna varia con le fasi lunari seguendo un ciclo di circa 29 giorni che è determinato dalla rivoluzione della Luna intorno alla Terra, e che è in relazione con la ritmicità tidale. Poiché tramite il ciclo lunare si può predire con estrema precisione quando si verificherà la marea primaverile più alta e quando quella più bassa, il moscerino marino Clunio marinus coordina la schiusa delle uova, l'accoppiamento e la deposizione con il ciclo lunare. Un altro famoso esempio è dato dal pesce marino Leuresthes tenuis che depone le uova sulle spiagge sabbiose della California durante le maree primaverili, usando le fasi della Luna come parametro temporale. Anche la deposizione delle uova di molti vermi marini viene sincronizzata con il ciclo lunare. Forse il più famoso di questi esempi è offerto dal verme Eunice viridis, il paIolo, che vive nella barriera corallina del Pacifico Meridionale. Durante il periodo che va da ottobre a dicembre, nell'ultimo giorno dell'ultimo quarto del ciclo lunare, un segmento del corpo di questi vermi si stacca, inizia a nuotare e raggiunge la superficie dove rilascia le uova e gli spermi, e dove avviene la fecondazione delle uova. La sincronizzazione di questo evento in tutta la popolazione assicura il pieno successo riproduttivo. Anche in questo caso studi condotti in laboratorio hanno dimostrato che questi ritmi persistono in condizioni costanti, cioè sono generati da un orologio biologico che ha un periodo simile al ciclo lunare.
Ritmi circannuali
l ritmi annuali sono determinati dal moto di rivoluzione della Terra intorno al Sole, hanno un periodo di l anno e sono tanto più pronunciati quanto maggiore è la distanza dall'equatore. Anche in questo caso è stato dimostrato che in molte specie di uccelli e di mammiferi i ritmi stagionali (annuali) dipendono da un orologio biologico endogeno circannuale. Nei mesi invernali alcuni mammiferi delle zone temperate entrano in un stato di sonno profondo durante il quale l'attività metabolica è ridotta (ibernazione). Tale stratagemma adattativo permette a questi animali di superare le avverse condizioni ambientali del periodo invernale. l periodi di ibernazione e attività si susseguono con periodicità annuale in natura, e persistono in laboratorio, in condizioni costanti, mantenendo per anni la stessa periodicità. Molte specie di uccelli sfuggono al rigore invernale delle regioni più fredde migrando verso le regioni più meridionali in cui il clima è più mite. l ritmi associati con queste migrazioni hanno spesso una forte componente endogena che è sotto il controllo di un orologio circannuale. Il saltimpalo Saxicola torquata può essere citato come un ottimo esempio di questa ritmicità. Quando viene mantenuto in laboratorio, in condizioni costanti, continua a mostrare un ciclo, con un periodo di circa 12 mesi, nella crescita e nel declino delle gonadi così come nella muta delle penne. Anche molte specie d'insetti che vivono in aree geografiche con forti variazioni stagionali possiedono un orologio biologico circannuale che regola l'attività e la diapausa.
Caratteristiche generali degli oscillatori circadiani
Un oscillatore circadiano è una parte di un organismo vivente (una cellula, un tessuto o un organo) capace, quando è isolata e mantenuta in condizioni costanti, di generare autonomamente un'oscillazione che ha un periodo di circa 24 ore.
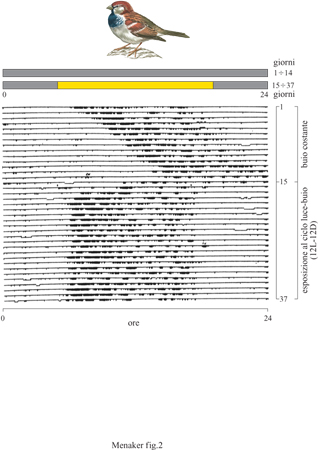
Quando un organismo è tenuto in condizioni costanti, eliminando le variazioni dovute all'ambiente, i ritmi circadiani dell'organismo persistono con un periodo simile, ma nella maggioranza dei casi diverso dalle 24 ore (fig. 2). Questo risultato dimostra che il ritmo è autonomo dall'ambiente ed è endogeno; il periodo di un ritmo in tali condizioni è detto periodo difree-running (corsa libera). Esso è direttamente correlato con quello dell'oscillatore biologico che lo genera, e perciò viene usato dagli studiosi di cronobiologia come una misura dell'orologio biologico stesso. Inoltre, dato che il periodo di free-running, per una data specie, è determinato geneticamente, tale parametro può essere utilizzato per valutare come particolari mutazioni genetiche influenzino l'orologio stesso.
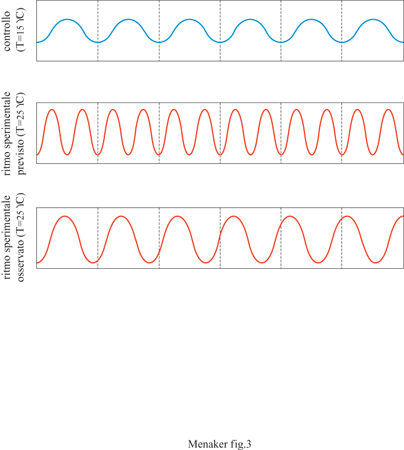
Un'altra caratteristica importante dei ritmi circadiani è la compensazione delle variazioni di temperatura: il periodo di free-running di un ritmo è influenzato soltanto in maniera modesta dalle variazioni della temperatura interna dell'organismo (fig. 3). Per la maggior parte dei processi biologici la velocità con cui il processo avviene raddoppia o triplica aumentando la temperatura di 10 °C. Al contrario, nei ritmi circadiani degli organismi sinora studiati, la velocità cambia di un fattore pari a circa 0,85÷1,3, ovvero il periodo dell'oscillazione cambia poco con il variare della temperatura. La compensazione delle variazioni di temperatura ha un significato adattativo importante, soprattutto negli organismi eterotermi (cioè quasi tutti gli organismi viventi esclusi uccelli e mammiferi) che non sono capaci di regolare in maniera fisiologica la loro temperatura corporea. Ovviamente, un orologio biologico in cui la velocità dell'oscillazione fosse significativamente influenzata dalla temperatura sarebbe di poca utilità in organismi in cui la temperatura corporea varia quotidianamente in maniera considerevole.
Per essere utili, gli orologi circadiani, come i nostri orologi da polso, devono essere sincronizzati con la rotazione della Terra intorno al suo asse. Il processo con cui un ritmo circadiano si sincronizza con un ciclo ambientale (di solito l'alternanza del giorno e della notte) è chiamato sincronizzazione (entrainment, v. figura 2). Data la sua importanza, molti ricercatori si sono dedicati allo studio dei meccanismi che sono alla base della sincronizzazione, e di conseguenza si possiedono molte informazioni su questo fenomeno. Benché numerosi fattori ambientali mostrino regolari variazioni giornaliere, solo alcuni di essi sono in grado di sincronizzare i ritmi circadiani. È stato dimostrato che la luce è il parametro ambientale più usato da tutti gli organismi, uomo incluso, per la sincronizzazione dei ritmi circadiani. Molti organismi possono essere sincronizzati dai cicli della temperatura, e in alcuni uccelli e mammiferi anche il cibo e le interazioni sociali possono essere usati per tale scopo.
Il termine specifico per la definizione di uno stimolo capace di sincronizzare un ritmo è zeitgeber (parola tedesca che significa "che dà il tempo"). Un ritmo circadiano è sincronizzato quando il suo periodo è uguale a quello del suo stimolo sincronizzatore (v. figura 2). Un ritmo sincronizzato mantiene sempre una relazione fissa con il suo zeitgeber la cui caratteristica fondamentale è la capacità, durante il processo di sincronizzazione, di alterare il periodo di free-running del ritmo circadiano in oggetto. Se tale periodo è più lungo rispetto a quello dello zeitgeber, quest'ultimo lo accorcerà, mentre se il periodo di free-running è più corto, lo allungherà.
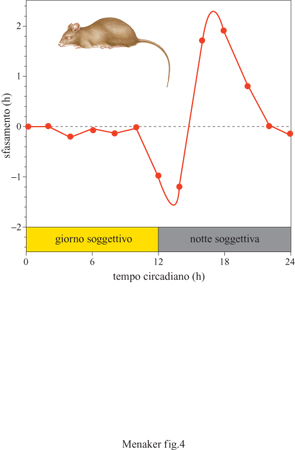
Il processo di sincronizzazione è stato analizzato in dettaglio e attualmente sono disponibili modelli che lo descrivono mediante un formalismo matematico (Pittendrigh e Daan, 1976). La sincronizzazione dell'orologio biologico viene raggiunta per mezzo di un cambiamento immediato della fase che può essere prodotto da un impulso di luce, così come da un ciclo di luce; il suo effetto dipende dal momento del ciclo circadiano in cui si verifica il singolo impulso. La risposta dell'organismo allo stimolo luminoso dipende, dunque, dalla fase temporale in cui esso si verifica (fig. 4), e la sua rappresentazione grafica è detta PRC (Phase Response Curve, curva di risposta di fase). La forma generale della PRC è simile per tutti gli organismi ma le sue caratteristiche dipendono da fattori quali la durata e l'intensità dello stimolo, la storia dell'organismo (cioè le condizioni ambientali a cui l'organismo è stato soggetto nella sua intera esistenza prima del test) e la specie. Se per un dato organismo si conoscono la PRC e il periodo di free-running, è possibile allora predire come e con quale fase l'organismo si sincronizzerà con un qualsiasi ciclo luce-buio scelto dallo sperimentatore. In natura gli zeitgeber naturali (principalmente i cicli di luce e di temperatura) mantengono gli organismi sincronizzati con l'ambiente azzerandoli giornalmente. La forma della PRC e il periodo di free-running determinano la relazione di fase fra l'organismo e il suo ambiente naturale, determinano cioè se l'organismo è diurno o notturno.
Diffusione e localizzazione degli orologi circadiani
A partire dagli anni Cinquanta i ritmi biologici sono stati studiati sotto vari aspetti: l'aspetto descrittivo, quello formale (matematico), quello fisiologico e, di recente, quello genetico e molecolare. All'inizio, la maggiore attenzione è stata rivolta alla loro descrizione. Questi studi iniziali dimostrarono che la ritmicità circadiana è molto diffusa nei protozoi, nei funghi, nelle piante e negli animali. l ritmi circadiani sono stati studiati in maniera particolareggiata in Euglena, Paramecium e Gonyaulax. Nel fungo Neurospora la crescita e la formazione delle spore sono regolate da ritmi circadiani, e per questo motivo tale organismo è stato, e continua a essere, un modello per lo studio della biologia molecolare e della genetica dei ritmi circadiani. l movimenti fogliari e dei petali e la secrezione del nettare rappresentano solo pochi esempi della miriade di ritmi circadiani presenti nelle piante. Fra gli animali pluricellulari, la ritmicità circadiana è stata descritta per tutti i raggruppamenti sistematici: dai celenterati fino ai mammiferi, uomo incluso.
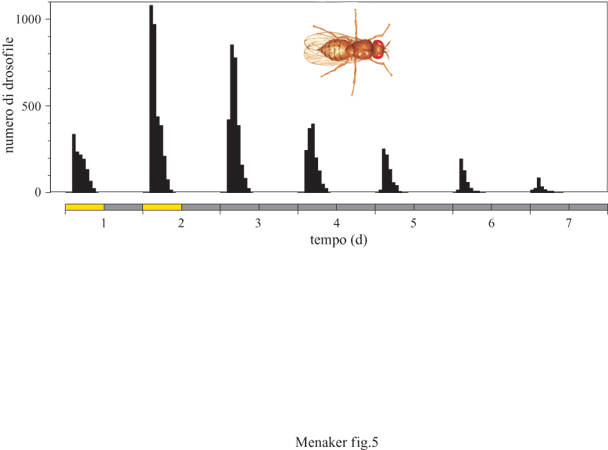
Degno di nota è anche il fatto che i ritmi circadiani sono stati descritti a livello di popolazione. Infatti, anche se ogni animale di una popolazione può avere un certo tipo di attività solo una volta nella vita, la distribuzione temporale di queste attività individuali può essere ritmica. Un esempio da ricordare in questa sede è il ritmo circadiano con cui i moscerini della frutta (Drosophila) emergono dal loro involucro pupale (fig. 5). Anche le nascite nella specie umana mostrano questo tipo di ritmicità essendo concentrate nelle ore mattutine. Di recente i ritmi circadiani sono stati descritti anche nei cianobatteri, dimostrando perciò la loro presenza anche nei procarioti.
Localizzazione anatomica degli orologi biologici
Negli organismi pluricellulari gli orologi biologici responsabili del controllo dei ritmi circadiani sono localizzati in particolari strutture che, salvo rare eccezioni, fanno parte del sistema nervoso. Sono stati sinora adottati tre approcci sperimentali per identificare le strutture contenenti un oscillatore circadiano: la lesione, la coltura in vitro e il trapianto.
Osservando gli effetti di una lesione in un'area del cervello o della rimozione di una determinata struttura su un particolare tipo di ritmo, è possibile determinare se tale struttura è coinvolta o meno nella sua regolazione. Nonostante presenti alcuni svantaggi, questa tecnica è stata utilizzata per una localizzazione preliminare degli orologi biologici in molte specie. Per esempio, l'orologio biologico che controlla la maggior parte dei ritmi circadiani dei mammiferi, costituito dai nuclei soprachiasmatici dell'ipotalamo, è stato localizzato per la prima volta utilizzando questa tecnica (Stephan e Zucker, 1972). Tuttavia, i risultati che si ottengono con le lesioni sono difficili da interpretare, e in ogni caso non dimostrano che la struttura lesionata è davvero un orologio biologico.
L'interpretazione dei risultati è meno ambigua quando si analizzano contemporaneamente i dati ottenuti effettuando lesioni e quelli provenienti da colture e da trapianti. Con l'avvento di alcune nuove tecniche è divenuto possibile, in alcuni casi, rimuovere parti anatomiche, mantenerle in vitro in condizioni costanti per molti giorni, e contemporaneamente misurare qualche parametro potenzialmente ritmico, per esempio l'attività elettrica, o il rilascio di un ormone, o la sintesi di un peptide. Se in queste condizioni il parametro misurato mantiene una propria ritmicità, la struttura contiene un orologio biologico. Con questo tipo di approccio è stato possibile dimostrare la presenza di un orologio biologico nella retina e nella ghiandola pineale dei vertebrati non mammiferi, come pure nella retina e nei nuclei soprachiasmatici dell'ipotalamo dei mammiferi.
Nonostante questi esperimenti confermino la presenza di un orologio circadiano nel tessuto mantenuto in coltura, essi non stabiliscono la sua funzione di orologio nell'organismo intatto. l trapianti, invece, possono fornire informazioni utilissime per stabilire il ruolo di un organo nell'ambito dell'organizzazione temporale dell'organismo studiato. Per esempio, nei roditori la lesione dei nuclei soprachiasmatici dell'ipotalamo determina la scomparsa della maggior parte dei ritmi circadiani, mentre il trapianto di questa struttura nervosa ripristina il ritmo circadiano dell'attività locomotoria, ma non altri ritmi. Inoltre, è stato dimostrato che se si trapiantano i nuclei soprachiasmatici dell'ipotalamo di un animale affetto da una mutazione genetica, che altera il periodo del ritmo dell'attività locomotoria, in un animale non affetto dalla mutazione, quest'ultimo presenterà un ritmo di attività locomotoria con un periodo simile a quello dell'animale donatore. Altri esperimenti con i trapianti hanno permesso di dimostrare che la ghiandola pine aie del passero domestico è un orologio biologico, così come il lobo ottico degli scarafaggi e il cervello del moscerino della frutta (Drosophila) e del baco da seta.

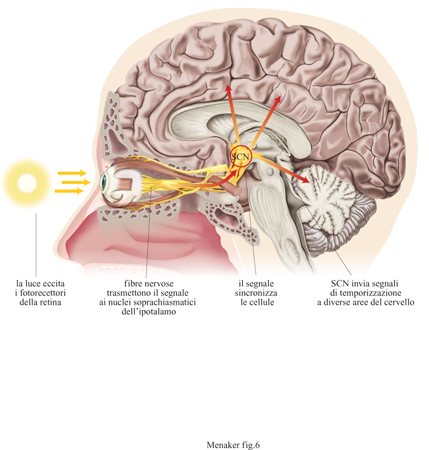
Nella tabella (tab. 2) sono menzionate tutte le strutture anatomiche in cui è stata dimostrata, in maniera diretta o indiretta, la presenza di un orologio circadiano, anche se non necessariamente il suo ruolo funzionale: risulta evidente che negli organismi pluricellulari sono presenti varie parti anatomiche (tessuti o organi) che contengono un orologio biologico, o almeno oscillatori che possono potenzialmente agire come orologi. Di solito negli animali gli orologi biologici sono stati localizzati nel sistema nervoso, ma studi recenti hanno evidenziato alcune eccezioni, mostrando che essi possono essere localizzati anche al di fuori di questo. Per esempio, in Drosophila è stato visto che i tubi malpighiani, che negli insetti equivalgono ai reni, contengono oscillatori circadiani (Giebultowicz e Hege, 1997). Inoltre, sembra che in questo insetto gli oscillatori circadiani siano localizzati anche nella proboscide e sui bordi delle ali (Plautz et al., 1997). Fra i vertebrati, la lucertola Iguana iguana possiede almeno cinque parti anatomiche contenenti un orologio biologico: la ghiandola pineale, l'occhio parietale, le retine e un altro sito, non ancora identificato, nel cervello (Tosini e Menaker, 1998). Generalizzando, si può affermare che nei vertebrati non mammiferi vi sono molte strutture capaci di oscillare con un periodo circadiano. Nei mammiferi, invece, l'orologio biologico più importante è localizzato, come già detto, nei nuclei soprachiasmatici dell'ipotalamo (fig. 6), e un altro orologio circadiano, la cui funzione è per lo più sconosciuta, è stato localizzato nella retina (Tosini e Menaker, 1996).
Gli orologi degli organismi pluricellulari sono eterogenei e nella maggior parte dei casi il tipo di cellula che genera la ritmicità non è stato ancora identificato. L'unica eccezione è l'identificazione delle cellule responsabili della variazione dell'attività elettrica nell'occhio del mollusco marino Bulla gouldiana (Michel et al., 1993). In questo animale l'orologio è stato localizzato in alcune centinaia di cellule elettricamente interconnesse chiamate neuroni basali della retina (BRN, Basai Retinal Neurons). Inoltre, un singolo neurone di questo tipo mantiene un'attività elettrica ritmica anche quando viene isolato e posto in coltura. L'orologio situato negli occhi dei roditori è localizzato nella retina neurale che, naturalmente, contiene molti tipi di cellule. Nella rana Xenopus laevis è presente un orologio biologico, localizzato nei fotorecettori, che regola la sintesi della melatonina (Cahill e Besharse, 1993). Anche alcune cellule della ghiandola pineale dei pesci, delle lucertole e degli uccelli contengono, forse, oscillatori circadiani, così come alcuni neuroni dei nuclei soprachiasmatici dell 'ipotalamo dei mammiferi (Welsh et al., 1995).
Organizzazione dei sistemi circadiani
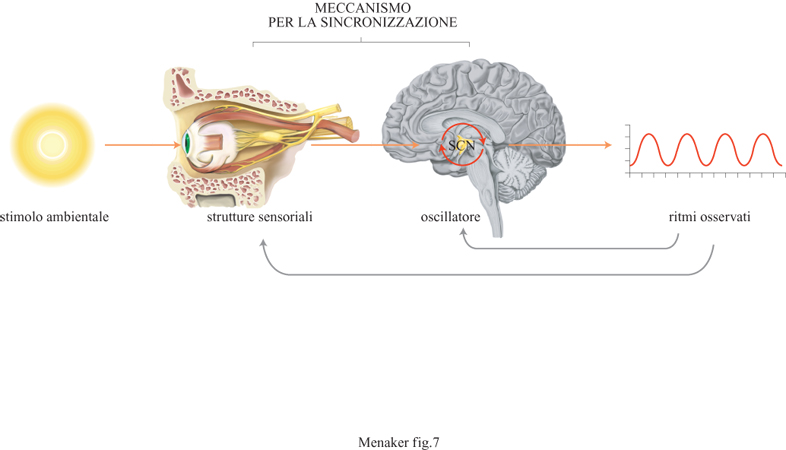
Da quanto detto in precedenza risulterà chiaro al lettore che in un organismo sono presenti molti orologi biologici che interagiscono fra di loro, con il resto dell'organismo e con l'ambiente esterno. Cosa sappiamo di questa organizzazione ciro cadiana? Un sistema circadiano, per essere organizzato ir maniera funzionale, deve essere costituito da tre componen· ti: gli oscillatori che generano il ritmo, una via sensoriale d input per la sincronizzazione degli oscillatori con l'ambiente esterno, e un output proveniente dall'oscillatore e capace d controllare i ritmi a livello dell'organismo (fig. 7). Il modo in cui queste parti sono assemblate può variare moltissimo fra diversi organismi a seconda delle strutture e dei percors neuronali coinvolti. Tale variabilità rende molto difficih discutere l'organizzazione circadiana in termini generali per questa ragione limiteremo la nostra trattazione all'organizzazione del sistema circadiano nei vertebrati.
L'organizzazione circadiana dei vertebrati è resa possibile da un asse circadiano costituito dalla retina, dai nuclei soprachiasmatici dell'ipotalamo, dalla ghiandola pineale e dalle interconnessioni esistenti fra queste strutture. Tali strutture interagiscono per controllare i ritmi circadiani, tramite segnali in uscita che coinvolgono sia gli impulsi nervos sia gli ormoni. La sincronizzazione di queste strutture con il ciclo luce-buio avviene tramite i segnali luminosi che vengono percepiti da speciali fotorecettori, dettifotorecettori circadiani, presenti nella retina (nei mammiferi e probabilmente in tutte le altre classi di vertebrati), nella ghiandole pineale (in tutte le classi di vertebrati non mammiferi) e nel cervello di tutti i vertebrati eccetto i mammiferi (fotorecettori cerebrali profondi), in aree non ancora localizzate. Come risulta chiaro da questa breve descrizione, le medesime strutture spesso contengono sia gli oscillatori circadiani sia i fotorecettori che le connettono con il mondo esterno. Un'organizzazione simile è diffusa ampiamente tra gli invertebrati.
Meccanismi che generano l'oscillazione circadiana
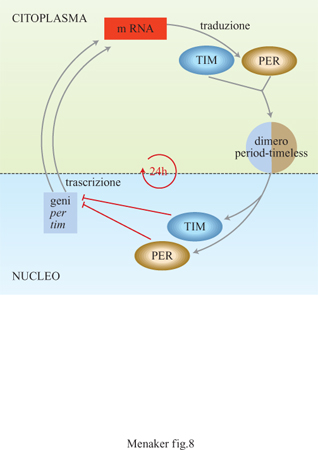
La ritmicità circadiana è stata studiata a livello genetico e molecolare in Neurospora e in Drosophila; i risultati di queste ricerche hanno fornito importanti indicazioni sui meccanismi subcellulari che generano questa oscillazione (Dunlap, 1996). Negli anni Settanta R.J. Konopka e S. Benzer (1971) iniziarono uno screening genetico per identificare le mutazioni che influiscono sul sistema circadiano (v. il saggio di R. Greenspan, nel l volume). l loro studi consentirono di identificare tre alleli mutanti di un singolo gene che fu chiamato period (per, da periodo). l moscerini che possedevano l'allele peri avevano un periodo di free-running molto più lungo del normale, quelli con l'allele per8 avevano un periodo molto più corto, mentre i moscerini con l'allele per0 erano completamente aritmici. Il gene per è stato localizzato successivamente sul cromosoma X, in seguito clonato e, attualmente, oggetto di studio da parte di molti gruppi di ricerca che si occupano della genetica e della biologia molecolare dei ritmi circadiani. l livelli dell'RNA messaggero di questo gene e della proteina da esso codificata, PER, hanno entrambi un ritmo circadiano ma c'è un ritardo di 6÷8 ore fra la sintesi dell'RNA e la produzione della proteina stessa. Recentemente in Drosophila è stato scoperto un altro gene mutante che è stato chiamato timeless (tim, cioè senza tempo) perché i moscerini mutanti sono aritmici (Sehgal et al., 1994). l moscerini omozigoti per questa mutazione perdono non solo la ritmicità nel comportamento, ma anche la ritmicità circadiana nell'espressione dell'RNA messaggero del gene per; ciò suggerisce che l'espressione ritmica della proteina PER è importante per il mantenimento dell'oscillazione circadiana. In questi moscerini la proteina PER è incapace di rientrare nel nucleo della cellula (Vosshall et al., 1994), e quindi non può regolare la propria trascrizione come si ritiene che avvenga in condizioni normali. Queste osservazioni hanno consentito di ipotizzare che l'interazione fra le proteine PER e TlM produca un dimero che permette loro di rientrare nel nucleo della cellula e quindi di regolare la propria trascrizione. Tale processo è considerato un elemento importante nel meccanismo che genera l'oscillazione circadiana (fig. 8).
Altri organismi, compreso l'uomo (Tei et al., 1997), possiedono un gene ortologo a per di Drosophila (derivano cioè da un gene ancestrale comune), suggerendo che il meccanismo molecolare capace di generare l'oscillazione circadiana si è conservato durante l'evoluzione fino ai mammiferi. Anche nei mammiferi (criceti e topi) sono state identificate due mutazioni che alterano i ritmi circadiani (Ralph e Menaker, 1988; Vitatema et al., 1994) e, nel caso dei topi, è stato clonato il gene responsabile, denominato Clock (King et al., 1997). l meccanismi molecolari responsabili dell'oscillazione circadiana sono attualmente oggetto di intenso studio e si spera di chiarirne il funzionamento nei prossimi dieci anni.
Significato adattativo dei ritmi circadiani
Il significato adattativo dei ritmi circadiani deve essere rilevante, considerata la loro diffusione negli organismi e la loro presenza in molti livelli dell'organizzazione biologica. Tuttavia, la loro ampia diffusione e la diversità delle funzioni che li regolano rendono difficile assegnare loro un ruolo generale. Per questo motivo, la discussione del valore adattativo dei ritmi circadiani riguarderà alcuni casi specifici in cui tale significato è stato realmente dimostrato. Per dare un ordine alla discussione separiamo il significato adattativo dei ritmi circadiani in due ampie categorie.
Mantenimento della sincronia fra l'organismo e il mondo esterno
Il più ovvio dei significati adattativi dei ritmi circadiani è la sincronizzazione delle funzioni ritmiche dell'organismo con i cicli ambientali (il ciclo della luce, della temperatura e dell'umidità) e con i cicli degli altri organismi che occupano la stessa nicchia ecologica. La presenza di un orologio intemo sincronizzato con l'ambiente esterno permette all'organismo di predire in modo accurato gli eventi che si verificheranno nel proprio ambiente, poiché la relazione di fase tra le oscillazioni interne e quelle esterne è regolata dalla selezione naturale, e ciò conferisce un notevole vantaggio. Questo vantaggio non sarebbe disponibile se l'ambiente si limitasse a costringere gli eventi ad avvenire.
l processi biologici sincronizzati, che richiedono molto tempo per essere portati a termine, possono essere messi in fase con i cicli ambientali in modo tale che il risultato finale del processo avvenga in un tempo critico più avanzato. Se, per esempio, la sopravvivenza di un animale che ha un ampio home range (area che l'animale occupa, ma che non difende) dipende dal suo ritorno alla tana di giorno, esso deve iniziare il suo viaggio di ritorno prima di percepire la luce dell'alba. Se ciò non accadesse, l'animale si potrebbe trovare lontano dalla sua tana al sopraggiungere delle condizioni sfavorevoli e, quindi, in una situazione pericolosa per la propria sopravvivenza.
È importante ricordare che anche se gli organismi usano di solito, come stimolo sincronizzatore, la caratteristica più stabile dell'ambiente, il significato adattativo di un particolare ritmo può essere ricercato nella sua relazione con altre caratteristiche ambientali. Per esempio, la variazione giornaliera del tasso di umidità nell'ambiente può non essere sufficientemente regolare per essere utilizzata come segnale sincronizzatore, ma al tempo stesso l'organismo può assumere una relazione di fase adattativa con essa sincronizzando si con il ciclo di illuminazione, che a sua volta può avere una relazione di fase statisticamente affidabile con il ciclo dell 'umidità.
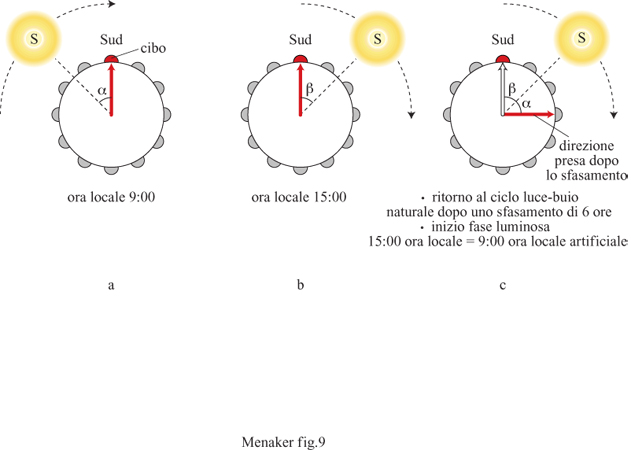
Numerosi studi hanno evidenziato l'utilizzo, da parte degli organismi, dei ritmi circadiani come 'orologi consultati continuamente'. Le api visitano ogni fiore in un'ora diversa del giorno in modo da arrivare nel momento in cui il fiore secerne il nettare. Sia le api sia i fiori sono sincronizzati dal ciclo lucebuio, e le api consultano il proprio orologio interno durante tutto il giorno, presumibilmente perché hanno imparato quali sono i fiori che secernono il nettare nelle diverse ore del giorno. La sincronizzazione di questi eventi riveste un'importanza adattativa notevole per entrambi gli organismi, poiché permette alle api di ottenere la massima quantità di cibo con il minimo sforzo e alle piante, che si servono delle api come mezzo per l'impollinazione, di concentrare la produzione del nettare, operazione metabolicamente dispendiosa, solo in determinate ore del giorno, con un notevole risparmio di energie. Molte specie di uccelli compiono navigazioni complesse che dipendono da orologi interni consultati durante tutto il giorno, così come la navigazione umana dipende da cronometri accurati. Un uccello sincronizzato in laboratorio con un ciclo luce-buio che ha una fase diversa rispetto al ciclo luce-buio presente in natura (fig. 9), una volta liberato, commetterà un errore nella direzione di navigazione proporzionale allo sfasamento fra il ciclo artificiale e quello naturale (v. i saggi di F. Papi e di E. Gwinner, in questo volume).
Anche la sincronizzazione dei cicli annuali di riproduzione con i cicli annuali dell'ambiente esterno è un esempio del valore adattativo della ritmicità biologica. La risposta fisiologica che sia le piante sia gli animali mostrano al variare delle ore di luce (cioè del fotoperiodo) è conosciuta con il nome difotoperiodismo: esso dipende dalla capacità dell'organismo di misurare il tempo, cioè di misurare la durata del periodo di luce in modo da distinguere i giorni con molte ore di luce da quelli con poche ore di luce. Una serie di studi ha dimostrato che la misurazione del fotoperiodo è ottenuta per mezzo di un ritmo circadiano di sensibilità a stimoli indotti dalla luce. La fioritura nelle piante, la diapausa negli insetti, la crescita delle gonadi negli uccelli e in alcuni mammiferi rappresentano risposte fotoperiodiche indotte da uno stimolo luminoso che si verifica in determinati momenti del ciclo circadiano interno dell'organismo. Lo stesso stimolo luminoso fornito in un momento diverso del ciclo circadiano non produce alcun effetto su questi processi. Il ritmo circadiano di sensibilità al segnale luminoso è sincronizzato dal ciclo luce-buio con una relazione di fase tale che nei giorni con molte ore di luce lo stimolo luminoso cadrà sulla parte della curva del ciclo che induce una risposta, mentre nei giorni caratterizzati da poche ore di luce il segnale luminoso cadrà nella parte insensibile.
Una differenza di fase tra le attività consente alle specie che occupano la stessa nicchia ecologica di ridurre la competizione per le stesse risorse energetiche. Per esempio, molte specie di pipistrelli e di uccelli sono insettivori e occupano l'ambiente aereo, ma nella maggior parte dei casi esse non competono per il cibo poiché le loro attività predatorie mantengono una sincronizzazione diversa con il ciclo della luce. Nelle foreste tropicali, molte specie di rane emettono il richiamo di accoppiamento principalmente durante la notte, ma ogni specie ha una definita nicchia temporale. Tale separazione temporale può essere raggiunta solo se l'organismo possiede un sistema capace di misurare il tempo in maniera accurata.
Mantenimento della sincronia tra i ritmi nel medesimo organismo
Come già menzionato, in un organismo, molti processi fisiologici sono ritmici e mantengono una precisa relazione di fase tra loro. Ciò è ovviamente di cruciale importanza e certamente di notevole valore adattativo per l'organismo stesso. l substrati per le reazioni biochimiche, le risorse energetiche, e gli enzimi specifici devono essere presenti al momento giusto e al posto giusto per consentire l'integrazione di organi, cellule e tessuti in un organismo funzionalmente efficiente. Tale integrazione è possibile solo se i processi che variano ritmicamente vengono controllati in modo tale da essere in fase tra loro. Sebbene sia un campo di indagine recente, vi sono già prove significative che documentano il valore adattativo dell'organizzazione temporale interna. Concludendo, possiamo affermare che, probabilmente, la sincronizzazione di un certo comportamento con gli eventi nell'ambiente esterno ha molti valori adattativi e, più in generale, che il significato adattativo dei ritmi circadiani, e degli altri ritmi, va ricercato nel controllo che essi esercitano sull'organizzazione biologica e temporale interna di tutti gli organismi.
Bibliografia citata
CAHILL, G.M. BESHARSE, J.C. (1993) Circadian clock function localized in Xenopus retinal photoreceptors. Neuron, 10, 573-577.
DUNLAP, J.C. (1996) Genetic and molecular analysis of circadian rhythms. Annu. Rev. Genet., 30, 579-601.
GIEBULTOWICZ, J.M., HEGE, D.M. (1997) Circadian clock in Malpighian tubules. Nature, 386, 664.
HOFFMANN, K. (1954) Versuche zu der im Richtungsfinden der Vogel enthaltenen Zeitschatzung. Z. Tierpsychol., 11, 453-475.
KING, D.P. et al. (1997) Positional cloning ofthe mouse circadian Clock gene. CelI, 89, 641-653.
KONOPKA, R.J., BENZER, S. (1971) Clock mutants of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA, 68, 2112-2116.
MENAKER, M. (1968) Extraretinallight perception in the sparrow, I: entrainment of the biological clock. Proc. Natl. Acad. Sci. USA, 59, 414-421.
MICHEL, S., GEUSZ, M.E., ZARITSKY, U., BLOCK, G.D. (1993) Circadian rhythm in membrane conductance express in isolated neurons. Science, 259, 239-241.
PITTENDRIGH, C.S. (1954) On the temperature independence in the clock-system controlling emergence in Drosophila. Proc. Natl. Acad. Sci. USA, 40, 1118-1129.
PITTENDRIGH, C.S., DAAN, S. (1976) A functional analysis of circadian pacemakers in nocturnal rodents. IV. Entrainment: pacemaker as clock. J. Comp. Physiol., 106, 291-331.
PLAUTZ J.D., KANEKO, M., HALL, J.C., KAY, S.A. (1997) Independent photoreceptive circadian clocks throughout Drosophila. Science, 278, 1632-1635.
RALPH, M.R., MENAKER, M. (1988) A mutation of the circadian system in golden hamsters. Science, 241, 1225-1227.
SEHGAL, A., PRICE, J., MAN, B., YOUNG, M. (1994) Loss of circadian behavioral rhythms and per oscillations in the Drosophila mutant timeless. Science, 263, 1603-1606.
STEPHAN, F.K., ZUCKER, I. (1972) Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc. Natl. Acad. Sci. USA, 69, 1583-1586.
TEI, H., OKAMURA, H., SHIGEYOSHI, Y., FUKUHARA, C., OZAWA, R., HIROSE, M., SAKAKI, Y. (1997) Circadian oscillation of a manunalian homologue of the Drosophila period gene. Nature, 389, 512-516.
TOSINI, G., MENAKER, M. (1996) Circadian rhythms in cultured manunalian retina. Science, 272, 419-421.
TOSINI, G., MENAKER, M. (1998) Multioscillatory circadian organization in a vertebrate, Iguana iguana. J. Neurosci., 18, 1105-1114.
VITATERNA, M.H., KING, D.P., CHANG, A.M., KORNHAUSER, J.M., LOWREY, P.L., McDONALD, J., DOVE, W.F., PINTO L.H., TUREK, F.W., TAKAHASHI, J.S. (1994) Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior. Science, 264, 719-725.
VOSSHALL, L.B., PRICE, J.L., SEHGAL, A., SAEZ L., YOUNG, M.W. (1994) Block in nuclear localization of period protein by a second clock mutation, timeless. Science, 263, 1606-1609.
WELSH, D.K., LOGOTHETIS, D.E., MEISTER, M., REPPERT, S.M. (1995) IndividuaI neurons dissociated from rats suprachiasmatic nucleus express independently phased firing rhythms. Neuron, 14, 697-706.
Bibliografia generale
ASCHOFF, J. Handbook of behavioral neurobiology. Biological rhythms. New York, Plenum Press, 1981.
KLEIN, D.C., MOORE, R.Y., REPPERT, S.M. Suprachiasmatic nucleus: the mind's clock. New York, New York University Press, 1991.
MOORE-EDE, M.C., SULZMAN, F.M., FULLER, C.A. The clocks that time uso Physiology of the circadian timing system. Cambridge, Mass., Harvard University Press, 1982.
PITTENDRIGH, C.S. (1993) Temporal organization: reflection of a darwinian clock-watcher. Annu. Rev. Physiol., 55, 16-54.
WETTERBERG, L. Light and biological rhythms in man. New York, Pergamon Press, 1995.