
Razza
Razza
di Guido Modiano
Razza
sommario: 1. Definizione. 2. Criteri di classificazione. 3. Genetica delle razze: a) livello descrittivo o statico; b) livello storico o dinamico; c) la struttura attuale della specie umana come problema storico. 4. I due tipi di evoluzione, genetica e culturale: a) analogie e differenze; velocità relative; b) interazioni fra i due processi evolutivi, quello biologico e quello culturale. 5. Il concetto di razza e i suoi limiti: a) limiti e difetti delle definizioni operative di razza; b) critiche al concetto di razza come prespecie. 6. Le modalità di diversificazione della nostra specie: il ruolo dei vari fattori evolutivi. 7. Diversità genetica e uguaglianza umana: a) scienza e decisioni etico-politiche; b) argomenti cari ai razzisti; c) variabilità dell'intelligenza; d) gli incroci tra razze diverse. □ Bibliografia.
1. Definizione
Le razze sono gruppi sufficientemente numerosi di individui (o di popolazioni) della stessa specie, che si presentano attualmente diversi per un insieme di caratteri ereditari, essendo vissuti in condizioni di isolamento reciproco di durata e grado notevoli, anche se generalmente indeterminati.
Si tratta, come si vede, di una definizione tutt'altro che soddisfacente, perché vaga e imprecisa in tutti i suoi aspetti, e quindi applicabile - ed effettivamente applicata - in modo più o meno arbitrario e soggettivo. Ma - a parte la considerazione che, di fatto, è proprio questo il criterio operativo con cui si procede alle suddivisioni all'interno delle specie - entità vaghe è giusto siano definite in modo vago. In casi del genere sarebbero semmai criticabili definizioni rigorose ed esatte, e proprio in quanto tali.
Si tornerà in seguito sulla validità e sui limiti del concetto di razza, ma conviene sottolineare fin da ora una delle sue componenti essenziali, quella storica. Una definizione delle razze solamente come ‛gruppi di individui della stessa specie che differiscono tra loro per un insieme di caratteri ereditari', che non tenesse conto cioè di come queste differenze si sono venute a creare storicamente, sarebbe incompleta e snaturerebbe il senso di questa suddivisione. Infatti il concetto di isolamento ‛sufficiente' (per grado e durata) implica che, insieme alle differenze constatabili che costituiscono la base della classificazione, se ne siano accumulate anche altre più o meno numerose e importanti.
Nelle pagine seguenti si farà riferimento alla specie umana non solo perché la suddivisione in razze di gran lunga più nota e più ricca riguarda la nostra specie e tiene conto sia dei caratteri ereditari biologici che di quelli culturali, ma soprattutto perché le implicazioni e il significato del termine razza, per quel che riguarda il problema del razzismo, sono esclusivi dell'uomo. Si può dire che pochi concetti biologici sono altrettanto rilevanti sul piano del comportamento sociale e politico. Anzi, la consapevolezza, talora esasperata, dei rischi connessi con una sua eventuale strumentalizzazione, ha rappresentato uno degli ostacoli principali in questo campo, già di per sé molto complesso, rendendo spesso gli ‛addetti ai lavori' non desiderosi di affrontarlo oppure incapaci di farlo in maniera obiettiva e neutrale, cioè nell'unico modo scientificamente valido. La posizione all'estremo opposto - quella dei razzisti, per intenderci - ha condotto senza eccezione a tali assurdi scientifici (come, per esempio, il concetto di ‛purezza della razza'), che non è degna nemmeno di essere presa in considerazione: per essere ammessa a un dibattito scientifico - sia pure per esservi criticata - una teoria deve raggiungere uno standard minimo e soddisfare certi requisiti, tra cui il non essere in palese contrasto con la realtà obiettiva e l'uso di un linguaggio razionale che non violi le regole più elementari della logica.
2. Criteri di classificazione
In linea di principio qualsiasi insieme di caratteri che presentino una forte componente ereditaria e siano distribuiti in modo diverso in gruppi diversi di individui della stessa specie può essere utilizzato per suddividere questa specie in razze.
Nel caso dell'uomo si possono chiaramente individuare tre criteri distintivi: 1) quello dell'antropologia fisica classica, parzialmente sovrapposto a un criterio geografico (Negri o Africani, Caucasoidi o Europoidi, Amerindi, ecc.); 2) quello genetico in senso stretto; 3) quello culturale. Ciascuno di essi è a sua volta costituito da un insieme di sottocriteri. Per esempio, tra i caratteri genetici è fondamentale la distinzione in caratteri fortemente selettivi e caratteri neutri o quasi neutri; per i caratteri culturali il tipo di suddivisione a cui si può arrivare non è certo lo stesso se si adotta un criterio glottologico, religioso, ecc., o una loro particolare combinazione. La distinzione tra i due criteri biologici, quello antropologico classico e quello genetico, non è di principio: evidentemente, anche i caratteri antropologici hanno una determinazione genetica. Essa è però complessa e per lo più non delucidata affatto a livello formale e tanto meno a livello molecolare. Al contrario l'antropologia ‛genetica' si basa su caratteri riconducibili allo schema mendeliano semplice, come i gruppi sanguigni e i polimorfismi proteici.
Non ha alcun senso domandarsi quale dei tre criteri distintivi sia il più valido o il più interessante. Ciascuno ha il suo significato e non è intercambiabile con gli altri due.
Il criterio antropologico classico è in questo momento l'unico che, sia pure con i suoi limiti e le sue vistose imperfezioni, ha permesso di suddividere in modo autonomo (cioè senza l'ausilio di altri criteri) la nostra specie in razze. Gli altri due criteri non hanno finora portato ad alcuna classificazione di carattere generale: il procedimento che in genere si segue quando si utilizzano caratteri genetici o culturali consiste nello studiare comparativamente come essi si distribuiscano in gruppi umani già definiti secondo l'antropologia fisica e geografica, che fornisce senza dubbio, almeno per ora, il più importante - se non l'unico - punto di riferimento dell'antropologia generale. D'altra parte, se limitiamo questa analisi ai soli criteri biologici, è facile constatare che il criterio genetico presenta su quello morfologico classico un vantaggio di estrema importanza: la possibilità di esprimere i caratteri differenziali in esame in termini di frequenze geniche permette di passare dalla fase puramente descrittiva ed essenzialmente statica, caratteristica fino a pochi anni fa dell'antropologia fisica, a quella dinamica, cioè interpretativa, della genetica di popolazione. Si passa cioè da una visione qualitativa, come poteva essere concepita al tempo di Darwin, all'attuale concezione quantitativa della microevoluzione, cioè dell'evoluzione all'interno della specie, scaturita dalla sintesi di darwinismo e mendelismo realizzata da R. A. Fisher, J. B. S. Haldane e S. Wright.
Si può finalmente tentare non solo di descrivere l'attuale differenziazione intraspecifica, ma di capire come ci si è arrivati: in una parola, tracciarne la storia. Non sarà mai una storia rigorosamente esatta - intendiamoci - ma lo saranno almeno i suoi fondamenti. Inoltre i progressi della genetica sono così rapidi che il numero - e il nostro grado di comprensione - dei caratteri differenziali utilizzabili aumentano a un ritmo precedentemente inimmaginabile, rendendo così sempre migliori le prospettive di riuscire a ricostruire, nelle sue linee generali, la storia della nostra specie. Si tratta in un certo senso di una lotta contro il tempo, dato che la maggior parte dell'attuale differenziazione entro la specie umana è destinata a diluirsi e forse a scomparire del tutto in tempi relativamente brevi.
Per tutti questi motivi nelle pagine che seguono noi ci proponiamo di discutere soprattutto gli aspetti genetici del problema rappresentato dal significato - da un punto di vista biologico - del termine razza in rapporto alla specie umana.
3. Genetica delle razze
La genetica delle razze può essere discussa a due livelli: descrittivo e di genesi delle razze. Ad ambedue questi livelli il fatto che i caratteri differenziali in esame siano esprimibili come frequenze di caratteri ‛alternativi' (frequenze geniche e aplotipiche) invece che come caratteri ‛continui', come quelli dell'antropologia fisica, costituisce un vantaggio notevolissimo.
a) Livello descrittivo o statico
Operando a livello descrittivo ci si propone: 1) di individuare i caratteri genetici che possono essere utilizzati come ‛marcatori antropologici' e di valutarne l'interesse e la validità a questo fine, e 2) di stabilire i criteri che si possono adottare - usando appunto questi marcatori - per tentare di classificare e di suddividere i vari gruppi che costituiscono la specie umana. Ci si propone, cioè, di fotografare il grado di differenziazione attuale della nostra specie individuando anche i modi migliori per esprimerlo in termini quantitativi, senza però addentrarsi nei processi storici che hanno condotto al quadro che si sta osservando.
1. I geni come marcatori antropologici. - Un gene può non essere di alcuna utilità per distinguere due popolazioni; questo accade se in entrambe esiste uno stesso e unico allele di quel gene, oppure se in tutte e due questo gene è polimorfico ma con le stesse frequenze degli stessi alleli comuni (in questo caso si tratta di eterogeneità intrapopolazioni, cioè polimorfismo, e non interpopolazioni, cioè politipismo). In tutti gli altri casi i geni possono contribuire a distinguere popolazioni diverse, e fungono pertanto da marcatori antropologici.
α) Sostituzioni alleliche parziali. La situazione più comune è quella di un gene polimorfico in tutte le popolazioni in esame e con gli stessi alleli comuni, che presentano però frequenze differenti. Rientrano in questo gruppo, fra gli altri, quasi tutti i sistemi di gruppo sanguigno, i polimorfismi biochimici eritrocitari e sierici e quelli delle immunoglobuline (v. sangue: Genetica del sangue) e i polimorfismi degli antigeni della istocompatibilità.
È chiaro che nessun gene di questo tipo può servire ad assegnare a un gruppo un singolo individuo: solo insiemi più o meno grandi di individui possono essere attribuiti a questa o a quella popolazione con un certo livello definito di verosimiglianza.
Solo quando un allele comune è proprio di una sola popolazione diventa talvolta possibile attribuire singoli individui a quella popolazione. È il caso dei polimorfismi cosiddetti ‛privati' (in cui una piccola popolazione, oltre all'allele o agli alleli che compaiono anche nelle altre, ne presenta uno suo particolare) e degli alleli limitati a certe razze principali (per esempio l'allele AcPc della fosfatasi acida si trova solo tra i Caucasici). La quota di individui attribuibili singolarmente alla popolazione cui appartengono è, in base alla legge di Hardy-Weinberg, (1-p2), se p è la frequenza globale degli alleli diversi da quello proprio della popolazione.
β) Sostituzioni alleliche complete. Si può immaginare una sola situazione semplice in cui, con l'esame di un solo gene, si potrebbero correttamente classificare tutti gli individui di una popolazione, uno per uno. Questo gene dovrebbe presentare una sola forma allelica in una popolazione e una sola forma allelica (diversa dalla prima) anche nelle altre. Se, come si reputa che accada di regola, questa differenza allelica consiste in una singola sostituzione amminoacidica, essa costituisce quella che si chiama ‛unità di evoluzione' (cioè fissazione di una sostituzione amminoacidica), e allora le differenze di frequenze alleliche inferiori a 1 che, come si è detto prima, sono le più comuni, devono essere considerate come ‛frazioni di unità di evoluzione'. Nei casi in cui si sia appunto verificata una sostituzione allelica completa tra due popolazioni al loro interno omogenee, si può dire che l'eterogeneità relativa a quel gene consiste di una sola componente, quella ‛tra' popolazioni: si ha cioè politipismo senza polimorfismo.
A livello di intere razze principali non si conosce nemmeno un caso in cui si sia arrivati a questo punto, mentre situazioni in cui ci si è molto vicini o addirittura ciò si è verificato sono state descritte per popolazioni limitate. Per esempio, tra gli Ebrei del Kurdistan, un allele Gd- ha una frequenza dell'ordine di 0,70-0,75, mentre esso è del tutto assente nella maggior parte del resto dell'umanità. L'allele βE raggiunge in certe popolazioni dell'Asia sudorientale (in Birmania) frequenze di circa 0,55, cioè è presente in circa l'80% degli individui, mentre è praticamente assente (trascurando le migrazioni) in tutto il resto dell'umanità: si può quindi asserire che l'ascendenza di un soggetto portatore di un βE risale sicuramente all'Asia sudorientale, mentre un individuo di questa parte del mondo solo di rado è privo di questo marcatore.
Un caso estremo è quello dell'allele Fy del sistema di gruppo sanguigno Duffy, che non produce nessun tipo di antigene Duffy noto e che quindi allo stato omozigote FyFy conferisce resistenza assoluta al Plasmodium vivax, l'agente patogeno della quartana (l'antigene Duffy è necessario per la penetrazione di questo plasmodio nel globulo rosso, per cui si suppone che sia il suo recettore specifico di membrana o, quantomeno, che sia con questo associato). Questo è l'unico allele del sistema Duffy che si trova tra i Negri del Gambia, dove la malaria da Plasmodium vivax non è stata mai osservata, mentre esso è praticamente assente nei Caucasici (che non sono resistenti a questo plasmodio); questi ultimi presentano invece due forme alleliche (Fya e Fyb), entrambe molto comuni, che producono ambedue un antigene Duffy, il Duffya e il Duffyb. Ci si trova quindi di fronte a un esempio di eterogeneità massima tra popolazioni, una delle quali, inoltre, è considerevolmente polimorfica al suo interno.
Vale la pena di sottolineare il fatto che, di questi tre esempi di eterogeneità assoluta, o quasi, tra popolazioni, due con certezza (gli alleli Gd- e Fy) e uno molto probabilmente (il βE) sono stati causati da un agente selettivo (in tutti e tre i casi la malaria).
Resta comunque il fatto che, allo stato attuale delle nostre conoscenze, sono pochi i geni che ci permettono, da soli, di attribuire (o di escludere) singoli individui a (o da) questo o quel gruppo razziale. È tuttavia ragionevole sospettare che tra i geni responsabili delle differenze razziali dell'antropologia fisica ve ne siano molti di questo tipo.
2. Misure di eterogeneità genetica e potere discriminante dei geni (diversità tra individui e tra gruppi di individui). - Le unità della genetica di popolazione sono il ‛gamete' e l'‛individuo', quindi gli indici di eterogeneità per un certo gene sono rappresentati dalla probabilità che un gamete scelto a caso sia diverso per quel gene da un altro gamete anch'esso estratto a caso dalla stessa popolazione (grado di eterozigosi), oppure che due individui qualsiasi abbiano genotipo diverso sempre per quel gene.
Le unità della genetica di una specie sono invece ‛gruppi' di gameti o di individui di quella specie (siano essi chiamati popolazioni o razze o demi), quindi gli indici di eterogeneità sono necessariamente più complessi. In entrambi i casi, comunque, gli indici di eterogeneità devono essere definiti in modo tale che, per ogni gene, al loro aumento corrisponda un aumento della probabilità che, esaminando quel gene, si sia in grado di distinguere tra un'unità (gamete, individuo, popolazione) e un'altra.
Gli indici genetici di eterogeneità tra gruppi rappresentano un progresso notevole rispetto a quelli precedenti, perchè il loro impiego rende possibile misurare allo stesso tempo l'eterogeneità sia tra i gruppi che all'interno di un singolo gruppo. Come problema non è certo nuovo; nuovo è il fatto che si sia trovato il modo di risolverlo.
Considereremo in breve qui di seguito i principali criteri usati per valutare l'eterogeneità tra gruppi di individui.
α) Deficit di eterozigoti. Se due alleli A1 e A2 di un gene A hanno le frequenze pi e qi in una popolazione i, e pj e qj in un'altra popolazione j (che considereremo per semplicità altrettanto numerosa) e le due popolazioni sono isolate riproduttivamente l'una dall'altra, la frequenza totale attesa degli eterozigoti Hettot, è:
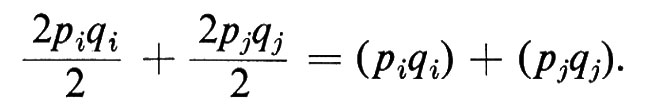
Dato che le frequenze medie degli alleli A1 e A2 sono rispettivamente:
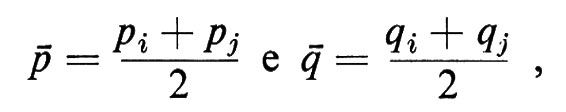
se le due popolazioni si fondessero in un'unica popolazione mendeliana, la frequenza 2äö degli eterozigoti sarebbe:

In condizioni di isolamento riproduttivo questo valore ‛massimo' di frequenza totale di eterozigoti si verifica solo se le frequenze degli alleli A1 e A2 sono le stesse nelle due popolazioni; l'isolamento è privo di conseguenze quando tiene separate popolazioni identiche. Ma, se esiste una eterogeneità genetica interpopolazioni (pi ≠ pj), la frequenza totale degli eterozigoti è solo una frazione di 2äö; tanto più piccola quanto più diverse sono le due popolazioni. Quindi il grado di eterogeneità tra popolazioni può essere espresso, per un certo gene, come deficit relativo di eterozigoti:
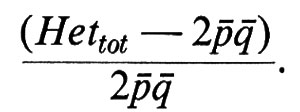
Se si indica con ö la frequenza globale dell'allele meno comune (q ≤ 0,5), la differenza tra due popolazioni (qi − qj) può assumere valori compresi tra 0, se qi = qj, e 2ö; se qi = 2ö e qj = 0; quest'ultimo caso si verifica quando l'allele meno comune si trova solo in una delle due popolazioni (massima eterogeneità interpopolazioni). A sua volta 2ö può arrivare fino a 1, se ö = 0,5 (v. sotto).
Per bassi valori di ö (ö〈 0,05) il valore massimo del deficit relativo di eterozigoti coincide quasi perfettamente con −ö; con il crescere di ö questo valore aumenta, fino a raggiungere −2ö quando ö = 0,5 (per ö → 0 il deficit relativo tende a −ö; per ö → 0,5 il deficit relativo tende a −2ö, cioè a 1).
Per esempio, se l'allele A2 ha una frequenza media ö = = 0,01, il massimo possibile deficit ‛assoluto' di eterozigoti (cioè A2 si trova in una sola delle due popolazioni dove ha la frequenza di 0,02) è solo 0,0002 (cioè: 0,0196 − 0,0198) che, espresso rispetto a 2äö; che è 0,0198, cioè come deficit ‛relativo', corrisponde a un valore di 0,0101 che è quasi identico a ö; che è un numero piccolo; vengono a mancare, cioè, ‛solo pochi dei pochi' eterozigoti che ci sarebbero se non ci fosse isolamento riproduttivo. Se invece ö è 0,5, il massimo deficit ‛assoluto' di eterozigoti è 0,5 che, espresso rispetto a 2äö; cioè come deficit ‛relativo', corrisponde a un valore di 1 che è anche uguale a 2äö; vengono cioè a mancare ‛tutti i molti' eterozigoti che ci sarebbero se non ci fosse isolamento riproduttivo.
Se si considera che l'eterogeneità genetica totale è la somma di due componenti, quella intrapopolazioni e quella interpopolazioni, e che la prima è l'Hettot (cioè la somma delle frequenze degli eterozigoti esistenti all'interno delle due popolazioni), mentre la seconda corrisponde al deficit di eterozigoti (cioè all'aumento di eterozigoti che si verificherebbe se scomparisse la barriera riproduttiva tra le due popolazioni), il significato di questa relazione risulta evidente: nel caso di geni con scarsa variabilità (ö → 0), quasi tutta la scarsa eterogeneità esistente è di tipo intrapopolazioni anche nella condizione di massima eterogeneità possibile tra popolazioni. Se invece l'allele A2 è nel suo insieme molto comune (ö → 0,5), sono possibili tutti i casi compresi tra questi due estremi: 1) assenza di eterogeneità interpopolazioni (qi = qj = ö), e allora non si ha deficit di eterozigoti, cioè l'intera eterogeneità genetica 2äö è di tipo intrapopolazioni (da questo punto di vista, infatti, è come se le due popolazioni costituissero una singola popolazione suddivisa in due parti eguali); 2) massimo grado di eterogeneità interpopolazioni (qi = 1,0; qj = 0,0; ä = ö = 0,5; (qi − qi) = 2ö = 1,0; deficit relativo di eterozigoti = 2ö = 1,0), e allora tutta quanta la cospicua eterogeneità del gene A è di tipo interpopolazioni, e infatti ciascuna delle due popolazioni è monomorfica, cioè al suo interno è omogenea. È quindi evidente che geni poco variabili non possono in nessun caso marcare molto distintamente due ‛popolazioni' pur potendo riuscire utili per marcare alcuni ‛individui' di una popolazione; mentre quelli molto variabili possono farlo se soddisfano all'ulteriore requisito che i loro alleli comuni siano distribuiti in modo molto ineguale, cioè se tendono a essere politipici più che polimorfici.
β) Grado di diversità tra gameti o tra individui estratti dalla stessa o da differenti popolazioni. Quanto maggiore è la quota di eterogeneità genetica che ha, per così dire, segregato, cioè suddiviso una popolazione in due popolazioni diverse, tanto maggiore è la frequenza di eterozigoti che si otterrebbero con incroci interpopolazioni rispetto a quelli che si otterrebbero con incroci intrapopolazioni. L'eterogeneità (relativa a un determinato gene) tra gameti o individui estratti da due popolazioni può essere la stessa che si troverebbe se essi provenissero dalla stessa popolazione solo nel caso che queste due popolazioni siano identiche per quel gene, altrimenti è maggiore. Quindi l'incremento assoluto o relativo di probabilità che i gameti o gli individui risultino diversi per il fatto di averli estratti da due popolazioni invece che dalla stessa può costituire un indice ulteriore di eterogeneità interpopolazioni. L'eterogeneità totale tra l'insieme dei gameti delle due popolazioni resta costante, e pari a 2äö; quanto più le due popolazioni diventano uniformi al loro interno e quanto più l'eterogeneità diventa di tipo interpopolazioni, tanto più aumenta - di un valore corrispondente - il grado di eterozigosi tra gli zigoti risultanti dall'unione di gameti di popolazioni diverse.
La situazione limite è quella della politipia completa, cioè di popolazioni monomorfiche per alleli diversi; in questo caso nessuno degli incroci all'interno di una popolazione darebbe origine a eterozigoti e l'intera progenie di tutti gli incroci misti sarebbe costituita da eterozigoti. Come si è accennato prima, verosimilmente questo è quanto accade per molti dei geni non identificati che determinano i caratteri evidenziati dall'antropologia fisica: per esempio è perlomeno ragionevole supporre che per alcuni dei geni che controllano la pigmentazione della cute si sia fissato in certe razze un allele e in altre razze un altro allele, dato che nella progenie di Caucasici non si trova mai un soggetto con la pelle scura come quella degli Africani e, viceversa, non si trova mai un soggetto normale con la pelle chiara nella progenie di Africani.
L'indice di eterogeneità tra gruppi più comunemente usato è l'indice di Nei; siccome, però, questo indice è lievemente più complesso di quelli appena discussi, si ritiene opportuno rimandare, per una sua trattazione esauriente, a testi specializzati (v., per esempio, Cavalli-Sforza e Bodmer, 1971).
b) Livello storico o dinamico
È questo, evidentemente, un approccio molto più ambizioso di quello semplicemente descrittivo precedentemente illustrato, perchè si propone di spiegare come si sia arrivati all'attuale struttura genetica della specie umana. Il suo scopo ultimo consiste quindi nella ricostruzione della struttura originaria della nostra specie e nel tentativo di predirne per estrapolazione il destino, almeno nelle sue grandi linee, dopo aver individuato le forze che hanno condotto alle trasformazioni attuali.
I processi in gioco sono talmente complicati che si rende necessario introdurre un certo schematismo, trattando separatamente eventi che spesso rappresentano aspetti complementari dello stesso fenomeno (come la formazione e la scomparsa di gruppi razziali per mescolamento di razze diverse), anche a costo di apparire troppo semplicistici. La realtà è in effetti molto più complessa e sfumata di quanto possa risultare dalla seguente trattazione.
1. Come si formano le razze genetiche (individuate, cioè, con criteri genetici). - I meccanismi evolutivi responsabili della formazione di entità subspecifiche del tipo delle razze coincidono almeno in parte con quelli della speciazione, differendone sul piano quantitativo per il fatto di non aver proceduto abbastanza innanzi da produrre nuove specie. Il presupposto necessario - lo stesso in entrambi i casi - è l'isolamento riproduttivo, conditio sine qua non perché possa aver luogo un'evoluzione divergente, a sua volta possibile solo se esiste una variabilità genetica. L'evoluzione da sola evidentemente non crea diversificazione nel senso di molteplicità di gruppi geneticamente diversi: ‛una' entità riproduttiva rimane ‛una' entità riproduttiva, anche se diversa da come era prima. È con l'isolamento riproduttivo di gruppi inizialmente identici o comunque molto simili (perché derivati dalla scissione in frazioni della stessa unità) che si crea la premessa perché si formino, a causa dell'evoluzione divergente, altrettanti gruppi diversi tra loro, oltre che da quello da cui tutti si sono originati.
L'isolamento riproduttivo è dovuto a barriere che possono essere di varia natura, per esempio geografiche. Per i gruppi simpatrici sempre, e per gli altri molto spesso, le prime barriere riproduttive sono etologiche - nel caso dell'uomo si preferisce parlare di barriere sociali, culturali, politiche, religiose, di casta ecc.; queste barriere possono essere molto efficienti e preparare il terreno perché si formino in seguito, e in tempi molto più lunghi, barriere genetiche e fisiologiche.
L'evoluzione divergente procede attraverso gli stessi meccanismi elementari dell'evoluzione in generale (v. evoluzione) e il grado finale di divergenza è la risultante dell'azione dei vari fattori in gioco che si sovrappongono al grado di isolamento (che può anche non essere assoluto). Da una parte agiscono i fattori deterministici, cioè selettivi, come la selezione stabilizzante e la selezione adattativa ad ambienti diversi, e dall'altra quelli stocastici, come la mutazione, la deriva genetica e la commistione genetica con altri gruppi. Qui di seguito sono rappresentati in modo schematico alcuni dei possibili destini di geni polimorfici in gruppi inizialmente identici che si sono separati riproduttivamente.
Per semplicità non si sono rappresentati i possibili effetti di eventuali altri fattori, come la commistione genetica (o genetic admixture) tra le popolazioni che stanno evolvendo in modo indipendente, e tra queste popolazioni e altre ancora che non sono mostrate negli schemi. Su quest'ultimo meccanismo, che ha certamente giocato un ruolo fondamentale nella formazione della maggior parte delle razze (anzi di tutte, purché si risalga abbastanza a ritroso lungo la loro storia), torneremo in seguito (v. sotto, 2). Un altro possibile fattore di divergenza che non si è preso in considerazione è il cosiddetto ‛effetto del fondatore' (una popolazione derivata da un piccolo gruppo può presentare con frequenza elevata uno o più alleli altrove rari, perché il piccolo gruppo dei ‛fondatori' può aver portato con sé per caso uno di questi alleli; proprio a causa della limitatezza del gruppo, questo allele è stato certo molto frequente almeno in un primo momento - tutti gli alleli presenti in una popolazione molto piccola hanno necessariamente una frequenza relativa piuttosto elevata - e talvolta rimane frequente anche in seguito).
Nel complesso, la diversificazione genetica, cioè la formazione di nuove razze, è la risultante degli effetti di forze contrastanti, che la ostacolano oppure la favoriscono.
α) Fattori che si oppongono alla diversificazione genetica. Oltre, naturalmente, agli scambi riproduttivi, uno dei fattori principali che si oppongono alla diversificazione genetica è la selezione stabilizzante, che tende a mantenere lo status quo opponendosi a variazioni, anche casuali, della struttura genetica iniziale. Anche processi selettivi che tendano a modificare le frequenze geniche iniziali possono ostacolare la diversificazione genetica, se agiscono nello stesso senso in popolazioni diverse. Questi fenomeni di ‛non divergenza evolutiva' si devono ipotizzare soprattutto nel caso di popolazioni isolate fra loro, ma che si sono evolute in presenza di agenti selettivi comuni: in altre parole, a storie ambientali simili spesso corrispondono fenotipi simili. È però molto difficile, in pratica, distinguere tra somiglianze fenotipiche dovute a ‛non divergenza' (alla cui base c'è di regola una somiglianza molto stretta, o addirittura una completa identità genetica) e somiglianze genetiche prodotte da ‛convergenza' evolutiva, e solo in alcuni casi si è riusciti a individuare con precisione l'origine di tali somiglianze.
Per esempio, è noto da circa trent'anni che la talassemia (Th) e l'enzimopenia per la glucosio-6-fosfatodeidrogenasi (Gd- (v. sangue: Genetica del sangue) sono molto frequenti nelle popolazioni esposte per tempi molto lunghi alla malaria, ma è solo da poco che si è dimostrato che entrambe queste caratteristiche genetiche selezionate dalla malaria (in quanto conferiscono resistenza a questo fattore selettivo) sono molto eterogenee, nel senso che gli alleli Th comuni nella popolazione sarda non sono gli stessi di quelli trovati nelle popolazioni del delta padano e questi, a loro volta, sono diversi da quelli delle popolazioni negre e da quelli delle popolazioni dell'Asia sudorientale. Lo stesso discorso vale per gli alleli Gd-. È evidente che ci si trova di fronte, in casi di questo genere, a fenomeni di convergenza evolutiva, oltretutto parziale: gli alleli Th e Gd- di queste popolazioni sono simili per il loro effetto selettivo finale, la resistenza alla malaria, che è quello evolutivamente rilevante, ed entro questi limiti è giusto parlare di convergenza evolutiva e di ‛razze malariche', cioè di razze che, avendo tutte avuto una lunga storia malarica, hanno ora caratteristiche genetiche che conferiscono resistenza a questa malattia. Ma, al di là del risultato selettivo e al di là di un livello fenotipico relativamente semplice (fenotipi talassemici ed enzimopenici) la somiglianza cessa e con essa il grado di convergenza: ogni popolazione si è creata per conto suo i propri alleli Th e Gd-. Processi di convergenza evolutiva come questi sono il frutto dell'azione combinata della selezione e del caso. La selezione è responsabile del fatto che nelle zone malariche e non altrove siano molto diffusi alleli Th e Gd-; mentre è presumibilmente solo, o quasi solo, per caso che in Sardegna ci siano proprio quei dati alleli Th e Gd-, diversi da quelli di altre popolazioni ‛malariche'.
Nella maggior parte dei casi, invece, non è nota la base molecolare dei caratteri genetici che rendono simili tra loro popolazioni molto diverse per numerosi altri aspetti, per cui non siamo in grado di valutare in che misura la loro somiglianza sia dovuta a non divergenza e in che misura sia invece dovuta a convergenza evolutiva. Questo vale, ad esempio, per l'intensa pigmentazione della pelle degli Africani e degli aborigeni australiani.
In linea di massima, si può affermare che vere identità si possono formare solo per non divergenza, mentre è quasi impossibile che popolazioni diverse si costruiscano ciascuna per conto proprio nello stesso identico modo la base genetica per un fenotipo complesso, quando questo fenotipo può essere determinato da numerosi genotipi. In generale, fenomeni cospicui di convergenza evolutiva come i ‛polimorfismi malarici' stanno a indicare che non c'erano molte altre scelte adattative a disposizione: se ci fossero state altre strade evolutive a portata di mano, è presumibile che esse sarebbero state imboccate (tanto più che questi adattamenti sono ben lontani dall'ottimale; v. sotto, cap. 4, § b) e ora non esisterebbe una convergenza di grado così elevato, sia pure solo a livello fenotipico.
β) Fattori che causano diversificazione genetica. Popolazioni separate riproduttivamente possono divergere nel corso della loro evoluzione a causa di due fattori: il caso, cioè la deriva genetica, e la selezione. I due modi di divergere che ne risultano differiscono per molti aspetti, di cui uno, particolarmente interessante, riguarda le relazioni che si possono stabilire tra i gradi di divergenza dei vari geni che stanno evolvendo contemporaneamente in popolazioni isolate fra loro.
Se la spinta evolutiva è costituita dalla selezione, non ci si attende di trovare alcuna correlazione tra i gradi di divergenza mostrati dai vari geni (a meno che siano spinti dallo stesso fattore di selezione, come nel caso dei geni Th e Gd-).
Al contrario, geni diversi che evolvono contemporaneamente sotto l'azione della sola deriva genetica in popolazioni separate dovrebbero tendere a divergere nella stessa misura. Infatti, dato che la divergenza globale (cioè accumulatasi dal momento della separazione riproduttiva tra le varie popolazioni) dipende, per ogni gene soggetto esclusivamente alla deriva genetica, solo dalla ‛storia demografica' di queste popolazioni - e questa storia è stata certamente la stessa per l'intero genoma -, ne consegue che la deriva ha avuto la stessa opportunità di agire su tutti i geni. Non importa che non sappiamo quanto grande sia stata questa opportunità (perché non conosciamo le ‛storie demografiche' delle popolazioni che si stanno confrontando): possiamo lo stesso esser certi che tutti i geni sono stati sottoposti a una identica deriva. Ci si attende allora che le frequenze di tutti gli alleli ‛neutri' che evolvono indipendentemente in popolazioni diverse divergano fra loro nella stessa misura, purché queste divergenze siano opportunamente standardizzate riferendole, per ogni allele (la cui frequenza sia p), alla varianza p (1 − p) della sua frequenza.
In conclusione, se si misurano i gradi di divergenza (esprimendoli, per esempio, come distanze genetiche) tra varie popolazioni per un certo numero di polimorfismi, è possibile individuare quali di essi si sono comportati come neutri: sono quelli per i quali si giunge a stime ragionevolmente concordanti; e quali quelli le cui frequenze alleliche attuali sono state determinate anche dalla selezione: sono quelli per i quali le distanze genetiche tra le varie popolazioni sono risultate molto differenti da quelle trovate per i polimorfismi concordanti. Per esempio, le distanze genetiche tra Africani e altri gruppi sono risultate particolarmente grandi per gli alleli ‛malarici' Th, Gd- e Fy, proprio come ci si attendeva.
2. Come scompaiono le razze genetiche. - Quando si parla di scomparsa di una razza non è più possibile tener distinte le sue due componenti, quella biologica e quella culturale. Nella maggioranza dei casi, una buona parte della serie di caratteri ereditari distintivi che individuano una razza sono sia biologici sia culturali, ma non sempre è così: per esempio, gli Ebrei, se sono una razza, lo sono solo dal punto di vista culturale, dato che sul piano genetico e antropologico formano un gruppo estremamente eterogeneo, composto da un insieme di sottogruppi, ciascuno dei quali è in genere molto più simile alla popolazione con la quale ha vissuto a lungo, rappresentandone una minoranza, che agli altri sottogruppi ebraici. In generale, ci si può trovare di fronte a un ampio spettro di possibilità, da quella che una determinata razza sia individuabile quasi esclusivamente per alcuni suoi caratteri distintivi biologici a quella che sia individuabile quasi soltanto per alcuni suoi caratteri distintivi culturali; fra questi due casi estremi si colloca tutta una gamma di situazioni intermedie, che sono poi le più comuni.
Anche per quel che riguarda la scomparsa delle razze, il ruolo giocato da queste due componenti può essere molto diverso. Talora esse seguono un destino parallelo, cioè i marcatori biologici che contraddistinguono una razza in via di estinzione scompaiono di pari passo con la sua cultura e le sue tradizioni; in certi casi possono presentare un andamento simile non solo i processi quantitativi della scomparsa biologica e di quella culturale di una razza, ma perfino - sul piano formale - i meccanismi che li provocano. Infatti una razza può essere cancellata più o meno completamente attraverso due meccanismi: la soppressione vera e propria, del tipo ‛soluzione finale', e il mescolamento globale con una o più razze, che porta, insieme alla scomparsa delle razze coinvolte, anche alla formazione di una o più razze nuove. Ebbene, in questi casi la componente biologica e quella culturale tendono di regola a seguire lo stesso destino: quando a livello fisico, cioè biologico, si attua la soppressione vera e propria di un gruppo etnico, come nel caso degli Indiani del Nordamerica nel secolo scorso e degli Armeni e degli Ebrei in questo secolo, si tende a sopprimerne, nel contempo, anche la cultura o quantomeno il tessuto sociale; quando invece le componenti biologiche distintive di una razza scompaiono per mescolamento, anche quelle culturali tendono a scomparire allo stesso modo, cioè, in realtà, più che a scomparire, a mescolarsi e a interagire con le caratteristiche culturali dei gruppi etnici con i quali si viene attuando la fusione. Si generano così nuove razze, sia biologiche che culturali, come è accaduto per i Coloured del Sudafrica risultati dalla fusione di Negri, Orientali e Caucasici.
Però, come si è detto prima, non sempre nell'estinzione di una razza la componente biologica e quella culturale scompaiono di pari passo. Per esempio, nelle popolazioni dell'America Latina l'estinzione culturale è stata estremamente più accentuata di quella biologica, almeno a giudicare da alcuni dei parametri culturali più significativi, come la lingua e la religione (ma si deve essere cauti nel valutare il grado di scomparsa di una cultura solo con questi criteri).
Presumibilmente, l'evento che più spesso è stato responsabile della formazione di nuove razze attraverso il meccanismo della scomparsa di razze preesistenti che hanno mescolato i loro patrimoni ereditari - sia biologici che culturali - è stata la migrazione, più o meno pacifica. Di regola è molto difficile, quando ci si trova di fronte all'avvenuta diffusione di una certa cultura, effettuare una stima attendibile, sia pure approssimativa, della componente biologica e della componente culturale di quella migrazione: verificare cioè quanto le idee si sono diffuse per via demica (cioè attraverso la migrazione di gruppi di individui, e quindi di geni, di entità paragonabile a quelli delle popolazioni che hanno ricevuto la nuova cultura) e quanto per via culturale (al limite, anche un solo individuo può introdurre l'agricoltura in un intero popolo).
In un solo caso, finora, questo problema è stato affrontato in modo adeguato: è noto che l'agricoltura si è diffusa nel Vecchio Mondo partendo da un centro situato nella Mesopotamia, dove sarebbe sorta circa 10.000 anni fa. Lo studio della distribuzione delle frequenze geniche per un gran numero di marcatori antropologici nelle popolazioni europee ha portato all'individuazione di un cline cumulativo disposto lungo l'asse di diffusione di questa innovazione culturale, e questo evidentemente indica che insieme a essa si sono diffusi i geni dei suoi inventori, i quali non hanno soltanto diffuso la loro cultura, ma si sono spostati in massa essi stessi, mescolando il proprio patrimonio genetico con quello delle popolazioni che via via incontravano (v. Menozzi e altri, 1978).
α) Stime del grado di mescolamento genetico (cioè del contributo relativo di razze diverse alla formazione di razze miste nuove). Il patrimonio genetico di un gruppo etnico può, a differenza della sua cultura, essere espresso in termini quantitativi, sotto forma di frequenze geniche e aplotipiche. Quindi siamo in grado di seguire e misurare l'andamento e l'entità della scomparsa di una razza solo per la sua componente genetica. Quello che accade alla sua componente culturale, invece, non è suscettibile, almeno per il momento, di essere descritto in termini quantitativi.
Quando una popolazione si estingue semplicemente perché gli individui che la costituiscono non lasciano progenie (perché vengono soppressi o per qualsiasi altro motivo), allora non esistono problemi nel descrivere l'andamento del fenomeno da un punto di vista genetico quantitativo esso è descritto adeguatamente dall'andamento dei parametri demografici di quella popolazione, anzi addirittura si identifica con esso (il patrimonio genetico potenziale di una popolazione non è altro, in prima approssimazione, che il numero di individui appartenenti a tale popolazione che non hanno ancora superato l'età feconda).
Ma una razza può ‛scomparire' (in realtà non si tratta di una vera e propria scomparsa) anche in un altro modo: mescolandosi geneticamente con altre. In questi casi, che sono molto più comuni e interessanti dei precedenti, il patrimonio genetico della razza scomparsa non è andato perduto, è solo trasformato: le razze che si sono mescolate non esistono effettivamente più come tali, sebbene i loro patrimoni genetici continuino a esistere sotto forma di una parte del patrimonio genetico della nuova razza generatasi dal loro mescolamento (si tratta di un processo simile a quello che si verifica se si mescolano due soluzioni, una di idrato di sodio e l'altra di acido cloridrico: non scompaiono né gli ioni sodio né quelli cloro, ma la soluzione risultante non è né acida né basica ed è quindi completamente diversa dalle soluzioni da cui è derivata).
L'entità del mescolamento (spesso indicata con il termine ‛commistione genetica') è un parametro che rappresenta il contributo genetico dato alla formazione della nuova razza da parte della razza minoritaria. Per esempio, se una razza M (mista) risulta dall'unione di due razze A e Be il loro contributo relativo è 90 e 10, si parla di commistione genetica da parte di B, che può essere stimata, utilizzando un qualsiasi marcatore genetico che presenti frequenze alleliche diverse nelle due razze che si sono mescolate, con la seguente formula:
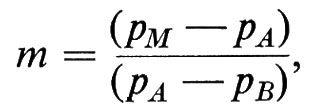
dove pM, pA e pB sono le frequenze dell'allele in esame rispettivamente nella popolazione mista, nella A e nella B. E evidente che, sebbene in linea di principio qualsiasi allele per il quale si verifichi la condizione pA ≠ pB possa essere utilizzato per stimare m, si prestano in pratica a tale scopo solo alleli per i quali pA è ‛mòlto' diverso da pB; la situazione ideale è quella in cui non solo pA è molto diverso da pB, ma in più pA = 0; in questo caso infatti tutti gli alleli di quel tipo (la cui frequenza è pM) che si trovano nella popolazione mista M sono necessariamente derivati da B.
β) Motivi di incertezza nelle stime di m. Queste stime sono sempre notevolmente approssimate, in primo luogo perché dipendono da tre stime di frequenze geniche ciascuna delle quali ha un suo proprio errore che, indicando la frequenza con p, è:
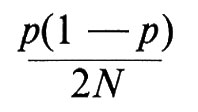
(dove 2N è il numero di geni studiati, se N sono gli individui che costituiscono il campione in esame); in secondo luogo perché sono soggette a fonti di errore non valutabili.
Una fonte di errore consiste nel fatto che l'unica frequenza che si può stimare in modo veramente diretto è pM; pA e pB, invece, sono in realtà, più che stimate, inferite, e questo per due motivi: 1) le popolazioni A e B di partenza non sono quasi mai note con certezza (per esempio nella stima della commistione genetica caucasica nella popolazione nera degli Stati Uniti non è del tutto chiaro cosa si debba intendere esattamente per Africani, cioè per popolazione A, e cosa per Caucasici, cioè per popolazione B, visto che di regola gli alleli che si utilizzano a questo scopo non hanno affatto le stesse frequenze in tutti i Caucasici e in tutti gli Africani); 2) l'immissione di geni B in una popolazione A si attua quasi sempre nel corso di molte generazioni, per cui non è possibile sapere quali fossero le frequenze pA e pB nelle popolazioni A e B quando hanno cominciato a mescolarsi e si deve quindi assumere che già allora fossero le stesse che troviamo oggi.
Un'altra importante possibile causa di errore dipende dal fatto che la formula con cui si calcola m è valida solo se l'unico meccanismo che ha influenzato pA durante la sua trasformazione in pM stato la commistione genetica da parte della popolazione B. In particolare, la selezione non deve avere accelerato nè frenato lo spostamento, causato dalla commistione genetica, di pA verso il valore pB. Se ciò si fosse verificato, si giungerebbe a sovrastimare o, rispettivamente, a sottostimare m.
La miglior garanzia contro tutte queste fonti di errore deriva dal fatto che la commistione genetica è uguale per tutti i geni (il contributo di B al patrimonio genetico di M non avviene per singoli geni, ma per gameti, cioè per genomi completi), per cui ci si attende che tutte le stime di m, ciascuna riferentesi a un determinato allele, relative a una certa popolazione mista M, risultino uguali. Quello che in effetti si trova è che, se si esamina un numero adeguato di geni (per esempio una quindicina), quasi tutti gli m risultano molto simili tranne due o tre che risultano molto diversi dagli altri. Si conclude allora che la stima giusta è quella intorno alla quale si raccolgono la maggioranza degli m, e che gli m molto diversi riguardano geni le cui frequenze, oltre che per commistione genetica, sono cambiate anche per effetto della selezione. Anzi, questo costituisce un ulteriore approccio indiretto al problema di individuare quali polimorfismi sono stati soggetti anche alla selezione e quali solo alla deriva genetica (v. È a, 2). Con uno studio di questo genere si è giunti a una stima di m di circa 0,10 per la commistione genetica caucasica tra i neri viventi in Georgia (S.U.A.), basata sui risultati concordanti ottenuti su 12 geni, mentre per i geni βS, Gd-, T e Hp si è arrivati a valori molto più elevati (~0,45), il che rappresenta un'ottima conferma della validità di questo metodo, perché gli alleli βS e Gd- sono notoriamente soggetti a intensa selezione.
c) La struttura attuale della specie umana come problema storico
Supponiamo che la specie umana sia suddivisa in un certo numero di gruppi ben individuati, che potremmo chiamare razze. Anche se ci si trovasse di fronte a una situazione così chiara, sorgerebbe comunque il problema di cercare di ricostruire come ci si è arrivati.
Si tratta di un problema storico, non diverso in linea di principio da tutti gli altri. Anche in questo caso si devono, in una prima fase, individuare, descrivere e comprendere i fenomeni elementari del processo storico in esame, che potremmo chiamare processi fondamentali e comuni; quelli, per intendersi, che nella storia nel senso usuale sono le guerre, le rivoluzioni, le riforme, le invasioni, il caso ecc. e che, parlando di origine delle razze, sono la selezione, la deriva genetica, le migrazioni ecc. In una seconda fase si deve ricostruire il processo storico vero e proprio, individuandone il reale svolgimento, cioè le effettive interazioni tra i processi elementari che lo hanno determinato.
Delle caratteristiche generali dei fattori responsabili della microevoluzione, cioè della genesi delle razze, si è discusso in precedenza (v. anche evoluzione), e si è visto che, pur essendo ben lontani dall'averli compresi a fondo, ne abbiamo tuttavia una visione complessiva abbastanza soddisfacente.
Nella ricostruzione storica della struttura attuale della specie umana siamo invece molto più indietro, anche perché non è affatto chiaro nemmeno il punto di arrivo: il modo in cui suddividiamo in questo momento la nostra specie non è certo l'unico possibile né è particolarmente soddisfacente. Comunque sono stati fatti dei tentativi di costruire un albero evolutivo all'interno della nostra specie, il cosiddetto minimal evolutionary tree di Cavalli-Sforza (v. evoluzione: La moderna teoria dell'evoluzione), utilizzando a questo scopo solo polimorfismi genetici neutri, che, per quanto si è detto sopra, dovrebbero presentarsi con distanze genetiche, tra popolazioni isolate riproduttivamente, proporzionali alla durata dell'isolamento. È probabile che si compiranno grandi progressi in questa direzione nei prossimi anni, perché i polimorfismi nuovi che si possono utilizzare per queste ricerche stanno aumentando rapidamente non solo di numero, ma anche sul piano qualitativo: fino a solo due anni fa sarebbe stato inconcepibile raccogliere dati che non riguardassero i geni strutturali delle proteine - una frazione esigua e non rappresentativa dell'intero genoma - anzi delle proteine eritrocitarie o sieriche, in ogni caso ematiche. È evidente che sta ora per diventare possibile studiare altri materiali biologici e soprattutto regioni di DNA con significati funzionali diversi, per esempio regolativi. E non è affatto escluso che, prendendo in considerazione nuovi geni, si modificheranno alcune delle opinioni attuali, forse radicalmente: questo è già accaduto quando dai caratteri dell'antropologia classica si è passati ai gruppi sanguigni e ai polimorfismi proteici (v. cap. 5, § a, 1).
4. I due tipi di evoluzione, genetica e culturale
a) Analogie e differenze; velocità relative
In nessuna specie la componente culturale dell'evoluzione - e quindi anche della diversificazione intraspecifica - gioca un ruolo tanto importante come nell'uomo. Le due evoluzioni, quella genetica e quella culturale, sono connesse l'una con l'altra in modo indissolubile e inestricabile e si svolgono contemporaneamente, anche se non parallelamente.
L'evoluzione genetica, pur nella sua complessità, è incomparabilmente più semplice di quella culturale: ciascun individuo è connesso con le generazioni precedenti attraverso due soli genitori, anzi attraverso i due soli gameti che ha ricevuto da questi genitori, e con le generazioni successive solo attraverso i gameti che trasmette ai suoi figli; la trasmissione dei caratteri ereditari genetici è solo ‛verticale' ed è inoltre concentrata nel tempo e nello spazio, il che permette, almeno a livello di singoli geni, di arrivare a formulazioni semplici come le leggi di Mendel e di Hardy-Weinberg, cioè di impostare i problemi dell'evoluzione in modo rigorosamente quantitativo. Ma quanti genitori e figli ‛culturali' ha ciascuno di noi? Sul piano culturale ogni persona si trova a far parte di una rete inestricabile di relazioni, che sono, oltre tutto, bidirezionali: la trasmissione dei caratteri culturali è anche ‛orizzontale', come quella delle malattie infettive.
Tuttavia molti dei fattori evolutivi e di diversificazione intraspecifica che operano nell'evoluzione genetica sono chiaramente individuabili, almeno in prima approssimazione, anche nell'evoluzione culturale. Anche nella genesi delle ‛razze culturali' giocano un ruolo preminente, anzi insostituibile, l'isolamento e la variabilità, nonché processi fondamentalmente simili, almeno da un punto di vista formale, alla deriva genetica e alla selezione. Si tratta essenzialmente di sostituire termini come ‛patrimonio e struttura genetica' con ‛insieme di tradizioni e cultura'; ‛alleli' con ‛idee e convinzioni alternative'; ‛gameti', come veicoli di patrimoni genetici, con gli infiniti tipi di veicoli della trasmissione culturale; ‛deriva genetica' con ‛deriva culturale' (il caso ha ampia opportunità di agire anche nella trasmissione dei caratteri culturali); ‛selezione stabilizzante' con ‛conformismo' o con ‛dittatura'; ‛selezione dirompente' con ‛rivoluzione'; ‛mutazioni' con ‛nuove idee'.
Dove veramente l'evoluzione genetica e quella culturale differiscono in modo estremo è nella velocità. La velocità dell'evoluzione genetica, cioè dei cambiamenti nel tempo del patrimonio genetico, ha come unità di tempo la generazione ed è commisurata sulla scala delle ere geologiche, perché si è plasmata come risposta a variazioni ambientali che si verificavano con velocità di questo ordine di grandezza. Pur non potendolo dimostrare, è giusto presumere che, se le ere geologiche si fossero succedute a distanza di secoli e se ciononostante esistessero delle forme di vita sulla Terra, i tempi della loro evoluzione genetica sarebbero stati i secoli invece che i milioni di anni.
La velocità dell'evoluzione culturale è immensamente più grande: rivoluzioni culturali come quella agricola, industriale ed ecologica si sono succedute a distanze ravvicinate, anzi sempre più ravvicinate.
I meccanismi che rendono l'evoluzione culturale tanto più veloce di quella genetica sono fondamentalmente due: l'entità dei fattori in gioco e il fatto che l'evoluzione culturale è, almeno in una certa misura, lamarckiana.
Si è detto prima che sia nell'evoluzione genetica sia in quella culturale operano processi selettivi. Nel caso dell'evoluzione genetica questi si concretano in differenze di idoneità biologica (fitness darwiniana): individui con genotipi diversi hanno in media, per questo motivo, differenti numeri di figli. Nel caso dell'evoluzione culturale, individui con idee diverse hanno, per questo motivo, idoneità culturali diverse. Ma, per quanto grande possa essere il vantaggio selettivo, cioè l'incremento di fitness conferito da un genotipo particolarmente favorevole, esso sarà sempre minuscolo rispetto a quello che può essere associato a certi casi di elevata idoneità culturale: Priamo con le sue cento mogli ha avuto di certo pochissimi figli rispetto alle decine o centinaia di milioni di figli culturali di Gesù Cristo, di Confucio e di Galileo. Fattori selettivi di questa entità non sono nemmeno concepibili a livello biologico e lo stesso, evidentemente, vale per la deriva genetica e, ancora di più, per le conseguenze dell'immigrazione: Pizarro con le sue poche decine di compagni ha distrutto una cultura ed è riuscito a sostituire la religione locale con la propria, ma non ha certo sostituito il patrimonio genetico degli Incas con quello degli Spagnoli.
Una delle cause principali della lentezza dell'evoluzione biologica è il suo procedere in modo darwiniano invece che lamarckiano. Ebbene, anche da questo punto di vista l'evoluzione culturale è molto più rapida: essa può procedere in tutti e due i modi perché non solo possono essere selezionate idee adattative preesistenti, ma diventano ereditabili tutte quelle che si siano formate adattativamente nel corso della vita di un individuo. Infatti, in linea di principio, non c'è nulla che vieti all'evoluzione culturale di generare nuove idee in funzione di determinate situazioni, cioè di ‛mutare' in modo adattativo. Questa differenza fondamentale si può esprimere anche asserendo che, mentre l'evoluzione genetica è soltanto di tipo variazionale (consiste, cioè, in una variazione nel tempo delle frequenze relative di quelle unità - esse stesse immutabili in tutto il corso della loro esistenza - che sono i genotipi degli individui), viceversa l'evoluzione culturale è allo stesso tempo variazionale e trasformazionale, dato che evolvono le unità stesse della popolazione che sta evolvendo. Infatti queste unità, nel corso della loro vita, vanno incontro a trasformazioni, cioè non restano identiche a se stesse, proprio come accade, per esempio, nell'evoluzione delle stelle, ciascuna delle quali si modifica e ha quindi una sua propria evoluzione trasformazionale, che è alla base dell'evoluzione variazionale delle popolazioni di stelle (v. Lewontin, 1979).
b) Interazioni fra i due processi evolutivi, quello biologico e quello culturale
Questi due processi si svolgono interagendo fra loro in continuazione in modi molto complessi e in misura molto elevata. Alcune di queste interazioni sono ovvie e non particolarmente interessanti: certe decisioni culturali, come, ad esempio, quella in base alla quale gli Eschimesi non si trasferiscono nell'Africa equatoriale, hanno una base genetica, nella misura in cui l'adattamento degli Eschimesi al clima freddo è geneticamente determinato.
Molto più interessante è invece esaminare quale può essere stato l'impatto dell'evoluzione culturale sulla preesistente evoluzione biologica e sul suo modo generale di procedere. Si conoscono vari esempi di adattamenti genetici a condizioni ambientali create dall'evoluzione culturale; i polimorfismi ‛malarici' (per gli alleli Th, Gd- e per quello della falcemia, cioè il βS) sono uno di questi, perché la malaria è diventata oloendemica nelle zone climaticamente adatte alla zanzara in seguito all'avvento dell'agricoltura, che ha provocato l'addensamento della popolazione e ha richiesto estesi diboscamenti, entrambe condizioni che, sia pure con meccanismi diversi, favoriscono la diffusione di questa malattia. Lo stesso vale, presumibilmente, per molte malattie infettive diffusesi a causa dell'urbanizzazione, come la tubercolosi e il morbillo, alle quali popolazioni mai esposte in precedenza si sono mostrate particolarmente vulnerabili. Un altro caso interessante - anche se non dimostrato in modo conclusivo come quello della malaria - è la deficienza di lattasi dopo il primo anno di vita all'incirca. Eccetto che nei Caucasici - che hanno adottato l'allevamento del bestiame (per cui il latte è diventato un alimento importante anche oltre il periodo dell'allattamento) varie decine di migliaia di anni fa - questa deficienza enzimatica costituisce quasi la regola dopo la prima infanzia. Ma non è tanto a livello di effetti singoli - per quanto importanti possano essere - che si esercita l'influenza dell'evoluzione culturale su quella biologica: quello che più conta è il suo effetto ‛generale', che si esplica in almeno due modi.
Il primo di questi modi è il ‛carattere di urgenza' che devono assumere tutte le risposte genetiche quando il fattore che le ha provocate è di origine culturale: i ritmi in gioco sono molto più sostenuti di quelli naturali a cui, fino a tempi molto recenti, l'evoluzione genetica ‛si era abituata' a rispondere. Non deve quindi meravigliarci il fatto che, contrariamente alla regola che le risposte adattative agli ambienti naturali sono quanto di più perfetto si possa immaginare, le risposte adattative ad ambienti creati da innovazioni culturali hanno tutte in comune la provvisorietà, l'approssimazione, la precarietà. Sono, in una parola, adattamenti di emergenza a cui non è stato dato il tempo di diventare ottimali. Se torniamo ai polimorfismi ‛malarici' - che sono gli unici compresi abbastanza bene da poter essere discussi - risulta subito evidente che essi costituiscono un esempio tipico di risposta adattativa di emergenza, che ciascuna popolazione esposta bruscamente (cioè nel giro di secoli e forse in certi casi perfino di decenni) a un fattore selettivo intenso come la malaria ha messo a punto per suo conto. Non erano a disposizione né molto tempo né grandi numeri (le popolazioni a quel tempo erano relativamente piccole) per andare a reperire la risposta ideale; sono così venuti fuori i geni Th e Gd- che proteggono solo parzialmente e solo alcuni individui, e che comportano un elevato ‛carico genetico'. Si tratta inoltre di geni che possono essere prodotti facilmente, come tutti i geni cui corrisponde una perdita di funzione, sia pure con certe caratteristiche peculiari che la rendano evolutivamente accettabile. Per quanto poco ottimali questi adattamenti genetici possano essere, resta sempre il fatto che, molto verosimilmente, le popolazioni umane esposte per tempi molto lunghi a un'intensa endemia malarica non sarebbero sopravvissute se essi non fossero stati adottati. È certo comunque che hanno servito allo scopo e che non c'erano molte altre scelte possibili.
Il rilasciamento della selezione biologica è il secondo effetto generale dell'evoluzione culturale su quella genetica. Che esso sia in atto nessuno lo mette in dubbio. Quello che invece si può discutere è quanto grande tale rilasciamento possa essere stato e, soprattutto, potrà essere, problema che si collega con altre questioni ancora, cioè per quanto tempo esso abbia già avuto l'opportunità di agire e su che frazione dell'intero patrimonio genetico. Non siamo in grado di dare risposte precise a queste domande, tuttavia è certo che si deve scartare l'opinione superficiale secondo cui solo in quest'ultimo secolo, con i grandi progressi della medicina, la selezione biologica ha cominciato a diventare meno stringente: già tra gli Spartani, pur con il loro monte Taigeto, essa era incomparabilmente più blanda di quanto può esserlo anche adesso in uno dei pochi popoli di cacciatori e raccoglitori tuttora esistenti. Un grandissimo numero di imperfezioni fisiche con base almeno parzialmente genetica, che da tempi molto lunghi, in certe culture, hanno avuto un peso trascurabile o comunque non tale da ridurre l'idoneità dei loro portatori (ernie, daltonismo, ecc.), hanno costituito di sicuro degli svantaggi biologici considerevoli in condizioni di vita primitive. Dal momento che questi caratteri genetici lievemente sfavorevoli hanno cessato di essere controselezionati da molto tempo in gran parte della specie umana, mentre ciò si è verificato solo di recente in certe popolazioni e in alcune ancora non si è verificato affatto, è ragionevole supporre - ed è stato infatti ipotizzato - che questi caratteri siano attualmente più diffusi nelle popolazioni di più antica civiltà sedentaria e che sempre di più tenderanno a diffondersi nella nostra specie.
Nel complesso si può concludere non solo che in questo momento l'evoluzione culturale sta procedendo nella nostra specie a una velocità molto superiore a quella dell'evoluzione biologica (tanto che ormai il destino dell'uomo dipende soprattutto dalla sua evoluzione culturale per tempi lunghi e solo da essa per tempi brevi), ma che è l'ambiente creato dalla cultura l'unico ancora in grado di mettere a dura prova la nostra capacità di adattamento genetico. L'evoluzione culturale modifica l'ambiente, anche quello fisico, a una velocità cui l'evoluzione genetica non può tener dietro, ed è quindi compito dell'evoluzione culturale stessa - e solo di questa - trovare, e in modo lamarckiano, le risposte adattative.
Considerato l'impatto che una cultura avanzata può esercitare sull'andamento generale dell'evoluzione biologica, non è escluso che le strutture genetiche di popolazioni primitive possano avere dei caratteri differenziali comuni e diversi nel loro insieme da quelli delle popolazioni che per tempi più lunghi sono vissute in ambienti ‛meno naturali'. Queste ultime, presumibilmente, hanno più geni sfavorevoli, non solo del tipo di quelli dovuti al rilasciamento della selezione, ma anche di quelli accumulatisi come risposta di emergenza a variazioni ambientali di origine culturale, come i geni che conferiscono resistenza ad agenti patogeni, ma a un certo prezzo, rappresentato dal ‛carico genetico' a essi associato. È evidente che solo i geni del primo tipo possono essere considerati sfavorevoli in senso assoluto, mentre gli altri lo sono solo se mancano gli agenti patogeni e le condizioni favorevoli allo sviluppo delle malattie da essi provocate, mentre quando queste malattie sono diffuse risultano utili, se non addirittura indispensabili alla sopravvivenza della popolazione: basti pensare alle decimazioni da morbillo e tubercolosi che si sono verificate in popolazioni primitive quando sono venute a contatto con popolazioni europee.
5. Il concetto di razza e i suoi limiti
Una specie può essere suddivisa in razze sulla base di due tipi di definizioni. Il primo comprende le definizioni che potremmo chiamare ‛operative' (una delle quali è stata adottata e brevemente illustrata nel cap. 1), perché contengono, in modo più o meno esplicito, i criteri che si devono seguire e i risultati che si devono ottenere per poter affermare di avere individuato delle razze distinte.
Il secondo tipo di definizioni si riferisce, più che ai procedimenti da seguire per identificare, caratterizzare e descrivere le razze, al loro significato tassonomico ed evolutivo. Le razze potrebbero per esempio essere considerate sottospecie o prespecie, cioè come tappe intermedie del lungo processo che dà luogo alla comparsa di specie diverse, tutte derivate dalla stessa; indubbiamente c'è molto di vero in questa concezione: se le specie fossero consistite di un singolo insieme di individui senza alcuna barriera riproduttiva al loro interno, avrebbero potuto evolvere (diventare cioè diverse da come erano prima), ma non si sarebbe potuta verificare la formazione di due o più specie da una sola, fenomeno che invece è certamente esistito, esiste tuttora ed è di sicuro uno dei più importanti dell'evoluzione filogenetica.
Entrambi questi concetti di razza sono facilmente criticabili da molteplici punti di vista, a tal punto che al termine ‛razza' non si può attribuire che una validità scientifica molto relativa da qualunque angolatura esso venga considerato.
Non si vuole suggerire che entità di questo genere non esistano, né che termini come ‛razza', ‛gruppi etnici', ‛demi', ‛popolazioni' - che si riferiscano cioè alla variabilità obiettivamente constatabile all'interno della maggior parte delle specie - vadano abbandonati. Sono talmente utili che non se ne potrebbe fare a meno (come ha detto Dobzhansky ‟se le razze non esistessero, occorrerebbe inventarle"). Si vuole invece proporre che, nel continuare a usarli, non ci si dimentichi mai di avere a che fare con concetti ed entità vaghi e validi entro limiti molto ristretti, che devono scoraggiare qualsiasi tipo di estrapolazione che non sia ampiamente giustificata.
a) Limiti e difetti delle definizioni operative di razza
Molti di questi limiti - ma i meno seri - risultano ovvi dalle definizioni stesse, come quella fornita nel cap. 1. È evidente infatti che un gruppo di individui, per essere considerato una razza, deve essere ‛sufficientemente numeroso' e deve essere distinguibile non per uno, ma per ‛un insieme' di caratteri ereditari. Ma cosa si debba intendere di preciso per ‛sufficientemente numeroso' e per ‛un insieme' di caratteri ereditari lo si può decidere solo in modo arbitrario, e altrettanto arbitrario è il grado di diversità che due gruppi di individui devono presentare, rispetto ai caratteri prescelti, per poter essere considerati due razze diverse. Soggettivi e sfumati, come i criteri con i quali attribuiamo a un corso d'acqua il nome di ruscello o di torrente o di fiume (per rifarsi al paragone del celebre genetista Curt Stern), sono anche i confini tra i concetti di razze minori o sottorazze e razze principali, come Caucasici, Negri e Mongolici.
Ma se al concetto di razza si potessero rivolgere solo critiche di questo genere la sua validità non ne verrebbe a soffrire granché: il concetto di fiume non è invalidato dal fatto che la sua distinzione da quello di torrente è, in una certa misura, arbitraria. E questo perché, al di fuori di questo aspetto quantitativo impreciso, il concetto di fiume è perfettamente chiaro.
Per il significato del termine razza, invece, quel grado di vaghezza, arbitrarietà e imprecisione che disturba a prima vista chi ne legge la definizione non è che la punta di un iceberg: ben altri e più gravi motivi di perplessità appaiono a chi si preoccupi di prendere in esame il significato di fondo del termine. Per l'arbitrarietà si potrebbe rimediare in modo relativamente facile fissando valori ragionevoli che creino distinzioni precise pur nell'ambito di un continuum; non sarebbe poi tanto grave se tutto si riducesse al fatto che alcuni chiamano razze minori gli stessi gruppi che altri chiamano razze maggiori o principali. L'avere a che fare con grandezze continue invece che discontinue rende solo più complicato - ma non insolubile - un problema. Non è invece altrettanto facile trovare soluzioni soddisfacenti quando il problema è ‛intrinsecamente' complesso o addirittura insolubile per ragioni di principio. E questa è appunto la situazione in cui ci si trova quando si cerchi di suddividere una specie, soprattutto quella umana, in razze, quando si cerchi cioè di mettere in pratica una loro definizione operativa.
1. Critica di fondo a qualsiasi definizione operativa di razza. - Il vero limite del concetto di razze come gruppi di individui con caratteristiche ereditarie diverse è la pressoché completa mancanza di correlazione tra i vari tipi di classificazioni possibili. Ogni grande criterio di suddivisione (antropologico fisico, genetico e culturale) genera una classificazione a sé stante. Non solo, ma non danno risultati concordanti, spesso nemmeno parzialmente, neppure i sottocriteri di un criterio principale: sono possibili numerose classificazioni puramente antropologiche del tutto diverse, e numerose suddivisioni genetiche della nostra specie nettamente contrastanti fra loro oltre che con quelle suggerite dall'antropologia; per non parlare delle difficoltà che si incontrano nell'antropologia culturale, che sono di gran lunga maggiori per motivi troppo ovvi perché valga la pena di discuterne.
Quando è cominciata lentamente a sorgere l'antropologia genetica, in questi ultimi cinquant'anni, molti hanno creduto ottimisticamente che essa, con la sua maggiore rigorosità quantitativa e con la perfetta conoscenza delle basi genetiche formali dei caratteri presi in esame, avrebbe perfezionato e corretto il quadro tracciato in precedenza dall'antropologia fisica. Si credeva in sostanza di guardare le stesse cose, sia pure con uno strumento più rigoroso, più obiettivo e più sensibile. In particolare ci si aspettava che razze giudicate molto distanti sul piano antropologico fisico sarebbero risultate molto distanti anche adottando criteri genetici: se si sostituisce un termometro rozzo con uno migliore ci si aspetta di trovare una buona correlazione tra le due serie di misure e si può allora procedere a correggere i risultati palesemente errati del primo termometro. Ma se il secondo strumento, invece di misurare con maggiore precisione la stessa cosa, ne misura un'altra fondamentalmente diversa - se, per restare nello stesso paragone, e un barometro invece di un termometro - allora si troveranno risultati del tutto incompatibili. Ebbene, quando si sono confrontati i risultati dell'antropologia fisica con quelli della genetica di popolazione ci si é trovati di fronte a discrepanze gravissime, almeno sul piano quantitativo, tali da dare l'impressione che i caratteri presi in considerazione con i due criteri fossero ‛intrinsecamente' diversi. Popolazioni indiscutibilmente molto distanti da un punto di vista antropologico, che appartengono cioè a ‛razze principali' differenti, sono separate da distanze genetiche solo di poco superiori a quelle intercorrenti tra popolazioni antropologicamente affini; inoltre, in media, il numero di geni per i quali un francese ha un genotipo diverso da quello di un giapponese è maggiore, ma solo di poco, del numero di geni per i quali differiscono fra loro un francese da un altro francese o un giapponese da un altro giapponese. E tutto ciò è la regola, non l'eccezione.
Questo notevole grado di indipendenza tra i diversi criteri o sottocriteri di suddivisione può essere mostrato facilmente in vari modi, per esempio confrontando carte di una stessa area geografica, come un continente, suddivise con criteri diversi: glottologico, religioso, politico, antropologico fisico, genetico e così via. Se si sovrappongono l'una all'altra mappe di questo genere disegnate su materiale trasparente, in modo da poterle confrontare con un unico colpo d'occhio, si constata un disaccordo quasi assoluto tra di esse. Un'altra osservazione pertinente è che lo spettro di variazione di molti caratteri antropologici è quasi altrettanto ampio all'interno di una razza di quanto lo è nell'intera specie: per esempio, l'intervallo tra la statura media della razza africana più bassa (i Pigmei) e quella della più alta (i Nilotici) è praticamente uguale a quello dell'intera nostra specie. Lo stesso vale per l'indice cefalico e per altri caratteri ancora.
Gli unici caratteri che mostrano un certo grado di coerenza, che tendono cioè a variare in modo correlato, sono quelli sorti come risposte adattative a un fattore ecologico comune molto stringente, come un clima freddo particolarmente rigido.
Una delle discrepanze più appariscenti tra criteri di classificazione diversi è quella che esiste tra la classificazione antropologica e quella genetica. È vero che i minimal evolutionary trees (cioè i dendrogrammi; v. evoluzione: La moderna teoria dell'evoluzione) costruiti sulla base di quest'ultima per render conto dell'attuale struttura della nostra specie sono, in sostanza, in accordo con quelli dell'antropologia, ma le distanze antropologiche sono talmente più grandi di quelle genetiche che questi alberi evolutivi sembrano costruiti addirittura su scale diverse.
Non c'è dubbio, infatti, che l'affermazione che Caucasici, Africani e Mongolici sono molto diversi sotto certi aspetti non è frutto della fantasia degli antropologi, ma è semplicemente un dato di fatto; a tal punto che ogni singolo individuo può, nella grandissima maggioranza dei casi e anche sulla base di singoli caratteri antropologici, essere assegnato correttamente e senza difficoltà a uno di questi tre gruppi. È altrettanto vero però che la distanza genetica tra questi gruppi - stimata su un gran numero di marcatori - è relativamente modesta e certo non tale da permettere di classificare ogni singolo individuo, tanto meno utilizzando singoli marcatori genetici.
Presumibilmente due sono i motivi principali che hanno reso tanto grandi le distanze antropologiche: i caratteri antropologici sono caratteri ‛esterni' e sono stati scelti proprio perché distribuiti in modo molto eterogeneo.
Cavalli-Sforza e Bodmer hanno suggerito che il fatto che l'antropologia abbia scelto i suoi marcatori tra i caratteri morfologici esterni, a differenza dei marcatori sierologici e biochimici della genetica di popolazione, può avere contribuito ad aumentare le distanze antropologiche rispetto a quelle trovate con approcci genetici. Secondo questi autori, i caratteri esterni sono esposti agli occhi dell'antropologo nella stessa maniera in cui lo sono ‛agli occhi' della selezione: è molto più ragionevole supporre che risenta in modo adattativo delle condizioni climatiche un carattere cutaneo, come il colore della pelle, oppure scheletrico, come la conformazione corporea, piuttosto che un carattere interno come un antigene di gruppo sanguigno. In altre parole, gli antropologi avrebbero scelto i loro marcatori non tra caratteri qualsiasi, ma tra caratteri che, per gli stessi motivi che li rendevano particolarmente accessibili alla loro osservazione, erano anche particolarmente esposti all'ambiente esterno, cioè alla selezione. Questa, agendo in modo diverso su individui esposti ad ambienti diversi, avrebbe quindi causato la loro diversificazione razziale in senso adattativo. In questo modo l'antropologia ha misurato le distanze ‛ambientali' (più precisamente le distanze tra le conseguenze genetiche delle diversità di ambiente) più che le distanze genetiche ‛medie'. Ogni insediamento umano, e specialmente quelli in ecotipi estremi, ha dovuto mettersi molto in fretta l'abito adatto al suo ambiente, pena l'estinzione, e una risposta rapida e adeguata non si sarebbe potuta realizzare altro che con processi selettivi, certo non per azione del puro caso su una variabilità genetica a sua volta prodotta a caso. La selezione ha agito per i caratteri connessi con l'adattamento ad ambienti estremi (caldo, freddo, malaria, altitudine ecc.) con efficienza e rapidità paragonabili a quelle messe in atto dalla selezione artificiale nella genesi delle razze degli animali domestici, per i quali solo pochi millenni sono stati sufficienti a creare razze che differiscono, per i caratteri selezionati - e presumibilmente solo, o soprattutto, per quelli - in misura cospicua e appariscente, quale si osserva di regola solo tra gruppi sistematici molto lontani (basti pensare alle varietà di forme oggi esistenti all'interno di specie come quella del cane o del bue).
I genetisti invece hanno lavorato su caratteri interni e perciò relativamente protetti dall'azione dell'ambiente e meno suscettibili di evolvere in funzione di esso.
In sostanza, le divergenze evolutive riguardanti caratteri genetici come i polimorfismi sierologici e biochimici sono da attribuirsi quasi esclusivamente alla deriva genetica, mentre quelle dei caratteri antropologici sono state causate non solo dalla deriva genetica, ma anche - anzi soprattutto - dalla selezione, che è un fattore evolutivo molto più rapido ed efficiente. Se la microevoluzione nell'ambito della specie umana potesse ricominciare daccapo, è presumibile che si arriverebbe di nuovo a una sua diversificazione antropologica simile, almeno nelle grandi linee, a quella esistente (non è certo per caso che gli Eschimesi hanno caratteristiche antropologiche che li rendono resistenti al freddo), mentre la distribuzione degli alleli neutri o quasi neutri dell'antropologia genetica risulterebbe alla fine del tutto diversa da quella che conosciamo; infatti essa non è altro che una delle numerosissime distribuzioni possibili, pressoché equivalenti dal punto di vista evolutivo.
Gli antropologi hanno scelto i caratteri su cui concentrare la loro attenzione non solo tra quelli morfologici esterni, ma in più, nell'ambito di questo gruppo di caratteri preferenzialmente adattativi, hanno privilegiato quelli che meglio si prestavano a distinguere una razza dall'altra (cioè quelli con grande eterogeneità intergruppi e il più possibile omogenei all'interno dei gruppi), considerando quindi i caratteri poco distintivi come poco adatti ai loro scopi e quelli che si presentavano identici in tutti i gruppi umani come affatto inadatti per definizione. E, per una scienza che ha per scopo principale, o perlomeno come punto di partenza obbligato, la suddivisione della specie umana in gruppi, è proprio quello che si doveva fare e quindi non c'è nulla da eccepire: non sarebbe certo il caso di proporre come carattere antropologico il numero di occhi o di nasi per individuo. Tuttavia il fatto che questo sia stato un procedimento perfettamente giustificato e adeguato per quel determinato scopo non vuol dire che l'insieme dei caratteri studiati dagli antropologi sia rappresentativo dei caratteri umani; significa anzi il contrario: siccome un carattere si considera antropologicamente utile (o, più in breve, antropologico) solo se è ‛particolarmente diversificato' tra i vari gruppi umani, non è certo da un insieme di caratteri scelti con un criterio di questo genere, cioè dai caratteri antropologici, che si può valutare il grado ‛medio' di variabilità esistente tra le razze umane.
La maggior parte dei polimorfismi genetici, invece, e in particolare quelli biochimici, sono stati scoperti esaminando geni strutturali scelti con criteri puramente tecnici (cioè solo perché si disponeva di tecniche adatte a studiarli dal punto di vista della genetica di popolazione), quindi da un punto di vista che sostanzialmente prescindeva dal fatto che essi presentassero o meno una variabilità intra- o interpopolazioni: solo dopo averli studiati veniva accertato quali di questi geni fossero variabili e quali no, e il grado e il tipo della loro eventuale variabilità. È evidente allora che, almeno in prima approssimazione, il campione prescelto dovrebbe essere rappresentativo dei caratteri umani e che la variabilità interrazziale (o distanza genetica) trovata al suo interno dovrebbe costituire una stima attendibile del grado ‛medio' di diversificazione genetica dell'uomo, anche se valida solo per quella parte del genoma che è costituita da geni strutturali. Secondo questa stima, la variabilità genetica della nostra specie è grandissima, ma solo una ‛piccola' parte di essa è una variabilità tra gruppi; la maggior parte consiste in variabilità individuale intragruppi, cioè in polimorfismi genetici.
Da altre discipline, infine, come la biochimica e la farmacologia, sembrerebbe di poter ricavare un quadro della variabilità umana ancora diverso, che si può - o almeno si poteva fino a poco tempo fa - riassumere, sia pure semplicisticamente, dicendo che tale variabilità è poco più che trascurabile.
Questi tre quadri molto contrastanti della variabilità della nostra specie sono elencati qui di seguito in modo schematico.

Il quadro più rispondente alla realtà è certamente quello suggerito dalla genetica, perché esso è derivato da osservazioni programmate ed effettuate proprio con lo scopo di stimare la variabilità genetica e quindi con un approccio, almeno apparentemente, esente da vizi sistematici.
Gli altri due modi di valutare la variabilità genetica (nel suo complesso e nelle sue due componenti) sono invece dei sottoprodotti di ricerche che, dirette ad altri scopi, non erano per principio interessate a evitare errori sistematici che la riguardassero. Ne è risultato che l'antropologia, che ha studiato solo caratteri con eterogeneità intergruppi molto maggiore di quella intragruppi, è arrivata a sovrastimare la prima e a sottostimare la seconda, e che la biochimica e la farmacologia, per le quali la variabilità genetica di qualunque tipo non era altro che un'indesiderabile complicazione, hanno avuto la tendenza a minimizzarla.
Sembra quindi ragionevole concludere che la nostra specie è costituita da un insieme di gruppi molto diversi fra loro per i caratteri esterni, cioè antropologici, e molto simili per quelli interni, cioè sierologici e biochimici; e che, viceversa, gli individui di ogni gruppo sono molto simili fra loro per i caratteri esterni (tutti con lo stesso ‛abito antropologico' frutto della stessa ‛storia ambientale') e molto eterogenei quelli interni. In altre parole, le razze non sono affatto così diverse come ci suggeriscono gli abiti che indossano. Questo, fra l'altro, ci mostra che non c'è nulla da stupirsi per non aver mai trovato una riduzione di fertilità negli incroci interrazziali, nemmeno in quelli tra le razze più lontane: è chiaro che la suddivisione dell'uomo è molto meno profonda, dal punto di vista genetico, di quella che si osserva in molte altre specie (per esempio, di Ditteri) e che viene a buon diritto considerata precorritrice della scissione in specie diverse.
2. Ciò che rimane valido del concetto di razze definite con un criterio operativo. - La mancanza di correlazione tra le classificazioni suggerite dai vari criteri, nonché tra le distanze valutabili con essi, porta necessariamente alla conclusione che non è mai lecito estrapolare; nemmeno all'interno di un criterio principale, come quello antropologico, o genetico o culturale.
Poiché le razze si sono differenziate con meccanismi adattativi e per azione del caso, e solo i primi hanno una grossa componente deterministica, l'unica affermazione generale ragionevole è che razze simili per ‛storia ambientale' tendono a essere simili anche geneticamente. Ma anche qui si individuano subito due limiti: le razze tendono a essere simili solo per quei caratteri che hanno un significato adattativo verso le caratteristiche ambientali comuni, e inoltre, nell'ambito di questi caratteri, la somiglianza riguarda di regola il loro fenotipo più che il genotipo (come nel caso delle razze ‛malariche', nelle quali sono diffusi fenotipi del tipo della talassemia e della enzimopenia per la G6PD, dovuti però a genotipi diversi nei diversi gruppi).
b) Critiche al concetto di razza come prespecie
Non c'è dubbio che tutte le specie sono state per un certo periodo delle razze che possono essere chiamate, con pieno diritto, prespecie. In moltre, molte delle razze attuali sono destinate a diventare specie e sono quindi anch'esse delle prespecie. È questo l'aspetto delicato della definizione di prespecie: essa è operativa solo in teoria, dato che potrebbe essere applicata solo dopo aver constatato il verificarsi di un evento futuro e incerto. Che una razza, dopo essersi isolata da tutte le altre, diventi una specie, è una possibilità, oltre tutto nemmeno probabile. Non è certo una legge biologica, come lo è per un embrione svilupparsi e diventare un individuo maturo. L'embrione può quindi a buon diritto essere considerato un preindividuo, mentre non sarebbe altrettanto giusto considerare il termine razza come equivalente al termine prespecie. È proprio qui che la diversificazione tra razze, in quanto non programmabile, differisce dalla differenziazione.
È chiaro quindi che il termine prespecie, se riferito a una determinata razza, può essere usato solo a posteriori, cioè dopo avere constatato che essa come tale (senza essersi fusa con altre razze, diventando quindi una entità diversa) è diventata una specie; se lo si riferisce invece a razze attuali, allora va usato in senso generico, dicendo che alcune di esse - ma non sappiamo quali - sono delle prespecie.
Il concetto di prespecie ha, come si vede, un valore molto teorico, che non può essere di molto aiuto nel suddividere una specie in sottogruppi, salvo che nei casi in cui si sia effettivamente trovata una forte barriera riproduttiva tra i gruppi in esame. Per la specie umana, in cui ciò risulta non essersi verificato, almeno a livello biologico, il termine prespecie sarebbe del tutto ingiustificato per ciascuna delle sue razze, tanto più che, come effetto della sua evoluzione culturale più recente, le già piccole distanze genetiche oggi esistenti al suo interno sono destinate a ridursi e probabilmente a scomparire in tempi relativamente brevi.
Resta comunque il fatto che, sul piano tassonomico, esistono differenze qualitative e non semplicemente quantitative tra razza e specie: al livello di razze, la diversificazione è per definizione ancora reversibile, attraverso il loro eventuale mescolamento, mentre quando ha varcato la soglia della specie è diventata, sempre per definizione, irreversibile (trascurando i possibili fenomeni di introgressione). Ne segue che la specie umana è monofiletica. Infatti è da considerare del tutto inverosimile che due bonae species nel senso tassonomico della parola (cioè incapaci di dare origine a ibridi fertili), evolvendo indipendentemente l'una dall'altra, convergano geneticamente a un punto tale da diventare capaci di incrociarsi e generare ibridi fertili, cioè, per dirla con altre parole, si fondano in una specie sola. La diversificazione della specie umana è quindi tutta da attribuirsi a fenomeni di divergenza evolutiva iniziatisi nell'unico gruppo di individui che è diventato Homo sapiens.
6. Le modalità di diversificazione della nostra specie: il ruolo dei vari fattori evolutivi
Si è visto prima che i fattori responsabili della diversificazione all'interno della nostra specie sono (una volta soddisfatta la conditio sine qua non dell'isolamento riproduttivo tra gruppi inizialmente simili, che potremmo chiamare prerazze) la selezione, la deriva genetica e il mescolamento con altri gruppi. È evidente che, pur disponendo di conoscenze abbastanza approfondite su ciascuno di essi, non siamo in grado di inferire la storia biologica della nostra specie se non in modo approssimativo. Tuttavia è possibile tracciare nelle loro linee generali i processi attraverso i quali è presumibile si siano originate le razze umane.
All'inizio deve essere esistito un unico gruppo di individui progenitori della nostra specie. Da esso, in tempi successivi, si devono essere staccati vari gruppi; alcuni hanno evidentemente avuto il tempo e l'opportunità di diversificarsi fino a dare origine alle razze umane, quali le vediamo oggi. Senza dubbio ciascuno di questi processi di diversificazione avrà avuto la sua storia particolare; tuttavia, è ragionevole supporre che tutti abbiano seguito un iter e obbedito a regole entro certi limiti abbastanza simili. Per esempio, per quanto si è detto prima, è presumibile che se due gruppi, dopo essersi separati riproduttivamente, hanno dato origine a due razze diverse, questo si sia verificato per opera di una selezione diversificante e solo in misura molto minore per effetto della deriva genetica. E questo, a sua volta, implica che la condizione necessaria affinché due gruppi inizialmente uguali si siano diversificati fino a diventare due razze distinte deve essere stata che, per effetto della loro separazione, le loro evoluzioni indipendenti si siano svolte in ambienti diversi, siano state cioè guidate, almeno per quei caratteri che ora li distinguono l'uno dall'altro, da fattori selettivi diversi. In altre parole, è lecito supporre che la causa vera della formazione di nuove razze sia stata la differenza tra gli ambienti che sono stati via via occupati dalla specie umana, mediata dalle spinte selettive differenziali che questi ambienti hanno evocato, e quindi che la deriva genetica abbia di regola giocato un ruolo secondario.
Se si accettano queste premesse, si può in prima approssimazione accettare anche l'ipotesi che di regola le razze abbiano subito il seguente processo di formazione. Due o più gruppi derivanti da uno stesso gruppo, e quindi inizialmente uguali (trascurando i possibili ‛effetti del fondatore'), si separano riproduttivamente andando a occupare habitat diversi ed esponendosi quindi ad ambienti diversi. Da questo momento essi iniziano a evolvere indipendentemente sotto l'effetto della selezione, della deriva genetica ed eventualmente del mescolamento con altri gruppi diversi. Questa evoluzione indipendente porterà alla loro diversificazione per tutti i caratteri genetici che presentino un certo grado di variabilità. Questa diversificazione, però, non procederà affatto nello stesso modo per tutti i caratteri. Al contrario, quei pochi caratteri che hanno a che fare con l'adattamento differenziale dei vari gruppi agli ambienti in cui si trovano a vivere sono subito sottoposti alla selezione, che, in tempi relativamente brevi (anche di solo poche migliaia di anni) e in modo più o meno stringente, farà sì che ogni gruppo acquisti una struttura genetica compatibile con il suo ambiente e diversa da quella degli altri gruppi. Durante questo breve periodo esisterà, all'interno di ogni gruppo che sta mettendo a punto il suo adattamento genetico, una cospicua variabilità, prevalentemente sotto forma di polimorfismi transeunti, ma anche sotto forma di polimorfismi destinati a diventare bilanciati che si stanno avvicinando alle frequenze di equilibrio. Una volta conseguito l'adattamento, rimarrà, di questa grande variabilità, solo la quota associata a questi ultimi polimorfismi. Naturalmente, nel tempo in cui si verificano questi eventi agirà anche la deriva genetica, ma in maniera stocastica e quindi molto più lentamente. In compenso però essa opererà su un numero molto maggiore di geni, cioè su tutti quelli che non sono coinvolti nell'adattamento differenziale. Immaginiamo adesso, per semplificare (forse in modo esagerato) le cose, che, trascorsa questa prima rapida fase di adattamento genetico che ha creato una grande diversificazione razziale, ma solo per i caratteri adattativi differenziali, le strutture genetiche dei vari gruppi non siano ulteriormente perfezionabili rispetto agli ambienti in cui vivono. È evidente che da quel momento in poi la selezione tenderebbe a mantenere lo status quo, a mantenere cioè le differenze razziali adattative comportandosi all'interno di ogni gruppo da agente stabilizzante e uniformante in proporzione all'entità del vantaggio che offre al gruppo quella particolare risposta adattativa. La deriva genetica invece continuerà indisturbata ad accumulare a caso differenze intergruppi per tutti i geni, compresi quelli adattativi, con l'unica limitazione, per quanto riguarda questi ultimi, che le sostituzioni alleliche casuali non devono comportare una riduzione dell'adattamento genetico del gruppo (è verosimile, per esempio, che in una popolazione esposta alla malaria, nella quale la talassemia abbia raggiunto una certa frequenza ottimale, sia permesso alla deriva di sostituire più o meno completamente un allele talassemico con un altro; mentre è molto meno verosimile che essa possa far variare la frequenza globale della talassemia).
Nel complesso, quindi, è presumibile che l'attuale diversificazione razziale sia il risultato, per i caratteri adattativi differenziali, di un processo bifasico: un primo periodo breve, per così dire ‛di emergenza', durante il quale hanno fatto in tempo a diversificarsi - e in maniera cospicua - solo i geni implicati nell'adattamento differenziale; e una seconda fase, di durata indefinita, in cui la selezione differenziale agisce non più da fattore dirompente bensì da fattore stabilizzante (al livello fenotipico rilevante per l'adattamento), mentre la deriva genetica continua il suo lavoro di diversificazione lento e regolare. ‛Regolare', naturalmente, se riferito al suo effetto medio su tutti i geni, perché, trattandosi di un processo stocastico, il suo effetto su ogni singolo gene è invece per definizione del tutto casuale (è proprio sull'assunto che la diversificazione media prodotta dalla deriva genetica aumenti con velocità costante che si basano i tentativi di ricostruzioni storiche della struttura attuale della nostra specie; v. cap. 3, § c). Questo processo è rappresentato in forma di diagrammi nella fig. 3.
Vale la pena di sottolineare un aspetto della variabilità intergruppi, cioè tra razze, che potrebbe non apparire chiaro dagli schemi della fig. 3. Sebbene la fig. 3B mostri che questa variabilità a lungo andare diventa molto elevata anche per geni non adattativi, si deve egualmente concludere che le uniche differenze interrazziali che contano sono quelle generate dalle diversità ambientali. Infatti la variabilità rilevante è quella ‛media' per gene (v. fig. 3A; v. tabella a p. 1043); quella globale è grande solo perché i geni sono molti: il dire che la variabilità genetica intergruppi è grande perché lo è la variabilità ‛globale' sarebbe come dire che due persone con capigliature molto simili hanno capelli molto diversi perché la somma delle differenze riscontrate confrontando capello per capello è elevata. E questo vale a maggior ragione nel caso della rilevanza biologica di queste differenze, in quanto quelle accumulate per opera della deriva genetica riguardano per definizione caratteri neutri o quasi neutri.
Il quadro appena illustrato è quello a cui si è giunti con l'analisi antropologica e con quella genetica, approfondita quel tanto che era permesso da tecniche semplici e di routine, quali gli esami ematologici standard (determinazioni emocromocitometriche ed esame dello striscio di sangue) e le analisi elettroforetiche. Esaminando a questo livello i caratteri adattativi come quelli ‛malarici' (per esempio la talassemia) si arriva alla conclusione che l'agente selettivo, una volta raggiunto il suo effetto, impedisce ulteriori variazioni che comporterebbero una diminuzione della idoneità biologica globale della popolazione.
L'analisi fine delle basi molecolari di questi caratteri adattativi, che per ora è stata possibile solo per i caratteri ‛malarici' (v. cap. 3, § b, 1), ha però mostrato che di regola l'uniformità del carattere adattativo è tale solo nella misura in cui esso determina il vantaggio selettivo corrispondente. Al di là di questo vincolo, cioè purché questa condizione sia soddisfatta, esiste un considerevole grado di variabilità all'interno del carattere adattativo: in una parola, al fenotipo richiesto e mantenuto dalla selezione corrispondono non uno ma vari genotipi. E questa variabilità la si trova tanto ‛all'interno' di popolazioni adattate alla terzana maligna che - anzi ancora di più - ‛tra' popolazioni adattate a questo agente selettivo.
Dato che solo i polimorfismi ‛malarici' sono stati studiati a livello molecolare, a rigore l'affermazione che i caratteri adattativi sono solo apparentemente omogenei dovrebbe essere limitata a questi polimorfismi. Ma, visto che l'unica serie di caratteri adattativi finora studiata ha mostrato questa eterogeneità nascosta, è quanto meno ragionevole supporre di non avere scoperto un fenomeno raro. In altre parole, appare verosimile che l'alto grado di somiglianza antropologica riscontrabile per caratteri adattativi come il colore della pelle tra popolazioni esposte a un ambiente simile, ma certamente rimaste a lungo separate - nasconda in effetti un grado elevato di eterogeneità rilevabile solo spingendo l'analisi di questi caratteri a un livello paragonabile a quello cui è giunto lo studio delle talassemie, delle emoglobinosi e dell'enzimopenia per la G6PD. Quando questo verrà fatto - e non c'è motivo di dubitare che lo si farà - sarà possibile misurare l'effetto della deriva genetica sui caratteri adattativi, cioè dal momento in cui i corrispondenti fattori di selezione hanno cominciato ad agire, e si potrà tentare di distinguere le somiglianze antropologiche e genetiche dovute a convergenza evolutiva da quelle dovute a non divergenza (v. cap. 3, § b, 1).
7. Diversità genetica e uguaglianza umana
In un famoso libro, intitolato appunto Diversità genetica e uguaglianza umana, Dobzhansky ha trattato questo argomento in modo magistrale non solo dal punto di vista scientifico, ma soprattutto per l'altissimo grado di equilibrio, imparzialità e obiettività. Si tratta di un titolo particolarmente felice, che esprime in forma stringatissima la vera essenza del problema.
a) Scienza e decisioni etico-politiche
Il libro di Dobzhansky ha tuttavia dato origine a molti equivoci, il primo dei quali è di carattere molto generale, perché concerne la funzione e la responsabilità della scienza non solo riguardo alla diversità umana e alle sue eventuali implicazioni (e già non sarebbe poco), ma addirittura riguardo a tutte le grandi decisioni etiche dell'umanità. Esso consiste nel credere - o nel comportarsi come se si credesse - che alla scienza tocchi l'onere di dimostrare la validità etica di determinati comportamenti invece che semplicemente quello di fornire dati esatti e pertinenti. In questo contesto quindi, visto che è ovvio per tutti che non è possibile dimostrare scientificamente questioni riguardanti la dignità e i diritti umani, si richiede alla scienza di fornire dati che costituiscano una giustificazione razionale di decisioni etiche prese a priori e comunque indipendentemente da essa. Così, da parte delle ideologie razziste si è preteso che la scienza producesse su ordinazione le prove che esistono razze superiori e razze inferiori; più tardi, inorriditi dalle conseguenze concrete di queste ideologie, ci si è rivolti di nuovo alla scienza pretendendo questa volta che essa fornisse - di nuovo su ordinazione - le prove dell'uguaglianza umana o quanto meno minimizzasse il valore biologico delle diversità che innegabilmente esistono. Il ruolo della scienza è invece completamente diverso e consiste nell'analizzare nel modo più imparziale e distaccato possibile la realtà obiettiva, cioè, in questo contesto, nel descrivere senza riserve mentali, pregiudizi e reticenze le caratteristiche della grandissima diversità esistente all'interno della nostra come di tutte le altre specie.
È invece dovere etico dell'uomo quello di accettare, e soprattutto attuare, il principio dell'uguaglianza umana svincolandolo completamente dai risultati della scienza, che anzi, se investita da una tale responsabilità morale - che non le compete - e guidata da una forte carica emotiva, diventerebbe fatalmente una pseudoscienza. Il minimizzare la diversità genetica è cadere in un tranello razzistico, perché equivale ad ammettere implicitamente che una tale diversità giustificherebbe una discriminazione sociale. Così si esprime a questo proposito la dichiarazione sulla razza dell'UNESCO (1952): ‟Parità di opportunità e uguaglianza di fronte alla legge non dipendono in alcun modo, come principi etici, dall'asserzione che gli esseri umani siano di fatto uguali per dote naturale".
Lo studio scientifico della diversità umana fa parte di un processo suddiviso in tre fasi. Nella prima fase si deve decidere se e quanto occuparsi di questo tipo di problemi. Alcuni, ad esempio Bodmer e Cavalli-Sforza (v., 1970), hanno sostenuto che il problema della diversità genetica per l'intelligenza non debba ricevere negli Stati Uniti un alto grado di priorità fino a quando il clima razziale di questo paese non sia mutato. Nella seconda fase, esclusivamente tecnica, si effettua lo studio scientifico del problema. La terza fase riguarda le decisioni e le iniziative da prendere nei riguardi di questa diversità.
È evidente che la prima, e soprattutto la terza, di queste fasi, sono di natura etico-politica. In particolare l'esperienza ci ha insegnato che nella realtà solo questa terza fase ha avuto un peso: quando si è presa la decisione di attuare una politica razzistica non la si è certo fatta dipendere da una dimostrazione e da un avallo scientifico, che del resto non è stato difficile procurarsi (cfr. il Manifesto sulla difesa della razza, del 1938).
b) Argomenti cari ai razzisti
Come si è accennato all'inizio, non c'è da spendere molte parole sugli argomenti sostenuti dai razzisti che, più che argomenti, sono slogan privi di qualsiasi base scientifica e razionale. Ne discuteremo alcuni solo per dimostrare quanto si è appena asserito e perché il fatto di essere irrazionali non ha loro impedito di svolgere un ruolo importante e di raggiungere apici di grande celebrità (un ruolo lo svolgono ancora, ma non più in forma ufficiale). Del resto, solo di rado l'irrazionalità ha costituito un serio ostacolo per le ideologie e per la loro traduzione in pratica. È molto più sostenibile, caso mai, la tesi opposta.
α) Razze ‛superiori' e razze ‛inferiori'. Hanno un senso questi attributi? Se applicati tout court, sono evidentemente un'assurdità che si può definire addirittura patetica, se si pensa che i razzisti, a qualsiasi razza appartengano, definiscono la propria superiore e le altre inferiori: l'etnocentrismo è un male molto diffuso.
Se invece ci si riferisce a caratteri specifici, allora questi gradi di qualità esistono senz'altro. Forse la nozione più certa e più chiara che abbiamo sulle diversità razziali è che esse hanno un'origine adattativa: quindi niente di più ovvio che gli Eschimesi siano geneticamente molto superiori ai Negri per il modo in cui sono adattati al freddo, e che si verifichi l'opposto per la vita nei climi torridi tropicali ed equatoriali. Si può anzi generalizzare affermando che di regola ogni razza nel proprio ambiente è superiore a tutte le altre: la progenie di qualsiasi razza sopraggiunta in un ambiente già occupato da una razza indigena, di regola, sul piano genetico, non ha che da guadagnare da incroci interrazziali, se è destinata a restare in quell'ambiente. In altre parole, caratteri che siano fissati, o anche semplicemente molto frequenti in una razza sono certamente buoni se quella razza si trova nel suo ambiente e cattivi in un ambiente diverso; caratteri utili nell'habitat originario (ad esempio, gli alleli ‛malarici' come il βS, i Th e i Gd ) possono diventare dannosi per le popolazioni che li posseggono in caso di mutamento di ambiente. Nessuna razza possiede con frequenza elevata alleli sfavorevoli in qualunque condizione, comprese quelle del suo stesso ambiente attuale o passato, perché la selezione ne avrebbe impedito l'accumulo.
β) Purezza della razza. Anche questo argomento, come del resto il precedente, è difficile da controbattere razionalmente, per il semplice motivo che non significa nulla. Se ciascuna razza avesse i propri alleli e questi fossero assenti nelle altre, allora avrebbe un senso discutere circa la desiderabilità e opportunità di introdurre nel pool genico di una razza altri alleli, per essa del tutto nuovi, provenienti da un'altra razza. Anzi questa domanda avrebbe un senso non solo etico, ma anche biologico. Ma parlare di purezza della razza nella situazione che ben conosciamo, in cui, salvo poche eccezioni, tutte le razze hanno gli stessi alleli e differiscono solo perché le loro frequenze sono diverse, è come dire che si vuole preservare la purezza di una soluzione di cloruro di sodio al tre per cento e che quindi occorre proteggerla da eventuali mescolamenti con una soluzione di cloruro di sodio al due e mezzo per cento.
Anche l'eventuale obiezione che le frequenze alleliche proprie delle varie razze sono quelle ottimali nel loro ambiente, per cui un cambiamento, causato da incroci con razze aventi frequenze alleliche diverse, potrebbe essere dannoso, è priva di valore. Infatti sono ormai molte le prove che per la grande maggioranza dei polimorfismi genetici (cioè per tutti salvo quelli bilanciati) le frequenze alleliche che si riscontrano attualmente sono il prodotto del caso, tanto che le varie razze mostrano distanze genetiche all'incirca proporzionali alla durata della loro segregazione (v. cap. 3, § c; v. cap. 6).
Per alcune ‛razze', infine, il concetto di purezza rasenta addirittura il ridicolo, come nel caso della ‛razza italica', che può essere presa addirittura come esempio classico di crogiuolo di razze, come risulta non solo dalla storia, ma è documentato anche geneticamente (per esempio, si trovano in Sicilia con frequenza apprezzabile geni tipicamente africani come il βS e il GdA-).
γ) Le caratteristiche di ogni individuo coincidono con quelle ‛proprie della sua razza'. Sostenere un argomento di questo genere è come sostenere che esiste solo una diversità tra le razze; quella entro le razze, che invece sappiamo essere molto maggiore, è considerata inesistente. Questo ‛errore' deriva dall'avere estrapolato a tutti i caratteri quanto si osserva per i soli caratteri antropologici classici (per i quali ciò effettivamente è in gran parte vero, per i motivi discussi in dettaglio nel cap. 6: per esempio tutti i Caucasici hanno la pelle più chiara di quella di tutti i Negri).
Valga ad esempio il quoziente di intelligenza (QI). Se si è raggiunta la convinzione che l'intelligenza di un certo gruppo di individui è inferiore a quella di un altro gruppo (per esempio, QI = 80 e 100, rispettivamente), ci si comporta come se tutti gli individui del primo gruppo avessero un QI di 80 e tutti quelli del secondo un QI di 100, cioè come se ciascuno dei soggetti del primo gruppo fosse meno intelligente di ciascuno di quelli del secondo. Si tratta di un ‛errore' statistico addirittura banale, che consegue però il risultato voluto, quello di discriminare in base alla razza invece che in base al carattere in questione, che serve solo da pretesto. Infatti, poiché i QI sono ampiamente dispersi intorno alle loro medie, le due distribuzioni sono ampiamente sovrapposte e quindi, se si volesse discriminare veramente in base all'intelligenza, si dovrebbero scartare indiscriminatamente tutti gli individui sotto una certa soglia, per esempio 90, accettando così molti degli appartenenti al primo gruppo e scartandone molti del secondo. Invece il risultato della discriminazione per gruppo è, ovviamente, che si eliminano soggetti molto intelligenti del primo gruppo a favore di soggetti ipodotati del secondo. Invece di valutare ogni individuo per quello che è, gli si attribuisce aprioristicamente, per estrapolazione, il valore medio attribuito alla sua razza.
I fautori di questa logica adottano per coerenza anche il procedimento opposto, compiendo l'‛errore' che, per la gravità delle sue conseguenze, è forse il più nefasto di tutti e che consiste nell'attribuire le proprietà caratteristiche di un singolo a tutta la sua razza. Da qui a considerare un intero gruppo razziale compartecipe e corresponsabile addirittura di un'azione - considerata, a torto o a ragione, criminale - che sia stata compiuta da un suo singolo membro, in certi casi nemmeno contemporaneo, il passo è molto breve: la storia passata e recente di tutti i tipi di razzismo pullula di esempi del genere. (V. anche razzismo).
c) Variabilità dell'intelligenza
È questo senza alcun dubbio l'argomento più scottante di un campo già di per sé delicato qual è lo studio delle razze. Ma non è questo l'unico motivo per cui si è scritto tanto su questo problema e ci sono state polemiche tanto feroci: un'altra ragione è che se ne sa così poco e il problema stesso è talmente complicato, che molti possono esprimere la loro opinione senza dimostrarla, ma anche senza che la si possa ‛falsificare' in modo assolutamente inconfutabile; ciò è invece possibile, almeno in linea di principio, per i problemi che si prestano a un approccio scientifico rigoroso, per i quali il livello delle opinioni viene superato abbastanza facilmente e si può raggiungere la certezza che una certa ipotesi sia vera oppure falsa. Il problema della variabilità dell'intelligenza è invece talmente complesso che non se ne prevede affatto una soluzione a breve scadenza; fra l'altro, la ricerca di questa soluzione non è certo facilitata dall'intervento di numerosi autori che usano come criteri di giudizio della validità di certe ipotesi le implicazioni delle stesse e tendono a considerare valide solo le ipotesi che hanno delle implicazioni a loro gradite. Per esempio, tra i criteri usati per controbattere l'ipotesi che la variabilità dell'intelligenza abbia una componente genetica c'è stato spesso l'argomento che, se così fosse, allora esisterebbe un certo grado di predestinazione. Ma nessuno ha mai potuto dimostrare che essa non esista per niente. Il fatto che l'esistenza di ‛un certo grado di predestinazione' possa riuscire scomoda per alcune ideologie, che negano a priori l'‛era scritto' (in questo caso nel DNA), dovrebbe essere - ma spesso purtroppo così non è stato - irrilevante al fine di accertare quale sia effettivamente la verità a questo proposito.
Nello studio della natura della variabilità dei caratteri ci si può trovare di fronte a un ampio spettro di situazioni. A un estremo stanno i caratteri ‛genetici' (come per esempio i gruppi sanguigni), dove questo attributo sta a indicare non tanto che sono caratteri ereditari, quanto che, per il fatto di essere discontinui, di essi si sa anche come sono trasmessi geneticamente (sono tutti caratteri mendeliani unifattoriali).
Ci sono poi i caratteri ‛fisici' quantitativi (cioè con variabilità continua). Di molti di essi è possibile misurare in modo rigoroso la variabilità fenotipica, perché sono fissi e perfettamente definibili (come per esempio la statura). Tuttavia, essendo la loro variabilità continua, diventa praticamente impossibile analizzarli accuratamente: sappiamo solo che tale variabilità ha una componente genetica di tipo polifattoriale, ma, appunto per questo motivo, ignoriamo come questi caratteri siano trasmessi (cioè quanti geni e quanti alleli siano coinvolti e come si esprimano).
Nel caso dell'intelligenza, o, per meglio dire, del QI, ci troviamo di fronte a un carattere non solo continuo, ma di cui è addirittura difficile misurare la variabilità fenotipica per molti motivi: il carattere misurato non è stabile (essendo influenzato dall'esperienza); lo strumento usato per misurarlo non è esatto; si usa un unico strumento tarato su una razza (la caucasica) per misurare il QI e la sua variabilità in altre razze (non esistono QI assolutamente ‛liberi da influenze culturali'). Infine nessuno può dire che cosa il QI rappresenti del patrimonio psichico di un individuo (forse dovremmo avere un QI più elevato per poterlo capire). È facile rendersi conto del perché si sappia tanto poco di sicuro sulla componente genetica di una variabilità che non si riesce a misurare e a comprendere in modo soddisfacente nemmeno a livello fenotipico.
Le uniche razze studiate piuttosto estesamente sono quella negra e quella caucasica. Quello che è stato accertato è facile a dirsi; quello che è stato ipotizzato molto meno.
1. Dati di fatto. - Di sicuro si sa solo che il QI è un carattere molto variabile, tanto fra le due razze meglio studiate che all'interno di ognuna di esse, e che, con questo test messo a punto sui Caucasici, il QI medio di questi ultimi è di circa 15 punti più elevato di quello dei Negri che vivono nella stessa zona degli Stati Uniti. Esso inoltre presenta nella parte settentrionale degli Stati Uniti valori più elevati che nel Sud, sia tra i Caucasici che tra i Negri, tanto che il QI medio di questi ultimi viventi negli Stati del Nord è più alto di quello medio dei Caucasici degli Stati del Sud. Questo per quanto riguarda la variabilità fenotipica.
Circa la componente genetica di questa variazione, è stato dimostrato con certezza che, all'interno della razza caucasica, essa esiste. Sulla sua grandezza è invece molto difficile esprimere un giudizio. Infatti è vero che essa appare a prima vista considerevole (per esempio la somiglianza tra i QI dei gemelli monozigoti è risultata molto elevata anche se erano stati allevati separatamente), ma è perfettamente possibile che essa sia stata sovrastimata persino negli studi più rigorosi (che sono pochi), perché non si è potuto tener conto nemmeno in questi della possibilità, tutt'altro che inverosimile, che esista una covarianza tra intelligenza e ambiente culturale, cioè che in media si sia verificata la tendenza da parte del gemello più intelligente a vivere in un ambiente intellettualmente più stimolante e la tendenza contraria da parte del gemello meno dotato. In questo caso, gemelli identici a lungo andare avranno accentuato la loro somiglianza aggiungendovi un'ulteriore causa, quella della somiglianza del loro ambiente culturale. Il risultato finale sarebbe evidentemente che l'effetto dell'identità del loro genotipo sarebbe sopravvalutato. Del resto è intuitivo, anche senza addentrarsi in dettagli statistici, che la validità della stima della componente ereditaria di un certo carattere, ottenuta con il metodo gemellare, si basa sull'assunto che le differenze tra gli ambienti a cui sono esposti i gemelli monozigoti siano in media eguali a quelle dei gemelli dizigoti.
Non sono stati effettuati studi sufficientemente estesi anche tra i Negri, e non ne sono stati tentati affatto per accertare in modo diretto se le differenze tra i Caucasici e i Negri avessero una base genetica (il disegno sperimentale necessario è in pratica inattuabile).
2. Ipotesi. - Abbandoniamo ora il terreno solido dei fatti e passiamo a quello infido delle ipotesi. Anzitutto una premessa indispensabile: per i caratteri quantitativi in generale, e per il QI ancora più che per gli altri, non si può parlare solo di componente genetica e di componente ambientale della loro variabilità. Occorre aggiungere la componente di interazione, che presumibilmente è quella predominante: a parità di variabilità genetica, due gruppi potrebbero mostrare una variabilità fenotipica tutta di natura ambientale (cioè, in questo caso, essenzialmente culturale), per cui in circostanze molto sfavorevoli tutti gli individui resterebbero, chi più chi meno, al di sotto delle loro potenzialità genetiche la cui variabilità sarebbe quindi irrilevante. Viceversa, in condizioni ambientali ottimali, tutti esprimerebbero il massimo delle loro potenzialità genetiche e la variabilità fenotipica osservabile, non essendo stato l'ambiente per nulla limitante, sarebbe tutta da attribuirsi a una variabilità genetica. Questi sono, evidentemente, quadri estremi e pertanto irrealistici, e la verità di sicuro non coincide con nessuno dei due (in condizioni uniformemente sfavorevoli si dovrebbe trovare una variabilità fenotipica ridotta rispetto a quella esistente in un ambiente, sia pure uniforme, ma favorevole. Questo è appunto quanto si è constatato confrontando la variabilità del QI nell'ambito di famiglie negre e di famiglie caucasiche: le prime mostravano una dispersione di valori molto minore). In conclusione il concetto di ereditarietà genetica della variabilità del QI è relativo; qualsiasi valore si sia attribuito alla sua componente genetica - non importa quanto esatto e accurato esso sia - ha una validità limitata alle circostanze ambientali (fisiche e culturali) nelle quali lo si è misurato. Non sono lecite estrapolazioni, se non estremamente limitate e con grandissima cautela. E invece è proprio in questo campo che sono state proposte le più ardite estrapolazioni, come quella di credere che l'elevata componente genetica della variabilità del QI trovata all'interno della razza caucasica costituisca una prova valida che questa componente è elevata anche per le differenze trovate tra razza caucasica e razza negra (v. Jensen, 1972 e 1973).
Chiarito questo punto, si possono considerare in breve le ipotesi più ragionevoli che sono giustificate dai pochi dati sicuri attualmente a disposizione.
3. Genetica della variabilità del QI - È presumibile che, come è stato dimostrato per i Caucasici, qualunque altra razza o gruppo umano che viva in condizioni ambientali e culturali ottimali o quasi ottimali mostrerebbe al suo interno un elevata variabilità del QI; è presumibile, inoltre, che una certa quota di questa variabilità avrebbe una base, sia genetica sia derivante dall'interazione tra genotipo e ambiente, il cui peso relativo è ragionevole supporre particolarmente elevato nelle zone estreme della distribuzione (al di sotto di certi valori molto bassi sono spesso in gioco con un ruolo determinante cause genetiche come, per esempio, l'idiozia fenilpiruvica, o anche disturbi dello sviluppo embrionale; per i valori elevatissimi, invece, non si hanno dati altrettanto certi, ma è assurdo pensare che qualunque famiglia avrebbe potuto diventare la famiglia Bach e che ognuno sia un potenziale Einstein o Mao Tse-tung).
Il problema del ruolo dei fattori genetici nel determinismo delle differenze tra razze per i rispettivi QI medi è certo di gran lunga il più rilevante dal punto di vista sociopolitico ed è certo quello che ha suscitato le polemiche più accese a livello scientifico. Si può dire anzi che esso costituisce l'unico caso in cui alcuni autori (v. Jensen, 1972 e 1973) sono giunti a conclusioni di tipo razzistico partendo da dati raccolti ed elaborati con criteri scientifici (in tutti gli altri casi, come si è visto prima, non si è curata nemmeno l'apparenza, per cui la confutazione delle tesi razzistiche non si è svolta a livello scientifico, ma ben al di sotto).
Ridotta all'essenziale, la critica che si deve muovere a Jensen è che, non disponendo di alcun dato diretto sull'esistenza o meno di una componente genetica della differenza di QI medio tra Caucasici e Negri, ha creduto di poterne fare a meno. Come si è accennato prima, egli ha semplicemente estrapolato l'alta ereditabilità genetica trovata ‛all'interno' della razza caucasica alla differenza ‛interrazze' osservata tra Caucasici e Negri. Non si vuole qui sostenere che nessuna estrapolazione sia lecita nella scienza (si tratta di uno degli strumenti concettuali più potenti e, giustamente, più utilizzati), ma che è giustificato ricorrervi solo quando ci sono dei buoni motivi. In questo caso, invece, non solo questi buoni motivi mancano, ma esistono addirittura delle controindicazioni precise e inconfutabili. Non si tratta di un partito preso per invalidare le conclusioni di Jensen (se fossero dimostrate lo si dovrebbe riconoscere, indipendentemente dalle proprie convinzioni sociali e politiche), ma di un atteggiamento scientifico di critica che, se si avessero a disposizione dati analoghi, si dovrebbe manifestare anche in campi diversi come, per esempio, quello delle varietà di piante coltivate (cioè in assenza di qualsiasi carica emotiva).
Un argomento tendente a dimostrare che alcune razze sarebbero geneticamente inferiori alla razza caucasica sul piano intellettuale, che ha avuto molto seguito fino a pochi anni fa e anche tra ben noti genetisti (v. Darlington, 1969 e 1972), è il seguente. Poiché le caratteristiche di comportamento, e non solo quelle di struttura, così specializzate negli animali domestici e in particolare nei cani, hanno sicuramente una base genetica, frutto della selezione artificiale da parte dell'uomo; poiché anche le razze umane sono state condizionate per tempi molto lunghi da ambienti (anche culturali) molto diversi, perché allora non credere che quello che si è verificato nelle razze canine non sia avvenuto anche nella nostra specie?
C'è un motivo molto importante per supporre che l'evoluzione del comportamento nell' uomo si svolga in modo assai diverso da quello degli altri animali. In questi ultimi essa non può essere che genetica; nell'uomo potrebbe essere genetica, ma anche culturale. Poiché ogni risposta adattativa segue la via di minor resistenza e l'adattamento per via culturale è appunto, senza alcun dubbio, la via di minor resistenza, sembra ragionevole credere che le differenze tra le razze dovute a caratteristiche comportamentali abbiano tutte o quasi tutte una base culturale. Oltre tutto non è stata mai dimostrata la benché minima correlazione tra quella che si potrebbe chiamare ‛idoneità culturale' (v. cap. 4, § a) e l'idoneità biologica, né c'è alcun motivo valido di supporre che essa esista, mentre l'esistenza di questa correlazione è una condizione necessaria affinché un adattamento di tipo culturale ne provochi anche uno di tipo genetico.
In conclusione, è stata trovata una certa differenza tra razza caucasica e razza negra per una certa proprietà detta QI, misurata con un test messo a punto per i Caucasici, che è correlata in modo imprecisato con ciò che chiamiamo intelligenza e in modo assolutamente indefinito con il complesso delle proprietà psichiche dell'uomo. Sebbene la variabilità di questo carattere all'interno della razza caucasica sia in parte determinata geneticamente, ci sono ragioni molto valide per supporre che la differenza tra Caucasici e Negri sia prevalentemente, se non esclusivamente, culturale, anche se dati pertinenti diretti mancano del tutto. In queste condizioni, il comportamento più saggio sembrerebbe consistere nel continuare a raccogliere dati accurati senza lasciarsi fuorviare da idee preconcette di nessun genere.
d) Gli incroci tra razze diverse
È interessante discutere l'argomento degli incroci tra razze diverse non solo da un punto di vista strettamente biologico, ma considerando anche l'atteggiamento assunto al riguardo dai genetisti, perché è illustrativo di quanto può essere fuorviata la scienza da fatti e da pregiudizi che dovrebbero esserle del tutto estranei.
Nel corso di questi ultimi secoli e soprattutto in quello attuale si sono verificati su vasta scala incroci tra razze anche molto lontane. Né la comune osservazione e nemmeno studi ad hoc - necessariamente non rigorosi data la natura del problema, sebbene ragionevolmente accurati - hanno messo in evidenza alcun effetto cospicuo di questo mescolamento razziale. Si può quindi affermare con certezza che gli incroci interrazziali hanno conseguenze non molto rilevanti, e comunque spesso troppo piccole per essere apprezzabili; probabilmente alcune saranno benefiche, altre dannose e altre ancora (verosimilmente la grande maggioranza) né dannose né benefiche, attribuendo a tali qualifiche un valore relativo a certe determinate circostanze ambientali. Non si direbbe proprio di trovarsi di fronte a un problema vitale della biologia, né sul piano teorico né su quello pratico. E invece esso ha suscitato e continua a suscitare un interesse che non è esagerato definire morboso.
Un interesse per questo argomento (non è il caso di chiamarlo problema, perché lo è solo per coloro che vogliono considerarlo tale) sarebbe stato giustificato ai tempi in cui questa enorme mole di dati non era disponibile e si poteva pertanto supporre che incroci tra razze diverse dessero origine a una prole molto peculiare (per esempio molto peggiore oppure molto migliore di quella generata dai comuni incroci intrarazziali). Ma adesso che tutto ciò si può escludere, il continuare a nutrire preoccupazioni al riguardo è segno inequivocabile di razzismo; e infatti coloro che sostengono, malgrado l'evidenza contraria, la necessità di preoccupazioni di tal genere fino al punto di giungere a interventi legali al riguardo, non hanno però avanzato proposte analoghe per incroci che di sicuro sono molto pericolosi (per esempio, incroci tra consanguinei o tra eterozigoti per lo stesso gene letale recessivo) ma concernono individui della stessa razza per essi, evidentemente, è meglio un figlio anormale da due cugini che uno normale da due persone di razza diversa. È chiaro quindi che i fautori di provvedimenti tendenti a limitare la libertà di scelta del partner con cui generare i propri figli non sono mossi dalla motivazione di proteggere il patrimonio genetico delle future generazioni, ma vogliono semplicemente che non si verifichino incroci interrazziali. Questo, per loro, è il fine, non il mezzo.
Il periodo d'oro del razzismo, la cui conseguenza più ovvia è l'avversione per gli incroci interrazziali, è tutto compreso in questo secolo, con una punta massima tra le due guerre mondiali e un brusco calo di popolarità subito dopo la seconda, in seguito al genocidio degli Ebrei. Esso non poteva non interagire con la genetica, la ‛scienza degli incroci', sorta e sviluppatasi in questo stesso periodo.
Dall'esame dell'atteggiamento ufficiale dei genetisti si ricava una storia (v. Provine, 1973) molto istruttiva e poco edificante per la genetica, caduta in pieno nell'equivoco - ma si tratta veramente di equivoco, o piuttosto di una delle innumerevoli forme di opportunismo e di conformismo mascherate da impegno politico? - di credere che il compito della scienza non sia tanto quello di accertare la verità, comunicarla quando ci si è riusciti e di riconoscere di avere fallito quando non ci si è riusciti, quanto quello di fornire la giustificazione scientifica della validità etica delle correnti di pensiero e delle decisioni sociali e politiche del momento. Caso mai sono queste ultime che si devono adeguare ai fatti accertati; e anche l'accertamento di questi fatti non deve avere niente a che vedere con alcuna ideologia. Tutto questo dovrebbe essere ovvio a tal punto da rendere superfluo l'illustrarlo e il discuterlo, ma la storia che considereremo ora in breve mostra che non è affatto inutile ribadirlo. Infatti, salvo lodevoli eccezioni (come, per esempio, W. E. Castle), la genetica si è limitata in ogni periodo a fornire l'avallo alle ideologie del momento affermando di possedere le prove scientifiche che le giustificavano. La sua posizione ufficiale ha di regola coinciso con mirabile coerenza con la posizione più comoda, e spesso è stata condizionata dai fatti in misura talmente trascurabile che gli stessi dati che in una certa epoca venivano considerati come la prova di certe tesi sono stati poi citati come prova della tesi opposta.
Già ai suoi albori, cioè in un'epoca che si può definire, di fatto se non cronologicamente, premendeliana (tra Mendel e la riscoperta delle sue leggi), la genetica dava per scontato, in assenza della benché minima prova, che la razza caucasica era geneticamente più intelligente delle altre. Ed era questo appunto che ci si aspettava da essa: alla fine dell'Ottocento il colonialismo è al suo apice.
Con l'inizio del Novecento si entra in epoca mendeliana. Il razzismo raggiunge il suo massimo sviluppo nel periodo compreso tra le due guerre mondiali. Malgrado che in questo stesso periodo la genetica conosca un rigoglio eccezionale, ciò non impedisce che si affermino quasi senza opposizione (almeno così risulta dalla letteratura genetica dell'epoca) idee come le seguenti: caratteri come la criminalità e il nomadismo sono caratteri mendeliani unifattoriali; gli incroci tra i Bianchi e i Negri sono una forma di altruismo dei primi - che si abbassano - verso i secondi - che si elevano - col risultato di una perdita netta di intelligenza da parte della società; la prole di questi incroci è esposta a forti rischi di essere disarmonica in base alla sorprendente teoria (che ha avuto molti seguaci) che i caratteri quantitativi sono ereditati settorialmente, per cui la prole di un genitore alto e di un genitore basso potrebbe ereditare braccia lunghe da uno e gambe corte dall'altro o presentare addirittura gravi discrepanze tra i visceri e lo scheletro che li contiene (‟large frames and inadequate viscera"). Va da sé che la prole di individui di statura anche altrettanto diversa, ma appartenenti alla stessa razza, non correrebbe rischi di questo genere.
Si tenga conto che alcune di queste affermazioni ‛scientifiche' erano basate su dati che - solo che fossero stati guardati senza idee preconcette - avrebbero dimostrato il contrario.
Verso la fine degli anni venti la mole di dati che escludeva che gli incroci interrazziali causassero in modo preferenziale disarmonie morfologiche cospicue era diventata inoppugnabile. Si fece strada allora l'opinione che essi provocassero disarmonie endocrine e mentali serie: qualcosa di grave dovevano pur causare!
Dopo una fase di agnosticismo, in cui finalmente si riconosce di non disporre di dati sufficienti per trarre conclusioni sicure, pur continuando a considerare il problema degli incroci interrazziali come uno dei problemi centrali - se non addirittura ‛il problema' - della biologia umana, si giunge alla seconda guerra mondiale e allo sterminio degli Ebrei che, pur essendo stato uno dei più atroci crimini della storia, non poteva certo fornire alcun dato pertinente o rilevante per il problema biologico delle eventuali conseguenze degli incroci tra razze diverse. E di nuovo i biologi umani, riuniti in una commissione sotto l'egida dell'UNESCO presieduta da A. Montagu con lo scopo esplicito di condannare il razzismo, si lasciano trascinare in dichiarazioni molto poco scientifiche, anche se ispirate a motivazioni che sul piano morale sono certo migliori delle precedenti. Tanto che molti genetisti si rifiutano giustamente di firmare questa dichiarazione del 1949. Essa contiene infatti affermazioni di questo genere: ‟Studi biologici hanno fornito prove a favore dell'idea etica della fratellanza universale". Espressioni come queste gettano discredito sulla validità scientifica di tutto il contesto di cui fanno parte e devono essere lasciate in esclusiva ai razzisti che sono obbligati a ricorrerci quando vogliono dare una veste pseudoscientifica alle loro tesi. Non è il caso di mettersi sul loro stesso piano né, fortunatamente, è necessario, per essere antirazzisti, dire delle assurdità. Durante tutto il periodo della seconda guerra mondiale e del genocidio degli Ebrei non erano stati prodotti dati scientifici nuovi rilevanti per il problema dell'incrocio tra razze diverse e meno che mai misurazioni della fratellanza umana; quindi anche in questo caso la posizione scientifica ufficiale di molti biologi umani - diametralmente opposta a quella sostenuta con vigore fino a non molti anni prima con gli stessi dati scientifici - era dettata da considerazioni estranee alla scienza e, per l'ennesima volta, in accordo addirittura troppo perfetto con quello che la società voleva sentirsi dire.
Viene allora promulgata dall'UNESCO nel 1951 una seconda dichiarazione, più razionale e meno emotiva della precedente. In sostanza, essa afferma che non vi è alcuna prova che gli incroci interrazziali siano potenzialmente dannosi, per cui qualsiasi forma di proibizione di incroci tra razze diverse non avrebbe alcuna giustificazione biologica. Erano passati più di venti anni da quando erano diventati disponibili dati a sufficienza per poter esprimere chiaramente e umilmente questo semplice concetto. È evidente che in argomenti come questi è molto difficile attenersi ai fatti.
bibliografia
Biasutti, R., Razze e popoli della terra, Torino 1959.
Bodmer, W. F., Cavalli-Sforza, L. L., Intelligence and race, in ‟Scientific American", 1970, CCXXIII, 4, pp. 19-29 (tr. it.: Intelligenza e razza, in ‟Le scienze", 1971, VI, 30, pp. 36-47).
Bodmer, W. F., Cavalli-Sforza, L. L., Genetics, evolution and man, San Francisco 1976 (tr. it.: Genetica, evoluzione e uomo, Milano 1977).
Cavalli-Sforza, L. L., Evoluzione: la moderna teoria dell'evoluzione, in Enciclopedia del Novecento, vol. II, Roma 1977, pp. 871-884.
Cavalli-Sforza, L. L., Bodmer, W. F., The genetics of human populations, San Francisco 1971.
Darlington, C. D., The evolution of man and society, London 1969.
Darlington, C. D., Race, class and culture, in Biology and the human sciences (a cura di J. W. S. Pringle), Oxford 1972, pp. 95-120.
Dobzhansky, Th., Genetic diversity and human equality, New York 1973 (tr. it.: Diversità genetica ed uguaglianza umana, Torino 1975.
Hiernaux, J., The people of Africa, London 1974.
Ingle, D. J., Fallacies in arguments on human differences, in Human variation, the biopsychology of age, race and sex (a cura di R. T. Osborne, C. E. Noble e N. Weyl), New York 1978, pp. 5-27.
Jensen, A. R., Genetics and education, New York 1972.
Jensen, A. R., Educability and group differences, New York 1973.
Lévi-Strauss, Cl., Antropologia, in Enciclopedia del Novecento, vol. I, Roma 1975, pp. 202-214.
Lewontin, R. C., The genetic basis of evolutionary change, New York 1974.
Lewontin, R. C., Models of natural selection, in Mathematical models in biology. International ‛Vito Volterra' symposium, Roma 1979.
Luzzatto, L., Studies of polymorphic traits for the characterization of populations. African populations south of the Sahara, in ‟Israel journal of medical sciences", 1973, IX, pp. 1181-1194.
Luzzatto, L., Genetics of red cells and susceptibility to malaria, in ‟Blood", 1979, LIV, pp. 961-976.
Menozzi, P., Piazza, A., Cavalli-Sforza, L. L., Synthetic maps of human gene frequencies in Europeans, in ‟Science", 1978, CCI, pp. 786-798.
Modiano, G., Grado e natura della variabilità genetica: sua rilevanza biologica, in Colloquio di genetica di popolazioni, Accademia Nazionale dei Lincei, Roma 1976, pp. 63-116.
Montalenti, G., Evoluzione: l'evoluzionismo nella cultura del XX secolo, in Enciclopedia del Novecento, vol. II, Roma 1977, pp. 865-871.
Mourant, A. E., Köpec, A. C., Domaniewska-Sobczak, K., The distribution of the human blood groups and other polymorphisms, London-New York 1976.
Provine, W. B., Geneticists and the biology of race crossing, in ‟Science", 1973, CLXXXII, pp. 790-796.
UNESCO, The race concept, Paris 1951.
Workman, P. L., Gene flow and the search for natural selection in man, in ‟Human biology", 1968, XL, pp. 260-279.