
Selezione
Selezione
di John M. Thoday
SOMMARIO: 1. Introduzione. □ 2. Tipi di selezione. □ 3. Selezione naturale e artificiale e idoneità biologica. □ 4. Selezione in un singolo locus genico. □ 5. Polimorfismo: a) esempi di polimorfismi transeunti; b) esempi di polimorfismi che dipendono dalla frequenza. □ 6. La selezione in condizioni multigeniche: a) selezione direttiva e variabili continue; b) selezione stabilizzante; c) selezione diversificante. □ Bibliografia.
1. Introduzione.
Il concetto di selezione comparve per la prima volta con Empedocle di Agrigento: nel tentativo di spiegare sia la genesi della vita, sia l'organizzazione e l'adattamento degli organismi, questi postulò che la vita si fosse originata nel fango per sviluppo spontaneo di tutti i diversi possibili tipi di arti, tronchi e teste. Successivamente arti, tronchi e teste si sarebbero uniti a caso costituendo una gran varietà di organismi, alcuni vitali e altri no: quelli vitali sarebbero stati in grado di sopravvivere mentre gli altri sarebbero stati eliminati. Empedocle spiegava così l'organizzazione dei sopravvissuti attraverso la selezione naturale e l'incapacità di riprodursi di quelli male organizzati, nonché, incidentalmente, l'origine di alcuni mostri mitologici.
Il concetto di selezione naturale divenne importante solo quando, verso la metà del XIX secolo, Ch. Darwin e A. R. Wallace pubblicarono le loro opere fondamentali sull'evoluzione (v. evoluzione). La teoria dell'evoluzione però esisteva già da tempo: il nonno di Darwin, E. Darwin, e in seguito J.-B. de Lamarck avevano con chiarezza ipotizzato che le specie si fossero originate attraverso la discendenza per modificazioni successive, ma non era stata avanzata alcuna ipotesi, tranne quella di Lamarck e altre a essa collegate, sul meccanismo che poteva determinare tali modificazioni. Darwin e Wallace compresero entrambi che, se esistevano variazioni ereditabili, queste dovevano influenzare le probabilità dei vari organismi di contribuire alle future generazioni. Questo processo, per mezzo del quale la riproduzione differenziale di organismi con patrimonio genetico diverso porta a una modificazione delle caratteristiche genetiche di una popolazione, è fondamentalmente ciò che noi chiamiamo selezione.
Nè Darwin nè Wallace proposero una spiegazione per l'insorgere delle variazioni sulle quali dovrebbe operare la selezione; capirono però che dal momento che gli organismi sono in grado di riprodursi in numero maggiore di quanto può essere tollerato dalle risorse ambientali, deve esistere una qualche forma di eliminazione. Da ciò deriva che, se esiste una variazione genetica di ciò che noi chiamiamo idoneità biologica (fitness), allora la riproduzione differenziale può condurre a una modificazione delle caratteristiche genetiche di una popolazione.
Lo stesso Darwin, incapace di scoprire l'origine delle variazioni sulle quali la selezione può operare, divenne in seguito sostanzialmente un lamarckiano e suppose che una delle cause delle variazioni ereditabili fosse da ricercarsi nelle conseguenze dell'uso o del disuso delle capacità dell'organismo, conseguenze che, se sono ereditabili, possono determinare la comparsa di una progenie meglio adattata rispetto ai genitori. Darwin però suppose anche che negli organismi vi fossero fonti di modificazioni delle caratteristiche ereditabili che ‟nella nostra ignoranza chiamiamo spontanee", modificazioni che oggi indichiamo con il termine di mutazioni. Darwin propose quindi i due principî fondamentali della moderna teoria dell'evoluzione che, negando la possibilità di ereditare gli effetti dell'uso e del disuso, ma ammettendo la variazione per mutazione e la riproduzione differenziale delle varie forme originate per mutazione, spiega appunto come la selezione naturale sia in grado di determinare quali forme mutanti sopravviveranno con maggiore frequenza. Da allora molte ricerche sono state condotte sulla selezione e sui suoi effetti oltre che sulla mutazione, sulla sua frequenza e sui vari suoi tipi. In alcuni casi si è trattato di analisi teoriche e matematiche, in altri di esperimenti di laboratorio o di esami di popolazioni naturali; ma oltre a ciò una grande massa di informazioni si è ottenuta dalla genetica applicata all'allevamento delle piante e degli animali, settori nei quali viene estesamente impiegata la selezione artificiale invece di quella naturale.
2. Tipi di selezione.
Lo stesso Darwin aveva già identificato una selezione naturale, una artificiale e un terzo tipo, intermedio fra i due precedenti, attuato inconsapevolmente dall'uomo, il quale, anche quando non sta deliberatamente selezionando in vista di un certo fine, può, senza rendersene conto, influire, favorevolmente o sfavorevolmente, sui suoi raccolti coltivando campioni non casuali. Un esempio di selezione inconsapevole e sfavorevole si è verificato nel passato nelle foreste, quando i boscaioli abbattevano gli alberi che fornivano il migliore legname lasciando che la foresta si riproducesse naturalmente con quegli alberi che essi avevano di fatto rifiutati; un procedimento di questo tipo, ripetuto per varie generazioni, può provocare lo scadimento della qualità del legname. Un altro esempio di selezione inconsapevole e sfavorevole è fornito dalle conseguenze del trattamento delle erbe da pascolo da parte di coloro che le coltivano per raccogliere i semi da mettere in commercio per la semina di nuovi pascoli. Gli agricoltori richiedono erbe con molte foglie e una lunga stagione di produzione, mentre i produttori di semi trattano le erbe da pascolo come piante da raccolto e quindi selezionano piante con breve stagione di produzione e periodo di maturazione uniforme, il che è corretto per il grano e l'orzo, ma del tutto errato per le erbe da pascolo. Un esempio di selezione inconsapevole, ma favorevole, potrebbe essere quello rappresentato dall'origine delle nostre piante di cereali giganti: infatti piante che crescono molto producono una quantità maggiore di semi, e l'agricoltura sviluppandosi può aver operato una selezione inconsapevole a favore di quelle piante che per costituzione genetica presentavano un buon adattamento e un'alta produttività nelle condizioni di coltivazione dei popoli primitivi.
Secondo altri criteri di classificazione la selezione, oltre che in naturale e artificiale, può essere distinta in direttiva, stabilizzante e diversificante, come vedremo successivamente. La selezione, inoltre, può dipendere dalle frequenze geniche e dalla densità di popolazione. Ancora, v'è un tipo di selezione, preso in considerazione da Darwin e frequentemente designato come selezione sessuale, in cui genotipi diversi hanno probabilità diverse di realizzare l'accoppiamento più adatto.
3. Selezione naturale e artificiale e idoneità biologica.
La distinzione fra selezione naturale e artificiale dipende dai fattori che sono determinanti per l'idoneità biologica: con tale termine ci riferiamo al differente contributo che genotipi diversi danno alle generazioni successive; è infatti solamente tramite questa diversa partecipazione di differenti genotipi che può aver luogo l'evoluzione genetica.
La selezione naturale è ovviamente implicata nell'evoluzione naturale, e in questo caso ciò che determina le varie idoneità biologiche sono i differenti livelli di adattamento dei diversi genotipi di una popolazione alle circostanze ambientali presenti in quel momento. Nella selezione artificiale invece è lo sperimentatore o l'allevatore di piante o animali che impone idoneità biologiche diverse ai diversi genotipi. Cosi in una popolazione in cui alcuni individui sono più alti di altri e in cui le differenze di altezza sono, almeno in parte, determinate geneticamente, lo sperimentatore può allevare solo gli individui più alti della media oppure quelli estremamente alti; in questo caso egli impone un'idoneità biologica vantaggiosa agli individui più alti, poiché accresce la loro probabilità di fornire progenie alle generazioni successive. La selezione naturale può ovviamente continuare a operare anche negli esperimenti di selezione artificiale, poiché i diversi genotipi presenti negli individui che il ricercatore sceglie come genitori o riscontrabili nella progenie che ne discende possono possedere idoneità biologiche diverse nelle condizioni ambientali adottate per allevare gli animali e le piante. Ciò che però le due forme di selezione hanno in comune è che le modificazioni nel corso delle generazioni saranno determinate dal grado di varietà genetica e dalla differente probabilità dei diversi genotipi di una popolazione di fornire progenie alle generazioni successive. In ambedue i casi le ineguali idoneità biologiche di geni o di combinazioni di geni diversi determinano la direzione e l'entità della variazione che avverrà per effetto della selezione naturale o artificiale.
4. Selezione in un singolo locus genico.
In una popolazione di animali o di piante comuni vi possono essere una o più forme di un gene per ogni posizione sui cromosomi; se in una popolazione vi sono due forme (alleli) di un gene, vi possono essere tre genotipi. Per esempio, nell'uomo vi è un locus genico per il quale vi sono due alleli di un gene, che possono essere denominati A e a, i quali influenzano lo sviluppo di pigmento nella pelle: se vi è pigmento nella cute deve essere presente l'allele A, mentre la presenza del solo allele a è caratteristica di un soggetto privo di pigmento nella pelle, nei peli e nelle iridi, denominato albino. Quando avviene la riproduzione, i due alleli, che occupano posizioni corrispondenti in una coppia di cromosomi, si separano durante la formazione di uova o spermi, cosicché l'uovo e lo spermio che si uniscono per dare luogo a un uovo fecondato contengono, ognuno, solo un rappresentante della coppia di geni A e a, dal momento che uova e spermi possono contenere o A o a, ma non ambedue. Al momento della fecondazione si formano uova fecondate che ovviamente contengono due geni, uno proveniente dall'uovo, l'altro dallo spermio: se ambedue contenevano A, l'individuo cui l'uovo fecondato darà origine sarà AA e avrà pigmentazione normale; se invece sia l'uovo sia lo spermio contenevano a, l'individuo avrà costituzione aa e sarà albino; se infine l'uovo conteneva A e lo spermio a, o viceversa, l'individuo sarà Aa, e avrà normale pigmentazione dal momento che possiede A, ma sarà in grado di produrre gameti contenenti a. Sono gli individui di questo tipo, detti ‛eterozigoti', che di solito, accoppiandosi fra loro, hanno figli albini. Infatti se due eterozigoti si uniscono, ci si attende che 1/4 dei loro figli sia albino e che i rimanenti 3/4 siano normali, come è indicato nello schema seguente ove è riportata la tipologia della discendenza dei matrimoni fra eterozigoti:
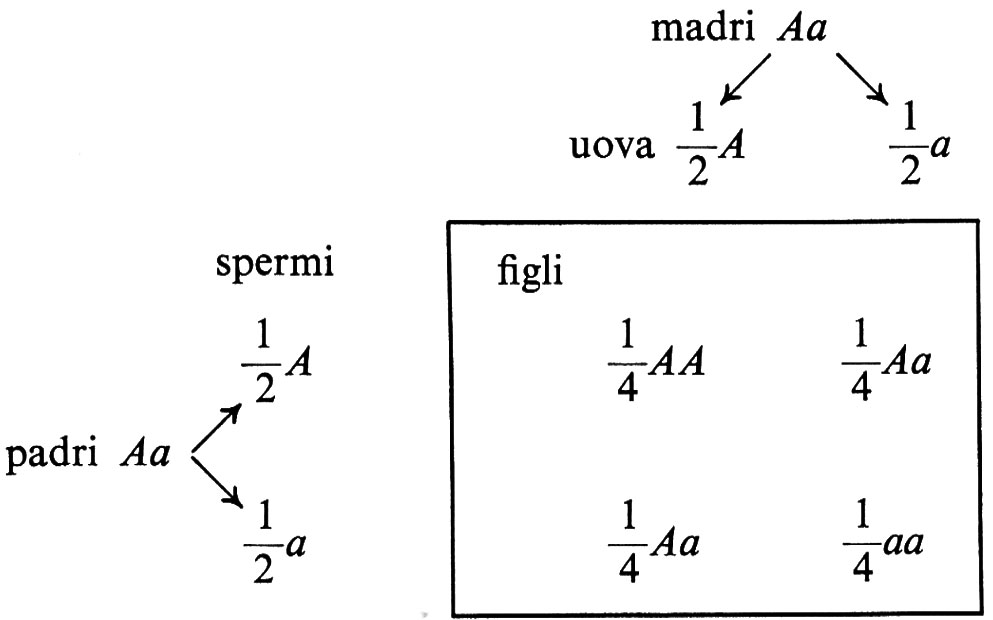
Quindi le uova fecondate saranno per 1/4 AA, per 1/2 Aa e per 1/4 aa e pertanto si svilupperanno da esse individui per 3/4 non albini e per 1/4 albini. Estendiamo ora questo schema all'intera popolazione: di tutte le uova in essa presenti una quota, che indichiamo con p, sarà A, mentre il restante, che chiamiamo q, sarà a; analogamente accade per gli spermi. In questo modello semplice p e q costituiscono tutti i geni presi in considerazione nella popolazione e quindi p + q = 1. Quindi, quando le p uova, che sono A, si combinano con gli spermi A avremo p2 uova fecondate AA, e pq uova fecondate Aa; d'altro canto, quando le q uova, che sono a, si combinano con gli spermi, avremo qp uova fecondate Aa e q2 uova fecondate aa. In una popolazione sono quindi prevedibili p2 individui AA, 2pq (cioè pq +qp) individui Aa e q2 individui aa. Tali individui a loro volta daranno origine, nel formare la generazione successiva, a p uova A e a q uova a, e analogamente a p spermi A e a q spermi a, come era nella generazione precedente. Quindi, se nulla viene a disturbare le frequenze geniche della popolazione, le frequenze degli individui AA, Aa e aa rimarranno inalterate nel corso delle generazioni. Si determina invece una selezione se uno dei tre genotipi di fatto si riproduce meno efficacemente degli altri. L'albinismo, come si è detto, è un carattere ‛recessivo': ciò vuol dire che non riusciamo a distinguere gli individui AA da quelli Aa, osservando il pigmento presente nella loro pelle. Ora, gli individui albini (aa) si riproducono un po' meno degli altri: da una parte sono soggetti a cancro cutaneo, dall'altra hanno difficoltà sociali, ad esempio nella ricerca del coniuge, che possono determinare uno svantaggio nella riproduzione. L'idoneità biologica degli individui aa sarà quindi inferiore a quella degli individui AA e Aa: in altri termini a ogni bambino generato da individui AA o Aa corrisponde, in media, meno di uno generato dagli albini. Quindi l'idoneità biologica dei tre genotipi può essere definita 1 per gli individui AA e per quelli Aa e 1-s per gli individui aa, dove s è lo svantaggio selettivo che colpisce i genotipi aa. Nella teoria della selezione s è detto coefficiente di selezione.
È ora facile vedere che in questa situazione, dal momento che gli individui aa contribuiscono alla generazione successiva un po' meno degli altri, vi sarà una perdita relativa di geni a; quindi, se la frequenza dei geni a in una certa generazione è q, in quella successiva sarà un po' meno di q: in realtà è possibile dimostrare che la perdita di geni corrisponde a (q2s)2, dal momento che vi sono q2 individui aa e ognuno di essi possiede 2 geni a. Se ciò accade, la frequenza dei geni a, cioè q, diminuirà con il passare delle generazioni con una velocità che può essere calcolata, se è noto s, fino al momento in cui tutti i geni presenti nella popolazione saranno A. È questo un esempio di selezione naturale che agisce sulla frequenza dei geni di un singolo locus: un esempio di popolazione che sta perdendo un gene secondo questa modalità è illustrato nella fig. 1.
Il modello sopra descritto è ovviamente molto semplice: bisogna perciò prendere in considerazione qualche piccola complicazione. Se infatti le cose fossero così semplici come nel nostro esempio, il gene svantaggiato dovrebbe con il tempo scomparire completamente, così che alla fine nella nostra popolazione non sarebbe più possibile trovare albini. Vi è però un altro fenomeno, che noi chiamiamo ‛mutazione', che può modificare le frequenze geniche in una popolazione. Si dice che vi è stata una mutazione quando un gene A diviene improvvisamente un gene a e questo si replica nelle divisioni cellulari fedelmente, come accadeva quando era A. La mutazione, almeno in parte, è un processo reversibile, così che in una popolazione in cui siano contemporaneamente presenti geni A e a, avviene regolarmente che una piccolissima parte di geni A diventa a, mentre una piccola quota di geni a diventa A. Nella popolazione vi è perciò da un lato un aumento di geni a che corrisponde alla quota del gene A moltiplicata per la frequenza con cui i geni A diventano a, frequenza che chiamiamo u, e pertanto la velocità di perdita di geni A da parte della popolazione è pu; d'altro canto, sempre a opera della mutazione, vi è un aumento di geni A, o perdita di geni a, che corrisponde a qv, dove v è la frequenza con cui i geni a diventano A. In conclusione, se non vi è selezione, se cioè gli individui AA, Aa e aa hanno tutti la medesima idoneità biologica, è prevedibile che con il tempo la popolazione raggiunga una situazione bilanciata o (equilibrio di Hardy-Weinberg) quando la velocità di perdita di geni A (incremento di geni a) corrisponde esattamente alla velocità di perdita di geni a (incremento di geni A), quindi quando pu = qv. Se invece la selezione è operante, l'equilibrio è più complesso, perché in questo caso abbiamo sia la perdita di geni A per mutazione, sia la perdita di geni a per selezione. Quindi per selezione e mutazione operanti contemporaneamente, l'equilibrio sarà raggiunto quando pu, la velocità dell'aumento di geni a, sarà uguale a qv + 2sq2, cioè alla velocità di perdita di geni a a opera della mutazione e della selezione.
Possiamo capire meglio il significato di tutto ciò facendo un esempio numerico. Sperimentalmente si è visto che circa ogni centomila uova o spermi possiamo trovare un gene mutato spontaneamente in un dato locus: adottiamo quindi questo valore come frequenza di mutazione. Supponiamo poi che s sia realmente molto piccolo, dell'ordine di 1/100 o 0,01; ciò significa che ogni 100 figli generati da individui AA e ogni 100 figli generati da individui Aa ne nascono, in media, solo 99 da individui aa: una differenza nell'idoneità biologica molto difficile, se non impossibile, da misurare. Sostituendo questi valori nell'equazione dell'equilibrio, si trova che q corrisponde all'incirca a 0,04 o 4%, e quindi q2 = 0,0016 o circa 2 su 10.000. È possibile quindi verificare che una popolazione con questa frequenza di mutazione e sottoposta a una pressione selettiva così piccola all'equilibrio conterrà alcuni mutanti, ma ben pochi, qualunque sia stata la loro frequenza in origine. Ciò serve a dimostrare che è la selezione e non la mutazione a determinare fondamentalmente le frequenze geniche di una popolazione; e del resto, poiché la teoria che abbiamo descritto implica che una popolazione ad alta frequenza di geni sfavorevoli tende gradualmente a perderli per opera della selezione, possiamo dire che proprio quest'ultima determina fondamentalmente la direzione dell'evoluzione. La mutazione è ovviamente la fonte della differenza genetica su cui la selezione può operare, ma non è la frequenza di mutazione verso una qualche particolare direzione che determina sostanzialmente le caratteristiche future di una popolazione. La mutazione sarà un fattore determinante delle frequenze geniche solo nel caso che le diverse forme di un gene in un particolare locus cromosomico abbiano in realtà trascurabili effetti differenziali sull'idoneità biologica.
Tale dimostrata impossibilità della frequenza di mutazione a determinare la direzione dell'evoluzione può indurre il lettore a chiedersi donde questa in sostanza derivi, dal momento che è ovvio che la selezione può operare solo se vi sono geni diversi tra i quali effettuare una ‛scelta'. Nei riguardi dell'evoluzione, il ruolo della mutazione è di importanza pari a quello svolto dalla selezione, in quanto fonte dei geni nuovi sui quali può operare la selezione stessa. Ma l'evoluzione continua di una specie non dev'essere considerata dipendente sostanzialmente dall'insorgere in qualsiasi momento di variazioni mutazionali mai avvenute in precedenza e in grado di fornire geni migliori di quelli già esistenti; dobbiamo piuttosto ritenere che essa dipenda da modificazioni delle condizioni ambientali di vita per cui geni, comparsi per precedenti mutazioni e che non avevano fornito un vantaggio selettivo, vengono a trovarsi in circostanze tali da risultare vantaggiosi e da potersi perciò diffondere.
5. Polimorfismo.
Le condizioni teoriche elementari esaminate finora, che appartengono alla genetica delle popolazioni sviluppata intorno al 1930 da R. A. Fisher, J. B. S. Haldane e s. Wright, implicano che, per quanto riguarda singoli loci genici, una popolazione dovrebbe essere omogenea a meno che due alleli diversi di un locus genico siano veramente neutri per quel che riguarda la loro idoneità biologica. Al contrario, ricerche su popolazioni naturali hanno spesso dimostrato, e in alcuni casi ciò è noto da molto tempo, che possono esistere contemporaneamente più forme geniche. Quando due geni per lo stesso locus coesistono in una popolazione e il più raro dei due ha una frequenza troppo alta per essere spiegata in base alla pressione di mutazione, una frequenza il cui limite massimo è spesso considerato pari al 10%, si dice che la popolazione in questione è polimorfica. Tale concetto è stato ampiamente discusso da E. B. Ford, il quale ha distinto due grandi categorie: il ‛polimorfismo transeunte' e il ‛polimorfismo bilanciato'. Il polimorfismo è detto transeunte se la popolazione è considerata nel suo processo di evoluzione, quando cioè un gene in precedenza anormale per la popolazione è divenuto vantaggioso e la sua frequenza sta quindi progressivamente crescendo, mentre quella del gene in precedenza normale si riduce. Tali polimorfismi non presentano problemi particolari: sono semplicemente esempi di evoluzione in corso. Se invece il polimorfismo è stabile, cioè se due geni alternativi per lo stesso locus sono mantenuti a un certo livello di equilibrio per periodi di tempo abbastanza lunghi, ci troviamo chiaramente in una situazione non prevista dalle condizioni di equilibrio tra selezione e mutazione, illustrate prima. Esistono due diverse ipotesi per spiegare l'esistenza dei polimorfismi di questo tipo: alcuni ritengono che i geni rilevanti siano neutri e che quindi il polimorfismo non abbia alcun significato per l'organismo; altri pensano invece che il polimortismo sia attivamente mantenuto dalla selezione.
La selezione può in linea di principio mantenere i polimorfismi, ed effettivamente in qualche caso ciò è stato dimostrato. La selezione raggiunge questo risultato con due diversi meccanismi: nel primo l'eterozigote, con costituzione Aa, come abbiamo già visto nel caso dell'albinismo, può essere più idoneo di ambedue gli omozigoti (AA o aa) e, quando ciò accade, la selezione opera attivamente mantenendo ambedue i geni nella popolazione. Il secondo meccanismo comporta in qualche modo una ‛selezione dipendente dalle frequenze', in cui cioè le idoneità biologiche relative dei geni alternativi dipendono dalla loro frequenza di modo che se A è comune e a è raro, a è il più idoneo dei due geni e, viceversa, se A è raro e a è comune, A è il più idoneo. Vi è poi una terza situazione nella quale può essere mantenuto il polimorfismo bilanciato di un locus: ciò accade quando l'ambiente è eterogeneo per ciò che riguarda lo spazio o il tempo. Per esempio, un gene può essere avvantaggiato in una stagione dell'anno e l'altro in un'altra stagione e, se l'organismo presenta molte generazioni nel corso dell'anno, si può verificare una ‛selezione ciclica': ambedue i geni saranno presenti in ogni momento, ma uno è più frequente d'inverno, l'altro in primavera. Casi di selezione di questo tipo sono stati studiati da N. W. Timoféeff-Ressovsky nelle coccinelle che presentano forme differenti, con macchie più o meno numerose, a frequenza diversa nelle varie stagioni. Un altro esempio molto conosciuto è fornito dalle forme alternative di cromosomi in Drosophila pseudo-obscura, studiate da Th. Dobzhansky, alcune delle quali variano la loro frequenza relativa in modo ciclico durante l'anno (v. fig. 2). Peraltro non è certo che la situazione di selezione ciclica mantenga necessariamente un polimorfismo permanente, poiché una popolazione potrebbe perdere uno dei geni nella stagione in cui esso è svantaggioso: per evitare quest'eventualità, sarebbe necessaria un'idoneità superiore degli eterozigoti.
Anche condizioni ambientali eterogenee per ciò che riguarda lo spazio possono determinare in una popolazione una situazione in cui in una particolare zona è favorito un gene mentre in un'altra è favorito un gene alternativo. Generalmente in situazioni di questo tipo si parla di selezione diversificante. Talvolta è denominata selezione da nicchia ecologica quella in cui geni diversi sono favoriti in nicchie ecologiche diverse. La possibilità, peraltro dubbia, che condizioni di questo tipo mantengano un polimorfismo senza che sia implicato anche il fattore frequenza, è comunque verosimile: infatti le dimensioni delle diverse nicchie ecologiche, espresse come numero di membri delle specie rilevanti che la nicchia è in grado di ospitare, possono determinare una situazione in cui, se diviene troppo frequente il genotipo favorito per una data nicchia, non è più possibile che questa venga occupata da tutti gli individui di quel genotipo, che devono quindi tentare di vivere in una nicchia diversa, nella quale sono svantaggiati. Quindi, se un gene e troppo frequente rispetto alle dimensioni relative della nicchia, diviene svantaggiato; se invece è meno frequente di quanto sia compatibile con le dimensioni della nicchia, è favorito. Tutte le situazioni di questo tipo saranno prese in considerazione nel paragrafo che riguarda la selezione diversificante, quando passeremo in rassegna casi reali di selezione naturale o artificiale.
a) Esempi di polimorfismi transeunti.
L'esempio più classico di polimorfismo transeunte, l'osservazione cioè, in natura, della modificazione di una frequenza genica per un singolo locus, è fornito dal fenomeno noto come melanismo industriale. Dalla metà del secolo scorso in poi, nei paesi industrializzati, numerose popolazioni di Insetti sono cambiate notevolmente: infatti le forme a colorazione scura, di solito estremamente rare, sono divenute sempre più comuni, così che in alcune aree, per esempio le zone industriali dell'Inghilterra centrale, le popolazioni di Insetti sono ora costituite praticamente solo da individui a colorazione scura. H. B. D. Kettlewell (v. mimetismo) ha studiato attentamente una di queste specie, Biston betularia, una falena le cui forme melaniche o scure si sono diffuse rapidamente nelle regioni industriali inglesi a partire dal 1848. Cinquant'anni dopo, in alcune zone industriali avevano raggiunto il 90 e persino il 95%. Sostanzialmente, la forma melanica differisce dalla ‛tipica' per una singola sostituzione genica, ed è stata formulata l'ipotesi che il principale fattore determinante l'idoneità biologica relativa delle due forme fosse la predazione da parte di uccelli nelle zone non inquinate, poichè le forme originali delle falene erano molto ben mimetizzate. Infatti, quando l'inquinamento industriale eliminò i licheni dagli alberi dove le falene riposano durante il giorno, il mimetismo divenne insufficiente, e la forma tipica, risultando facilmente identificabile, fu predata dagli uccelli; al contrario, le rare forme nere si mimetizzarono meglio nel nuovo ambiente e acquisirono quindi un'idoneità biologica relativa migliore di quella della forma originaria (v. fig. 3, A e B). Gli ornitologi rifiutarono questa ipotesi, poiché nessuno aveva mai visto uccelli predare le falene sui tronchi degli alberi durante il riposo diurno. N. Tinbergen, lavorando con Kettlewell, riuscì a dimostrare, con riprese cinematografiche al rallentatore, che gli uccelli si muovevano troppo in fretta perché il fenomeno potesse essere osservato a occhio nudo (v. fig. 3 C). Kettlewell poi, ponendo un numero uguale di falene tipiche e melaniche su tronchi d'albero in una zona industriale inquinata e in una non inquinata e osservando quanti insetti di ciascuna delle due forme venivano catturati dagli uccelli, dimostrò elegantemente una predazione differenziale molto rilevante, poiché la forma melanica è catturata più spesso nei boschi indenni, mentre quella tipica è predata di più sugli alberi delle zone inquinate (v. tab. I; v. Kettlewell, 1955 e 1956).
Inoltre, dopo aver allevato moltissime falene di entrambi i tipi e dopo averle opportunamente marcate, Kettlewell le liberò nei boschi, ricatturandole poi con trappole: riuscì così a confermare una notevole differenza di sopravvivenza e al tempo stesso dimostrò che, in condizioni naturali, notevoli pressioni selettive venivano esercitate in favore delle falene chiare nei boschi non inquinati e in favore di quelle scure nei boschi inquinati. Quest'esempio di polimorfismo transeunte viene poi ulteriormente complicato dal fatto che le forme scure sono un po' più vigorose di quelle chiare, ma tale superiorità non riesce a compensare il loro svantaggio nel mimetismo quando i tronchi degli alberi, in zone non inquinate, sono ricoperti da licheni. La fig. 4 mostra come variano in Inghilterra le frequenze delle forme melaniche.
Sono state compiute numerose ricerche sperimentali che dimostrano come la selezione modifichi le frequenze geniche in popolazioni allevate in laboratorio. Nella fig. 5, desunta dalle indagini di Ph. L'Héritier e G. Teissier (v., 1937), sono illustrati i due primi esperimenti di laboratorio sulla selezione naturale: si tratta di studi ormai classici, condotti su Drosophila, che dimostrano gli effetti della selezione su una singola differenza genica. Nel primo esperimento il gene anormale Bar scompare progressivamente; nel secondo il gene ebony, sebbene per effetto della selezione presenti una frequenza progressivamente ridotta a un valore piuttosto basso, raggiunge alla fine un livello stabile al quale è ancora presente, così che nella popolazione si determina un polimorfismo stabile. Ciò deriva probabilmente da un certo vantaggio posseduto dall'eterozigote, ma anche in questo esempio, comunque, nel periodo durante il quale il gene ebony continua a essere ampiamente soppiantato dal suo allele normale, si può dire che la popolazione è in una situazione di polimorfismo transeunte.
b) Esempi di polimorfismi che dipendono dalla fre quenza.
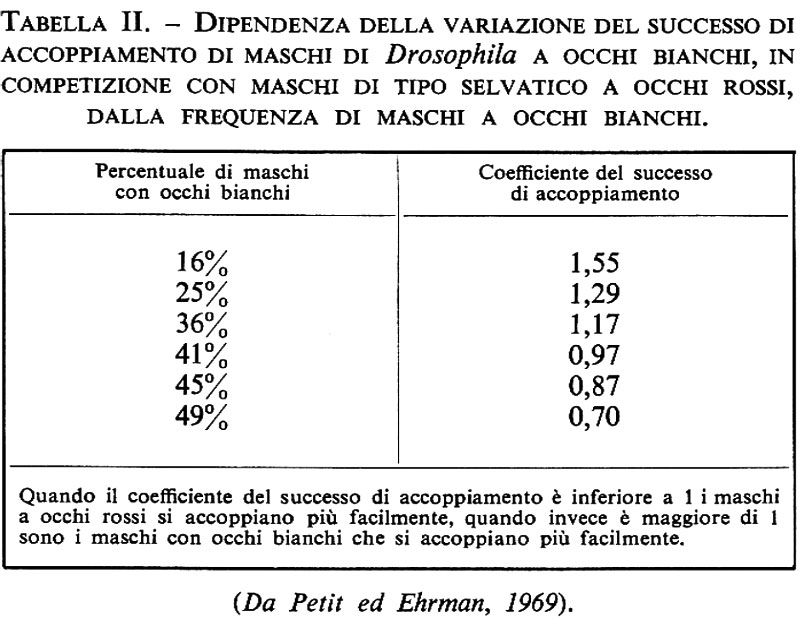
Si conoscono numerosi casi in cui le idoneità biologiche relative di geni allelici dipendono dalla frequenza. Una dipendenza dalla frequenza è quasi certamente presente in situazioni di selezione diversificante, che prenderemo in esame in seguito. Un caso singolare di dipendenza dalla frequenza è quello denominato ‛effetto del tipo raro', scoperto in Francia da Cl. Petit e studiato successivamente da L. Ehrman in America, riguardante il successo relativo dell'accoppiamento di moscerini con genotipi diversi: per alcune differenze geniche è stato chiaramente dimostrato che l'allele più raro è preferito nell'accoppiamento, come risulta dalla tab. II. Come abbiamo già accennato, si vede chiaramente che un effetto di questo tipo dovrebbe contribuire a mantenere un equilibrio intermedio stabile delle frequenze geniche per i loci interessati.
Tabella 2
6. La selezione in condizioni multigeniche.
Possiamo definire un organismo come un insieme di parti correlate tra loro, i cui attributi sono interdipendenti: è un modo diverso per dire che gli organismi sono organizzati. Proprio in considerazione di tale organizzazione si deve ritenere poco probabile che le differenze che si riscontrano in natura possano essere ricondotte a una singola coppia di geni, i cui effetti sull'idoneità biologica siano completamente indipendenti da ciò che accade in tutti gli altri loci genici. La vera situazione in natura dev'essere perciò estremamente complessa. I modelli teorici che prendono in esame e studiano le conseguenze prevedibili nell'attribuire idoneità biologiche diverse a determinate coppie di geni sono estremamente utili, ma non possono sostituire l'osservazione di situazioni reali, in cui molti geni interagiscono nei loro effetti sull'idoneità biologica. Inoltre molte variabili che è possibile studiare, come l'abilità nell'eseguire test di intelligenza, nell'uomo, o la statura, sempre nell'uomo, il numero di setole in una zona particolare della superficie esterna, in Drosophila, la quantità nei raccolti, la fertilità nelle piante o negli animali, sono caratteri che possono essere influenzati da molti geni. Caratteri di questo tipo possono essere ovviamente di notevole importanza per determinare l'idoneità biologica di un individuo o per l'applicazione pratica negli allevamenti.
Tutti questi caratteri presentano ciò che noi chiamiamo variazione continua: sono molto importanti nella genetica applicata e devono essere di grande rilievo nell'evoluzione, dal momento che la teoria dell'evoluzione, così come è stata elaborata da Darwin, sostiene di necessità che la maggior parte delle modificazioni evolutive sono relativamente lente e comportano variazioni minime delle caratteristiche degli organismi piuttosto che cambiamenti drastici. Ciò è ovvio, poiché l'organismo è così integrato che un improvviso e drastico cambiamento di una qualsiasi delle sue caratteristiche dovrebbe necessariamente rompere tale integrazione e determinare una riduzione di idoneità biologica. D'altro canto è possibile che piccole modificazioni di un carattere determinino un piccolo aumento dell'idoneità biologica e questo può andare ad aggiungersi ad altri caratteri, o determinarne modificazioni, così che, per esempio, un organo come l'occhio può evolversi come una unità funzionante. Invece una modificazione considerevole di qualsiasi parte dell'occhio sarebbe chiaramente causa di difficoltà: per esempio un grosso cambiamento nel solo cristallino renderebbe impossibile mettere a fuoco un'immagine sulla retina, mentre piccoli cambiamenti del cristallino possono migliorare l'occhio e successivi piccoli cambiamenti della forma della retina, in rapporto alla nuova forma del cristallino, possono ulteriormente perfezionarlo. Quindi cambiamenti minimi ma costanti di tutte le diverse parti, se i geni le cui frequenze stanno cambiando hanno effetti molti piccoli, possono contribuire all'evoluzione di un organo o di un organismo in modo integrato. É perciò ovvio, come è stato detto prima, che i vari stadi evolutivi attraversati da un organismo dall'inizio della sua esistenza devono aver corrisposto ad altrettanti organismi così ben organizzati da potersi riprodurre e lasciare una discendenza.
Prima di esaminare le modalità dell'azione della selezione sui diversi caratteri degli organismi, dobbiamo precisare in che modo descriverli. La descrizione è fatta statisticamente e puo essere illustrata con una distribuzione di frequenze, come nella fig. 6. Se ora prendiamo in considerazione una curva di distribuzione come quella della figura, possiamo vedere che la selezione può agire in vari modi: tutti comunque implicano idoneità biologiche diverse per le diverse zone della curva di distribuzione. In effetti prenderemo in considerazione tre tipi di selezione in relazione a caratteri con variabilità continua: selezione direttiva, selezione stabilizzante e selezione diversificante.
Selezione direttiva. Questo tipo di selezione si manifesta se gli individui che nel grafico occupano una delle zone laterali sono più adatti degli individui della zona opposta, cioè contribuiscono più di questi alla generazione successiva. A determinare la selezione direttiva possono concorrere vari tipi di relazione tra il valore del carattere e l'idoneità biologica: può ad esempio accadere che l'idoneità cresca regolarmente con il valore del carattere; o invece che solo gli individui che sono al di sopra di un particolare valore della distribuzione siano del tutto idonei, nel qual caso si parla di selezione troncante. Il troncamento è comune negli esperimenti di selezione artificiale, quando lo sperimentatore decide di allevare solo, ad esempio, il 5o% o il 100% della parte superiore della distribuzione, imponendo così una forte selezione in favore di un cambiamento della media. La fig. 6 si riferisce a un esempio di selezione di questo tipo e dimostra come la distribuzione del carattere nella popolazione vada modificandosi con il passare delle generazioni.
Selezione stabilizzante. Si ha selezione stabilizzante quando in vicinanza della media della popolazione vi è il massimo di idoneità biologica, che si riduce invece con l'allontanamento da questo valore medio, in ambedue le direzioni. Nella fig. 7 è riportato come esempio un esperimento di selezione stabilizzante condotto su una popolazione di Drosophila.
Selezione diversificante. Si dice infine che vi è selezione diversificante quando i massimi di idoneità biologica sono più di uno e il valore medio non coincide con un carattere ottimale; in questo modo ambedue le estremità della distribuzione contengono individui più idonei di quelli che si trovano nella zona centrale. Questa situazione è analoga a quella della selezione da nicchia che abbiamo esaminato in precedenza, in cui più di un genotipo è favorito perché vi è più di un ambiente e i genotipi intermedi non si adattano bene a nessun ambiente. La fig. 8 mostra i risultati di una selezione diversificante condotta per due generazioni; è opportuno confrontarla con le figure che illustrano gli effetti della selezione stabilizzante e della selezione direttiva.
a) Selezione direttiva e variabili continue.
Sono stati eseguiti numerosissimi esperimenti di selezione artificiale per modificare le caratteristiche di popolazioni sia per fini applicativi, sia per ricerca pura; quest'ultima è stata in gran parte condotta su Drosophila melanogaster e ha consentito di trarre alcune conclusioni di valore generale. Innanzitutto, se imponiamo una selezione direttiva, in genere le popolazioni rispondono positivamente, e ciò significa che vi è di solito una variabilità genetica nell'ambito della quale la selezione può operare le sue scelte. In secondo luogo è raro che la risposta sia tale che la progenie degli individui estremi selezionati sia anch'essa lontana dalla media allo stesso grado dei genitori. Ciò accade perché parte della varietà della popolazione deriva da una variazione dei fattori ambientali e non da una variazione genetica, onde è possibile stabilire il concetto dell'ereditabilità di una tale variabile continua, corrispondente a una misura che indica quale quota della varietà presentata da una popolazione consegue a varietà genetica. Gli stessi esperimenti di selezione consentono di calcolare l'ereditabilità, della quale sono state proposte anche numerose altre modalità di misurazione, più elaborate, ma ognuna con i propri punti deboli. Un'analisi completa di questi e di altri problemi analoghi è stata compiuta da Falconer (v., 1960), cui si rimanda per maggiori dettagli. Vari tipi di risposta alla selezione direttiva sono presentati nella fig. 9: a rappresenta una risposta costante che diminuisce di velocità fino a che la popolazione raggiunge un livello al quale la risposta cessa; b rappresenta una risposta con un inizio analogo cui segue un periodo di rapida accelerazione e quindi un plateau; in c si ha un risultato piuttosto comune: il tentativo di mantenere la risposta di partenza determina l'estinzione della popolazione. Quest'ultima risposta è una forma particolare di una conseguenza pressoché generale della selezione artificiale: nel rispondere alla selezione la popolazione diviene sempre meno vitale, sempre meno fertile e trova sempre maggiore difficoltà a conservarsi. Alcune popolazioni si riprendono da questa riduzione di vitalità, altre invece non vi riescono. Vi sono state molte discussioni sulle ragioni di tale riduzione di vitalità: essa può essere conseguenza degli effetti dannosi degli stessi geni responsabili della modificazione del carattere sottoposto a selezione, oppure dei cambiamenti della frequenza di altri geni legati a quelli direttamente sottoposti a selezione per associazione genetica a livello cromosomico. Ambedue le spiegazioni sono probabilmente vere, anche se è stato chiaramente dimostrato che quella basata sull'associazione è in alcuni casi la più valida. Ovviamente questa non è comunque la sola ragione per cui una linea dovrebbe cessare di rispondere alla selezione: le linee possono trovarsi in tale condizione senza aver raggiunto un basso livello di vitalità. Ciò accadrebbe se tutti i geni le cui frequenze possono essere aumentate dalla selezione fossero ‛giunti alla fissazione', cioè fossero divenuti omozigoti, dopo l'eliminazione dalla popolazione di geni allelici che operano in senso opposto. Che questa sia talvolta la spiegazione è chiaramente dimostrato dall'incrocio di linee selezionate diverse (v. fig. 9, d): si ottengono in questo modo linee ibride che rispondono alla selezione meglio di quanto facevano ambedue le linee parentali.
L'associazione tra geni che influenzano direttamente il carattere sottoposto a selezione è un altro elemento importante: in tal modo infatti è spesso possibile spiegare le apparenti novità prodotte dalla selezione e i casi di risposte accelerate alla selezione stessa (v. fig. 10).
Sono stati effettuati numerosi tentativi per dimostrare con esperimenti di selezione l'ipotesi genetico-quantitativa, con qualche limitato successo. In generale le misure di ereditabilità e le previsioni sulla risposta che daranno una, due o tre generazioni di selezione si accordano abbastanza bene con i risultati, ma la teoria ha i suoi limiti in quanto, per renderla matematicamente trattabile, si sono dovute prefissare alcune condizioni, il che ha senza dubbio dato origine a qualche difficoltà. In particolare si assume di solito che i geni che influenzano il carattere da selezionare non siano associati sullo stesso cromosoma, onde a lungo andare le previsioni possono risultarne viziate.
Un risultato consueto, anche se non universale, fornito da esperimenti di selezione artificiale, mostra la tendenza di un determinato carattere, spinto dalla selezione in una certa direzione, a tornare verso i suoi valori originali se la spinta selettiva viene rimossa; esso indica che la selezione naturale sta agendo sul carattere stesso in modo opposto alla selezione artificiale. Queste osservazioni, se appaiono in accordo con quelle concernenti gli effetti della selezione sulla vitalità e la fertilità degli organismi, sono anche indicative della tendenza manifestata dalla selezione naturale a mantenere la media dei caratteri in questione al valore osservato nella popolazione di partenza. Quanto finora è stato detto ci induce a prendere in esame la selezione stabilizzante.
b) Selezione stabilizzante.
In generale è stato dimostrato che la selezione stabilizzante, con il passare delle generazioni, riduce la varietà della popolazione. Ciò può accadere in due modi, ed è naturalmente possibile che ambedue siano cpntemporaneamente attivi nella popolazione: nel primo caso la selezione può ridurre la quota di varietà genetica di una popolazione; nel secondo possiamo immaginare che la selezione stabilizzante operi in modo che gli individui di una popolazione giungano a possedere meccanismi di sviluppo talmente organizzati da essere sottoposti all'influenza delle variabili ambientali in misura minore rispetto alla popolazione originaria.
È noto che la selezione stabilizzante agisce in natura in vari casi: uno dei più rilevanti è quello illustrato nella fig. 11, ricavata dai risultati di M. N. Karn e L.S. Penrose (v., 1952), la quale mostra che la selezione stabilizzante agisce nell'uomo in funzione del peso alla nascita.
Il concetto di selezione stabilizzante è molto importante: le prime ipotesi sulla selezione, nell'ambito della teoria dell'evoluzione, prendevano per lo più in considerazione modificazioni nel tempo e quindi la teoria della selezione naturale e quella dell'evoluzione per selezione naturale erano discusse prevalentemente in termini che noi oggi definiremmo di selezione direttiva. Tuttavia l'acquisizione che la mutazione è costantemente in grado di cambiare le caratteristiche dei geni di una popolazione, consentì di comprendere che la selezione non ha come solo scopo quello di modificare le caratteristiche di una popolazione, ma anche quello di mantenere costanti tali caratteristiche, se non è richiesto un cambiamento. Secondo K. Mather la selezione stabilizzante in una situazione multigenica può determinare in una popolazione un'organizzazione genetica particolare, che presenta vantaggi notevoli. Un esempio di questo tipo di organizzazione si può avere considerando due loci genici di uno stesso cromosoma, ciascuno dei quali abbia due geni alternativi che possiamo denominare + e -, indicando con tali segni la direzione in cui i due geni tendono a spingere il carattere sottoposto a selezione stabilizzante. Se consideriamo ad esempio la statura umana, possiamo supporre che per un locus si possano avere tre genotipi, +/+, +/−, −,−, ai quali corrispondono tre diverse stature, + una unità, zero, − una unità, e che anche per l'altro locus si possano avere due geni con effetti identici. In complesso esistono i seguenti genotipi diversi:
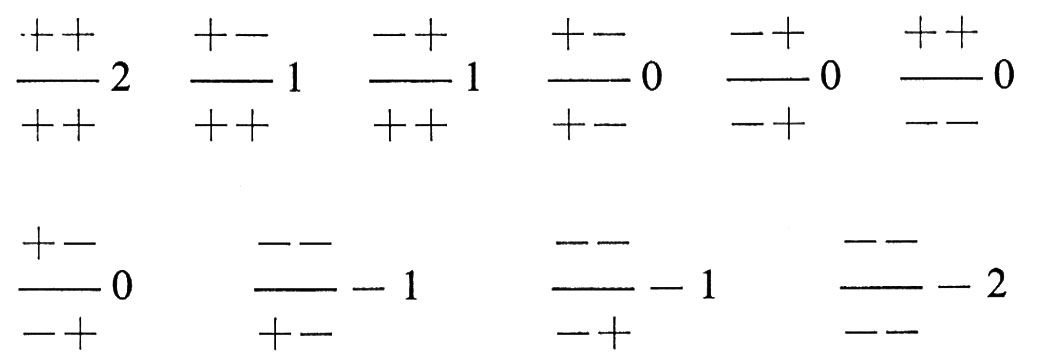
Le cifre accanto ai genotipi indicano le deviazioni della statura dei vari genotipi dalla statura ottimale della popolazione, che viene indicata con O. Perciò i genotipi con deviazioni negative danno origine a individui più bassi, quelli con deviazioni positive a individui più alti del valore ottimale. È evidente che vi sono vari genotipi che determinano una statura ottimale:

Se i vari genotipi presenti nella popolazione si accoppiano a caso, potranno talvolta accoppiarsi fra loro anche i genotipi
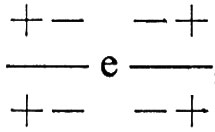
, dando origine a una progenie con genotipo
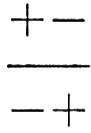
Questo particolare tipo di eterozigote,
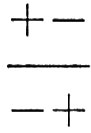
, se i due loci sono situati in stretta vicinanza nello stesso cromosoma, produrrà uova o spermi i cui genotipi saranno nella grande maggioranza dei casi + - o - +; questi, accoppiati con spermi e uova simili provenienti da altri individui, daranno ovviamente luogo a genotipi
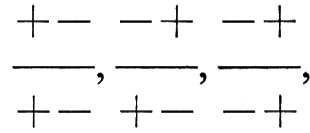
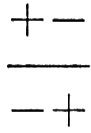
i quali sono tutti del tipo ottimale. Si prenda ora in considerazione un altro genotipo,
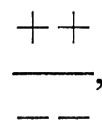
il quale determina anch'esso un fenotipo ottimale: questo produce gameti che sono per metà ++ e per metà − −, i quali accoppiati con uova e spermi + - o - + danno luogo a individui che deviano dalla statura ottimale. Quindi il genotipo
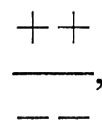
sebbene presenti un fenotipo ottimale, genera una progenie che non è ottimale, mentre il genotipo
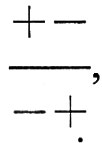
sebbene contenga i medesimi geni, genera una progenie per la maggior parte ottimale. In sostanza il fatto che i geni siano associati sui cromosomi può influire sul grado di stabilizzazione di una popolazione, se è in azione una selezione stabilizzante; in realtà si può ipotizzare che tale selezione elimini praticamente tutti i cromosomi + + e - -, lasciando solo i tre genotipi
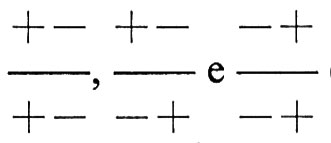
come caratteristici della popolazione. Tale popolazione tenderà comunque a generare una certa varietà genetica, perché, durante la formazione di uova e spermi, un individuo con genotipo
, mediante il fenomeno del crossing-over, produrrà una piccola frazione di uova e spermi che sono + + o − −. L'entità di questa frazione è determinata dal grado di associazione dei due geni, di quanto cioè sono collocati vicini sul cromosoma: se lo sono molto, tale frazione è piccola. Il mantenimento di un tale sistema genetico bilanciato consente alla popolazione di conservare una certa flessibilità genetica, nonostante questo costi la produzione per crossing-over di una piccola frazione di rari genotipi che sono relativamente poco idonei biologicamente. Ciò garantisce, comunque, che qualora in tempi successivi le circostanze ambientali cambino tanto che la statura precedentemente ottimale cessi di essere tale, la popolazione è ancora in grado di evolvere verso un nuovo optimum. Questi sistemi poligenici bilanciati, come li ha chiamati Mather, costituiscono quindi un compromesso fra la necessità immediata o a breve termine di mantenere stabile l'eredità e quella a lungo termine di conservare la capacità di attuare cambiamenti. La nostra convinzione che sistemi di questo tipo siano effettivamente operanti nelle popolazioni naturali è confortata dall'osservazione generale che campioni di popolazioni in condizioni di laboratorio rimangono abbastanza stabili se non sono esposti a selezione, ma sono, ciò nonostante, molto sensibili alla selezione artificiale. È stato poi dimostrato, esaminando le popolazioni che si ottengono dopo selezione, che la ricombinazione del tipo descritto nella fig. 10 si determina in risposta alla selezione. Test specifici dimostrano poi che la ricombinazione anche quando avvenga tra coppie di cromosomi presi dalla medesima popolazione naturale può generare varietà. Tutto ciò sta in sostanza a dimostrare che combinazioni bilanciate di cromosomi eterozigoti devono essere molto comuni nelle popolazioni naturali. Si ritiene che una delle cause di questa situazione sia la selezione stabilizzante, sebbene si discuta molto su quanta parte della eterozigosi naturale debba esserle attribuita.
c) Selezione diversificante.
Un esempio delle conseguenze immediate dell'esposizione di una popolazione a selezione diversificante è già stato fornito nella fig. 8. Numerosi esperimenti di selezione diversificante, condotti utilizzando come carattere per lo più il numero di setole in Drosophila o, talvolta, caratteristiche comportamentali, o la stagione della fioritura nelle piante, hanno dimostrato che la selezione diversificante può: 1) aumentare la variabilità di una popolazione (v. fig. 7); 2) mantenere stabili i polimorlismi (v. fig. 12); 3) produrre e mantenere differenze fra due popolazioni fra le quali vi sia un alto livello di scambio di geni (v. fig. 13); 4) dividere una popolazione in due parti fra le quali vi sia un notevole isolamento riproduttivo (v. fig. 14).
Tre di questi effetti sono risultati di notevole interesse teorico. Innanzitutto, la scoperta che la selezione diversificante può mantenere stabile un polimorfismo ha reso ancora più complessa la teoria sui polimorfismi, anche se in realtà ha reso più facile, in linea di principio, spiegare molte situazioni polimorfiche naturali. Infatti in precedenza si riteneva che il mantenimento di un polimorfismo stabile dovesse dipendere dal vantaggio dell'eterozigote, cioè dal fatto che questo avesse un'idoneità biologica maggiore di ambedue gli omozigoti. La selezione diversificante, con la sua capacità di creare e mantenere un polimorfismo, ha chiaramente dimostrato che questo non è l'unico meccanismo possibile.
La seconda conseguenza importante degli esperimenti di selezione diversificante è stata la dimostrazione che due sottogruppi di una popolazione possono divergere l'uno dall'altro, se sottoposti a selezione in direzioni diverse, anche se in ogni generazione essi si scambiano una buona parte dei loro geni. In precedenza si riteneva invece generalmente che un intenso scambio reciproco di geni impedisse a due sottopopolazioni di diversificarsi, perché la migrazione genica riusciva a vanificare la divergenza eventualmente prodotta dalla selezione. Ora ovviamente dobbiamo considerare non solo il grado di migrazione genica tra le due sottopopolazioni, ma anche l'intensità della selezione che tende a far divergere le popolazioni: vi dev'essere un punto di equilibrio da un lato del quale la selezione è, rispetto alla migrazione, più efficace, dall'altro meno. Nel primo caso le due popolazioni tendono a divergere, mentre nel secondo tendono a convergere. In questo contesto va considerato anche che la migrazione di geni da una popolazione a un'altra non è necessariamente casuale. Possiamo ad esempio immaginare una situazione nella quale una popolazione può perdere per migrazione una forma allelica più dell'altra, ad esempio più A che a, mentre un'altra popolazione, adiacente alla prima, può perdere più a che A in quanto le condizioni in cui si trovano le due popolazioni sono tali che, mentre gli individui con genotipo AA della prima popolazione trovano congeniale una componente ambientale, gli individui con genotipo aa della seconda trovano invece adatta una diversa zona dell'ambiente. È quindi ovvio che una popolazione perde geni A per guadagnare a con la migrazione, mentre l'opposto accade nell'altra popolazione; vi sarà quindi ‛migrazione selettiva'. Questa non interessa necessariamente qualsiasi selezione naturale che agisca contemporaneamente su due popolazioni, ma può comunque essere in linea di principio una potente sorgente di divergenza genetica fra due popolazioni. Thoday e Gibson (v., Environmental ..., 1970, p. 990) hanno dimostrato sperimentalmente in una situazione campione che questa migrazione selettiva può far divergere due sottopopolazioni e hanno sottolineato come il loro esperimento ricordi un aspetto delle popolazioni umane. Possiamo infatti considerare le classi sociali come sottopopolazioni distinte per quanto riguarda qualsiasi caratteristica geneticamente determinata che promuova la mobilità sociale: è noto che coloro che posseggono un quoziente di intelligenza maggiore tendono a salire lungo la scala sociale, mentre coloro che l'hanno più basso tendono a discendere. Dal momento che le variazioni del quoziente intellettivo sono in gran parte determinate geneticamente, i geni implicati mostreranno una migrazione selettiva tra le classi: quindi, secondo una certa previsione teorica, le classi possono divergere come risultato della migrazione selettiva.
La dimostrazione che la selezione diversificante può far divergere sottopopolazioni nonostante vi sia fra loro un notevole scambio di geni ha contribuito a chiarire alcuni problemi che riguardano popolazioni naturali e che sono stati oggetto di studio da parte di genetisti ecologi. A. D. Bradshaw ad esempio ha dimostrato che l'erba Agrostis tenuis, a impollinazione anemofila, ha selezionato genotipi resistenti al piombo sulla superficie di miniere di piombo abbandonate. Finché non vennero compiuti esperimenti di selezione diversificante la teoria esistente non consentiva di spiegare questo fenomeno, poiché l'impollinazione anemofila e le estesissime popolazioni lontane dalle miniere e sensibili al piombo avrebbero dovuto provocare una notevole migrazione di geni entro l'area delle miniere stesse impedendo, secondo le possibili previsioni, a genotipi speciali di stabilirsi nelle aree minerarie. Si è ora dimostrato chiaramente che in questo fenomeno è implicata un'intensissima selezione diversificante.
Il terzo e più controverso risultato degli esperimenti di selezione diversificante riguarda la divisione di una popolazione in due sottopopolazioni tra le quali lo scambio di geni sia limitato o addirittura assente: questa situazione corrisponde in natura a quella descritta come l'origine delle specie. Classicamente (v. Mayr, 1940) l'unica ipotesi che si riteneva valida sull'evoluzione delle specie era la seguente: due popolazioni della stessa specie, isolate da qualche barriera geografica quale una catena montagnosa o un gran fiume o una barriera di ghiacci durante le glaciazioni, davano origine a due popolazioni fra le quali non vi era alcuno scambio di geni. In queste condizioni di isolamento le due popolazioni avrebbero dovuto divergere perché esposte alla selezione in ambienti diversi, e tale divergenza poteva proseguire fintanto che le loro costituzioni genetiche divenissero tali che, in caso di ibridazione, gli ibridi sarebbero risultati biologicamente poco idonei. Esse potevano addirittura giungere a uno stadio nel quale non era loro possibile riconoscersi per incrociarsi e dar luogo a ibridi. Perciò, anche se la barriera che le divideva fosse caduta, ad esempio alla fine della glaciazione, con il ritirarsi dei ghiacciai, le due popolazioni avrebbero potuto invadere le reciproche aree senza incrociarsi tra loro, dimostrando così di essere due specie chiaramente distinte. Le dimostrazioni che provengono dalla sistematica e dalla geografia delle piante e degli animali illustrano chiaramente che in gran parte l'origine delle specie è avvenuta in questo modo, vale a dire allopatricamente. È comunque rimasto a lungo controverso se la genesi di specie diverse possa avvenire anche con altre modalità.
Una di queste modalità, in realtà di tipo semiallopatrico, è stata proposta da Dobzhansky, il quale ha formulato l'ipotesi che talvolta, quando la barriera viene a cadere, le due popolazioni situate ai suoi due lati possano non aver raggiunto un grado di divergenza sufficiente a impedirne l'incrocio quando esse si incontrano nuovamente. Tuttavia queste popolazioni possono essere divenute sufficientemente divergenti perché gli ibridi che ne risultano siano meno biologicamente idonei dei non ibridi. La selezione può quindi intervenire guidando le scelte nell'accoppiamento, poiché gli individui che generano ibridi disperdono parte del loro sforzo riproduttivo nel produrre una progenie meno idonea e conseguentemente quelli che non formano ibridi contribuiscono alle generazioni successive con una progenie più numerosa. Perciò, se vi è una qualche variazione genetica che influenza la scelta nell'accoppiamento o il meccanismo di impollinazione delle piante, sarà utilizzata dalla selezione: in tal modo le due popolazioni, che erano divenute divergenti secondo una modalità allopatrica, dopo essersi nuovamente incontrate formeranno una barriera specifica simpatricamente.
Il terzo possibile meccanismo di speciazione è invece simpatrico. In questo caso si suppone che, in un ambiente eterogeneo, due parti di una popolazione simpatrica divengano selezionate per nicchie ecologiche diverse all'interno della stessa regione e in tal modo divergano: tale divergenza perdura fino a quando gli ibridi nati da genitori appartenenti a nicchie diverse divengono scarsamente adatti ad ambedue le nicchie e quindi biologicamente meno idonei dei non ibridi. A questo punto, come ipotizzato dalla teoria di Dobzhansky, può iniziare a divenire efficace la selezione a favore di barriere fra gli accoppiamenti. La differenza fra questo ultimo meccanismo e quello della vera speciazione simpatrica sta semplicemente nel fatto che nel meccanismo proposto da Dobzhansky la divergenza iniziale ha luogo in una condizione allopatrica.
È ovvio che gli esperimenti di selezione artificiale non sono in grado di dimostrare se in natura è stato operante o no uno dei vari meccanismi, ma possono dimostrare che la selezione è in grado di produrre un certo tipo di conseguenze e inoltre che le popolazioni sottoposte a selezione posseggono la capacità genetica di rispondere in un certo modo. In particolare gli esperimenti di selezione artificiale, condotti con l'impiego della selezione diversificante, hanno chiaramente dimostrato che una popolazione può divergere anche se vi è un'intensa ibridazione tra le due sottopopolazioni: tali esperimenti dimostrano così che, mentre deve ritenersi senz'altro errata l'ipotesi originale che sosteneva l'impossibilità di una divergenza in assenza di isolamento, la speciazione simpatrica è invece in linea di principio possibile, malgrado non si riesca in realtà a dimostrarne l'effettivo verificarsi in natura. L'esperimento di Thoday e Gibson (v. fig. 14) è stato compiuto proprio per verificare più a fondo questa possibilità: è diverso da precedenti esperimenti sulla selezione diversificante in quanto, dopo la selezione di due classi, ai moscerini è stata lasciata libertà di scelta nell'accoppiamento e nell'incrocio. Il risultato è stato una rapida divergenza con una rapida riduzione della frequenza di incroci ibridi, sebbene tutti i moscerini si accoppiassero in un piccolo ambiente. Numerose ripetizioni di questo esperimento, con l'impiego di altri ceppi, non hanno però dato gli stessi risultati, anche se diverse sono le ragioni di questa discrepanza (v. Thoday e Gibson, The probability..., 1970). Occorre qui mettere in rilievo che la dimostrazione che la selezione artificiale di un certo tipo può produrre un certo risultato, anche una sola volta, è una prova sufficiente che il risultato stesso è una possibile conseguenza di questo tipo di selezione. Non ci si può attendere che la medesima conseguenza si abbia con altre popolazioni, dal momento che queste possono non possedere la stessa varietà genetica di quelle usate negli esperimenti riusciti positivamente. Ciò che ignoriamo è quanto spesso potremmo ottenere tale risultato se potessimo ripetere numerossissime volte questo esperimento, utilizzando ogni volta popolazioni diverse. Comunque, Beardmore e Baldawi (v. Thoday, 1972) hanno ottenuto risultati assolutamente identici a quelli di Thoday e Gibson, utilizzando una popolazione diversa: essi hanno così confermato che la speciazione simpatrica è una possibilità che non dev'essere trascurata da coloro che studiano l'evoluzione.
Bibliografia.
Beardmore, J. A., Developmental stability in constant and fluctuating temperatures, in ‟Heredity", 1960, XIV, pp. 411-422.
Dobzhansky, Th., A directional change in the genetic constitution of a natural population of Drosophila pseudoobscura, in ‟Heredity", 1947, I, pp. 53-64.
Dobzhansky, Th., Genetics and the origin of species, New York 1951.
Falconer, D. S., Introduction to quantitative genetics, New York 1960.
Karn, M. N., Penrose, L. S., Birth weight and gestation time in relation to maternal age, parity and infant survival, in ‟Annals of eugenics", 1952, XVI, pp. 147-164.
Kettlewell, H. B. D., Selection experiments on industrial melanism in the Lepidoptera, in ‟Heredity", 1955, IX, pp. 323-342.
Kettlewell, H. B. D., Further selection experiments on industrial melanism in the Lepidoptera, in ‟Heredity", 1956, X, pp. 287-301.
Kettlewell, H. B. D., Survey of the frequencies of Biston betularia L. (Lep.) and its melanic forms in Great Britain, in ‟Heredity", 1958, XII, pp. 51-72.
L'Héritier, Ph., Teissier, G., Étude d'une population de Drosophiles en équilibre, in ‟Compte rendu de l'Académie des Sciences", 1933, CXCVII, pp. 1765-1767.
L'Héritier, Ph., Teissier, G., Élimination des formes mutantes dans les populations de Drosophiles. Un cas des Drosophiles ‛Bar', in ‟Compte rendu de la Société de Biologie", 1937, CXXIV, pp. 880-882.
Mather, K., Human diversity, Edinburgh 1964.
Mayr, E., Systematics and the origin of species, New York 1940.
Merrel, D. J., The modern theory of evolution: evolution and genetics, New York 1962.
Millicent, E., Thoday, J. M., Effects of disruptive selection. IV. Geneflow and divergence, in ‟Heredity", 1961, XVI, pp. 199-217.
Petit, Cl., Ehrman, L., Sexual selection in Drosophila, in ‟Evolutionary biology", 1969, III, pp. 177-223.
Thoday, J. M., Disruptive selection, in ‟Proceedings of the Royal Society of Biology", 1972, CLXXXII, pp. 109-254.
Thoday, J. M., Gibson, J. B., Environmental and genetical contributions to class difference: a model experiment, in ‟Science", 1970, CLXVII, pp. 990-992.
Thoday, J. M., Gibson, J. B., The probability of isolation by disruptive selection, in ‟American naturalist", 1970, CIV, pp. 219-230.
Timoféeff-Ressosvsky, N. W., Mutations and geographical variation, in The new systematics (a cura di J. Huxley), Oxford 1940, pp. 73-136.
Tinbergen, N., Behaviour, systematics and natural selection, in ‟Ibis", 1959, CI, pp. 318-330.