
Piante transgeniche e loro applicazioni nelle agrobiotecnologie
Piante transgeniche e loro applicazioni nelle agrobiotecnologie
La produzione di piante transgeniche, cioè l'introduzione di geni provenienti da altri organismi in cellule vegetali e la successiva rigenerazione di piante a partire dalle cellule trasformate, è una nuova tecnologia ricca di grandi potenzialità che ha rivoluzionato la biologia vegetale e sta per avere un notevole impatto sull'agricoltura. Con questa tecnologia,è possibile non solo studiare in che modo i diversi geni sono attivi nei vegetali ma anche trasformare le piante in fabbriche a basso costo per produrre nuove qualità di amido, grassi, plastiche ed enzimi di impiego farmaceutico e industriale. È possibile rendere le piante resistenti agli erbicidi e agli agenti inquinanti come l'ozono e utilizzarle per l'accumulo, lo smaltimento senza pericolo e la riconversione di inquinanti di origine industriale. Le piante coltivate possono essere modificate al fine di renderle resistenti agli stress ambientali e alle malattie o di facilitare la produzione di ibridi. Le implicazioni sociali ed etiche della produzione di piante transgeniche sono discusse brevemente al termine di questo saggio.
Piante transgeniche
Negli anni Ottanta i primi rapporti scientifici sulla produzione di piante transgeniche (un processo detto anche trasformazione) hanno spalancato un nuovo mondo di possibilità di ricerca per i biologi vegetali. Questo settore della biologia è stato letteralmente rivoluzionato dalla possibilità di produrre piante transgeniche, cioè di introdurre nuovi geni nel patrimonio genetico dei vegetali o di sopprimere l'espressione di geni in esso presenti. Con questa tecnologia è possibile acquisire nuove conoscenze sul metabolismo delle piante, sulla funzione delle diverse cellule, sullo sviluppo, sulla fisiologia e sulla risposta a stimoli ambientali come la siccità, il freddo o la salinità del suolo. Inoltre, la trasformazione delle piante coltivate può permettere una produzione più efficiente di cibo, mangime per animali, fibre e prodotti farmaceutici a beneficio dell'intera umanità (tab. I). La trasformazione delle piante richiede una serie di tecnologie che sono, al giorno d'oggi, ampiamente in uso nelle università, nelle industrie e negli istituti di ricerca. Le tecnologie di base non sono particolarmente complesse e possono essere applicate facilmente anche in laboratori con un modesto sostegno finanziario (come nel caso dei paesi in via di sviluppo); i benefici derivanti dalla produzione di piante transgeniche non sono, perciò, necessariamente limitati ai paesi sviluppati. Tuttavia, attualmente solo poche specie vegetali, come il tabacco e la patata, sono facilmente trasformabili, mentre la trasformazione delle più importanti piante coltivate, come il mais, il frumento e la soia, è ancora complicata e scarsamente efficiente.
Introduzione di geni nel genoma della pianta
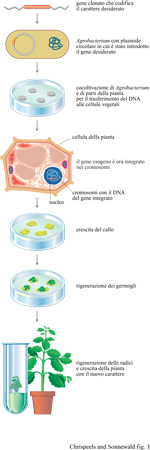
La produzione di piante transgeniche richiede diversi passaggi. lnnanzitutto, è necessario un sistema di trasformazione efficiente. Il più usato, la trasformazione mediata da agrobatterio, si basa su un meccanismo naturale di trasferimento di geni che avviene quando il batterio Agrobacterium tumefaciens, che causa nelle piante la galla, o tumore del colletto, trasferisce una breve sequenza di DNA plasmidico al genoma della pianta infettata (Chilton et al., 1977). Una scoperta fondamentale è stata che ceppi virulenti di A. tumefaciens contengono un grande plasmide extracromosomico (cioè non facente parte del singolo cromosoma del batterio), che è stato chiamato Ti (Tumor-inducing, induttore di tumore). L'evento molecolare che causa la formazione del tumore è il trasferimento e l'integrazione stabile di un preciso frammento del plasmide Ti nel genoma della pianta infettata. Il frammento, che è stato chiamato T-DNA (Transferred DNA, DNA trasferito), contiene alcuni geni, tre dei quali producono enzimi coinvolti nella via biosintetica di alcuni ormoni vegetali. L'aumento di sintesi di ormoni che ne deriva causa la crescita tumorale. Dunque, la formazione di tumori dovuta a infezione da agro batterio necessita del trasferimento di geni da un batterio a una pianta superiore e può, perciò, essere considerata un esempio di ingegneria genetica naturale. L'osservazione che grandi porzioni di T-DNA possono essere eliminate e sostituite con frammenti di DNA completamente diversi (Zambryski et al., 1983) ha aperto la strada all'introduzione di qualunque gene nelle cellule vegetali. L'introduzione simultane a del gene in esame e di geni codificanti marcatori, dominanti e selezionabili (Herrera-Estrella et al., 1983), l'eliminazione dal T-DNA dei geni che presiedono alla sintesi di ormoni e lo sviluppo del sistema di trasformazione che utilizza frammenti di foglie detti dischi fogliari (Horsch et al., 1985), hanno facilitato la produzione e la selezione di piante transgeniche e determinato lo sviluppo esplosivo della biologia molecolare delle piante. In ogni procedura di trasformazione solo una piccola percentuale delle cellule sottoposte al trattamento viene effettivamente trasformata; i marcatori dominanti selezionabili, come, per esempio, la resistenza a un antibiotico, permettono la riproduzione delle cellule trasformate ma non di quelle non trasformate, che vengono uccise quando al mezzo di coltura è aggiunto l'antibiotico. L'eliminazione dei geni che presiedono alla sintesi degli ormoni permette che le cellule trasformate diano origine a una nuova pianta, invece che continuare ad accrescersi come callo, cioè un ammasso indifferenziato di cellule, tumorale (fig. 1). Sebbene il sistema di trasferimento di geni mediato dall'agrobatterio sia il più conveniente per produrre piante transgeniche, esso non è applicabile a ogni specie vegetale: per esempio, non si è riusciti a trasformare con l' agro batterio la maggior parte delle Monocotiledoni. Si stanno, quindi, compiendo sforzi consistenti nel tentativo di sviluppare sistemi di trasferimento diretto di DNA, senza l'ausilio di plasmidi e batteri. Le tecniche alternative di trasformazione comprendono il microbombardamento con particelle ricoperte di DNA, l'uso di vettori virali, la trasformazione di protoplasti (cellule vegetali isolate e private della parete cellulare), tramite elettroporazione o trattamento con polietilenglicol (ambedue i procedimenti aumentano la permeabilità della membrana plasmatica e dunque facilitano l'introduzione di DNA), la microiniezione di DNA in protoplasti e la macroiniezione o l'infiltrazione di DNA in germogli vegetativi o in tessuti fiorali.
Promotori tessutospecifici e cellulospecifici nella costruzione di geni chimerici
La trasformazione ha spesso come obiettivo l'introduzione nella pianta ospite di una nuova via metabolica o la modificazione di una via già esistente. A causa della complessa organizzazione delle piante superiori e della distribuzione delle vie metaboliche in distinti comparti subcellulari, la manipolazione dei processi metabolici richiede l' ausilio di diversi strumenti molecolari. A seconda del problema specifico che si desidera risolvere mediante la trasformazione, il gene introdotto può dover essere espresso in maniera tessuto specifica o cellulospecifica (cioè in un solo tipo di tessuto o di cellula) o in appropriate condizioni ambientali o di sviluppo della pianta. l geni introdotti sono, per questo motivo, solitamente chimere, formate da quattro frammenti di DNA fusi in maniera appropriata: l) la sequenza che codifica la proteina che presiede alla funzione che si vuole trasferire; 2) una sequenza che codifica un peptide che indirizza al comparto subcellulare appropriato; 3) un promotore all'estremità 5' del DNA, cioè all'inizio della chimera; 4) una sequenza terminale all'estremità 3'.
Il promotore è l'elemento del gene che ne controlla la specificità di espressione; il promotore 35S è, finora, il più usato negli esperimenti di trasformazione delle piante. Questo promotore virale assai efficace fa parte del genoma del virus del mosaico del cavolfiore (CaMV, Cauliflower Mosaic Virus), dove presiede alla sintesi di un RNA virale che ha una costante di sedimentazione 35S. La regione del promotore 35S, comprendente la cassetta TATA e circa 350 paia di basi a monte di essa, è sufficiente per ottenere alti livelli di trascrizione di una qualunque sequenza di DNA fusa immediatamente a valle. Il promotore 35S è definito costitutivo perché promuove un'espressione generalizzata in tutta la pianta, tuttavia le sequenze poste sotto il suo controllo non sono espresse in modo uguale in ogni organo. Questo promotore è stato usato con successo per produrre piante tolleranti ai pesticidi o resistenti ai patogeni, ma è scarsamente usato nello studio delle vie metaboliche. Recentemente, sono stati isolati numerosi promotori con diverse specificità di espressione. Un gene batterico, codificante l'enzima β-glucuronidasi (GUS), è spesso usato per costruire chimere in esperimenti volti a determinare la specificità dei diversi promotori. L'enzima GUS non è presente nei vegetali e in una appropriata e semplice reazione biochimica produce un composto di colore blu. Poiché per questa reazione si possono utilizzare tessuti, organi o piante intatte, è facile identificare in piante transgeniche la specificità di un qualunque promotore fuso a monte del gene GUS: i tessuti in cui il promotore è attivo sintetizzano l'enzima e dunque, se è somministrato il substrato adatto, si colorano di blu. Sono stati isolati promotori specifici per organi e tessuti, come per esempio le foglie, i semi, i tuberi di patata, i frutti e i meristemi.
La fotosintesi avviene soprattutto nelle foglie mature, completamente espanse, composte da diversi tessuti e tipi di cellule. l geni foglia-specifici maggiormente studiati codificano per le seguenti proteine: la subunità minore della ribulosio-l,5-bifosfato-carbossilasi-ossigenasi, la proteina legante la clorofilla a/b e la proteina di 10 kDa del complesso che sviluppa l'ossigeno del foto sistema II. L' introduzione in piante transgeniche del gene GUS, posto sotto il controllo dei promotori di questi geni, ha dimostrato che la loro espressione è inducibile dalla luce e avviene in diversi tipi di cellule: del mesofillo, di guardia e compagne. Sono stati isolati altri promotori che permettono l'espressione esclusivamente nelle cellule di guardia e in quelle del mesofillo. È possibile alterare il trasporto di prodotti della fotosintesi usando il promotore floema-specifico del gene codificante l'enzima saccarosiosintetasi o il promotore del gene rolC di Agrobacterium rhizogenes. Sebbene la specificità dei differenti promotori sia utile negli studi sul metabolismo fogliare, differenze di sviluppo o di condizioni ambientali possono, tuttavia, complicare l'interpretazione dei dati ricavabili dall'analisi delle piante transgeniche. l promotori inducibili da agenti applicabili dall'esterno sarebbero ideali per l'espressione controllata di geni eterologhi. Molti promotori vegetali sono inducibili da metaboliti endogeni che hanno funzione di segnali molecolari, come gli acidi abscissico e salicilico. Questi promotori sono, però, di limitata utilità poiché: l) i livelli naturali dei metabolitisegnale e la risposta al segnale dei vari tessuti o cellule possono variare; 2) i metaboliti-segnale modificano l'espressione di altri geni oltre a quello in esame; 3) i geni inducibili dai metaboliti descritti finora sono controllati da meccanismi complessi che comprendono fattori ambientali e condizioni di sviluppo. La costruzione in laboratorio di nuovi promotori che rispondano a induttori chimici, ma non ai prodotti presenti naturalmente nelle piante, potrebbe consentire di aggirare questi problemi.
Indirizzo delle proteine all'appropriato comparto subcellulare
Oltre a una adeguata espressione cellulospecifica o tessutospecifica, è quasi sempre necessario indirizzare le proteine esogene al comparto subcellulare appropriato. A parte poche proteine sintetizzate dai plastidi e dai mitocondri, le proteine sono codificate dal DNA nucleare e sintetizzate nel cito sol. Il trasporto dei polipeptidi verso la giusta destinazione subcellulare dipende dalla presenza di segnali di indirizzo e le proteine destinate a rimanere nel citosol mancano di tali segnali. Le proteine secrete dalla cellula o destinate al vacuolo sono sintetizzate come preproteine, contenenti un peptide con un segnale idrofobico N-terminale, che permette l'introduzione del polipeptide nascente nel lume del reticolo endoplasmatico (Chrispeels, 1991). Un sistema di vescicole in movimento trasporta le proteine dal reticolo endoplasmatico al complesso di Golgi e da qui all'esterno della cellula o al vacuolo. La secrezione non richiede ulteriori segnali, mentre per il trasporto al vacuolo sono necessari segnali aggiuntivi. Anche le proteine mitocondriali e plastidiali codificate dal genoma nucleare sono sintetizzate nel citosol come precursori, con specifici peptidi-segnale transienti, cioè che sono successivamente rimossi. Per indirizzare proteine esogene verso i mitocondri o i plastidi è sufficiente fondere la sequenza che codifica il peptide transiente opportuno a quella codificante la proteina presa in esame.
Sovraespressione o sottoespressione di geni specifici
Le piante possono essere trasformate per introdurre nuovi geni, e dunque proteine e funzioni, o per aumentare o limitare l'espressione di funzioni esistenti. L'aggiunta di un nuovo gene, solitamente, ha come risultato la presenza nella pianta transgenica dell 'mRNA da esso trascritto e della proteina tradotta da tale mRNA. Tuttavia, quando una pianta è trasformata con una copia aggiuntiva di un gene in essa già presente, il risultato può essere la sovraespressione o la sottoespressione di tale gene. Il fenomeno di sottoespressione è stato definito soppressione senso (sense suppression, per distinguere questo fenomeno da quello della antisense suppression, v. oltre) e il suo meccanismo di funzionamento non è ancora ben chiaro. Un esempio di soppressione senso è il risultato della trasformazione del pomodoro con una chimera codificante i trascritti parziali dei geni per gli enzimi poligalatturonasi e pectinaesterasi, sotto il controllo del promotore 35S di CaMV: il risultato è la simultanea sottoespressione di ambedue i geni nelle piante transgeniche. Una tecnica ampiamente usata per provocare la sottoespressione di un gene è la trasformazione di piante con un costrutto antisenso, in cui la sequenza del gene studiato è posta, rispetto al promotore, in senso opposto a quello naturale (antisenso anziché senso) (Mol et al., 1990). Lo scopo è di bloccare il flusso di informazione genetica dal DNA all'mRNA e poi alla proteina, tramite la sintesi di una catena di RNA antisenso, cioè complementare all'mRNA naturale trascritto dal gene di cui si vuole inibire l'espressione. La catena antisenso causa la riduzione quantitativa dell'mRNA naturale complementare e, di conseguenza, provoca una ridotta sintesi della proteina da esso codificata, ma anche in questo caso l'esatto meccanismo del fenomeno non è conosciuto. L'inibizione di geni vegetali tramite costrutti antisenso permette una riduzione estremamente variabile dell'espressione della proteina in oggetto, l'inibizione di attività enzimatiche codificate da famiglie multigeniche e, tramite l'uso di opportuni promotori, l'inibizione di enzimi in modo tessuto- e cellulospecifico. Una delle prime applicazioni di questa strategia antisenso è stata la riduzione della biosintesi di etilene nei frutti di pomodoro, al fine di estenderne il tempo di possibile permanenza sui banconi di vendita. Questo approccio è stato, inoltre, utilizzato per inibire la sintesi di amido in tuberi di patata.
l risultati ottenuti tramite le piante transgeniche devono sempre essere interpretati con cautela. lnnanzitutto, utilizzando le procedure standard di trasformazione, l'integrazione del nuovo gene avviene in un sito casuale del genoma della pianta ospite. Il sito può far parte di un qualunque gene della pianta, la cui interruzione, con la conseguente mancata espressione, può dar luogo a fenotipi inattesi e del tutto non correlati con l'attività del nuovo gene introdotto. Per escludere gli effetti di questa mutagenesi da inserzione è necessario analizzare diverse piante transgeniche derivanti da eventi indipendenti di trasformazione con lo stesso costrutto. La stessa procedura è necessaria per escludere gli effetti della variazione somaclonale, cioè della variabilità dovuta al fatto che le cellule che formano un callo sono piuttosto instabili geneticamente e, dunque, le singole piante rigenerate possono non essere geneticamente identiche. In secondo luogo, l'analisi delle piante transgeniche trasformate con costrutti antisenso è complicata dalla grande variabilità nell'espressione organo-specifica dei geni in oggetto. In terzo luogo, nelle generazioni di piante transgeniche successive alla prima, il gene inserito può essere reso silente e, quindi, la sua espressione può andare persa. Un altro problema è costituito dal fatto che quando un enzima è espresso ectopicamente (cioè in un tipo di cellule o un comparto subcellulare differenti da quelli naturali) i risultati nella fisiologia dell'ospite possono essere inattesi e difficili da interpretare. Infine, la sovraespressione di enzimi di origine vegetale non sempre risulta in un aumento dell' attività enzimatica nella pianta transgenica. L'enzima può essere soggetto a controllo da parte di metaboliti o a controllo postraduzionale (per esempio, tramite fosforilazione). Per annullare l'effetto dei meccanismi endogeni di controllo è possibile esprimere enzimi mutati aventi nuove caratteristiche di regolazione (Stark et al., 1992). In alternativa, agli enzimi vegetali possono essere preferiti i loro omologhi batterici o animali, se esistenti, che possono essere regolati in modo diverso e, quindi, sfuggire ai sistemi di controllo operanti nelle piante.
Piante transgeniche e produzione di nuovi amidi, oli, plastiche, enzimi e prodotti farmaceutici
Modificazione della qualità e della quantità dell'amido
L'amido, la maggiore riserva di carboidrati delle piante vascolari, è composto da polimeri del glucosio (glucani) a catena lineare (amilosio) e ramificata (amilopectina). A seconda della specie, l'amilosio costituisce dal 10 al 25% dell' amido. La lunghezza delle regioni lineari dell' amilopectina è compresa tra i 5 e i 100 residui di glucosio. Alcuni di questi polisaccaridi possono essere fosforilati, come nel caso dell'amido di patata. Le proprietà fisicochimiche importanti per l'uso tecnologico-alimentare dell'amido (la temperatura di gelatinizzazione, la retrogradazione e la viscosità) sono influenzate dalle dimensioni dei granuli e dalla loro omogeneità, dal rapporto fra amilosio e amilopectina, dalla distribuzione delle catene a basso e alto peso molecolare e dal contenuto di lipidi, ceneri e fosforo. L'amido che è attualmente in commercio è estratto da un numero limitato di piante, soprattutto mais e patata.
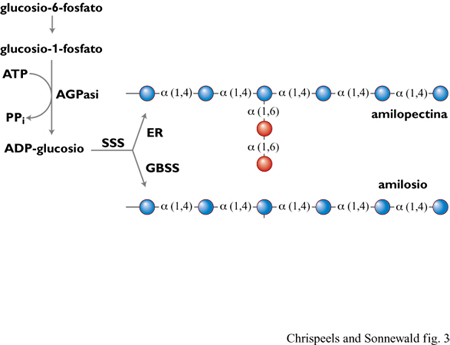
La biosintesi dell'amido richiede tre enzimi, presenti esclusivamente nei plastidi: ADP - gluco siopirofosforilasi (AGPasi), amidosintetasi, nelle due forme, solubile (SSS, Soluble Starch Synthase) e associata ai granuli (GBSS, GranuleBound Starch Synthase), ed enzima di ramificazione (fig. 3). A partire da ADP-glucosio, sintetizzato dal glucosio-I-fosfato grazie all'enzima AGPasi, l'enzima amidosintetasi catalizza la formazione di un legame α(1-4) fra il residuo di glucosio e una catena di glucano preesistente, forse legata a una proteina; la natura di questa proteina e la via bio sintetica che porta al suo legame con i residui di glucosio sono, tuttavia, sconosciute nei vegetali. Usando la catena di glucano α(I-4) come substrato, l'enzima di ramificazione catalizza la formazione di ramificazioni α(1-6) nei polimeri di amido (Martin e Smith, 1995).
Sono state studiate, in alcune specie, mutazioni naturali che alterano il rapporto quantitativo tra amilosio e amilopectina. L' enzima amidosintetasi si trova sia associato ai granuli di amido sia in forma solubile. Mutanti privi di amilosio sono caratterizzati dalla mancanza di amidosintetasi associata ai granuli di amido, mentre la mancanza di isoforme dell' enzima di ramificazione causa la formazione di amido ad alto contenuto di amilosio (Martin e Smith, 1995). Con l'eccezione dei mutanti waxy (letteralmente: cero so ) di mais, che sono privi di amilosio poiché mancano di GBSS, gli altri mutanti della sintesi di amido non sono ampiamente sfruttati commercialmente, poiché la loro significativa riduzione di resa in campo non è compensata dalla possibilità di vendita a un prezzo proporzionalmente più elevato. Grazie alla facilità di trasformazione della patata, gli approcci molecolari per alterare la qualità o quantità di amido si sono concentrati su questa pianta (tab. 2).
L'importanza dell'enzima AGPasi nella sintesi dell'amido è stata dimostrata tramite l'uso di costrutti antisenso. Una riduzione dell'attività di questo enzima al 5% del normale causa la formazione di tuberi quasi totalmente mancanti di amido (meno del 5% di quello presente normalmente). Il crollo nell'accumulo di amido è accompagnato da un aumento di zuccheri solubili (soprattutto saccarosio e glucosio). L'esperimento opposto è stato fatto da D.M. Stark e collaboratori (1992), esprimendo in piante di patata transgeniche il gene glgC di Escherichia coli, che codifica un mutante di AGPasi con proprietà allosteriche alterate, sotto il controllo di un promotore tubero-specifico. La localizzazione guidata dell' enzima batterico nei plastidi causa, nei tuberi delle piante transgeniche, un aumento nella quantità di amido fino al 50% in più del normale.
I tentativi di modificare la qualità dell' amido si sono focalizzati soprattutto sul rapporto fra amilosio e amilopectina. Sono state seguite due strategie: la repressione-antisenso di geni endogeni per la sintesi di amido e la sovraespressione di geni batterici coinvolti nella sintesi di glicogeno, un polimero simile all'amilopectina. Shewmaker e collaboratori (1994) hanno introdotto nella patata un gene chimerico, formato dal promotore tubero-specifico della patatina, una proteina endogena, dalla regione genica che codifica il peptide-segnale transiente per l'indirizzamento ai plastidi della subunità minore dell'enzima ribulosiobifosfatocarbossilasi di soia e dal gene glgA di E. coli che codifica l'enzima glicogenosintetasi. I tuberi delle piante transgeniche hanno un ridotto accumulo di amido e un aumento di zuccheri solubili. La grandezza dei granuli, il rapporto amilosio/amilopectina e il contenuto di fosforo dell'amido sono anch'essi ridotti e l'amilopectina è più ramificata che nei tuberi di piante non trasformate. Questi cambiamenti alterano le proprietà fisico-chimiche dell'amido, quali la viscosità della pasta e la gelatinizzazione.
La funzione delle diverse isoforme dell'enzima amidosintetasi è stata determinata usando una strategia antisenso. Nella patata, l'isoforma più studiata di questo enzima è GBSSI (Granule-Bound Starch Synthase I, amidosintetasi associata ai granuli I). Un costrutto antisenso di questa isoforma, introdotto nella patata, causa l'inibizione completa dell'enzima e la formazione di amido privo di amilosio. Una seconda isoforma GBSSII (Granule-Bound Starch Synthase lI) è stata recentemente donata da una banca di cDNA da tuberi e il suo contributo alla sintesi dell'amido è stato studiato in piante trasformate con il costrutto antisenso. Sebbene il livello di trascritto e della proteina corrispondente siano marcatamente ridotti, non è stata osservata nessuna differenza apparente nel contenuto di amido o nel rapporto amilosio/amilopectina. La funzione di GBSSII rimane, perciò, ancora ignota.
Oltre alle isoforme di amidosintetasi legate ai granuli d'amido, sono state identificate tre forme solubili di questo enzima in estratti di tuberi di patata. Una di esse è stata donata e, mediante la tecnica antisenso, se ne è ridotta l'espressione a livelli non misurabili. Il risultato è una modificazione della struttura dell'amido. Flipse e collaboratori (1996) hanno utilizzato una strategia combinata per alterare la qualità dell'amido. Un mutante di patata mancante di amilosio (amI, amylosefree) è stato trasformato con costrutti senso o antisenso del gene codificante l'enzima di ramificazione, allo scopo di modificare il grado di ramificazione dell'amilopectina. Con entrambi i costrutti si è avuta come risultato la completa soppressione dell'mRNA e dell'enzima endogeni. Il crollo dell'attività enzimatica non si è tradotto in un aumento del contenuto di amilosio o in un'alterazione delle ramificazioni di amilopectina, ma in un cambiamento delle proprietà fisico-chimiche. Dunque, l'amido delle piante trasformate è diverso da quello del mutante amI, anche se le basi strutturali di tali differenze non sono ben chiare. Nell'insieme, i risultati sopra descritti dimostrano che è possibile alterare sia la qualità sia la quantità di amido. Malgrado ciò, fatta eccezione per GBSSI, non è ancora stato possibile assegnare chiaramente una funzione alle altre isoforme di amidosintetasi o all' enzima di ramificazione presenti in tuberi di patata. Ulteriori ricerche sulla formazione dei granuli potranno fornire, in un futuro che si spera vicino, le basi scientifiche per la produzione di amidi 'su misura'.
Semi con nuovi grassi nelle piante transgeniche
I grassi e gli oli, collettivamente chiamati trigliceridi o triacilgliceroli (TAG), sono esteri organici del glicerolo con tre molecole di acidi grassi. Nei semi e in altri organi della pianta, dove sono presenti in abbondanza, i TAG hanno soprattutto funzione di riserva. La lunghezza della catena di carbonio degli acidi grassi varia fra gli 8 e i 20 residui, ma le catene a 16 o 18 residui sono le più comuni nelle piante superiori. Si conoscono più di 200 tipi di acidi grassi di origine vegetale, ma solo quelli contenenti gli acidi laurico, palmitico, stearico, oleico, linoleico e linolenico sono sfruttati commercialmente. Circa il 90% dell'olio vegetale prodotto è usato per l'alimentazione umana, il rimanente è utilizzato per fini non alimentari. Il maggiore mercato nonalimentare per gli oli vegetali è quello della produzione di saponi, detergenti e altri tensioattivi. La lunghezza delle catene degli acidi grassi e la loro saturazione sono proprietà importanti dei TAG e ne determinano l'uso specifico. Per esempio, le catene di media lunghezza hanno proprietà ideali come tensioattivi. Attualmente, le maggiori fonti di acidi grassi di media lunghezza per uso commerciale sono gli oli di cocco e di palma.
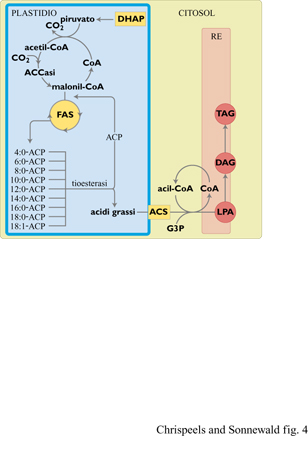
Nelle cellule animali, la biosintesi degli acidi grassi avviene nel citosol, ma nelle piante superiori questo processo è localizzato nei plastidi (fig. 4). Il primo passaggio obbligato nella biosintesi degli acidi grassi è la formazione di malonil-coenzima A (malonil-CoA) a partire da acetilCoA e bicarbonato, reazione catalizzata dall'enzima acetilCoA-carbossilasi (ACCasi). Durante i cicli di allungamento di questa corta catena di acido grasso, catalizzati da una serie di enzimi che, nell'insieme, costituiscono il complesso acido grasso sintasi (FAS, Fatty Acid Synthase), le catene aciliche sono legate covalentemente a una proteina trasportatrice delle catene aciliche (ACP, Acyl Carrier Protein). Le reazioni di allungamento possono terminare con il trasferimento della catena al glicerolo-3-fosfato (G3P) e questi acilgliceroli sono utilizzati per la sintesi dei lipidi delle membrane plastidiali. In alternativa, tioesterasi specifiche possono idrolizzare l'acil-ACP neosintetizzato, originando un acido grasso libero e ACP. Gli acidi grassi liberi sono trasportati fuori dai plastidi ed esterificati ad acil-CoA grazie a una acil-CoA-sintetasi localizzata sulla membrana esterna dei plastidi. Gli acil-CoA sono trasportati nel reticolo endoplasmatico, probabilmente tramite proteine di trasporto, e in questo comparto sono utilizzati per la sintesi di TAG di riserva e fosfolipidi delle membrane non-plastidiali. La prima reazione che avviene nel reticolo endoplasmatico è la formazione di acido lisofosfatidico (LPA) a partire da acil-CoA e glicerolo-3-fosfato, catalizzata dall' enzima glicerolo- 3 - fosfatoaciltransferasi. In seguito, l'acido lisofosfatidico è convertito a diacilglicerolo (DAG) attraverso l'azione degli enzimi acido lisofosfatidicoaciltransferasi (che produce acido fosfatidico) e acido fosfatidicofosfatasi. Il diacilglicerolo può essere convertito in un fosfolipide (per es. fosfatidilcolina) o usato per la formazione di TAG. Oltre ai processi appena descritti, gli acidi grassi possono subire reazioni di desaturazione e idrossilazione, durante la loro sintesi o dopo il trasporto al reticolo endoplasmatico.
La lunghezza delle catene di acido grasso è determinata dall' azione di tioesterasi specifiche che idrolizzano l'acido grasso-ACP. Le tioesterasi della maggior parte delle piante coltivate idrolizzano 16:0-ACP (16:0 indica che l'acido grasso ha 16 atomi di carbonio e 0 doppi legami) e 18:1ACP; tuttavia le tioesterasi di altre piante idrolizzano acilACP con catena di media lunghezza, causando l'accumulo di acidi grassi di media lunghezza negli oli dei semi. Il ruolo cruciale delle tioesterasi è stato dimostrato isolando da Umbellularia californica il gene codificante una tioesterasi che idrolizza specificamente il lauril-ACP (12:0-ACP) e introducendolo in piante transgeniche di Arabidopsis e di colza (V oelker et al., 1992). L'olio isolato dai semi di queste piante transgeniche contiene dal 25 al 40% di acido laurico, fornendo una fonte alternativa di acidi grassi a media lunghezza rispetto agli oli di cocco e di palma. Recentemente lo stesso gruppo di ricerca ha isolato da Cuphea hookeriana un clone di cDNA che codifica una acil-ACP-tioesterasi. l semi di questa pianta accumulano soprattutto gli acidi grassi con catene 8:0 e 10:0. L'introduzione del clone di cDNA in piante transgeniche di colza provoca l'accumulo di grassi con catene 8:0 e 10:0 e una riduzione di acido linoleico (18:2) e linolenico (18:3). Oltre all'attività delle tioesterasi, anche l'attività dell'acil-CoA-desaturasi è stata alterata mediante trasformazione, al fine di modificare i livelli di insaturazione delle catene.
Manipolazione della struttura della lignina e digeribilità degli alimenti per animali
La lignina, il secondo composto organico più abbondante presente sulla Terra dopo la cellulosa, è un polimero complesso derivante dalla polimerizzazione ossidativa di tre composti fenolici: gli alcoli coniferilico, sinapilico e cumarilico, che differiscono tra di loro nel numero di gruppi metilici. La lignina è localizzata principalmente nei tessuti vascolari delle piante, dove agisce come materiale di rinforzo e riempimento fra le fibrille di cellulosa nelle spesse pareti secondarie di taluni tipi di cellule. Oltre a questo ruolo di rinforzo, la lignina partecipa alla difesa della pianta dall'attacco di agenti patogeni e dalla predazione da parte di erbivori. Gli alimenti per animali ad alto contenuto di lignina sono generalmente meno digeribili. La composizione della lignina varia da pianta a pianta e dipende dalla presenza e dalla specificità degli enzimi O-metiltransferasi (OMT), che determinano il tipo di monomero prodotto. l diversi acidi fenolici sintetizzati da OMT sono attivati dall'enzima 4-cumarato-CoA-ligasi, che forma un tioestere mediante il legame al CoA, e sono successivamente ridotti mediante due reazioni catalizzate dagli enzimi cinnamil-CoA-reduttasi e cinnamilalcoldeidrogenasi, generando i tre alcoli sopra citati che costituiscono i monomeri di lignina. l primi tentativi di modificare la struttura della lignina sono stati condotti mediante l'espressione senso o antisenso delle OMT, nel tentativo di alterare la ripartizione dei flussi bio sintetici fra le vie che conducono alla produzione dei differenti monomeri e mediante l'espressione antisenso di cinnamilalcoldeidrogenasi, con lo scopo di ridurre la produzione degli alcoli e permettere, perciò, l'accumulo delle rispettive aldeidi. Le soppressioni antisenso di OMT l e di cinnamilalcoldeidrogenasi in piante transgeniche di tabacco non hanno portato a cambiamenti nella quantità di lignina ma ne hanno alterato la composizione, aprendo la strada alla manipolazione in vivo della qualità di questo composto.
Produzione di plastica biodegradabile e di polimeri del fruttosio

Oltre a cercare di manipolare alcuni dei prodotti endogeni delle piante, si è anche tentato di guidare il flusso dei composti del carbonio verso la produzione di nuove biomolecole. Uno dei primi esempi della sintesi di un nuovo biopolimero, in piante transgeniche, è stata la produzione di acido poli-3-idrossibutirrico (PHB, Poly-3-Hydroxy Butyrate) in Arabidopsis (fig. 5). A partire da acetil-CoA, la sintesi di PHB richiede tre reazioni enzimatiche. La prima è la condensazione di due molecole di acetil-CoA a formare acetoacetil-CoA ed è catalizzata dall'enzima 3-chetotiolasi. In seguito, una acetoacetil-CoA-reduttasi NADPH-dipendente riduce l'acetoacetil-CoA a 3-idrossibutirril-CoA, che è infine polimerizzato a PHB dall'enzima PHB-sintasi.
La produzione di PHB è stata studiata nel citosol e nei plastidi di piante di Arabidopsis. Nelle cellule vegetali, acetil-CoA è presente nel citosol, nei plastidi, nei mitocondri e nei perossisomi. Poiché nel citosol è presente anche acetoacetil-CoA, per permettervi la produzione di PHB è stato sufficiente esprimere gli enzimi reduttasi e sintasi della suddetta via metabolica. Le piante transgeniche esprimenti ambedue gli enzimi accumulano quantità molto limitate di PHB (0,1 % del peso secco della pianta) e subiscono un forte rallentamento della crescita. Questo rallentamento può essere dovuto a un abbassamento del livello di acetil-CoA citosolico, fino a quantità insufficienti per sostenere la sintesi di composti essenziali alla crescita. I plastidi sono ricchi di acetil-CoA, utilizzato per la biosintesi degli acidi grassi e possono, dunque, essere un comparto subcellulare conveniente per la sintesi di PHB. A questo scopo, si sono dovuti indirizzare verso i plastidi di piante transgeniche tutti e tre gli enzimi della via di sintesi del polimero. La quantità di PHB prodotto nelle piante trasformate in questo modo raggiunge un massimo del 14% del peso secco delle foglie senescenti, senza avere alcuna conseguenza sulla crescita della pianta (Nawrath et al., 1994). Da questi risultati si possono trarre due conclusioni principali: l) il flusso dei composti del carbonio può essere indirizzato verso la produzione di una quantità significativa di polimeri non vegetali; 2) è necessario considerare con attenzione la compartimentazione subcellulare delle vie metaboliche.
In modo simile, i tentativi di produrre fruttani (polimeri del fruttosio) in piante che normalmente non accumulano questi composti ha rivelato differenze dovute alla scelta dei comparti di accumulo. I fruttani vegetali sono polimeri del fruttosio (da 5 a 60 residui), terminanti con un singolo residuo di D-glucosio unito da un legame β(2-1). I fruttani sono ampiamente diffusi nel regno vegetale, ma sono prodotti anche da molti microrganismi. Tuttavia, nei batteri la lunghezza della catena prodotta è solitamente molto maggiore (più di 100.000 residui di fruttosio) che nelle piante. Per verificare se il flusso biosintetico dei composti del carbonio possa essere guidato verso la sintesi di fruttani, il gene SacB di Bacillus subtilis, che codifica l'enzima che catalizza la formazione di polimeri di fruttosio utilizzando saccarosio come substrato, è stato introdotto nel tabacco e nella patata, due piante che normalmente non accumulano fruttani. Il gene batterico è stato modificato per permettere la secrezione o il trasporto nel vacuolo dell'enzima codificato. La secrezione dell' enzima ha causato nelle piante transgeniche una grave necrosi, mentre il trasporto nel vacuolo ha permesso l'accumulo di fruttani (fino al 30% del peso secco delle foglie) senza provocare visibili alterazioni fenotipiche nelle piante (Van der Meer et al., 1994).
Applicazione delle piante nella produzione di enzimi di uso industriale
Dal punto di vista commerciale, le piante potrebbero anche diventare un conveniente sistema di produzione di enzimi e altre proteine eterologhe. La produzione di enzimi in grande scala è attualmente ottenuta tramite microrganismi cresciuti in fermentatori. Tuttavia, questa tecnologia comporta una serie di svantaggi e non tutte le proteine eucariotiche mantengono le loro funzioni se prodotte in microrganismi. Possono insorgere problemi di sterilità e di contaminazione da tossine e le modificazioni postraduzionali delle proteine, spesso necessarie al loro corretto funzionamento, possono essere differenti fra i microbi e gli eucarioti superiori. Inoltre, la sovraespressione di proteine nei batteri può dar luogo alla formazione dei cosiddetti corpi di inclusione, dai quali può essere molto difficile solubilizzare la proteina in esame. Questi problemi hanno indotto i biotecnologi a esaminare il possibile impiego di piante transgeniche come bioreattori per la produzione di enzimi di interesse industriale. La produzione nelle piante può presentare una serie di vantaggi rispetto a quella nei microrganismi, come batteri o lieviti. Poiché le piante sono fotoautotrofe (utilizzano la luce per produrre l'energia di cui hanno bisogno e non richiedono sostanze organiche nel terreno di crescita), i costi energetici, necessari alla produzione di enzimi eterologhi nelle piante, risultano molto contenuti. Inoltre, la superficie di coltivazione utilizzata per una determinata pianta è agevolmente modificabile, in modo da adattare facilmente, in termini quantitativi, la produzione alla domanda del prodotto. Le piante operano alcune delle modificazioni postraduzionali delle proteine che avvengono negli animali e, perciò, è più probabile che producano enzimi stabili e in forma attiva. Inoltre, le proteine espresse nei semi possono mantenere la propria attività durante la conservazione prolungata degli stessi. È possibile produrre proteine in piante che sono già normalmente coltivate e sottoposte a trasformazioni dall'industria alimentare, o da altre industrie, e purificare l'eventuale enzima eterologo come prodotto industriale secondario. Un altro vantaggio risiede nella possibilità di modificare sostanze normalmente prodotte dalla pianta esprimendo un enzima eterologo. Tuttavia, affinché la produzione di proteine eterologhe nelle piante sia utile dal punto di vista commerciale, è necessario che una serie di prerequisiti sia soddisfatta. In particolare: l) la pianta deve produrre una notevole quantità dell'enzima eterologo; 2) l'enzima deve essere stabile all'interno della pianta e 3) non deve perdere attività enzimatica durante il raccolto e le eventuali successive procedure di purificazione; 4) se la purificazione è necessaria, essa deve essere semplice ed economica; 5) la crescita della pianta non deve subire conseguenze negative a causa dell'espressione dell'enzima eterologo. Numerosi enzimi di interesse industriale sono stati espressi in piante transgeniche (Goddijn e Pen, 1995), nella maggior parte dei casi sotto il controllo di un promotore costitutivo. La quantità di proteina eterologa ottenuta varia dallo 0,1 al 4,0% delle proteine solubili totali della pianta. Malgrado i successi ottenuti, i costi di purificazione delle proteine ricombinanti dagli estratti vegetali rimangono uno dei problemi più critici di questa tecnologia. Per alcune applicazioni, tuttavia, può non essere necessaria la purificazione dell'enzima e può essere sufficiente ottenere materiale vegetale essiccato, contenente notevoli quantità della proteina ricombinante. Una fitasi di origine fungina è stata espressa in semi di tabacco usando un promotore seme-specifico e si è dimostrato che pollame sottoposto a una dieta integrata con i semi transgenici arricchiti di fitasi utilizza in modo più efficiente il fosforo contenuto nel mangime. Lo scopo di questo progetto è di ridurre la quantità di fosforo che deve essere aggiunto al mangime per animali, in modo da evitare l'inquinamento ambientale da fosfato presente negli escrementi.
Produzione di vaccini contro importanti malattie infettive
Vaccini a basso costo contro le malattie infettive (per esempio, il colera) sarebbero di enorme aiuto ai paesi poveri del terzo mondo. Avendo bilanci di spesa per la sanità di pochi dollari pro capite, questi paesi non possono permettersi di comprare i vaccini prodotti dalla ricerca scientifica dei paesi ricchi. Le piante transgeniche potrebbero essere una soluzione ideale per l'immunizzazione, sia attiva sia passiva. L'immunizzazione attiva consiste nella stimolazione del sistema immunitario dell'individuo. La stimolazione della produzione di immunoglobuline A (lgA) da parte del sistema immunitario delle mucose è ottenuta più facilmente attraverso la somministrazione orale dell'antigene che attraverso la somministrazione per iniezione sottocutanea o endovenosa. L'immunizzazione passiva è una strategia totalmente differente, in cui l'individuo riceve anticorpi che sono stati prodotti al di fuori del proprio corpo. Questi anticorpi potrebbero benissimo essere prodotti in piante, invece che nei costosi sistemi di colture cellulari utilizzati attualmente.
Le diarree provocate da infezioni batteriche sono una delle cause principali di mortalità infantile nei paesi in via di sviluppo. Queste malattie si sviluppano quando batteri come Vibrio cholerae, o ceppi enterotossici di Escherichia coli, colonizzano l'intestino tenue e producono enterotossine che si legano ai gangliosidi presenti sulla superficie delle cellule epiteliali. Le enterotossine sono proteine multimeriche, la cui struttura è stata determinata di recente e i cui geni sono stati donati. Il legame dell'enterotossina all'epitelio può essere inibito da anticorpi prodotti contro il polipeptide che interagisce con i gangliosidi. Questa inibizione è alla base della strategia di sviluppo dei vaccini orali, che stimolano la mucosa a produrre anticorpi i quali, secreti nell'apparato digerente, prevengono l'attacco delle tossine all' epitelio.
l primi esperimenti condotti con lo scopo di combattere ceppi di E. coli enterotossici hanno dato risultati molto promettenti. La tossina L T (heat-labile enterotoxin, tossina labile al calore) prodotta da questo ceppo di E. coli è formata da un polipeptide (LT-A) avente un peso molecolare di 27 kDa (subunità A), e da cinque polipeptidi (LT-B) con un peso di 11,6 kDa (subunità B). Il legame con i gangliosidi è mediato dalle subunità B. Lo scopo degli esperimenti è stato, perciò, di esprimere LT -B in piante transgeniche e di stabilire se potesse funzionare come vaccino orale. Haq e collaboratori (1995) hanno espresso LT-B in foglie di tabacco e in tuberi di patata transgenici. Sebbene sia stato utilizzato un forte promotore, i tessuti delle piante transgeniche accumulavano quantità molto basse dell'antigene (5 μg e 110 μg per grammo di proteina totale di foglia di tabacco o di microtubero di patata, rispettivamente). Tuttavia, i livelli di accumulo sono stati sufficienti per stimolare una risposta immunitaria in topi nutriti con queste piante. Un topo nutrito con 5 grammi di tubero transgenico riceve una dose di l5÷20 JLg di antigene LT-B ricombinante (durante un lasso di tempo di 2 ÷ 6 ore) e produce anticorpi specifici contro LT-B nel siero (lgG) e nelle mucose (lgA). Sarà necessario stabilire se questi anticorpi siano in grado o meno di proteggere gli animali dalle infezioni batteriche. Se la risposta sarà positiva, i ricercatori dovranno cercare un metodo migliore di somministrazione del vaccino, poiché le patate crude non sono un cibo appetibile per l'uomo. Il gruppo di ricerca sta puntando la sua attenzione sulla banana, un frutto tropicale facile da diffondere, ampiamente coltivato e appetibile, che sarebbe un veicolo ideale per un vaccino orale.
L'uso di virus vegetali come sistemi ad alta resa per la presentazione di epitopi eterologhi ha aperto la possibilità di produrre elevate quantità di nuovi vaccini nelle piante. Utilizzando la malaria come malattia modello, il virus del mosaico del tabacco (TMV) è stato modificato mediante ingegneria genetica al fine di ottenere la presentazione di epitopi antigenici del plasmodio della malaria sulla superficie del virus. In seguito a infezione di piante con TMV, le particelle virali potrebbero essere ottenute in quantità sufficienti dai tessuti vegetali infettati sistemicamente.
Come accennato sopra, le piante transgeniche potrebbero anche essere usate per produrre anticorpi da utilizzare in strategie di immunizzazione passiva. Tenendo presente questo scopo, sono stati espressi nelle piante anticorpi modificati tramite ingegneria genetica. Un esempio promettente è il successo ottenuto nell'espressione di un anticorpo che riconosce il fattore di adesione alla superficie cellulare di mutanti di streptococco, l'agente che causa le carie dentali (Ma et al., 1995). In questo caso, le piante sarebbero particolarmente adatte come fonte di anticorpi, poiché il trattamento della carie richiede l'uso di grandi quantità di immunoglobuline per ogni paziente.
Manipolazione delle proprietà agronomiche e di quelle successive alla raccolta
Manipolazione della produzione fotosintetica del saccarosio

L'assimilazione foto sintetica del carbonio è uno dei fattori determinanti per la crescita delle piante e per la resa dei raccolti. I prodotti primari della foto sinte si, i triosofosfati, sono utilizzati nel cloroplasto per la biosintesi dell'amido o esportati nel citosol come precursori per la sintesi del saccarosio. Le foglie mature producono con la foto sintesi composti organici in eccesso rispetto ai propri bisogni metabolici. Questi composti sono esportati verso gli organi non foto sintetizzanti o che, in ogni caso, non producono abbastanza composti organici per le proprie necessità metaboliche o per la propria attività di accumulo di riserve. Poiché, nella maggior parte delle piante, il saccarosio è il principale composto in cui viene indirizzato il carbonio fissato dalla fotosintesi, le attività enzimatiche coinvolte nella produzione fotosintetica del saccarosio sono particolarmente importanti per la resa dei raccolti. Si ritiene che le reazioni chiave di regolazione della bio sintesi del saccarosio siano l'interconversione di fruttosio-l,6-difosfato (FBP) e fruttosio-6- fosfato (F6P) e la formazione di saccarosio-6- fosfato (S6P) da UDP-glucosio (UDPG) e F6P (fig. 6). Per scoprire se queste siano realmente le reazioni chiave di regolazione nel flusso metabolico e nella partizione del carbonio foto sintetico, gli enzimi che catalizzano queste reazioni sono stati sovraespressi o sottoespressi in piante transgeniche. L'interconversione di FBP e F6P coinvolge tre attività enzimatiche: l'enzima fruttosio-l,6-difosfatasi citosolica (citFBPasi) catalizza la reazione da FBP a F6P mentre l' enzima fosfofruttochinasi (PFK) catalizza la reazione nella direzione opposta; un terzo enzima, pirofosfato frutto sio-6- fosfato-l- fosfotrasferasi (PFP), catalizza la reazione in ambedue le direzioni. Un solo enzima, la saccarosiofosfatosintasi (SPS), è invece responsabile della formazione di S6P, che è poi convertito a saccarosio attraverso l'azione dell' enzima saccarosiofosfatofosfatasi (SPP). Per superare la regolazione esercitata a livello della interconversione di FBP e F6P sono state tentate diverse strategie. Le attività degli enzimi FBPasi e PFP sono sotto controllo allosterico da parte del metabolita-segnale fruttosio2,6-difosfato (F2,6BP) che stimola la PFP e inibisce la FBPasi. Per determinare l'effettiva importanza di F2,6BP nella regolazione del metabolismo del carbonio, è stata introdotta in piante transgeniche di tabacco una forma modificata dell'enzima fosfofrutto- 2-chinasi, isolata dal fegato di ratto (Scott et al., 1995). La forma modificata utilizzata non ha attività fosfatasica, mentre mantiene l'attività chinasica. Le piante transgeniche ottenute posseggono un livello di F2,6BP che è 2 o 3 volte più elevato del normale: ciò inibisce il flusso di assimilati verso la formazione di saccarosio e lo stimola verso la formazione di amido, confermando il ruolo regolatorio ipotizzato per F2,6BP.
Come risultato della defosforilazione di FBP da parte di PFP, in presenza di fosfato inorganico, si formano F6P e pirofosfato inorganico. Potrebbe, perciò, essere possibile spostare l'equilibrio di questa reazione verso F6P rimuovendo pirofosfato dall'equilibrio e, in questo modo, contrastare il controllo esercitato da F2,6BP. Inoltre, viene prodotto pirofosfato anche durante la conversione di glucosio-l- fosfato (GlP) a UDP-glucosio (UDPG), reazione che produce l'altro substrato per la sintesi di S6P e che è catalizzata dall' enzima UDP-glucosiopirofosforilasi (UGPasi). Perciò, introducendo in piante transgeniche una nuova pirofosfatasi che idrolizzi pirofosfato nel cito sol, il flusso metabolico del carbonio dovrebbe essere stimolato verso la formazione di saccarosio. Come atteso da queste premesse, l'espressione costitutiva dell'enzima pirofosfatasi citosolica di E. coli, in foglie di piante transgeniche di tabacco e patata, causa un incremento (di un fattore che varia da 3 a 4) nel rapporto tra zuccheri solubili e amido accumulati. Inaspettatamente, tuttavia, queste piante transgeniche mostrano crescita ritardata e ridotta formazione di radici, suggerendo che, oltre ad aumentare la produzione di saccarosio, il nuovo enzima altera l'esportazione di prodotti della fotosintesi dalle cellule del mesofillo agli altri tessuti della pianta (Lerchl et al., 1995).
L'effetto di una riduzione di citFBPasi è stato studiato in un mutante di Flaveria linearis, in cui l'attività di questo enzima è ridotta del 75% (Micallef e Sharkey, 1996), e in piante transgeniche di patata che, esprimendo un costrutto antisenso del gene codificante l'enzima, hanno un'attività di citFBPasi che varia tra il 9 e il 55% del normale (Zrenner et al., 1996). Un'inibizione dell'attività al di sotto del 20% del normale causa un accumulo dell'acido-3-fosfoglicerico (3-PGA), di triosofosfati e di FBP nelle foglie. Il risultato è che la velocità di assimilazione del carbonio, misurata come scambio di gas, è saturata con una quantità di luce più bassa e la velocità di fotosintesi in condizioni saturanti di luce e COz è ridotta. La misurazione dei flussi fotosintetici del carbonio, tramite marcatura radioattiva di frammenti di foglie con 14COZ' ha rivelato una riduzione del 53-65% nella sintesi di saccarosio e solo del 18-24% in quella di amido. Malgrado questi cambiamenti, le concentrazioni di saccarosio, allo stato stazionario, nelle foglie mature non sono alterate. L'amido si accumula a un livello più che triplo rispetto al normale ed è degradato durante la notte. Questi risultati forniscono un forte sostegno all'ipotesi che gli esosi o gli esosofosfati prodotti dalla degradazione dell'amido sono esportati dai cloroplasti, aggirando, in questo modo, la limitazione nella sintesi di saccarosio causata dall'inibizione di citFBPasi. In accordo con questa ipotesi, la crescita delle piante e la resa di tuberi rimangono inalterate. Da questi dati si può concludere che una ridotta capacità di produzione foto sintetica di saccarosio può essere efficacemente compensata senza effetti negativi sulla resa dei raccolti in serra o in camere di crescita, poiché la pianta può modificare la propria strategia di esportazione di carbonio dal sito della foto sintesi.
Diverse evidenze sperimentali suggeriscono che l' enzima saccarosiofosfatosintasi (SPS) catalizza la reazione limitante la velocità di sintesi del saccarosio (Stitt e Sonnewald, 1995). lnnanzitutto, variazioni dell'attività di SPS sono spesso correlate a variazioni nella velocità di sintesi e traslocazione del saccarosio. In secondo luogo, l'attività di SPS può essere modulata da diversi meccanismi che interagiscono in modo gerarchico come: l) la regolazione allosterica da metaboliti; 2) la fosforilazione postraduzionale reversibile della proteina; 3) la regolazione sviluppo-specifica dell'espressione del gene che codifica SPS, che determina la quantità di enzima prodotto. Nelle foglie di spinacio e nei tuberi di patata, l'enzima è attivato dal glucosio-6fosfato e inibito dal fosfato inorganico. Inoltre, l'enzima di spinacio esiste in due forme che hanno cinetiche di attività distinte e un diverso grado di fosforilazione (Huber e Huber, 1992). La forma altamente fosforilata è caratterizzata da bassa affinità per il substrato (fruttosio-6- fosfato) e l'effettore (glucosio-6- fosfato), mentre la forma meno fosforilata ha alta affrnità per le due molecole.
Due strategie, la sovraespressione eterologa e la repressione antisenso, sono state usate per studiare l'effetto di alterazioni dell' attività di SPS sulla crescita della pianta. Gli scienziati della società Calgene, in California, hanno trasformato piante di pomodoro con una sequenza codificante SPS di mais, posta sotto il controllo di un promotore foglia-specifico di tabacco o del promotore costitutivo 35S CaMV. Il livello di attività di SPS nelle piante transgeniche è risultato fino a sei volte maggiore del normale (Galtier et al., 1995). Nelle foglie mature l'elevata attività di SPS provoca una riduzione nell'accumulo dell'amido e un aumento del saccarosio (v. figura 6). Questo è dovuto a una stimolazione nella partizione del flusso di carbonio verso la produzione di saccarosio; si è quindi concluso che SPS gioca un ruolo cruciale nella partizione del flusso di carbonio. Le piante transgeniche sovraesprimenti SPS, coltivate in camere di crescita, non mostrano sofferenza e la produzione totale di materia secca non è alterata; tuttavia, il rapporto radice/parte aerea è modificato in favore della parte aerea (Galtier et al., 1995). L'incremento di attività dell'SPS non causa un aumento significativo nella velocità di fotosintesi all'aria, ma le piante trasformate con SPS, se cresciute a concentrazioni di anidride carbonica atmosferica elevata, mostrano una velocità di foto sintesi più elevata del 20% rispetto ai controlli non trasformati. Si può, perciò, concludere che un aumento nella capacità di bio sintesi del saccarosio può modificare l'acclimatazione ad alte concentrazioni di anidride carbonica. Si è però osservato che tale incremento nella velocità di fotosintesi non causa un aumento significativo nella produzione di frutti. Non si è potuta, perciò, stabilire una stretta correlazione tra velocità di fotosintesi nelle foglie e rendimento produttivo, in modo simile a quanto osservato nel caso discusso sopra delle piante di patata trasformate con un costrutto antisenso di citFBPasi.
L'effetto di una diminuzione nell'attività dell'SPS è stato studiato in piante di patata trasformate con un gene SPS chimerico antisenso o senso (cosoppressione). In foglie mature si è ottenuta una riduzione di attività dell' enzima del 60-70%. La riduzione nella quantità di enzima prodotto è in parte compensata dal suo stato più attivo. La riduzione di attività enzimatica non causa una riduzione della velocità di fotosintesi, tuttavia il rapporto fra saccarosio e amido è modificato in favore di quest'ultimo. Il ridotto accumulo di zuccheri solubili è causato da una riduzione quantitativa dell' aliquota di carbonio indirizzata verso la formazione del saccarosio. La ridotta attività di SPS nelle piante transgeniche non causa alcun fenotipo visibile e non influenza la produzione di tuberi. Si può concludere che, nel complesso, la sovra- e sottoespressione di SPS provoca i cambiamenti attesi per quanto riguarda la partizione del carbonio. Tuttavia, rimane dubbio se la velocità di produzione foto sintetica di saccarosio sia il fattore che determina il rendimento finale dei raccolti: altri processi, come il caricamento (loading) del floema o l'utilizzazione di saccarosio nei tessuti non produttori potrebbero costituire i fattori limitanti.
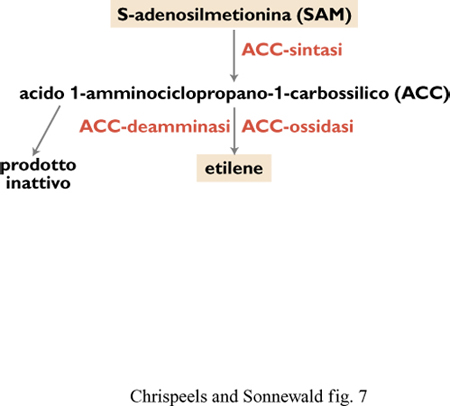
Riduzione delle perdite di frutti e ortaggi dopo la raccolta Il processo di maturazione dei frutti è associato ad alterazioni di colore, consistenza, sapore, aroma e suscettibilità ad agenti patogeni. Il ben noto intenerimento dei frutti durante la maturazione è accompagnato da processi di degradazione delle pareti cellulari. l cambiamenti a livello biochimico e molecolare associati alla maturazione dei frutti sono stati studiati in modo approfondito nel pomodoro. La maturazione di molti frutti, come i pomodori, le mele e le banane, è controllata dall' etilene, la cui sintesi aumenta notevolmente durante questo processo. l geni codificanti gli enzimi coinvolti nella degradazione della parete cellulare, nel cambiamento di colore e nella sintesi di etilene sono stati clonati e sono state create piante transgeniche difettose in aspetti diversi della maturazione dei frutti. Le strategie si sono concentrate sull'inibizione della biosintesi di etilene e della degradazione della parete cellulare. A partire da Sadenosilmetionina (SAM) la biosintesi di etilene richiede due attività enzimatiche: ACC-sintasi, che catalizza la formazione di acido l-amminociclopropano-l-carbossilico (ACC), e ACC-ossidasi, che converte l'ACC in etilene (fig. 7).
cDNA codificanti questi enzimi sono stati isolati da diverse specie vegetali, tra cui il pomodoro, e le attività enzimatiche sono state inibite in pomodori transgenici da due diversi gruppi di ricerca, in Gran Bretagna e negli Stati Uniti. Per ambedue gli enzimi, la repressione antisenso causa una notevole riduzione nel contenuto di etilene e un ritardo nella maturazione dei frutti staccati dalla pianta. Un diverso approccio per sopprimere la sintesi di etilene è stato seguito dai ricercatori della società Monsanto. Operando una selezione su 600 specie di batteri del suolo, basata sulla capacità di crescere in presenza di ACC come unica fonte di azoto, si è identificato in due specie di Pseudomonas l'enzima ACCdeamminasi, capace di metabolizzare ACC in un prodotto inattivo. La trasformazione delle piante di pomodoro con il gene codificante questo enzima causa una riduzione dei livelli di etilene e un ritardo significativo nella maturazione. La riduzione della sintesi di etilene aumenta marcatamente la durata dei pomodori sugli scaffali dei negozi. Si spera di poter metter in vendita questi frutti dopo averli raccolti nel momento in cui cominciano ad avere un colore rosato, diversamente dalla normale pratica di raccolta di frutti di colore verde, che sono poi esposti all' etilene per promuovere il processo di maturazione.
Il processo di intenerimento del frutto di pomodoro è accompagnato dalla demolizione della lamella mediana della parete cellulare, che è costituita principalmente da pectine (un gruppo di polisaccaridi, composti soprattutto da acido poligalatturonico parzialmente esterificato con gruppi metilici). Dati biochimici e genetici suggeriscono che l'enzima poligalatturonasi (PG) svolga un ruolo importante in questo processo. Per saggiare questa ipotesi sono state create piante con soppressione antisenso, aventi meno dell' 1% della normale attività della PG. Un'analisi iniziale dei frutti non ha messo in luce alcuna differenza nel loro intenerimento, tuttavia un'ispezione più accurata della loro solidità durante l'immagazzinamento e un'analisi dei frutti completamente maturi hanno rivelato lievi differenze (Grierson e Schuch, 1993). Sulla base di questi risultati si è concluso che la PG non è un determinante principale dell'intenerimento dei frutti di pomodoro. Effetti opposti sulla solidità, durante l'invecchiamento dei pomodori, sono stati osservati in piante transgeni che con soppressione antisenso aventi ridotta attività dell'enzima pectinametilesterasi (PME). Durante la conservazione i frutti transgenici perdono quasi completamente la loro integrità (Tieman e Handa, 1994). Il ruolo della PME nel metabolismo della parete cellulare non è ben chiaro: si è ipotizzato che l'enzima renda le pectine più suscettibili all'azione della PG durante la maturazione dei frutti sulla pianta e, in effetti, nei frutti transgenici con soppressione antisenso il peso molecolare delle pectine è più elevato del normale. La disintegrazione dei frutti transgenici isolati dalla pianta potrebbe essere dovuta a un effetto indiretto: è stato proposto che il più elevato livello di metilesterificazione della pectina, dovuto all'inibizione della PME, possa causare modificazioni nell' architettura della parete cellulare, fra cui una riduzione della formazione di legami crociati dovuta agli ioni calcio e che queste alterazioni, a loro volta, rendano la parete maggiormente suscettibile all'azione sinergica dei diversi enzimi idrolitici in essa presenti.
Oltre all'eccessivo intenerimento, il cambiamento di colore dei frutti e degli ortaggi verso il bruno riduce la loro appetibilità per i consumatori ed è, dunque, un problema prioritario per i produttori e l'industria alimentare. Nei tuberi di patata, i danni meccanici durante raccolta e immagazzinamento provocano la formazione di 'macchie nere' causate dall' accumulo di polifenoli. L'enzima polifenolossidasi è ritenuto responsabile delle reazioni iniziali che conducono all'accumulo di polifenoli nei tessuti danneggiati. In accordo con quest'ipotesi, l'inibizione dell'attività della polifenolossidasi, tramite strategia antisenso, provoca in piante transgeniche di patata un'inibizione dell'imbrunimento dei tuberi dopo la raccolta (Bachem et al., 1994).
Oltre a essere un'importante fonte alimentare di carboidrati, i tuberi di patata sono anche usati come materiale da cui ottenere svariati prodotti di trasformazione. L'applicazione industriale delle patate è, però, limitata poiché è necessario inibire la germogliazione dei tuberi durante il loro immagazzinamento mediante trattamento con agenti chimici inibitori o conservazione a bassa temperatura. La seconda soluzione causa l'accumulo di zuccheri riducenti (come i monosaccaridi glucosio e fruttosio), non compatibile con l'industria delle patate soffiate e delle patate fritte: la reazione fra zuccheri riducenti e amminoacidi durante la frittura dà un indesiderabile colore marrone alle patate fritte. Si pensa che l'accumulo di zuccheri riducenti indotto dalle basse temperature sia causato da uno squilibrio fra idrolisi dell'amido e attività glicolitica. Come risultato di questo squilibrio, l'enzima saccarosiofosfatosintasi produce saccarosio, che è successivamente idrolizzato dall'enzima invertasi dando origine agli zuccheri riducenti glucosio e fruttosio. Per verificare se l'accumulo di zuccheri riducenti sia realmente correlato all'attività dell'invertasi, sono state create piante di patata transgeniche con soppressione antisenso, aventi una ridotta attività di invertasi acida solubile ed è stato studiato l'accumulo di zuccheri solubili (zuccheri riducenti e saccarosio) nei tuberi conservati al freddo. Sebbene non sia stata trovata correlazione fra il livello totale di attività invertasica e l'accumulo di zuccheri riducenti al momento della raccolta dei tuberi dalla pianta, si è osservata una forte correlazione tra il rapporto zuccheri riducenti-saccarosio e l'attività invertasica solubile dopo la conservazione al freddo. Sulla base di questi dati si è concluso che le invertasi non controllano la quantità totale di zuccheri solubili in tuberi conservati al freddo ma sono coinvolte nella regolazione del rapporto fra zuccheri riducenti e saccarosio.
Piante transgeniche con migliore resistenza agli stress abiotici
Stress da sali
Le piante sono immobili, per cui non possono sfuggire a condizioni ambientali sfavorevoli come la siccità, il freddo o la presenza di un eccesso di sali nel suolo. Perciò esse hanno sviluppato dei meccanismi per proteggersi dagli stress abiotici che causano disidratazione delle cellule (Bartels e Nelson, 1994). Un meccanismo comune è l'accumulo di composti a basso peso molecolare, osmoticamente attivi e non tossici (osmoprotettori). l più noti sono zuccheri, alcoli, l'amminoacido prolina e composti ammonici quatemari (per esempio, glicinbetaina). Per aumentare la concentrazione intracellulare di glicinbetaina, nel tentativo di migliorare la tolleranza agli stress, G. Lilius e collaboratori (1996) hanno introdotto in piante di tabacco un gene di E. coli codificante l'enzima colinadeidrogenasi. La crescita delle piante transgeniche in condizioni non stressanti è risultata normale. Tuttavia, aumentando la concentrazione di NaCI nel mezzo di coltura fino a 300 mM, il peso secco delle piante transgeniche è risultato incrementato dell'80% rispetto alle piante non trasformate, indicando un aumento della tolleranza allo stress salino. M.C. Tarczynski e collaboratori (1993) hanno studiato l'effetto dell'alcole mannitolo introducendo nel tabacco il gene di E. coli codificante l'enzima mannitolo-l-fosfatodeidrogenasi. Le piante transgeniche risultanti accumulano mannitolo, normalmente non presente nel tabacco, e tollerano un'elevata concentrazione di sale nel terreno. L'espressione dello stesso gene in Arabidopsis causa un aumento nella germinabilità dei semi in condizioni di alta salinità. Poiché, in questo caso, l'accumulo di mannitolo non è elevato, è improbabile che un effetto osmotico diretto da parte dell'alcole sia la causa della tolleranza agli stress. Non possono, tuttavia, essere esclusi effetti di destabilizzazione sulle membrane cellulari o su enzimi specifici: è nota da tempo l'importanza dell'integrità delle membrane e della loro composizione lipidica in presenza di stress.
La risposta delle piante agli stress, oltre ai cambiamenti metabolici e all'accumulo di composti protettivi a basso peso molecolare, comporta l'attivazione della trascrizione di un vasto gruppo di geni e, quindi, la sintesi di nuove proteine. Le proteine che si accumulano negli embrioni durante gli stadi terminali dello sviluppo del seme (LEA, Late Embryogenesis Abundant) sono state inizialmente identificate nel cotone. In seguito, queste proteine sono state trovate in molte altre specie, non solo nei semi ma anche in altre parti della pianta. Poiché vi è correlazione fra l' espressione di proteine LEA e varie condizioni di stress, si è ipotizzato che questo gruppo di proteine possa essere essenziale per la sopravvivenza delle piante in condizioni stressanti. La proteina LEA HV Al di orzo è stata espressa in riso transgenico. Il risultato è stato un aumento della tolleranza alla siccità e alla salinità. Per questo motivo, le proteine LEA sono utilizzabili per il miglioramento genetico delle piante coltivate, al fine di ottenere una maggiore tolleranza agli stress.
Stress da freddo
Studi genetici e biochimici hanno rivelato una correlazione positiva fra tolleranza al freddo e presenza di un'alta percentuale di acidi grassi insaturi nella membrana dei cloroplasti. È stato proposto che il grado di insaturazione sia determinato dall' enzima presente nei plastidi glicerolo- 3fosfatoaciltransferasi (GPAT). Per verificare questa ipotesi, N. Murata e collaboratori (1992) hanno prodotto piante transgeniche di tabacco esprimenti GPAT di zucca o di Arabidopsis. Le piante esprimenti l' enzima di Arabidopsis hanno mostrato un aumento della tolleranza al freddo, mentre quelle trasformate con l'enzima di zucca sono risultate meno resistenti dei controlli non trasformati. Le differenze nella tolleranza sono correlate con il grado di insaturazione degli acidi grassi misurato nelle piante transgeniche, fornendo una prova diretta dell'importanza della composizione lipidica nel determinare il grado di resistenza allo stress da freddo.
Stress da siccità
Il trealosio, un disaccaride non riducente composto da due molecole di glucosio, è presente in molti animali e microrganismi. Fra i vegetali, solo un limitato numero di piante tolleranti l'essiccazione contiene questa sostanza. Il trealosio è stato descritto come un agente stabilizzante delle proteine e delle membrane durante l' essiccazione e lo stress da alta temperatura, svolgendo così un ruolo protettivo. Commercialmente, il trealosio è usato come additivo per migliorare la qualità e il sapore di cibi essiccati e trattati industrialmente e come stabilizzante di vaccini, ormoni e componenti del sangue ma, a causa degli alti costi di produzione, il suo impiego come additivo per cibi non è comune.
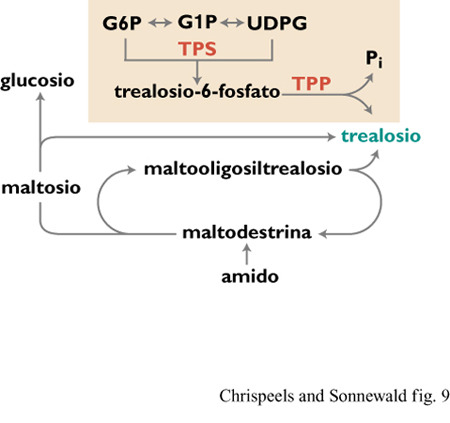
Le possibili vie bio sintetiche del trealosio sono varie (fig. 9). Nel lievito e in E. coli la biosintesi di questo composto richiede due enzimi: trealosio-6-fosfatosintetasi (TPS), che converte glucosio-6-fosfato e UDP-glucosio a trealosio-6- fosfato, e trealosio-6- fosfato fo sfatasi (TPP), che converte trealosio-6-fosfato a trealosio e fosfato inorganico. Durante un congresso internazionale ad Amsterdam, i ricercatori della società Calgene hanno comunicato di avere introdotto in piante transgeniche i geni di E. coli codificanti questi enzimi. Le piante risultanti accumulano quantità molto limitate di trealosio. Una collaborazione fra due laboratori scandinavi ha avuto, recentemente, come risultato la creazione di piante transgeniche di tabacco esprimenti trea10sio-6-fosfatosintetasi di lievito. Queste piante accumulano quantità di trealosio non molto elevate ma comunque significative (circa l ÷ 3 mg per grammo di campione secco), e mostrano un aumento marcato e geneticamente ereditabile nella tolleranza alla siccità.
Piante transgeniche resistenti a insetti infestanti e microrganismi patogeni
Le malattie provocate da funghi, batteri e virus e i danni causati da insetti e nematodi sono responsabili di sostanziali perdite nei raccolti, malgrado gli sforzi considerevoli compiuti per generare varietà resistenti e nonostante gli investimenti economici in pesticidi e altri programmi di controllo degli infestanti. Attualmente, gli insetticidi chimici applicati tramite irrorazione sono il sistema principale di lotta agli insetti infestanti e, a livello mondiale, gli agricoltori spendono 3÷5 miliardi di dollari all'anno per questo scopo. Si stanno praticando sforzi per diminuire la dipendenza dagli insetticidi usando sistemi di lotta integrati e utilizzando varietà resistenti. Una delle cause principali delle notevoli perdite nei raccolti, malgrado i costosi sforzi compiuti, risiede nella diffusa pratica della monocoltura ripetuta, cioè la coltivazione di una singola specie, con un patrimonio genetico uniforme, ripetuta anno dopo anno sullo stesso terreno. Questa pratica favorisce la diffusione di microrganismi e insetti nocivi. Sfortunatamente, il trattamento estensivo con pesticidi chimici aumenta la pressione selettiva e, come risultato, insetti e ceppi patogeni resistenti evolvono con maggiore rapidità. Il problema è ancor più complicato dal fatto che i pesticidi eliminano anche gli organismi che normalmente limitano la diffusione degli insetti infestanti e dei patogeni. Vi sono numerosi esempi di specie di insetti che non costituivano un problema 30 o 40 anni fa ma che ora sono diventati dei seri infestanti, proprio a causa dell'uso dei pesticidi. Tuttavia, sembra improbabile che si passerà all'adozione di pratiche agricole che si basino maggiormente sulla rotazione dei raccolti o sull 'uso di varietà aventi patrimonio genetico misto e, dunque, è probabile che si intensifichino sempre più gli sforzi per utilizzare l'ingegneria genetica al fine di creare varietà di piante coltivate resistenti a patogeni e infestanti.
Semi resistenti agli insetti
Tutte le piante contengono sostanze chimiche naturali di difesa che le proteggono da animali nocivi e da microrganismi patogeni; questa è la base di tutti gli sforzi di miglioramento genetico per creare piante resistenti. l geni che rendono resistente una varietà selvatica possono essere trasferiti a una coltivata mediante incrocio, se ambedue le varietà appartengono alla stessa specie, come, per esempio, il frumento selvatico e quello coltivato. Il trasferimento di geni tra specie diverse richiede, invece, le tecniche del DNA ricombinante e di trasformazione.
l semi sono la risorsa più importante di una pianta poiché rappresentano la generazione successiva e sono, perciò, naturalmente ben difesi contro la possibilità di essere aggrediti da insetti e mammiferi. Un meccanismo fondamentale di difesa naturale dei semi consiste nell'accumulo, alloro interno, di alte quantità di lectine e inibitori di enzimi digestivi. Le lectine sono proteine che riconoscono e legano carboidrati. Le lectine vegetali resistono alla digestione nel tubo digerente degli animali e, grazie alle loro proprietà di riconoscimento, si legano al rivestimento dell'intestino. Entrano, poi, rapidamente nelle cellule mediante endocitosi e agiscono come molecole-segnale che alterano la normale fisiologia dell'animale. Gli inibitori degli enzimi digestivi formano semplicemente un complesso 1:1 con l'enzima bersaglio, inattivandolo. Inibitori della α-amilasi e di diverse proteasi sono stati isolati da semi e caratterizzati. Questi due tipi di inibitori ostacolano la digestione del cibo ingerito dagli insetti e dall 'uomo e sono responsabili della necessità di cuocere a lungo certi semi, come per esempio i fagioli, che ne contengono elevate quantità.

Il gene codificante l'inibitore della α-amilasi di fagiolo è stato usato per produrre i primi semi transgenici aventi nuovi tipi di resistenza all'attacco di insetti. Sebbene i semi delle leguminose contengano elevate quantità di lectine e di inibitori enzimatici, essi sono danneggiati da alcuni bruchidi (una famiglia di coleotteri) specialmente durante la conservazione successiva al raccolto. Questi insetti depositano le uova sui semi; le larve che ne nascono si rintanano nel seme, nutrendosi di esso. Evidentemente, alcune specie di bruchidi hanno sviluppato evolutivamente dei meccanismi per superare le difese del seme che sono, invece, adeguate per resistere all'attacco di altre specie. Il fagiolo, Phaseolus vulgaris, è suscettibile all'attacco di due bruchidi presenti in America Latina, Acanthoscelides obtectus e Zabrotes subfasciatus, ma è resistente a bruchidi asiatici come Callosobruchus maculatus e Callosobruchus chinensis. La resistenza è causata dalla presenza nel fagiolo di una proteina, chiamata inibitore della α-amilasi, che inibisce l'enzima α-amilasi presente nel tubo digerente dei bruchidi asiatici, ma non l'enzima dei bruchidi latinoamericani. Quando il gene che codifica questo inibitore è stato isolato ed espresso in quantità elevate nei semi di due altri legumi, il pisello e l'azuki, i semi transgenici sono diventati resistenti a C. maculatus e C. chinensis (fig. 10). I piselli hanno acquisito, inoltre, resistenza al bruchide del pisello, che attacca i semi in via di sviluppo sulla pianta (Shade et al., 1994).
L'endotossina di Bocillus thuringiensis
Il batterio Bacillus thuringiensis (Bt) produce grandi quantità di proteine tossiche, chiamate endotossine. Queste proteine avvelenano le larve di insetti che si nutrono delle foglie di piante su cui cresce il batterio. Non è ancora chiaro l'esatto meccanismo d'azione delle endotossine, ma per diventare attive esse devono essere parzialmente digerite dalle proteasi dell'apparato digerente della larva. Esistono centinaia di ceppi di Bt e la maggior parte di essi produce tossine che sono specifiche solo per un numero limitato di ordini di insetti, come i lepidotteri (per es. farfalle e tignole), i coleotteri (per es. scarabei e maggiolini) o i ditteri (per es. zanzare). I primi esperimenti in cui un gene codificante l'endotossina è stato introdotto in piante di tabacco o pomodoro hanno mostrato una riduzione significativa nei danni causati alle foglie dalle larve di lepidotteri. Successivamente, il gene è stato modificato mediante tecniche di ingegneria genetica e la nuova versione è in grado di produrre in piante transgeniche una quantità di endotossina 500 volte più elevata rispetto ai primi esperimenti. Questo importante miglioramento ha aperto la via all'impiego di svariati geni codificanti endotossine da diversi ceppi di B. thuringiensis per creare patate resistenti alla dorifora, cotone resistente a Heliotis zea, un lepidottero nottuide, mais e riso resistenti alla piralide del mais, un lepidottero che scava la sua tana nel fusto e nelle foglie. Le tossine di Bt agiscono solo sugli insetti e non sono nocive per l'uomo; inoltre, usando promotori specifici è possibile far esprimere le tossine esclusivamente nell'organo, come il fusto o la radice, soggetto all'attacco degli insetti, evitando così l'espressione negli organi utilizzati dall'uomo, come i semi.
Il problema dell'evoluzione della tolleranza alle tossine
Una notevole preoccupazione che riguarda l'uso dei geni codificanti tossine, o di ogni altro gene, per controllare gli insetti infestanti, è la probabile comparsa di insetti che possono neutralizzare o tollerare questi veleni. L'esperienza acquisita con le varietà di piante coltivate, ottenute con le tecniche classiche di miglioramento genetico tramite incrocio, ha mostrato che la resistenza basata su di un singolo gene non è di lunga durata e viene meno dopo 4 ÷ 8 anni, per la comparsa di insetti tolleranti. La resistenza crea, infatti, una fortissima pressione selettiva a cui gli insetti rispondono evolvendosi. Per esempio, è stato scoperto che la resistenza alla tossina di Bt può insorgere da modificazioni nelle molecole che legano la tossina alla superficie delle cellule epiteliali del tratto digerente della larva. Esperimenti di selezione con una sola tossina mostrano che gli insetti possono evolvere resistenza ad ampio spettro contro più tossine di Bt. In uno studio recente, è stato dimostrato che in Heliothis virescens la normale frequenza di alleli conferenti resistenza alle tossine di Bt è approssimativamente 10-³: si pensa, quindi, che sotto forte pressione selettiva tale resistenza evolva rapidamente.
Sono state proposte diverse strategie per ovviare al problema della comparsa di insetti resistenti in seguito al trasferimento di geni nelle piante transgeniche. Una è la creazione di 'rifugi', cioè piccole parti del campo, coltivate con piante non transgeniche, in cui gli insetti possano trovare rifugio, in modo da ridurre la pressione selettiva dovuta alle piante transgeniche resistenti. Un'altra strategia consiste nella miscelazione dei semi transgenici con una piccola percentuale di semi non trasformati, in modo da avere in ogni coltivazione un certo numero di piante suscettibili all'attacco degli insetti. Tuttavia, non esiste nessuna prova che queste strategie funzionino e, in effetti, è stata già ottenuta qualche prova della loro scarsa efficacia, specialmente con gli insetti che si muovono da una pianta all'altra. Una strategia per ottenere resistenza duratura agli insetti infestanti consiste nel trasferire alla stessa pianta diversi geni, codificanti diverse proteine, ciascuna delle quali conferisce resistenza grazie a un meccanismo differente da quello delle altre. Una strategia alternativa è quella di adottare i principi della lotta integrata, tentando di ridurre la popolazione di insetti nocivi al di sotto del livello minimo di danno economico, senza, tuttavia, eliminarla del tutto; a tal fine si possono mescolare semi transgenici e non, con una percentuale di non transgenici piuttosto elevata, fino al 50% a seconda del particolare insetto. È poco probabile, però, che gli agricoltori adottino una simile strategia se non vi sono obbligati da appositi atti legislativi.
Piante transgeniche resistenti a virus
l virus sono organismi estremamente piccoli in grado di causare malattie in qualunque altro organismo: mammiferi, pesci, insetti, piante, alghe e perfrno batteri. Ogni virus ha solitamente un numero limitato di ospiti e un virus che causa una malattia in una specie vegetale può infettare qualche specie simile, ma di solito non infetta piante geneticamente distanti. Sicuramente i virus delle piante non infettano la specie umana. l virus, che sono molto più piccoli di una singola cellula, sono formati da una o poche molecole di acido nucleico racchiuse da un involucro proteico (rivestimento o capside). L'acido nucleico può essere RNA o DNA e costituisce il genoma del virus. Il genoma virale codifica pochissime proteine (solitamente da 4 a 8) e dunque i virus non si possono riprodurre al di fuori dell'ospite e ne utilizzano i meccanismi bio sintetici per la propria riproduzione. l virus passano da una pianta all'altra soprattutto grazie agli insetti, come, per esempio, le mosche bianche che si nutrono della linfa elaborata. Gli insetticidi sono stati, finora, l'unico strumento per combattere i virus delle piante, che causano enormi perdite nei raccolti di molte regioni tropicali e subtropicali, mentre gli sforzi per creare piante resistenti ai virus tramite incroci e selezione sono stati di scarso successo. Le piante infettate da virus sono spesso rachitiche e hanno foglie giallognole: i nomi coloriti di molti virus, come' striatura gialla del mais', 'mosaico del tabacco' o 'accartocciamento fogliare della patata', derivano dall'aspetto della pianta infettata.

Le più importanti proteine codificate dal genoma virale sono la proteina di rivestimento o del capside, la DNA o RNA-replicasi e la proteina di movimento. Quest'ultima modifica le connessioni fra le cellule vegetali per permettere al virus di trasferirsi da una cellula all'altra. Per creare piante transgeniche resistenti a virus sono stati utilizzati i geni virali che codificano queste proteine. La strategia è stata denominata resistenza derivata da patogeni (pathogenderived resistance), in quanto i geni che conferiscono resistenza provengono dal virus stesso. È possibile rendere le piante resistenti alla maggior parte delle classi di virus trasformandole con costrutti portanti il gene virale che codifica la proteina del capside, dopo averlo posto sotto il controllo di un forte promotore costitutivo. I primi esperimenti condotti nel 1987 mostrarono che piante di pomodoro, trasformate con la proteina del capside del virus del mosaico del tabacco, erano resistenti all'infezione da parte di questo virus. Il meccanismo attraverso cui agisce la resistenza non è ancora conosciuto. Nel caso di alcuni virus, l'eccesso di proteina di rivestimento nel citoplasma della cellula ospite può prevenire, indirettamente, la moltiplicazione virale; per altri virus, invece, ciò che è cruciale per la resistenza è un livello elevato di mRNA codificante la proteina di rivestimento, mentre la proteina stessa non è necessaria. Nel 1995 l'industria sementiera Asgrow ha introdotto in commercio una varietà di zucca resistente al virus del mosaico giallo della zucchina e al virus II del mosaico del cocomero (fig. 12). Questi virus possono ridurre del 20 ÷ 80% la resa dei raccolti di zucche e di altre cucurbitacee.
In Cina è in commercio da diversi anni tabacco proveniente da piante transgeniche resistenti al virus del mosaico del tabacco, grazie alla trasformazione con la proteina virale di rivestimento. Varietà di patate, cetrioli e pomodori resistenti a vari virus entreranno in commercio, nei prossimi anni, nei paesi occidentali e forse anche altrove.
La pathogen-derived resistance può essere ottenuta anche introducendo in piante transgeniche il gene della replicasi virale. Non sempre è necessario introdurre l'intera sequenza di questo gene: possono essere resistenti anche piante trasformate con geni troncati o modificati in modo che la proteina codificata non sia una replicasi attiva. Ancora una volta il meccanismo di resistenza non è chiaro, ma può essere connesso a possibili fenomeni di silenziamento genico dovuto a soppressione senso ed essere causato dagli elevati livelli di mRNA, più che da quelli proteici. In una particolare forma di soppressione senso detta cosoppressione, elevati livelli di mRNA trascritto da un determinato gene scatenano la degradazione totale degli mRNA della stessa specie e, dunque, la soppressione totale dell' espressione del gene. Questa strategia non è ancora arrivata alla fase di commercializzazione, ma sono state eseguite prove in campo con patate che esprimono il gene della replicasi del virus dell'accartocciamento fogliare della patata. I risultati hanno mostrato che queste piante sono immuni all'attacco del virus, che causa perdite nei raccolti in tutto il mondo.
Creazione di piante resistenti a funghi e batteri
Gli sforzi per produrre piante transgeniche resistenti a malattie dovute a funghi e batteri non hanno raggiunto gli stessi successi ottenuti per la resistenza a insetti e virus. Tuttavia, le conoscenze riguardo alla resistenza alle malattie si sono, recentemente, molto ampliate e ciò porterà allo sviluppo di nuove strategie per la produzione di piante transgeniche. L'infezione da parte di funghi o batteri induce nelle piante la sintesi di un certo numero di proteine; fra queste vi sono enzimi delle classi delle chitinasi e delle β-glucanasi, che sono in grado di degradare la parete cellulare dei funghi o dei batteri e, dunque, costituiscono parte della risposta di difesa della pianta. I primi esperimenti di trasformazione di piante con singoli geni, che codificano chitinasi o glucanasi, non hanno prodotto risultati incoraggianti ma, più recentemente, coesprimendo nel pomodoro transgenico due geni che codificano rispettivamente una chitinasi e una glucanasi, sono state prodotte piante resistenti a un fungo che causa la fusariosi. In modo simile, piante di tabacco che esprimono simultaneamente geni di orzo codificanti le chitinasi e le glucanasi mostrano un aumento della resistenza a Rhizoctonia solani, un fungo che infetta diverse piante, tra cui il pomodoro (Jach et al., 1995). Il maggior problema è che questo tipo di approccio consiste praticamente in tentativi alla cieca, poiché non si sa quasi nulla riguardo alla struttura delle pareti cellulari di questi funghi o all'esatta specificità delle glucanasi e delle chitinasi. Per produrre piante resistenti bisogna utilizzare geni che codificano enzimi molto attivi e non specifici, oppure enzimi non molto attivi ma molto specifici per la parete cellulare dei funghi. Sono state anche prodotte piante transgeniche che esprimono diverse forme di lisozima, un enzima che degrada la parete dei batteri, costituita da peptidoglicani (polimeri contenenti carboidrati e corte catene peptidiche). Il lisozima del batteriofago T4, espresso nei tuberi di patata, conferisce resistenza a Erwinia carotovora, un fungo responsabile della marcescenza dei tuberi.
Un altro approccio è basato sulla scoperta di una numerosa famiglia di peptidi, chiamati difensine (fig. 14), che hanno una potente e ampia attività antifungina in vitro. Un gruppo di ricerca belga ha recentemente espresso nelle piante di tabacco un gene codificante una difensina del seme di ravanello (Raphanus sativus) e ha mostrato che le piante transgeniche sono resistenti ad Alternaria longipes, un fungo che attacca le foglie. Le difensine sono molto diffuse nel regno vegetale e si stanno compiendo ulteriori indagini sulla possibilità di utilizzare questi peptidi in piante transgeniche (Terras et al., 1995).
Lo sviluppo delle conoscenze di base consentirà, senza dubbio, di tentare nuove strategie per conferire resistenza a funghi e batteri patogeni. Per esempio, cominciano a essere chiarite le basi molecolari di un fenomeno chiamato resistenza sistemica acquisita. Quando parte di una pianta è infettata da un patogeno, e questo è tenuto sotto controllo dai sistemi di difesa endogeni, l'intera pianta diventa resistente a un successivo attacco da parte di patogeni, anche diversi dal primo infettante. Il nome di questo fenomeno deriva dal fatto che le foglie non infettate rispondono a un segnale sistemico. Intermedi re attivi dell'ossigeno (ROl, Reactive Oxygen Intermediates), quali l'acqua ossigenata, fanno parte del sistema di segnali di risposta sistemica che è attivato nella pianta. È possibile, ottenendo un aumento nel livello di ROI mediante ingegneria genetica, conferire maggiore resistenza ai patogeni. Si può elevare il livello del ROl H2O2 esprimendo un enzima che genera questo composto, come per esempio l'enzima glucosiossidasi. Piante transgeniche di patata esprimenti tale enzima, di origine fungina, hanno elevati livelli di H2O2 e sono maggiormente resistenti a batteri e funghi patogeni (Wu et al., 1995).
Piante resistenti ai nematodi
l nematodi sono minuscoli animali che vivono nel suolo (in un chilo di terreno ve ne possono essere circa 500). Molti nematodi si nutrono di microrganismi ma altri sono parassiti che causano alle piante gravi danni valutabili globalmente, in termini economici, intorno ai 100 miliardi di dollari l'anno. l nematodi possono invadere le radici e lì moltiplicarsi, o semplicemente attaccarsi a esse e succhiarne la linfa. In ogni caso, danneggiano il sistema radicale e causano perdite nei raccolti. Quando un nematode invade o si attacca a una radice ne modifica notevolmente le cellule e causa la formazione di cellule giganti da cui trae nutrimento, fornito indirettamente dalle foglie foto sintetizzanti della parte aerea. Gli scienziati stanno tentando due strategie per combattere i nematodi. La prima consiste nell'esprimere, nelle radici, proteine che uccidano il nematode quando comincia a nutrirsi o che impediscano a esso di assumere nutrienti dalla pianta. I nematodi parassiti posseggono, ovviamente, proteasi con cui digeriscono le proteine vegetali; piante transgeniche che esprimono un gene del fagiolo dall'occhio (Vigna unguiculata), codificante un inibitore della tripsina, o un gene del riso, codificante un inibitore delle tiolproteasi, limitano lo sviluppo di Globodera pallida, un nematode che costituisce un serio problema per la patata. La seconda strategia consiste nell'indurre ablazione (uccisione mirata di cellule specifiche) delle cellule invase dai nematodi, in modo da prevenire lo sviluppo di cellule giganti nelle radici. Si sta studiando la possibilità di indurre ablazione utilizzando i promotori dei geni indotti nelle cellule giganti da cui i nematodi traggono nutrimento. Quando queste cellule cominciano a svilupparsi, vengono attivati geni specifici controllati da promotori che rispondono alla presenza del nematode; tramite costruzione di chimere, nelle quali proteine in grado di uccidere la cellula (per esempio, potenti ribonucleasi) sono poste sotto il controllo di questi promotori, è possibile ottenere l'ablazione delle cellule giganti in via di formazione. Poiché un nematode non può spostarsi dal sito di attacco, con la tecnica sopra descritta si priverebbe l'animale del suo nutrimento.
Ingegneria genetica e sostanze chimiche nocive nell'ambiente
Creazione di piante resistenti agli erbicidi
Per evitare che le piante coltivate soffrano a causa della competizione da parte di erbe infestanti, gli agricoltori hanno a disposizione diverse strategie, come la rimozione manuale o meccanica delle erbe infestanti, la rotazione dei raccolti, l'uso di coltivazioni ad alta densità e il controllo mediante sostanze chimiche. Quest'ultimo approccio, praticato mediante l'applicazione di erbicidi, è ora largamente diffuso in molti sistemi agricoli poiché è economicamente conveniente e richiede scarsa mano d'opera. Tutti gli erbicidi sono veleni che possono inquinare l'ambiente; quindi, gli erbicidi che sono rapidamente degradati e non sono tossici per gli animali o gli eco sistemi del suolo sono accettabili in agricoltura, mentre l'uso di erbicidi e altre sostanze chimiche che non soddisfino questi requisiti dovrebbe essere bandito.

{kind=link}
{kind=link}
Da lungo tempo i chimici organici cercano di identificare erbicidi specifici che abbiano un ampio spettro di tossicità verso molte piante ma che non siano nocivi nei confronti di una o più specie coltivate. La tolleranza è, solitamente, dovuta a una più rapida detossificazione dell' erbicida da parte della pianta tollerante rispetto a quella delle piante che vengono uccise. La detossificazione degli erbicidi e di altri xenobiotici (sostanze chimiche prodotte dall'uomo che entrano in contatto con le piante e altri organismi) comporta l'ossidazione da parte dell'enzima citocromo-P450-monoossigenasi, o la coniugazione da parte degli enzimi glutatione-Stransferasi o glicosiltransferasi e il successivo confmamento in un vacuolo. La pianta muore se le cellule non detossificano l'erbicida con velocità sufficiente, per cui esso inibisce un enzima chiave del metabolismo o blocca una via essenziale di trasduzione dei segnali. L'impiego diffuso di erbicidi sottopone le popolazioni di erbe infestanti a enormi pressioni selettive, con il conseguente emergere di varietà resistenti. La resistenza può essere il risultato di una mutazione nell' enzima bersaglio dell' erbicida o dello sviluppo di un più efficiente meccanismo di detossificazione. Una volta comprese le basi biochimiche della tolleranza agli erbicidi non è stato difficile creare piante transgeniche resistenti: tutto ciò che occorre sono i geni codificanti gli enzimi di detossificazione. Questi possono essere isolati dalle piante, specialmente dalle erbe infestanti resistenti, ma ancor più facilmente dai batteri, soprattutto quelli del suolo, che sono in grado di metabolizzare molti xenobiotici. Proprio dai batteri, i ricercatori della società Calgene (Stalker et al., 1996) hanno isolato un gene che hanno poi utilizzato per produrre cotone resistente al bromoxynil (BXN™cotton). Il bromoxynil (3,5-dibromo-4-idrossibenzonitrile) è un erbicida che uccide le latifoglie mediante inibizione della fotosintesi. Molte monocotiledoni, come frumento, orzo, avena e cipolle, hanno tolleranza al bromoxynil, poiché contengono enzimi che convertono il gruppo benzonitrile in acido benzoico, con un processo che coinvolge due reazioni enzimatiche. l batteri svolgono la stessa azione grazie a un singolo enzima chiamato nitrilasi. Il gene codificante questo enzima è stato isolato dal batterio Klebsiella ozaenae e introdotto in piante di tabacco, cotone e altri vegetali, sotto il controllo di un forte promotore costitutivo. Le piante di cotone transgenico risultanti sono resistenti al bromoxynil, anche quando l'erbicida è applicato in quantità 20 volte superiori a quelle solitamente utilizzate in campo (11,2 kg/ha invece di 0,56 kg/ha). Queste piante transgeniche tollerano applicazioni multiple dell'erbicida e possono essere trattate in ogni momento del loro ciclo vitale (fig. 15).
Altre società agrobiotecnologiche hanno introdotto geni per la tolleranza agli erbicidi in diverse e importanti piante coltivate. Questa strategia non è priva di detrattori e si è ipotizzato che il diffondersi dell'uso di piante transgeniche di questo tipo condurrà a un maggiore impiego di diserbanti chimici. Una tale conseguenza non è, però, inevitabile: è attualmente pratica comune compiere un trattamento con erbicidi prima che le nuove piantine coltivate emergano dal terreno, al fine di uccidere le erbe infestanti già presenti (trattamento pre-emergenza); utilizzando piante resistenti agli erbicidi tale trattamento può essere evitato. L'esperienza indica che, negli USA, un buon controllo delle erbe che infestano il cotone richiede, attualmente, da 3,2 a 3,4 kg/ha di erbicida, nella stagione di crescita. Usando cotone BXN™ e Buctril® (una marca di bromoxynil) è possibile ridurre la quantità a 1,7 kg/ha. Bisogna, però, tenere presente che l'area sottoposta a trattamento potrebbe diventare molto estesa nel caso che la tolleranza fosse introdotta nelle specie arboree. Attualmente, i diserbanti non sono usati nelle nuove piantagioni di alberi ma potrebbero esserlo se saranno prodotte specie transgeniche resistenti.
Piante transgeniche ed eliminazione dei rifiuti tossici dall'ambiente
Il suolo e le acque di scarico sono spesso contaminati da metalli pesanti tossici come cadmio, piombo e mercurio o da svariati composti organici dotati di elevata tossicità. La decontaminazione del suolo tramite tecniche convenzionali costa circa 500.000 dollari per ettaro e, a causa di questi costi elevati, non si sono fatti molti progressi a livello mondiale nella pulizia dei suoli contaminati. Un nuovo approccio, chiamato fitodecontaminazione (phytoremediation), è stato recentemente proposto per questo problema. La strategia prevede l'uso di piante coltivate per rimuovere dal suolo i metalli pesanti. Le piante possono, in seguito, essere raccolte e bruciate e potrebbe perfino essere possibile recuperare in questo modo metalli di valore come il nichel.
Da questo punto di vista, le piante possono essere considerate come pompe che usano l'energia solare per estrarre e concentrare minerali presenti nell'ambiente. Le piante assorbono dal terreno non solo i loro principali nutrienti minerali (azoto, fosforo, potassio, calcio, zolfo e magnesio), ma anche metalli pesanti essenziali per la crescita e lo sviluppo (ferro, manganese, zinco, rame, molibdeno e nichel). Molte piante, tuttavia, assorbono anche metalli pesanti di cui non hanno bisogno, se questi sono presenti nel suolo; fra questi sono compresi anche metalli tossici come cadmio, cromo, piombo, argento, selenio e mercurio. L'accumulo di questi elementi è nocivo per la maggior parte delle piante; esistono tuttavia gli 'iperaccumulatori', che tollerano elevate concentrazioni di metalli tossici e che costituiscono un'eccezione. Le cellule di queste piante hanno evoluto meccanismi per accumulare metalli pesanti nei vacuoli, all'interno dei quali essi non possono arrecare danni. Sebbene la ricerca di base su questi meccanismi di tolleranza sia solo agli inizi, le prime osservazioni indicano che i metalli pesanti inducono nelle piante la sintesi di piccoli peptidi ricchi dell'amminoacido cisteina, chiamati fitochelatine. l complessi fitochelatina-metallo pesante sono, poi, trasportati attraverso la membrana del vacuolo grazie a una speciale proteina trasportatrice. Il vacuolo è un esteso comparto subcellulare, usato soprattutto per l'accumulo di riserve. Le piante immagazzinano nei vacuoli metaboliti di riserva (per esempio, saccarosio), proteine di riserva, nutrienti minerali, ioni (nitrato, potassio, fosfato) e sostanze chimiche di difesa. Inoltre, i vacuoli possono funzionare da siti di deposito di molecole nocive, come i metalli pesanti o gli xenobiotici. I geni codificanti gli enzimi necessari alla sintesi di fitochelatine sono stati isolati e presto sarà possibile produrre piante transgeniche che sintetizzano grandi quantità di questi peptidi. Invece il gene codificante la proteina di trasporto vacuolare è stato, finora, identificato solo nel lievito. Elevando l'espressione di questo gene è stato possibile creare ceppi di lievito in grado di iperaccumulare cadmio. Un approccio di questo tipo dovrebbe funzionare anche nelle piante, ma si deve attendere l'isolamento del corrispondente gene vegetale.
Una strategia totalmente differente, per trattare il mercurio, si basa sulla volatilizzazione dei metalli pesanti. Il mercurio è tossico per le cellule perché si combina con i gruppi solfidrilici che spesso fanno parte dei siti attivi degli enzimi. Alcuni batteri sono in grado di tollerare elevate concentrazioni di mercurio poiché possiedono la reduttasi dello ione mercurico, un enzima che riduce i cationi mercurici (Hg²+) a mercurio metallico (Hgo) volatile. La trasformazione di piante di Arabidopsis con il gene codificante questo enzima ha portato a piante che crescono normalmente in mezzi di coltura contenenti cloruro di mercurio, nei quali la crescita di esemplari non trasformati è, invece, completamente arrestata (Rugh et al., 1996).
La volatilizzazione potrebbe essere utilizzata anche per il selenio. Le proprietà chimiche di questo metallo sono simili a quelle dello zolfo e le piante che accumulano zolfo, come la colza (Brassica), spesso accumulano anche selenio. Brassica juncea, una pianta indiana che è in grado di tollerare la siccità e lo stress salino, è capace di ridurre di circa il 50% la concentrazione di selenio nel suolo, fino alla profondità di 1 m, se coltivata e raccolta per diversi anni.
Per avere successo, la tecnica di fitodecontaminazione dovrà associare le strategie metaboliche delle specie iperaccumulanti con le qualità delle piante coltivate normalmente nell'area da trattare. Molte piante iperaccumulanti sono piccole e insolite e producono una biomassa limitata. Un'efficiente fitodecontaminazione richiede piante con una buona crescita vegetativa e in grado di essere facilmente raccolte e processate. Questa tecnologia dovrà, dunque, necessariamente fondarsi su piante transgeniche e i geni responsabili del successo degli iperaccumulatori dovranno essere trasferiti a piante più facilmente gestibili.
Protezione delle piante dai danni dell'ozono e degli altri stress ossidativi
L'industrializzazione e la diffusione dell 'uso dell' automobile sono le cause principali dell'inquinamento da ozono; le estese perdite di foreste e raccolti causate dagli elevati livelli di ozono sono un esempio tangibile degli effetti dello stress ossidativo sulle piante. Meno visibili, ma ugualmente gravi, sono le perdite causate dal danno ossidativo che costituisce un effetto secondario degli stress da freddo e da calore. In presenza di stress ambientali, le cellule accumulano ROI con conseguente danneggiamento ossidativo delle funzioni metaboliche. A causa della natura delle reazioni fotosintetiche, la cellula vegetale è permanentemente a rischio di danno da ROI e il cloroplasto, dove avviene la fotosintesi, è una delle principali fonti di questi composti. Le piante hanno sviluppato evolutivamente meccanismi per proteggersi dai ROl, ma a volte i sistemi di protezione risultano insufficienti, specialmente se lo stress avviene in presenza di un'alta intensità di luce.
In condizioni di alta intensità luminosa avviene un'eccessiva riduzione del foto sistema l, ma il processo di riduzione dell' anidride carbonica non è in grado di tenere lo stesso passo: l'eccesso di elettroni è deviato direttamente verso l'ossigeno, creando ROI. Questi possono danneggiare il cloroplasto, o passare nel citosol e provocare danni ad altri componenti della cellula. Quando le piante sono sottoposte all'azione dell'ozono, la maggior parte del danno causato dai ROI è invece extracellulare. Tuttavia, bisogna tener presente che parte dei composti diffonde anche nel citoplasma. Le piante si difendono dai ROI utilizzando due reazioni enzimatiche che convertono questi composti a prodotti innocui. La prima reazione è catalizzata dall' enzima superossidodismutasi e produce perossido di idrogeno (H2O2), che è esso stesso un ROI. La seconda reazione distrugge H2O2 attraverso l'azione dell'enzima ascorbatoperossidasi, che catalizza la conversione dell'acido ascorbico a radicali mono-deidroascorbato con la simultanea demolizione di H2O2. l radicali sono riconvertiti ad acido ascorbico dall'azione della ferredossina o di altre reduttasi solubili. Poiché i ROI possono essere presenti in ogni comparto cellulare, forme diverse di questi enzimi sono localizzate nei cloroplasti, nei mitocondri e nel citosol.
Numerose osservazioni suggeriscono che un aumento nei livelli di questi enzimi, ottenuto mediante ingegneria genetica, potrebbe aiutare le piante ad affrontare gli stress ossidativi. lnnanzitutto, in presenza di stress l'attività di questi enzimi aumenta e l'aumento è correlabile allo sviluppo di tolleranza. In secondo luogo, le piante esposte a un certo tipo di stress acquisiscono maggiore tolleranza anche verso altri tipi, un fenomeno noto come tolleranza crociata. Infine, alcuni erbicidi uccidono le piante elevando marcatamente i ROI quando queste sono esposte alla luce: mutanti che tollerano questi erbicidi hanno più alti livelli di enzimi che distruggono i ROI e, quindi, tollerano maggiormente lo stress. Queste osservazioni sono alla base di esperimenti di ingegneria genetica che potranno fornire una verifica diretta dell'ipotesi che gli enzimi descritti giochino un ruolo centrale nella protezione dagli stress. Il gene codificante la superossidodismutasi è stato modificato per permettere l' accumulo dell' enzima nei cloroplasti, i principali produttori di ROI. Quando questo gene è stato espresso in piante di tabacco e patata, il livello di superossidodismutasi nei cloroplasti è aumentato di 2 ÷ 4 volte, e questo aumento, seppur modesto, ha conferito una considerevole protezione dallo stress: piante di tabacco transgenico esposte per 7 giorni a livelli ambientali di ozono, simulanti le fluttuazioni naturali osservate in aree industriali, mostrano danni da ozono (lesioni necrotiche) sul 6 ÷ 7% della superficie fogliare, rispetto al 22% osservato nelle piante di controllo non trasformate (Van Camp et al., 1994). Queste piante transgeniche sono anche maggiormente resistenti all'effetto dell'associazione fra stress fotoossidativo e stress da freddo o gelo. Per esempio, le foglie di piante di controllo mantenute a 3 °C per 6 ore alla luce e poi riportate a temperatura normale sono risultate incapaci di compiere la foto sintesi, mentre l'apparato fotosintetico delle piante transgeniche esposte allo stesso trattamento è risultato essere totalmente protetto dai danni ossidativi e in grado di riprendere normalmente la propria attività.
Miglioramento genetico delle piante
Per migliaia di anni, gli agricoltori hanno modificato il patrimonio genetico delle piante attraverso una selezione deliberata o, inconsapevolmente, attraverso le pratiche di coltivazione da essi adottate. Durante il 20° secolo, le scienze della genetica e della statistica hanno posto basi più razionali per il miglioramento genetico delle piante.
Produzione di ibridi: geni per creare sterilità maschile e per ristabilire fertilità nella progenie
Nel 1908 lo statunitense G.H. Schull, lavorando al miglioramento genetico del mais, riferì di avere ottenuto risultati sbalorditivi incrociando piante di mais di due differenti linee geneticamente pure (omozigoti). Il raccolto ottenuto, a partire dai semi della linea ibrida (eterozigote) risultante dall'incrocio delle due linee pure, era di quattro volte superiore rispetto a quelli ottenibili dalle linee pure di partenza. Questo inatteso vigore delle piante eterozigoti è diventato famoso come lussureggiamento degli ibridi, o eterosi, cosicché gli ibridi di mais di questo tipo hanno giocato un ruolo importante nel costante aumento di rendimento di questa pianta in tutto il mondo. Il lussureggiamento degli ibridi è maggiore nelle piante a impollinazione incrociata (impollinazione fra due individui distinti), mentre non è altrettanto pronunciato nelle piante autoimpollinanti, come il frumento. Tuttavia, anche nel secondo caso la disponibilità di ibridi aumenterebbe la resa dei raccolti del 20 ÷ 30%. La produzione efficiente ed economicamente vantaggiosa di semi ibridi richiede linee di partenza maschio-sterili (che non producono polline funzionante per la riproduzione) e linee ripristinatrici della fecondità, cioè aventi geni che ripristinino la fertilità maschile nella progenie dell'incrocio. La maschio-sterilità permette di controllare la riproduzione sessuale: i semi ibridi sono prodotti da piante maschio-sterili, fecondate con polline delle linee ripristinatrici della fecondità. Le piante che cresceranno dai semi ibridi non saranno maschio-sterili e quindi produrranno semi poiché la fertilità è stata ripristinata. L'identificazione di linee mutanti maschio-sterili di mais e di linee con geni ripristinatori ha reso facile e poco costosa la produzione di ibridi. Prima dell'introduzione di questo sistema, i fiori maschili (il mais ha fiori maschili e femminili separati) erano rimossi a mano da metà delle piante, con costi molto elevati. Per molte specie è più difficile che per il mais rimuovere a mano i fiori maschili o, se i due sessi non sono separati, asportare la parte maschile del fiore. In molti casi non esiste un sistema genetico di linee maschio-sterili e ripristinatrici; ne risulta che la produzione di semi ibridi è spesso economicamente impossibile.
Una collaborazione fra ricercatori della Plant Genetic System di Gent, in Belgio, e la University of Califomia di Los Angeles, negli Stati Uniti, ha superato questi problemi inserendo, mediante ingegneria genetica, un sistema di maschio-sterilità/ripristino di fertilità in diverse piante coltivate. La chiave del successo è stata l'ablazione delle cellule dell' organo riproduttore maschile (stame) che produce il polline. L'ablazione è stata ottenuta mediante una potente ribonucleasi batterica, codificata dal gene barnase di Bacillus amyloliquefaciens. Quando questo gene è posto sotto il controllo di un promotore stame-specifico ed è espresso in piante transgeniche, le piante risultanti sono maschio-sterili, poiché la ribonucleasi uccide le cellule che dovrebbero produrre gli stami. Di conseguenza i fiori di queste piante non hanno stami.
Il ripristino della fertilità dopo incrocio è stato ottenuto usando un gene che codifica un inibitore della ribonucleasi batterica (barstar), posto sotto il controllo di un forte promotore costitutivo e introdotto in una pianta della stessa specie, ma di una diversa linea rispetto a quella trasformata con barnase (Mariani et al., 1992). Quando le piante maschio-sterili che esprimono barnase sono impollinate con polline proveniente dalle piante che esprimono barstar, i semi risultanti generano piante che producono polline (e dunque semi), poiché barstar annulla l'azione di barnase. Il gene codificante la ribonucleasi funziona da gene conferente maschio-sterilità e quello codificante barstar da gene ripristinatore. Il sistema è stato provato in piante di frumento e di colza e sembra molto promettente: le prove in campo con colza transgenica mostrano un aumento di resa del 20 ÷ 30% degli ibridi rispetto alle linee pure autofecondate.
Produzione di semi asessuati
Alcune piante di interesse agricolo, come la patata e la canna da zucchero, si propagano in modo vegetativo o asessuato, ma la maggior parte delle piante coltivate si propaga tramite semi che derivano dalla riproduzione sessuale. La caratteristica distintiva della riproduzione sessuale è il fatto che appaiono nuove combinazioni di geni nella generazione successiva e, quindi, i semi di piante ibride generano sempre popolazioni non uniformi di piante. Tutte le piante propagate a partire da tessuti vegetativi sono, invece, geneticamente identiche, indipendentemente dal fatto che la pianta di partenza sia una linea pura, un ibrido o una varietà selvatica. Questo è un grande vantaggio, poiché gli agricoltori desiderano una popolazione di piante che diano una resa uniformemente elevata. È possibile combinare la facilità con cui i semi sono raccolti, conservati e manipolati con i vantaggi di uniformità della propagazione vegetativa o clonale? Alcune specie vegetali sono caratterizzate dalla formazione di semi senza fecondazione (apomissi) in aggiunta ai semi normali. I semi che si formano per apomissi sono generati nell' ovario del fiore a causa di una stimolazione ormonale che provoca la divisione cellulare nel tessuto ovarico. Il patrimonio genetico di questi semi prodotti in assenza di fecondazione è, perciò, identico a quello della pianta madre e tutti i semi prodotti per apomissi sono geneticamente identici. I semi apomittici sono portatori di grandi speranze per l'agricoltura e sono particolarmente adatti per la produzione di piante 'd' élite', risultanti dalla combinazione fra modifiche introdotte tramite tecnologie di ingegneria genetica e una serie di susseguenti incroci. Il problema è che non si sa ancora come si possano costringere le piante di interesse agricolo a produrre semi apomissici. La ricerca è attualmente indirizzata verso l'identificazione dei geni responsabili della normale insorgenza dell'apomissi in alcune specie non coltivate. A partire da questi, si dovrebbe essere in grado di identificare geni equivalenti in piante coltivate e di alterarne l'espressione, in modo che anche le piante di interesse agrario producano semi apomittici. Sebbene questo obiettivo sembri ancora confinato in un lontano futuro, è ragionevole ritenere che esso sarà convenientemente raggiunto.
Geni che controllano la fioritura
Nella maggior parte delle piante la fioritura è controllata da fattori endogeni, come l'orologio biologico che regola lo sviluppo, e da fattori esogeni, come la lunghezza del periodo giornaliero di luce. Le piante devono raggiungere una certa dimensione o livello di maturazione prima di essere in grado di rispondere ai fattori estrinseci che inducono la fioritura. Il tempo necessario per raggiungere questa maturazione varia, a seconda delle specie, da alcune settimane a decine di anni. Durante la crescita vegetativa, i meristemi apicali producono in continuazione nuove foglie e nuovo tessuto che va ad aggiungersi al fusto sotto stante. La fioritura avviene quando il meristema del germoglio apicale è convertito a meristema fioraIe. In alternativa, il meristema apicale può prima originare meristemi laterali che formano germogli laterali vegetativi seguiti da meristemi fiorali. In ambedue i casi, il destino del meristema è modificato e il cambiamento è mediato dall'azione di geni che controllano l'identità dei meristemi, come Apetalai (APi) e Lea.fY (LFY). Quando questi geni sono inattivati a causa di una mutazione, i meristemi che dovrebbero produrre fiori continuano semplicemente a produrre nuovi tessuti vegetativi. Partendo da questa osservazione, si è ipotizzato che, ponendo questi geni sotto il controllo di un forte promotore costitutivo, sia possibile ottenere l'effetto opposto: la conversione prematura dei meristemi vegetativi a meristemi fiorali.
I ricercatori del Salk Institute for Biological Sciences a San Diego, California, hanno posto il gene LFY sotto il controllo del promotore costitutivo CaMV 35S e hanno introdotto il gene chimerico in un albero perenne, che ha un tempo di maturazione di anni. Le piantine transgeniche, rigenerate mediante coltura di tessuti dai germogli trasformati, hanno mostrato una fioritura estremamente precoce: in pochi mesi di coltura si sono sviluppati fiori dalle ascelle fogliari. Se risulterà che questi fiori precoci sono fertili e in grado di produrre semi, le piante trasformate con LFY saranno veramente molto utili per chi lavora al miglioramento genetico delle specie arboree: la lunghezza del periodo di crescita vegetativa di molte specie di alberi (5 ÷ 10 anni) costituisce, attualmente, l'ostacolo principale per il loro miglioramento genetico tramite incrocio.
Problemi di regolamentazione e problemi sociali connessi all'ingegneria genetica delle piante
I vantaggi e i pericoli dell'ingegneria genetica delle piante, specialmente per quanto riguarda i prodotti destinati all'alimentazione, sono stati oggetto di intenso dibattito dalla fine degli anni Ottanta. La discussione ha coinvolto associazioni per la difesa dei consumatori, rappresentanti dell'industria, scienziati, legislatori ed esperti di etica. Nessun'altra innovazione nella produzione di cibo è mai stata sottoposta a un'analisi critica ugualmente approfondita. Nel corso di questo dibattito, molte delle domande poste dagli oppositori hanno ricevuto risposte soddisfacenti. Sfortunatamente, attivisti dei gruppi contrari alla produzione di piante transgeniche, non soddisfatti di sollevare semplicemente obiezioni e porre domande, hanno occupato e danneggiato laboratori di ricerca e serre o distrutto piante transgeniche coltivate in campo. Questo è avvenuto sporadicamente in California e Olanda, più sistematicamente in Germania, dove per esempio dal gennaio all'agosto 1996 si sono contati tredici episodi di distruzione di piantagioni sperimentali, alcune delle quali avevano proprio lo scopo di valutare i possibili rischi per l'ambiente causati dalle nuove piante. Vi sono state anche proteste pacifiche come, per esempio, le opposizioni giudiziarie al conferimento di brevetti e pesanti boicottaggi dei primi alimenti prodotti da piante transgeniche apparsi sul mercato. Vista l'esperienza, è ragionevole pensare che la messa in pratica effettiva delle grandi potenzialità della trasformazione delle piante dipenderà soprattutto da due fattori: l) come i governi regolamenteranno la produzione di cibo e di altri prodotti ottenibili dalle piante transgeniche; 2) se l'opinione pubblica accetterà o no tali prodotti.
Regolamentazione da parte dei governi
Negli Stati Uniti, tre agenzie governative regolamentano la produzione di alimenti prodotti mediante ingegneria genetica: la Food and Drug Administration (FDA), lo United States Department of Agriculture (USDA) e l'Environmental Protection Agency (EPA). La FDA ha sempre sostenuto la posizione secondo cui i prodotti dell'ingegneria genetica non richiedono regolamentazione aggiuntiva rispetto a quella necessaria per ogni prodotto alimentare o medicinale. L'USDA e l'EPA hanno, invece, sostenuto sin dall'inizio che sarebbe stato necessario regolamentare la coltivazione delle nuove piante prodotte grazie alle tecniche di DNA ricombinante. Una posizione simile, in favore di nuove regole, è stata presa dall'Unione Europea. La posizione 'regolamentarista' si basa sul concetto secondo il quale le piante prodotte tramite ingegneria genetica sarebbero più pericolose, e avrebbero proprietà meno prevedibili, rispetto a quelle prodotte secondo le tecnologie tradizionali. Questa visione è in disaccordo con quella che riscuote il consenso degli scienziati, dell'industria, del mondo accademico e degli istituti di ricerca governativi. Il Field Trial Permits Service (APHIS) dell'USDA, addetto alla regolamentazione delle piante transgeniche, tratta queste come nuovi infestanti potenzialmente pericolosi, sebbene non vi sia un ragionevole schema logico che suggerisca come una pianta transgenica possa realmente diventare pericolosa, solo per il fatto di essere transgenica. I permessi dell'USDA per le prove in campo richiedono che queste includano una valutazione approfondita dell'impatto ambientale: per le centinaia di permessi accordati non è stato osservato nessun impatto significativo sull'ambiente. Agli scienziati dell'industria e delle istituzioni accademiche l'enorme quantità di lavoro d'ufficio richiesto per ottenere un permesso sembra tempo sprecato. Gli scienziati sostengono che il processo di regolamentazione dovrebbe focalizzarsi sui rischi tangibili associati alla comparsa di una nuova pianta e non sul metodo utilizzato per produrla. L'USDA attualmente utilizza un sistema efficiente di notificazione per sei importanti piante coltivate che sono state modificate per determinate caratteristiche e questa pratica ha reso meno oneroso il carico di lavoro burocratico per gli scienziati. In Europa il sistema di regolamentazione nei confronti delle biotecnologie applicate all'agricoltura è più macchinoso che negli Stati Uniti. I permessi per le prove in campo sono più difficili da ottenere e le procedure variano da paese a paese. Per ottenere un permesso di produzione (rilascio illimitato), è necessario fare domanda in uno dei paesi dell'Unione Europea (UE). La domanda è valutata da due commissioni, una nazionale e una dell'VE. Per quest'ultima, il permesso è accordato se la maggioranza dei paesi vota a favore. Una volta superati questi ostacoli, la pianta può essere coltivata in ogni paese dell'UE, compresi quelli i cui rappresentanti in commissione hanno espresso voto contrario.
Una fonte di notevole preoccupazione, sia in Europa sia in America Settentrionale, riguarda il problema delle etichettature speciali. È necessario, o perfino auspicabile, marcare con etichette speciali i prodotti alimentari derivanti dalle piante transgeniche esposti sugli scaffali di negozi, mercati e supermercati? Un'etichetta speciale fornisce delle informazioni utili per permettere al consumatore di prendere una decisione con cognizione di causa, o semplicemente aumenta la paura dell'ignoto? Avremmo dovuto in passato etichettare in modo simile le nuove varietà di piante alimentari che sono state prodotte tramite irradiazione con raggi X o coltura di tessuti? In sostanza, è importante il metodo utilizzato per creare una nuova varietà o il rischio noto costituito dal prodotto ottenuto? Gli scienziati sostengono che il metodo non sia importante e i governi stanno cominciando a condividere questa posizione. La proposta di etichette speciali è stata respinta negli Stati Uniti e in altri paesi, ma in Europa la questione non è ancora stata risolta. Le nuove varietà prodotte mediante ingegneria genetica contengono un certo numero di nuovi geni e proteine, ma lo stesso vale, e in misura maggiore, per le varietà prodotte con le tecnologie tradizionali di incrocio e selezione: in realtà, mediante le tecnologie tradizionali sono trasferite porzioni ben più grandi di DNA. L'unico problema che può sorgere riguarda le proteine allergeniche. Molti prodotti alimentari, anche di uso estremamente comune, contengono allergeni e gli individui allergici a determinati alimenti evitano di assumerli. Per esempio, alcune persone sono allergiche ai prodotti a base di farina di frumento. Se una proteina nota per essere un allergene fosse trasferita dal frumento al pomodoro sarebbe giusto etichettare quei particolari pomodori transgenici scrivendo chiaramente che essi contengono una proteina di frumento, che costituisce un potenziale agente scatenante allergie. L'enfasi, in questo caso, è posta sul pericolo potenziale e non sul metodo utilizzato per produrre la nuova varietà di pomodoro. Considerando la questione da un punto di vista genetico, non vi è alcuna valida ragione scientifica per raggruppare tutte le piante prodotte mediante ingegneria genetica sotto un'unica categoria etichettabile in modo particolare. Nuove varietà di piante già in commercio e piante completamente nuove compaiono in continuazione nei negozi e nei mercati e sono acquistate dai consumatori senza alcuna necessità di etichette particolari. Dal punto di vista dell'industria di lavorazione dei prodotti agricoli a fini alimentari non vi è dubbio che l'obbligo di etichette speciali sarebbe un serio disincentivo all'impiego di piante transgeniche: un simile obbligo richiederebbe la separazione delle fabbriche che trattano piante transgeniche da quelle che trattano piante ottenute con tecniche tradizionali, o la duplicazione delle linee di produzione all'interno di una stessa fabbrica.
Problemi ecologici
Un'altra serie di problemi riguarda gli aspetti ecologici della coltivazione di piante transgeniche. Possono queste piante diventare infestanti incontrollabili? Possono i nuovi geni trasferirsi in popolazioni di piante selvatiche a causa di incroci spontanei fra la varietà coltivata e le varietà selvatiche? Quale sarà l'effetto della diffusione dell'uso di erbicidi e pesticidi sull'evoluzione di erbe e insetti infestanti?
Non vi è ragione di pensare che le piante transgeniche possano facilmente diventare infestanti incontrollabili. Molte specie vegetali esotiche che in passato hanno invaso nuovi territori sono diventate dei seri infestanti, come per esempio il tumbleweed (Salsola kali), l'erba arrotolata e trasportata dal vento che è arrivata negli USA come contaminante di partite di semi provenienti dall'Europa e ha invaso le pianure del nord negli anni Settanta del 19° secolo; in questo senso, però gli organismi prodotti mediante ingegneria genetica non sono da considerare esotici. Essi piuttosto rappresentano specie già presenti in una determinata zona, alle quali sono stati aggiunti uno o due nuovi geni che non conferiscono alla specie in questione un vantaggio per sopravvivere in quel particolare ecosistema.
Tuttavia, nel caso in cui nella stessa area di coltivazione di una determinata pianta transgenica crescano anche piante selvatiche a essa imparentate, i geni introdotti mediante ingegneria genetica possono trasferirsi alle varietà selvatiche. Un simile evento è stato recentemente documentato in Danimarca per quanto riguarda la colza (Brassica napus) transgenica e l'infestante Brassica campestris (Mikkelsen et al., 1996). Lo stesso problema può presentarsi per il girasole negli USA, per il mais e i fagioli in Messico, per il pomodoro in America Meridionale e in tutti gli altri casi in cui sono presenti varietà selvatiche di specie coltivate. Non è scontato che un simile trasferimento di geni sarebbe dannoso per l'ecosistema naturale o per gli agricoltori. Questo tipo di trasferimento avviene già ora, solo che i donatori sono costituiti dalle varietà di élite prodotte mediante tecniche tradizionali. È molto improbabile che il trasferimento di geni crei dei superinfestanti, poiché i caratteri trasferiti non conferiscono alcun vantaggio selettivo in un ecosistema naturale.
Per quanto riguarda il problema della possibile evoluzione di organismi infestanti (erbe, insetti, batteri, ecc.), l'esperienza sufficientemente ampia sull'uso di pesticidi chimici permette di sostenere che la pressione selettiva conseguente all'uso di queste sostanze favorisce l'emergere di infestanti resistenti. Vi sono moltissimi esempi di erbe infestanti resistenti agli erbicidi e di insetti resistenti agli insetticidi. Questa tendenza può solo subire un'accelerazione. Dati i costi di produzione delle varietà transgeniche, sarà interesse delle ditte produttrici istruire gli agricoltori riguardo ai metodi di gestione dei loro campi, al fine di permettere che l'impiego di queste varietà rimanga conveniente il più a lungo possibile; infatti, una volta emersi infestanti resistenti, la coltivazione della varietà transgenica tollerante cessa di essere conveniente.
Problemi etici
Un terzo gruppo di questioni riguarda i problemi etici connessi alla decisione di permettere il brevetto di forme di vita e quelli del giusto compenso. Secondo le legislazioni dei paesi industrializzati, chi inventa qualcosa ha il diritto di essere protetto, in modo da poter trarre un profitto dalla propria invenzione. Il problema è se questo diritto si possa estendere alle forme viventi e, in caso affermativo, se gli agricoltori o altri debbano essere ricompensati per avere mantenuto comunità di piante in cui piante selvatiche contenenti geni utili hanno trovato rifugio. Le industrie biotecnologiche dei paesi sviluppati insistono che gli organismi viventi possono essere brevettati e che debba essere permesso trarre profitto da queste invenzioni. In genere, i governi degli stessi paesi hanno sostenuto questa posizione. Ma chi è il proprietario dei geni o delle piante da cui essi provengono? Sono questi patrimonio dell'umanità, a disposizione gratuita di chi desideri fame uso, o le specie e le varietà indigene sono, invece, risorse che appartengono a chi le mantiene? Il fatto è che la maggior parte delle piante che forniscono cibo all'umanità proviene da paesi poveri e in via di sviluppo dell'Africa, America Latina e Asia, mentre la maggior parte dell'impiego commerciale di queste risorse genetiche ha luogo nei paesi sviluppati. Le grandi industrie dei paesi ricchi sarebbero felicissime di vendere a quelli poveri le versioni migliorate degli stessi semi che hanno avuto origine proprio in questi ultimi. Trovare una soluzione adeguata ed equa non sarà facile.
Bibliografia citata
BACHEM, C.W.B., SPECKMANN, G.-J., VANDERLINDE, P.C.G., VERHEGGEN, F.T.M., HUNT, M.D., STEFFENS, J.C., ZABEAU, M. (1994) Antisense expression of polyphenol oxidase genes inhibits enzymatic browning in potato. Bio/TechnoIogy, 12, 1101-1105.
BARTELS, D., NELSON, D. (1994) Approaches to improve stress tolerance using molecular genetics. Plant CelI Environ., 17, 659-667.
CHILTON, M.- D., DRUMMOND, M.H., MERLO, D.J., SCIAKY, D., MONTOYA, A.L., GORDON, M.P., NESTER, E.W. (1977) Stable incorporation of plasmid DNA into higher plant cells: the molecular basis of crown gall tumorigenesis. CelI, 11, 263-271.
CHRISPEELS, M.J. (1991) Sorting of proteins in the secretory system. Annu. Rev. Plant Physiol. PIant Mol. Biol., 42, 1-24.
FLIPSE, E., SUURS, L., KEETELS, C.lA.M., KOSSMANN, J., JACOBSEN, E., VISSER, R.G.F. (1996) Introduction of sense and antisense cDNA for branching enzyme in the amylose-free potato mutant leads to physico-chemical changes in starch. PIanta, 198, 340-347.
GALTIER, N., FOYER, C.H., MURCHIE, E., ALRED, R., QUlCK, P., VOELKER, T.A., THÉPPENIER, C., LASCÈVE, G., BETSCHE, T. (1995) Effects of light and atmosphere carbon dioxide enrichment on photosynthesis and carbon partitioning in leaves of tomato (Lycopersicum esculentum L.) plants over-expressing sucrose phosphate synthase. J Exp. Bot., 46, 1335-1344.
GODDIJN, O.J.M., PEN, J. (1995) Plants as bioreactors. Trends Biotechnol., 13, 379-387.
GRIERSON, D., SCHUCH, W. (1993) Contro I ofripening. Phil. Trans. Roy.Soc.London,342,241-250.
HAQ, T.A., MASON, H.S., CLEMENTS, J.D., ARNTZEN, C.J. (1995) Oral immunization with a recombinant bacterial antigen produced in transgenic plants. Science, 268, 714-716.
HERRERA-ESTRELLA, L., DE BLOCK, M., MESSENS, E., HERNALSTEENS, J.-P., V AN MONTAGU, M., SCHELL, J. (1983) Chimeric genes as dominant selectable markers in plant cells. EMBO J., 2, 987-995.
HORSCH, R.B., FRY, J.E., HOFFMANN, N.L., EICHHOLTZ, D., ROGERS, S.G., FRALEY, R.T. (1985) A simple and generaI methodfortransferring genes into plants. Science, 227, 1229-1231.
HUBER, J.L.A., HUBER, S.c. (1992) Site-specific serine phosphorylation of spinach leaf sucrose-phosphate synthase. Biochem. J., 283, 877-882.
JACH, G., GORNHARDT, B., MUNDY, J., LOGEMANN, J., PINSDORF, E., LEAH, R., SCHELL, J., MAAS, C. (1995) Enhanced quantitative resistance against fungal disease by combinatorial expression of different barley antifungal proteins in transgenic tobacco. Plant J., 8, 97-109.
LERCHL, J., GEIGENBERGER, P., STITT, M., SONNEWALD, U. (1995) Inhibition of long-distance sucrose transport by inorganic pyrophosphatase can be complemented by phloem-specific expression of cytosolic yeast-derived invertase in transgenic plants. Plant CelI, 7, 259-270.
LILIUS, G., HOLMBERG, N., BULOW, L. (1996) Enhanced NaCI stress tolerance in transgenic tobacco expressing bacterial choline dehydrogenase. Bio/TechnoIogy, 14, 177-180.
MA, J.K.-C., HIATT, A., HEIN, M., VINE, N.D., WANG, F., STABILA, P., VAN DOLLEWEERD, C., MOSTOV, K., LEHNER, T. (1995) Generation and assembly of secretory antibodies in plants. Science, 268, 716-719.
MARIANI, C., GOSSELE, V., DE BEUCKELEER, M., DE BLOCK, M., GOLDBERG, R.B., DE GREEF, W., LEEMANS, J. (1992) A chimaeric ribonuclease-inhibitor gene restores fertility to male sterile plants. Nature, 357, 384-387.
MARTIN, C., SMITH, A.M. (1995) Starch biosynthesis. Plant CelI, 7, 971-985.
MICALLEF, B.J., SHARKEY, T.D. (1996) Genetic and physiological characterization of Flaveria linearis plants having a reduced activity of cytosolic fructose-I ,6-bisphosphatase. Plant CelI Environ., 19, 1-9.
MIKKELSEN, T.R., ANDERSEN, B., JORGENSEN, R.B. (1996) The risk of crop transgene spread. Nature, 380, 31.
MOL, J.N.M., VAN DER KROL, A.R., VANTUNEN, A.J., VAN BLOKLAND, R., DE LANGE, P., STUlTJE. A.R. (1990) Regulation of plant gene expression by antisense RNA FEBS Lett., 268, 427-430.
MURATA, N., ISHIZAKI-NISHIZAWA, O., HIGASHI, S., HAYASHI, H., TASAKA, Y., NISHIDA, I. (1992) Genetically engineered alteration in the chilling sensitivity of plants. Nature, 356, 710-713.
NAWRATH, C., POIRIER, Y., SOMMERVILLE, C. (1994) Targeting ofthe polyhydroxybutyrate biosynthetic pathway to the plastids of Arabidopsis thalianaresults inhigh-Ievels of polymer accumulation. Proc. Natl. Acad. Sci. USA, 91, 12760-12764.
RUGH, C.L., WILDE, H.D., STACK, N.M., THOMPSON, D.M., MEAGHER, A, MEAGHER, R.B. (1996) Mercuric ion reduction and resistance in transgenic Arabidopsis thaliana plants expressing a modified bacterial merA gene. Proc. Natl. Acad. Sci. USA, 93, 3182-3187.
SCOTT, P., LANGE, A.J., PILKIS, S.J., KRUGER, N.J. (1995) Carbon metabolism in leaves oftransgenic tobacco (Nicotiana tabacum L.) containing elevatedfructose 2,6-bisphosphate levels. PlantJ., 7, 461-469.
SHADE, R.E., SCHROEDER, H.E., PUEYO, J.J., TABE, L.M., MURDOCK, L.L., HIGGINS, T.l.V., CHRISPEELS, M.J. (1994) Transgenic pea seeds expressing the IX-amylase inhibitor ofthe common bean are resistant to bruchid beetles. Bio/Technology, 12, 793-796.
SHEWMAKER, C.K., BOYER, C.D., WIESENBORN, D.P., THOMPSON, D.B., BOERSIG, M.R., OAKES, J.V., STALKER, D.M. (1994) Expression of Escherichia coli glycogen synthase in the tubers of transgenic potatoes (Solanum tuberosum) results in a highly branched starch. Plant Physiol., 104, 1159-1166.
STALKER, D.M., KISER, J.A., BALDWIN, G., COULOMBE, B., HOUCK, C.M. (1996) Cotton weed control using the BXN™ system. In Herbicide-resistant crops: agricultural, environmental, economic, regulatory, and technical aspects, voI. 6, a c. di Duke, S.O., Boca Raton (Florida), CRC Press Inc., pp. 93-105.
STARK, D.M., TIMMERMANN, K.P., BARRY, G.F., PREISS, J., KISHORE, G .M. (1992) Regulation of the amount of starch in planttissues by ADP glucose pyrophosphorylase. Science, 258, 287-292.
STITT, M., SONNEWALD, U. (1995) Regulation of metabolismin transgenic plants. Annu. Rev. Plant Physiol., Plant Mol. Biol., 46, 341-368.
TARCZYNSKI, M.C., JENSEN, R.G., BOHNERT, J.H. (1993) Stress protection of transgenic tobacco by production of the osmolyte mannitol. Science, 259, 508-510.
TERRAS, F.R.G. et al. (1995) Small cysteine-rich antifungal proteins from radish: their role in host defense. Plant CelI, 7, 573-588.
TIEMAN, D.M., HANDA, AK. (1994) Reductionin pectinmethylesterase activitymodifies tissue integrity and cationlevels inripening tornato (Lycopersicum esculentum Mill.) fruits. Plant Physiol., 106, 429-436.
VAN CAMP, W., WILLEKENS, H., BOWLER, C., VAN MONTAGU, M., INZÉ, D., REUPOLD-POPP, P., SANDERMANN, H.Jr., LANGEBARTELS, C. (1994) Elevated levels of superoxide dismutase protect transgenic plants against ozone damage. Bio/Technology, 12, 165-168.
VAN DER MEER, I.M., EBSKAMP, M.J.M., VISSER, R.G.F., WEISBEEK, P.I., SMEEKENS, S.C.M. (1994) Fructan as a new carbohydrate siuk in transgenic potato plants. Plant CelI, 6, 561-570.
VOELKER, T.A., WORRELL, A.C., ANDERSON, L., BLEIBAUM, J., FAN, C., HA WKINS, D.J., RADKE, S.E., DA VIES, H.M. (1992) Fatty acid biosynthesis redirected to medium chains in transgenic oilseed plants. Science, 257, 72-74.
WU, G., SHORTT, B.J., LAWRENCE, E.B., LEVINE, E.B., FITZSIMMONS, K.C., SHAH, D.M. (1995) Disease resistance conferred by expression of a gene encoding H₂O2-generating glucose oxidase in transgenic potato plants. Plant CelI, 7, 1357-1368.
ZAMBRYSKI, P., JOOS, H., GENETELLO, C., LEEMANS, J., VAN MONTAGU, M., SCHELL, J. (1983) Ti plasmid vectorforthe introduction of DNA into plant cells without alteration of their normal regeneration capacity. EMBO J., 2, 2143-2150.
ZRENNER, R., KRAUSE, K.-P., APEL, P., SONNEWALD, U. (1996) Reduction of the cytosolic fructose-I,6-bisphosphatase in transgenic plants limit photosynthetic sucrose biosynthesis with no impact on plant growth and tuber yield. Plant J., 9, 671-681.
Bibliografia generale
ALLEN, R.D. (1995) Dissection of oxidative stress using transgenic plants. Plant Physiol., 107, 1049-1054.
CHRISPEELS, M.J., SADAVA, D. Plants, genes and agriculture. Boston, Jones and Barlett, 1994.
CHRISTOU, P. (1992) Genetic transformation of crop plants using microprojectile bombardment. PIant J., 2, 275-281.
CONRAD, U., FIEDLER, U. (1994) Expression of engineeredantibodies in plant cells. PIant Mol. Biol., 26, 1023-1030.
CUNNINGHAM, S.D., BERTI, W.R., HUANG, J.W. (1995) Phytoremediation of contaminated soils. Trends Biotech., 13, 393-397.
HOOYKAAS, P.J.J., SCHILPEROORT, R.A. (1992) Agrobacterium and plant genetic engineering. Plant Mol. Biol., 19, 15-38.
MASON, H.S., ARNTZEN, C.J. (1995) Transgenic plants as vaccine production systems. Trends Biotechnol., 13, 388-392.
NAWRATH, C., POIRIER, Y., SOMMERVILLE, C. (1995) Plant polymers for biodegradable plastics: cellulose, starch and polyhydroxyalkanoates. Mol. Breeding, I, 105-122.
OHLROGGE, J.B. (1994) Design of new plant products: engineering of fatty acid metabolismo Plant Physiol., 104, 821-826.
YODER, J.I., GOLDSBROUGH, A.P. (1994) Transformation systems for generating marker-free transgenic plants. Bio/Technology, 12, 263-267.