
Simmetrie in biologia
Simmetrie in biologia
di Mario Ageno
SOMMARIO: 1. Introduzione. □ 2. Simmetrie geometriche nel mondo dei viventi: a) animali (Metazoi); b) piante (metafiti); c) funghi; d) protisti (Eucarioti unicellulari); e) Procarioti; f) Virus. □ 3. Simmetrie funzionali. □ 4. Struttura fisica e adattamenti evolutivi. □ 5. Le simmetrie assenti (asimmetrie): a) la chiralità molecolare della biosfera; b) altri esempi. □ Bibliografia.
1. Introduzione.
Il concetto di simmetria che trova impiego in campo biologico è, per ora almeno, quasi esclusivamente quello classico. Ogni simmetria viene pertanto caratterizzata mediante un certo numero di elementi geometrici - piani, assi, centro - che definiscono le operazioni rispetto alle quali una certa ‛forma' biologica rimane invariata. Non mancano tuttavia casi in cui questo concetto elementare viene generalizzato. Si riconoscono simmetrie nella dimensione temporale e si parla anche di simmetria per operazioni di ricoprimento in cui uno spostamento rigido è combinato con una legge di crescita o di sviluppo. Un esempio ben noto è offerto dalla conchiglia di Nautilus, che si sviluppa parallelamente a un piano, attorno a un centro, secondo una spirale logaritmica.
Mentre le simmetrie che conservano le distanze possono esser tutte facilmente descritte e catalogate, più difficile si presenta il problema della teorizzazione di queste simmetrie generalizzate. Tutto dipende da quali tipi di leggi di sviluppo, o anche semplicemente di corrispondenza, siamo disposti ad accettare come base di una simmetria. Al limite, ogni legge che fissi quantitativamente come sono disposte nello spazio le une rispetto alle altre o come crescono nel tempo le varie parti di un organismo (e le parti di ciascuna parte) può rientrare nel concetto generalizzato di simmetria. In tal caso, peraltro, tale concetto viene a saldarsi con una particolare accezione del concetto più generale di ordine: quella secondo cui la parola ‛ordine' è portatrice di valore, in quanto implica appunto l'esistenza di una legge (e quindi di una simmetria).
In pratica, lo studio delle simmetrie in biologia trova ben presto un ostacolo nella difficoltà di identificare relazioni e leggi rigorosamente quantitative in sistemi estremamente complessi, e soprattutto di cogliere gli invarianti essenziali di forme che pur si presentano come caratteristiche. La nostra ancor grande ignoranza dei meccanismi dello sviluppo embrionale e della differenziazione, in una parola la nostra limitata conoscenza dei viventi, non ci consente inoltre di capire, il più delle volte, l'origine e la natura delle simmetrie riconosciute. Al posto di una soddisfacente teoria generale, dobbiamo quindi contentarci, per ora, di un elenco delle simmetrie più evidenti, procedendo in modo quasi esclusivamente empirico.
Possiamo tuttavia fare subito esplicitamente alcune osservazioni di un certo rilievo. In primo luogo, occorre porre l'accento sul fatto che il carattere essenzialmente irreversibile, in particolare evolutivo, dei processi biologici non impedisce che esistano simmetrie biologiche semplici e generalizzate: in particolare coinvolgenti leggi di sviluppo. Rientrano per esempio nel quadro lo sviluppo esponenziale di una coltura batterica in un mezzo liquido omogeneo, nonché le relazioni tra crescita e forma e le relazioni tra forme corrispondenti, messe in evidenza più di mezzo secolo fa da D'Arcy Thompson (v., 1917) in un libro giustamente famoso. Su di esse tuttavia ancor oggi sappiamo dire ben poco. Si consideri, ad esempio, la legge di sviluppo di una foglia, la cui forma è specie-specifica: essa è sicuramente vincolata da condizioni d'invarianza quantitativamente ben definite, ma assai difficili da individuare.
Una seconda osservazione importante che possiamo fare fin d'ora è che le simmetrie biologiche non sono quasi mai perfette. Basta considerare, tanto per fare un esempio, come sia solo apparente la simmetria bilaterale nell'uomo, e ciò non solo per quanto riguarda la posizione reciproca di numerosi organi interni, ma anche e soprattutto per la struttura e la funzionalità del cervello. Solo in tempi recenti misure molto accurate hanno messo definitivamente in evidenza differenze sistematiche di volume e di forma tra l'emisfero destro e il sinistro. Si sa tuttavia fin dalla metà del secolo scorso che esiste una differenza funzionale importante tra l'emisfero sinistro, che svolge un ruolo dominante per tutto ciò che riguarda le funzioni linguistiche, e l'emisfero destro, che prevale nelle funzioni percettive e nell'analisi di eventi non legati al linguaggio (v. emisferi cerebrali: Dominanza cerebrale).
Non sembra tuttavia che la violazione o rottura parziale delle simmetrie biologiche possa essere facilmente ricondotta a quella stessa spiegazione che vale per la rottura delle simmetrie in fisica. Qui i vari tipi di interazione tra particelle sono caratterizzati essenzialmente dall'ordine di grandezza delle energie in gioco. Le interazioni più energiche hanno spesso caratteri di simmetria che non sono posseduti da interazioni meno energiche. Quando interazioni dei due tipi sono contemporaneamente presenti, le simmetrie dipendenti dalle interazioni più energiche sussistono come simmetrie approssimate, delle quali le conseguenze delle interazioni meno energiche rappresentano soltanto più o meno piccole violazioni. Questo tipo di discorso certamente non vale come spiegazione della rottura delle simmetrie biologiche: qui le interazioni tra le particelle costituenti sono sempre e solo interazioni elettromagnetiche, e non sembra neppure che le simmetrie biologiche possano essere in modo semplice e diretto ricondotte a esse.
È importante osservare a questo proposito che le simmetrie biologiche si manifestano essenzialmente a livello macroscopico. I fenomeni biologici elementari (quelli di cui si occupa la biologia molecolare, per intenderci) coinvolgono sistematicamente oggetti le cui dimensioni li pongono all'interfaccia tra il campo quantistico, ove dominano le conseguenze del principio di indeterminazione, e il campo classico, ove la legge dei grandi numeri rende tutti i processi descrivibili mediante teorie deterministiche. È il ‛campo delle dimensioni intermedie', in cui le macromolecole biologicamente significative si comportano di regola più o meno come bacchette o granuli in sospensione nell'acqua, in modo dunque ormai descrivibile con ottima approssimazione mediante la meccanica classica, mentre solo nei processi chiave queste bacchette o granuli danno luogo alla ristrutturazione di sistemi di legami covalenti, a deformazioni nella distribuzione di carica elettronica, al passaggio di elettroni o protoni attraverso barriere di potenziale, in una parola a fenomeni che la fisica classica non è in grado di descrivere.
Le simmetrie biologiche si manifestano di regola solo a livelli organizzativi superiori e quindi in pieno campo classico: si tratta dunque certamente di simmetrie di tipo completamente diverso da quelle fondamentali di cui tratta la fisica. Ne segue che anche la violazione o rottura così frequente di queste simmetrie non può non avere cause essenzialmente diverse.
La terza e ultima delle osservazioni importanti che ci proponevamo di fare è che, oltre alla presenza di determinate simmetrie, costituisce talora un problema biologico sicuramente importante anche la completa ‛assenza' negli organismi di una simmetria che, in base a considerazioni scientifiche generali, sembrava lecito attendersi. Il caso di gran lunga più noto, ma non certo il solo, è quello che va sotto il nome di ‛asimmetria molecolare della biosfera'. Di ciò parleremo a suo tempo.
2. Simmetrie geometriche nel mondo dei viventi.
Vediamo ora di illustrare le precedenti considerazioni generali con un rapido excursus nel mondo della sistematica, per elencare le principali simmetrie concretamente presenti nel mondo della vita. Cercheremo alla fine di trarre qualche conclusione generale sulla loro natura e origine.
a) Animali (Metazoi).
Non esistono tipi animali asimmetrici. Gli elementi di simmetria sono sempre facilmente riconoscibili, anche se la simmetria è spesso imperfetta e più o meno profondamente violata. A eccezione di Celenterati, Ctenofori ed Echinodermi, tutti i tipi hanno simmetria bilaterale semplice: un unico piano di simmetria che passa per i due poli anteriore e posteriore dell'organismo, dividendo quest'ultimo in due metà specularmente simmetriche. In alcuni molluschi (Gasteropodi) la simmetria bilaterale primitiva è mascherata dalla torsione del sacco dei visceri, avvenuta secondariamente, con conseguente asimmetria della conchiglia univalve, generalmente a spirale elicoidale.
I Celenterati sono gli unici metazoi diblastici. Appartengono a questo tipo idre, polipi e meduse. Il loro corpo richiama la morfologia della gastrula e sembra un sacco a doppia parete. La simmetria è di regola ‛raggiata'. Esiste un asse tra la bocca e il polo opposto: le parti del corpo sono tutte disposte attorno a tale asse come i raggi di una ruota. Nei polipi la simmetria, oltre che raggiata, può essere ‛bilaterale doppia': esistono in tal caso due piani di simmetria tra loro ortogonali, ciascuno dei quali divide l'animale in due metà specularmente simmetriche.
Negli Ctenofori, metazoi triblastici, la simmetria è bilaterale doppia e spesso può interpretarsi come rottura di una primitiva simmetria raggiata. Gli Ctenofori sono animali marini dal corpo gelatinoso trasparente, tutti luminescenti. Il più noto è il cosiddetto cinto di Venere (Cestus Veneris), che talora può anche oltrepassare il metro di lunghezza.
Gli Echinodermi, infine, allo stadio larvale hanno simmetria bilaterale doppia, che nelle forme adulte si trasforma di regola in simmetria pentaradiata per la formazione attorno alla bocca di una corona di braccia che facilitano la cattura del cibo. Sono invertebrati marini, tra i quali i più noti sono i ricci e le stelle di mare.
Si osservi che negli animali strutturalmente più semplici la simmetria è raggiata. Via via che gli animali diventano più complessi si ha rottura sempre più decisa della simmetria primitiva e la simmetria diventa bilaterale doppia e infine bilaterale. La stessa simmetria bilaterale, quasi sempre perfetta nei Pesci, è spesso violata negli Uccelli e nei Rettili ove si hanno casi di sviluppo asimmetrico degli organi interni. Imperfetta nei Mammiferi, è ormai soltanto apparente nell'uomo, le cui due metà, destra e sinistra, sono strutturalmente e funzionalmente diverse.
b) Piante (metafiti).
Al contrario di ciò che avviene nel regno animale, nel regno delle piante la maggior parte delle specie non presenta simmetria d'insieme evidente: ogni specie ha una sua struttura caratteristica, ma ogni singolo esemplare è essenzialmente asimmetrico e differisce nei particolari da ogni altro esemplare. Il singolo esemplare si presenta come un tema fondamentale continuamente riprodotto con variazioni occasionali importanti. Viene fatto di pensare a una simmetria di base (caratterizzata ad esempio dal sistema di ramificazione) continuamente violata nel corso dello sviluppo, tanto da renderne alla fine irriconoscibili i caratteri. Ciò può forse mettersi in relazione con la staticità della pianta, che è in grado di reagire alle variazioni occasionali dell'ambiente solo adattando a esse, nella velocità e nella forma, il suo sviluppo.
L'assenza di una simmetria d'insieme non impedisce che si riconoscano spesso in una pianta simmetrie locali di parti o di organi speciali. Nel fusto delle piante (per esempio nel tronco e nei rami degli alberi) si riconoscono spesso regioni a simmetria cilindrica quasi perfetta. Le foglie sono per la maggior parte dotate di un piano di simmetria perpendicolare alle due pagine, superiore e inferiore, tra loro diverse, e passante per il punto di attacco (forma dorsoventrale con simmetria bilaterale).
Simmetrie più complesse si riscontrano nella distribuzione delle foglie lungo i fusti e in quei sistemi di foglie metamorfosate che costituiscono il fiore delle Fanerogame. La distribuzione delle foglie lungo il fusto (fillotassi) è caratteristica della specie: lungi dall'essere casuale, segue alcune regole generali che definiscono un ordine assai rigoroso. In primo luogo, i punti di inserimento delle foglie sul fusto appartengono tutti a un piccolo numero di linee longitudinali (parallele all'asse del fusto) che prendono il nome di ‛ortostiche': le ortostiche definiscono attorno all'asse una simmetria raggiata. Facendo passare due piani longitudinali per le ortostiche di due foglie vicine, si individua un angolo diedro che ha per spigolo l'asse, la cui sezione normale prende il nome di ‛angolo di divergenza'. Il suo valore è caratteristico della specie: esso viene di regola espresso come frazione propria dell'angolo giro.
Per ciò che concerne i valori della divergenza, è stata sviluppata fin dal secolo scorso una complicata numerologia, solo in parte veramente fondata su dati di osservazione. Si ripeterebbero con frequenza particolarmente elevata i valori della serie:
1/2, 1/3, 2/5, 3/8, 5/13, 8/21, 13/34, ... ,
detta serie principale della fillotassi. Si tratta, come si vede, della successione di Fibonacci, in cui ogni numeratore (o denominatore) si ottiene facendo la somma dei due numeratori (o denominatori) precedenti. I successivi termini di questa successione oscillano tra 1/2 e 1/3, avvicinandosi sempre più a un valore limite pari a 137°30′28′′.
Le foglie possono poi essere isolate (alterne), cioè disposte una per nodo, oppure riunite in verticilli di due o più foglie per nodo, ugualmente spaziate lungo l'angolo giro. Nel caso di foglie alterne, collegando il punto di inserimento nel fusto di ciascuna foglia con quello della foglia più vicina, si ottiene una linea spirale immaginaria che unisce tutte le foglie successive di un fusto e che prende il nome di ‛spirale generatrice'. Si chiama ‛ciclo' il percorso lungo tale linea che congiunge due foglie poste immediatamente l'una sotto l'altra, sulla medesima ortostica. Se m/n è la divergenza, il ciclo comprende m giri della spirale generatrice ed è l'ennesima foglia seguente quella che si pensa venga a trovarsi sulla stessa ortostica.
Quando la tassia è verticillata, le foglie di due verticilli sovrapposti non sono mai situate sulla stessa ortostica: i due verticilli sono sfasati l'uno rispetto all'altro di un angolo pari a metà dell'angolo di divergenza. Relativamente frequente nella regione delle foglie normali la formazione di verticilli bimembri alternati tra loro: la divergenza in questo caso vale 1/2 (cioè 180°) e i verticilli consecutivi sono sfasati di 90° ciascuno rispetto al precedente. Questa disposizione delle foglie è detta ‛decussata'.
I verticilli di più membri sono caratteristici dei fiori cosiddetti ciclici. Anche in questo caso, quando il numero dei membri per verticillo non cambia, essi sono alternati. Tuttavia nei verticilli fiorali sovrapposti il numero dei membri spesso cambia, soprattutto nel passaggio dalle foglie corolline alle staminali e da queste ai carpelli. Talora manca un verticillo, che la simmetria generale del fiore farebbe prevedere. Sia il numero dei verticilli fiorali, sia quello dei membri per verticillo può variare tra ampi limiti. Nella maggior parte delle Dicotiledoni quest'ultimo numero è tuttavia 5 o 4 (fiori pentameri e tetrameri) e nelle Monocotiledoni è di regola 3 (fiori trimeri). Esistono anche fiori in cui le foglie fiorali sono disposte a spirale (fiori aciclici) o in parte a spirale e in parte in verticilli (fiori emiciclici). Esempi sono offerti dall'ordine delle Policarpiche (ranuncoli, ninfee, magnolie, ecc.).
La simmetria del fiore (ci riferiamo in particolare alle Angiosperme) è di regola la simmetria raggiata, che è da ritenersi corrispondente alla struttura originaria: è infatti quella che si avvicina di più alla disposizione delle foglie nella regione vegetativa. È però frequente il caso di fiori ‛zigomorfi' o simmetrici, cioè dotati di un solo piano di simmetria che divide il fiore in due metà specularmente simmetriche: tali i fiori delle orchidee, delle Labiate, delle Fumaracee, ecc. Che la zigomorfia tragga origine da una rottura della simmetria raggiata è in ogni caso reso evidente dalla complicata struttura di questi fiori, i cui elementi presentano sempre metamorfosi e riduzioni: si tratta di regola di un adattamento all'impollinazione entomofila. È molto raro il caso di fiori completamente asimmetrici; tuttavia nelle orchidee la simmetria bilaterale è talora rotta da una torsione del ricettacolo. Una prova singolare del fatto che la simmetria raggiata è veramente quella originaria è offerta dal fenomeno delle pelorie, fiori anomali a simmetria raggiata cui dà luogo eccezionalmente una pianta a fiori zigomorfi. In molti gruppi può, del resto, osservarsi direttamente un'evoluzione graduale dall'actinomorfismo (simmetria raggiata) allo zigornorfismo, alla completa irregolarità del fiore.
La disposizione regolare delle foglie è particolarmente evidente nelle sezioni degli apici vegetativi e delle gemme, soprattutto in quelle raccorciate, ove le protuberanze fogliari sono molto fitte. Soprattutto l'esame degli apici vegetativi mostra che le nuove protuberanze fogliari si serrano regolarmente contro le preesistenti, occupando via via tutto lo spazio libero: il posto dove compariranno le nuove foglie è determinato dalla posizione delle vecchie. Si è cercato di giustificare la serie principale della fillotassi col fatto che la relativa disposizione delle foglie permetterebbe la migliore utilizzazione di tutto lo spazio disponibile, in modo che le singole foglie si rechino a vicenda il minimo impedimento possibile nella funzione della nutrizione. In realtà non bisogna dimenticare che, se le simmetrie fogliari sono indiscutibili, esse sono spesso rese imperfette da condizioni locali che possono provocare delle modificazioni della disposizione primitiva, determinando talora anche delle disposizioni irregolari. In generale, poi, non ci si può attendere misure di divergenza più precise di un grado, il che rende vane numerologie complicate. La fillotassi infine è costante per una data specie, ma solo entro certi limiti: essa dipende fino a un certo punto anche dalle condizioni in cui la pianta cresce. Non di rado la disposizione delle foglie cambia gradualmente nel corso dello sviluppo, per cui è differente nella pianta giovane e nella pianta adulta.
c) Funghi.
Nel regno dei funghi, i casi di simmetria più importanti riguardano i corpi fruttiferi dei Basidiomiceti. La maggior parte di essi ha, com'è ben noto, simmetria raggiata, il più delle volte imperfetta a causa delle condizioni particolari di crescita. Il micelio non presenta di regola particolari simmetrie, salvo il caso di Lactarius deliciosus (agarico delizioso o lapacendro buono) che, in zone pianeggianti dei boschi di Conifere, dà talora luogo ai cosiddetti ‛cerchi delle streghe'.
d) Protisti (Eucarioti unicellulari).
Le forme dei Protozoi sono estremamente varie e un gran numero di tipi diversi di simmetria è rappresentato tra essi e tra le alghe unicellulari.
Sono ben noti i mirabili disegni delle teche delle Diatomee, la cui simmetria è bilaterale oppure raggiata. Quest'ultima può dar luogo a teche circolari o poligonali, con asse di simmetria di ogni ordine: ternario, quaternario, ecc. Vi sono diatomee con teche circolari divise in 25 settori.
Simmetrie più complesse sono presentate dai Foraminiferi, i cui gusci possono essere mono- o politalami. Nei primi, la camera può essere sferica o allungata, ramificata o incurvata a spirale. Nei politalami, le logge successive possono essere disposte secondo una serie rettilinea, oppure secondo una spirale piana o tridimensionale.
Anche i Radiolari presentano una grande varietà di forme: sferica, allungata, a campana, a ghianda e così via. La simmetria è sferica o raggiata attorno a un asse.
e) Procarioti.
I Batteri hanno di regola forme tondeggianti (cocchi) o allungate (bacilli). Spesso hanno un asse di simmetria cilindrica e un piano di simmetria perpendicolare all'asse (bacilli, clostridi). Talora il piano di simmetria manca (plectridi). Se il batterio ha la forma di un bastoncello rastremato e incurvato esiste un solo piano di simmetria (vibrioni), ma può anche mancare qualunque elemento di simmetria.
Infine, il batterio allungato può essere ritorto a elica attorno a un asse, come nel caso delle Leptospire e del treponema.
f) Virus.
Particolarmente interessanti sono le simmetrie presentate dalle particelle virali, in quanto direttamente legate al modo di aggregarsi delle proteine globulari che costituiscono il capside. Le forme più comuni sono a bastoncello o isometriche (quasi sferiche). Come esempio delle prime, citiamo il virus del mosaico del tabacco (TMV), il cui capside è formato da 2.130 molecole proteiche globulari (capsomeri) disposte a elica attorno a una cavità centrale. I virus isometrici hanno invece capsidi di forma poliedrica, molto spesso un icosaedro regolare, come il virus iridescente di Tipula paludosa (TIV). Sono anche frequenti forme poliedriche non regolari, come quelle dei batteriofagi T2 e T4 di Escherichia coli, i cui capsidi sembrano essere icosaedri allungati. Molti batteriofagi di forma icosaedrica sono dotati di coda, utilizzata per iniettare l'acido nucleico del virus nel batterio ospite: la particella virale nel suo complesso ha quindi simmetria raggiata attorno a un asse di figura che è un asse quinario.
Si conoscono anche virus le cui particelle hanno struttura più complessa e i cui elementi di simmetria non sono evidenti. Non bisogna dimenticare che non è facile determinare la forma tridimensionale di una particella virale in base alle osservazioni fatte al microscopio elettronico: le procedure preparative causano molto spesso deformazioni difficilmente valutabili. Ad esempio, è stata molte volte, da parte di numerosi ricercatori, segnalata indipendentemente la scoperta di particelle virali di forma ottaedrica: sembra tuttavia che si tratti di artefatti e che la forma effettiva delle particelle osservate fosse quella solita dell'icosaedro regolare.
Va notata la frequenza con cui compare, nelle simmetrie biologiche, l'asse quinario, che non compare in cristallografia.
3. Simmetrie funzionali.
Dello stesso tipo elementare delle simmetrie geometriche sopra considerate sono le simmetrie relative alla dimensione temporale, che si riscontrano molto frequentemente negli organismi viventi. Si tratta di variazioni periodiche nell'andamento di determinati processi, tali cioè che una traslazione per un intervallo di tempo ben determinato riporta il processo a una fase indistinguibile da quella iniziale.
I fenomeni periodici che si svolgono negli organismi sono di tre tipi diversi: 1) si tratta in alcuni casi di processi periodici regolati esclusivamente da forze endogene, che ne determinano tra l'altro il periodo, in modo indipendente da eventuali periodicità delle condizioni ambientali. Tali il battito cardiaco e il ciclo della respirazione polmonare negli animali superiori; 2) in altri casi, il processo biologico è regolato, nella sua periodicità, esclusivamente dalla periodicità di certi processi ambientali, senza il concorso apprezzabile di forze endogene. Tale il processo fotosintetico nelle piante, che si arresta necessariamente con il venir meno della luce solare e riprende ogni giorno con la sua ricomparsa; 3) c'è infine una terza categoria di processi biologici periodici, la cui periodicità è assicurata essenzialmente da forze endogene, ma che vengono contemporaneamente influenzati da periodicità ambientali, che li pilotano al sincronismo. Sono i cosiddetti ‛ritmi biologici', tra i quali i più comuni e sicuramente accertati sono quelli circadiani. Esistono probabilmente anche ritmi biologici circannuali (v. ritmi biologici).
Mentre le prime due categorie comprendono processi particolari, ciascuno dei quali dispone di un meccanismo indipendente che rappresenta una determinata risposta a un determinato problema esistenziale, i ritmi circadiani costituiscono un fenomeno molto più generale, la cui origine e natura sono molto meno evidenti. Anzitutto, essi sono caratterizzati da un periodo proprio di 24 h in natura, mentre in condizioni ambientali artificiali costanti convenientemente scelte la periodicità continua almeno per un certo tempo, in generale con un periodo proprio un po' diverso: prossimo a 24 h, ma sicuramente non coincidente con tale valore.
Ritmi circadiani si osservano in tutte le specie viventi, meno che nei Procarioti, che a quanto sembra ne sono privi. Riguardano le più diverse funzioni organiche, come l'attività motoria, il sonno degli animali, i movimenti delle foglie nelle piante, l'emissione di luce da parte di certi organismi unicellulari, la trasformazione delle pupe in immagini in molti insetti, l'attività di molti enzimi, il volume nucleare di molte cellule, la sintesi del DNA e così via. Sono riscontrabili sia a livello della singola cellula eucariotica, sia a livello dell'organismo multicellulare. È dimostrato che hanno un fondamento genetico: si sono ottenuti, per esempio in Drosophila, mutanti aritmici o con periodo circadiano radicalmente diverso dal normale. Il periodo di una determinata oscillazione circadiana in condizioni ambientali costanti (cioè non pilotata dall'alternanza giorno-notte) si mantiene costante con grande precisione, anche se non è caratteristico della specie e nello stesso individuo può subire variazioni sia spontanee sia indotte da agenti esterni. Esso è praticamente indipendente dalla temperatura, essendo i relativi Q10 compresi tra 0,9 e 1,2, mentre i Q10 dei processi metabolici ordinari sono dell'ordine di 3 o 4. Un certo numero di fenomeni periodici ambientali è in grado di pilotare i ritmi biologici, trascinandoli al sincronismo. Segnali luminosi e il passaggio da un ambiente a un altro possono causare degli spostamenti di fase e dei transienti, mentre interventi di carattere chimico sono di regola del tutto inefficaci.
Questa complessa fenomenologia, per i cui dettagli rinviamo alla letteratura specializzata, ha fatto per un pezzo supporre che ogni organismo contenga un oscillatore centrale, un master clock, cioè un organo realizzante un'oscillazione pilota, da cui dipenderebbero tutti i ritmi propri dell'organismo in questione. È tuttavia necessario dir subito che nessuna delle numerosissime ricerche all'uopo intraprese ha consentito di individuare questo master clock. Non solo, ma quando con interventi esterni si è riusciti a bloccare per un certo tempo qualche ritmo circadiano, al termine del blocco esso ha sempre ripreso con la stessa fase che avrebbe avuto se il blocco non ci fosse stato: si è dovuto quindi concludere che si trattava non del fantomatico master clock (rimasto intoccato), ma di un'oscillazione da questo pilotata. Solo trattamenti praticamente letali hanno potuto interrompere ‛ogni' oscillazione, senza peraltro che fosse possibile individuare l'oscillatore pilota.
4. Struttura fisica e adattamenti evolutivi.
Dopo questa rapida rassegna delle principali simmetrie che compaiono ai diversi livelli di organizzazione biologica, dovremmo ora affrontare il problema della loro natura e origine. Diciamo subito che, per ora, non soltanto non esiste una teoria generale delle simmetrie biologiche universalmente accettata, ma neppure il problema stesso dell'esistenza di queste simmetrie è stato enunciato e affrontato se non in casi particolari. Dobbiamo pertanto limitarci a poche considerazioni di carattere generale, nel tentativo di individuare l'orientamento che potrà domani portare a una migliore comprensione del problema.
Le simmetrie che abbiamo precedentemente elencato sembrano potersi suddividere in due categorie, anche se probabilmente la linea di demarcazione è tutt'altro che netta. Ci sono anzitutto entità biologiche i cui caratteri di simmetria dipendono chiaramente dalla ‛struttura fisica' delle parti che le compongono. L'esempio migliore è quello offerto dai capsidi dei piccoli virus, che, come si è detto, possono essere a bastoncello o isometrici. La scelta della forma dipende evidentemente da come i singoli capsomeri si aggregano gli uni agli altri al momento della polimerizzazione. La fisica di questi processi (che è poi anche la fisica della comparsa e scomparsa di organelli e strutture intracellulari, come ad esempio quelle dell'apparato mitotico) incomincia oggi a essere abbastanza chiara. Si tratta di processi ‛diretti' dalle variazioni di entropia, in cui l'aumento decisivo di entropia che determina l'aggregazione è fornito dalla destrutturazione dell'acqua, che si ha quando due regioni complementari idrofobe della superficie di due macromolecole, già esposte al mezzo circostante, entrano in contatto tra loro.
Per ciò che riguarda le forme possibili per il capside di una particella virale, una teoria limitata ai piccoli virus nudi, cioè privi di involucro, è stata proposta da A. Klug e D.L.D. Caspar nel 1960. Essa non è una teoria generale delle architetture virali: capsidi poliedrici non regolari, come quelli del T2 e del T4, non vi rientrano.
Le ipotesi su cui si basa la teoria di Klug e Caspar sono le seguenti: 1) il capside è formato da unità morfologiche tutte uguali tra loro, i capsomeri, ciascuno dei quali comprende una o più catene polipeptidiche (anche tra loro diverse) per le quali non è necessario postulare particolari condizioni di simmetria; 2) i capsomeri hanno forma globulare e sono in grado di stabilire tra loro legami specifici (ovviamente non covalenti) tra punti ben definiti della loro superficie. I legami a priori possibili sono in numero assai limitato e dipendono dalla natura delle proteine costituenti, cioè in definitiva dal patrimonio ereditario del virus cui appartengono.
Queste due ipotesi sembrano bastare da sole a giustificare l'esistenza dei due tipi fondamentali di capside, elicoidale e poliedrico.
La cosa è evidente per ciò che riguarda i capsidi elicoidali. In un capside elicoidale infatti i capsomeri sono tutti in posizioni equivalenti: ogni unità può essere sovrapposta alla successiva mediante un'unica operazione, consistente in una rotazione attorno all'asse dell'elica, accompagnata da una traslazione parallela a tale asse. Non vi è alcuna restrizione per il numero dei capsomeri per giro dell'elica, numero che (come nel TMV) può benissimo non essere intero. La lunghezza del capside non è determinata dalla simmetria, ma è potenzialmente illimitata: ciò che la determina è la lunghezza della molecola di acido nucleico che esso contiene.
Meno immediata è la giustificazione che la teoria di Klug e Caspar dà per l'esistenza di capsidi isometrici. La domanda da porre è: ‟Per quale ragione un capside isometrico (quasi sferico) deve ammettere un qualche tipo di simmetria?" Per rispondere, supponiamo di aver di fronte a noi il capside già costruito. I capsomeri sono tutti uguali tra loro, ed essendo presumibilmente asimmetrici, ciascuno di essi ha in sostanza un solo modo possibile per legarsi a tutti i capsomeri vicini. Ne segue che ciascun capsomero si trova rispetto ai capsomeri vicini in una posizione affatto equivalente a quella di ciascun altro: vi è un solo movimento che porta un determinato capsomero a coincidere con uno determinato dei capsomeri vicini e tale movimento porta l'intero capside a coincidere con se stesso. I movimenti di ricoprimento del capside con se stesso sono rotazioni attorno al centro di una sfera. La teoria dei gruppi finiti di rotazioni attorno a un punto porta a individuare tutti i tipi di simmetria possibili per un capside che soddisfi alle ipotesi di Klug e Caspar.
Le prime due soluzioni da considerare sono quelle che corrispondono al gruppo ciclico Cp e al gruppo diedrico Dp: esse portano a un capside formato da un numero qualsiasi di unità identiche, disposte simmetricamente attorno a un asse come gli spicchi di un'arancia (simmetria raggiata). Tali unità possono essere asimmetriche (gruppo ciclico) o presentare un piano di simmetria perpendicolare all'asse (gruppo diedrico) come appunto gli spicchi di un'arancia. Queste due prime soluzioni possibili non vengono prese in considerazione da Klug e Caspar: in effetti, nessuno dei virus conosciuti presenta capsidi di questo tipo.
Altre tre soluzioni possibili corrispondono rispettivamente ai seguenti gruppi finiti di rotazioni: tetraedrico (T), ottaedrico (O), icosaedrico (I), e le unità equivalenti asimmetriche di cui risulta costituito il capside nei tre casi sono rispettivamente in numero di 12, 24 e 60.
A titolo di esempio, si consideri un icosaedro regolare. Su ciascuna delle venti facce triangolari si riportino, in prossimità dei vertici e in posizioni simmetriche, tre figure asimmetriche. Si ha un totale di 60 figure in posizioni del tutto equivalenti, che si possono assumere come rappresentazione grafica delle 60 unità strutturali asimmetriche equivalenti di un capside icosaedrico. Naturalmente, il numero dei capsomeri può essere anche minore di 60. Si possono, per esempio, avere solo 12 capsomeri disposti nei vertici dell'icosaedro, ciascuno dei quali tuttavia deve avere simmetria di rotazione quinaria (e comprendere quindi cinque unità polipeptidiche identiche). Si possono anche avere solo 20 capsomeri, uno per faccia dell'icosaedro, ciascuno dotato di simmetria ternaria.
Il fatto singolare che la teoria di Klug e Caspar non spiega è che, come Cp e Dp, anche i due gruppi Te O non sono rappresentati tra i capsidi dei virus fino a oggi conosciuti, mentre, come si è visto, sono molto frequenti, soprattutto tra i piccoli virus, i capsidi a simmetria icosaedrica. Ciò fa pensare che vi sia qualche condizione, non specificata nelle ipotesi di partenza, che favorisce in modo praticamente esclusivo la simmetria icosaedrica rispetto a quelle tetraedrica e ottaedrica. Potrebbe trattarsi del fatto che un contenitore icosaedrico, tra tutti i contenitori a forma di poliedro regolare, è quello più vicino alla forma sferica e quindi realizza il rapporto minimo tra superficie e volume, consentendo, a parità di volume del genoma, un'economia di materiale proteico.
La prima difficoltà che s'incontra è che un gran numero di virus icosaedrici di dimensioni medie e grandi hanno capsidi costituiti da un numero di capsomeri, e quindi di unità strutturali, molto maggiore di 60. Non è possibile realizzare capsidi del genere mediante unità strutturali tutte tra loro uguali e disposte tutte nello stesso modo rispetto alle unità contigue.
Klug e Caspar hanno sviluppato la loro teoria in modo da poterla applicare anche a questi virus poliedrici di maggiori dimensioni. Rilassando il meno possibile la condizione di equivalenza tra tutte le unità strutturali, interpretano i relativi capsidi come derivati dall'icosaedro per divisione delle facce in un certo numero di triangoli minori. Si ottengono in tal modo dei poliedri detti icosideltaedroidi, che tanto più si avvicinano alla forma sferica quanto maggiore è il numero delle unità che li costituiscono. La microscopia elettronica ha confermato in parecchi casi questa estensione della teoria.
Mentre dunque i piccoli virus isometrici hanno simmetria icosaedrica perfetta, nei virus di dimensioni maggiori la simmetria icosaedrica ancora sussiste, ma non è più basata sull'equivalenza di tutti gli elementi strutturali. I capsomeri non sono più tutti uguali tra loro, alcuni essendo costituiti da sei, altri (nei vertici) da cinque unità strutturali. La coda dei batteriofagi, l'allungamento dell'icosaedro nella testa dei fagi della serie T di E. coli rappresentano altre, più radicali, rotture della simmetria base, rotture che vanno evidentemente messe in relazione col particolare patrimonio ereditario del virus.
Questo ci porta naturalmente a parlare della seconda delle due categorie in cui avevamo diviso gli elementi di simmetria presenti nel mondo biologico: elementi di simmetria che non dipendono più esclusivamente dalla struttura fisica delle parti costituenti, ma che dipendono in modo essenziale da adattamenti evolutivi.
Gli adattamenti evolutivi, il raggiungimento di una maggiore complessità organizzativa, hanno come prima conseguenza la rottura di simmetrie preesistenti. Ciò però non significa che tutti gli elementi di simmetria vadano perduti: alcuni scompaiono, mentre i rimanenti vengono a dipendere sempre più direttamente dal patrimonio ereditario del relativo organismo.
È probabile che le simmetrie che noi oggi osserviamo negli organismi unicellulari siano ormai tutte imputabili ad adattamenti evolutivi, sulla base però di una simmetria sferica originaria, dovuta essenzialmente a fenomeni fisici, quali la tensione superficiale e la pressione osmotica.
Una parte delle simmetrie che si osservano negli organismi multicellulari sono ovviamente legate alla geometria della divisione cellulare: forze fisiche e patrimonio specifico ereditario cooperano nel realizzarle, come nel caso della simmetria cilindrica dei tronchi e della simmetria diedrica di molti frutti. Altre simmetrie degli organismi multicellulari sono invece legate al complicato e per noi ancora oscuro processo di sviluppo dell'embrione e il loro carattere adattativo è evidente. Un'ipotetica simmetria sferica originaria si trasforma per successive rotture in simmetria diedrica e poi in simmetria ciclica e in simmetria bilaterale doppia e bilaterale semplice. La simmetria raggiata negli animali è il risultato di un adattamento evolutivo in vista della miglior soluzione del problema della cattura delle prede, mentre la simmetria bilaterale semplice è chiaramente un adattamento che facilita il moto degli animali in presenza del campo gravitazionale terrestre.
Vi sono tuttavia anche organismi in cui ogni simmetria è andata perduta e che sono perciò divenuti completamente asimmetrici per adattamento all'ambiente in cui vivono. Tali per esempio, tra i Protozoi, le amebe, la cui forma cambia continuamente col formarsi di sempre nuovi pseudopodi, mentre i vecchi vengono retratti. Asimmetrici divengono nel corso dello sviluppo postembrionale alcuni pesci bentonici (Pleuronettidi), che poggiano sul fondo marino con uno dei lati del corpo (il sinistro o il destro, secondo la specie). Anche la maggior parte delle conchiglie dei Gasteropodi è asimmetrica, la loro simmetria essendo quella di una spirale che può essere destrorsa o sinistrorsa, il verso essendo determinato geneticamente. Dell'asimmetria delle piante e degli animali superiori abbiamo già parlato. Recentemente, la genetica delle asimmetrie ha incominciato a essere un campo di ricerca promettente.
Ci resta da affrontare il problema della natura e dell'origine delle simmetrie funzionali, cioè in particolare dei ritmi biologici. Qui le ipotesi avanzate sono molte e vanno dalla proposta di variabili periodiche ambientali ‛nascoste', che piloterebbero in ogni caso i processi biologici, alle più varie identificazioni del master clock.
Tra queste ultime, ha avuto particolare fortuna negli ultimi anni l'idea che il master clock abbia a che fare con qualcuno dei cicli regolativi a controreazione presenti nella cellula, cicli che, come tutti i circuiti a controreazione mal regolati, possono dar luogo a oscillazioni periodiche. Si tratta ovviamente di un'ipotesi improbabile per molte ragioni.
La giustificazione sul piano evolutivo dei circuiti di regolazione del metabolismo cellulare consiste nell'economia di materiali di sintesi che essi consentono. La loro risposta non è determinata soltanto da fattori endogeni, ma anche dalle condizioni ambientali in cui la cellula viene a trovarsi: per esempio, dalla presenza o meno nell'ambiente di certe sostanze. Il loro Q10 è quello normale dei processi metabolici. Oscillazioni sono state osservate in vari casi, ma i periodi sono risultati di qualche minuto, cioè di un ordine di grandezza diverso rispetto a quelli dei ritmi circadiani. Infine, eventuali oscillazioni di un ciclo regolativo rappresentano un fenomeno parassita che diminuisce l'efficacia della regolazione: non si capisce quindi come oscillazioni del genere siano potute diventare un elemento funzionale stabile del sistema e quale vantaggio evolutivo possa giustificare un fatto del genere.
Altre ipotesi, come quelle di Bünning e Moser e quella della Sweeney, hanno chiamato in causa le membrane cellulari, supponendo ch'esse svolgano un ruolo fondamentale in un processo oscillatorio identificabile col master clock. Si tratta tuttavia di idee piuttosto vaghe, scarsamente fondate, per ora, su dati osservati.
Un diverso orientamento prende le mosse dall'ipotesi che il fantomatico master clock, che nessuno ha mai trovato, non esista affatto e che il complesso dei ritmi circadiani di un organismo non sia altro che il risultato della più conveniente ‛organizzazione temporale' di tutti i processi che si svolgono in esso. Il punto chiave sarebbe la differenza tra una cellula procariotica, a organizzazione centralizzata, nella quale non esistono ritmi circadiani, e una cellula eucariotica, la cui organizzazione è decentrata, dato che compiti specifici sono affidati a organelli cellulari largamente autonomi. Gli eventi interni si svolgono qui spesso lungo linee processuali parallele e, almeno parzialmente, indipendenti. È ragionevole pensare che la loro programmazione temporale particolareggiata possa subire anche ampie variazioni, purché naturalmente i metaboliti essenziali siano in definitiva sempre pronti in quantità sufficiente al momento della richiesta e siano sempre possibili le risposte essenziali di adattamento immediato all'ambiente.
Per ciascuna delle reazioni chimiche che si svolgono nell'organismo in esame, si possono individuare delle condizioni di optimum: condizioni in cui per esempio l'attività degli enzimi coinvolti è massima e massima è la velocità di reazione. Se però si confrontano con le condizioni effettive in cui quella reazione si svolge di fatto nell'organismo, ci si accorge di regola che in natura si è lontani dall'optimum. Ciò è dovuto al fatto che inevitabilmente un gran numero di reazioni si svolgono contemporaneamente e quindi nelle stesse condizioni ambientali. Le condizioni esterne sono quelle che sono e l'organismo non ha potere su di esse. Per ciò che riguarda le condizioni interne, la specie ha dovuto necessariamente ricorrere a un compromesso: le varie reazioni contemporanee hanno infatti condizioni di optimum diverse. La specie ha realizzato quelle condizioni di compromesso che rappresentano un massimo di vantaggio dal punto di vista evolutivo. Questo compromesso chiama in causa l'organizzazione temporale interna, in quelle cellule in cui i diversi processi godono di una larga autonomia. Una specie del genere otterrà un notevole vantaggio evolutivo alternando e sfasando opportunamente nel tempo i vari processi, in modo da poter realizzare, in tempi diversi, condizioni ambientali interne per ciascuno di essi più vicine alle condizioni di optimum.
È chiaro poi che se le condizioni esterne (illuminazione, temperatura, ecc.) sono già per loro conto periodiche, un ulteriore vantaggio evolutivo si avrà se l'organismo accorda in qualche modo la sua organizzazione temporale interna con la variazione periodica dei parametri esterni.
5. Le simmetrie assenti (asimmetrie).
Giungiamo così all'ultimo degli argomenti che vogliamo trattare: quello delle simmetrie che a priori ci si aspetterebbe di vedere rispettate nel mondo biologico e che invece risultano assenti. Non alludiamo al fatto che nei singoli organismi le simmetrie specifiche risultano sempre più o meno profondamente violate dalla variabilità occasionale, non ereditaria, legata alle particolari condizioni di vita. Queste violazioni casuali non annullano di regola completamente le simmetrie specifiche, anche se le rendono spesso quasi irriconoscibili: tutto ciò che si può dire è che esse cooperano a creare l'individualità di ciascun organismo.
a) La chiralità molecolare della biosfera.
Le simmetrie ‛assenti' sono veramente del tutto assenti e non per variabilità occasionale. Come abbiamo già detto, la più importante riguarda le strutture biologiche a livello molecolare. Vediamo con precisione di che cosa si tratta.
Com'è ben noto, tutte le molecole che non sono sovrapponibili alla loro immagine speculare, e che per ciò sono dette molecole ‛chirali', esistono in due forme, o tipi, che sono appunto specularmente simmetriche: i due ‛enantiomeri' della sostanza in questione. Una molecola costituita da un atomo di carbonio cui siano legati quattro atomi o gruppi tutti diversi tra loro, se non possiede un piano di simmetria, è (come subito si verifica) una molecola chirale.
I due opposti enantiomeri di una sostanza a molecola chirale vengono distinti l'uno dall'altro (e caratterizzati con le lettere D ed L premesse al nome della sostanza in questione) in base a una complicata serie di regole che tengono conto dei numeri atomici e delle masse di tutti i gruppi coinvolti: questa caratterizzazione è fatta in modo da dar ragione del comportamento dei due enantiomeri nelle reazioni chimiche. Inoltre, i due enantiomeri sono di regola entrambi otticamente attivi: una soluzione di uno di essi in un solvente inattivo fa ruotare in un verso il piano di polarizzazione di un fascetto di luce polarizzata rettilineamente; un'identica soluzione dell'altro fa ruotare il piano di polarizzazione dello stesso angolo, ma in verso opposto. Se la rotazione avviene nel senso delle lancette dell'orologio per un osservatore che riceve la luce, l'attività è detta positiva (+); se la rotazione avviene invece in senso opposto a quello delle lancette dell'orologio, l'attività è detta negativa (−). Il tipo di chiralità di un enantiomero (D o L) e il segno della sua attività ottica (+ o −) sono caratteristiche tra loro indipendenti della molecola. Esistono, tanto per fare un esempio, una L(+) alanina e una L(−)serina.
Se in laboratorio procediamo alla sintesi di una sostanza a molecola chirale a partire da sostanze tutte a molecola achirale (cioè dotata di simmetria rispetto a un piano), i due opposti enantiomeri vengono sintetizzati ambedue contemporaneamente e con probabilità uguali: il prodotto è una miscela dei due in parti uguali, miscela che risulta otticamente inattiva e che viene detta ‛racemica'.
In natura esiste un gran numero di sostanze a molecola chirale. Esistono anche molte sostanze, come il quarzo e il clorato di sodio, che pur non avendo molecole chirali (ed essendo otticamente inattive in soluzione) formano cristalli otticamente attivi: in questo caso l'attività ottica dipende da come sono disposti i vari atomi nel reticolo cristallino. Nel mondo inorganico, quando è presente un enantiomero di una certa sostanza, anche l'altro enantiomero è sempre presente e con la stessa abbondanza. Se si trovano dei cristalli otticamente positivi di una certa specie, si ritrovano anche i corrispondenti cristalli negativi e nello stesso numero. Così, in media, nel mondo inorganico non esiste un tipo di chiralità preferenziale, nè a livello delle molecole nè a livello dei cristalli: il mondo inorganico è sostanzialmente racemico.
Ben diversa è la situazione nel mondo della vita. La maggior parte delle sostanze presenti negli organismi viventi è a molecola chirale, ma tali sostanze non sono presenti con ambedue gli enantiomeri nello stesso organismo e neppure con un enantiomero in certi organismi e con l'enantiomero opposto in certi altri. Una determinata sostanza a molecola chirale è di regola presente con lo stesso enantiomero in tutta la biosfera (di qualche eccezione a questa regola parleremo tra poco). Per esempio, tutti gli amminoacidi che entrano a far parte delle proteine biologiche, a eccezione della glicina, hanno molecola chirale: le proteine di tutti gli organismi conosciuti contengono esclusivamente amminoacidi della forma L. Analogamente, gli zuccheri che entrano a far parte delle molecole degli acidi nucleici sono esclusivamente della forma D.
Ecco dunque una simmetria che a priori sembrerebbe del tutto logico attendersi e che in realtà è completamente assente nel mondo biologico. La biosfera, lungi dall'essere racemica, è molecolarmente asimmetrica. È chiaro che si tratta di un fatto che richiede una spiegazione, ma un secolo di discussioni e di ricerche non è bastato per trovarla.
Com'è ben noto, è stato Pasteur a richiamare l'attenzione sul problema, dimostrando contemporaneamente che non si tratta di un problema chimico, ma di un problema biologico e più precisamente di un problema relativo all'origine della vita sulla Terra. Separando manualmente al microscopio i cristalli destrorsi da quelli sinistrorsi di un tartrato otticamente inattivo, egli dimostrò prima di tutto che le sostanze otticamente attive presenti negli organismi danno anch'esse luogo a due opposti enantiomeri, come le sostanze otticamente attive inorganiche. Il fatto che ‛oggi' ogni organismo sia in grado di utilizzare o di sintetizzare uno solo dei due opposti enantiomeri non ha nulla di strano: quell'organismo è costituito da sostanze tutte chirali presenti con un solo enantiomero, ha già in sé l'asimmetria molecolare ed è quindi del tutto naturale che si comporti in tal modo. Ma prima che la vita comparisse sulla Terra, l'ambiente era molecolarmente simmetrico o racemico: come si è prodotta l'asimmetria molecolare che oggi è comune a tutti gli organismi? Come è avvenuta la prima sintesi asimmetrica, in un ambiente in cui non esisteva asimmetria molecolare?
Pur avendo un'idea molto chiara del problema e avendo tentato anche azzardati esperimenti a scopo orientativo, Pasteur non riuscì a trovare la soluzione e dopo di lui non riuscirono a trovarla diverse generazioni di ricercatori. A parte le interpretazioni vitalistiche che non sono mancate, ma che ovviamente sono prive di reale potere esplicativo, le ipotesi che sono state proposte si possono dividere in due gruppi. Da un lato ci sono le ipotesi che tendono a dare del fenomeno una spiegazione deterministica: se una di esse fosse la spiegazione corretta, in ogni altro pianeta in cui si determinassero condizioni simili a quelle vigenti sulla Terra primitiva, favorevoli alla vita, questa evolverebbe con un tipo di asimmetria molecolare identico a quello della biosfera terrestre. Dall'altro lato stanno invece le ipotesi che tendono a dare dello stesso fenomeno una spiegazione probabilistica: se una di esse fosse la spiegazione valida, le biosfere dei pianeti in cui esiste la vita mostrerebbero asimmetria molecolare ora dell'uno ora dell'altro tipo con probabilità uguali. La vita nell'Universo sarebbe dopo tutto, in media, racemica.
Le prime spiegazioni proposte per l'asimmetria molecolare della biosfera sono state di tipo deterministico. L'idea base era quella di attribuire l'instaurarsi dell'asimmetria nei viventi a un processo prebiologico o per lo meno abiologico, in cui un qualche agente fisico ‛asimmetrico' avrebbe portato al prevalere di uno dei due opposti enantiomeri e allo scomparire dell'altro. Per esempio, nel 1894, van't Hoff credette di aver trovato la causa di un processo del genere nella radiazione elettromagnetica circolarmente polarizzata, che si produrrebbe in vari modi alla superficie della Terra a partire dalla radiazione naturale proveniente dal Sole. Cotton aveva dimostrato che i due isomeri ottici di una stessa sostanza assorbono diversamente luce polarizzata circolarmente (per esempio destrorsa), se la sua lunghezza d'onda è prossima a quella di una banda di assorbimento (dicroismo circolare). Se l'energia dei quanti è sufficiente per spezzare qualche legame molecolare, il più assorbente dei due enantiomeri verrà allora distrutto più rapidamente dell'altro e una soluzione della sostanza in questione, inizialmente racemica, potrà diventare, attraverso un prolungato irraggiamento, otticamente attiva. Solo nel 1929 W. Kuhn dimostrò sperimentalmente la realtà del fenomeno, che è però di scarsissima entità. La vera difficoltà sta tuttavia nel fatto che mai nessuno è riuscito a dimostrare che possa formarsi in natura un'apprezzabile componente di radiazione elettromagnetica circolarmente polarizzata prevalentemente in un verso, né per riflessione della luce solare sull'acqua sotto un angolo prossimo all'angolo di Brewster, nè per effetto del campo magnetico terrestre, nè in qualsiasi altro modo. L'ipotesi di van't Hoff ha pertanto dovuto essere abbandonata.
Nel 1957, tuttavia, tale ipotesi è stata riesumata da F. Vester in una nuova forma. Vester ha proposto che la radiazione elettromagnetica responsabile della fotolisi selettiva di un enantiomero sia stata la radiazione di frenamento degli elettroni emessi nelle radioattività β naturali. Tale radiazione è infatti polarizzata circolarmente in verso sinistrorso a causa della non conservazione della parità nelle interazioni deboli. Che l'effetto esista è provato indirettamente dagli esperimenti di Kuhn. Esso è tuttavia così piccolo che fino a oggi non ha potuto esser messo sicuramente in evidenza, neppure ricorrendo a sorgenti β di grande intensità. C'è un'ovvia sproporzione quantitativa tra la causa proposta e l'effetto che si vorrebbe con essa giustificare.
Si è anche osservato che la non conservazione della parità nelle interazioni deboli potrebbe avere come conseguenza una piccolissima differenza tra le energie di legame delle due molecole enantiomeriche di una stessa sostanza chirale, a causa d'interazioni d'ordine superiore tra i nuclei atomici e gli elettroni periferici. La valutazione quantitativa dell'effetto, in base ai dati che si hanno sui limiti entro cui è conservata la parità nelle interazioni elettromagnetiche, sembra tuttavia escludere ogni possibilità di spiegare per questa via l'asimmetria molecolare della biosfera.
Alcuni dei tentativi di spiegazione su basi probabilistiche fanno ricorso a processi fisici o chimici, altri collegano l'affermarsi dell'asimmetria molecolare della biosfera con gli stessi processi selettivi che hanno portato alla comparsa dei primi organismi e accompagnato la loro evoluzione. Nel 1898 K. Pearson ha dimostrato che ogni volta che una miscela racemica subisce un processo fisico o chimico si verificano nelle percentuali presenti dei due enantiomeri delle fluttuazioni statistiche, che possono funzionare da innesco per la selezione definitiva (nella stessa sostanza o in un'altra da essa derivata) del tipo di asimmetria che in esse prevale. È anche stata provata sperimentalmente l'esistenza di processi sia fisici, quale la cristallizzazione da soluzione soprassatura, sia chimici, quale l'autocatalisi e la catalisi mutua, in cui uno dei due enantiomeri viene selezionato a caso, a seconda della simmetria delle molecole che casualmente danno inizio al processo. Questo, una volta iniziato, si svolge poi come un processo a scatto (di tipo autorigenerativo). Processi del genere possono oggi avere un certo interesse anche dal punto di vista industriale, per la produzione di enantiomeri separati di sostanze chirali. Per ciò che concerne la possibilità di spiegare con essi l'asimmetria molecolare della biosfera, però, è da porre in rilievo come essi possano funzionare da selettori di chiralità solo in un ambiente molto limitato, come l'apparecchio di laboratorio. In grande, nell'oceano primitivo ad esempio, proprio per il suo carattere probabilistico, un processo del genere non può che iniziare in più punti, lontani tra loro, in modi indipendenti: il prodotto, in media, non potrà in ogni caso che essere racemico.
Diversamente si presentano i tentativi di spiegazione che fanno dell'asimmetria molecolare della biosfera un fenomeno esclusivamente biologico. A sostegno di una siffatta spiegazione stanno, oltre al fallimento di tutti i tentativi di teorie a base fisico-chimica, anche i fatti seguenti.
1 . Non è vero che gli enantiomeri della chiralità ‛sbagliata' siano sempre del tutto assenti nel mondo biologico. Per esempio, amminoacidi della forma D sono presenti come elementi strutturali nelle pareti batteriche, anche se sono del tutto assenti nelle proteine e negli organismi superiori.
2. Esiste nel fegato dei Mammiferi un enzima, la Damminoacidossidasi, la cui funzione sembra essere quella di distruggere quegli amminoacidi della forma D che venissero eventualmente ingenti dall'organismo. La presenza di questo enzima fa pensare che la sparizione quasi completa dal mondo biologico dei D-amminoacidi sia dopotutto un fatto relativamente recente.
3. Sono stati trovati nel meteorite di Murchison degli amminoacidi di produzione naturale abiologica che sono racemici. Ciò significa che, in 4,6 miliardi di anni, la fotolisi selettiva, se esiste, non l'ha spuntata contro la lentissima reazione di racemizzazione spontanea degli amminoacidi.
Chi ha per primo concepito la chiralità della biosfera terrestre come rottura di una preesistente simmetria, concomitante con il sorgere e l'affermarsi della vita sulla Terra, è stato G. Wald in un lavoro del 1957. Egli è partito dall'idea, verificata d'altra parte sperimentalmente, che da una miscela racemica di monomeri chirali tendano a formarsi per polimerizzazione prevalentemente due tipi di polimeri: quelli formati da monomeri tutti della forma L e quelli formati da monomeri tutti della forma D. Questi sono infatti molto più stabili di tutti i polimeri misti e la velocità di polimerizzazione è per essi di regola assai maggiore.
Wald ne dedusse che, con la comparsa della vita sulla Terra, debbono essersi formate coppie di popolazioni che diremo enantiomeriche: tali cioè che nella prima comparisse una certa serie di sostanze a molecola chirale ciascuna in una sola e ben determinata forma (D per alcune, L per altre), mentre nell'altra popolazione compariva la stessa serie di sostanze a molecola chirale, ciascuna tuttavia nella forma opposta. Le due popolazioni avrebbero per un certo tempo potuto evolvere l'una indipendentemente dall'altra. La selezione naturale avrebbe poi eliminato una di esse, avendo l'altra a un certo punto conseguito un certo non specificato vantaggio evolutivo sulla prima.
Come mai la stessa forma di chiralità si è poi imposta in tutte, senza eccezioni, le specie viventi? Si potrebbe pensare che la selezione sia avvenuta prima che ci fosse una chiara e netta suddivisione dei viventi in specie distinte. La risposta però sarebbe insoddisfacente, poiché tutto fa pensare che la vita possa essersi originata sulla Terra anche più volte, in tempi, luoghi e forme diverse. Di fatto, Wald fa notare come le lunghe e complesse catene alimentari che collegano specie a specie assicurino un deciso vantaggio evolutivo a quelle forme di chiralità che rendono la preda utilizzabile (dal punto di vista alimentare) da parte del predatore. Ciò porta evidentemente al prevalere e al definitivo affermarsi della stessa forma di chiralità in tutta la biosfera. Questa in sintesi la soluzione del problema proposta da Wald. Soluzione che ha però un punto debole. Perché una delle due popolazioni enantiomeriche acquisti sull'altra un vantaggio evolutivo e la elimini, occorre che le due popolazioni siano in qualche modo in competizione tra loro e possono esserlo solo per il rifornimento di una qualche sostanza necessaria per la loro alimentazione presente nell'ambiente in quantità limitata e che, per essere utilizzabile da entrambe, deve essere necessariamente achirale. In tal caso tuttavia, e spesso con maggiore probabilità di un'estinzione concorrenziale di una delle due, può verificarsi il caso di un'evoluzione divergente che, senza estinzioni, porta all'eliminazione della concorrenza. Questa possibilità, che Wald non considera, toglie interesse alla soluzione da lui proposta.
Il problema può tuttavia essere affrontato da un altro punto di vista. Innanzi tutto è da osservare che le coppie di popolazioni enantiomeriche ipotizzate da Wald non possono essere mai esistite. Per convincersene, si pensi a come deve essersi svolto il processo che ha portato alla comparsa, in lagune ai margini dei continenti, delle prime forme di vita. I gas atmosferici erano tutti a molecola achirale e pertanto il brodo primitivo, contenente tra l'altro tutti i precursori delle molecole biologicamente significative, non poteva essere che un brodo racemico. A un certo punto, in qualche modo che ancora non conosciamo (anche se numerose ipotesi sono state fatte in proposito), è incominciata nel brodo la sintesi dei polimeri lineari che hanno alla fine realizzato il ciclo proteine-acidi nucleici-proteine, che è ancor oggi il processo fondamentale della vita. Tale processo deve aver avuto inizio da oligomeri delle due specie fatti di pochi elementi, per passare gradualmente a polimeri via via più lunghi, fino a raggiungere alla fine le lunghezze attuali. All'inizio doveva trattarsi di un processo di sintesi largamente casuale, anche se si può pensare che già allora un tipo di polimeri indirizzasse almeno parzialmente la crescita dell'altro. Solo gradualmente e con l'aumentare delle lunghezze il processo deve essere diventato più preciso, fino ad assumere il carattere quasi deterministico odierno.
Ora, per lunghezze ancora molto modeste, il numero dei possibili polimeri lineari (sia nel caso degli acidi nucleici, sia in quello delle proteine) è talmente grande, da rendere praticamente impossibile che nell'oceano primitivo abbiano potuto casualmente sintetizzarsi dei polimeri identici e di chiralità opposta. Si consideri per esempio che di proteine diverse, tutte costituite da venti residui di amminoacido, ne esistono ben 2020 ≈ 1026.
È quindi chiaro che nel brodo primitivo racemico l'asimmetria molecolare si è sviluppata ‛spontaneamente', man mano che progrediva il processo di sintesi di polimeri sempre più lunghi. E questo non per l'intervento di agenti fisici esterni o come risultato di un qualche processo di autocatalisi: semplicemente per l'impossibilità pratica, per mancanza di tempo e di materia prima, di sintetizzare i due opposti enantiomeri di ogni tipo di polimero lineare. Naturalmente, possono benissimo essersi formate in tal modo, in zone diverse della superficie terrestre, popolazioni ‛diverse' di protorganismi, ciascuna asimmetrica e priva della popolazione enantiomerica corrispondente.
Col tempo, queste popolazioni devono essere venute a contatto tra loro ed essersi trovate a competere per materiale nutritivo, che proveniva allora tutto dall'atmosfera. Ciò deve aver provocato un certo numero di estinzioni e processi di evoluzione divergente per eliminare la concorrenza. Il passo critico deve tuttavia essere stato il passaggio dagli eterotrofi primitivi ai primi organismi autotrofi, certamente fotosintetici, che, ormai indipendenti dal limitato rifornimento di materiali nutritivi dall'atmosfera, debbono essersi rapidamente diffusi in modo prodigioso.
Ci sono molti indizi che fanno pensare che la fotosintesi sia effettivamente un processo molto antico. Quella eseguita oggi dalle piante verdi era già sicuramente attiva 1,8 miliardi di anni fa, quando grandi quantità di ossigeno hanno incominciato a riversarsi nell'atmosfera terrestre. Ma la liberazione di ossigeno per deidrogenazione dell'acqua presumibilmente durava già da molto tempo, anche se l'ossigeno invece di riversarsi nell'atmosfera veniva fissato dal ferro in soluzione, che passando da ferroso a ferrico precipitava. Se, come sembra possibile, le cosiddette formazioni striate di ferro sono la testimonianza rimasta di un processo del genere e dato che molte di esse risalgono a più di 3 miliardi di anni fa, si deve retrodatare fino almeno a tale epoca la comparsa della forma più sofisticata di fotosintesi. D'altra parte, in strati sedimentari molto antichi, sicuramente fino a 2,7 miliardi di anni (Soudan Shale, Michigan) e probabilmente fino a 3,2 miliardi di anni (Fig Tree Chert, Africa sudorientale), sono stati trovati due idrocarburi isoprenoidi, che si ritengono derivati dalla demolizione della molecola della clorofilla. Infine Hodgson e Ponnamperuma nel 1968 hanno dimostrato che negli esperimenti alla Miller, assieme ai precursori delle molecole biologicamente significative, vengono sintetizzate anche delle porfirine, classe di composti alla quale appartiene la dorofilla (v. biofisica).
Tutto ciò porta alla conclusione che la comparsa di una forma anche primitiva di fotosintesi deve aver costituito la prima effettiva differenziazione tra gli organismi eterotrofi primitivi, e aver rappresentato un vantaggio evolutivo decisivo. Le popolazioni coeve non fotosintetiche, di fronte alla rapidissima moltiplicazione dei primi organismi fotosintetici, possono aver evitato l'estinzione solo se capaci di utilizzare i prodotti sintetizzati dagli organismi fotosintetici, o vivendo sui loro rifiuti e sulle loro spoglie, o adattandosi a vivere in simbiosi con loro, o trasformandosi in predatori ai loro danni.
Com'è chiaro, ciò significa il definitivo prevalere di un unico e ben determinato tipo di asimmetria molecolare in tutta la biosfera: quella dei primi organismi fotosintetici.
b) Altri esempi.
Quella molecolare non è l'unica simmetria ‛assente' nel mondo della vita. Un caso singolare è quello della legge di corrispondenza tra triplette di nucleotidi nell'RNA messaggero e residui di amminoacidi nelle relative proteine, legge di corrispondenza che è nota sotto il nome di ‛codice genetico'. Il codice genetico è presumibilmente universale e presenta molte regolarità, sia per ciò che riguarda i gruppi di triplette sinonime (corrispondenti cioè tutte allo stesso amminoacido), sia per ciò che riguarda le proprietà chimiche degli amminoacidi simbolizzati da questi gruppi di triplette. Ciò fa pensare a una simmetria originaria, che ha subito molteplici rotture. Ma queste rotture sono tali da introdurre una singolare asimmetria nella legge. Caratterizzando, come si fa di solito, i quattro tipi di nucleotidi dell'mRNA con l'iniziale del nome della base che in ciascuno di essi compare (Adenina, Uracile, Guanina, Citosina), si ha che gli otto gruppi di triplette:
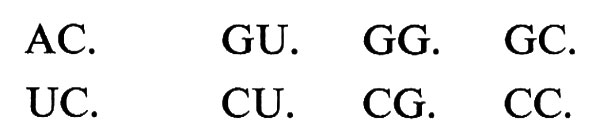
(dove il punto indica il terzo nucleotide della tripletta, che può essere uno qualsiasi) comprendono ciascuno quattro triplette sinonime. Gli altri otto gruppi di triplette invece:
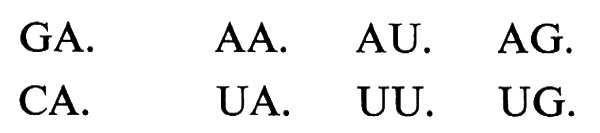
comprendono ciascuno triplette di diverso significato. Se il fatto che le quattro triplette caratterizzate dagli stessi nucleotidi in prima e seconda posizione non hanno sempre lo stesso significato dipende dalla rottura di una simmetria, si tratta di una rottura singolarmente ‛simmetrica', che richiama alla mente l'assenza negli organismi della molecola specularmente simmetrica di ogni molecola chirale in essi presente.
L'ultima simmetria ‛assente' a cui vogliamo accennare riguarda, prevalentemente almeno, la specie Homo sapiens. Negli esemplari della specie si ha, di regola, dominanza della mano destra, cui corrisponde la già citata dominanza dell'emisfero cerebrale sinistro per ciò che riguarda le funzioni linguistiche. Eccezioni a questa regola sono ben note e abbastanza frequenti, ma restano eccezioni in tutte le popolazioni umane. Si pone pertanto il problema di quale sia l'origine di questa dominanza e del perché non esistano popolazioni di mancini, nelle quali le eccezioni siano rappresentate dai destri, tanto più che sembra vi siano indizi che la stessa dominanza prevalga anche presso talune scimmie antropomorfe. È un problema dello stesso tipo di quello della chiralità della biosfera.
Bibliografia.
Ageno, M., L'asimmetria molecolare della biosfera, in Programmi di biofisica, Torino 1979, pp. 56-79.
Ageno, M., I ritmi biologici, in Programmi di biofisica, Torino 1979, pp. 80-118.
Bentley, R., Molecular asymmetry in biology, 2 voll., New York 1969-1970.
Bünning, E., The physiological clock, New York 19733.
Caspar, D. L. D., Klug, A., Physical principles in the construction of regular viruses, in ‟Cold Spring Harbor Symposia on quantitative biology", 1962, XXVII, pp. 1-24.
Denes, F., Umiltà, C. (a cura di), I due cervelli. Neuropsicologia dei processi cognitivi, Bologna 1978.
Klug, A., Caspar, D. L. D., The structure of small viruses, in Advances in virus research (a cura di K. M. Smith e M. A. Lauffer), vol. VII, New York 1960, pp. 225-325.
Lauffer, M. A., Entropy-driven processes in biology, Berlin 1975.
Lawless, J. G., Peterson, E., Amino acids in carbonaceous chondrites, in Cosmochemical evolution and the origin of life (a cura di J. Orò, S. L. Miller, C. Ponnamperuma e R. S. Young), vol. II, Dordrecht-Boston 1974, pp. 3-8.
Miller, S. L., Production of amino acids under possible primitive earth conditions, in ‟Science", 1953, CXVII, pp. 528-529.
Pengelley, E. T. (a cura di), Circannual clocks, New York 1974.
Sweeney, B. M., Rhytmic phenomena in plants, London 1969.
Thompson, D'A.W., On growth and form, 2 voll., London 1917, 19422.