
Paleontologia dell'occhio
Paleontologia dell'occhio
I trilobiti, un gruppo di artropodi marini comparsi durante l'esplosione di forme di vita avanzata che avvenne nel Cambriano, hanno lasciato un'impressionante testimonianza fossile dei primi organi visivi. La struttura della superficie visiva di questi occhi antichi si è conservata nei suoi tratti salienti, in quanto le lenti degli occhi composti dei trilobiti erano costituite di calcite cristallina e la struttura nella quale gli occhi erano racchiusi si è anch'essa mineralizzata. È possibile individuare nel corso dell'evoluzione dei trilobiti alcuni stadi che hanno portato a un miglioramento delle funzioni visive, culminante nello sviluppo di lenti capaci di correggere l'aberrazione sferica. Sebbene non si sia conservato nulla delle strutture interne degli occhi dei trilobiti, l'ottimizzazione delle funzioni visive dei loro apparati diottrici è una testimonianza eloquente dell'aumento parallelo della complessità dei fotorecettori e del sistema nervoso in risposta alla pressione selettiva. Sebbene i trilobiti si siano estinti, alcune forme attuali di vita hanno sviluppato perfetti apparati diottrici del tutto simili a quelli che si erano evoluti in questo gruppo di animali.
L'occhio e i requisiti per la visione
Come si è potuto apprendere dai resti fossili, la vista, indubbiamente una delle più evolute ed essenziali funzioni della vita animale, si è sviluppata molto presto. l più antichi occhi conosciuti appartengono ai trilobiti, un gruppo di artropodi che fiorì nei mari a partire dal Cambriano inferiore fino al Permiano superiore, lungo un arco di tempo di circa 325 milioni di anni. Gli occhi di questi animali sono occhi composti, come quelli degli insetti e dei crostacei, e compaiono già negli esemplari più antichi. Sono presenti in paia, posizionati lateralmente lungo la testa dell'animale. Prima di procedere nella descrizione di ciò che si è trovato e di valutare che cosa manchi nei resti fossili dei trilobiti, è importante presentare una panoramica sulla visione così come noi la conosciamo dalle forme di vita attualmente esistenti.
L'occhio composto rappresenta una delle due strategie per la visione; l'altra, che si è sviluppata nei cefalopodi e nei vertebrati altrettanto presto nella storia della vita, assomiglia alla struttura di una macchina fotografica. Sebbene richiedano due tipi di occhio strutturalmente diversi, le due strategie si basano su meccanismi del tutto simili.
l requisiti basilari per la visione consistono nella capacità di percepire e reagire alla luce, di localizzare la fonte da cui questa proviene e avvertire il movimento, di percepire forma e profondità, cioè la distanza relativa degli oggetti, di distinguere i colori e, in molti casi, la polarizzazione della luce. Ai livelli più bassi di complessità, è necessario, perché ciò si verifichi, che vi sia un sensore o un rilevatore di luce, cioè un qualche dispositivo che risponda allo stimolo luminoso. Inoltre, devono essere presenti congegni ottici direzionali, o apparati diottrici, in grado di trasferire uno schema luminoso, l'oggetto che si vede, al sensore della luce e, infine, un 'cervello', cioè una struttura neurale in grado di ricevere e coordinare il segnale ricevuto e di ricomporre come immagine l'oggetto visto. A un livello più alto, dove viene richiesta una percezione più precisa e versatile, si rende necessario un ulteriore perfezionamento di tali strutture. Per poter percepire il movimento, gli occhi devono essere in grado di seguire l'evoluzione dello stimolo visivo durante il tempo. Per poter percepire la profondità, il sistema visivo deve avere la capacità di comparare le immagini ottenute da diversi punti di vista. Per poter distinguere il colore e la polarizzazione della luce, gli occhi devono avere delle strutture sensoriali ancora più complesse.
Strutture fondamentali dell'occhio
l fotorecettori, dispositivi sensibili alla luce, simili in tutti i tipi di animali e situati generalmente nella parte più interna dell'occhio, agiscono in maniera analoga alla pellicola di una macchina fotografica: reagiscono chimicamente alla luce. In tutti gli invertebrati hanno la forma di un'ordinata rete di microtubuli impilati che, nei vertebrati, vengono sostituiti da dischi. Queste strutture contengono dei pigmenti, molecole di rodopsina, che vengono alterati chimicamente dalla luce. La reazione chimica all'interno della rodopsina genera una serie di impulsi elettrici che sono convogliati al cervello da un nervo ottico.
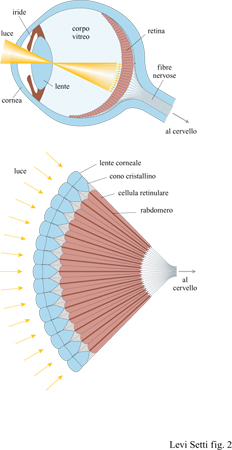
In tutti i tipi di occhio, il legame tra il fotorecettore e il mondo esterno è costituito da un'interfaccia ottica, la struttura trasparente che raccoglie e concentra la luce dal mondo esterno, cioè l'oggetto, e trasporta il disegno luminoso, cioè l'immagine, ai fotorecettori (fig. 1).
Qualunque sia la struttura dell'interfaccia ottica, una singola lente o un insieme di lenti, l'immagine deve essere frammentata in più elementi puntiformi in modo che, nella rete dei fotorecettori, ogni singola unità possa percepire una parte di ciascuna immagine. Qui risiede il denominatore comune di tutti gli schemi visivi: l'immagine deve essere campionata in piccoli frammenti dalla rete dei fotorecettori e, fino a un certo punto, quanto più è fine la rete dei fotorecettori, tanto più è grande la capacità di distinguere i dettagli. Vi è un limite pratico a quanto possano essere piccoli gli elementi di questa rete, in quanto l'apertura dell'interfaccia ottica causa fenomeni di diffrazione che, comunque, confondono un po' l'immagine. La fotografia e la televisione sono due esempi familiari di tale meccanismo, che sembra essere fondamentale per la ricostruzione di ogni immagine.
Due approcci funzionali alla visione
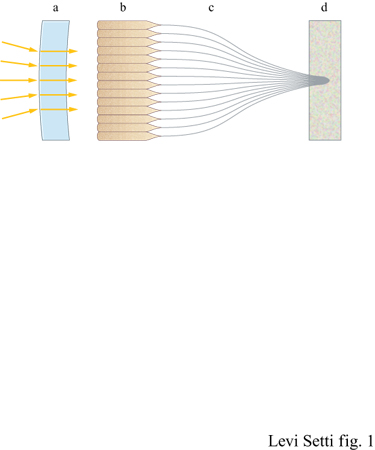
l due sistemi visivi, quello dell' occhio 'a macchina fotografica' e quello dell' occhio composto, possono essere considerati come due soluzioni equivalenti al problema di campionare l'immagine in elementi discreti (fig. 2).
Nell'occhio 'a macchina fotografica', i fotorecettori sono organizzati in una fitta schiera che copre la superficie più interna della cavità oculare, la retina. Le cellule sensibili alla luce sono chiamate coni e bastoncelli. Nella sua forma più semplice, la retina è illuminata attraverso un piccolo foro, mentre nelle forme più avanzate, quali l'occhio umano, il foro è sostituito da una lente focale provvista di un'apertura variabile che mette a fuoco l'immagine e regola il livello di illuminazione per ottimizzare la risposta della retina. La lente proietta un'immagine continua ma i fotorecettori, che nella retina umana sono costituiti da circa 30 milioni di coni e 100 milioni di bastoncelli, scindono l'immagine in numerosi elementi discreti puntiformi e la campionano. La corteccia visiva, che fa parte del sistema nervoso, ricostruisce poi i numerosi stimoli discreti dando luogo alla percezione dell'immagine finale. In questo tipo di occhio la messa a fuoco è realizzata da una lente a distanza focale variabile oppure da una lente mobile o ancora, in pochi casi, da una retina mobile. Altre strutture dell'occhio, inclusi la cornea e il corpo vitreo che riempie la cavità oculare, sono accessori dell'interfaccia ottica. Normalmente servono due occhi di questo tipo per misurare le distanze e per ottenere una visione stereo scopica, sebbene i movimenti oculari unitamente a una memoria dell'immagine possano a volte sostituire la visione stereoscopica. In vari tipi di occhi animali viene realizzata una scansione del campo visivo per consentire il campionamento dell'immagine. Un esempio estremo di questo tipo di occhio si trova in Copilia quadrata, un microscopico organismo marino nel quale un singolo fotorecettore a tutto campo effettua una scansione delle immagini proiettate da un paio di lenti. Dalla sorprendente complessità della retina, che compie una sorta di lettura parallela delle informazioni visive in arrivo, al recettore a scansione, che compie una lettura seriale o sequenziale, tutti gli occhi 'a macchina fotografica' svolgono, di fatto, lo stesso tipo di funzione.
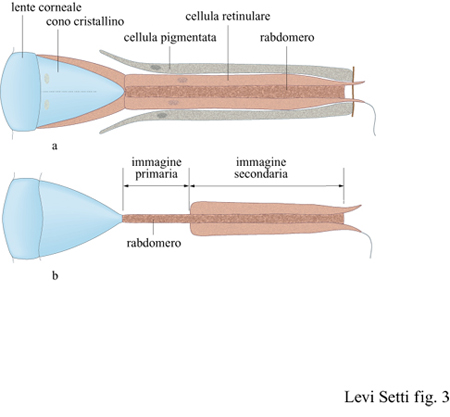
Nel secondo schema per la visione, l'occhio composto, è la stessa interfaccia ottica, e non la retina, a frammentare l'immagine in elementi puntiformi. Dato l'incredibile numero di specie animali che lo hanno adottato (basti pensare agli insetti), non vi sono dubbi sul successo di questo sistema. La superficie visiva di questo tipo di occhi è composta da una schiera di lenti separate o da altri dispositivi capaci di raccogliere la luce e filtrare i raggi, con ciascun elemento della superficie visiva connesso a un proprio fotorecettore. I moduli di cui sono fatti gli occhi composti si chiamano ommatidi: ciascuno di essi ha la forma di un cilindro trasparente che termina all'estremità che si affaccia all'esterno, la parte distale, con una lente cornea, con un cono cristallino o con una combinazione dei due (fig. 3).
Sull'altra estremità, quella prossimale o interna, di ciascun ommatidio vi è un fotorecettore, o rabdomero, collegato mediante un neurone al sistema nervoso e infine al 'cervello'. Con questo tipo di organizzazione, mediante un fascio di neuroni, dalla rete dei singoli fotorecettori emerge una lettura parallela degli stimoli elettrici. Il mosaico di elementi puntiformi che provengono dall'interfaccia ottica deve poi essere coordinato da un qualche tipo di cervello (la struttura centrale) per produrre la percezione dell'immagine. La rete dei recettori è relativamente rozza se paragonata a quella della retina degli occhi 'a macchina fotografica', ma la funzionalità di questo sistema è essenzialmente la stessa. La principale differenza consiste nella struttura dell'interfaccia ottica e nei fenomeni ottici che tale struttura può originare. Generalmente, un occhio composto ha un campo visivo insolitamente ampio, in quanto gli ommatidi sono tutti disposti in maniera da puntare in direzioni leggermente differenti. In realtà il campo visivo di una coppia di occhi composti può sovrapporsi nella zona frontale e dorsale, rendendo possibile la visione stereoscopica lungo l'intero ambiente circostante. In linea di principio, una parziale visione stereoscopica può anche essere disponibile separatamente per ogni singolo occhio composto, nella regione in cui il campo visivo degli ommatidi vicini si sovrappone. Un vantaggio ulteriore dell'occhio composto, raramente riconosciuto, sta nell'intrinseca ampiezza della profondità di campo. Dal momento che questo tipo di occhio rappresenta uno strumento ottico che, con buona approssimazione, può essere considerato come una fila bidimensionale di piccole aperture stenopeiche, esso non ha bisogno della messa a fuoco o dell' accomodazione visiva per ottenere contemporaneamente immagini relativamente precise di oggetti sia vicini sia lontani.
La testimonianza fossile degli occhi dei trilobiti
Forse non è una coincidenza che l'approccio modulare alla costruzione dell'occhio sia comparso per la prima volta negli artropodi, e in particolare nei trilobiti, in quanto queste creature mostrano un notevole grado di modularità, cioè un'omologia seriale, nella costruzione del loro intero corpo. Come l'occhio composto, i trilobiti erano costituiti da una sequenza di segmenti virtualmente identici, ognuno provvisto originariamente di tutte le strutture necessarie per vivere. Gli occhi di questi animali sono spesso ben conservati grazie al fatto che le lenti, come il resto dell'esoscheletro, erano costituite da calcite, materiale dotato di una notevole capacità di conservazione, specialmente nel calcare. Sebbene in qualche circostanza le lenti siano andate incontro a una ricristallizzazione per diagenesi dopo la morte, in molti casi la struttura e la mineralogia originarie delle lenti si sono potute conservare praticamente intatte. Ciò è però vero solo per le lenti e per le regioni adiacenti della cuticola che costituiscono l' interfaccia ottica, mentre le parti morbide che giacciono al di sotto della superficie della lente decadono in fretta, senza lasciare traccia, poco dopo la morte. Anche così, si ha una mole notevole di informazioni circa la struttura, la crescita e l'evoluzione di questi occhi (Clarkson, 1975; 1979) e circa la funzione e l'ottica del sistema visivo dei trilobiti. Nei trilobiti si distinguono due tipi principali di occhi, indicati come oloeroali e sehizoeroali, ed è stato proposto un possibile terzo tipo, l'abatocroale.
Occhi olocroali

Nella maggior parte dei trilobiti gli occhi sono olocroali, cioè con piccole e numerose lenti di calcite, in diretto contatto le une con le altre e fittamente assemblate (fig. 4); queste lenti sono di solito tonde o esagonali.
Un unico sottile strato di calcite copre la fila di lenti, che costituisce una sorta di cornea continua con la superficie esterna della cuticola. Sotto la superficie visiva si trova spesso uno zoccolo stretto, il solco oculare, che può essere provvisto di fossette sensoriali. Direttamente sopra la superficie visiva si estende parte della sutura facciale, una linea di fragilità che permette al vecchio esoscheletro di rompersi in varie parti durante le mute. Dall'altro lato della sutura si trova il lobo palpebrale, esso stesso parte della regione centrale della testa, il eranidium, che forma una sorta di 'palpebra superiore' con una protuberanza che si curva esternamente.
Gli occhi olocroali si trovano nella famiglia Olenellidi, i primi trilobiti del Cambriano, e, sebbene la superficie visiva non si sia in molti casi conservata, sono estremamente abbondanti i resti di occhi olocroali di vario tipo databili dal Cambriano inferiore alla fine del Permiano. Nelle prime forme, eccetto che per alcuni individui giovani, la superficie visiva veniva rilasciata durante la muta o con la morte e soltanto il lobo palpebrale e il solco oculare definiscono il profilo dell'occhio.
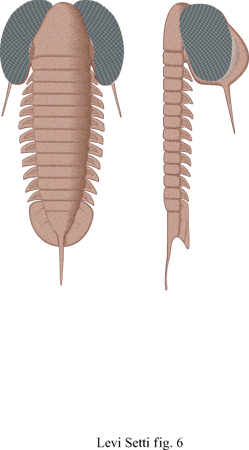
Verso la fine del Cambriano e durante l'epoca postcambriana, la superficie visiva si fonde con il solco oculare e si conserva come parte dell'esoscheletro mineralizzato. Molti occhi olocroali sono reniformi e sessili, con al massimo poche centinaia di lenti; ma durante l'Ordoviciano compaiono parecchi tipi differenti di occhi. La nicchia pelagica viene colonizzata, probabilmente per la prima volta, e nei trilobiti in grado di nuotare, come quelli del genere Opipeuter (fig. 6) e Cyclopyge, gli occhi diventano innnensi e in alcuni casi fusi anteriormente. Nella maggior parte dei trilobiti del periodo successivo all'Ordoviciano gli occhi, se del tipo olocroale, sono generalmente reniformi con lenti ben sviluppate e questo tipo di organizzazione conservativa persiste fino alla fine del Permiano.
Durante la crescita dei trilobiti a partire dai primi stadi di vita, gli occhi compaiono sul margine anterolaterale; con le mute successive essi migrano verso l'interno e dorsalmente, portando con sé la sutura facciale. Le lenti sono ubicate in una zona generativa che giace sotto la sutura facciale e nuove lenti sono aggiunte continuamente sotto quelle preesistenti. L'occhio maturo presenta una superficie curva, e il campo visivo angolare sotteso dipende dall'entità della curvatura. Se la superficie visiva è solo leggermente incurvata nel piano verticale, il campo visivo sotteso può avere un'ampiezza latitudinale di non più di 30° dall'orizzontale. Un occhio fortemente incurvato, d'altra parte, sottenderà un campo visivo panoramico, che può estendersi da sotto l'orizzontale fino a 90° sopra. Se tali occhi presentano una curvatura pronunciata anche nel piano orizzontale, i campi visivi dei due occhi possono sovrapporsi anteriormente e posteriormente. L'organizzazione delle lenti in questo tipo di occhi è notevole da molti punti di vista (Clarkson, 1975). Su scala piccola, le lenti sono organizzate in disegni esagonali, in maniera molto simile alle celle di un alveare. Su scala più grande, emerge un disegno di spirali logaritmiche che si intersecano, simili all'organizzazione dei flosculi in un girasole gigante.

Le proprietà delle spirali logaritmiche o equiangolari hanno affascinato i naturalisti e i poeti per secoli (D'Arcy Thompson, 1942). In natura si verifica la loro formazione ogni volta che un processo di crescita, cioè un'espansione lineare, si combina con la necessità di conservare la circolarità delle forme, cioè la sinnnetria rotazionale. La crescita degli occhi dei trilobiti, in particolare nelle varietà con occhi olocroali, è stata dominata da questo tema. Ogni lente è un cristallo singolo di calcite, composto da lamelle radiali disposte intorno al proprio asse cristallografico c (fig. 8); presumibilmente la lente cresceva per aggiunta di nuova calcite all'estremità esterna di ogni lamella.
L'asse cristallografico c è orientato lungo l'asse della lente, perpendicolarmente alla superficie visiva, rendendo minimi in tal modo i fenomeni di birifrangenza. Nei trilobiti con guscio sottile le lenti sono generalmente sottili e biconvesse, mentre nelle forme che presentano un guscio spesso le lenti formano delle colonne prismatiche allungate, con terminazioni semicircolari. La natura e l'orientamento dei cristalli di ciascun elemento sono evidenti nella sezione sottile della figura (fig. 9), tagliata lungo la superficie visiva di una delle forme più recenti, dove sono visibili i piani di sfaldamento che attraversano i prismi con un angolo costante rispetto all'asse di ciascun elemento. Il tracciamento dei raggi mostra che le lenti olocroali, qualunque sia la loro forma, mettono a fuoco circa alla stessa distanza relativa al di là della lente; ciò che è importante dal punto di vista ottico è la convessità relativa della superficie della lente. Circa la struttura interna degli occhi non si sa nulla, ma probabilmente questi erano provvisti sotto ogni lente di unità simili all'ommatidio e funzionavano come gli occhi dei crostacei (Fordyce e Cronin, 1993).
Occhi schizocroali
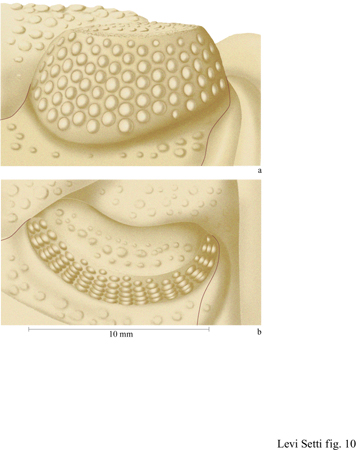
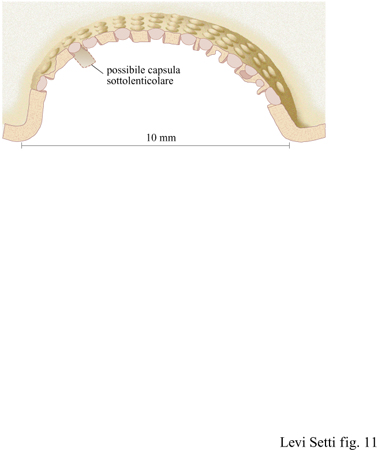
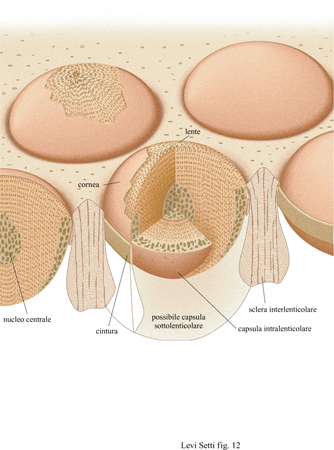
Solo un gruppo di trilobiti aveva occhi schizocroali: il sottordine Phacopina, apparso nell'Ordoviciano inferiore ed estintosi alla fine del Devoniano. Questi occhi sono unici; non vi è nulla di simile nel regno animale. Generalmente grandi, hanno una superficie visiva ricurva, le lenti sono notevolmente ampie, fortemente biconvesse e separate l'una dall' altra da una cuticola, uguale a quella del restante esoscheletro (fig. 10). Queste lenti sono organizzate normalmente in un sistema regolare di fitti assemblaggi a forma di esagono orientati in modo tale da formare delle righe rettilinee verticali, le righe dorsoventrali, e sono generalmente molto meno numerose di quelle degli occhi olocroali. Solo quando gli occhi schizocroali vengono osservati in sezione diventano più evidenti le differenze rispetto a quelli olocroali. Ogni lente biconvessa è posizionata all'estremità esterna di una cavità cilindrica infossata e sulla sua superficie esterna vi è una sottile cornea di calcite. Questa si immerge lungo la cuticola e in rari casi si può vedere che continua in un cilindro affusolato, che si stringe nella parte inferiore (figg. 11, 12). Le lenti stesse sono costituite di due elementi, sono cioè lenti doppie.

Gli occhi schizocroali crescono mediante un sistema di sostituzioni delle lenti, analogamente agli occhi olocroali, con spirali logaritmiche che si intersecano; al crescere dell'occhio la superficie visiva si espande verso il basso. In ognuna delle file dorsoventrali, tuttavia, le lenti sono graduate in dimensione, divenendo via via più larghe verso la base dell'occhio e solitamente verso la fronte e il dorso. Se le lenti fossero state tutte della stessa dimensione, come succede per gli occhi olocroali, esse sarebbero state disposte in righe parallele, ma l'allargamento verso il basso avrebbe creato dello spazio in eccesso. Su una superficie in espansione è geometricamente impossibile impacchettare in sequenza regolare lenti aventi tutte la stessa dimensione ed è probabile che si verifichino alcune anomalie durante la crescita. Questo è comune negli occhi olocroali (v. figura 4), ma si verifica anche nel caso delle prime forme di trilobiti che presentano occhi schizocroali, quelli appartenenti al genere Ormathops dell'Ordoviciano inferiore (fig. 14) dove, su una superficie per il resto regolare, in alcune aree sono evidenti lacune o irregolarità nella disposizione, oppure si verifica che le lenti collocate in righe parallele giacciono troncate contro altri blocchi. L'aumento verso il basso della spaziatura tra le lenti, in molte specie con occhi schizocroali, è una soluzione semplice al problema della compattazione delle lenti.
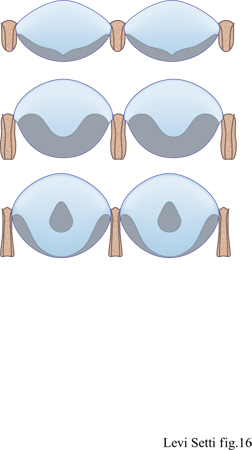
Come per gli occhi olocroali, le lenti schizocroali sono costituite da lamelle, che si irradiano dall'asse cristallografico c. In sezioni normali rispetto al piano principale, è evidente come ogni lente sia in realtà un sistema di due lenti, in cui una capsula intralenticolare prossimale si connette con l'unità disposta più in alto lungo la superficie ondulata (Clarkson, 1968). Dall'orientamento dei piani di sfaldamento nelle unità superiori, è anche evidente che l'asse c del cristallo di calcite che componeva le lenti era orientato lungo l'asse della lente (Clarkson e LeviSetti, 1975; Levi-Setti, 1993). La struttura di queste lenti doppie è stata descritta per la prima volta quasi cento anni fa da G. Lindstrom (1901), un eminente paleontologo svedese, che pensava che tali strutture potessero essere un artefatto dovuto a processi diagenetici. La loro natura primaria, tuttavia, è stata interpretata (Levi-Setti, 1975; Clarkson e Levi-Setti, 1975) in relazione alla funzione di correzione dell'aberrazione sferica dovuta allo spessore delle lenti e tale descrizione, che sarà discussa successivamente in maniera più approfondita, è stata confermata ampiamente da alcuni studi recenti (Horváth, 1989a; Horváth e Clarkson, 1993). Per quanto riguarda l'organizzazione delle lenti nei trilobiti della famiglia Facopidi, sono presenti almeno tre schemi anche se, molto probabilmente, il loro funzionamento si basa sugli stessi principi ottici. Le lenti presenti in alcuni dei primi Facopidi, come quelli del genere Dalmanitina dell'Ordoviciano, hanno una prima superficie apparentemente a forma di iperbole e la capsula piuttosto appiattita, con una depressione centrale; quelli del genere Crozonaspis a loro contemporanei, d'altra parte, avevano lenti più convesse e la capsula era più spessa, con una superficie superiore ondulata (fig. 16). Nei Facopidi del Devoniano medio la capsula era fatta di calcite compatta e si assottigliava verso l'esterno, fino a scomparire nella zona prossimale. Sembra esservi, tuttavia, un nucleo denso centrale addizionale, la cui funzione attualmente è sconosciuta (Miller e Clarkson, 1980). Dopo la muta, le lenti si rigeneravano; nei Facopidi si è potuto dedurre come ciò avvenisse osservando una serie di individui caratterizzati da un guscio sottile, morti durante la muta o immediatamente dopo. Nei primissimi stadi postecdisi la cuticola era ancora flessibile e ogni lente aveva la forma di un piccolo cono sospeso al centro della cornea. Successivamente la lente si allargava verso la periferia, ispessendosi e assumendo una forma tondeggiante e infine acquisendo la caratteristica superficie ondulata. La capsula e il nucleo si differenziavano per ultimi. G. Horváth e E.N.K. Clarkson (1993) hanno dimostrato come queste lenti fossero ottimizzate per la correzione dell'aberrazione sferica in tutti gli stadi dello sviluppo, come sarà discusso ulteriormente più avanti.
l primi occhi schizocroali derivarono quasi certamente da antenati con occhi olocroali per pedomorfosi, il mantenimento di caratteri giovanili di un antenato in un discendente adulto. Nei pochi esemplari con occhi olocroali di cui sono noti i primi stadi di sviluppo, quali Olenus del Cambriano e Paladin del Carbonifero (v. figura 4c), le lenti erano relativamente larghe e separate l'una dall' altra da una cuticola; questi sono, in effetti, miniature di occhi schizocroali (Clarkson e Zhang, 1991). A mano a mano che l'occhio si sviluppa, scompaiono le cuticole interstiziali e l'occhio diventa olocroale. Il mantenimento di questa struttura giovanile nello stadio adulto, per pedomorfosi, sarebbe il primo e più importante passo nell'origine degli occhi schizocroali. Nelle prime specie di Facopidi come Ormathops, l'uguaglianza nella dimensione delle lenti derivava probabilmente da precursori con occhi olocroali; nelle forme più tardive, la regolarità nell 'impacchettamento, presumibilmente importante per il corretto funzionamento dell'occhio, era ottenuta mediante un semplice incremento nel diametro delle lenti. Generalmente, si ritiene (Campbell, 1975; Clarkson, 1979) che negli occhi schizocroali una capsula ocellare relativamente piccola, piuttosto che un ommatidio, giacesse sotto ogni lente (v. figura 12). Si è tentati di ipotizzare che questo tipo di occhio possa rappresentare un aggregato di semplici oculari in uno stadio di transizione dall'occhio composto all'occhio 'a macchina fotografica'. Tale assunzione comporta (Clarkson e Levi-Setti, 1975; Fordyce e Cronin, 1989) che dietro ogni lente vi sia una sottile retina, piuttosto che un singolo fotorecettore simile a un ommatidio, composto di un piccolo insieme di cellule retiniche, come negli occhi olocroali e in quelli degli artropodi attuali. Questo può spiegare la separazione tra le lenti, e la necessità di formare un'immagine perfetta nelle lenti degli occhi schizocroali. In effetti, se ci fosse stato un singolo rabdomero nella capsula al di sotto della lente, allora la risoluzione spazi aie sarebbe stata estremamente ridotta a causa del numero esiguo di elementi puntiformi generati dalle lenti spaziate, presenti in questo tipo di occhi. Un tale passo all'indietro avrebbe sfidato la sofisticata ottimizzazione delle funzioni ottiche dimostrata dalle strutture doppie degli occhi schizocroali, le quali possono essere sfruttate appieno solo mediante il campionamento dell'immagine da parte di una schiera di fotorecettori ad alta risoluzione del tipo della retina.
Occhi abatocroali
Alcuni autori sostengono che gli occhi degli Eodiscidi del Cambriano medio e inferiore costituiscono un terzo tipo di occhio, intermedio tra quelli olocroale e schizocroale, l' occhio abatocroale. Questi occhi, descritti per la prima volta da P.A. Jell (1975) in Pagetia del Cambriano medio australiano, assomigliano superficialmente ai piccoli occhi schizocroali. Hanno un numero relativamente piccolo di lenti biconvesse separate, circa 50 ÷ 70, disposte in modo semiregolare, ma in cui spesso non si trova materiale cuticolare tra le lenti; come suggerisce Jell, è possibile che ognuno di questi elementi avesse una sua propria cornea, ancorata al margine di ciascuna lente. Tuttavia, in uno studio condotto su Eodiscidi del Cambriano inferiore, X.G. Zhang e E.N.K. Clarkson (1990) sollevano qualche dubbio circa la possibilità che l'occhio abatocroale fosse realmente un tipo differente di organo visivo. Questi autori hanno esaminato gli Eodiscidi cinesi Shizhudiscus e Neocobboldia chinlinica. In Shizhudiscus sono presenti occhi olocroali normali con lenti in contatto tra loro, mentre in Neocobboldia chinlinica le lenti sono circolari e separate. La disponibilità di una serie ontogenetica completa per entrambe le specie ha permesso di ipotizzare che gli occhi di Neocobboldia potessero essere in realtà derivati da un normale occhio olocroale per pedomorfosi, in maniera analoga a quella ipotizzata per l'origine degli occhi schizocroali dei Facopidi. Zhang e Clarkson (1990) hanno suggerito che l'occhio abatocroale potrebbe essere un tipo di occhio schizocroale, sebbene meno specializzato di quello che si trova nei Facopidi. A favore di questo punto di vista è la scoperta che le lenti di Neocobboldia, sebbene non presentassero una struttura doppia, erano provviste di una superficie prossimale asferica con una protuberanza centrale, del tutto simile a quella delle unità superiori delle lenti dei primi occhi schizocroali di Dalmanitina dell'Ordoviciano, che si credono originate allo stesso modo per neotenia (Clarkson e Levi-Setti, 1975). La neotenia si riferisce a una riduzione nel tasso di sviluppo di un particolare carattere, in questo caso dell'occhio, rispetto a quelli del resto dell'individuo. Come sarà discusso più avanti, una particolare funzione ottica potrebbe essere associata a questo tipo di lenti, in relazione ai primi passi nel processo di ottimizzazione delle funzioni visive.
Principi dell'ottica e ottimizzazione della visione negli occhi dei trilobiti
È stato rilevato inizialmente da R. Levi-Setti (1975) ed elaborato successivamente da Clarkson e Levi-Setti (1975) che il profilo delle unità superiori delle strutture a doppia lente osservato negli occhi schizocroali di alcuni Facopidi presenta una stretta somiglianza con quello delle lenti asferiche e aplanari derivate da René Descartes (1637) e Christiaan Huygens (1690). Queste spesse lenti di vetro, funzionanti in aria, trasmettono un fascio di raggi luminosi che emerge da una sorgente puntiforme, cioè l'oggetto, fino a un punto di fuoco, cioè il punto di formazione dell'immagine, soddisfacendo la cosiddetta condizione stigmatica per un sistema ottico. Tale sistema non soffre di aberrazione sferica, per cui l'immagine di un oggetto puntiforme non è più un punto, ma una distribuzione diffusa di punti lungo l'asse ottico. Descartes derivò la sua costruzione geometricamente, applicando la legge della rifrazione da lui scoperta, attribuita indipendentemente anche a W. Snellius. Invece, Huygens basava la sua costruzione sul principio dell' ottica ondulatoria che porta il suo nome, per il quale la condizione stigmatica si raggiunge quando tutti i raggi compiono il loro percorso dalla sorgente puntiforme al punto di formazione dell'immagine attraverso le lenti nello stesso tempo, compreso il percorso lungo la retta che unisce l'immagine all'oggetto, che è il più breve e quindi quello che, in linea di principio, richiede il tempo minore. In quest'affermazione si può riconoscere un fondamentale principio ottico, il principio di Fermat del tempo minimo, secondo il quale il percorso effettivo che compie un raggio di luce tra due punti è quello che si attraversa nel minor tempo. Riformulando il principio di Fermat, applicato alla costruzione di Huygens, in termini di cammino ottico, grandezza ottenuta moltiplicando la distanza geometrica percorsa da un raggio luminoso per l'indice di rifrazione del mezzo, si può affermare che tutti i raggi provenienti da una sorgente puntiforme, che attraversano una qualsiasi sequenza di mezzi rifrangenti, convergeranno al punto di formazione dell'immagine, se percorrono cammini ottici minimi e identici. Nel caso di un diottro, cioè un'interfaccia che separa due mezzi con indici di rifrazione n1 e n2, si avrebbe quindi un'equazione del tipo
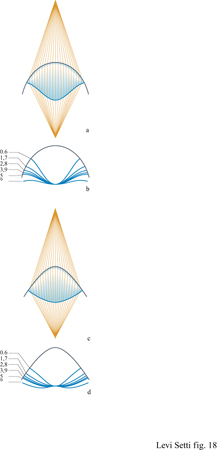
l1n1 + l2n2 = costante dove l1 e l2 sono le distanze geometriche percorse nei due mezzi. Tale equazione definisce la forma dell'interfaccia, appartenente a una famiglia di curve denominate ovali cartesiani. Per esempio, un fascio di raggi paralleli in aria, cioè dalla sorgente verso l'infinito, convergerà in un punto nel vetro quando l'interfaccia convessa dell'ovale cartesiano è un ellissoide. Analogamente, i raggi che provengono da una sorgente puntiforme in aria convergeranno in un punto di formazione dell'immagine all'infinito nel vetro, ossia rendendo i raggi paralleli, quando l'interfaccia convessa dell'ovale carte siano è un iperboloide. Se abbiamo a che fare con due interfacce, come nel caso delle lenti spesse prese in considerazione da Descartes e Huygens, una o entrambe le interfacce devono essere ovali cartesiani per soddisfare la condizione stigmatica. In questo modo, per esempio, una lente convessa a doppia iperbole è una lente perfetta. Nella costruzione di Huygens, la prima superficie riflettente è sferica e la superficie di uscita è un ovale carte siano dal profilo ondulato. In una ricostruzione con il computer delle 'lenti ustorie' di Descartes e Huygens, si può vedere (v. fig. 18) come la superficie ondulata di uscita cambi in funzione della distanza dall'oggetto, per due tipi diversi della prima superficie. In una rielaborazione moderna del principio di Fermat in forma variazionale è ancora permessa, senza influire sul risultato, una serie di traiettorie che deviano leggermente dalla costruzione ideale. Bisogna tuttavia notare che, applicando il principio di Fermat per calcolare il profilo di una superficie, non è necessario che l'interfaccia costituisca una transizione repentina tra due mezzi con indici di rifrazione differenti. Un cambiamento graduale nelle proprietà rifrangenti non influenzerà il risultato finale, come in un miraggio dove un cuscino d'aria d'intensità variabile si comporta, per un osservatore lontano, come una superficie speculare.
Ottica delle doppie lenti schizocroali

Sulla base della discussione precedente sull'ottica cartesiana e huygensiana, è interessante considerare la superficie prossimale ondulata delle unità superiori nelle strutture delle lenti doppie dei Facopidi come un ovale carte siano, che renderebbe tale unità una lente stigmatica e aplanare o, in altre parole, una lente che forma un'immagine priva dello sfocamento causato dall' aberrazione sferica. Una differenza rilevante, però, tra la situazione cui si riferiscono le costruzioni in figura (v. figura 18), valide per lenti di vetro funzionanti in aria, e la situazione presente effettivamente nei trilobiti, sta nella natura dei mezzi coinvolti: calcite orientata (indice di rifrazione n = 1,66 lungo l'asse c) per le unità superiori, acqua del mare (n = 1,33) anteriormente alle lenti, fluidi corporei (n = 1,35) posteriormente. Inoltre, vi sono le capsule intralenticolari con un indice di rifrazione sconosciuto. Nelle nostre prime ricostruzioni empiriche, condotte mediante tracciamento dei raggi e modelli sperimentali una soluzione che rendeva aplanatico l'insieme dei doppietti per esempio nel profilo dei Crozonapsis - l'indice di rifrazione della capsula aveva un valore intermedio tra quello della calcite e quello dei fluidi corporei. La sua funzione sembrava principalmente quella di mettere a fuoco i raggi più periferici. Sebbene la composizione di questa unità sia sconosciuta, il fatto che si sia preservata come fossile suggerisce che fosse coinvolto un componente minerale, probabilmente un composto stratificato di calcite e materiale organico. Una ricostruzione più rigorosa fatta da G. Horváth (1989a) per il profilo, tra gli altri, di Crozonaspis, mediante un modello matematico che consente di fare delle simulazioni al calcolatore, ha prodotto un indice di rifrazione n = l,53 per le capsule. La sua funzione è stata studiata ulteriormente da Horváth (1996), relativamente alla sua capacità di aumentare la trasmittanza delle lenti doppie rispetto a quelle singole di Huygens. Alcuni tracciamenti dei raggi, ottenuti con l'ausilio del calcolatore e basati sul suddetto indice di rifrazione, sono presentati in figura (fig.19), dove viene mostrata una lente divisa, con una struttura a lente doppia perfettamente funzionante a sinistra e come unità singola sulla destra (v. figura 19a). Nella metà sinistra, la forma dell'interfaccia corretta è determinata dalla condizione stigmatica del punto di fuoco. Nella metà destra, in assenza di tale correzione, i raggi che emergono intersecano l'asse sopra una distribuzione di punti focali differenti dando luogo a una forte aberrazione sferica. Nella figura 19b si vede come la forma dell'interfaccia vari in funzione della posizione dell'oggetto, la curva più bassa per un oggetto all'infinito, quella più alta per un oggetto posto a distanza uguale a quella dello spessore delle lenti. Degno di nota è il parametro f/numero, cioè il rapporto tra la distanza focale e l'apertura della lente, che in queste lenti funzionanti in acqua è pari circa a f/l,l, paragonabile a quello delle lenti delle migliori macchine fotografiche reperibili attualmente in commercio. Sebbene sia stato notato che il profilo delle unità superiori delle lenti di Dalmanitina socialis somigliasse a quello che si ottiene utilizzando le costruzioni di Descartes (Levi-Setti, 1975; Clarkson e Levi-Setti, 1975), non è stata possibile la sua riproduzione utilizzando il tracciamento dei raggi mediante il calcolatore di Horváth (1989a), per l'assunzione che tutti i raggi debbano convergere in un unico fuoco. Su questo punto si tornerà di nuovo più avanti, quando si discuterà della funzione ottica delle lenti degli occhi abatocroali.
Le lenti giganti degli occhi schizocroali, comparate con quelle degli occhi olocroali, con il loro potere eccezionale di assorbire la luce suggeriscono che gli animali che possedevano questo tipo di occhi vivessero in un ambiente molto poco illuminato o che fossero animali notturni, e che le loro larghe lenti biconvesse cercassero di raccogliere quanta più luce possibile. Siccome la superficie esterna delle lenti schizocroali è molto convessa, le lenti adiacenti nello stesso occhio potrebbero essere state usate per la visione stereoscopica (Stockton e Cowen, 1976). Questo spiegherebbe le file verticali dorsoventrali, la cui alta separazione angolare potrebbe tornare utile a tale proposito, così come la necessità di una disposizione regolare di queste lenti nella superficie visiva. I trilobiti devono riprodurre le loro lenti dopo ogni muta, giacché le lenti della muta precedente restano con il vecchio esoscheletro. Come già detto nella precedente descrizione degli occhi schizocroali, inizialmente le nuove lenti sono piccole e coniche per poi allargarsi verso la periferia della cornea e svilupparsi ulteriormente. La funzione ottica delle lenti è ottimizzata fm dai primi stadi dello sviluppo; viene inoltre raggiunta un'efficacia massima con una superficie della lente prossimale del tipo di Huygens per la correzione dell'aberrazione sferica, aumentando così la sua efficienza nel raccogliere la luce e nel trasferire il contrasto. Durante i primissimi stadi dello sviluppo l'occhio era miope, ma in seguito divenne in grado di vedere distintamente a una distanza che andava da qualche millimetro fino a qualche centimetro dalla superficie visiva (Horváth e Clarkson, 1993). Dopo la muta e durante il periodo in cui la cuticola rimane flessibile, i trilobiti non erano in grado di muoversi molto, come accade per parecchie specie di artropodi immediatamente dopo la muta. In questa fase, gli animali dovevano probabilmente tenersi nascosti, forse rimanendo parzialmente sottoterra o subendo mute notturne; in tale situazione era molto importante avere occhi estremamente sensibili alla luce per scorgere i predatori, come pure era importante la possibilità di ottenere un'immagine chiara al più presto dopo l'ecdisi. La maturazione ottica negli occhi schizocroali differisce da quella che si riscontra in alcuni diottri degli attuali occhi composti, dove la maturazione stessa è relativamente lenta e la qualità dell'immagine, negli stadi immediatamente successivi all'ecdisi, è estremamente scadente, mentre con la maturazione dell'occhio le lenti diventano più efficaci. Questa differenza è probabilmente correlata ai differenti stili di vita postecdisi dei trilobiti e degli artropodi attuali.
Occhi abatocroali dei trilobiti: formazione dell'immagine mediante lenti bifocali
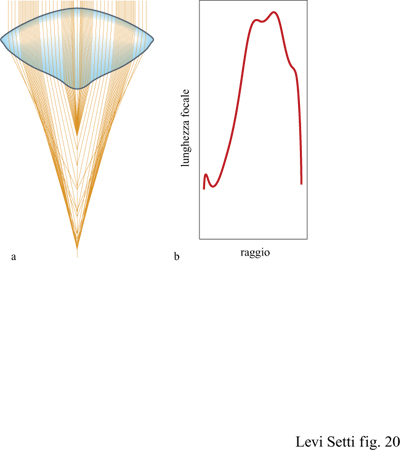
Siccome la classificazione dell'occhio di tipo abatocroale rimane incerta, il suo sistema ottico è stato studiato in maniera dettagliata, con particolare riferimento ai trilobiti Neocobboldia chinlinica le cui lenti, come notato precedentemente, evidenziano un qualche tipo di correzione ottica. Una sezione attraverso il piano verticale principale delle lenti è stata ricostruita sulla base di fotografie ottenute con il microscopio elettronico a scansione (SEM, Scanning Electron Microscope) e il profilo della lente distale, cioè l'entrata, e prossimale, cioè l'uscita, è stato digitalizzato mediante un analizzatore di immagine (fig. 20). Un tracciato computerizzato dei raggi parassiali è mostrato in figura 20a, dove si assume che la superficie distale della lente di calcite era immersa in acqua di mare con un indice di rifrazione n = 1,33, mentre l'interno era in contatto con il citoplasma con un indice di rifrazione n = 1,35; sono indicate due regioni di convergenza per i raggi che attraversano la regione periferica della lente e la protuberanza centrale che possiede un maggiore potere di rifrazione, come si può anche apprezzare quantitativamente dal grafico della lunghezza focale in funzione della distanza del raggio radiale (v. figura 20b). La lente abatocrale di Neocobboldia è quindi realmente bifocale, con una regione centrale che ha una lunghezza focale minore rispetto alla zona anulare e una transizione abbastanza rapida tra le due regioni. Un'aberrazione sferica consistente colpisce la regione più periferica della lente, a meno che non ci sia un filtro di pigmenti schermanti sotto e intorno alla lente, come negli artropodi attuali. Il rialzo relativamente ripido del profilo prossimale e la differenza considerevole nell'indice di rifrazione tra la lente (calcite: n = 1,66) e il materiale sotto stante la lente (probabilmente fluido corporeo: n = 1,35) determinavano una grande riflessione interna sulla superficie prossimale rifrangente, riducendo la trasmittanza e così anche l'efficacia dell'occhio nel raccogliere la luce. Vista la somiglianza di questo tipo di lente con quello schizocroale di Dalmanitina, che non è conforme al tipo huygensiano presente nelle altre lenti del tipo schizocroale, si è tentati di associare anche a quest'ultima una funzione bifocale. Tale ipotesi è attualmente oggetto di studio.
Gli occhi abatocroali vengono classificati come un terzo tipo di occhio dei trilobiti, distinti da quello schizocroale e olocroale; inoltre essi acquisiscono un particolare significato evolutivo in quanto forniscono l'evidenza di un primo passo, benché non ancora ottimale, verso la correzione dell'aberrazione sferica delle lenti, che verrà raggiunta pienamente nei gruppi successivi di trilobiti. È in qualche modo sorprendente trovare, proprio all'inizio dell'evoluzione dei trilobiti, un tale tentativo di ottimizzazione delle funzioni visive e, come per le lenti doppie ottimizzate dei Facopidi, si potrebbe speculare su quale sia stata la pressione selettiva alla quale tale fenomeno è stato sottoposto. Da questa prospettiva, altri fattori, oltre alla correzione dell'aberrazione sferica, potrebbero contribuire a migliorare i processi visivi.
Per esempio, la profondità del campo, cioè l'intervallo di distanze alle quali gli oggetti possono essere chiaramente distinti con un sistema diottrico-fotorecettore, acquisisce rilevanza particolare per l'osservazione dell'ambiente circostante e per la salvaguardia della vita. Se infatti gli occhi abatocroali di Neocobboldia erano provvisti di una retina in miniatura, come supposto per l'occhio schizocroale, allora la visione bifocale può essere considerata come una risposta al bisogno di una profondità di campo maggiore; la lente bifocale non avrebbe avuto alcun valore se le lenti individuali avessero posseduto soltanto un singolo fotorecettore.
Evoluzione dell'ottica delle lenti dei trilobiti
Lenti doppie grandi e aplanatiche, cioè corrette dall'aberrazione sferica, apparvero relativamente tardi neII'evoluzione dei trilobiti (Clarkson, 1975; 1979) ed erano presenti unicamente negli occhi schizocroali dei Facopidi; l'occhio ancestrale dei trilobiti conteneva solo piccole lenti singole di calcite. Questo è vero anche per gli occhi del tipo abatocroale, che si svilupparono indipendentemente e che rappresentano uno stadio evolutivo intermedio tra gli occhi olocroali e quelli schizocroali. Gli occhi di tipo abatocroale di Neocobboldia chinlinica, a differenza di quelli schizocroali e olocrali, avevano lenti bifocali che sembrano essere state corrette, in parte, dall'aberrazione sferica. Sulla base dei dati che si sono accumulati sul sistema visivo dei trilobiti si può proporre il seguente panorama evolutivo dell'ottica delle loro lenti:
l) il primo stadio dello sviluppo si verificò nell' occhio olocroale ancestrale e consistette nel minimizzare l'influsso della birifrangenza nella singola lente corneale. Questo problema si risolse mediante un orientamento corretto dell' asse c dei cristalli di calcite, in modo tale che questo fosse sempre allineato parallelamente all'asse ottico delle lenti, che era a sua volta normale alla superficie visiva dell'occhio. Le piccole lenti olocroali non erano altrettanto aplanatiche. A giudicare dalle loro dimensioni, esse dovrebbero aver avuto un potere di rifrazione limitato e, quindi, non essere state sottoposte a pressione selettiva per correggere l'aberrazione sferica.
2) Un secondo passo evolutivo fu la comparsa dell'occhio abatocroale in cui erano presenti lenti più grandi di quelle dell'occhio olocroale, probabilmente associate a un ambiente visivo poco illuminato. Senza nessuna correzione, le grandi lenti abatocroali potrebbero aver sofferto di un'aberrazione sferica notevole. La forma effettiva delle lenti garantiva un certo grado di correzione dell'aberrazione sferica, e la protuberanza centrale sul profilo della lente prossimale la rendeva bifocale e, di conseguenza, aumentava l'effettiva profondità di campo. La riflessione interna potrebbe avere in qualche modo ridotto l'efficienza di queste lenti nel raccogliere la luce. Questo tipo di occhi è confinato ai trilobiti della famiglia Eodiscidi del Cambriano medio e inferiore. Molti di questi animali persero i loro occhi del tutto e il gruppo si estinse non lasciando discendenti. A giudicare dal diametro medio di circa 20 ÷ 30 μm, le lenti abatocroali dovrebbero essersi approssimate ai limiti della diffrazione. Ciononostante, ci deve essere stata ancora qualche pressione selettiva per correggere l'aberrazione sferica e determinare lo sviluppo delle lenti bifocali, le quali tuttavia potrebbero essere riapparse in alcuni dei primi esempi di occhi schizocroali.
3) Nel complesso, lo sviluppo evolutivo di gran lunga più rilevante si trova nel tipo schizocroale. Negli occhi dei Facopidi sia la struttura ordinata nella disposizione delle lenti, sia il perfetto profilo huygensiano dell'interfaccia sfericamente corretta suggeriscono una risposta a pressioni selettive che vanno ben oltre la semplice necessità di raccogliere la massima quantità di luce. Anche se, a causa della maggior concentrazione di luce su un piano focale ben definito, la correzione dell'aberrazione sferica potrebbe aver ridotto il livello di soglia per la percezione della luce e la presenza delle cavità interne alle lenti potrebbe aver migliorato il potere di acquisizione della luce da parte delle lenti riducendo la riflessione interna, altri artropodi marini hanno sviluppato soluzioni estremamente più semplici per portare al limite massimo la raccolta della luce, per esempio la presenza dei coni cristallini in Limulus (Levi Setti et al., 1975). Nella struttura degli occhi schizocroali, il passo evolutivo più importante, di fatto, potrebbe essere costituito dalla maggiore complessità dei fotorecettori e della rete neurale che beneficia di una visione stereoscopica e di un'immagine definita dell'ambiente circostante: in altre parole lo sviluppo di un vero e proprio, ancorché primitivo, cervello. A causa dell'estinzione dei trilobiti, l'occhio schizocroale rimane un punto d'arrivo nella storia evolutiva. Tuttavia, i principi dell'ottica hanno guidato l'evoluzione convergente e sistemi visivi ottimizzati in maniera simile si sono sviluppati in altre forme di vita alle quali accenneremo.
Riduzione degli occhi e cecità nei trilobiti
Gli occhi composti sono strutture primarie presenti in molti trilobiti: sono particolarmente evidenti nelle specie in grado di arrotolarsi fino a formare una palla per proteggersi e presumibilmente funzionavano come primi sensori di allarme, rilevando l'avvicinarsi dei predatori. Sembrerebbe quindi strano che, indipendentemente, in molti gruppi di trilobiti gli occhi si siano ridotti o scomparsi del tutto. In questi casi, la perdita di tali strutture è correlata all'adozione di ambienti di vita privi di luce, nelle profondità marine, o disabitati; tuttavia in alcune specie completamente cieche si sono sviluppati organi di senso di un tipo completamente differente, diventati successivamente i dispositivi principali di controllo dell' ambiente, come per esempio nei Trinucleidi dell'Ordoviciano dove le ampie e infossate frange cefaliche costituivano probabilmente un organo vibrosensorio.
L'esempio più evidente della riduzione progressiva dell'occhio fino alla cecità finale si trova nei Proetidi del Devoniano medio e superiore, dotati di occhi olocroali, e nei Facopidi, dotati di occhi schizocroali. In alcune linee distinte di questi gruppi, la sutura facciale migra verso l'esterno e lateralmente e gli occhi si riducono. Una delle migliori testimonianze di tale fenomeno proviene dai reperti rinvenuti in Germania e nella Francia meridionale (Feist e Clarkson, 1989; Feist, 1991), nei quali può essere seguita una perdita progressiva degli occhi lungo alcuni milioni di anni; anche in alcune specie di trilobiti cinesi è stata osservata una parallela e contemporanea perdita di tali strutture. Dal momento che ogni fase successiva nella riduzione degli occhi ricorda una condizione più giovanile della forma ancestrale, questo processo sembra essere sotto controllo pedomorfico. l trilobiti che andavano incontro a tale trasformazione sono così diventati idonei a una modalità di vita bentonica, quando il fondo marino roccioso, durante il Siluriano e il Devoniano inferiore, venne sostituito in gran parte del mondo da una morbida coperta di fango. In questo contesto la riduzione degli occhi e la cecità sembrano essere più adattative che degenerative.
Altri sistemi visivi fossili: morfologia e diottrica delle macchie oculari negli ostracodi estinti
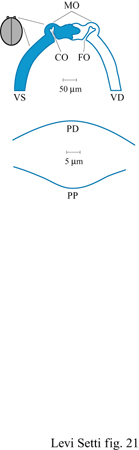
Come nei trilobiti, gli ostracodi fossili possedevano degli apparati diottrici che in vita erano composti di calcite, cosicché i loro occhi si sono conservati minutamente. D'altra parte, gli ostracodi fossili hanno molti parenti attuali i cui sistemi visivi sono ben noti. l tubercoli ottici o macchie oculari sono strutture a guscio posizionate nella regione anterodorsale delle valve di alcuni ostracodi fossili e attuali provvisti di occhi (Kontrovitz e Myers, 1984). Queste strutture (fig. 21) sono piccole, circolari o ovali, trasparenti e convesse verso l'esterno e hanno un diametro di 30 ÷ 60 μm. La lente di queste macchie oculari è costituita, come negli ostracodi moderni, da calcite trasparente orientata, che permette alla luce di entrare nell'occhio all'apice distale di una cavità oculare piuttosto peduncolata. La cavità oculare, in cui passano gli assoni, è un canale nel guscio bivalve che giace tra un'apertura interna, chiamata fossa oculare, e la superficie distale della macchia oculare. La superficie distale della lente è convessa e approssimativamente sferica; la superficie prossimale della lente è ondulata con una convessità centrale e concavità anteriori e posteriori (v. figura 21b). Superficialmente, questa geometria è simile alla forma delle lenti superiori negli occhi schizocroali dei trilobiti. Secondo M. Kontrovitz e J.H. Myers (1984), le lenti delle macchie oculari degli ostracodi concentrano la luce nello spazio oculare sotto stante. Nel fossile Echinocythereis jacksonensis, i raggi luminosi parassiali sono focalizzati a una distanza che cade dietro la cavità oculare, a meno che i raggi non vengano riflessi dal piano coroidale dell' occhio (Kontrovitz e Myers, 1988).
La comparazione di una specie attuale, Echinocythereis margaritifera, con Echinocythereis jacksonensis dell'Oligocene evidenzia una morfologia analoga della macchia oculare e, presumibilmente, anche un funzionamento simile (Kontrovitz e Myers, 1984). Il genere Echinocythereis è di particolare interesse perché la morfologia delle macchie oculari è correlata ai livelli di luce che, a loro volta, sono in relazione alla profondità delle acque. Per quei sedimenti in cui erano presenti ostracodi forniti di occhi è quindi disponibile un metodo per ricostruire la profondità delle acque (Kontrovitz e Myers, 1988), a meno che questi ostracodi non fossero specie notturne. R.H. Benson (1984) riporta una diminuzione generale nella dimensione delle macchie oculari degli ostracodi fossili che è proporzionale all'aumento della profondità dell' acqua, fino a scomparire alla profondità di 600÷900 m. Kontrovitz e Myers (1984; 1988) hanno dimostrato che nell'oceano, a una profondità superiore a 300 m, gli occhi degli ostracodi podocopidi avrebbero avuto una scarsa utilità, a causa sia della loro conformazione sia dei bassi livelli di luce. Eventuali modificazioni al sistema macchia oculare-piano coroidale, in effetti, avrebbero potuto far ben poco per migliorare la visione a profondità così elevate.
Mediante tracciamento dei raggi, Myers e Kontrovitz (1988) sono stati in grado di stabilire che le macchie oculari degli ostracodi, sia fossili sia attuali, sono più o meno corrette dall'aberrazione sferica mediante il profilo ondulato huygensiano della lente di calcite prossimale, come si verifica nelle lenti corneali aplanatiche dei trilobiti con occhi schizocroali. Nonostante ciò, restano molti dubbi sulla presenza negli ostracodi fossili e attuali di un sistema visivo altamente sviluppato. Il piccolo numero di rabdomeri, 9 ÷ 18 per capsula, che si riscontra nelle forme attuali e che era probabilmente presente in quelle fossili, impedirebbe una vista ad alto potere risolutivo (Kontrovitz e Myers, 1984). D'altra parte, il valore dij1numero del sistema lentespecchio degli ostracodi podocopidi, simile a quello dell'occhio di Pecten (v. oltre), era limitato in un intervallo di 0,50 ÷ 0,25, il più piccolo conosciuto per qualsiasi organismo. Questi animali, inoltre, sembrano essersi adattati bene in ambienti scarsamente illuminati (Kontrovitz e Myers, 1988). Lo scopo della correzione dall'aberrazione sferica delle macchie oculari negli ostracodi potrebbe quindi essere semplicemente un aumento nella capacità di raccogliere la luce in ambienti bui.
Correzione ottica negli invertebrati attuali
Fatta eccezione per l'uso delle lenti di cristalli di calcite, in molti invertebrati attuali si riscontrano somiglianze di vario grado con le strutture degli occhi schizocroali dei trilobiti, alcune anche tra i non artropodi. Questo tipo di occhi ha in comune con quelli dei trilobiti schizocroali almeno una delle tre caratteristiche tipiche: unità ottiche ben separate, presenza di lenti doppie, correzione dell'aberrazione sferica. La struttura attuale che più si avvicina alle caratteristiche esterne peculiari degli occhi schizocroali dei trilobiti è l'occhio composto del maschio adulto degli insetti pterigoti strepsitteri, che si presenta sfaccettato a forma di mora, con ommatidi ben separati (Strohm, 1910; Roesch, 1913).
Alcuni occhi attuali possiedono lenti doppie che sono aplanatiche come quelle presenti nei trilobiti con occhi schizocroali. Uno di questi sistemi visivi attuali è l'occhio composto di Notonecta, la cimice d'acqua che nuota all'indietro (Schwind, 1980). In ogni ommatidio, entrambe le lenti sono omogenee dal punto di vista ottico, con indici di rifrazione differenti. Tra le due lenti c'è un sottile strato di transizione a forma di campana che riduce l'aberrazione sferica della lente, tanto in aria quanto in acqua (Horváth, 1989b). Le larve di Perga, un imenottero della famiglia dei Tentredinidi, hanno solo un paio di ocelli composti che presentano lenti circolari biconvesse: la lente consiste di due strati otticamente omogenei con indici di rifrazione differenti. Al di sotto delle lenti è presente una retina ampia (Meyer-Rochow, 1974). Utilizzando il tracciamento dei raggi è stato possibile evidenziare che gli ocelli sono corretti dall'aberrazione sferica.
In Pecten, un mollusco bivalve marino, sono presenti lungo i margini del mantello circa 60 occhi ben separati, di diametro pari a circa l mm (Land, 1965). Un'immagine reale si forma nell'occhio per rifrazione attraverso una lente spessa e per riflessione sulla membrana argentea, una struttura multistratificata composta di cristalli di guanina, che funziona come un riflettore estremamente efficace. La membrana argentea sferica riveste interamente la zona posteriore dell'occhio. La superficie posteriore della lente omogenea è sferica, quella frontale è a forma di campana. Questo profilo frontale corregge dall'aberrazione sferica come le piastre di correzione nel telescopio astronomico Schmidt (Horváth e Varju, 1993). La retina occupa lo spazio tra la lente e la membrana argentea ed è costituita di due strati principali di fotorecettori (Land, 1966). L'immagine si forma sulla retina distale, che è coinvolta anche nella percezione del movimento. La retina prossimale, dove non si forma l'immagine, controlla il livello d'illuminazione.
Ringraziamenti
R. Levi-Setti è grato alla Physical Sciences Division dell'università di Chicago per il sostegno a questa attività multidisciplinare. A Budapest questo lavoro è stato sostenuto da un finanziamento ricevuto da G. Horváth dalla Hungarian National Scientific Research Foundation (OTKA F-012858).
Bibliografia citata
BENSON, R.H. (1984) Estimating greater paleodepths with ostracodes, especially in past thermospheric oceans. Palaeogeog. Palaeoclimatol. Palaeoecol. Amsterdam, 48, 104-107.
CAMPBELL, K.S.W. (1975) The functional anatomy of trilobites: musculature and eyes. J. Proc. R. Soc. N. S. w., 108, 168-188.
CLARKSON, E.N.K. (1968) Structure of the eye of Crozonaspis struvei (Trilobita, Dalmanitidae, Zeliszkellinae). Senck. Lethaea, 49, 383-391.
CLARKSON, E.N.K. (1975) The evolution of the eye in trilobites. Fossils and Strata, 4, 7-31.
CLARKSON, E.N.K. (1979) The visual system of trilobites. Palaeontology, 22, 1-22.
CLARKSON, E.N.K., LEVI-SETTI, R. (1975) Trilobite eyes and the optics of Des Cartes and Huygens. Nature, 254, 663-667.
CLARKSON, E.N.K., ZHANG, X. (1991) Ontogeny of the Carboniferous trilobite Paladin eichwaldi shunnerensis (King, 1914). Trans. R. Soc. Edinburgh: Earth Sci., 82, 277-295.
D'ARCY THOMPSON, W. (1942) On growth and form, Cambridge, Cambridge University Press.
DES CARTES, R. (1637) Oeuvres de Des Cartes: La Geometrie, Livre 2, pp. 134. Leyden, J. Maire.
FEIST, R. (1991) The Late Devonian trilobite crises. Hist. Biol. 5, 197-214.
FEIST, R., CLARKSON, E.N.K. (1989) Environmentally controlled phyletic evolution, blindness and extinction in Late Devonian tropidocoryphine trilobites. Lethaia, 22, 359-373.
FORDYCE, D., CRONIN, T.W. (1989) Comparison of fossilized schizochroal compound eyes of phacopid trilobites with eyes of modern marine crustaceans and other arthropods. J. Crustacean Biol., 9, 554-569.
FORDYCE, D., CRONIN, T.W. (1993) Trilobite vision: a comparison of schizochroal and holochroal eyes with the compound eyes of modern arthropods. Paleobiology, 19, 288-303.
HORVÁTH, G. (1989a) Geometric optics of trilobite eyes: a theoretical study of the shape of the aspherical interface in the cornea of schizochroal eyes of phacopid trilobites. Math. Biosciences, 96, 79-94.
HORVÁTH, G. (1989b) Geometric optical optimisation of the corneal lens of Notonecta glauca. J. Theor. Biol., 139, 389-404.
HORVÁTH, G. (1996) The lower lens unit in schizochroal trilobite eyes reduces reflectivity: on the possible optical function of the intralensar bowl. Hist. Biol., 12, 83-92.
HORVÁTH, G., VARJU, D. (1993) Theoretical study ofthe optimal shape of the front profile of the lens in the eye of the scallop Pecten. Bull. Math. Biol., 55, 155-174.
HORVÁTH, G., CLARKSON, E.N.K. (1993) Computational reconstruction of the probable change of form of the corneal lens and maturation of optics in the post-ecdysial development of the schizochroal eye of the Devonian trilobite Phacops rana milleri Stewart 1927. J. Theor. Biol., 160, 343-373.
HUYGENS, C. (1690) Traite de la Lumiere. Leiden, Pierre van der Aa.
JELL, P.A. (1975) The abathochroal eye of Pagetia: a new type of trilobite eye. Fossils and Strata, 4, 33-43.
KONTROVITZ, M., MYERS, J.H. (1984) A study of the morphology and dioptics of some ostracode eyespots. Trans.-Gulf Coast Ass. Geol. Soc. 34, 369-372.
KONTROVITZ, M., MYERS, J.H. (1988) Ostracode eyes as paleoenvironmental indicators: physicallimits of vision is some podocopids. Geology, 16, 293-295.
LAND, M.F. (1965) Image formation by a concave reflector in the eye of the scallop Pecten maximus. Journal of Physiology, 179, 138-153.
LAND, M.F. (1966) Activity in the optic nerve of Pecten maximus in response to changes in light intensity, and to pattern and movement in the optical environment. Journal of Experimental Biology, 45, 83-99.
LEVI-SETTI, R. (1975) Trilobites: a photographic atlas, Chicago, Univ. Chicago Press.
LEVI-SETTI, R. (1993) Trilobites, 2a ed. Chicago, Univ. Chicago Press.
LEVI-SETTI, R., PARK, D.A., WINSTON, R. (1975) The corneal cones of Limulus as optimised light concentrators. Nature, 253, 115-116.
LINDSTROM, G. (1901) Researches on the visual organs of the trilobites. K. Svensk. Vetensk. Akad. Handl., 34, 1-86. MEYER-ROCHOW, V.B. (1974) Structure and function of the larval eye ofthe sawfly. J. Insect Physiol., 20,1565-1591.
MILLER, J., CLARKSON, E.N.K. (1980) The post-ecdysial development of the cuticle and the eye of the Devonian trilobite Phacops rana milleri Steward 1927. Proc. R. Soc. Lond., B 288, 461-480.
MYERS, J.H., KONTROVITZ, M. (1988) Geometric optics of some ostracode eyes. In Evolutionary biology of Ostracoda., a c. di Hanai T., Ikeya N., Ishizaki K. Proc. 9th Int. Symp. on Ostracoda. Elsevier, Amsterdam and Koansha, Tokyo, pp. 187-193.
ROESCH, P. (1913) Beitriige zur Kenntnis der Entwicklungsgeschichte der Strepsipteren. Jenaer Zeits. Naturwiss., 50, 97-146.
SCHWIND, R. (1980) Geometrical optics of the Notonecta eye: adaptation in optical environment and way of life. J. Comp. Physiol., 140, 59-68.
STOCKTON, W.L., COWEN, R. (1976) Stereoscopic vision in one eye: paleophysiology of the schizochroal eye of trilobites. Paleobiology, 2, 304-315.
STROHM, K. (1910) Die zusanunengesetzten Augen der Miiunchen von Xenos rossii. Zoologischer Anzeiger, 36, 156-159.
ZHANG, X.G., CLARKSON, E.N.K. (1990) The eyes oflower Cambrian eodiscid trilobites. Palaeontology, 33, 911-932.
Bibliografia generale
CLARKSON, E.N.K. Invertebrate palaeontology and evolution., 4a ed. Chapman & Hall, 1998.
LEVI-SETTI, R. Trilobites, 2a ed. Chicago, University of Chicago Press, 1993.