
Sistemi ambientali e loro componenti ecologiche
Sistemi ambientali e loro componenti ecologiche
L'ecologia dei sistemi ambientali (Iandscape ecology) è una disciplina che studia la complessità ambientale analizzando l'importanza dei rapporti spaziali tra le diverse componenti (individui, popolazioni, comunità, mosaici ambientali) del mondo reale. Il sistema ambientale, definito come rappresentazione spaziale di un ecosistema, viene strutturato dalla sovrapposizione degli ambienti così come vengono percepiti dalle diverse specie o modificati dai processi ecologici, quali le esondazioni e gli incendi. Il modo con cui un organismo percepisce l'ambiente in cui vive è specie-specifico e possiamo perciò definire un ambito geobotanico, uno animale e uno antropico. La complessità dei sistemi ambientali viene affrontata su scala spaziale mediante i sistemi informativi territoriali, e i dati vengono analizzati con procedure geostatistiche integrate dalla matematica dei frattali. L'ecologia dei sistemi ambientali è applicata soprattutto alla gestione dei paesaggi a scala umana e, in particolare, alla valutazione, gestione e conservazione della natura. È in questi campi che l'ecologia dei sistemi ambientali trasferisce al meglio le conoscenze sul funzionamento dei mosaici ambientali.
Introduzione
Negli ultimi decenni è apparso sempre più evidente che la dimensione spaziale degli ecosistemi doveva essere allargata per meglio comprendere i complessi processi che interessano la biosfera e più in generale l'ecosfera. La percezione della complessità del pianeta Terra si è radicata sia nella ricerca scientifica che nel più vasto pubblico e la globalizzazione dei problemi ha reso necessario ampliare la scala di analisi e di interpretazione dei fenomeni. Ampliando la scala spazi aie si è reso necessario riconsiderare la dimensione temporale e quindi creare un quadro di riferimento potente e flessibile in grado di muoversi cambiando la scala spazi aie e temporale da un sistema a un altro. La ricerca ecologica legata, fino alle soglie degli anni Ottanta, a concezioni di omogeneità ambientale, di equilibrio ecosistemico e di separazione dell'uomo dai processi ambientali, ha subito, proprio in quest'ultimo decennio, un profondo cambiamento. Infatti, non appena la scala spaziale è stata ampliata, la ricerca ecologica ha iniziato a considerare l'ambiente come un sistema eterogeneo, dove la varietà dei processi ecologici si estrinseca in stati di equilibrio dinamico (flussi omeoretici) e dove i processi condizionati dall'uomo diventano parte integrante dei meccanismi ambientali. L'ecologia dei sistemi ambientali studia la complessità ambientale analizzando soprattutto l'importanza dei rapporti spaziali tra le diverse componenti del mondo reale.
L'eterogeneità sembra il pattem dominante della maggior parte dei sistemi ambientali e alla sua configurazione è riconosciuto un ruolo sempre maggiore nel condizionamento dei processi ecologici. Un sistema ambientale presenta caratteri eterogenei che vengono espressi mediante entità discrete o tessere, che compongono un mosaico strutturalmente e geograficamente distinto. l caratteri salienti di queste tessere quali la dimensione, la forma, il tipo di vegetazione, la ricchezza biologica, l'abbondanza degli organismi e il flusso dei nutrienti contribuiscono alla strutturazione e al mantenimento della complessità dei sistemi ambientali.
Il sistema ambientale
Un sistema ambientale viene definito come la rappresentazione spaziale dell'ecosistema. Si passa così da un'entità descritta solamente sotto l'aspetto funzionale (l'ecosistema) a un'entità definita nello spazio (il sistema ambientale). Un sistema ambientale implica un approccio abbastanza nuovo e ricco di significato. Infatti, aumentando le dimensioni geografiche vengono inglobati un numero crescente di processi e la relativa diversità biologica. Di fatto, si tratta di un sistema di ambienti ciascuno specie-specifico e delimitato dalla specie che ne riconosce i confini e i caratteri a essa favorevoli e con il quale interagisce a una scala speciespecifica. Un sistema ambientale è, quindi, l'insieme di più sistemi specie-specifici che possono sovrapporsi in varia misura tra loro e che concorrono alla complessità ambientale. Questa visione dell'ambiente è caratterizzata da una grande quantità d'informazione che si traduce in un insieme di meccanismi di retro azione che concorrono a regolare e a calibrare i vari processi e alla fine rendono possibile la vita di un gran numero di organismi, creando ruoli ridondanti. Tutto ciò costituisce una premessa alle capacità autopoietiche del sistema stesso, cioè capacità 'creative' che consentono, attraverso meccanismi omeoretici e omeostatici, di adattarsi alle nuove condizioni.
Quando si considera un sistema ambientale, è necessario sapere estrarre le componenti che interessano senza trascurare il suo insieme. Tale approccio diventa nei fatti transdisciplinare e l'analisi di un sistema ambientale concorre in grande misura alla comprensione della complessità ecologica di un'area geografica. Le dimensioni di un sistema ambientale possono variare da pochi centimetri a decine di chilometri; quando si analizzano ampie aree il sistema ambientale viene definito anche paesaggio. Questo termine, che richiama immediatamente l'immagine di una grande area, viene inteso in senso scenografico e geografico in quasi tutte le culture occidentali. Tuttavia ogni organismo percepisce in modo specie-specifico un proprio paesaggio che addirittura può cambiare a seconda del contesto fisiologico in cui tale organismo si trova in quel momento (v. oltre). Il paesaggio viene quindi percepito dall'uomo come il proprio ambiente; di conseguenza, lo studio del suo funzionamento viene definito ecologia del paesaggio e di fatto questa dimensione è la più vicina ai complessi processi che l'uomo, attraverso la propria cultura, ha attivato su un substrato naturale.
Sono state date molte definizioni a un paesaggio percepito dall 'uomo risultato di approcci culturali e scientifici differenti. Il paesaggio è il carattere complessivo di una regione (von Humboldt, 1807); è un'area eterogenea composta da un cluster di ecosistemi interagenti che si ripetono in maniera simile attraverso una regione (Forman e Godron, 1986); rappresenta la totalità di entità fisiche, ecologiche e geografiche che integrano tutti i processi naturali e umani e i loro pattem (Naveh, 1987); è una particolare configurazione di topografia, copertura di vegetazione, uso del suolo e insediamenti umani che sono delimitati da processi naturali e culturali e attività antropiche (Green et al., 1996). W. Haber (1996, comunicazione personale) definisce un paesaggio come una porzione di terra di cui abbiamo la percezione, senza osservare troppo da vicino una singola componente che ci appare familiare.
Attualmente la maggior parte dell'ecologia dei sistemi ambientali è dedicata ai paesaggi modificati dall'uomo sebbene i suoi principi e gli strumenti di analisi possano essere validamente impiegati anche nello studio dei paesaggi non direttamente influenzati dall'uomo. L'ecologia dei sistemi ambientali (landscape ecology), studia quindi sistemi complessi e spesso richiede il riferimento a un organismo o a un gruppo di organismi per essere meglio compresa (Tumer et al., 1995). Di conseguenza il paesaggio percepito dall'uomo risulterà differente dal paesaggio percepito da un coleottero (Wiens e Milne, 1989) e quando si parla di paesaggio umano bisogna necessariamente introdurre anche la componente culturale a lui associata.
Il contributo delle diverse discipline all'ecologia dei sistemi ambientali
Proprio per le sue caratteristiche di disciplina complessa il corpo teorico dell'ecologia dei sistemi ambientali è stato costruito attraverso una serie di integrazioni e di convergenze teoriche e applicative in cui la teoria insulare (MacArthur e Wilson, 1967) e l'ecologia geografica (MacArthur, 1984) occupano una posizione preminente. Queste teorie, per prime, hanno sottolineato l'importanza dell'isolamento e delle dimensioni delle isole nel determinare la ricchezza di specie e la loro abbondanza. Isole piccole e distanti dalla terraferma possiedono un numero inferiore di specie rispetto alle isole più vicine e di dimensioni maggiori. Inoltre, il rischio di estinzione e la probabilità di colonizzazione sono tra loro inversamente correlate. Più un'isola è piccola maggiore è il rischio di estinzione mentre si abbassa la probabilità che una specie la ricolonizzi. Questa teoria può essere efficacemente impiegata anche nella descrizione delle cosiddette isole terrestri, cioè di ambienti che per effetto della frammentazione si sono ridotti di dimensioni e sono stati circondati da ambienti ostili per determinate specie.
Sistemi ambientali ed eterogeneità
Un sistema ambientale è costituito da due componenti fondamentali: la componente geografica o corologica e la componente topologica, cioè funzionale. Di fatto queste due componenti non possono essere separate tra loro senza ridurre drasticamente il livello di informazione che ne deriva quando sono considerate assieme. l sistemi ambientali sono per defrnizione ambienti eterogenei, sono cioè formati da elementi differenti in vario rapporto tra di loro. Questa eterogeneità è un pattem molto importante perché di fatto determina la diversità ecologica di qualsiasi regione. L'eterogeneità può essere costituita sia da strutture rilevabili fisicamente come i boschi o i coltivi, sia da processi come il differente comportamento termico dei versanti e il livello di idoneità ambientale specie-specifica. Comunque, il livello di eterogeneità varia a seconda dell'arrangiamento spaziale delle singole parti componenti. Lo spazio è quindi una componente importante per determinare la diversità delle differenti forme di vita. Per questo l'arrangiamento spaziale delle varie entità, dei loro processi e dei relativi pattem può essere considerato uno dei principali e più influenti paradigmi per introdurre la dimensione spaziale come elemento di per sé importante. La presentazione dei sistemi ambientali come componenti di una gerarchia 'nidificata' (un sistema gerarchico è definito 'nidificato' quando il livello più alto contiene tutti gli altri livelli) incorporando il concetto di scala ha contribuito in maniera determinante a legare tra loro i diversi paradigmi (Allen e Starr, 1982; O'Neill et al., 1986) e le diverse teorie (Delcourt e Delcourt, 1988).
Contemporaneamente la possibilità di misurare la complessità e i relativi pattern di molti sistemi naturali attraverso una geometria non euclidea, cioè impiegando i frattali (Mandelbrot, 1975), ha fatto intravedere un approccio più convincente alla complessità, integrando informazioni in scarsa relazione tra loro. Le nuove teorie sulla eterogeneità (Kolasa e Pickett, 1991) e il ruolo dei regimi di disturbo nei processi ecologici (Pickett e White, 1985) rappresentano significativi passi in avanti in cui il paradigma degli ecotoni (Hansen e Di Castri, 1992), i processi a essi associati quali la connettività e la connessione (Merriam, 1984) e i modelli delle metapopolazioni (Gilpin e Hanski, 1991) sono stati ampiamente incorporati nell'ecologia dei sistemi ambientali (v. oltre). Il riconoscimento di una struttura eterogenea dei sistemi ambientali ha consentito di ampliare e di adattare il modello demografico source and sink (sorgente e scarico) di H.R. Pulliam (1988) assegnando nuovi ruoli alle singole tessere di un sistema ambientale (v. oltre).
Lo studio dell'ecologia dei sistemi ambientali
La complessità dei temi trattati e il contemporaneo contributo di differenti discipline ha portato a considerare un sistema ambientale un'entità che cambia caratteri proprio a seconda dell'approccio. Questa caratteristica è stata ampiamente sottolineata nei principali testi di ecologia dei sistemi ambientali (Forman e Godron, 1986; Naveh e Lieberman, 1994; Forman, 1995; Zonneveld, 1995) dai quali traspare, comunque, una ben differenziata e spesso divergente visione di questa disciplina. Infatti, dalla visione antropocentrica e globalizzante di Z. Naveh e A.S. Lieberman, si passa alla visione più geografica e applicata alla classificazione tipologica di I.S. Zonneveld e, infine, alla visione più geometrica di R.T.T. Forman e M. Godron dove è lo spazio su grande scala a condizionare i processi.
È tempo di riconciliare le diverse concezioni di sistema ambientale e di integrarle considerando le differenti componenti biologiche (Farina, 1998). Storicamente questa disciplina è nata dalle scienze geografiche umane e la prima descrizione di famiglie di pattern e di processi era strettamente legata alla sfera antropica. Il recente sviluppo di studi, soprattutto nell'America Settentrionale, sull'arrangiamento spaziale di pattern e sui processi che interessano il suolo, la vegetazione e la componente animale oltre quella antropica, ha consentito di fare un poderoso balzo in avanti. Devono infatti essere considerate tre prospettive nell'ecologia dei sistemi ambientali: quella umana, quella geobotanica e, infine, quella animale.
La prospettiva umana consente di disaggregare e riaccorpare un sistema ambientale secondo entità funzionali che hanno un significato per l'uomo. La prospettiva geobotanica tiene in considerazione la distribuzione nello spazio delle componenti abiotiche e biotiche, dal paesaggio del suolo a quello 'percepito' dalle piante e alla distribuzione delle entità vegetali quali i boschi e le praterie. Per le piante la 'percezione' viene intesa come la capacità di incorporare le informazioni dall'ambiente circostante. Ciò è in relazione diretta con l'ampiezza dell'adattamento, del tasso di colonizzazione ed estinzione e con la sopravvivenza agli stress naturali e a quelli indotti dall'uomo.
La prospettiva animale considera il modo in cui ciascuna specie percepisce il proprio ambiente. È evidente che una farfalla non può utilizzare gli stessi parametri ambientali di un cervo o di un pesce. Per questo, la sua vita si organizzerà e avrà influenza su una parte ben precisa di un sistema ambientale. La combinazione delle interazioni espresse da una comunità animale costituisce un fattore di condizionamento non trascurabile del sistema ambientale considerato nella sua complessità. In quest' ottica rientra anche l'uomo nella sua duplice identità di specie biologica e di organismo intelligente.
Le prospettive appena descritte non sono in contrasto tra loro poiché ciascuna esplora un dominio di pattern e di processi che di fatto sono le componenti dell'intero sistema biologico ed ecologico; inoltre esistono molti più aspetti in comune, quali la percezione dello spazio e l'arrangiamento spaziale di pattern e di processi, che differenze. Così, alla fine, la strategia comune degli ecologi dei sistemi ambientali è quella di inserire alla più adatta scala spaziotemporale questi pattern e processi.
La dimensione umana di un sistema ambientale è probabilmente la più complicata perché sovrappone la componente animale e culturale dell'uomo. Questa dimensione è in relazione con processi che hanno una scala spaziale e temporale molto ampia. La dimensione biologica dell'uomo può essere comparata a quella degli animali ma la componente culturale dell'umanità è unica, e questa è la componente più interagente con un sistema ambientale, soprattutto per l'attitudine dell'uomo a utilizzare tecnologie che finiscono per superare i limiti fisici e biologici dei sistemi naturali in cui opera.
Le tre prospettive sopra discusse non possono essere affrontate separatamente ma la loro componibilità è legata alla capacità di utilizzare un approccio interdisciplinare se non transdisciplinare. La complessità di un sistema ambientale è così grande che anche un approccio come quello descritto sopra non basta a spiegare tutti i processi ma, se non altro, ha la possibilità di gettare nuova luce su tale complessità e di delineare i limiti delle proprie capacità esplicative.
L'ecologia dei sistemi ambientali è, quindi, una delle discipline ecologiche fra le più promettenti, altamente differenziata al proprio interno ma in grado di esprimere una sola 'anima' legata alla dimensione frnita delle aree di studio. Questa disciplina consente una più precisa sovrapposizione delle informazioni provenienti dal mondo reale con i processi virtuali che oggi la cultura postindustriale sta esasperando in un processo di globalizzazione.
Al momento è molto alto il rischio di considerare l'ecologia dei sistemi ambientali solamente da un punto di vista antropocentrico e questo potrebbe portare a vederla come una disciplina dogmatica priva di fondamenti teorici e di risultanze sperimentali. Peraltro appare inaccettabile e riduttivo considerare l'ecologia dei sistemi ambientali solo come un'ecologia a scala ampia. In realtà uno dei punti di maggiore forza di questa disciplina è proprio quello di essere in grado di trasferire le informazioni attraverso differenti famiglie di processi che si verificano a scale spaziotemporali differenti e di legare le conoscenze che derivano dalla ricerca ecologica puntuale con azioni di progettazione e di programmazione territoriali.
Teorie e modelli incorporati nell'ecologia dei sistemi ambientali
La teoria gerarchica e la teoria della percolazione insieme a due modelli di dinamica di popolazione, il modello della metapopolazione e il modello source and sink, partecipano a creare un potente quadro di riferimento per l'ecologia dei sistemi ambientali (v. oltre). Formulati in diversi contesti questi elementi teorici hanno come punto comune l'interpretazione della complessità di un sistema ambientale. È innegabile il loro ruolo nella creazione di un corpo disciplinare omogeneo che rappresenta un ponte paradigmatico attraverso la complessità di un sistema ambientale.
Teoria gerarchica e struttura di un sistema ambientale
La teoria gerarchica (Allen e Starr, 1982; O'Neill et al., 1986; Allen e Hoekstra, 1992) rappresenta un utile strumento per esplorare molti pattern e processi attraverso diversi livelli di scale spaziotemporali. Considerando la complessità come un attributo intrinseco di un paesaggio, il paradigma della gerarchia spiega come le differenti componenti localizzate a certe scale entrino in contatto con altre visibili a differenti scale di risoluzione. La teoria gerarchica considera un sistema come componente di un sistema più ampio considerato come composto da sotto sistemi. Muovendosi da un sistema a un altro le caratteristiche dei fenomeni cambiano come, per esempio, la classificazione di un paesaggio, transitando tra livelli gerarchici di dimensioni crescenti: ecotopo (unità elementare di un sistema ambientale costituita da componenti biotiche e abiotiche), microchore (distribuzione spaziale di ecotopi), mesochore (sistema ambientale costituito da un insieme di microchore), macrochore (mosaico di sistemi ambientali), megachore (insieme di elementi geografici a scala di diversi km).
Un bacino idrografico è un esempio di sistema gerarchico composto da sottobacini che, a loro volta, sono l'aggregazione di impluvi minori. Un sistema esiste indipendentemente dalle sue componenti e generalmente è in grado di esprimere una propria capacità autorganizzativa e di trasmissione delle informazioni, cioè di essere un sistema cibernetico. Un paesaggio ha una sua complessità e per poterla comprendere a fondo è necessario focalizzarsi su un determinato livello di organizzazione. Esistono innumerevoli livelli gerarchici e quindi altrettanti sistemi tra loro variamente 'nidificati'. Il comportamento in un certo sottosistema condiziona verso il basso e verso l'alto i sistemi vicini. La velocità con cui avvengono i processi e quindi la scala temporale sono specifici di ogni livello. Per questo, per passare da un livello a un altro è necessario modificare la risoluzione. La teoria gerarchica risulta molto utile per comprendere, per esempio, gli effetti dei differenti regimi di disturbo. Così il fuoco appare un elemento a forte impatto distruttivo per il 'sistema albero', e un elemento di 'creatività' ecologica per l'intera foresta in quanto determina nuove condizioni per molte specie di piante e animali.
La teoria della percolazione
La teoria della percolazione è stata formulata nello studio del comportamento dei fluidi attraverso un mezzo (Stauffer, 1985) e ha recentemente trovato interessanti applicazioni per la modellistica dei sistemi ambientali (Gardner et al., 1987). A differenza dei processi di diffusione per i quali ogni particella in un liquido si muove in tutte le direzioni, il processo di percolazione interessa regioni fmite del mezzo in cui il fluido stesso permane. Quando il fluido occupa circa lo 0,5928% dell'intera superficie della matrice questo valore è detto soglia di percolazione (Ziff, 1986); tale stato garantisce, per esempio, alle celle di una matrice di essere in contatto tra loro da un lato all'altro della matrice stessa. Questo limite teorico è comunque un importante riferimento quando si studiano pattern reali quali l'espansione di un'area urbana o l'espansione del bosco per effetto della successione secondaria.
L'importanza di questa teoria nello studio del comportamento di un sistema ambientale è evidente se si pensa che attorno alla soglia critica il contagio, gli incendi delle foreste e le esplosioni demografiche di agenti patogeni presentano i loro stati iniziali di diffusione (Tumer, 1987). Questa teoria è inoltre efficacemente utilizzata nello studio del comportamento animale in un sistema eterogeneo. Infatti, la teoria della percolazione è stata ampiamente testata usando i modelli neutri, cioè matrici quadrate i cui elementi sono riempiti a caso. l modelli neutri prevedono una completa indipendenza spaziale tra le varie celle e gli habitat non mostrano autocorrelazione. Questi modelli considerano sia la scala di risoluzione che il comportamento delle singole celle della matrice stessa simulando, per esempio, la percezione delle varie specie rispetto all'ambiente in cui sono inserite. Poiché la percezione dell'ambiente è specie-specifica, è intuitivo che a parità di celle, che rappresentano un certo ambiente favorevole, ciascuna specie potrà avere la percezione dell'ambiente come habitat percolante, cioè connesso o meno. Questi modelli possono essere applicati per prevedere il limite oltre il quale un ambiente che va incontro a frammentazione perde alcune sue caratteristiche importanti, come la connettività.
Il modello della metapopolazione
L'aumento della frammentazione forestale è una tendenza generale negli ambienti naturali che ha creato appezzamenti di boschi isolati. La frammentazione ha aumentato il rischio di estinzione delle popolazioni relitte. Si può assumere che quando una popolazione vive in ambienti frammentati ed eterogenei percepisca il sistema ambientale in cui vive come un mosaico in cui ambienti favorevoli si alternano ad ambienti più o meno ostili e che i contatti tra le diverse sottopopolazioni siano assicurati solamente da processi di emigrazione e immigrazione. Il rischio di estinzione locale e quello di ricolonizzazione dipendono dalla capacità di mantenere uno scambio di individui. Il successo della ricolonizzazione dipende da molti fattori quali la capacità di dispersione; queste popolazioni sono quindi considerate come componenti di una metapopolazione. Il termine "metapopolazione" è stato introdotto per la prima volta da R. Levins (1970) per descrivere una popolazione di popolazioni (Gilpin e Hanski, 1991; Hanski e Gilpin, 1991). Le metapopolazioni sono sistemi in cui il tasso di estinzione e di ricolonizzazione creano flussi di individui che assicurano la connessione genetica tra le sottopopolazioni.
Il modello della metapopolazione è in stretta relazione con la teoria insulare (MacArthur e Wilson, 1967) ma se ne differenzia per la ridotta ostilità dei sistemi terrestri in cui le metapopolazioni si sviluppano. Nella teoria insulare la distanza tra le diverse isole rappresenta il maggiore meccanismo di isolamento e la distanza dalla terraferma è uno dei fattori determinanti il tasso di estinzione e di ricolonizzazione. Nel modello della metapopolazione, invece, le varie sottopopolazioni sono in rapporto costante tra loro; di conseguenza gli scambi di individui, e quindi del patrimonio genico, devono essere mantenuti elevati. Il modello della metapopolazione trova utile impiego se applicato ad aree frammentate di cui si vogliono conoscere le dinamiche e le probabilità di estinzione o di ricolonizzazione di popolazioni isolate.
Il modello source and sink

Il modello source and sink è stato elaborato da H.R. Pulliam (1988) per studiare le risposte ambientali in alcune specie di uccelli. Secondo questo modello, che si adatta molto bene ad ambienti eterogenei dove le risorse non sono uniformemente distribuite, una popolazione presenta caratteri source quando il rapporto tra le nascite e le morti è sempre superiore all'unità. In altre parole in una popolazione con caratteri source vi è sempre un surplus di individui che tendono ad allontanarsi. Al contrario, una popolazione di tipo sink presenta un rapporto tra le nascite e le morti inferiore all'unità; detta popolazione può persistere solamente se a essa si vanno ad aggiungere nuovi arrivati. Questo modello può essere utilizzato anche per caratterizzare le tessere di un ambiente eterogeneo a seconda delle loro capacità di sostenere una popolazione source o sink (fig. 1). Appare quindi estremamente utile nella valutazione ambientale anche perché è strettamente legato al modello della metapopolazione. l caratteri source o sink di una tessera dipendono, inoltre, dalle dimensioni stesse delle tessere. In genere larghe tessere di un ambiente hanno caratteristiche prevalentemente source rispetto a quelle di piccole dimensioni. l caratteri source o sink di una tessera dipendono anche dalla stagionalità delle risorse e dal ruolo che queste risorse giocano per la sopravvivenza di una specie ai vari livelli di aggregazione. Infatti, questo modello può essere esteso proficuamente anche al di fuori dell'ambito della demografia di popolazioni. In questo modo se la presenza di risorse in una certa tessera garantisce la sopravvivenza di una specie, possiamo di fatto considerla una tessera source.
Esistono varianti del modello source and sink: per esempio, A.R. Watkinson e W.I Sutherland (1995) hanno coniato il termine pseudo-sink per descrivere un ambiente che apparentemente presenta caratteri sink, ma in realtà, anche se l'immigrazione cessa, le popolazioni si stabilizzano su livelli minimi senza scomparire del tutto. Vi sono inoltre casi speciali in cui ambienti con caratteri sink appaiono così attrattivi per le specie da assumere l'apparenza di ambienti source (Pulliam, 1996). È il caso di molti ambienti modificati dall'uomo che offrono molte risorse ma non garantiscono, per esempio, sufficiente protezione dai predatori o in cui l'uomo stesso produce, nel periodo sensibile per una certa specie, un disturbo tale da vanificare il successo riproduttivo o addirittura interromperlo. Ovviamente, va detto che questo modello non consente sempre di desumere la qualità di un ambiente, soprattutto se si utilizza il criterio demografico. Un ambiente con caratteri sink può avere in un certo momento più individui di un ambiente source. Vi sono, infatti, popolazioni in cui la maggior parte degli individui si trova in tessere di tipo sink.
Gli ecotoni
Il carattere eterogeneo di un sistema ambientale determina il contatto tra tessere con caratteristiche differenti (Pickett e White, 1985; Hansen et al., 1992). Le zone di contatto vengono chiamate ecotoni. Gli ecotoni furono descritti da F.E. Clements (1905) come zone di tensione dove le specie delle comunità adiacenti si incontrano. Lo stesso autore fu il primo a utilizzare il termine come parola di derivazione greca composta da οiκoς e τovoς.
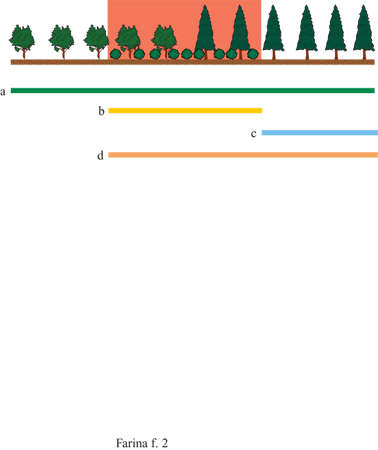
Vi sono molte definizioni degli ecotoni a seconda che si prendano in considerazione pattem, processi o veri e propri habitat. Gli ecotoni sono quindi anche habitat in cui si incontrano specie che possiedono caratteristiche tali da poterutilizzare al meglio le qualità transitorie di queste strutture che di fatto sono ambienti di frontiera (Ricklefs, 1973). In realtà, sebbene il concetto di ecotono sia facilmente comprensibile, lo studio presenta una certa difficoltà legata alle proprietà scalari dell'ecotono stesso. Per esempio, a mano a mano che ci si avvicina a un ecotono diventa sempre più difficile distinguerne la struttura che si era apprezzata a distanza maggiore. Lo studio degli ecotoni presenta difficoltà dovute non solo alle loro caratteristiche scalari ma anche al fatto che ogni specie biologica, interagendo in maniera specie-specifica con l'ambiente, percepisce e quindi colloca gli ecotoni in posizioni differenti (fig. 2). Quindi gli ecotoni che possono essere apprezzati dall'analisi di una foto aerea o dall'analisi di una carta geologica non sempre hanno effetti su tutte le specie.
Gli ecotoni sono strutture importanti per il funzionamento di un sistema ambientale. Infatti, nelle zone di contatto tra tessere differenti avviene il trasferimento di nutrienti, acqua, spore, semi e animali. Per il loro carattere di transizione appaiono inoltre ottimi indicatori dei cambiamenti climatici legati all'aumento di CO2 e dell'inquinamento atmosferico. Gli ecotoni sono caratterizzati inoltre dall'avere tassi di produttività primaria e secondaria elevati perché, in genere, beneficiano delle caratteristiche degli ambienti costituenti. Negli ambienti modificati dall'uomo gli ecotoni costituiti da boschetti, siepi, filari e bordi di campi, rappresentano spesso aree-rifugio per molte specie sensibili al regime di disturbo dell'uomo.
Lungo i fiumi e nelle aree lagunari gli ecotoni sono le aree di congiunzione tra l'ambiente terrestre e quello acquatico intercettando il flusso di nutrienti che si muove tra questi due ambienti. L'importanza degli ecotoni è stata da tempo riconosciuta (Clements, 1897; Livingston, 1903; Griggs, 1914) e, sebbene in vari periodi autorevoli ecologi ne abbiano evidenziato l'importanza (Weaver e Clements, 1929; Odum, 1959; Daubenmire, 1968; Ricklefs, 1973), solo recentemente sono stati effettuati studi sistematici su questo argomento (Di Castri et al., 1988; Naiman e Decamps, 1990; Holland et al., 1991; Hansen e Di Castri, 1992; Risser, 1995).
Studiare il funzionamento di un sistema ambientale impiegando il paradigma degli ecotoni sembra oggi un approccio molto promettente per comprendere come la materia, l'energia e gli organismi interagiscono con l'eterogeneità ambientale e, quindi, con la complessità stessa di un sistema ambientale. Non a caso la ricchezza e il movimento degli organismi, il flusso e l'accumulo della materia e dell'energia e, infine, la diffusione del disturbo sono fortemente influenzati dai bordi del mosaico ambientale. Lo studio degli ecotoni diventa, quindi, fondamentale per comprendere i processi ecologici, soprattutto se l'oggetto di studio è un sistema ambientale che, per definizione, è una parte del mondo reale su cui siamo interessati a compiere investigazioni. L'approccio dell'ecologia ecosistemica che si focalizza soprattutto sui meccanismi che agiscono in un ecosistema selezionando opportunamente l'interno di aree omogenee, sembra sostanzialmente diverso da quello dell'ecologia dei sistemi ambientali che considera il funzionamento di sistemi ecologici inseriti in un contesto geografico.
È possibile stabilire una classificazione degli ecotoni a seconda della loro origine. M.M. Holland (1988) ha proposto quattro tipi di ecotoni: quelli creati e mantenuti dal disturbo umano (le siepi, le arature o le erbe falciate); quelli creati e mantenuti dai processi naturali come la successione ecologica o la transizione tra ambiente terrestre e acquatico creata dalle dighe costruite dai castori; quelli creati da processi naturali e mantenuti dall'uomo, come le lagune costiere o le boscaglie ripariali; infine, quelli creati dall 'uomo e mantenuti dai processi naturali, come le aree paludose attorno a invasi artificiali. In genere, vi sono sostanziali differenze tra ecotoni naturali ed ecotoni prodotti dall 'uomo. Nel primo caso si tratta sempre di aree di transizione con gradienti poco marcati, mentre, in quelli creati dall 'uomo i bordi appaiono molto evidenti e le caratteristiche tra le tessere cambiano bruscamente.
Caratteristiche degli ecotoni
Sebbene di difficile approccio, è comunque possibile individuare le strutture e le funzioni degli ecotoni. La dimensione di un ecotono è data dall'area di transizione tra un sistema e un altro. La forma può essere lineare, come negli ecotoni creati dall 'uomo, circolare o convoluta. Questa caratteristica determina il tasso di trasferimento dell'informazione, dell'energia e del materiale attraverso gli ecotoni stessi.
La struttura biologica è determinata dalla biomassa o dalla densità degli organismi dominanti. Il contrasto strutturale è determinato dalla differenza tra la struttura dell'ecotono e quella degli ambienti. L'eterogeneità interna è rappresentata dalla variabilità dei cambiamenti all'interno dell'ecotono. La densità dell'ecotono è data dalla quantità di superficie ecotonale presente in una certa area. Vi sono, inoltre, importanti variabili funzionali quali la stabilità che misura il grado di suscettibilità al cambiamento da parte di un ecotono. La resilienza misura invece la capacità di un ecotono di tornare alle condizioni iniziali dopo un disturbo.
Fattori esterni e fattori interni concorrono alla formazione degli ecotoni. I fattori esterni sono determinati da processi che intervengono in un sistema ambientale, i fattori interni rappresentano di fatto le discontinuità. Molte specie animali, per esempio, sono in grado di creare condizioni ostili per altre; inoltre, a livello intraspecifico la territorialità può assumere caratteri ecotonali simili a un cambiamento delle condizioni vegetazionali in un mosaico ambientale. Alcune specie di piante modificano a tal punto il pH del suolo che, di fatto, creano condizioni ecotonali più o meno ostili per altre piante. L'ombra stessa prodotta da una chioma è in grado di ridurre le capacità di crescita di altre piante. Molte specie invasive quali il topinambur (Helianthus tuberosus) crescono con popolamenti così fitti da impedire ad altre specie di insediarsi.
Molte specie animali creano, con la loro attività, degli ecotoni. È il caso del campagnolo delle nevi (Microtus nivalis), una piccola arvicola che vive nelle praterie montane oltre il limite della vegetazione arborea. La sua incessante attività alimentare, che non si arresta neanche in inverno, condiziona fortemente la distribuzione del mirtillo (Vaccinium spp.) dei cui fusticini sempreverdi si nutre, soprattutto durante la stagione fredda (Farina et al., 1986). Allo stesso modo talpe, formiche e termiti modificano con lo scavo e la costruzione di domi di terra o di nidi epigei la copertura della vegetazione alterando anche la microtopografia del suolo.
In definitiva, la presenza degli ecotoni può essere rilevata sia a livello di megascala, come una regione bioclimatica, che a livello di microscala, cioè in pochi centimetri quadrati di terreno. Il ruolo degli ecotoni in un sistema ambientale è abbastanza chiaro. Essi rappresentano delle membrane semipermeabili attraverso le quali si verificano i processi di diffusione passiva, come il movimento delle masse d'aria o dell'acqua che sposta materia e organismi, o i processi di diffusione attiva attraverso i quali gli organismi spostano di fatto biomassa da una parte all'altra di un sistema ambientale modificando anche l'informazione.
La presenza di ecotoni contribuisce senz'altro alla stabilità di un sistema garantendo la presenza di una complessità necessaria a ridurre il rischio di un brusco degrado e di perdita delle capacità autopoietiche del sistema stesso. Così un ecotono ripariale (rappresentato dalla vegetazione della sponda fluviale) riduce gli effetti delle esondazioni durante i periodi di piena trattenendo, tra l'altro, grande quantità di sostanza organica che viene successivamente utilizzata nella catena trofica di questi sistemi. Vi sono organismi che vivono esclusivamente in aree ecotonali come la maggior parte degli anfibi. Per esempio, la grande ricchezza di specie della barriera corallina è legata ai caratteri ecotonali di tali strutture. Inoltre, è proprio nelle aree di incontro delle grandi masse d'acqua marine che si hanno le maggiori concentrazioni di specie planctoniche.
Lungo gli ecotoni l'abbondanza e la diversità animale sono più alte delle aree adiacenti ma, ovviamente, questo è vero anche per il rischio di predazione che aumenta in questi ambienti lineari. Insomma, queste strutture possono giocare svariati ruoli, anche se da sole non rappresentano paradigmi sufficientemente potenti da spiegare la complessità e quindi il funzionamento di un sistema ambientale. Ma se a questo paradigma si appaia, per esempio, il modello source and sink o la teoria gerarchica allora molta informazione può essere desunta dalla loro considerazione.
I paesaggi culturali
La maggior parte dei sistemi ambientali terrestri è stata profondamente modificata dall'attività dell 'uomo che ha costruito città, tagliato foreste, modificato il paesaggio vegetale, costruito strade e autostrade e indotto modificazioni del clima. Secondo la prospettiva dell'ecologia dei sistemi ambientali queste modificazioni avvengono a carico dell'arrangiamento spaziale delle tessere, modificano la frammentazione di grandi tessere forestali e aumentano le aree di transizione, cioè gli ecotoni.
Le caratteristiche emergenti di tali sistemi modificati sono date dall'aumento della frammentazione delle originali coperture forestali e dall'aumento della eterogeneità ambientale. In ambienti così strutturati il ruolo dell'ecologia dei sistemi ambientali è sempre più importante perché è proprio per le sue capacità di studiare pattern e processi a scale diverse, considerando anche gli ambienti modificati dall 'uomo, che questa disciplina ecologica diventa un importante strumento di analisi. Questo approccio sta assumendo quindi un ruolo sempre più decisivo in molti settori dell'attività umana che hanno rapporti più diretti con l'ambiente e i suoi caratteri. Infatti, l'ecologia dei sistemi ambientali è in grado di seguire i processi ecologici attraverso un ampio intervallo di scale spaziali e temporali che esprimono la complessità dei sistemi stessi. Tale disciplina è, quindi, in grado di dirigere la gestione e la pianificazione di specie chiave, resti di foreste, reti di margini e di influenzare o indirizzare lo sviluppo urbano. In particolare può affrontare le problematiche di molti sistemi ambientali che, modificati dall'uomo, hanno assunto un elevato valore in termini di complessità, diversità biologica e valore scenico. Questi sistemi vengono chiamati paesaggi culturali, cioè paesaggi che sono stati sottoposti per lungo tempo al regime di disturbo prodotto dall'uomo e in cui la qualità e l'arrangiamento spaziale delle tessere componenti sono il risultato di complessi meccanismi di retro azione tra uomo e natura.
Un paesaggio culturale riflette le interazioni tra uomo e ambiente naturale e rappresenta un fenomeno complesso con identità tangibili e non tangibili (Plackter e Rossler, 1995). Nel 1991 il segretariato dell'UNESCO (United Nations Educational, Scientific and Cultural Organization) propose alcune linee guida per identificare questi paesaggi. l paesaggi culturali presentano, in genere, una struttura complessa formata da un mosaico a grana fine in cui i fisiotopi, cioè le unità fisiche di un paesaggio, sono ben localizzati e utilizzati in modo differente dall'agricoltura, dalla pastorizia e dalla forestazione. Nelle aree collinari e montane sono comuni le modificazioni dei pendii mediante il terrazzamento per facilitare la lavorazione del terreno e, al tempo stesso, per ridurre l'erosione e trattenere i nutrienti.
l paesaggi culturali possono rappresentare un buon modello per testare la possibilità di assicurare la presenza dell 'uomo senza distruggere le risorse e senza perturbare in modo irreversibile gli ambienti. Oggi ciò può apparire utopico per la sfera tecnologica, ma la lezione che deriva dai paesaggi culturali non può essere ignorata e tale modello andrebbe riproposto in molte regioni della Terra. Ogni paesaggio culturale è il prodotto delle interazioni tra l'uomo e l'ambiente su tempi molto lunghi e per questo quando l'attività umana si riduce, come accade sempre più spesso con l'abbandono delle pratiche agricole nelle regioni montane, la fragilità di questo sistema appare evidente e i paesaggi culturali perdono parte della loro complessità. Per esempio, i terrazzamenti abbandonati vanno incontro a un rapido degrado.
Generalmente ogni paesaggio culturale presenta differenze se comparato ad altri; tuttavia in genere presenta maggiori strutture lineari e spesso più ambienti aperti rispetto ai paesaggi indisturbati. In alcuni casi è completamente differente come, per esempio, in quello delle oasi del deserto che hanno una maggiore densità di palme rispetto al deserto circostante.
l paesaggi culturali sono in genere il risultato delle interazioni tra le popolazioni residenti e l'ambiente naturale, ma anche popolazioni nomadi quali i mongoli devono aver giocato un ruolo determinante nella caratterizzazione delle steppe. Intere regioni, come il Mediterraneo, possono essere considerate paesaggi culturali e le modificazioni prodotte dall'uomo possono ritenersi irreversibili sia per la diversità biologica sia per quella ecologica. Per questo è viva la preoccupazione per la scomparsa di queste importanti testimonianze che, tra l'altro, hanno un forte richiamo turistico. Bisogna valutare con attenzione ciò che spesso appare naturale e spontaneo; in regioni come quella mediterranea la diversità biologica è stata ridotta da migliaia di anni e non esistono più nuove specie per riempire le nicchie ecologiche lasciate vuote da specie che non sono riuscite ad adattarsi al disturbo arrecato dall'uomo. l paesaggi culturali sono fragili e necessitano della gestione umana per essere mantenuti; soprattutto nelle regioni a clima arido l'abbandono favorisce gli incendi con frequenze anomale che determinano la riduzione della complessità ecologica. Infatti, sebbene la maggior parte delle specie vegetali mostri un'elevata tolleranza al fuoco, è proprio la maggiore frequenza che impedisce l'avvio della successione ecologica con conseguente aumento del rischio di erosione durante i periodi di piogge intense.
La gestione dei sistemi ambientali

Attualmente la gestione delle specie sito per sito o la gestione degli ecosistemi utilizzando singole specie non è ritenuta una strategia efficace. L'ecologia dei sistemi ambientali considera i siti non isolati tra loro e rilevanti le interazioni che scaturiscono dalle relazioni tra le differenti componenti. La scala dei sistemi ambientali rappresenta uno dei più efficienti approcci per la gestione degli ecosistemi (fig. 5). Infatti, un sistema ambientale può essere considerato come un contenitore geograficamente definito dove non solo sono presenti pattem e processi spontanei ma anche pattem e processi indotti dall 'uomo e dalla sua cultura. Un bacino idrografico può essere considerato come un sistema ambientale ma è possibile individuare altre unità funzionali come l'ecotopo, le micro-, meso-, macro- e megachores. Per esempio, la scala di bacino idrografico appare un efficiente approccio per la gestione di un sistema ambientale. Il bacino idrografico può essere considerato un'unità multifunzionale di un paesaggio in cui il deflusso delle acque e il movimento dei nutrienti sono processi distintivi.
Nella gestione di un sistema ambientale possono essere utilizzate efficacemente le specie 'ombrello', cioè quelle specie la cui presenza condiziona il funzionamento dell'intero ecosistema. Così, nelle grandi praterie dell'America Settentrionale il bisonte ha rappresentato una specie chiave nel funzionamento del sistema prateria. Il pascolo, il calpestio e il deposito di escrementi hanno modificato l'intero ecosistema. Lo stesso ruolo è svolto da molti ungulati nelle savane africane.
Quando una specie chiave scompare l'intero sistema collassa modificandosi. Così la scomparsa del bisonte probabilmente è alla base della scomparsa di molte graminacee e della loro sostituzione con arbusti.
Principi per la conservazione delle riserve e dei parchi
Alcuni principi derivanti sia dalla teoria insulare (MacArthur e Wilson, 1967) sia dalla combinazione dei paradigmi dell'ecologia dei sistemi ambientali consentono di descrivere procedure rivolte alla conservazione di aree di rilevante importanza naturalistica. Per questo è importante stimare la superficie minima di un'area. Le dimensioni di una foresta tropicale, per esempio, devono essere grandi tanto da mantenere la diversità biologica. In ambienti temperati anche superfici di foresta modeste mantengono un'elevata biodiversità. Le aree contigue conservano più specie di foresta interna rispetto alle aree disgiunte anche se la superficie totale è equivalente. In ambienti forestali, tessere separate fra loro ma vicine presentano più specie rispetto a tessere distanti. Tessere disgiunte ma collegate da corridoi di aree protette sono più favorevoli che non tessere completamente isolate. A parità di valore un'area protetta di forma arrotondata è preferibile a un'area di forma allungata dove il rapporto tra il perimetro e l'area è elevato. Questi principi sottolineano l'importanza dell'area, la forma delle tessere, la connessione e lo sviluppo degli ecotoni.
Conservazione e regimi di disturbo
La struttura del mosaico ambientale è sempre più importante per la conservazione della natura. In particolare, i regimi di disturbo a grande scala sono fondamentali per mantenere i processi ecologici (Baker, 1992). Ovviamente, per poter attivare i regimi di disturbo sono necessari grandi spazi in modo da poter interessare solo una parte del sistema stesso. Infatti, come nel caso degli incendi, è necessario che alcune tessere sopravvivano in maniera da attuarsi uno shif ting mosaic (mosaico ambientale che cambia continuamente) e dare modo alle tessere disturbate di recuperare attraverso la successione secondaria. Così la conservazione di comunità climax può essere assicurata se queste sono circondate da un buffer (area tampone) di comunità successionali e quindi più giovani.
Può comunque sembrare un'utopia mantenere grandi aree non sottoposte all'attività umana e per questo il ruolo dell'ecologia dei sistemi ambientali è proprio quello di valutare le possibilità che hanno vari mosaici ambientali - composti da relitti di foreste, coltivazioni, sistemi fluviali, aree urbane e infrastrutture - di sopportare una ragionevole biodiversità prendendo in considerazione molte variabili. Una delle regole principali nella delimitazione di un'area protetta è quella di garantire un'area minima in cui le dinamiche dei disturbi possano attuarsi. In altre parole, l'area dovrebbe contenere gruppi di tessere sottoposte a un regime di disturbo in grado di assicurare una struttura del sistema relativamente stabile a scala temporale bassa. In questo modo dovrebbe venirsi a creare uno shifting mosaic che fornisca le massime potenzialità per assicurare la complessità ecologica. I confini di un parco o di una riserva dovrebbero essere scelti in maniera che coincidano con l'inizio o la fine dei disturbi come, per esempio, i limiti delle esondazioni fluviali, o i punti dove il fuoco si arresta spontaneamente. È bene evitare di identificare una riserva o un parco con un solo tipo di habitat e, ancora, sarebbe opportuno estendere alcuni vincoli anche al di fuori dei limiti dei parchi (Halladay e Gilmour, 1995) in maniera che processi e specie possano completare le loro funzioni.
Il ruolo dei corridoi nella conservazione
Il corridoio, cioè una stretta fascia di un determinato ambiente che si incunea in ambienti differenti, trova molti consensi soprattutto tra i pianificatori ma solleva molte perplessità tra gli ecologi (Harrison, 1992). Come spesso accade, quando si studia un sistema complesso la funzione dei corridoi non sempre coincide con i pattern previsti, vale a dire siepi, filari di alberi e corsi d'acqua. In alcuni casi queste strutture sono veri corridoi, ma in altri pur avendo pattern ascrivibili a questo tipo non svolgono di fatto tale funzione.
Non vi è dubbio che soprattutto le specie più mobili utilizzino vie preferenziali per i loro spostamenti ma non sempre queste vie sono riconoscibili secondo i criteri di valutazione dell'uomo. Spesso sono le strutture morfologiche come le creste montane o i canyon ad attrarre gli animali, facilitando per esempio i loro movimenti. La lepre preferisce spostarsi utilizzando i sentieri degli animali domestici e lo stesso vale anche per la volpe e per molti altri piccoli mammiferi. Per alcuni grandi carnivori come il puma i corridoi, rappresentati da aree meno ricche di prede, vengono percorsi a una velocità maggiore delle aree di caccia. La conoscenza del territorio da parte di molti animali è fondamentale per la sopravvivenza sia per gli erbivori che si spostano da un pascolo a un altro, sia per i predatori che cercano le loro prede esplorando il loro territorio sistematicamente.
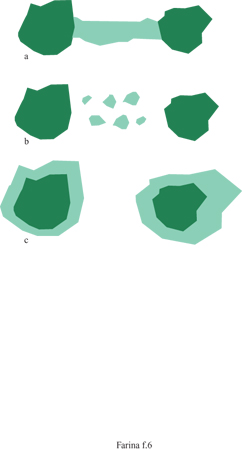
La conservazione di frammenti di habitat e popolazioni relitte Negli ambienti dominati dall'uomo è molto comune la presenza di zone frammentate che sono occupate da popolazioni più o meno isolate la cui sopravvivenza dipende dal grado di connettività fra le tessere (fig. 6). A livello locale possono verificarsi estinzioni delle popolazioni in certe tessere ma se l'estinzione non avviene contemporaneamente in tutte le tessere, la presenza di una certa specie è comunque garantita a livello di mosaico ambientale. Dimensione, forma e qualità delle tessere hanno una notevole influenza sul rischio di predazione e sul successo riproduttivo. Le dimensioni di una tessera non sono sufficienti per caratterizzarla qualitativamente, infatti due tessere di uguale dimensione, ma di forma diversa, esprimono un rapporto tra superficie e volume piuttosto diverso.
Nuove linee per la conservazione ecologica: la conservazione dei processi
Utilizzando i paradigmi dell'ecologia dei sistemi ambientali è più agevole e realistico attuare politiche di conservazione dei flussi e dei processi piuttosto che concentrarsi sulla conservazione di una determinata specie almeno quando non esistono specie di grande importanza da conservare. Spesso una specie è presente solo per periodi effimeri compresi tra pochi giorni e alcuni anni e, in genere, è legata a particolari stadi della successione ecologica. La rarità di una specie, inoltre, è spesso legata ai suoi cicli demografici; una specie può essere rara ai margini del proprio areale e molto comune nell'area baricentrica. Per esempio, la conservazione dei flussi migratori degli uccelli del paleartico occidentale non può essere affidata alla protezione di una singola specie ma all'individuazione delle aree utilizzate dai migratori per le loro soste lungo le rotte. La maggior parte dei migratori terrestri si ferma lungo la rotta migratoria in ambienti diversi da quelli di svernamento o di riproduzione e, come nel caso dei silvidi e dei turdidi, ama ambienti aperti come la savana con cespugli e prati-pascoli, fondamentali per la ricerca del cibo. Alcune specie, al contrario, sono specialiste durante la riproduzione ma diventano generaliste durante gli spostamenti migratori.
Strutture gerarchiche dei sistemi ambientali e conservazione della biodiversità
Considerando come gerarchico un sistema ambientale, le varie componenti sono caratterizzate da dinamiche differenti a seconda del livello di appartenenza. È quindi impossibile conservare questi sistemi senza tenere conto di tale strutturazione; con questo approccio, di fatto, si tende a conservare l'intera diversità di un sistema ambientale. La conservazione sia della diversità biologica sia di quella ecologica appare, quindi, come un obiettivo strategico una volta superati i falsi miti della conservazione delle specie. Con questa prospettiva l'economia e la diversità possono trovare un punto d'incontro e garantire soluzioni ecologicamente valide. In altre parole diventa prioritario trovare, in un mondo dominato dall'uomo, regimi di disturbo che 'mimino' i disturbi naturali responsabili delle dinamiche ambientali che siano altrettanto utili per il mantenimento di livelli economici soddisfacenti. Infatti, le crisi socioeconomiche delle varie società in diversi periodi e in diverse parti della Terra sono state in gran parte provocate da una improvvisa divergenza tra processi ecologici e processi socioeconomici.
Metodi per lo studio dei sistemi ambientali
L'elaborazione spaziale dei dati è un tema centrale dell'ecologia dei sistemi ambientali e per attuarla è necessario disporre di una serie di strumenti, anche molto sofisticati, che consentano di raccogliere e gestire l'informazione geografica e quella ecologica. Gli strumenti e le tecniche spaziano necessariamente dal rilevamento a distanza (remote sensing) ai sistemi informativi geografici (GlS, Geographic lnformation Systems).

Sono applicabili almeno due livelli principali di analisi; il primo riguarda lo studio della forma e della dimensione delle tessere che costituiscono un sistema ambientale, mentre, il secondo è rappresentato dallo studio delle caratteristiche stesse del mosaico ambientale (fig. 7). Di conseguenza, è importante il rapporto tra il perimetro e la superficie delle tessere, la loro forma e le loro dimensioni. L'analisi del mosaico ambientale si basa sul conteggio della diversità delle tessere, sulle loro adiacenze o sul loro livello di aggregazione. La distanza tra queste assume particolare importanza per valutare il grado di connettività che il sistema può garantire per un determinato organismo. Inoltre, la distanza implica, di fatto, l'aumento del rischio da predazione e una riduzione della probabilità di trasporto di vettori (intesi come forze fisiche o animali che spostano materiale o energia). Alcune tecniche provengono da metodi di analisi delle immagini elettroniche e si basano sulle caratteristiche dei singoli pixel. Queste analisi possono essere effettuate su dati disponibili in formato raster (rappresentazione di un processo mediante unità spaziali discrete, celle o pixel).

Un approccio interessante allo studio dei pattern spaziali deriva dall'applicazione del concetto di lacunarità in una distribuzione frattale (Mandelbrot, 1975). Un oggetto che ha una bassa lacunarità presenta invarianza quando viene traslato, mentre un oggetto che presenta un' elevata eterogeneità nella distribuzione delle lacune assume una diversa conformazione spaziale quando viene traslato (Farina, 1998). Di fatto la lacunarità risulta un approccio promettente per studiare l'eterogeneità ambientale. Poiché anche lo studio dei margini assume una particolare rilevanza, la complessità di un sistema ambientale può essere studiata non solo attraverso l'analisi della complessità delle tessere (eterogeneità) ma anche attraverso quella dei margini (fig. 8).
La struttura frattale dei sistemi ambientali
Gli organismi e le loro aggregazioni presentano una distribuzione spaziale che riflette l'eterogeneità dei sistemi ambientali. La distribuzione degli organismi è determinata, infatti, dall'ineguale distribuzione delle risorse e quindi delle biomasse. Per misurare questa complessità un nuovo e promettente strumento è costituito dalla geometria frattale (Mandelbrot, 1975).
Negli ambienti modificati dall'uomo a dominare sono le forme geometriche regolari o comunque lineari. Quando, al contrario, si considera un sistema naturale l'irregolarità delle forme impedisce un approccio di tipo euclideo, poco efficace per descrivere la complessità ambientale. L'impiego della geometria frattale è ritenuto molto promettente non solo per l'analisi di pattern ma anche di fenomeni come l'ineguale distribuzione degli organismi, i loro movimenti e più in generale il loro comportamento.
Il termine "frattale" è stato coniato da B.B. Mandelbrot nel 1975 per descrivere un oggetto irregolare in cui le irregolarità si presentano a tutte le scale. Un frattale può essere considerato un oggetto o un pattern che non ha dimensioni intere. Quando un oggetto frattale presenta strutture a grande scala che si ripetono a scale più piccole si dice che mostra un'autosimilarità. Si possono distinguere due differenti tipi di frattali: quelli regolari e quelli random. Nei primi l'autosimilarità è la stessa a ogni livello, cioè un oggetto è copia di se stesso a ogni scala. Alla seconda categoria appartengono i frattali naturali come le nubi, le linee di costa e l'abbondanza di organismi nello spazio. L'autosimilarità rilevata solo a livello statistico, invece, è un concetto scala-dipendente. La costa per esempio è un oggetto frattale e la sua lunghezza dipende dalla scala di risoluzione con cui viene misurata. La geometria frattale presenta un ampio intervallo di applicazioni (v. anche il saggio di L. Pietronero, Il semplice e il complesso dalla fisica alla biologia) nelle differenti discipline che comprendono le scienze naturali come la geologia, l'idraulica, la struttura del suolo, le dinamiche, il trasporto microbico e, infine, la struttura della vegetazione. In particolare l'analisi frattale è utile nello studio di quegli oggetti che presentano ambiguità dovute alle proprietà scalari. Così l'abbondanza di un organismo è in rapporto alla scala con cui vengono aggregate le informazioni, cioè il livello di risoluzione con cui queste distribuzioni vengono trattate.
I sistemi informativi territoriali
l sistemi informativi territoriali rappresentano la tecnologia per la gestione dei dati spaziali attraverso l'impiego di computer e di strumenti per l'input di dati (tavole di digitalizzazione, scanner) e la loro restituzione grafica (stampanti, plotter). In genere, oggi un buon GlS è rappresentato da una combinazione di cartografia digitale, rilevamento a distanza e gestione di data base (Burrough, 1986). Un GlS appare oggi indispensabile per compiere investigazioni quali i cambiamenti nell 'uso del suolo, i pattern della vegetazione, la distribuzione e lo spostamento degli animali in un sistema ambientale e la modellistica dei processi ecologici (Tomlin, 1990; Coulson et al., 1991; Maguire et al., 1991). Esistono GlS di tipo vettori aie e di tipo raster. l primi presentano una più precisa rappresentazione degli oggetti in quanto espressi dai loro stessi contorni, mentre i secondi scompongono l'oggetto in parti discrete o celle (pixel). Il vantaggio dell'utilizzo dei sistemi di tipo raster consiste principalmente nella facilità con cui le informazioni possono essere manipolate. Infatti, più celle, parte di una matrice, possono essere aggregate tra loro, confrontate con le loro vicine e riaggregate per livelli di risoluzione differenti. Inoltre, le informazioni derivanti dalle immagini satellitari sono in formato raster e possono essere manipolate con relativa facilità da un GlS in formato raster. Questo sistema ha però lo svantaggio di richiedere molta memoria per archiviare le informazioni.
Il rilevamento a distanza
Le strutture di un sistema ambientale sono in genere facilmente rilevabili da immagini da satellite, soprattutto quando si opera con sensori di 10m di risoluzione al suolo. In particolare, quando si analizza un'area geografica ampia, la risoluzione oggi disponibile, che varia dai lO ai 30 m, è sufficiente per ricavare informazioni sulla struttura stessa del paesaggio considerato. È infatti possibile ricavare molte informazioni se la scala geografica considerata risulta ampia. In particolare le tecniche di rilevamento a distanza sono in grado di evidenziare le zone di transizione tra i diversi ambienti. La complessità dei margini è un buon indicatore dei processi in atto in un'area determinata. Oggi, in associazione con le tecniche di rilevamento a distanza, vi è la possibilità di localizzare al suolo i pattem rilevati dai sensori montati sui satelliti. Queste tecniche di posizionamento su scala globale (GPS, Global Positioning Systems), messe a punto per fini militari e per la navigazione aerea e marittima, sono oggi disponibili a costi contenuti (Leick, 1990; Hofmann-Wellenhof et al., 1993) e trovano quindi anche in ecologia notevoli possibilità applicative (Trimble Navigation, 1994). Un gruppo di satelliti consente di identificare al suolo, attraverso un metodo di triangolazione, le coordinate geografiche in cui il ricevitore (rover) è collocato con approssimazioni che vanno da 25 metri fino a pochi centimetri a seconda della configurazione. Così, grazie ai GPS, è possibile disegnare aree, localizzare animali e seguime i movimenti. Allo stesso modo è possibile attraversare un ambiente e censire gli uccelli localizzando gli avvistamenti (Farina, 1998). Le informazioni vengono memorizzate in tempo reale e quindi trasferite, al termine della sessione di lavoro, in un GIS per ulteriori trasformazioni. Con questa tecnica è possibile seguire al suolo l'andamento di un pattem come un corso d'acqua, una linea di costa o un ecotono.
Bibliografia citata
ALLEN, T.F.H., STARR, T.B. (1982) Hierarchy. Perspectives Jor ecological complexity. Chicago-Londra, University of Chicago Press.
ALLEN, T.F.H., HOEKSTRA, T.W. (1992) Toward a unified ecology. New York, Columbia University Press.
BAKER, W.L. (1992) The landscape ecology of large disturbances in the design and management of nature reserves. Landscape Ecology, 7, 181-194.
BURROUGH, P.A. (1986) Principles of geographic information systems for land resources assessment. Oxford, Clarendon Press e Oxford-New York, Oxford University Press.
CLEMENTS, F .E. (1897) Peculiar zonal formations of the great plains. Am. Nat., 31, 968.
CLEMENTS, F.E. (1905) Research methods in ecology. Lincoln, University Publishing Co.
COULSON, R.N., LOVELADY, C.N., FLAMM, R.O., SPRADLING, S.L., SAUNDERS, M.C. (1991) In Quantitative methods in landscape ecology. The analysis and interpretation of landscape heterogeneity, a c. di Tumer M.G., Gardner R.H., New York, Springer-Verlag, pp. 153-172.
DAUBENMIRE, R. (1968) Plant communities. A textbook of plant synecology. New York, Harper & Row. DELCOURT, H.R., DELCOURT, P.A. (1988) Quatemary landscape ecology: relevant scales in space and time. Landscape ecol., 2, 23-44.
DI CASTRI, F., HANSEN, A.J., HOLLAND, M.M. (1988) A new look at ecotones: emerging intemational projects on landscape boundaries. Biology International, Special Issue,17, 1-163.
FARINA, A. (1998) Principles and methods in landscape ecology. Londra, Chapman & Hall.
FARINA, A., TOMASELLI, M., FERRARI, C. (1986) Studi sui rapporti tra vegetazione e Microtus nivalis (Martins) nell'Appennino reggiano. Prime osservazioni (Mammalia, Rodentia). Atti 51 Conv. UZI, Roma. BolI. Zool., 53, 86.
FORMAN, R.T.T. (1995) Land mosaics. The ecology of landscapes and regions. Cambridge, Cambridge Academic Press.
FORMAN, R.T.T., GODRON, M. (1986) Landscape ecology. New York, Wiley & Sons.
GARDNER, R.H., MILNE, B.T., TURNER, M.G., O'NEILL, R.V. (1987) Neutral models for the analysis of a broad-scale landscape pattems. Landscape Ecology, l, 19-28.
GILPIN, M., HANSKI, I., a c. di (1991) Metapopulation dynamics: empirical and theoretical investigations. Londra, Academic Press for the Liunean Society of London.
GREEN, B.H., SIMMONS, E.A., WOLTJER, I. (1996) Landscape conservation. Some steps towards developing a new conservation dimension. A draft report of the IUCN-CESP landscape Conservation Working Group. Department of Agriculture, Horticulture and Enviroument, Wye College, Ashford, Kent, UK.
GRIGGS, R.F. (1914) Observations on the behaviour of some species on the edges of their ranges. Bull. Torrey Bot. Club, 41, 25-49.
HALLADAY, D., GILMOUR, D.A., a c. di (1995) Conservation biodiversity outside protected areas. The role of traditional agroecosystems. Gland-Cambridge, IUCN.
HANSEN, A.J., DI CASTRI, F., a c. di (1992) Landscape boundaries. Consequences for biotic diversity and ecological flows. New York, Springer-Verlag.
HANSEN, A.J., DI CASTRI, F., NAIMAN, R.I. (1992) Ecotones: what and why? In Landscape boundaries. Consequences for biotic diversity and ecological flows, a c. di Hansen A.J., Di Castri F., New York, Springer-Verlag, pp. 9-46.
HANSKI, I., GILPIN, M. (1991) Metapopulation dynamics: brief history and conceptual domain. Biological J. Linnean Society, 42, 3-16.
HARRISON, R.L. (1992) Toward a theory of inter-refuge corridor design. Conservation Biology, 6, 293-295.
HOFMANN-WELLENHOF, B., LICHTENEGER, H., COLLINS, J. (1993) Global positioning system, theory and practice, 2a ed., Vieuna-New York, Springer-Verlag.
HOLLAND, M.M., (1988) SCOPE/MAB Technical consultations on landscape boundaries. Report of a SCOPE/MAP workshop ecotones. In A new look at ecotones: emerging international projects on landscape boundaries., a c. di Di Castri F., Hansen A.I., Holland M.M, Parigi, Intemational Union ofBiological Sciences.
HOLLAND, M.M., RISSER, P.G., NAIMAN, R.J. (1991) Ecotones. The role of landscape boundaries in the management and restoration of changing environments. Londra, Chapman & Hall.
HUMBOLDT, A. VON, (1807) Ideen zu einer geographie der pjlanzen nebst einem gemalde der tropenlander. Tubinga.
KOLASA, J., PICKETT, S.T.A., a c. di (1991) Ecological heterogeneity. New York, Springer-Verlag.
LEICK, A. (1990) GPS satellite surveying. New York, Wiley & Sons.
LEVINS, R. (1970) Extinction. In Some mathematical questions in biology. Lectures in mathematics in the life sciences, a c. di Gertenshaubert M., Providence, American Mathematical Society, pp. 77-107.
LIVINGSTON, B.E. (1903) The distribution of the upland societies of Kent County. Michigan Bot. Gaz., 35, 36-55.
MAcARTHUR, R.H. (1984) Geographical ecology, patterns in the distribution of species. Princeton, Princeton University Press.
MAcARTHUR, R.H., WILSON, E.O. (1967) The Theory of island biogeography. Princeton, Princeton University Press.
MAGUlRE, D.J., GOODCHILD, M.F., RHIND, D.W., a c. di (1991) Geographical information systems. Harlow, Longman Scientific & Technical e New York, Wiley.
MANDELBROT, B.B. (1975) Les objects fractals: forme, hasard et dimension. Parigi, Flammarion.
MERRIAM, G. (1984) Connectivity: a fundamental characteristic of landscape pattem. In Methodology in landscape ecological research and planning: proc. of the first international seminar of the International Association of Landscape Ecology (IALE). Roskilde, Danimarca, pp. 5-15.
NAIMAN, R.J., DEcAMPS, H. (1990) The ecology and management of aquatic-terrestrial ecotones. Parigi, Unesco; Park Ridge, The Parthenon Pub. Group.
NAVEH, Z. (1987) Biocybemetic and thermodynamic perspectives of landscape functions and land use pattems. Landscape Ecology, 1, 75-83.
NAVEH, Z., LIEBERMAN, A.S. (1994) Landscape ecology. Theory and application, 2a ed., New York, Springer-Verlag.
ODUM, E.P. (1959) Fundamentals of ecology, 2a ed., Filadelfia, Sannders.
O'NEILL, R.V., DEANGELIS, D.L., WAIDE, J.B., ALLEN, T.F.H. (1986) A hierarchical concept of ecosystems. Princeton, Princeton University Press.
PICKETT, S.T.A., WHITE, P.S. (1985) The ecology oJnatural disturbance and patch dynamics. Orlando, Academic Press.
PLACKTER, H., RossLER, M. (1995) Cultural landscapes: reconnecting culture and nature. In Cultural landscapes of universal value, a c. di Droste B. van, Plackter P., Rossler H., Jena-New York, G. Fischer.
PULLIAM, H.R. (1988) Source-sink, and population regulation. The American Naturalist, 132, 652-661.
PULLIAM, H.R. (1996) Sources and sinks: empirical evidence and population consequences. In Population dynamics in ecological space and time, a c. di Rhodes O.E., Chesser R.K., Smith, M.H., Chicago, The University of Chicago Press, pp. 45-69.
RICKLEFS, R.E. (1973) Ecology. Newton, Chiron Press.
RISSER, P.G., a c. di (1995) Understanding and managing ecotones. Ecology International, 22.
STAUFFER, D. (1985) Introduction of percolation theory. Londra, Taylor & Francis.
TOMLIN, C.D. (1990) Geographic information systems and cartographic modelling. Englewood Cliffs, Prentice-Hall.
TRIMBLE NAVIGATION (1994) GPS mapping systems, generaI reference. Sunnyvale, Trimble Navigation Ltd. TURNER, M.G. (1987) Spatial simulation of landscape changes in Georgia: a comparison of 3 transition models. Landscape Ecology, l, 29-36.
TURNER, M.G., GARDNER, R.H., O'NEILL, R.V. (1995) Ecological dynamics at broad scales. Bioscience, 29, suppl.
WATKINSON, A.R., SUTHERLAND, W.J. (1995) Source, sinks and pseudo-sinks. J. AnimaI Ecology, 64, 126-130.
WIENS, J.A., MILNE, B.T. (1989) Scaling oflandscape in landscape ecology, or landscape ecology from a beetle's perspective. Landscape Ecology, 3, 87-96.
WEAVER, J.E., CLEMENTS, F.E. (1929) Plant ecology. New York, McGraw-Hill Book Company.
ZIFF, R. (1986) Test scaling exponents for percolation-cluster perimeters. and ecology. Phys. Rev. Lett., 56, 545-548.
ZONNEVELD, I.S. (1995) Land ecology: an introduction to landscape ecology as a base for land evaluation, land management and conservation. Amsterdam, SPB Academic Publishing.